
Submitted:
21 October 2025
Posted:
22 October 2025
You are already at the latest version
Abstract
Fungal pathogens represent a major constraint to global agricultural productivity, causing a wide range of plant diseases that severely affect staple crops such as cereals, legumes, and vegetables. These infections result in substantial yield losses, deterioration of grain and produce quality, and significant economic impacts across the entire agri-food sector. Among phytopathogens, fungi are considered the most destructive, causing a wide range of diseases such as powdery mildew, rusts, fusarium head blight, smut, leaf spot, rots, late blight, and other fungal pathogens. Traditional plant protection methods do not always provide long-term effectiveness and environmental safety, which requires the introduction of innovative approaches to creating sustainable varieties. CRISPR-Cas (Clustered Regularly Interspaced Short Palindromic Repeats) technology opens up new opportunities for targeted genome editing, allowing the modification or silencing of susceptibility genes and thus increasing plant resistance to fungal infections. This review presents current achievements and prospects for the application of CRISPR-Cas technology to increase the resistance of major agricultural crops to fungal diseases. The implementation of these approaches contributes to the creation of highly productive and resistant varieties, which is crucial for ensuring food security in the context of climate change.
Keywords:
CRISPR-Cas
; fungal diseases
; main agricultural crops
; cereals
; legumes
; vegetables
1. Introduction
Plant diseases are one of the most significant threats to agriculture and global food security. Given that the current world population of 7.6 billion is expected to reach 8.6 billion by 2030, 9.8 billion by 2050, and 11.2 billion by 2100, combating crop diseases is crucial to ensuring food security and maintaining sufficient food production to meet growing global demand [1]. According to data from the Food and Agriculture Organization of the United Nations [2], diseases and pests cause 20–40% of global crop losses each year, equivalent to billions of dollars in economic losses. The economic impact of crop diseases goes far beyond direct crop losses. In countries where agriculture is the main source of income and employment, especially in developing countries, disease outbreaks cause catastrophic damage, particularly to small farmers, who often do not have access to modern plant protection products. These epiphytotics cause food prices to rise, disrupt trade chains, and provoke food shortages, exacerbating the problems of hunger and malnutrition [3]. Plant diseases also pose a serious obstacle to the implementation of the Zero Hunger program, as they cause losses of up to 40% of the global harvest each year [4], directly threatening food security and contributing to rising levels of hunger.
Diseases caused by fungal pathogens pose a serious threat, affecting major agricultural crops—cereals, legumes, and vegetables—and leading to significant crop losses and reduced product quality. Among plant diseases, fungal pathogens are considered the most destructive, causing a wide range of infections in major agricultural crops. In cereal crops, the most common fungal diseases are powdery mildew, rusts (leaf, stem, and yellow), fusarium head blight, smut, and helminthosporium root rot [5]. Legumes are often affected by fungi of the genera Ascochyta and Rhizoctonia, which cause ascochyta blight and fusarium wilt [6]. Vegetable crops suffer from a variety of fungal diseases, including powdery mildew, late blight, leaf spot, anthracnose, and various root rots [7].
Currently, the main strategy for combating plant diseases remains the use of chemical pesticides. However, the use of pesticides often has direct and indirect negative effects on the environment and other living organisms [8]. Moreover, due to the widespread and uncontrolled use of these substances, phytopathogens are gradually developing resistance to them [9]. Reducing the negative consequences of the chemicalization of agricultural systems requires the search for alternative solutions. One of the most effective and environmentally friendly approaches is the creation of disease-resistant varieties through plant breeding [10,11]. Traditional breeding, despite its past successes, faces a number of limitations: the complexity of introgression of resistance genes (R), limited genetic variability, the transfer of undesirable traits along with target traits, and the labor-intensive nature of the process [12,13]. In addition, the pace of traditional breeding does not always keep up with the rate of pathogen evolution and growing global demand for food, especially in the context of global climate change [14].
Alternative approaches to plant breeding, such as mutation breeding and transgenesis [15,16] are also used, but they have limitations that reduce their effectiveness and public acceptance (Figure 1).
