
Submitted:
21 October 2025
Posted:
22 October 2025
You are already at the latest version
Abstract
Although the digestion of dietary and endogenous proteins by the exocrine pancreatic proteases and peptidases in the small intestine luminal fluid is highly efficient for most proteins, it has been approximated that between 3 and 11 g of proteins and peptides are transferred from the small to the large intestine in humans. Here, the proteins and peptides are degraded by the bacterial proteases and peptidases, releasing amino acids in free form. These amino acids are utilized by the abundant population of bacteria, notably amino acids that the bacteria are unable to synthesize, and which can thus be considered as indispensable for these microorganisms. The metabolism of amino acids by colonic bacteria is related to the synthesis of proteins while some specific amino acids are used for DNA and RNA synthesis and in catabolic pathways allowing ATP synthesis and leading to the production of metabolites with documented roles in the metabolism and physiology of both commensal and pathogenic intestinal microorganisms. In the present review, we examine the recycling of undigested host’s protein by the large intestine microbiota and the metabolism of amino acids by the intestinal bacteria. We describe in addition how these metabolic pathways are involved in bacterial growth and communication, as well as in bacterial physiology in terms of virulence, resistance to detrimental environmental conditions, and capacity to form biofilms.
Keywords:
undigested proteins
; intestinal microbiota
; metabolic recycling
; bacterial anabolism
; bacterial physiology
1. Introduction
The intestine of mammals is well known to be inhabited by a complex mixture of microorganisms among which bacteria have been the subject of a vast majority of studies [1]. These microorganisms, which include bacteria, archaea, viruses and fungi are forming the intestinal microbiota. Briefly, archaea is a group of prokaryotes which, based on some of their different molecular, biochemical, and morphological characteristics when compared to bacteria, form a separate domain of the living world than bacteria [2]. Many of the archaea present in the intestinal content are methane-producing archaea called methanogens, which are thus operative to produce methane from hydrogen under anaerobic conditions, as those prevailing in the colon [3,4]. The human intestinal virome is largely dominated by a population of phages which, by infecting specific populations of intestinal bacteria, are one important element implicated in the bacterial composition of the intestinal microbiota [5,6]. Fungi, although representing a minor component of the microbiota, are eukaryotic organisms which can be involved in a competitive relationship with bacteria in the human gut [7]. Lastly, although classically not being considered as a part of the intestinal microbiota, protozoans in the intestine are eukaryotic organisms belonging to the group of protists, and as such, are components of the gut ecosystem [8].
In healthy individuals, the relatively rapid transit of the luminal content through the small intestine does not allow the development of a large concentration of bacteria in the proximal parts of the small intestine. In sharp contrast, the concentration of bacteria greatly increases in the ileum, and in the large intestine (cecum, colon and rectum) [9]. Indeed, in the human colon, the concentration of bacteria represents as much as 109-1012 colony-forming unit (CFU) per g of content. As a result, bacteria in human feces represent more than 50% of the total solid part [10].
The spectacular increase in the number of bacteria in the large intestine luminal fluid is notably related to the much slower transit of the intestinal content in the large than in the small intestine, allowing intense metabolism of the available substrates provided by the host. The transit time in the human colon is rather variable, averaging approximately 60 hours [11,12]. In healthy adults, the luminal contents of the distal parts of the intestinal tract are characterized by a low level of oxygen tension, thus promoting notably the development of large communities of predominant obligate anaerobes [13], coexisting with facultative anaerobes [14].
The substrates available for the growth of bacteria within the colonic fluid are mainly undigested substrates which are transferred from the ileum to the colon through the ileocecal junction [15]. Within the human large intestine, the cecum and the proximal (ascending) colon receive a fluid which is relatively rich in undigested substrates, while the bacteria in the transverse and distal (descending) colon received presumably more limited quantities of substrates, mainly because the available substrates have been already partly used in the more proximal parts of the large intestine. Regarding the protein part of the undigested substances moving from the ileum to the large intestine, although the metabolism of these compounds in the different parts of the large intestine is poorly documented, it appears that protein fermentation occurs mainly within the distal parts of the colon [16,17]. The process of protein fermentation within the large intestine is influenced by numerous parameters including substrate availability, transit time, pH and osmolarity. The ratio of available carbohydrates/proteins represent an important determinant for substrate utilization by the gut microbiota [18], and in humans, the higher the availability of complex carbohydrates (for a given quantity of proteins), the lower microbiota ferments protein [19,20].
When the fermentable carbohydrates are supplied in high amounts to the intestinal microbiota metabolic activity, the nitrogenous substrates are used for bacterial anabolism and bacterial growth. Conversely, proteins are degraded by intestinal bacteria when other sources of energy are scarce. Longer transit time and more alkaline pH are notably associated with higher level of protein fermentation [21,22]. Thus, the carbohydrates/proteins ratio influences the metabolic activity of the intestinal bacteria, and thus the amounts of amino acids metabolized in the different metabolic pathways [23].
2. Origin of Proteins Available for the Large Intestine Microbiota
In the human large intestine luminal fluid, amino acids in free form are usually not the main nitrogenous source directly available for the microbiota, since amino acid absorption by the enterocytes of the small intestine represents a highly efficient process [24]. Then, few amino acids are transferred from the small to the large intestine through the ileocecal junction [25]. Indeed, in the large intestine, the amino acids originate mainly from the proteins that have not been digested or not fully digested in the small intestinal content [26]. Dietary protein digestion is overall an efficacious process with digestibility in the small intestine being in most cases equal or even higher than 90% [27,28]. However, some proteins from dietary origin are more resistant to the digestion by the catalytic activities of proteases secreted by the exocrine pancreas. These proteins include for instance gluten which is relatively resistant to digestion [29]. It has been estimated from clinical studies performed in the last decades that between 1.5 and 5 g of nitrogen are transferred every day from the small to the large intestine [30,31,32,33]. Of note, the nitrogenous material found in the distal small intestine is not only originating from the diet, but for a major part from endogenous sources. In fact, it has been approximated that roughly 40% of the nitrogenous material transferred to the large intestine originate from alimentation, while the remaining 60% originate from the endogenous compartment [25]. Such nitrogenous material is not constituted exclusively by proteins and peptides, but also, although to a much lower extent, by amino acids (approximately 10%), and by minor quantities of urea and ammonia [33].
By using a classical conversion factor between nitrogen and proteins equal to 6.25, by estimating, as explained above, that 40% of the nitrogenous material transferred to the large intestine originates from undigested dietary proteins, and by considering that 90% of the nitrogenous material is made by proteins and peptides, it can be roughly estimated that approximately between 3 and 11 g of alimentary proteins escape digestion in the small intestine. This estimation fits rather well if we consider the mean protein consumption in geographical areas like European countries and the USA, which averages 85 g/day in adults [34,35], and the mean dietary protein digestibility in the small intestine with an average value of 90%. Indeed, from these data, we can calculate that an average value of 8.5 g of alimentary proteins would be recovered within the large intestine, a value in the 3-11 g range as calculated above [36].
3. Metabolism of Proteins by the Large Intestine Bacteria
3.1. Degradation of Proteins by the Bacterial Proteases and Peptidases and Transport of Peptides and Amino Acids in Bacteria
In the large intestine, the bacterial proteases and peptidases release amino acids from the luminal proteins [37]. Bacteria are equipped with a highly diverse set of proteases present in many common gut bacterial species such as Clostridium spp., Bacteroides spp., Lactobacillus spp. and many others. In fact, hundreds of different proteases have been identified in these bacterial species [38]. Some bacteria, such as lactic acid bacteria, have developed proteolytic systems which compensate for their limited or absent capacities to synthesize several amino acids [39]. Proteolytic systems in lactic acid bacteria gather both extracellular and membrane bound proteases (notably PrtP and CEP) that convert proteins into oligopeptides, and these processes are followed by importation into the bacterial cells via peptide transporters such as Opp, Dpp, and Dtp for oligopeptides, dipeptides, and di- and tripeptides, respectively. Finally, numerous intracellular peptidases in bacteria degrade the oligopeptides into shorter oligopeptides and amino acids in free form [40].
Amino acids and their derivatives can also be both imported and exported from the bacterial cells via transmembrane proteins, including the ATP-dependent ABC transporters, different families of channel proteins, and secondary carriers relying on the proton-motive force, ionic sodium motive force, and solute-solute exchange [41,42,43]. These amino acid and oligopeptide transporters have been notably studied in Escherichia coli [44]. The uptake of amino acids by the large intestine bacteria requires a significant fraction of the available metabolic energy, and as shown in lactic acid bacteria, the metabolic energy cost of amino acid uptake can be reduced by accumulating oligopeptides instead of the individual amino acids, followed by proton-motive-generating efflux of amino acids accumulated in excess in bacteria compared to metabolic needs [45]. The export of amino acids from intestinal bacterial cells into the luminal fluid may represent a way to supply amino acids to other bacterial species, thus representing a process of metabolic cooperation between different bacterial species.
The metabolic fate of amino acids present in free form in the large intestine luminal fluid has been studied. Although some measurable transfer of amino acids across the rodent colonic mucosa has been measured in vivo [46], amino acids, in sharp contrast with the situation prevailing in the small intestine, are not believed to be absorbed to any significant extent by the mammalian colonic epithelium [47,48,49], and if this proposition is correct, this means that amino acids entering the large intestine are almost lost for utilization by the host for protein synthesis and for utilization in other metabolic pathways [50].
However, some experimental arguments suggest that some amino acid absorption through the mammalian large intestine epithelium cannot be totally excluded [51]. Firstly, in the pig model, infusion of proteins or amino acids into the large intestine lumen indicate whole body N balance improvement which may suggest some amino acid absorption [52]. Furthermore, some absorption of amino acids from microbial origin through the pig colon has been suggested based on the appearance of 15N-labeled amino acids in the venous blood after infusion of 15N-labeled bacteria into the cecum [53]. In addition, some biochemical data indicate the presence of the ATBo+ neutral and cationic amino acid transporter in the colon [54], these transporters being localized at the apical pole of colonocytes [55]. Another neutral and cationic amino acid transporter, namely Bo+ system is expressed on the apical surface of colonic absorptive cells [56]. The hypothesis that some amino acids available within the large intestine luminal fluid would be used for local protein synthesis within the colonic epithelial cells is a possibility that has been little evaluated [57]. In the pig model, during the neonatal period, some amino acid absorption by the colon epithelium has been measured [58,59], suggesting a transient colonic absorption of amino acids in this experimental model.
Finally, it appears from the available data that amino acids within the colon large intestine luminal fluid in adults are kept available, presumably for their most part, for the metabolic activity of the microbiota in relationship with their growth and physiology as will be detailed in the following chapters of the present review.
3.2. The Indispensable Amino Acids for Intestinal Bacterial Species
The definition of indispensable amino acids in mammals (including humans) is related to the fact that among the amino acids required for metabolism and physiological functions in the tissues and organs, nine of them (namely isoleucine, leucine, valine, lysine, methionine, phenylalanine, threonine, tryptophan, and histidine) must be obligatory provided by the diet to meet the body’s requirement [60,61]. In other words, indispensable amino acids are those which cannot be synthesized in the body, or cannot be synthesized to any significant extent, when compared to the metabolic and physiological needs.
By analogy with the definition of indispensable amino acids in animals, it is tempting to propose the use of the same terminology with some nuances for the intestinal bacteria. Indeed, not all bacteria within the large intestine are able to synthesize the 20 usual amino acids and the two unusual amino acids (namely selenocysteine and pyrrolysine) they need for protein synthesis and for utilization in other metabolic pathways which are central for bacterial growth and physiology [23]. The amino acids which cannot be synthesized by the intestinal bacteria or cannot be synthesized in significant amount to cover the requirement must be supplied from the luminal fluid and imported within the bacterial cells.
Then, by deduction, the non-indispensable (or dispensable) amino acids for the large intestine bacteria are those which can be synthesized in sufficient amount to cover their metabolic and physiological requirements. The studies of amino acid biosynthesis in intestinal bacteria have been historically focused on few bacterial species including notably Escherichia coli, including the enterohemorrhagic E. coli (EHEC) which is responsible for severe colonic infection [62], Salmonella typhurium, a pathogen known to provoke diarrhea and inflammation in ileum and colon [63], and Bacillus subtilis, which is present in the human gut, and which have shown some protective effects in Citrobacter rodentium-associated colitis [64]. Obviously, the available data related to these few bacterial species cannot be considered as representative of amino acid anabolism for all the bacterial species present within the large intestine. With this reservation in mind, many of the metabolic pathways at the basis of amino acid metabolism are relatively well conserved across the bacterial lineages. The description of the metabolic pathways involved in the biosynthesis of the 22 amino acids in bacteria is outside the scope of this review but can be found in recent reviews dedicated to this topic [23,38].
The diversity of the metabolic capacities for the different bacteria living in gut can be illustrated by some typical examples. Clostridium perfringens, a common human enteropathogen [65] lacks for instance one or several genes involved in glutamate, arginine, histidine, lysine, methionine, serine, threonine, aromatic, and branched-chain amino acid biosynthesis [66], thus depending on the presence of these amino acids in the surrounding media for all the biochemical pathways involving amino acids, including protein synthesis. Then, these amino acids can be considered as indispensable for Clostridium perfringens.
Regarding Lactobacillus johnsonii, a human gut commensal [67], this bacterium also appears unable to synthesize almost all amino acids, due to the lack of complete biosynthetic pathways, thus exhibiting dependence on most amino acids either in free form or supplied in peptidic forms in the media [68]. Other bacteria, like Campylobacter jejuni, a foodborne pathogen which causes inflammation and enteritis in humans [69] and Enterococcus faecalis strains, which are resident intestinal bacteria associated with invasive infections and inflammatory bowel diseases [70], do not contain the whole biosynthetic pathways for some amino acids [71].
Here, it is important to recall that the sole presence of genes within a bacterial genome is not sufficient to establish their functionality [38]. For instance, the genes for all the 20 common amino acids have been identified in the bacterium Lactococcus lactis, a noninvasive and nonpathogenic organism [72], but despite this result, several amino acids, namely isoleucine, valine, leucine, histidine, methionine, and glutamate, need to be supplied to this bacterium for growth. This is because several genes coding for several enzymes involved in the biosynthetic pathways of these amino acids are nonfunctional due to point mutations [73,74]. This is also the case for Staphylococcus aureus, a common colonizer of the human gut [75], since two strains exhibit auxotrophy for arginine, valine, leucine, cysteine, and proline, despite the presence of the complete sets of genes corresponding to the different biochemical pathways responsible for the biosynthesis of these amino acids in the two strains studied [76]. Thus, for these bacteria, the five amino acids listed above can be considered as indispensable.
3.3. Utilization of Amino Acid for Macromolecule Synthesis in Bacteria
Amino acids in bacteria are well known to be utilized for the synthesis of macromolecules including not only proteins, but also for RNA and DNA synthesis (the main pathways for the metabolism of amino acids by the intestinal bacteria are summarized in Figure 1). To face changing environment, bacteria depend on strictly coordinated proteostasis networks that finally control processes such as the rates of protein synthesis and degradation [77]. One critical mechanism that is involved in the response of bacteria to changing environment (including changes in the concentrations of nutrient in the environment), is represented by the control of the expression of genes at the level of proteosynthesis. At each of the three major steps of translation, namely initiation, elongation, and termination, bacterial cells can tune the translation rate and thus the intracellular protein concentrations depending on their environment [78]. Of note, protein synthesis and protein turnover are dependent on post-translational modifications of a group of bacterial proteins [79].
Regarding RNA and DNA synthesis, three amino acids, namely glycine, aspartate, and glutamine, are used as precursors for the synthesis of the purine and pyrimidine rings. The metabolic pathways involved in pyrimidine biosynthesis from amino acids has been for instance studied in numerous bacterial species, and the results have shown that the synthesis of pyrimidines requires carbamylphosphate as a precursor. In Escherichia coli, Salmonella typhimurium, and other bacterial species including Pseudomonas aeruginosa (present in the human intestinal tract [80]) and Proteus mirabilis (which is associated with inflammation in Crohn’s disease [81]), carbamylphosphate is synthesized from glutamine and bicarbonate in an ATP-dependent reaction. In the first step of pyrimidine synthesis, carbamylphosphate and aspartate are condensed to form carbamylaspartate. This latter metabolite is then the precursor for uridine triphosphate and cytidine triphosphate in six and seven steps, respectively [82]. Interestingly, the pool of intracellular nucleotides in bacteria ultimately regulate protein synthesis at various stages of this process [83], showing interrelationships between the different metabolic pathways.