Achieving the goal of ending hunger by 2030 requires comprehensive approaches, including the development of disease-resistant crops using methods such as CRISPR-Cas (Clustered Regularly Interspaced Short Palindromic Repeats), and the implementation of sustainable disease and pest control strategies that enable the precise, rapid, and reproducible creation of genetically improved lines [17]. The integration of traditional breeding and genetic engineering using cloned R genes provides a sustainable pathway to creating crop varieties with a broad spectrum of disease resistance; the introduction of such genes effectively enhances their biotic resistance [18]. CRISPR-Cas technology is based on the system's ability to precisely target specific regions of the genome to create desired mutations (knockouts or knock-ins). This process is facilitated by a guide RNA (sgRNA), which recognizes the target region, and the Cas9 endonuclease, which cuts the DNA at the specified position. Modern genome editing platforms allow increasing plant resistance to diseases in various ways: by knocking out susceptibility genes (S-genes), replacing alleles through homology-directed repair, introducing resistance genes (R-genes), modifying the regulation of R/S gene expression, and through multiplex editing targeting multiple resistance and susceptibility factors simultaneously [19,20,21].
This review summarizes current and promising achievements that open up new opportunities for applying CRISPR-Cas technology to increase the resistance of major agricultural crops (cereals, legumes, and vegetables) to fungal diseases. This technology allows targeted modification or silencing of susceptibility genes, providing an effective and environmentally safe alternative to traditional plant protection methods. The use of CRISPR-Cas paves the way for the creation of sustainable varieties with high resistance and yield, which is key to ensuring global food security in the face of climate change.
2. CRISPR–Cas Genome Editing to Improve Fungal-Disease Resistance in Main Cereal Crops
2.1. Wheat
Wheat (Triticum aestivum L.) is one of the most important food crops, providing a significant portion of global grain consumption. However, its production faces growing threats related to climate change, the spread of new pathogens, and the declining effectiveness of traditional plant protection methods [22,23,24]. Previous studies have identified 33 susceptibility genes (S-genes) that act as negative regulators of resistance. It is proposed to use the CRISPR-Cas system to modify these S-genes, which will allow the creation of mutant lines with increased resistance to biotic stress and enrich the genetic resources of wheat for further selection [25].
Fusarium head blight (FHB), caused mainly by Fusarium graminearum, is a serious threat to global wheat production [26]. The most studied resistance locus, Fhb1, first discovered in Chinese germplasm, provides partial protection against FHB. The key susceptibility gene associated with this locus is TaHRC, which encodes a histidine-rich calcium-binding protein (Table 1). Its deletion affecting the start codon (TaHRC-S) confers resistance to infection [27]. The identity of TaHRC-R alleles in different genotypes indicates a single origin of the resistant allele, presumably associated with the Dahongpao haplotype. For targeted editing of TaHRC, a gRNA delivery system based on barley mosaic virus (BSMV) was developed in the varieties ‘Bobwhite’ and ‘Everest’ [28]. Additionally, TaHRC susceptibility genes were successfully edited using the CRISPR-Cas9 system delivered through interspecific hybridization of wheat with corn expressing Cas9 and gRNA. Among plants regenerated from haploid embryos in vitro, mutations at the target loci were found in 15–33% of samples, demonstrating the effectiveness of this approach [29].
The mycotoxin deoxynivalenol (DON) is of particular importance as a key factor in the virulence of F. graminearum. When comparing infection with the wild type and the Δtri5 mutant, which does not produce DON, approximately 400 differentially expressed genes activated by the mycotoxin were identified. Among them, TaNFXL1 was identified, whose expression level is higher in susceptible genotypes. Functional analysis using virus-induced gene silencing (VIGS) and CRISPR confirmed that TaNFXL1 is a negative regulator of resistance, and its knockout increases plant resistance to FHB [30]. Thus, targeting the TaHRC and TaNFXL1 genes using CRISPR-Cas is a promising strategy for increasing wheat resistance to fusarium head blight and reducing DON accumulation in grain.
Another necrotrophic pathogen, Parastagonospora nodorum, causes wheat Septoria nodorum blotch (SNB), which leads to significant yield losses and grain quality deterioration [31]. To edit the Tsn1 and Snn5 susceptibility genes to P. nodorum, Poddar et al. developed an approach based on the use of the Cas9–RNP (ribonucleoprotein) complex, which allows for the effective selection of gRNA and editing in wheat protoplasts and immature embryos without the introduction of exogenous DNA [32]. This method provides heritable mutations without the need for transgenic plant selection, opening up new opportunities for accelerated selection of resistant varieties. In addition, P. nodorum protoplasts were transformed with a Cas9–sgRNA complex targeting the Tox3 gene, which resulted in 100% editing efficiency. When co-transformed with an HDR cassette containing 1 kb homologous flanks, the frequency of homologous recombination exceeded 70%, and when microhomologies (50 bp) were used, it reached 25%. These results confirm the high accuracy and versatility of CRISPR-Cas9 technology for editing the P. nodorum genome [33].
Powdery mildew of wheat, caused by the obligate biotrophic fungus Blumeria graminis f. sp. tritici, is one of the most devastating diseases of the crop, causing up to 5% of global wheat yield losses annually [34]. Loss-of-function mutations in the Powdery Mildew Locus O (MLO) gene provide broad resistance to the disease but are often accompanied by reduced yield. Li et al. described the Tamlo-R mutant line with a 304 kb deletion in the MLO-B1 locus, characterized by high resistance while maintaining productivity. The deletion alters the chromatin structure and activates the tonoplast transporter TaTMT3B, which compensates for the negative effects of MLO knockout ([35]. Similar results were obtained when the TaEDR1 gene was suppressed by virus-induced silencing or RNA interference (RNAi), which increased wheat resistance to powdery mildew, indicating the role of TaEDR1 as a negative regulator of resistance. No off-target changes were detected in the TaEDR1 mutants obtained. Such plants showed resistance to powdery mildew and did not exhibit pathogen-induced cell death [36]. Knockout of homologues of this gene in wheat (TaMLO) using the CRISPR-SpCas9 system led to the formation of resistance to the powdery mildew pathogen B.tritici [37]. Thus, editing the MLO and TaEDR1 genes using CRISPR-Cas accelerates the introduction of resistance alleles into elite varieties, ensuring the creation of productive and resistant crops and providing valuable germplasm for further breeding.
Wheat rust (yellow, leaf, and stem rust) is one of the main causes of reduced yields in this crop. Yellow (Puccinia striiformis f. sp. tritici) and leaf (Puccinia triticina Erikss) rusts are among the most widespread and dangerous diseases of wheat and are the main factor negatively affecting wheat yield and quality, causing significant economic damage [38,39,40]. A study by Liu et al. showed that suppression of the TaMKP1 gene using CRISPR-Cas9 increases wheat resistance to yellow rust and powdery mildew, as well as increasing yield without infection. It has been established that TaMKP1 interacts directly with activated TaMPK3/4/6, and TaMPK4 interacts with TaPAL, indicating its role as a negative modulator of MAPK signaling [41]. Wang et al. identified the TaPsIPK1 gene, which encodes a cytoplasmic kinase-like receptor that contributes to susceptibility to P.tritici. The fungal effector PsSpg1 binds to TaPsIPK1, enhancing its kinase activity and inducing phosphorylation of the transcription factor TaCBF1d. Inactivation of TaPsIPK1 via CRISPR-Cas9 provides broad-spectrum resistance to the pathogen without negatively affecting agronomic traits, as confirmed by two years of field trials [42]. In addition, Liu et al. demonstrated that the TaGW2 gene, known as a negative regulator of grain weight, also reduces wheat resistance to leaf rust by ubiquitinating the TaSGT1 protein. Editing TaGW2 using CRISPR-Cas9 simultaneously increased grain weight and resistance to leaf rust disease [43].
Recent studies show that the CRISPR-Cas system is a powerful tool for targeted editing of wheat susceptibility genes and creating resistant lines without compromising yield. Modification of key genes such as Fhb1, TaHRC, and TaNFXL1 increases wheat resistance to fusarium head blight, while loss of function of Tsn1, Snn5, and Tox3 reduces susceptibility to SNB. Mutations in the MLO, TaEDR, and TaMKP1 genes provide broad resistance to powdery mildew, while changes in TaMKP1, TaGW2, and TaPsIPK1 contribute to increased resistance to leaf and yellow rust, respectively. The combination of CRISPR-Cas with VIGS, protoplast and RNP editing methods, as well as interspecific hybridization, opens up new opportunities for accelerated and accurate selection. These approaches not only enable the development of resistant and high-yielding varieties, but also form the scientific basis for the precise control of wheat immunity, expanding its genetic diversity and resistance to biotic stresses.