3.4. Utilization of Amino Acids for ATP Synthesis in Bacteria
Under aerobic conditions, some bacterial species can convert amino acids to their α-ketoacids counterparts via transamination and deamination reactions. These α-ketoacids are then oxidized in the citric acid cycle. In addition, some bacteria are equipped with a branched-chain keto acid dehydrogenase complex that yields ATP from the oxidized form of the branched-chain amino acids [84]. However, under strict anaerobic conditions, such as the one which prevails in the colon, or in the absence of suitable electron acceptors, strict or facultative anaerobic bacteria such as Clostridia and Fusobacteria can utilize amino acids as energy sources, thus fermenting amino acids in a wide range of reactions including transamination, oxidation, and reduction [85,86]. Thus, in the absence of oxygen and of other inorganic electron acceptors, the substrates for fermentation are used both as electron donors and as electron acceptors, thus resulting in low yield ATP production when compared to the yield of ATP production under aerobic conditions [87]. Many electron donors and acceptors can participate in the different biochemical pathways involved in protein fermentation, including α-ketoacids, molecular hydrogen (H2), as well as amino acids [88,89]. Bacterial amino acid fermentation results in the production of metabolic end products, including mainly short- and branched-chain fatty acids [84].
In addition, bacteria such as Lactic acid bacteria can use amino acid decarboxylation to generate ATP [90,91]. Amino acid decarboxylation presumably supports ATP production in condition of adverse environmental conditions. For instance, tyrosine decarboxylation in bacterial species equipped with this metabolic pathway results in the production of tyramine and carbon dioxide (CO2), and allows the production of ATP by a proton motive force composed of a pH gradient formed during proton consumption in the decarboxylation reaction, and by membrane potential, which itself results from electrogenic transport of tyrosine in exchange for tyramine [92].
In Clostridium species, including Clostridium difficile, specific metabolic pathways grouped under the name of Stickland metabolism is central for ATP production. Stickland reactions involves the coupled oxidation and reduction of two amino acids, with one amino acid acting as an electron donor, while another amino acid acts as an electron acceptor, then facilitating the production of ATP by substrate-level phosphorylation, and by maintaining the NADH/NAD+ pool [93]. The most efficient electron donors include amino acids such as alanine, valine, leucine, and isoleucine, while the most efficient electron acceptors include proline, glycine, and leucine [94]. Ornithine, a non-proteogenic amino acid can undergo Stickland metabolism to yield acetyl CoA, ammonia, and alanine, or can be alternatively converted to proline [95], thus participating in the production of ATP in the bacterial species equipped with these metabolic capacities. Stickland amino acid reactions are believed to provide metabolic resources to support bacterial growth when other substrates, such as carbohydrates, are in short supply [96].
Of note, amino acid utilization for ATP production may be operative in a preferential manner when compared to other ATP-producing substrates. For instance, Clostridium sticklandii, a non-pathogenic proteolytic clostridium, which is known to be dependent on amino acids for growth, is considered as a specialist for amino acid degradation. This bacterium has been studied in detail for its metabolic capacity to degrade amino acids thus allowing ATP production [97]. Clostridium sticklandii is characterized by numerous transporters for oligopeptides and amino acids, and with a battery of proteolytic enzymatic activities. This bacterium preferentially utilizes the amino acids arginine, threonine, cysteine, serine, and glycine for ATP production.
Interestingly, several amino acids are used by some bacterial species in specific conditions of growth. For instance, lysine degradation, which ends with the synthesis of butyrate, represents a major source of ATP for Clostridium sticklandii only in stationary growth phase.
3.5. Utilization of Amino Acids for the Synthesis of Bioactive Metabolites in Bacteria and Effects of These Compounds on Bacterial Growth and Physiology
Numerous amino acid-derived metabolites produced by different intestinal commensal and pathogenic bacteria have been shown to be active on their own metabolism and physiology, with associated effects on their virulence and growth capacities [98]. In addition, from recent studies, emerging roles of several amino acid-derived metabolites released in the surrounding media by some bacterial species have been presumed to act on other bacteria species, thus suggesting that amino acid bacterial metabolism may be involved in the regulation of bacterial physiology and likely as a means of communication between bacteria in different ecosystems.
Amino acid-derived polyamines facilitate bacterial growth, modulate bacterial virulence, promote bacterial acid resistance, and regulate biofilm formation
Polyamines are small aliphatic amines that are produced by bacteria primarily from amino acids. The main polyamines synthesized by bacteria are putrescine, spermidine, agmatine and cadaverine, but numerous polyamine derivatives are produced from these polyamines, and for several of them, by pathways that are not operative in eukaryotic cells [99,100]. Putrescine and spermidine are generally the most common polyamines found in bacteria, while agmatine and cadaverine are found at lower concentrations and spermine is rarely detected in most bacterial species [101]. Polyamines have been measured in the intestinal contents at concentrations from micro- to millimolar concentrations [1]. A part of polyamines within bacteria is associated with RNA, and such association is paralleled by effects of polyamines on transcription and translation [102,103,104]. The precursors for putrescine and spermidine synthesis in bacteria are the amino acids ornithine, arginine, and methionine, while agmatine is produced from arginine. Regarding cadaverine, this polyamine is produced from lysine [101]. The conversion of putrescine into spermidine requires the decarboxylated form of the methyl donor S-adenosylmethionine (Figure 2).
However, not all bacteria are equipped with the metabolic machinery allowing polyamine biosynthesis from amino acid precursors, thus indicating that these bacterial species may be dependent on the import of the polyamines present in the media [105]. These polyamines are likely originating from the polyamines synthesized and released by other members of the bacterial community. In fact, systems of polyamine uptake and release have been identified in numerous bacterial species. For instance, agmatine-putrescine antiporters have been characterized in Enterococcus faecalis, and such antiporters is used for importing agmatine and exporting putrescine [106]. Polyamine degradation pathways represent another important player in fixing the polyamine concentrations within the bacteria intracellular medium [107]. Thus, polyamine concentrations in bacteria are the net result of endogenous synthesis, import, export, and degradation of polyamines.
Polyamines participate in critical physiological functions in intestinal bacteria. The effects of polyamines on bacterial growth are rather heterogeneous depending on the species studied. For instance, Escherichia coli rendered deficient for polyamine synthesis can still grow, albeit at reduced rate, when compared with the wild-type counterpart, thus indicating that polyamines are not strictly required for growth of this bacterium [108]. Likewise, in the foodborne pathogen Salmonella enterica, polyamines depletion reduces but do not suppress bacterial cell growth [99]. In contrast, polyamine biosynthesis is essential for the growth of the pathogen Pseudomonas aeruginosa and of the foodborne pathogen Campylobacter jejuni [109,110].
Spermidine can modulate the production of bacterial toxins. Indeed, this polyamine reinforces for instance the production of colibactin [111]. Regarding cadaverine, this polyamine can attenuate virulence and reduce enterotoxin activity in Shigella spp. (Maurelli PNAS 1998). In addition, putrescine, cadaverine, spermidine, and spermine have been shown to modulate the virulence of Vibrio cholerae [112].
Polyamines can be incorporated into bioactive compounds, notably into bacterial siderophores. These compounds are secreted by bacteria and then scavenge iron in the extracellular medium; this latter compound being then supplied to bacteria via specific receptors [113]. Spermidine and cadaverine can be linked covalently to peptidoglycans, and this process presumably increase bacterial cell wall rigidity [114]. Cadaverine has been demonstrated to provide to Escherichia coli and to the foodborne pathogen Vibrio parahaemolyticus a mechanism involved in acid resistance [115,116].
Lastly, polyamines are involved in the regulation of biofilm formation. Briefly, biofilms can be viewed as a mixture of high-density bacterial (and archaeal) community contained within a self-producing protective matrix made of polysaccharides, proteins, nucleic acids, and lipids [117,118]. Such structures represent an important element for the modulation of bacterial growth in changing environment, like the situation which may happen in the large intestine [119]. Biofilms are found in intestine where elements of the intestinal microbiota contained within these structures are in close contact with the intestinal mucosal surface [120]. Research on biofilms is motivated by the fact that microorganisms (including the intestinal pathogens) in biofilms become much more resistant to antimicrobial treatment [121]. Biofilm development and quorum sensing have been shown to be interconnected [122,123]. In a few words, quorum sensing enables bacteria to collectively modify their behavior in response to changes in the cell density and species composition within the local environment [124]. The implication of different polyamines in the regulation of biofilm formation, either positive or negative, has been recently documented. The polyamine agmatine is involved in the process of biofilm formation in Bacillus subtilis [125]. Another polyamine, namely the uncommon polyamine norspermidine induces biofilm formation in association with increased cyclic di-GMP synthesis, this latter compound being able by itself to activate in Vibrio cholerae the expression of genes involved in the formation of biofilm components [126]. In an intriguing and interesting way, norspermine appears to act on Vibrio cholerae via a norspermidine sensor [127]. It has been confirmed that norspermidine biosynthesis is required for biofilm formation by Vibrio cholerae [128]. In contrast, spermine inhibits Vibrio cholerae biofilm formation [129]. The results presented above indicate that different polyamines can exert opposite effects on the bacterial capacity to form biofilms. The respective roles of the different polyamines (either synthesized endogenously by bacteria or taken up from the extracellular media) for the regulation of biofilm formation by different bacterial species among the intestinal microbiota remain unclear, thus requiring additional experimental works.
Cysteine-derived hydrogen sulfide increases bacterial respiration and growth, promotes biofilm formation, and modulates the effects of different antibiotics
Hydrogen sulfide (H2S) is produced in bacteria from different S-containing substrates including notably cysteine and inorganic sulfate [130]. Cysteine-degrading bacteria in gut include Fusobacterium, Clostridium, Escherichia coli, Salmonella, Klebsiella, Streptococcus, and Enterobacter which convert cysteine to H2S through the catalytic activity of cysteine desulfhydrase [131,132,133]. Sulfate-reducing bacteria include the following gut bacteria: Desulfobrio, Desulfobacter, Desulfobulbus, and Desulfomaculum [134,135,136]. Desulfovibrio represents the dominant genera of sulfate-reducing bacteria in intestine [137,138,139]. The amount of sulfate in the diet, as well as the capacity of the small intestine to absorb this compound appear to represent the main parameters which determine the amount of sulfate being transferred to the large intestine [140]. In volunteers, the ingestion of sulfate in supplement increases the fecal sulfide production rate [141]. It appears that overall, current concentrations of sulfate in the large intestine are adequate to support the growth of sulfate-reducing bacteria.
H2S in the colonic luminal fluid can exist in three forms: the H2S gas that is partly dissolved in the aqueous phase and represents a highly diffusible compound, hydrosulfide anion (HS-)and sulfide ion (S2-), this latter compound being present at a negligible level. Indeed, in the aqueous phase, H2S dissociates into HS- and S2- and H+ with pKa values being 7.04 and 11.96, respectively [142,143]. In healthy subjects, the pH at the colonic mucosal surface ranges between 7.2 and 7.5 in the descending colon and the rectum [119]. Thus, by considering a pH equal to 7.4, approximately one third of sulfide is in the form of H2S at equilibrium, while two-third are in the form of hydrosulfide anion. However, if the pH becomes more acidic, the H2S/HS- ratio increases in the large intestine. The concentrations of total H2S measured in the colonic luminal fluid are rather divergent according to the technics used, ranging from high micromolar to low millimolar concentrations [1]. Measurement in the feces obtained from human volunteers, indicate that 8% of sulfide is in the unbound form [144]. The identification of the compounds which bind sulfide in the colonic lumen, although obviously not exhaustive, points out several compounds from dietary origin that are not fully absorbed in the small intestine. These compounds include zinc [145], heme [146], and polyphenols [147].
Incidentally, H2S is involved as a precursor for the synthesis of cysteine and methionine in bacteria, and then, such anabolic pathways in gut bacterial species may influence sulfide concentration within the intestinal content. In bacteria, the synthesis of cysteine is made from serine in two steps, with the first step being catalysed by serine acetyltransferase in the presence of the co-substrate acetyl CoA, and the second step is then catalysed by cysteine synthase using H2S as co-substrate [148,149]. Not all bacteria are equipped with cysteine synthase, since for instance, Bifidobacterium longum, which is one of the most abundant bacteria in the infant and adult intestine [150], lacks the corresponding gene and may use alternative pathways involving succinylhomoserine and H2S as substrates for cysteine synthesis [151]. Regarding methionine synthesis in bacteria, this amino acid is produced through metabolic pathways which share the first three steps involved in threonine synthesis up to the synthesis of homoserine. Then homoserine is converted to methionine by several different pathways which are operative in different bacterial species. These pathways allow the conversion of O-succinylhomoserine and cysteine to cystathionine or convert O-acetylhomoserine (synthesized from aspartate in four steps) and H2S (or cysteine) to cystathionine. Cystathionine is then converted to homocysteine, the immediate precursor of methionine in the reaction catalysed by methionine synthase [152,153] (Figure 3).
H2S, at low concentration stimulates the respiration of the intestinal pathogen Mycobacterium tuberculosis and promotes its growth [154]. In contrast, H2S at excessive concentrations inhibits the terminal oxidase of the respiratory chain of Escherichia coli [155], such terminal oxidase representing one element of the supramolecular organization of the bacterial respiratory chain [156]. However, Escherichia coli is equipped with alternative bd-type oxidase, which is insensitive to the inhibitory effects of H2S, and the presence of this isoenzyme allows respiration and growth of this bacterium in H2S-rich environment [157]. Similarly, cyanide-insensitive oxidase in the intestinal pathogen Pseudomonas aeruginosa has been shown to confer tolerance to H2S regarding its respiration [158]. Thus, it appears that alternative solution for respiration exist in some intestinal bacteria to cope with increased sulfide luminal concentration.
H2S can also act on some intestinal bacterial species as a protective compound. In the pioneering work of Bachenheimer and Bennett, H2S produced by Desulfovibrio desulfuricans was shown to be the diffusible factor responsible for the protection of Pseudomonas aruginosa from heavy metal toxicity, such as mercury [159]. Incidentally, these important results published more than 6 decades ago indicated that a given bacterial species was able to produce a compound which was active towards another bacterial species. Similarly, H2S produced by Escherichia coli contributes to the protection of Staphylococcus aureus against mercuric toxicity [160]. Since these discoveries, but much more recently, in the context of the study of bacterial antibiotic resistance, H2S has emerged as a protective compound for Pseudomonas aeruginosa and Staphylococcus aureus against the action of different antibiotics [161,162]. Although the precise mechanisms of action involved in the protective action of H2S against the effects of antibiotics are not yet known, several experimental results have begun to shed light on these mechanisms. In fact, sequestration of Fe2+ ions by H2S counteracts oxidative stress triggered by some antibiotics in Escherichia coli [163]. Furthermore, H2S has been demonstrated to be involved in the maintenance of the bacterial redox homeostasis and to protect pathogenic Escherichia coli strains against the oxidative stress triggered by the antibiotic ampicillin [164]. Also of note, cystathionine-Ɣ-lyase has been discovered as the primary enzymatic activity which is responsible for H2S production in Staphylococcus aureus and Pseudomonas aeruginosa, and inhibition of this catalytic activity potentiates the efficiency of antibiotics against both bacterial species in invitro and in vivo models of infection [165]. These results suggest that endogenously formed H2S may participate in the resistance of some bacteria to the effects of some antibiotics.
However, H2S is apparently not a bacterial amino acid which limits the efficiency of antibiotics against all bacteria present within the intestine. For instance, in Acinetobacter baumannii, which incidentally is not an H2S-producer, H2S reinforces the effects of several classes of antibiotics [166]. Thus, the H2S-mediated protection or conversely sensitization to the effects of different antibiotics depends on the bacterial species considered, and likely on the antibiotics used. Finally, in another context, the implication of H2S in the resistance to infection by pathogenic bacteria has been suggested [167]. In this latter study, the endogenous production of sulfide is apparently involved in the enhanced capacity of intestinal commensal bacteria to counteract pathogen infection.
Regarding the establishment and restoration of colonic microbiota biofilms, H2S has been shown to intervene in such processes [168]. Fusobacterium nucleatum is an H2S producer, and the production of this metabolite in this bacterial species modulates the virulence and susceptibility to antibiotics [169]. Scavenging of H2S in several bacterial species (including Escherichia coli) which produce this compound potentiates both the bactericidal effects of several active compounds and disrupts the formation of the bacterial biofilms, thus indicating that the endogenous synthesis of H2S by these species represent one component involved in biofilm formation and presumably the associated resistance to bactericidal agents [170].
Arginine-derived nitric oxide inhibits bacterial energy metabolism and growth, and increases the dispersal of biofilm
Nitric oxide (NO) is produced from the amino acid arginine by nitric oxide synthases found in numerous bacteria [171], including several bacteria found in intestine such as Bacillus subtilis and Lactobacillus fermentum [172,173]. Nitric oxide, as documented for hydrogen sulfide, is a highly diffusible gaseous compound. Incidentally, it is worth noting that the nitric oxide synthase pathway is not the exclusive way of nitric oxide production in intestinal bacteria. In fact, intestinal bacteria such as Campylobacter jejuni and Pseudomonas aeruginosa can produce NO in the process of nitrite (NO2-) reduction catalyzed by nitrite reductase [174,175,176]. NO can also be formed by ammonia oxidizing bacteria, such as bacteria belonging to the Nitrospira genus [177], some of them being found in the human intestine [178]. The concentration of NO in the gas phase of the colon has been measured in volunteers and average approximately 20 p.p.m. [179].