2.2. Rice
Rice (Oryza sativa L.) is one of the world's most important cereal crops, ranking third in terms of production volume after corn and wheat, and serving as the staple food for nearly half of the world's population. Rice plants are affected by a variety of diseases caused by fungi, bacteria, viruses, and nematodes, which attack various parts of the plant and significantly reduce yields. Among fungal pathogens, the most dangerous and widespread are rice blast disease (Magnaporthe oryzae) and rice false smut (Ustilaginoidea virens), which affect plants at all stages of development and manifest themselves in the form of lesions on the leaves, neck, and panicle [44].
M. oryzae, the causative agent of rice blast disease, is one of the most destructive pathogens of rice. To accelerate the editing of its genome, a plasmid-free CRISPR-Cas9 method has been developed that uses ribonucleoprotein (RNP) Cas9–sgRNA complexes. Unlike toxic plasmid expression of Cas9, the RNP approach provides high accuracy and efficiency, including point substitutions and multiple mutations in the ALB1 and RSY1 genes, and allows the production of isogenic strains that do not contain foreign DNA [45]. In rice, OsERF922, which encodes the AP2/ERF transcription factor, regulates plant defense. Its knockout via CRISPR-SpCas9 increased resistance to M. oryzae without affecting growth and yield [46]. Similarly, a mutation in OsSEC3A, a subunit of the exocyst, caused activation of defense pathways and dwarfism, enhancing resistance to rice blast disease [47]. Wang et al. used the Mo_tRNAArg24-gRNA-Cas9 cassette to significantly improve gene editing efficiency in M. oryzae. When tested on five genes (Rei1, Ppg1, Bip1, Bip2, Dbf2) located on different chromosomes, the Mo_tRNAArg24-gRNA-Cas9 cassette demonstrated high gene editing efficiency in M. oryzae (66.7–100%) without Cas9 toxicity, including two previously uncharacterized genes, with a mutation frequency of 66.7–100% without Cas9 toxicity [48].
Functional knockout of Pi21 in the Nanjing 9108 variety [49] and co-editing of Pita, Pi21, and ERF922 (vector pC1300-2×35S:: Cas9-gPita-gPi21-gERF922) in line L1014 resulted in high mutation frequencies (75–85%) and a significant increase in resistance to rice blast disease [50]. Multiplex editing of TMS5, Pi21, and Xa13 created non-transgenic triple mutants tms5/pi21/xa13 that are resistant to blast and bacterial blight, demonstrating the potential of hybrid rice breeding [51]. The CRISPR-Cas9 system targeting Pi21 and OsSULTR3;6 in the susceptible variety 58B achieved >90% editing efficiency and created lines with increased resistance to rice blast and bacterial leaf streak while maintaining yield [52]. Similarly, mutations in Bsr-d1, Pi21, and ERF922 in the Longke638S line contributed to double resistance to blast and bacterial blight diseases without reducing productivity [53]. In the Dalixiang variety, Pi21 knockout increased resistance without altering grain quality [54], and editing Pid3 using A. tumefaciens yielded nine homozygous pid3 mutants without the Cas9 locus, demonstrating a similar effect [55].
The bsr-d1 allele, encoding a transcription factor from the Digu variety, provides broad resistance to M. oryzae by suppressing H₂O₂ degradation. Knocking out Bsr-d1 induced redox status, amino acid and fatty acid metabolism, and salicylic acid signaling pathways, and identified a new target gene, Perox3, which reduces resistance when overexpressed [56]. Recently, Zhang et al. developed dCas9-based CRISPR interference (CRISPRi) to suppress genes of the fungal pathogens M. oryzae and U. virens. The use of gRNAs targeting the MoATG3, MoATG7, and UvPal1 promoters showed that a single gRNA can effectively reduce expression and induce expected phenotypes, making CRISPRi a powerful tool for functional genomics of pathogens [57].