NO has been shown to be involved in the control of bacterial growth and in the biosynthesis of several secondary metabolites. Indeed, NO exerts bacteriostatic effects against several intestinal bacteria such as Salmonella enterica [180]. NO appears to exert its bacteriostatic action at least partially by inhibiting enzymatic activities involved in bacterial energy metabolism [181]. Apart from these effects on bacterial growth, NO has been involved for the synthesis of secondary metabolite in bacteria. For instance, NO is used as a building block for the synthesis of the 1,2,3 triazole moiety of 8-azoguanine [182], a member of the purine and pyrimidine antimetabolites, but it remains unknown if 8-azoguanine is effectively produced by any bacterial species belonging to the intestinal microbiota. NO intervenes in nitration reactions within bacteria [183]. In fact, rufomycin which is produced by bacteria in a NO-dependent nitration step targets proteolysis in Mycobacterium tuberculosis [184].
Lastly, NO allows the rapid dispersal of biofilm macrostructure by mechanisms involving NO-sensory proteins in many pathogenic bacteria including the intestinal pathogen Vibrio cholerae [185]. Accordingly, several NO donors have been tested recently on different bacterial species, and the results of these experiments show antimicrobial and antibiofilm effects of these compounds, notably on Fusobacterium nucleatum [186]. In the bacterial species Vibrio parahaemolyticus which may act as an intestinal pathogen, NO promotes the transcription of the quorum sensing regulatory gene opaR [187]. Although NO emerges as a gasotransmitter which is active on the regulation of biofilm formation, further works are required to test if this compound produced endogenously by intestinal bacteria reaches concentrations within the luminal fluid which are compatible with this effect.
Tryptophan-derived indole diminishes bacterial growth and bacterial virulence
Numerous Gram-positive and Gram-negative bacterial species can produce indole from the precursor tryptophan, notably the intestinal bacterial species Escherichia coli, Proteus vulgaris, Clostridium spp., and Bacteroides spp. [188,189,190]. The indole concentration has been measured in the feces from volunteers, thus reflecting the concentration of indole in the most distal segment of the large intestine, and has been found to range from 0.3 to 6.6 millimolar [1]. Indole diminishes the capacities of bacterial cells for motility and aggregation in Listeria monocytogenes; a bacterial species occasionally found within the intestine [191]. Indole is also able to diminish the virulence of intestinal bacterial species such as Pseudomonas aeruginosa and Salmonella enterica [192,193].
Incidentally, but importantly, indole is also active in diminishing virulence and growth of the fungal species Candida albicans [194], a microrganism commonly found in intestine [195]. This discovery reveals a new way of communication between intestinal bacteria and fungi. Lactic acid bacteria are also sensitive to the effect of indole since this amino acid-derived metabolite exerts bacteriostatic effect on these bacteria [196]. Lastly, indole may affect the bacterial metabolism since indole affects toxin production by Klebsiella oxytoca [197], a normal resident in the intestine which may become pathogenic according to the environmental context [198].
Tyrosine-derived p-cresol produced by Clostridium difficile gives competitive advantage to this bacterium over other bacteria
The bacterial metabolite p-cresol (4-methylphenol) is produced from the amino acid tyrosine by anaerobic bacteria, notably the ones which live in the large intestine luminal fluid [199]. Among these numerous bacteria, specific families of bacteria like Fusobacteriaceae, Enterobacteriaceae, Clostridium, and Coriobacteriaceae are active p-cresol producers [200,201]. The p-cresol concentrations measured in the human colonic contents are in the low millimolar range [1]. Of major interest, the capacity of Clostridium difficile to produce p-cresol is one important element which gives to this bacterial species a competitive advantage over other gut bacteria such as Escherichia coli, Klebsiella oxytoca, and Bacteroides thetaiotaomicron [202]. Clostridium difficile is well known to represent a major cause of intestinal infection and diarrhea in patients following treatment with antibiotics ([203]. Using a mouse model of Clostridium difficile infection, it has been observed that excessive p-cresol production affects the gut microbiota diversity. Also of note, by removing the capacity of Clostridium difficile to produce p-cresol, this bacterium was less able to recolonize the intestine after an initial episode of infection [202]. As expected, Clostridium difficile can tolerate p-cresol concentrations as high as 10 millimolar [204,205].
Tyrosine-derived skatole inhibits biofilm formation by enterohemorrhagic Escherichia coli
Skatole (3-methylindole) is a metabolite produced by bacteria from tryptophan [206,207]. Lactobacillus, Clostridium, and Bacteroides are known skatole producers [208]. Fecal skatole concentrations in healthy individuals are usually relatively low, averaging approximately 40 micromolar [1]. Skatole display a robust capacity to inhibit biofilm formation by the enterohemorrhagic Escherichia coli [209].
Glycine-derived betaine protects bacteria against osmotic stress
Although glycine betaine metabolism has been principally studied in bacteria which are not found in intestine, these studies indicate that this compound is derived from extracellular sources and/or from endogenous synthesis from glycine [210,211]. Glycine betaine is used by bacteria for protection against osmotic stress [212]. Notably, glycine betaine is an osmoprotectant in the intestinal bacteria Pseudomonas aeruginosa [213]. Such capacity may be relevant in case of increased luminal osmolarity within the colonic fluid [119].
Amino acid-derived glutathione is active against oxidative stress
Glutathione is synthesized from the three amino acids glutamate, cysteine, and glycine by numerous Gram-negative bacteria but only in few Gram-positive bacteria such as Listeria monocytogenes. Glutathione plays a key role in maintaining the proper oxidation state of protein thiols in bacteria and is involved in the response of bacteria to environment creating a situation of oxidative stress [214,215].
Histidine-derived histamine helps bacteria to survive in acidic media
Histamine, mainly known as a biogenic amine involved in the allergic disease, has been demonstrated to act on some bacteria. Production of histamine from the simple decarboxylation of the amino acid histidine has been demonstrated in numerous Gram-positive and Gram-negative bacterial strains [216], notably in bacteria belonging to the intestinal microbiota such as Pediococcus parvulus, Morganella morganii, Klebsiella pneumoniae, Enterobacter spp., Citrobacter freundii, and Hafnia alvei [217]. Recent studies show that histamine can improve the survival of bacteria in acidic condition. Generation of proton motive force by histidine decarboxylation has been demonstrated in Lactobacillus buchneri [218]. Furthermore, it has been elegantly shown that by expressing the functional histidine decarboxylase pathway in Lactococcus lactis, these bacteria survive longer in acidic medium (pH 3.0) than the control cells counterpart [219]. However, since the pH in the human large intestine content is only slightly acidic or near neutrality, ranging from 5.7 to 6.8 in cecum, while ranging from 6.1 to 7.5 in the descending colon and rectum [220], it remains to be demonstrated if such histamine-dependent capacity of bacteria to survive in acidic medium is relevant for the intestinal bacteria in this range of pH.
Amino acid-derived dopamine, serotonin, noradrenaline, and gamma-amino butyric acid are involved in the regulation of bacterial growth, virulence, and bacterial acid tolerance
Dopamine, primarily known as a neurotransmitter in animals, has been shown to be active on some bacteria. Dopamine is produced by many bacterial species present in the intestine such as Bacillus subtilis, Escherichia coli, Staphylococcus aureus, Proteus vulgaris, and Klebsiella pneumoniae [221]. Bacterial tyrosinases are the enzymatic activities which catalyse in bacteria the conversion of tyrosine to dihydroxyphenylalanine (DOPA), the direct precursor of dopamine [222]. Dopamine concentration has been measured in the mouse large intestine luminal content, averaging approximately one micromolar [223], but to the best of our knowledge, the concentration of dopamine in the human colonic fluid has not been determined. Dopamine accelerates the growth of Pseudomonas aeruginosa and Klebsiella pneumoniae [224]. Furthermore, dopamine acts as a siderophore-like iron chelator that promotes the growth of Salmonella enterica [225].
Serotonin (5-hydroxytryptamine), a compound which is well known to be a neurotransmitter in animals, is produced from the amino acid tryptophan is many intestinal bacterial species among which Propionibacterium, Lactobacillus, Lactococcus, Bifidobacterium, Streptococcus, Bacteroides, and Escherichia coli [217]. Serotonin appears to regulate either positively or negatively, depending on the bacterial species considered, the virulence of different bacteria. Serotonin enhances the virulence of Pseudomonas aeruginosa in in vitro and in vivo experiments; the in vivo experiments being performed using a model of rodent infection [226]. In this latter study, serotonin was found to regulate bacterial quorum sensing. As introduced above, quorum sensing is the process by which bacteria communicate, allowing groups of bacteria to synchronously modify the bacterial population density and composition in vicinal community [123].
Noradrenaline (norepinephrine) is produced from the amino acid tyrosine by several intestinal bacteria including Bacillus subtilis, Escherichia coli, and Proteus vulgaris [217]. This compound, known as neurotransmitter in animals, affects bacterial growth either positively or negatively, depending on the bacterial species studied. Noradrenaline is notably active on the growth of the following intestinal anaerobic bacteria: Klebsiella pneumoniae, Fusobacterium nucleatum, Pseudomonas aeruginosa, Enterobacter clocae, Shigella sonnei, and Staphylococcus aureus. In addition to its effect on bacterial growth, noradrenaline increases the virulence of several anaerobic bacteria such as Clostridium perfringens [227,228,229].
Gaba-amino butyric acid (known as GABA) is produced from the amino acid glutamate by several bacterial species present in the intestinal luminal fluid such as Lactobacillus and Bifidobacterium [230,231,232]. GABA, known as the main inhibitory neurotransmitter in the mammalian brain, is involved in tolerance of bacteria such as Bacteroides spp. to acidic media through the maintenance of the intracellular pH [233,234].
Tyrosine-derived 4-hydroxyphenylacetate inhibits bacterial growth and virulence
The bacterial metabolite 4-hydroxyphenylacetate is a metabolic intermediate which is produced from the amino acid tyrosine by intestinal bacterial species that synthesize phenol and p-cresol as metabolic end products [200]. The transport of 4-hydroxyphenylacetate from the extracellular media has been studied in Escherichia coli [235], thus raising the hypothesis that this compound would be released by some intestinal bacterial species and taken up by others. This compound inhibits the growth of the foodborne pathogen Listeria monocytogenes, an effect which is associated with alteration of the bacterial morphology and with a decrease of the expression of several genes involved in bacterial virulence [236].
Amino acid (and other substrate)-derived organic acids serve as energy substrates in bacteria and modulate their growth and virulence
During the catabolism of amino acids within bacteria, several organic acids, including succinate, oxaloacetate, formate, and lactate, are intermediary or terminal metabolites which may be produced in significant amounts [15,237]. These compounds are obviously not exclusively produced from amino acids but can originate from other substrates like carbohydrates [238,239].
Succinate oxidation by Escherichia coli was described more than seven decades ago [240]. Since then, several studies on that topic were performed, including a study showing capacity of Bacillus lactis to use succinate for respiration [241]. Succinate can be used by bacteria for ATP and reduced equivalent generation. Succinate is involved in the mycobacteria respiration, serving both as an intermediate in the tricarboxylic acid cycle and as an electron donor for the respiratory chain [242]. Of note, impaired succinate oxidation in the intestinal bacterium Mycobacterium tuberculosis prevents its growth, thus pointing out a central role of the oxidation of this metabolite for M. tuberculosis proliferation. In this latter bacterium, succinate dehydrogenase, which converts succinate to fumarate, acts as a central regulator of respiration [243]. Succinate produced by the gut microbiota promotes infection by the opportunistic intestinal pathogen Clostridium difficile [244], and extracellular succinate induces biofilm formation by this pathogen [245]. Also of major interest, Clostridium butyricum, a butyrate-producing bacterium found in the human gut, can prevent the proliferation of C. difficile by diminishing the succinate concentration within the large intestine luminal content [246]. Such decreased succinate concentration represents apparently the net result of the overall metabolic activity of the intestinal microbiota, suggesting subtle metabolic communication between intestinal bacteria.
The tricarboxylic acid cycle intermediate oxaloacetate, when produced by Escherichia coli, improves the survival of the amoeba parasite Entamoeba histolytica in the large intestine luminal fluid [247], thus suggesting bacterial-parasite communication between microorganisms in the large intestine. Entamoeba histolytica is the causative agent of human amoebiasis, an enteropathy affecting millions of humans worldwide [248].
Regarding formate, this organic acid is involved in energy metabolism in different bacterial species. Formate enhances respiration in the intestinal bacterial species Campylobacter jejuni [249]. However, at excessive concentrations, formate can reduce the growth of several sulfate-reducing bacteria [250], notably the species Desulfovibrio vulgaris found in the human intestine. Formate has been demonstrated to be secreted by the intestinal pathogen Shigella flexneri, promoting the expression of genes involved in its own virulence [251].
Lactic acid bacteria ferment several substrates predominantly to lactate [252]. Lactate is used as an oxidative substrate and an ATP source by many intestinal bacteria such as Salmonella and Campylobacter [253,254].
Lastly, concerning citrate, although the tricarboxylic acid cycle is generally not much prominent in bacteria, some intestinal bacterial species such as sulfate-reducing bacteria and Escherichia coli can use several amino acids (as well as other substrates), giving rise during their catabolism to the synthesis of metabolic intermediates of the tricarboxylic acid cycle such as oxaloacetate and succinate (see above), and also of citrate [255,256]. Citrate can also be obtained by bacteria from extracellular sources [257]. Endogenous sources of citrate in bacteria involves citrate synthase which converts oxaloacetate and acetyl-CoA to citrate, while importation from the extracellular media involves citrate transporters [258,259]. Citrate metabolism has been studied in bacterial species such as the intestinal bacteria Lactobacillus casei, and the results obtained show that the major metabolic end products formed are pyruvate, lactate, and acetoin [260]. In intestinal bacteria such as Klebsiella pneumoniae and Enterobacter faecium, citrate represents an oxidative and ATP-producing substrate [257,261]. Citrate metabolism in bacteria is at the origin of the generation of a membrane potential and a pH gradient that are involved in ATP synthesis [262,263]. In addition to its role in bacterial energy metabolism, citrate is a precursor for the synthesis of staphyloferrins. These compounds, which are part of the family of siderophores, enhance bacterial growth under condition of iron restriction [264].
4. Conclusion and Perspectives
The idea that the host is the main supplier of amino acids for the metabolism and physiology of bacteria contained within the intestinal luminal fluid originates from the fact that significant amounts of undigested proteins, either from dietary or endogenous origin, are transferred from the ileum to the large intestine. The utilization of these proteins, after proteolysis by the intestinal bacteria, may be viewed, as we did in the present review, as a form of recycling of amino acids which have not been used by the hosts. These amino acids can be utilized for bacterial metabolism and physiology. Indeed, numerous bacteria of the intestinal microbiota do not possess the metabolic capacity to synthesize the 22 amino acids required for protein synthesis and for utilization of specific amino acids in other metabolic pathways. Thus, these amino acids which must be available from the extracellular medium can be viewed as “bacterially indispensable” for the survival of the intestinal bacteria and for the maintenance of the bacterial mass within the large intestine in a context of regular excretion of bacteria in the feces.
In this paper, we have focused on the effects of the amino acid-derived metabolites which have been shown to be involved in the regulation of the intestinal bacteria growth and in the bacterial metabolism and physiology. A growing number of articles reports on the effects of such bacterial amino acid derivatives on the virulence of the intestinal bacteria, on their capacity to form biofilms, and on their ability to face changing luminal parameters such as osmolarity, pH, and oxidative stress generating environment. Effects of amino acid-derived metabolites on bacterial respiration and energy metabolism have also been reported, and such effects have been in some cases linked with effects on bacterial growth. Also of major interest, recent papers indicate that the metabolism of amino acids within intestinal bacteria leads to the production of compounds which are released in the extracellular medium and which can act on other bacterial species present in the medium, thus revealing new mode of communication between bacteria from different species. From few emerging results presented in this review, it appears that this type of communication is not exclusively related to communication between intestinal bacteria but is also related to communication between intestinal bacteria and other intestinal microorganisms such as parasites and fungi.
The quantities of the amino acid precursors derived from undigested proteins which are available for the metabolism of bacteria in the large intestine luminal fluid represent likely a parameter which will influence the concentrations of the bioactive bacterial metabolites derived from amino acids in this fluid. In support of this concept, it has been shown in a randomized controlled double-blind clinical trial that supplementation with either plant or animal protein (with different amino acid composition but given in the same quantity) in volunteers led to differences in the composition of the amino acid-derived bacterial metabolites in feces and urine [265].