In addition to the knockout strategy, a promising approach is precise base editing, which allows for the correction of defective alleles of resistance genes (R-genes). Precise base editing also opens up possibilities for the correction of R-genes. For example, Ren et al. used the cytidine editor rBE5 to correct a single nucleotide substitution (G>A) in the recessive allele pi-d2, restoring resistance to rice blast disease [58].

Table 1.
Using CRISPR-Cas technology to improve fungal-disease resistance in cereals.
| Pathogen | Target Gene | Delivery Method | Type of Editing | References |
| Wheat | ||||
| Fusarium graminearum | Fhb1 | Particle bombardment | deletion mutation | [27] |
| Fusarium graminearum | TaHRC |
Agrobacterium- mediated transformation |
BSMV-mediated gene editing | [28] |
| Fusarium graminearum | TaHRC |
Agrobacterium- mediated transformation |
Knockout | [29] |
| Fusarium graminearum | TaNFXL1 |
Agrobacterium- mediated transformation |
virus-induced gene silencing | [30] |
| Parastagonospora nodorum | Tsn1 and Snn5 | Cas9-RNP mediated editing | Knockout | [32] |
| Parastagonospora nodorum | Tox3 | ribonuclear protein (RNP) complex targeting | Knockout | [33] |
| Blumeria graminis f. sp. tritici | MLO-B1 | Cas9-RNP mediated editing | Deletion | [35] |
| Blumeria graminis f.sp. tritici | TaMLO | Particle bombardment | Knockout | [37] |
| Blumeria graminis f. sp. tritici | TaEDR1 | Particle bombardment | Knockout | [36] |
|
Blumeria graminis f. sp. tritici Puccinia striiformis f. sp. tritici |
TaMKP1 |
Agrobacterium- mediated transformation |
Knockout | [41] |
| Puccinia striiformis f. sp. tritici | TaPsIPK1 | Agrobacterium- | Knockout | [42] |
| Puccinia triticina Erikss | TaGW2 | mediated transformation | Knockout | [43] |
| Rice | ||||
| Magnaporthe oryzae | ALB1 and RSY1 | RNP targeting | RSY1 | [45] |
| Magnaporthe oryzae | OsERF922 | Protoplast transformation | Mutation | [46] |
| Magnaporthe oryzae | OsSEC3A | Protoplast transfection | Mutation | [47] |
| Magnaporthe oryzae | Rei1, Ppg1, Bip1, Bip2, Dbf2 | Protoplast transformation | Mutation | [48] |
| Magnaporthe oryzae | Pi21 |
Agrobacterium- mediated transformation |
Knockout | [49] |
| Magnaporthe oryzae | Pita, Pi21 and ERF922 |
Agrobacterium- mediated transformation |
Knockout | [50] |
| Magnaporthe oryzae | TMS5,Pi21 иXa13 |
Agrobacterium- mediated transformation |
Mutation | [51] |
| Magnaporthe oryzae | Pi21 иOsSULTR3;6 |
Agrobacterium- mediated transformation |
Knockout | [52] |
| Magnaporthe oryzae | Bsr-d1, Pi21иERF922 |
Agrobacterium- mediated transformation |
Knockout | [53] |
| Magnaporthe oryzae | Pi21 |
Agrobacterium- mediated transformation |
Knockout | [54] |
| Magnaporthe oryzae | Pid3 |
Agrobacterium- mediated transformation |
Knockout | [55] |
| Magnaporthe oryzae | Bsr-d1, Perox3 |
Agrobacterium- mediated transformation |
Knockout | [56] |
| Magnaporthe oryzae, Ustilaginoidea virens | MoATG3,MoATG7 иUvPal1 |
Agrobacterium- mediated transformation |
Knockout | [57] |
| Magnaporthe oryzae | rBE5 |
Agrobacterium- mediated transformation |
Base editing | [58] |
Recent studies show that CRISPR-Cas technology has significantly accelerated the development of disease-resistant rice varieties. Knockout of key susceptibility genes (Pi21, Bsr-d1, OsERF922, Pid3, etc.) has provided effective protection against M. oryzae without reducing yield and grain quality. The development of improved systems, such as plasmid-free RNP complexes and Mo_tRNAArg24-gRNA-Cas9-type cassettes, has increased the accuracy and efficiency of editing, while the emergence of CRISPRi and base editing has opened up opportunities for fine-tuning the expression and restoring the functionality of R-genes. These advances form the basis for precision breeding of rice and the creation of sustainable, productive, and environmentally safe varieties.