However, some reservations must be formulated regarding the interpretation of the available results and numerous questions need to be answered in future experimental and clinical works. Firstly, most studies have been performed in vitro with intestinal bacteria in a context that often does not fully mimic the conditions that prevail in the large intestine. Secondly, the amino acid-derived metabolites are often tested at concentrations which may not be the ones that are found in the human large intestine luminal fluid. Of note, the concentrations of bacterial metabolites within the large intestine luminal fluid may for some of them largely diverge among individuals, and further information are needed to determine the proportion of metabolites in their free (presumably bioactive) form and in forms bound to luminal compounds. In addition, the intracellular concentrations of metabolites in intestinal bacteria which are associated with bioactive effects remain unknown in almost all cases. Third, these amino acid derivatives are generally tested individually, thus not representing the complex mixture of these derivatives as found in the large intestine content in real life situation.
These reservations being made, we are facing here an exciting and stimulating area of research in which future works should help to better understand the way amino acid metabolism within the intestinal bacteria influence their growth and physiology, and thus finally the composition of the intestinal microbiota in terms of commensal and pathogenic microorganisms. This will certainly provide precious indications which will prove to be useful in terms of human health from both preventive and curative perspectives.
Funding
This research received no specific funding.
Informed Consent statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Blachier, F. Metabolism of Alimentary Compounds by the intestinal microbiota and Health; Springer Nature: Cham, Switzerland, 2023. [Google Scholar]
- Schleifer, KH. Classification of bacteria and archaea: past, present and future. Syst. Appl. Microbiol. 2009, 32, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Matijašić M, Meštrović T, Paljetak HČ, Perić M, Barešić A, Verbanac D. Gut microbiota beyond bacteria-mycobiome, virome, archaeome, and eukaryotic parasites in IBD. Int. J. Mol. Sci. 2020, 21, 2668. [Google Scholar] [CrossRef]
- Rigottier-Gois, L. Dysbiosis in inflammatory bowel diseases: the oxygen hypothesis. ISME J. 2013, 7, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Carding SR, Davis N, Hoyles L. Review article: the human intestinal virome in health and disease. Aliment. Pharmacol. Ther. 2017, 46, 800–815. [Google Scholar] [CrossRef]
- Sausset R, Petit MA, Gaboriau-Routhiau V, De Paepe M. New insights into intestinal phages. Mucosal. Immunol. 2020, 13, 205–215. [Google Scholar] [CrossRef]
- Sartor RB, Wu GD. Roles for Intestinal Bacteria, Viruses, and Fungi in Pathogenesis of Inflammatory Bowel Diseases and Therapeutic Approaches. Gastroenterology. 2017, 152, 327–339. [Google Scholar] [CrossRef]
- Burgess SL, Gilchrist CA, Lynn TC, Petri WA Jr. Parasitic Protozoa and Interactions with the Host Intestinal Microbiota. Infect. Immun. 2017, 85, e00101–17. [Google Scholar] [CrossRef]
- Marteau P, Pochart P, Doré J, Béra-Maillet C, Bernalier A, Corthier G. Comparative study of bacterial groups within the human cecal and fecal microbiota. Appl. Environ. Microbiol. 2001, 67, 4939–4942. [Google Scholar] [CrossRef] [PubMed]
- Stephen AM, Cummings JH. The microbial contribution to human faecal mass. J. Med. Microbiol. 1980, 13, 45–56. [Google Scholar] [CrossRef]
- Stephen AM, Wiggins HS, Cummings JH. Effect of changing transit time on colonic microbial metabolism in man. Gut. 1987, 28, 601–609. [Google Scholar] [CrossRef]
- Bharucha AE, Anderson B, Bouchoucha M. More movement with evaluating colonic transit in humans. Neurogastroenterol. Motil. 2019, 31, e13541. [Google Scholar] [CrossRef]
- Flint HJ, Scott KP, Louis P, Duncan SH. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Winter SE, Bäumler AJ. Why related bacterial species bloom simultaneously in the gut: principles underlying the ‘Like will to like’ concept. Cell. Microbiol. 2014, 16, 179–184. [Google Scholar] [CrossRef]
- Macfarlane GT, Cummings JH. The colonic flora, fermentation, and large bowel digestive function. In: Phillips SF, Pemberton JH, Shorter RG, editors. The large intestine: physiology, pathophysiology, and disease. Raven Press, New York, USA, 1991.
- Windey K, De Preter V, Verbeke K. Relevance of protein fermentation to gut health. Mol. Nutr. Food Res. 2012, 56, 184–196. [Google Scholar] [CrossRef]
- Korpela, K. Diet, Microbiota, and Metabolic Health: Trade-Off Between Saccharolytic and Proteolytic Fermentation. Annu. Rev. Food Sci. Technol. 2018, 9, 65–84. [Google Scholar] [CrossRef]
- Smith EA, Macfarlane GT. Enumeration of human colonic bacteria producing phenolic and indolic compounds: effects of pH, carbohydrate availability and retention time on dissimilatory aromatic amino acid metabolism. J. Appl. Bacteriol. 1996, 81, 288–302. [Google Scholar] [CrossRef]
- Birkett A, Muir J, Phillips J, Jones G, O’Dea K. Resistant starch lowers fecal concentrations of ammonia and phenols in humans. Am. J. Clin. Nutr. 1996, 63, 766–772. [Google Scholar] [CrossRef]
- Geboes KP, De Hertogh G, De Preter V, Luypaerts A, Bammens B, Evenepoel P, Ghoos Y, Geboes K, Rutgeerts P, Verbeke K. The influence of inulin on the absorption of nitrogen and the production of metabolites of protein fermentation in the colon. Br. J. Nutr. 2006, 96, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Cummings JH, Hill MJ, Bone ES, Branch WJ, Jenkins DJ. The effect of meat protein and dietary fiber on colonic function and metabolism. II. Bacterial metabolites in feces and urine. Am. J. Clin. Nutr. 1979, 32, 2094–2101. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane GT, Cummings JH, Macfarlane S, Gibson GR. Influence of retention time on degradation of pancreatic enzymes by human colonic bacteria grown in a 3-stage continuous culture system. J. Appl. Bacteriol. 1989, 67, 520–527. [Google Scholar]
- Blachier, F. Amino acid metabolism for bacterial physiology. In: The evolutionary journey of amino acids. From the origin of life to human metabolism. Springer Nature, Cham, Switzerland, 2025.
- Bröer, S. Intestinal Amino Acid Transport and Metabolic Health. Annu. Rev. Nutr. 2023, 43, 73–99. [Google Scholar] [CrossRef] [PubMed]
- Gaudichon C, Bos C, Morens C, Petzke KJ, Mariotti F, Everwand J, Benamouzig R, Daré S, Tomé D, Metges CC. Ileal losses of nitrogen and amino acids in humans and their importance to the assessment of amino acid requirements. Gastroenterology. 2002, 123, 50–59. [Google Scholar] [CrossRef]
- Blachier F, Andriamihaja M, Kong XF. Fate of undigested proteins in the pig large intestine: What impact on the colon epithelium? Anim. Nutr. 2021, 9, 110–118. [Google Scholar] [CrossRef]
- Baglieri A, Mahe S, Zidi S, Huneau JF, Thuillier F, Marteau P, Tome D. Gastro-jejunal digestion of soya-bean-milk protein in humans. Br. J. Nutr. 1994, 72, 519–532. [Google Scholar] [CrossRef]
- Bos C, Juillet B, Fouillet H, Turlan L, Daré S, Luengo C, N’tounda R, Benamouzig R, Gausserès N, Tomé D, Gaudichon C. Postprandial metabolic utilization of wheat protein in humans. Am. J. Clin. Nutr. 2005, 81, 87–94. [Google Scholar] [CrossRef]
- Yao CK, Muir JG, Gibson PR. Review article: insights into colonic protein fermentation, its modulation and potential health implications. Aliment.Pharmacol. Ther. 2016, 43, 181–196. [Google Scholar] [CrossRef]
- Gibson JA, Sladen GE, Dawson AM. Protein absorption and ammonia production: the effects of dietary protein and removal of the colon. Br. J. Nutr. 1976, 35, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Kramer, P. The effect of varying sodium loads on the ileal excreta of human ileostomized subjects J. Clin. Invest. 1966, 45, 1710–1718. [Google Scholar] [CrossRef]
- Smiddy FG, Gregory SD, Smith IB, Goligher J. Faecal loss of fluid, electrolytes, and nitrogen in colitis before and after ileostomy. Lancet. 7114.
- Chacko A, Cummings JH. Nitrogen losses from the human small bowel: obligatory losses and the effect of physical form of food. Gut. 1988, 29, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Dubuisson C, Lioret S, Touvier M, Dufour A, Calamassi-Tran G, Volatier JL, Lafay L. Trends in food and nutritional intakes of French adults from 1999 to 2007: results from the INCA surveys. Br. J. Nutr. 2010, 103, 1035–1048. [Google Scholar] [CrossRef]
- Pasiakos SM, Agarwal S, Lieberman HR, Fulgoni VL 3rd. Sources and Amounts of Animal, Dairy, and Plant Protein Intake of US Adults in 2007-2010. Nutrients. 2015, 7, 7058–7069. [Google Scholar] [CrossRef]
- Blachier, F. Amino acid metabolism in the large intestine and physiological consequences. In: The evolutionary journey of amino acids. From the origin of life to human metabolism. Springer Nature, Cham, Switzerland, 2025.
- Kaman WE, Hays JP, Endtz HP, Bikker FJ. Bacterial proteases: targets for diagnostics and therapy. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1081–1087. [Google Scholar] [CrossRef]
- Portune K, Beaumont M, Davila AM, Tomé D, Blachier F, Sanz Y. Gut microbiota role in protein metabolism and health-related outcomes: the two side of the coin. Trends Food Sci. Technol. 2016, 57, 213–232. [Google Scholar] [CrossRef]
- Pessione, E. Lactic acid bacteria contribution to gut microbiota complexity: lights and shadows. Front. Cell Infect. Microbiol. 2012, 2, 86. [Google Scholar] [CrossRef] [PubMed]
- Liu M, Bayjanov JR, Renckens B, Nauta A, Siezen RJ. The proteolytic system of lactic acid bacteria revisited: a genomic comparison. BMC Genomics. 2010, 11, 36. [Google Scholar]
- Saier MH, Jr. Families of transmembrane transporters selective for amino acids and their derivatives. Microbiology (Reading). 2000, 146, 1775–1795. [Google Scholar]
- Tanaka KJ, Song S, Mason K, Pinkett HW. Selective substrate uptake: The role of ATP-binding cassette (ABC) importers in pathogenesis. Biochim. Biophys. Acta Biomembr. 2018, 1860, 868–877. [Google Scholar] [CrossRef]
- Hosie AH, Poole PS. Bacterial ABC transporters of amino acids. Res. Microbiol. 2001, 152, 259–270. [Google Scholar] [CrossRef]
- Burkovski A, Krämer R. Bacterial amino acid transport proteins: occurrence, functions, and significance for biotechnological applications. Appl. Microbiol. Biotechnol. 2002, 58, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Konings, WN. The cell membrane and the struggle for life of lactic acid bacteria. Antonie Van Leeuwenhoek. 2002, 82, 3–27. [Google Scholar] [CrossRef]
- Chen Y, Dinges MM, Green A, Cramer SE, Larive CK, Lytle C. Absorptive transport of amino acids by the rat colon. Am. J. Physiol. 2020, 318, G189–G202. [Google Scholar]
- van der Wielen N, Moughan PJ, Mensink M. Amino acid absorption in the large intestine of humans and porcine models. J. Nutr. 2017, 147, 1493–1498. [Google Scholar] [CrossRef] [PubMed]
- Fuller, M. Determination of protein and amino acid digestibility in foods including implications of gut microbial amino acid synthesis. Br. J. Nutr. 2012, 108, S238–S246. [Google Scholar] [CrossRef]
- Darragh AJ, Cranwell PD, Moughan PJ. Absorption of lysine and methionine from the proximal colon of the piglet. Br. J. Nutr. 1994, 71, 739–752. [Google Scholar] [CrossRef] [PubMed]
- Schaafsma, G. The protein digestibility-corrected amino acid score. J. Nutr. 2000, 130, 1865S–1867S. [Google Scholar] [CrossRef]
- Metges, CC. Contribution of microbial amino acids to amino acid homeostasis of the host. J. Nutr. 2000, 130, 1857S–1864S. [Google Scholar] [CrossRef]
- Fuller MF, Reeds PJ. Nitrogen cycling in the gut. Annu. Rev. Nutr. 1998, 18, 385–411. [Google Scholar] [CrossRef]
- Niiyama M, Deguchi E, Kagota K, Namioka S. Appearance of 15N-labeled intestinal microbial amino acids in the venous blood of the pig colon. Am. J. Vet. Res. 1979, 40, 716–718. [Google Scholar] [CrossRef]
- Nakanishi T, Hatanaka T, Huang W, Prasad PD, Leibach FH, Ganapathy ME, Ganapathy V. Na+- and Cl--coupled active transport of carnitine by the amino acid transporter ATB(0,+) from mouse colon expressed in HRPE cells and Xenopus oocytes. J. Physiol. 2001, 532, 297–304. [Google Scholar] [CrossRef]
- Hatanaka T, Huang W, Nakanishi T, Bridges CC, Smith SB, Prasad PD, Ganapathy ME, Ganapathy V. Transport of D-serine via the amino acid transporter ATB(0,+) expressed in the colon. Biochem. Biophys. Res. Commun. 2002, 291, 291–295. [Google Scholar] [CrossRef]
- Ugawa S, Sunouchi Y, Ueda T, Takahashi E, Saishin Y, Shimada S. Characterization of a mouse colonic system B(0+) amino acid transporter related to amino acid absorption in colon. Am. J. Physiol. 2001, 281, G365–G370.
- Blachier F, Mariotti F, Huneau JF, Tomé D. Effects of amino acid-derived luminal metabolites on the colonic epithelium and physiopathological consequences. Amino Acids. 2007, 33, 547–562. [Google Scholar] [CrossRef] [PubMed]
- James PS, Smith MW. Methionine transport by pig colonic mucosa measured during early post-natal development. J. Physiol. 1976, 262, 151–168. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda FV, Smith MW. Different mechanisms for neutral amino acid uptake by new-born pig colon. J. Physiol. 1979, 286, 479–490. [Google Scholar] [CrossRef]
- Hou Y, Yin Y, Wu G. Dietary essentiality of “nutritionally non-essential amino acids” for animals and humans. Exp. Biol. Med. (Maywood). 2015, 240, 997–1007. [Google Scholar] [CrossRef]
- Mathai JK, Liu Y, Stein HH. Values for digestible indispensable amino acid scores (DIAAS) for some dairy and plant proteins may better describe protein quality than values calculated using the concept for protein digestibility-corrected amino acid scores (PDCAAS). Br. J. Nutr. 2017, 117, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Detzner J, Pohlentz G, Müthing J. Enterohemorrhagic Escherichia coli and a fresh view on Shiga toxin-binding glycosphingolipids of primary human kidney and colon epithelial cells and their toxin susceptibility. Int. J. Mol. Sci. 2022, 23, 6884. [Google Scholar] [CrossRef]
- Zhang YG, Singhal M, Lin Z, Manzella C, Kumar A, Alrefai WA, Dudeja PK, Saksena S, Sun J, Gill RK. Infection with enteric pathogens Salmonella typhimurium and Citrobacter rodentium modulate TGF-beta/Smad signaling pathways in the intestine. Gut Microbes. 2018, 9, 326–337. [Google Scholar]
- Jones SE, Knight KL. Bacillus subtilis-mediated protection from Citrobacter rodentium-associated enteric disease requires espH and functional flagella. Infect. Immun. 2012, 80, 710–719. [Google Scholar] [CrossRef]
- Shrestha A, Mehdizadeh Gohari I, Li J, Navarro M, Uzal FA, McClane BA. The biology and pathogenicity of Clostridium perfringens type F: a common human enteropathogen with a new(ish) name. Microbiol. Mol. Biol. Rev. 2024, 88, e0014023.
- Shimizu T, Ohtani K, Hirakawa H, Ohshima K, Yamashita A, Shiba T, Ogasawara N, Hattori M, Kuhara S, Hayashi H. Complete genome sequence of Clostridium perfringens, an anaerobic flesh-eater. Proc. Natl. Acad. Sci. U S A. 2002, 99, 996–1001. [Google Scholar] [CrossRef]
- Denou E, Rezzonico E, Panoff JM, Arigoni F, Brüssow H. A Mesocosm of Lactobacillus johnsonii, Bifidobacterium longum, and Escherichia coli in the mouse gut. DNA Cell. Biol. 2009, 28, 413–422. [Google Scholar] [CrossRef]
- Pridmore RD, Berger B, Desiere F, Vilanova D, Barretto C, Pittet AC, Zwahlen MC, Rouvet M, Altermann E, Barrangou R, Mollet B, Mercenier A, Klaenhammer T, Arigoni F, Schell MA. The genome sequence of the probiotic intestinal bacterium Lactobacillus johnsonii NCC 533. Proc. Natl. Acad. Sci. U S A. 2004, 101, 2512–2517. [Google Scholar] [CrossRef] [PubMed]
- Young KT, Davis LM, Dirita VJ. Campylobacter jejuni: molecular biology and pathogenesis. Nat. Rev. Microbiol. 2007, 5, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Fan TJ, Goeser L, Naziripour A, Redinbo MR, Hansen JJ. Enterococcus faecalis gluconate phosphotransferase system accelerates experimental colitis and bacterial killing by macrophages. Infect. Immun. 2019, 87, e00080–19. [Google Scholar] [CrossRef] [PubMed]
- Yu XJ, Walker DH, Liu Y, Zhang L. Amino acid biosynthesis deficiency in bacteria associated with human and animal hosts. Infect. Genet. Evol. 2009, 9, 514–517. [Google Scholar] [CrossRef]
- Gomes-Santos AC, de Oliveira RP, Moreira TG, Castro-Junior AB, Horta BC, Lemos L, de Almeida LA, Rezende RM, Cara DC, Oliveira SC, Azevedo VA, Miyoshi A, Faria AM. Hsp65-producing Lactococcus lactis prevents inflammatory intestinal disease in mice by IL-10- and TLR2-dependent pathways. Front. Immunol. 2017, 8, 30. [Google Scholar]
- Bolotin A, Wincker P, Mauger S, Jaillon O, Malarme K, Weissenbach J, Ehrlich SD, Sorokin A. The complete genome sequence of the lactic acid bacterium Lactococcus lactis ssp. lactis IL1403. Genome Res. 2001, 11, 731–753. [Google Scholar] [CrossRef]
- Godon JJ, Delorme C, Bardowski J, Chopin MC, Ehrlich SD, Renault P. Gene inactivation in Lactococcus lactis: branched-chain amino acid biosynthesis. J Bacteriol. 1993, 175, 4383–4390. [Google Scholar] [CrossRef]
- Alsultan A, Walton G, Andrews SC, Clarke SR. Staphylococcus aureus FadB is a dehydrogenase that mediates cholate resistance and survival under human colonic conditions. Microbiology (Reading). 2023, 169, 001314.
- Kuroda M, Ohta T, Uchiyama I, Baba T, Yuzawa H, Kobayashi I, Cui L, Oguchi A, Aoki K, Nagai Y, Lian J, Ito T, Kanamori M, Matsumaru H, Maruyama A, Murakami H, Hosoyama A, Mizutani-Ui Y, Takahashi NK, Sawano T, Inoue R, Kaito C, Sekimizu K, Hirakawa H, Kuhara S, Goto S, Yabuzaki J, Kanehisa M, Yamashita A, Oshima K, Furuya K, Yoshino C, Shiba T, Hattori M, Ogasawara N, Hayashi H, Hiramatsu K. Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet. 2001, 357, 1225–1240. [Google Scholar] [CrossRef]
- Njenga R, Boele J, Öztürk Y, Koch HG. Coping with stress: How bacteria fine-tune protein synthesis and protein transport. J. Biol. Chem. 2023, 299, 105163. [Google Scholar] [CrossRef]
- Tollerson R 2nd, Ibba M. Translational regulation of environmental adaptation in bacteria. J. Biol. Chem. 2020, 295, 10434–10445. [Google Scholar] [CrossRef]
- Macek B, Forchhammer K, Hardouin J, Weber-Ban E, Grangeasse C, Mijakovic I. Protein post-translational modifications in bacteria. Nat. Rev. Microbiol. 2019, 17, 651–664. [Google Scholar] [CrossRef]
- Hu Y, Qing Y, Chen J, Liu C, Lu J, Wang Q, Zhen S, Zhou H, Huang L, Zhang R. Prevalence, risk factors, and molecular epidemiology of intestinal carbapenem-resistant Pseudomonas aeruginosa. Microbiol. Spectr. 2021, 9, e0134421. [Google Scholar] [CrossRef] [PubMed]
- Zhang J, Hoedt EC, Liu Q, Berendsen E, Teh JJ, Hamilton A, O’ Brien AW, Ching JYL, Wei H, Yang K, Xu Z, Wong SH, Mak JWY, Sung JJY, Morrison M, Yu J, Kamm MA, Ng SC. Elucidation of Proteus mirabilis as a key bacterium in Crohn’s disease inflammation. Gastroenterology. 2021, 160, 317–330.e11. [Google Scholar] [CrossRef]
- Charlier D, Nguyen Le Minh P, Roovers M. Regulation of carbamoylphosphate synthesis in Escherichia coli: an amazing metabolite at the crossroad of arginine and pyrimidine biosynthesis. Amino Acids. 2018, 50, 1647–1661. [Google Scholar] [CrossRef]
- Leiva LE, Zegarra V, Bange G, Ibba M. At the crossroad of nucleotide dynamics and protein synthesis in bacteria. Microbiol. Mol. Biol. Rev. 2023, 87, e0004422. [Google Scholar] [CrossRef] [PubMed]
- Oliphant K, Allen-Vercoe E. Macronutrient metabolism by the human gut microbiome: major fermentation by-products and their impact on host health. Microbiome. 2019, 7, 91. [Google Scholar]
- Davila AM, Blachier F, Gotteland M, Andriamihaja M, Benetti PH, Sanz Y, Tomé D. Intestinal luminal nitrogen metabolism: role of the gut microbiota and consequences for the host. Pharmacol. Res. 2013, 68, 95–107. [Google Scholar] [CrossRef]
- Kim J, Hetzel M, Boiangiu CD, Buckel W. Dehydration of (R)-2-hydroxyacyl-CoA to enoyl-CoA in the fermentation of alpha-amino acids by anaerobic bacteria. FEMS Microbiol. Rev. 2004, 28, 455–468. [Google Scholar] [CrossRef]
- Buckel, W. Energy conservation in fermentations of anaerobic bacteria. Front. Microbiol. 2021, 12, 703525. [Google Scholar] [CrossRef]
- Macfarlane GT, Macfarlane S. Bacteria, colonic fermentation, and gastrointestinal health. J. AOAC Int. 2012, 95, 50–60. [Google Scholar] [CrossRef]
- Barker, HA. Amino acid degradation by anaerobic bacteria. Annu. Rev. Biochem. 1981, 50, 23–40. [Google Scholar] [CrossRef]
- Pessione A, Lamberti C, Pessione E. Proteomics as a tool for studying energy metabolism in lactic acid bacteria. Mol. Biosyst. 2010, 6, 1419–1430. [Google Scholar] [CrossRef]
- Fernández M, Zúñiga M. Amino acid catabolic pathways of lactic acid bacteria. Crit. Rev. Microbiol. 2006, 32, 155–183. [Google Scholar] [CrossRef] [PubMed]
- Pereira CI, Matos D, San Romão MV, Crespo MT. Dual role for the tyrosine decarboxylation pathway in Enterococcus faecium E17: response to an acid challenge and generation of a proton motive force. Appl. Environ. Microbiol. 2009, 75, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Stickland, LH. Studies in the metabolism of the strict anaerobes (Genus Clostridium): The reduction of proline by Cl. sporogenes. Biochem. J. 1935, 29, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Marshall A, McGrath JW, Graham R, McMullan G. Food for thought-The link between Clostridioides difficile metabolism and pathogenesis. PLoS Pathog. 2023, 19, e1011034. [Google Scholar]
- Pruss KM, Enam F, Battaglioli E, DeFeo M, Diaz OR, Higginbottom SK, Fischer CR, Hryckowian AJ, Van Treuren W, Dodd D, Kashyap P, Sonnenburg JL. Oxidative ornithine metabolism supports non-inflammatory C. difficile colonization. Nat. Metab. 2022, 4, 19–28. [Google Scholar] [CrossRef]
- Pavao A, Graham M, Arrieta-Ortiz ML, Immanuel SRC, Baliga NS, Bry L. Reconsidering the in vivo functions of Clostridial Stickland amino acid fermentations. Anaerobe. 2022, 76, 102600. [Google Scholar] [CrossRef]
- Fonknechten N, Chaussonnerie S, Tricot S, Lajus A, Andreesen JR, Perchat N, Pelletier E, Gouyvenoux M, Barbe V, Salanoubat M, Le Paslier D, Weissenbach J, Cohen GN, Kreimeyer A. Clostridium sticklandii, a specialist in amino acid degradation:revisiting its metabolism through its genome sequence. BMC Genomics. 2010, 11, 555. [Google Scholar]
- Blachier, F. Amino acid-derived bacterial metabolites in the colorectal luminal fluid: effects on microbial communication, metabolism, physiology, and growth. Microorganisms. 2023, 11, 1317. [Google Scholar] [CrossRef]
- Michael, AJ. Polyamines in eukaryotes, bacteria, and archaea. J. Biol. Chem. 2016, 291, 14896–14903. [Google Scholar] [CrossRef]
- Li B, Baniasadi HR, Liang J, Phillips MA, Michael AJ. New routes for spermine biosynthesis. J. Biol. Chem. 2025, 301, 108390. [Google Scholar] [CrossRef]
- Shah P, Swiatlo E. A multifaceted role for polyamines in bacterial pathogens. Mol. Microbiol. 2008, 68, 4–16. [Google Scholar] [CrossRef]
- Yoshida M, Kashiwagi K, Shigemasa A, Taniguchi S, Yamamoto K, Makinoshima H, Ishihama A, Igarashi K. A unifying model for the role of polyamines in bacterial cell growth, the polyamine modulon. J. Biol. Chem. 2004, 279, 46008–46013. [Google Scholar] [CrossRef]
- Chattopadhyay MK, Keembiyehetty CN, Chen W, Tabor H. Polyamines stimulate the level of the sigma38 subunit (RpoS) of Escherichia coli RNA polymerase, resulting in the induction of the glutamate decarboxylase-dependent acid response system via the gadE regulon. J. Biol. Chem. 2015, 290, 17809–17821. [Google Scholar] [CrossRef] [PubMed]
- Igarashi K, Kashiwagi K. Effects of polyamines on protein synthesis and growth of Escherichia coli. J. Biol. Chem. 2018, 293, 18702–18709. [Google Scholar] [CrossRef] [PubMed]
- Igarashi K, Kashiwagi K. Polyamine transport in bacteria and yeast. Biochem. J. 1999, 344, 633–642. [Google Scholar] [CrossRef]
- Driessen AJ, Smid EJ, Konings WN. Transport of diamines by Enterococcus faecalis is mediated by an agmatine-putrescine antiporter. J. Bacteriol. 1988, 170, 4522–4527. [Google Scholar] [CrossRef] [PubMed]
- Large, PJ. Enzymes and pathways of polyamine breakdown in microorganisms. FEMS Microbiol. Rev. 1992, 8, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay MK, Tabor CW, Tabor H. Polyamines are not required for aerobic growth of Escherichia coli: preparation of a strain with deletions in all of the genes for polyamine biosynthesis. J. Bacteriol. 2009, 191, 5549–5552. [Google Scholar] [CrossRef]
- Nakada Y, Itoh Y. Identification of the putrescine biosynthetic genes in Pseudomonas aeruginosa and characterization of agmatine deiminase and N-carbamoylputrescine amidohydrolase of the arginine decarboxylase pathway. Microbiology (Reading). 2003, 149, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Hanfrey CC, Pearson BM, Hazeldine S, Lee J, Gaskin DJ, Woster PM, Phillips MA, Michael AJ. Alternative spermidine biosynthetic route is critical for growth of Campylobacter jejuni and is the dominant polyamine pathway in human gut microbiota. J. Biol. Chem. 2011, 286, 43301–43312. [Google Scholar] [CrossRef]
- Chagneau CV, Garcie C, Bossuet-Greif N, Tronnet S, Brachmann AO, Piel J, Nougayrède JP, Martin P, Oswald E. The polyamine spermidine modulates the production of the bacterial genotoxin colibactin. mSphere. 2019, 4, e00414–19. [Google Scholar]
- Goforth JB, Walter NE, Karatan E. Effects of polyamines on Vibrio cholerae virulence properties. PLoS One. 2013, 8, e60765. [Google Scholar]
- Kramer J, Özkaya Ö, Kümmerli R. Bacterial siderophores in community and host interactions. Nat. Rev. Microbiol. 2020, 18, 152–163. [Google Scholar] [CrossRef]
- Hamana K, Saito T, Okada M, Sakamoto A, Hosoya R. Covalently linked polyamines in the cell wall peptidoglycan of Selenomonas, Anaeromusa, Dendrosporobacter, Acidaminococcus and Anaerovibrio belonging to the Sporomusa subbranch. J. Gen. Appl. Microbiol. 2002, 48, 177–180. [Google Scholar] [CrossRef]
- Samartzidou H, Mehrazin M, Xu Z, Benedik MJ, Delcour AH. Cadaverine inhibition of porin plays a role in cell survival at acidic pH. J. Bacteriol. 2003, 185, 13–19. [Google Scholar] [CrossRef]
- Tanaka Y, Kimura B, Takahashi H, Watanabe T, Obata H, Kai A, Morozumi S, Fujii T. Lysine decarboxylase of Vibrio parahaemolyticus: kinetics of transcription and role in acid resistance. J. Appl. Microbiol. 2008, 104, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley L, Costerton JW, Stoodley P. Bacterial biofilms: from the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Flemming HC, Wingender J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Blachier F, Beaumont M, Andriamihaja M, Davila AM, Lan A, Grauso M, Armand L, Benamouzig R, Tomé D. Changes in the luminal environment of the colonic epithelial cells and physiopathological consequences. Am. J. Pathol. 2017, 187, 476–486. [Google Scholar] [CrossRef]
- Probert HM, Gibson GR. Bacterial biofilms in the human gastrointestinal tract. Curr. Issues Intest. Microbiol. 2002, 3, 23–27. [Google Scholar]
- Roy R, Tiwari M, Donelli G, Tiwari V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence. 2018, 9, 522–554. [Google Scholar] [CrossRef] [PubMed]
- Solano C, Echeverz M, Lasa I. Biofilm dispersion and quorum sensing. Curr. Opin. Microbiol. 2014, 18, 96–104. [Google Scholar] [CrossRef]
- Mukherjee S, Bassler BL. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Banerji R, Kanojiya P, Saroj SD. Role of interspecies bacterial communication in the virulence of pathogenic bacteria. Crit. Rev. Microbiol. 2020, 46, 136–146. [Google Scholar] [CrossRef]
- Burrell M, Hanfrey CC, Murray EJ, Stanley-Wall NR, Michael AJ. Evolution and multiplicity of arginine decarboxylases in polyamine biosynthesis and essential role in Bacillus subtilis biofilm formation. J. Biol. Chem. 2010, 285, 39224–39238. [Google Scholar] [CrossRef]
- Prentice JA, Bridges AA, Bassler BL. Synergy between c-di-GMP and quorum-sensing signaling in Vibrio cholerae biofilm morphogenesis. J. Bacteriol. 2022, 204, e0024922. [Google Scholar] [CrossRef]
- Karatan E, Duncan TR, Watnick PI. NspS, a predicted polyamine sensor, mediates activation of Vibrio cholerae biofilm formation by norspermidine. J. Bacteriol. 2005, 187, 7434–7443. [Google Scholar] [CrossRef]
- Lee J, Sperandio V, Frantz DE, Longgood J, Camilli A, Phillips MA, Michael AJ. An alternative polyamine biosynthetic pathway is widespread in bacteria and essential for biofilm formation in Vibrio cholerae. J. Biol. Chem. 2009, 284, 9899–9907. [Google Scholar] [CrossRef] [PubMed]
- Sobe RC, Bond WG, Wotanis CK, Zayner JP, Burriss MA, Fernandez N, Bruger EL, Waters CM, Neufeld HS, Karatan E. Spermine inhibits Vibrio cholerae biofilm formation through the NspS-MbaA polyamine signaling system. J. Biol. Chem. 2017, 292, 17025–17036. [Google Scholar] [CrossRef]
- Blachier F, Davila AM, Mimoun S, Benetti PH, Atanasiu C, Andriamihaja M, Benamouzig R, Bouillaud F, Tomé D. Luminal sulfide and large intestine mucosa: friend or foe? Amino Acids. 2010, 39, 335–347. [Google Scholar] [CrossRef]
- Barton LL, Ritz NL, Fauque GD, Lin HC. Sulfur cycling and the intestinal microbiome. Dig. Dis. Sci. 2017, 62, 2241–2257. [Google Scholar] [CrossRef]
- Basic A, Blomqvist M, Dahlén G, Svensäter G. The proteins of Fusobacterium spp. involved in hydrogen sulfide production from L-cysteine. BMC Microbiol. 2017, 17, 61. [Google Scholar] [CrossRef] [PubMed]
- Linden, DR. Hydrogen sulfide signaling in the gastrointestinal tract. Antioxid. Redox. Signal. 2014, 20, 818–830. [Google Scholar] [CrossRef] [PubMed]
- Croix JA, Carbonero F, Nava GM, Russell M, Greenberg E, Gaskins HR. On the relationship between sialomucin and sulfomucin expression and hydrogenotrophic microbes in the human colonic mucosa. PLoS One. 2011, 6, e24447. [Google Scholar]
- Villanueva-Millan MJ, Leite G, Mathur R, Rezaie A, Fajardo CM, de Freitas Germano J, Morales W, Sanchez M, Rivera I, Parodi G, Weitsman S, Rashid M, Hosseini A, Brimberry D, Barlow GM, Pimentel M. Hydrogen Sulfide and Methane on Breath Test Correlate with Human Small Intestinal Hydrogen Sulfide Producers and Methanogens. Dig. Dis. Sci. 2025, in press.