2.3. Barley
Barley (Hordeum vulgare L.) is one of the most important cereal crops, widely used in brewing, feed production, and the food industry [59]. Barley production is limited by the impact of various pathogens that cause fungal, bacterial, and viral diseases. The most common diseases include dwarf rust, powdery mildew, fusarium, and various forms of smut [60].
The use of CRISPR-Cas technology opens up broad prospects for increasing barley's resistance to disease through targeted editing of genes involved in the formation of the plant's natural defense mechanisms against pathogens.
The efficiency of gene editing is significantly increased when Cas9/sgRNA is used in the form of ribonucleoprotein (RNP) complexes that induce double-strand breaks in DNA at target loci. Gene replacement or inactivation can be achieved by combining Cas9/sgRNA RNP with a PCR-amplified selective marker containing short homologous flanks of the barley target gene for fungal transformation. This approach eliminates the need for labor-intensive vector construction procedures and ensures high precision of targeted knockout [61]. Colonization of plants by fungal pathogens is a complex multistep process involving numerous genes in both the host plant and the pathogen. In recent years, particular attention has been paid to the study of susceptibility genes (S-genes), which are negative regulators of plant defence mechanisms. One of the best-known examples is the MLO locus, first described in barley as a susceptibility factor to powdery mildew [62].
Research on editing the barley genome using CRISPR-Cas technology is actively developing, but most of it focuses on improving key agronomic traits—yield, drought resistance, starch content, and disease resistance. Cas technology is actively developing, but most of it is focused on improving key agronomic traits—yield, drought tolerance, starch and protein content—as well as creating resistance to viral infections, including barley yellow dwarf virus (BYDV) and mosaic virus. At the same time, there's still not much work being done on making barley more resistant to fungal pathogens like rust, powdery mildew, fusarium, and smut. This is due to the complexity of molecular interactions in the plant-pathogen system and the insufficient study of susceptibility genes in barley. Nevertheless, given the successful application of CRISPR-Cas technology for editing similar S-genes in wheat and rice, the use of this approach to target homologous genes in barley is a promising direction for increasing the crop's resistance to fungal diseases and expanding its genetic potential.
3. CRISPR-Cas Genome Editing for Improvement Fungal Disease Resistance of Legumes
Soybean (Glycine max L.) is one of the most economically important leguminous crops, grown and consumed worldwide as a source of protein and oil for animal feed and human nutrition [63]. However, global soybean production is seriously threatened by many diseases, one of which is powdery mildew caused by the obligate biotrophic fungus Erysiphe diffusa [64].
Bui et al. developed CRISPR-Cas9 constructs with dual sgRNA, which were successfully transferred to the Vietnamese soybean variety DT26 through transformation using A. tumefaciens (Table 2). Various mutant forms of the GmMLO genes, including biallelic, chimeric, and homozygous, were detected in the T0 generation. The inheritance and segregation of CRISPR-Cas9-induced mutations were confirmed and validated in the T1 and T2 generations. From the six GmMLO genes in the soybean genome, the authors obtained triple knockout mutants Gmmlo02/Gmmlo19/Gmmlo23 and quadruple knockout mutants Gmmlo02/Gmmlo19/Gmmlo20/Gmmlo23 in the T2 generation. When infected with Erysiphe diffusa, a fungus that causes powdery mildew in soybeans, all mutant plants showed increased resistance to the pathogen, especially the quadruple mutant. The degree of powdery mildew infection in mutant soybeans was reduced by 36.4% compared to wild-type plants. In addition, no pleiotropic effects on the growth and development of soybeans were observed in CRISPR-Cas9 mutants under greenhouse conditions [65].