- Gibson GR, Macfarlane GT, Cummings JH. Occurrence of sulphate-reducing bacteria in human faeces and the relationship of dissimilatory sulphate reduction to methanogenesis in the large gut. J. Appl. Bacteriol. 1988, 65, 103–111. [Google Scholar] [CrossRef]
- Ohge H, Furne JK, Springfield J, Sueda T, Madoff RD, Levitt MD. The effect of antibiotics and bismuth on fecal hydrogen sulfide and sulfate-reducing bacteria in the rat. FEMS Microbiol. Lett. 2003, 228, 137–142. [Google Scholar] [CrossRef]
- Rowan FE, Docherty NG, Coffey JC, O’Connell PR. Sulphate-reducing bacteria and hydrogen sulphide in the aetiology of ulcerative colitis. Br. J. Surg. 2009, 96, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Kushkevych I, Dordević D, Alberfkani MI, Gajdács M, Ostorházi E, Vítězová M, Rittmann SKR. NADH and NADPH peroxidases as antioxidant defense mechanisms in intestinal sulfate-reducing bacteria. Sci. Rep. 2023, 13, 13922. [Google Scholar] [CrossRef] [PubMed]
- Florin T, Neale G, Gibson GR, Christl SU, Cummings JH. Metabolism of dietary sulphate: absorption and excretion in humans. Gut. 1991, 32, 766–773. [Google Scholar] [CrossRef]
- Lewis S, Cochrane S. Alteration of sulfate and hydrogen metabolism in the human colon by changing intestinal transit rate. Am. J. Gastroenterol. 2007, 102, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Blachier F, Andriamihaja M, Larraufie P, Ahn E, Lan A, Kim E. Production of hydrogen sulfide by the intestinal microbiota and epithelial cells and consequences for the colonic and rectal mucosa. Am. J. Physiol. 2021, 320, G125–G135. [Google Scholar]
- Bouillaud F, Blachier F. Mitochondria and sulfide: a very old story of poisoning, feeding, and signaling? Antioxid. Redox. Signal. 2011, 15, 379–391. [Google Scholar] [CrossRef]
- Jørgensen J, Mortensen PB. Hydrogen sulfide and colonic epithelial metabolism: implications for ulcerative colitis. Dig. Dis. Sci. 2001, 46, 1722–1732. [Google Scholar] [CrossRef]
- Suarez F, Furne J, Springfield J, Levitt M. Production and elimination of sulfur-containing gases in the rat colon. Am. J. Physiol. 1998, 274, G727–G733. [Google Scholar]
- Jensen B, Fago A. Reactions of ferric hemoglobin and myoglobin with hydrogen sulfide under physiological conditions. J. Inorg. Biochem. 2018, 182, 133–140. [Google Scholar] [CrossRef]
- Andriamihaja M, Lan A, Beaumont M, Grauso M, Gotteland M, Pastene E, Cires MJ, Carrasco-Pozo C, Tomé D, Blachier F. Proanthocyanidin-containing polyphenol extracts from fruits prevent the inhibitory effect of hydrogen sulfide on human colonocyte oxygen consumption. Amino Acids. 2018, 50, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Oldham KEA, Prentice EJ, Summers EL, Hicks JL. Serine acetyltransferase from Neisseria gonorrhoeae; structural and biochemical basis of inhibition. Biochem. J. 2022, 479, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Hicks JL, Mullholland CV. Cysteine biosynthesis in Neisseria species. Microbiology (Reading). 2018, 164, 1471–1480. [Google Scholar] [CrossRef]
- Yao S, Zhao Z, Wang W, Liu X. Bifidobacterium longum: protection against inflammatory bowel disease. J. Immunol. Res. 2021, 2021, 8030297. [Google Scholar] [CrossRef]
- Schell MA, Karmirantzou M, Snel B, Vilanova D, Berger B, Pessi G, Zwahlen MC, Desiere F, Bork P, Delley M, Pridmore RD, Arigoni F. The genome sequence of Bifidobacterium longum reflects its adaptation to the human gastrointestinal tract. Proc. Natl. Acad. Sci. U S A. 2002, 99, 14422–14427. [Google Scholar] [CrossRef]
- Rodionov DA, Vitreschak AG, Mironov AA, Gelfand MS. Comparative genomics of the methionine metabolism in Gram-positive bacteria: a variety of regulatory systems. Nucleic Acids Res. 2004, 32, 3340–3353. [Google Scholar] [CrossRef]
- Ferla MP, Patrick WM. Bacterial methionine biosynthesis. Microbiology (Reading). 2014, 160, 1571–1584. [Google Scholar] [CrossRef] [PubMed]
- Saini V, Chinta KC, Reddy VP, Glasgow JN, Stein A, Lamprecht DA, Rahman MA, Mackenzie JS, Truebody BE, Adamson JH, Kunota TTR, Bailey SM, Moellering DR, Lancaster JR Jr, Steyn AJC. Hydrogen sulfide stimulates Mycobacterium tuberculosis respiration, growth and pathogenesis. Nat. Commun. 2020, 11, 557. [Google Scholar] [CrossRef]
- Borisov VB, Forte E. Impact of hydrogen sulfide on mitochondrial and bacterial bioenergetics. Int. J. Mol. Sci. 2021, 22, 12688. [Google Scholar] [CrossRef]
- Melo AM, Teixeira M. Supramolecular organization of bacterial aerobic respiratory chains: From cells and back. Biochim. Biophys. Acta. 2016, 1857, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Forte E, Borisov VB, Falabella M, Colaço HG, Tinajero-Trejo M, Poole RK, Vicente JB, Sarti P, Giuffrè A. The terminal oxidase cytochrome bd promotes sulfide-resistant bacterial respiration and growth. Sci. Rep. 2016, 6, 23788. [Google Scholar] [CrossRef]
- Nastasi MR, Caruso L, Giordano F, Mellini M, Rampioni G, Giuffrè A, Forte E. Cyanide insensitive oxidase confers hydrogen sulfide and nitric oxide tolerance to Pseudomonas aeruginosa aerobic respiration. Antioxidants (Basel). 2024, 13, 383. [Google Scholar] [CrossRef]
- Bachenheimer AG, Bennett EO. The sensitivity of mixed population of bacteria to inhibitors. I. The mechanism by which Desulfovibrio desulfuricans protects Ps. aeruginosa from the toxicity of mercurials. Antonie Van Leeuwenhoek. 1961, 27, 180–188. [Google Scholar]
- Stutzenberger FJ, Bennett EO. Sensitivity of mixed populations of Staphylococcus aureus and Escherichia coli to mercurials. Appl. Microbiol. 1965, 13, 570–574. [Google Scholar] [CrossRef]
- Shatalin K, Shatalina E, Mironov A, Nudler E. H2S: a universal defense against antibiotics in bacteria. Science. 2011, 334, 986–990. [Google Scholar] [CrossRef]
- Pal VK, Bandyopadhyay P, Singh A. Hydrogen sulfide in physiology and pathogenesis of bacteria and viruses. IUBMB Life. 2018, 70, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Mironov A, Seregina T, Nagornykh M, Luhachack LG, Korolkova N, Lopes LE, Kotova V, Zavilgelsky G, Shakulov R, Shatalin K, Nudler E. Mechanism of H2S-mediated protection against oxidative stress in Escherichia coli. Proc. Natl. Acad. Sci. U S A. 2017, 114, 6022–6027. [Google Scholar] [CrossRef] [PubMed]
- Shukla P, Khodade VS, SharathChandra M, Chauhan P, Mishra S, Siddaramappa S, Pradeep BE, Singh A, Chakrapani H. “On demand” redox buffering by H2S contributes to antibiotic resistance revealed by a bacteria-specific H2S donor. Chem. Sci. 2017, 8, 4967–4972. [Google Scholar] [CrossRef]
- Shatalin K, Nuthanakanti A, Kaushik A, Shishov D, Peselis A, Shamovsky I, Pani B, Lechpammer M, Vasilyev N, Shatalina E, Rebatchouk D, Mironov A, Fedichev P, Serganov A, Nudler E. Inhibitors of bacterial H2S biogenesis targeting antibiotic resistance and tolerance. Science. 2021, 372, 1169–1175. [Google Scholar] [CrossRef]
- Ng SY, Ong KX, Surendran ST, Sinha A, Lai JJH, Chen J, Liang J, Tay LKS, Cui L, Loo HL, Ho P, Han J, Moreira W. Hydrogen Sulfide Sensitizes Acinetobacter baumannii to Killing by Antibiotics. Front. Microbiol. 2020, 11, 1875. [Google Scholar] [CrossRef]
- Stacy A, Andrade-Oliveira V, McCulloch JA, Hild B, Oh JH, Perez-Chaparro PJ, Sim CK, Lim AI, Link VM, Enamorado M, Trinchieri G, Segre JA, Rehermann B, Belkaid Y. Infection trains the host for microbiota-enhanced resistance to pathogens. Cell. 2021, 184, 615–627.e17. [Google Scholar] [CrossRef]
- Motta JP, Flannigan KL, Agbor TA, Beatty JK, Blackler RW, Workentine ML, Da Silva GJ, Wang R, Buret AG, Wallace JL. Hydrogen sulfide protects from colitis and restores intestinal microbiota biofilm and mucus production. Inflamm. Bowel. Dis. 2015, 21, 1006–1017. [Google Scholar] [CrossRef]
- Chen YW, Camacho MI, Chen Y, Bhat AH, Chang C, Peluso EA, Wu C, Das A, Ton-That H. Genetic determinants of hydrogen sulfide biosynthesis in Fusobacterium nucleatum are required for bacterial fitness, antibiotic sensitivity, and virulence. mBio. 2022, 13, 0193622. [Google Scholar]
- Sun J, Wang X, Gao Y, Li S, Hu Z, Huang Y, Fan B, Wang X, Liu M, Qiao C, Zhang W, Wang Y, Ji X. H2S scavenger as a broad-spectrum strategy to deplete bacteria-derived H2S for antibacterial sensitization. Nat. Commun. 2024, 15, 9422. [Google Scholar] [CrossRef]
- Sudhamsu J, Crane BR. Bacterial nitric oxide synthases: what are they good for? Trends Microbiol. 2009, 17, 212–218. [Google Scholar] [CrossRef]
- Adak S, Aulak KS, Stuehr DJ. Direct evidence for nitric oxide production by a nitric-oxide synthase-like protein from Bacillus subtilis. J. Biol. Chem. 2002, 277, 16167–16171. [Google Scholar] [CrossRef]
- Morita H, Yoshikawa H, Sakata R, Nagata Y, Tanaka H. Synthesis of nitric oxide from the two equivalent guanidino nitrogens of L-arginine by Lactobacillus fermentum. J. Bacteriol. 1997, 179, 7812–7815. [Google Scholar] [CrossRef] [PubMed]
- Sellars MJ, Hall SJ, Kelly DJ. Growth of Campylobacter jejuni supported by respiration of fumarate, nitrate, nitrite, trimethylamine-N-oxide, or dimethyl sulfoxide requires oxygen. J. Bacteriol. 2002, 184, 4187–4196. [Google Scholar] [CrossRef] [PubMed]
- Weingarten RA, Grimes JL, Olson JW. Role of Campylobacter jejuni respiratory oxidases and reductases in host colonization. Appl. Environ. Microbiol. 2008, 74, 1367–1375. [Google Scholar] [CrossRef]
- Silvestrini MC, Falcinelli S, Ciabatti I, Cutruzzolà F, Brunori M. Pseudomonas aeruginosa nitrite reductase (or cytochrome oxidase): an overview. Biochimie. 1994, 76, 641–654. [Google Scholar] [CrossRef]
- Daims H, Lebedeva EV, Pjevac P, Han P, Herbold C, Albertsen M, Jehmlich N, Palatinszky M, Vierheilig J, Bulaev A, Kirkegaard RH, von Bergen M, Rattei T, Bendinger B, Nielsen PH, Wagner M. Complete nitrification by Nitrospira bacteria. Nature. 2015, 528, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Zheng Y, Ke J, Song J, Li X, Kuang R, Wang H, Li S, Li Y. Correlation between daily physical activity and intestinal microbiota in perimenopausal women. Sports Med. Health Sci. 2024, 7, 230–236. [Google Scholar]
- Kastner J, Pfeffel F, Rajek A, Pezawas T, Hiesmayr M, Eichler HG. Nitric oxide concentration in the gas phase of the gastrointestinal tract in man. Eur. J. Clin. Invest. 1997, 27, 992–996. [Google Scholar] [CrossRef]
- Richardson AR, Payne EC, Younger N, Karlinsey JE, Thomas VC, Becker LA, Navarre WW, Castor ME, Libby SJ, Fang FC. Multiple targets of nitric oxide in the tricarboxylic acid cycle of Salmonella enterica serovar typhimurium. Cell Host Microbe. 2011, 10, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Stern AM, Zhu J. An introduction to nitric oxide sensing and response in bacteria. Adv. Appl. Microbiol. 2014, 87, 187–220. [Google Scholar]
- Zhao G, Guo YY, Yao S, Shi X, Lv L, Du YL. Nitric oxide as a source for bacterial triazole biosynthesis. Nat. Commun. 2020, 11, 1614. [Google Scholar] [CrossRef]
- Caranto, JD. The emergence of nitric oxide in the biosynthesis of bacterial natural products. Curr. Opin. Chem. Biol. 2019, 49, 130–138. [Google Scholar] [CrossRef]
- Choules MP, Wolf NM, Lee H, Anderson JR, Grzelak EM, Wang Y, Ma R, Gao W, McAlpine JB, Jin YY, Cheng J, Lee H, Suh JW, Duc NM, Paik S, Choe JH, Jo EK, Chang CL, Lee JS, Jaki BU, Pauli GF, Franzblau SG, Cho S. Rufomycin targets ClpC1 proteolysis in Mycobacterium tuberculosis and M. abscessus. Antimicrob. Agents Chemother. 2019, 63, e02204–18. [Google Scholar] [CrossRef]
- Anantharaman S, Guercio D, Mendoza AG, Withorn JM, Boon EM. Negative regulation of biofilm formation by nitric oxide sensing proteins. Biochem. Soc. Trans. 2023, 51, 1447–1458. [Google Scholar] [CrossRef]
- Poh WH, Rice SA. Recent Developments in nitric oxide donors and delivery for antimicrobial and anti-biofilm applications. Molecules. 2022, 27, 674. [Google Scholar] [CrossRef]
- Ueno T, Fischer JT, Boon EM. Nitric oxide enters quorum sensing via the H-NOX signaling pathway in Vibrio parahaemolyticus. Front. Microbiol. 2019, 10, 2108. [Google Scholar] [CrossRef]
- Keszthelyi D, Troost FJ, Masclee AA. Understanding the role of tryptophan and serotonin metabolism in gastrointestinal function. Neurogastroenterol. Motil. 2009, 21, 1239–1249. [Google Scholar] [CrossRef]
- Lee JH, Wood TK, Lee J. Roles of indole as an interspecies and interkingdom signaling molecule. Trends Microbiol. 2015, 23, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Roager HM, Licht TR. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef] [PubMed]
- Rattanaphan P, Mittraparp-Arthorn P, Srinoun K, Vuddhakul V, Tansila N. Indole signaling decreases biofilm formation and related virulence of Listeria monocytogenes. FEMS Microbiol. Lett. 2020, 367, fnaa116. [Google Scholar] [CrossRef]
- Lee J, Attila C, Cirillo SL, Cirillo JD, Wood TK. Indole and 7-hydroxyindole diminish Pseudomonas aeruginosa virulence. Microb. Biotechnol. 2009, 2, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Nikaido E, Giraud E, Baucheron S, Yamasaki S, Wiedemann A, Okamoto K, Takagi T, Yamaguchi A, Cloeckaert A, Nishino K. Effects of indole on drug resistance and virulence of Salmonella enterica serovar Typhimurium revealed by genome-wide analyses. Gut Pathog. 2012, 4, 5. [Google Scholar] [CrossRef]
- Oh S, Go GW, Mylonakis E, Kim Y. The bacterial signalling molecule indole attenuates the virulence of the fungal pathogen Candida albicans. J. Appl. Microbiol. 2012, 113, 622–628. [Google Scholar] [CrossRef]
- Talapko J, Juzbašić M, Matijević T, Pustijanac E, Bekić S, Kotris I, Škrlec I. Candida albicans-The virulence factors and clinical manifestations of infection. J. Fungi (Basel). 2021, 7, 79.
- Nowak A, Libudzisz Z. Influence of phenol, p-cresol and indole on growth and survival of intestinal lactic acid bacteria. Anaerobe. 2006, 12, 80–84. [Google Scholar] [CrossRef]
- Ledala N, Malik M, Rezaul K, Paveglio S, Provatas A, Kiel A, Caimano M, Zhou Y, Lindgren J, Krasulova K, Illes P, Dvořák Z, Kortagere S, Kienesberger S, Cosic A, Pöltl L, Zechner EL, Ghosh S, Mani S, Radolf JD, Matson AP. Bacterial indole as a multifunctional regulator of Klebsiella oxytoca complex enterotoxicity. mBio. 2022, 13, e0375221. [Google Scholar]
- Högenauer C, Langner C, Beubler E, Lippe IT, Schicho R, Gorkiewicz G, Krause R, Gerstgrasser N, Krejs GJ, Hinterleitner TA. Klebsiella oxytoca as a causative organism of antibiotic-associated hemorrhagic colitis. N. Engl. J. Med. 2006, 355, 2418–2426. [Google Scholar] [CrossRef]
- Bone E, Tamm A, Hill M. The production of urinary phenols by gut bacteria and their possible role in the causation of large bowel cancer. Am. J. Clin. Nutr. 1976, 29, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Saito Y, Sato T, Nomoto K, Tsuji H. Identification of phenol- and p-cresol-producing intestinal bacteria by using media supplemented with tyrosine and its metabolites. FEMS Microbiol. Ecol. 2018, 94, fiy125.
- Harrison MA, Kaur H, Wren BW, Dawson LF. Production of p-cresol by Decarboxylation of p-HPA by All Five Lineages of Clostridioides difficile Provides a Growth Advantage. Front. Cell. Infect. Microbiol. 2021, 11, 757599. [Google Scholar]
- Passmore IJ, Letertre MPM, Preston MD, Bianconi I, Harrison MA, Nasher F, Kaur H, Hong HA, Baines SD, Cutting SM, Swann JR, Wren BW, Dawson LF. Para-cresol production by Clostridium difficile affects microbial diversity and membrane integrity of Gram-negative bacteria. PLoS Pathog. 2018, 14, e1007191. [Google Scholar]
- Abt MC, McKenney PT, Pamer EG. Clostridium difficile colitis: pathogenesis and host defence. Nat. Rev. Microbiol. 2016, 14, 609–620. [Google Scholar] [CrossRef]
- Hafiz S, Oakley CL. Clostridium difficile: isolation and characteristics. J. Med. Microbiol. 1976, 9, 129–136. [Google Scholar] [CrossRef]
- Dawson LF, Donahue EH, Cartman ST, Barton RH, Bundy J, McNerney R, Minton NP, Wren BW. The analysis of para-cresol production and tolerance in Clostridium difficile 027 and 012 strains. BMC Microbiol. 2011, 11, 86. [Google Scholar] [CrossRef]
- Jensen MT, Cox RP, Jensen BB. 3-Methylindole (skatole) and indole production by mixed populations of pig fecal bacteria. Appl. Environ. Microbiol. 1995, 61, 3180–3184. [Google Scholar] [CrossRef]
- Yokoyama MT, Carlson JR. Microbial metabolites of tryptophan in the intestinal tract with special reference to skatole. Am. J. Clin. Nutr. 1979, 32, 173–178. [Google Scholar] [CrossRef]
- Whitehead TR, Price NP, Drake HL, Cotta MA. Catabolic pathway for the production of skatole and indoleacetic acid by the acetogen Clostridium drakei, Clostridium scatologenes, and swine manure. Appl. Environ. Microbiol. 2008, 74, 1950–1953. [Google Scholar] [CrossRef]
- Choi SH, Kim Y, Oh S, Oh S, Chun T, Kim SH. Inhibitory effect of skatole (3-methylindole) on enterohemorrhagic Escherichia coli O157:H7 ATCC 43894 biofilm formation mediated by elevated endogenous oxidative stress. Lett. Appl. Microbiol. 2014, 58, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Morbach S, Krämer R. Structure and function of the betaine uptake system BetP of Corynebacterium glutamicum: strategies to sense osmotic and chill stress. J. Mol. Microbiol. Biotechnol. 2005, 10, 143–153. [Google Scholar]
- Nau-Wagner G, Opper D, Rolbetzki A, Boch J, Kempf B, Hoffmann T, Bremer E. Genetic control of osmoadaptive glycine betaine synthesis in Bacillus subtilis through the choline-sensing and glycine betaine-responsive GbsR repressor. J. Bacteriol. 2012, 194, 2703–2714. [Google Scholar] [CrossRef] [PubMed]
- Zou H, Chen N, Shi M, Xian M, Song Y, Liu J. The metabolism and biotechnological application of betaine in microorganism. Appl. Microbiol. Biotechnol. 2016, 100, 3865–3876. [Google Scholar] [CrossRef]
- Wargo, MJ. Homeostasis and catabolism of choline and glycine betaine: lessons from Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2013, 79, 2112–2120. [Google Scholar] [CrossRef]
- Ku JW, Gan YH. Modulation of bacterial virulence and fitness by host glutathione. Curr. Opin. Microbiol. 2019, 47, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Masip L, Veeravalli K, Georgiou G. The many faces of glutathione in bacteria. Antioxid. Redox. Signal. 2006, 8, 753–762. [Google Scholar] [CrossRef]
- Landete JM, De las Rivas B, Marcobal A, Muñoz R. Updated molecular knowledge about histamine biosynthesis by bacteria. Crit. Rev. Food Sci. Nutr. 2008, 48, 697–714. [Google Scholar] [CrossRef]
- Dicks LMT. Gut bacteria and neurotransmitters. Microorganisms. 2022, 10, 1838. [Google Scholar] [CrossRef]
- Molenaar D, Bosscher JS, ten Brink B, Driessen AJ, Konings WN. Generation of a proton motive force by histidine decarboxylation and electrogenic histidine/histamine antiport in Lactobacillus buchneri. J. Bacteriol. 1993, 175, 2864–2870. [Google Scholar] [CrossRef]
- Trip H, Mulder NL, Lolkema JS. Improved acid stress survival of Lactococcus lactis expressing the histidine decarboxylation pathway of Streptococcus thermophilus CHCC1524. J. Biol. Chem. 2012, 287, 11195–11204. [Google Scholar] [CrossRef]
- Nugent SG, Kumar D, Rampton DS, Evans DF. Intestinal luminal pH in inflammatory bowel disease: possible determinants and implications for therapy with aminosalicylates and other drugs. Gut. 2001, 48, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef]
- Claus H, Decker H. Bacterial tyrosinases. Syst. Appl. Microbiol. 2006, 29, 3–14. [Google Scholar] [CrossRef]
- Asano Y, Hiramoto T, Nishino R, Aiba Y, Kimura T, Yoshihara K, Koga Y, Sudo N. Critical role of gut microbiota in the production of biologically active, free catecholamines in the gut lumen of mice. Am. J. Physiol. 2012, 303, G1288–G1295. [Google Scholar]
- Belay T, Sonnenfeld G. Differential effects of catecholamines on in vitro growth of pathogenic bacteria. Life Sci. 2002, 71, 447–456. [Google Scholar] [CrossRef]
- Dichtl S, Demetz E, Haschka D, Tymoszuk P, Petzer V, Nairz M, Seifert M, Hoffmann A, Brigo N, Würzner R, Theurl I, Karlinsey JE, Fang FC, Weiss G. Dopamine is a siderophore-like iron chelator that promotes Salmonella enterica serovar Typhimurium virulence in mice. mBio. 2019, 10, e02624–18. [Google Scholar]
- Knecht LD, O’Connor G, Mittal R, Liu XZ, Daftarian P, Deo SK, Daunert S. Serotonin activates bacterial quorum sensing and enhances the virulence of Pseudomonas aeruginosa in the host. EBioMedicine. 2016, 9, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Boyanova, L. Stress hormone epinephrine (adrenaline) and norepinephrine (noradrenaline) effects on the anaerobic bacteria. Anaerobe. 2017, 44, 13–19. [Google Scholar] [CrossRef]
- Lustri BC, Sperandio V, Moreira CG. Bacterial chat: intestinal metabolites and signals in host-microbiota-pathogen interactions. Infect. Immun. 2017, 85, e00476–17. [Google Scholar]
- O’Donnell PM, Aviles H, Lyte M, Sonnenfeld G. Enhancement of in vitro growth of pathogenic bacteria by norepinephrine: importance of inoculum density and role of transferrin. Appl. Environ. Microbiol. 2006, 72, 5097–5099. [Google Scholar] [CrossRef] [PubMed]
- Kim J, Lee MH, Kim MS, Kim GH, Yoon SS. Probiotic properties and optimization of gamma-aminobutyric acid production by Lactiplantibacillus plantarum FBT215. J. Microbiol. Biotechnol. 2022, 32, 783–791. [Google Scholar] [CrossRef]
- Barrett E, Ross RP, O’Toole PW, Fitzgerald GF, Stanton C. gamma-aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef]
- Nomura M, Nakajima I, Fujita Y, Kobayashi M, Kimoto H, Suzuki I, Aso H. Lactococcus lactis contains only one glutamate decarboxylase gene. Microbiology (Reading). 1999, 145, 1375–1380. [Google Scholar] [CrossRef]
- Otaru N, Ye K, Mujezinovic D, Berchtold L, Constancias F, Cornejo FA, Krzystek A, de Wouters T, Braegger C, Lacroix C, Pugin B. GABA Production by human intestinal Bacteroides spp.: prevalence, regulation, and role in acid stress tolerance. Front. Microbiol. 2021, 12, 656895. [Google Scholar] [CrossRef]
- Feehily C, Karatzas KA. Role of glutamate metabolism in bacterial responses towards acid and other stresses. J. Appl. Microbiol. 2013, 114, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Prieto MA, García JL. Identification of the 4-hydroxyphenylacetate transport gene of Escherichia coli W: construction of a highly sensitive cellular biosensor. FEBS Lett. 1997, 414, 293–297. [Google Scholar] [CrossRef] [PubMed]
- YujiaLiu, Shi C, Zhang G, Zhan H, Liu B, Li C, Wang L, Wang H, Wang J. Antimicrobial mechanism of 4-hydroxyphenylacetic acid on Listeria monocytogenes membrane and virulence. Biochem. Biophys. Res. Commun. 2021, 572, 145–150. [Google Scholar] [CrossRef]
- Dai ZL, Wu G, Zhu WY. Amino acid metabolism in intestinal bacteria: links between gut ecology and host health. Front. Biosci (Landmark Ed). 2011, 16, 1768–1786. [Google Scholar] [CrossRef] [PubMed]
- Endo A, Nakamura S, Konishi K, Nakagawa J, Tochio T. Variations in prebiotic oligosaccharide fermentation by intestinal lactic acid bacteria. Int. J. Food Sci. Nutr. 2016, 67, 125–132. [Google Scholar] [CrossRef]
- Liong MT, Shah NP. Production of organic acids from fermentation of mannitol, fructooligosaccharide and inulin by a cholesterol removing Lactobacillus acidophilus strain. J. Appl. Microbiol. 2005, 99, 783–793. [Google Scholar] [CrossRef]
- Goocher CR, Woodside EE, Kocholaty W. The influence of 2,4-dinitrophenol on the oxidation of acetate and succinate by Escherichia coli. J. Bacteriol. 1954, 67, 593–596. [Google Scholar] [CrossRef]
- Lemma E, Hägerhäll C, Geisler V, Brandt U, von Jagow G, Kröger A. Reactivity of the Bacillus subtilis succinate dehydrogenase complex with quinones. Biochim. Biophys Acta. 1991, 1059, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Adolph C, McNeil MB, Cook GM. Impaired succinate oxidation prevents growth and influences drug susceptibility in Mycobacterium tuberculosis. mBio. 2022, 13, e0167222. [Google Scholar]
- Hartman T, Weinrick B, Vilchèze C, Berney M, Tufariello J, Cook GM, Jacobs WR Jr. Succinate dehydrogenase is the regulator of respiration in Mycobacterium tuberculosis. PLoS Pathog. 2014, 10, e1004510. [Google Scholar]
- Ferreyra JA, Wu KJ, Hryckowian AJ, Bouley DM, Weimer BC, Sonnenburg JL. Gut microbiota-produced succinate promotes C. difficile infection after antibiotic treatment or motility disturbance. Cell Host Microbe. 2014, 16, 770–777. [Google Scholar] [CrossRef]
- Auria E, Deschamps J, Briandet R, Dupuy B. Extracellular succinate induces spatially organized biofilm formation in Clostridioides difficile. Biofilm. 2023, 5, 100125. [Google Scholar]
- Hagihara M, Ariyoshi T, Kuroki Y, Eguchi S, Higashi S, Mori T, Nonogaki T, Iwasaki K, Yamashita M, Asai N, Koizumi Y, Oka K, Takahashi M, Yamagishi Y, Mikamo H. Clostridium butyricum enhances colonization resistance against Clostridioides difficile by metabolic and immune modulation. Sci. Rep. 2021, 11, 15007. [Google Scholar] [CrossRef]
- Shaulov Y, Shimokawa C, Trebicz-Geffen M, Nagaraja S, Methling K, Lalk M, Weiss-Cerem L, Lamm AT, Hisaeda H, Ankri S. Escherichia coli mediated resistance of Entamoeba histolytica to oxidative stress is triggered by oxaloacetate. PLoS Pathog. 2018, 14, e1007295. [Google Scholar]
- Guillén, N. Pathogenicity and virulence of Entamoeba histolytica, the agent of amoebiasis. Virulence. 2023, 14, 2158656. [Google Scholar] [CrossRef]
- Kassem II, Candelero-Rueda RA, Esseili KA, Rajashekara G. Formate simultaneously reduces oxidase activity and enhances respiration in Campylobacter jejuni. Sci. Rep. 2017, 7, 40117. [Google Scholar] [CrossRef]
- Voskuhl L, Brusilova D, Brauer VS, Meckenstock RU. Inhibition of sulfate-reducing bacteria with formate. FEMS Microbiol. Ecol. 2022, 98, fiac003. [Google Scholar] [CrossRef]
- Koestler BJ, Fisher CR, Payne SM. Formate promotes Shigella intercellular spread and virulence gene expression. mBio. 2018, 9, e01777–18. [Google Scholar]
- Liu, SQ. Practical implications of lactate and pyruvate metabolism by lactic acid bacteria in food and beverage fermentations. Int. J. Food Microbiol. 2003, 83, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Sheridan PO, Louis P, Tsompanidou E, Shaw S, Harmsen HJ, Duncan SH, Flint HJ, Walker AW. Distribution, organization and expression of genes concerned with anaerobic lactate utilization in human intestinal bacteria. Microb. Genom. 2022, 8, 000739. [Google Scholar] [CrossRef] [PubMed]
- Weghoff MC, Bertsch J, Müller V. A novel mode of lactate metabolism in strictly anaerobic bacteria. Environ. Microbiol. 2015, 17, 670–677. [Google Scholar] [CrossRef]
- Thauer, RK. Citric-acid cycle, 50 years on. Modifications and an alternative pathway in anaerobic bacteria. Eur. J. Biochem. 1988, 176, 497–508. [Google Scholar] [CrossRef]
- Zampieri M, Hörl M, Hotz F, Müller NF, Sauer U. Regulatory mechanisms underlying coordination of amino acid and glucose catabolism in Escherichia coli. Nat. Commun. 2019, 10, 3354. [Google Scholar] [CrossRef]
- Sarantinopoulos P, Makras L, Vaningelgem F, Kalantzopoulos G, De Vuyst L, Tsakalidou E. Growth and energy generation by Enterococcus faecium FAIR-E 198 during citrate metabolism. Int. J. Food Microbiol. 2003, 84, 197–206. [Google Scholar] [CrossRef]
- Muñoz-Elías EJ, McKinney JD. Carbon metabolism of intracellular bacteria. Cell Microbiol. 2006, 8, 10–22. [Google Scholar] [CrossRef]
- Underhill SAM, Cabeen MT. Redundancy in Citrate and cis-Aconitate Transport in Pseudomonas aeruginosa. J. Bacteriol. 2022, 204, e0028422. [Google Scholar] [CrossRef]
- Mortera P, Pudlik A, Magni C, Alarcón S, Lolkema JS. Ca2+-citrate uptake and metabolism in Lactobacillus casei ATCC 334. Appl. Environ. Microbiol. 2013, 79, 4603–4612. [Google Scholar] [CrossRef]
- Pfenninger-Li XD, Dimroth P. NADH formation by Na(+)-coupled reversed electron transfer in Klebsiella pneumoniae. Mol. Microbiol. 1992, 6, 1943–1948. [Google Scholar] [CrossRef] [PubMed]
- Ramos A, Poolman B, Santos H, Lolkema JS, Konings WN. Uniport of anionic citrate and proton consumption in citrate metabolism generates a proton motive force in Leuconostoc oenos. J. Bacteriol. 1994, 176, 4899–4905. [Google Scholar] [CrossRef]
- Pudlik AM, Lolkema JS. Citrate uptake in exchange with intermediates in the citrate metabolic pathway in Lactococcus lactis IL1403. J. Bacteriol. 2011, 193, 706–714. [Google Scholar] [CrossRef]
- Sheldon JR, Marolda CL, Heinrichs DE. TCA cycle activity in Staphylococcus aureus is essential for iron-regulated synthesis of staphyloferrin A, but not staphyloferrin B: the benefit of a second citrate synthase. Mol. Microbiol. 2014, 92, 824–839. [Google Scholar] [CrossRef] [PubMed]
- Beaumont M, Portune KJ, Steuer N, Lan A, Cerrudo V, Audebert M, Dumont F, Mancano G, Khodorova N, Andriamihaja M, Airinei G, Tomé D, Benamouzig R, Davila AM, Claus SP, Sanz Y, Blachier F. Quantity and source of dietary protein influence metabolite production by gut microbiota and rectal mucosa gene expression: a randomized, parallel, double-blind trial in overweight humans. Am. J. Clin. Nutr. 2017, 106, 1005–1019. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic view of the metabolism of amino acids by the intestinal bacteria. Undigested dietary and endogenous proteins supplied by the host are degraded into amino acids by the bacterial proteases and peptidases. Amino acids can then be transported inside bacteria and serve together with the amino acid synthesized within the bacteria for the synthesis of proteins. The synthesis of RNAs and DNA requires specific amino acids while several amino acids can be utilized for ATP production and for the synthesis of bioactive metabolites. These latter amino acid-derived metabolites can exert effects on the metabolism and physiology of the producing bacteria (bacteria 1). The amino acids which cannot be synthesized by bacteria 1 can be considered as indispensable for these bacteria and thus need to be imported from the medium. Some among the bioactive metabolites generated by bacteria 1 can be exported in the medium and act on the metabolism and physiology of other bacterial species (such as bacteria 2). Of note, some amino acids synthesized by bacteria 1 can be exported in the media and used by bacteria 2 (and vice versa) thus indicating potential metabolic cooperation between different members of the intestinal bacterial community.
Figure 1.
Schematic view of the metabolism of amino acids by the intestinal bacteria. Undigested dietary and endogenous proteins supplied by the host are degraded into amino acids by the bacterial proteases and peptidases. Amino acids can then be transported inside bacteria and serve together with the amino acid synthesized within the bacteria for the synthesis of proteins. The synthesis of RNAs and DNA requires specific amino acids while several amino acids can be utilized for ATP production and for the synthesis of bioactive metabolites. These latter amino acid-derived metabolites can exert effects on the metabolism and physiology of the producing bacteria (bacteria 1). The amino acids which cannot be synthesized by bacteria 1 can be considered as indispensable for these bacteria and thus need to be imported from the medium. Some among the bioactive metabolites generated by bacteria 1 can be exported in the medium and act on the metabolism and physiology of other bacterial species (such as bacteria 2). Of note, some amino acids synthesized by bacteria 1 can be exported in the media and used by bacteria 2 (and vice versa) thus indicating potential metabolic cooperation between different members of the intestinal bacterial community.
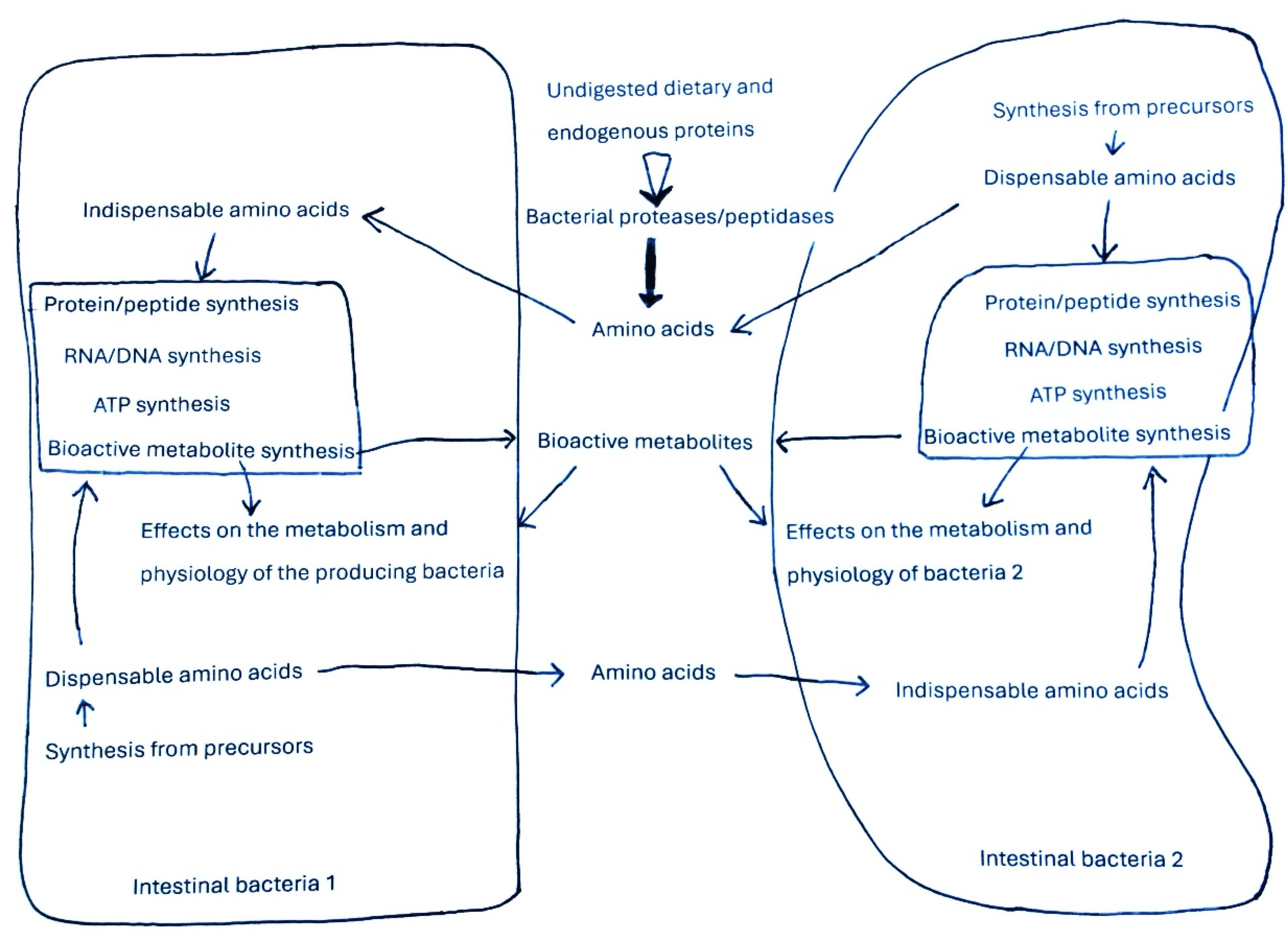
Figure 2.
Schematic view of the metabolism of polyamines by the intestinal bacteria. Arginine is the precursor for the synthesis of agmatine, while agmatine and ornithine are the precursors of putrescine. Putrescine and the methionine-derived metabolite decarboxylated S-adenosylmethionine are the precursors for the synthesis of spermidine, and to a lower extent of spermine. Lysine is the precursor of cadaverine. The concentrations of polyamines within bacteria are the net result of synthesis, degradation, uptake, and release in the medium. Polyamines can exert effects on the metabolism and physiology of the producing bacteria. Polyamines released in the medium can be taken up by other bacterial species where it can exert metabolic and physiological effects.
Figure 2.
Schematic view of the metabolism of polyamines by the intestinal bacteria. Arginine is the precursor for the synthesis of agmatine, while agmatine and ornithine are the precursors of putrescine. Putrescine and the methionine-derived metabolite decarboxylated S-adenosylmethionine are the precursors for the synthesis of spermidine, and to a lower extent of spermine. Lysine is the precursor of cadaverine. The concentrations of polyamines within bacteria are the net result of synthesis, degradation, uptake, and release in the medium. Polyamines can exert effects on the metabolism and physiology of the producing bacteria. Polyamines released in the medium can be taken up by other bacterial species where it can exert metabolic and physiological effects.
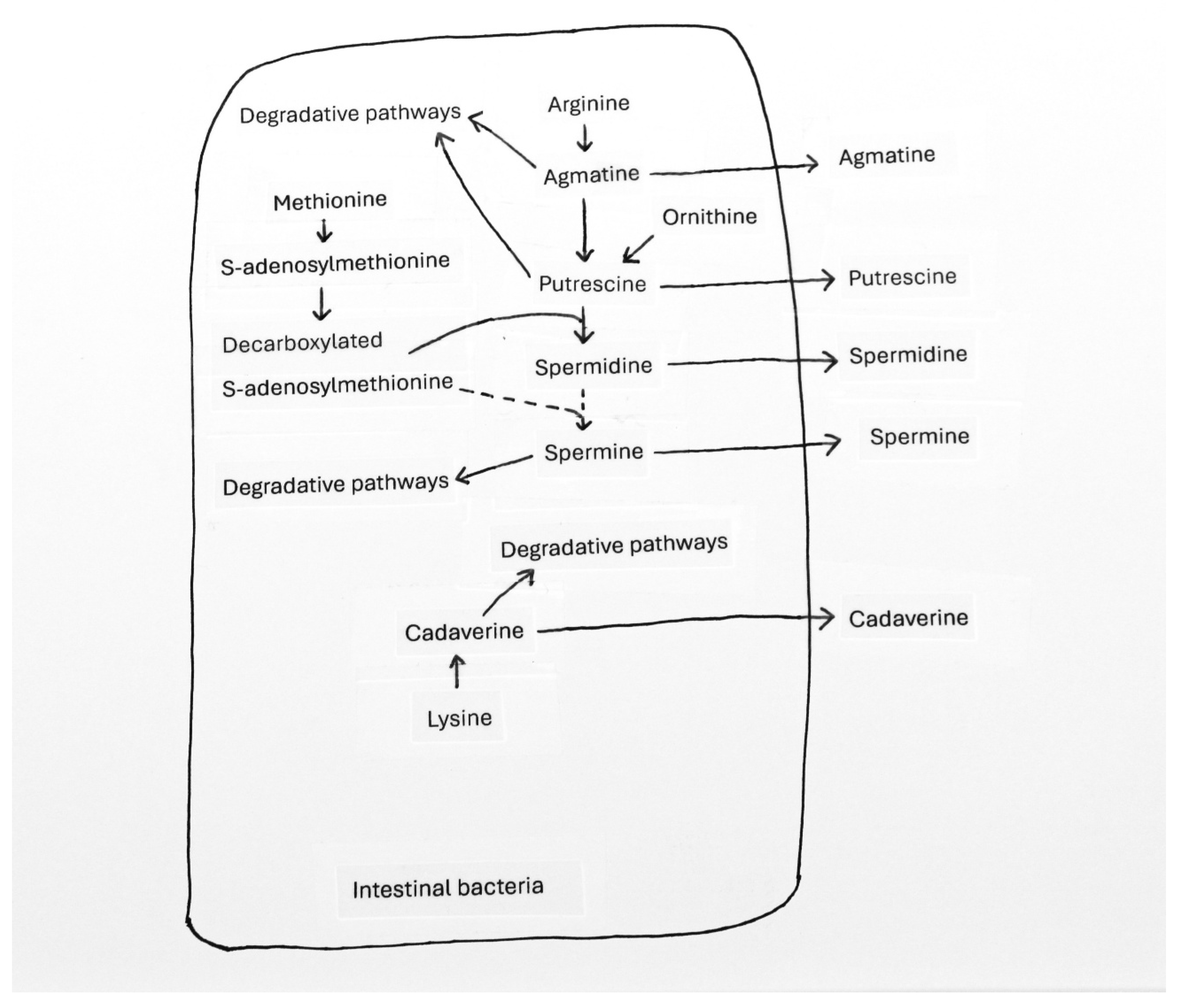
Figure 3.
Schematic view of the metabolism of hydrogen sulfide by the intestinal bacteria. Hydrogen sulfide (H2S) can be synthesized in bacteria mainly from cysteine and sulfate. H2S can affect the metabolism and physiology of the producing bacteria. H2S can diffuse from the producing cells and act on the metabolism and physiology of other bacteria (bacteria 2). H2S can be involved as precursor in the synthesis of cysteine and methionine in the bacteria equipped with the corresponding anabolic pathways.
Figure 3.
Schematic view of the metabolism of hydrogen sulfide by the intestinal bacteria. Hydrogen sulfide (H2S) can be synthesized in bacteria mainly from cysteine and sulfate. H2S can affect the metabolism and physiology of the producing bacteria. H2S can diffuse from the producing cells and act on the metabolism and physiology of other bacteria (bacteria 2). H2S can be involved as precursor in the synthesis of cysteine and methionine in the bacteria equipped with the corresponding anabolic pathways.
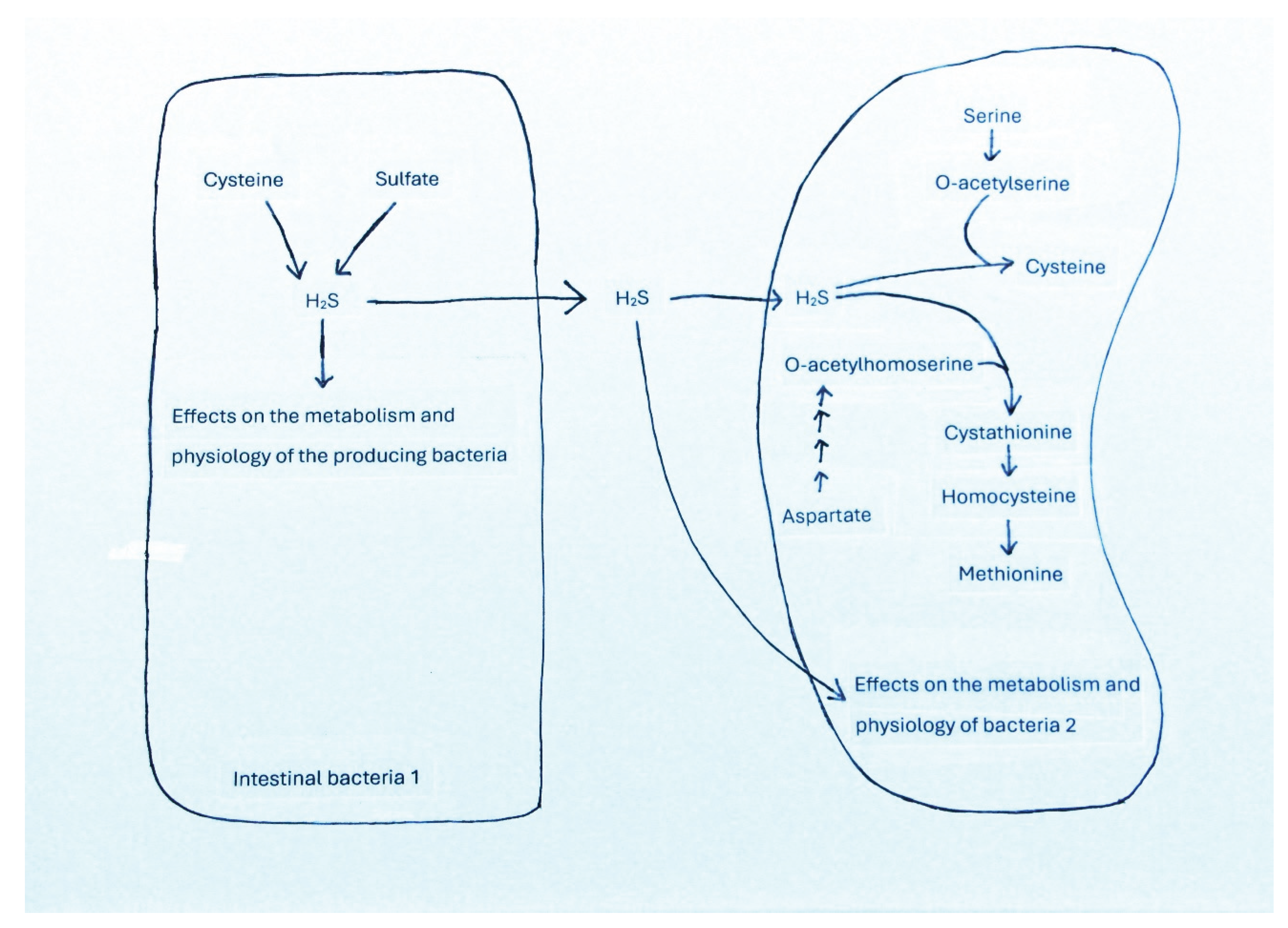
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