In another study, the CRISPR-Cas9 system was successfully introduced into the oomycete pathogen Phytophthora sojae, enabling rapid and efficient genome editing. Using the RXLR Avr4/6 effector gene as a target, the researchers observed that in the absence of a homologous template, double-strand breaks (DSBs) induced by Cas9 were repaired by non-homologous end joining (NHEJ), which mainly resulted in short insertions or deletions (indels). The Avr4/6 gene in P. sojae encodes a single RXLR-dEER effector protein of 123 amino acids, which determines avirulence in soybean plants carrying the Rps4 or Rps6 resistance genes. Initially thought to be two separate genes, Avr4 and Avr6 turned out to be a single locus, Avr4/6, located in a 24-kilobyte region of the genome [66].
Table 2.
Using CRISPR-Cas technology to improve fungal-disease resistance in legumes.
| Pathogen | Target Gene | Delivery Method | Type of Editing | References |
| Soybean | ||||
| Erysiphe diffusa | GmMLO02, GmMLO19, GmMLO20, GmMLO23 | Agrobacterium-mediated transformation | Knockout | [65] |
| Phytophthora sojae | RXLR Avr4/6 | (PEG)-mediated protoplast transformations | Knockout | [67] |
The authors note that most mutants were homozygous, presumably as a result of gene conversion caused by Cas9-mediated cleavage of non-mutant alleles. In the presence of donor DNA, homologous repair (HDR) was observed, which led to the replacement of Avr4/6 with the NPT II genome. By testing the specific virulence of several NHEJ mutants and HDR-mediated gene replacements in soybean, we confirmed the contribution of Avr4/6 to recognition by the soybean R gene loci Rps4 and Rps6, but also found an additional contribution of these two loci to resistance [67].
These studies demonstrate the effectiveness of CRISPR-Cas9 technology in enhancing soybean resistance to powdery mildew and investigating the mechanisms of P. sojae pathogenicity. Targeted editing of host susceptibility genes (GmMLO) and pathogen effectors (Avr4/6) provides valuable information and practical tools for creating disease-resistant soybean varieties.
4. CRISPR-Cas Genome Editing for Improvement Fungal Disease Resistance of Vegetables
4.1. Tomato
Tomato (Solanum lycopersicum L.) is one of the world's most important vegetable crops. According to data from the Food and Agriculture Organization of the United Nations (FAO) as of September 2025, it is cultivated on an area of 5.41 million hectares, with a total production of 192 million tons and an average yield of 35.5 tons per hectare. Pathogens establish compatible interactions by exploiting host factors encoded by plant susceptibility genes (S genes). Data indicate that inactivating plant S genes can confer broad-spectrum and persistent resistance [68].
Powdery mildew is a widespread fungal disease that causes significant economic damage to crops such as wheat, barley, and tomatoes, mainly in temperate climates [69]. It has been found that plants resistant to powdery mildew carry recessively inherited mutations with loss of function in the MLO gene [70][. In MLO mutant plants, powdery mildew pathogenicity is arrested at the stage of cell wall penetration and host cell invasion. As a result, fungal sporophytes cannot form haustoria in host cells, and fungal colonies do not develop. Thus, this gene has become the subject of interest in many studies on genome editing to obtain resistance to fungal diseases [71].
One of the first studies on the use of the CRISPR-Cas system with a knocked-out MLO gene for tomatoes was conducted in 2017 by the team of Nekrasov et al. In the study, the authors report the creation of the Tomelo tomato variety, which does not contain transgenic elements and is resistant to the fungal pathogen powdery mildew, using CRISPR-Cas9 technology in less than ten months (Table 3). The study also performed whole-genome sequencing to show that the resulting mutant line does not contain any foreign DNA sequences, but only has a deletion that is indistinguishable from natural mutations. The group also presented evidence that CRISPR-Cas9 is a highly accurate tool, as no off-target mutations were found in Tomelo. Thus, it was shown that mutations can be easily introduced into elite or locally adapted tomato varieties in less than a year with relatively minimal effort and investment [72].
Another group of researchers also obtained knockout lines for the SlMlo1 gene, demonstrating complete resistance to powdery mildew fungus. The results demonstrate the effectiveness of the CRISPR-Cas9 system for introducing targeted mutagenesis to rapidly develop pathogen-resistant tomato varieties, which is consistent with the results of a previous study. The study used the CRISPR-Cas9 system to obtain SlMlo1 knockout mutants by transforming tomatoes with Agrobacterium in the elite tomato line BN-86. The authors succeeded in creating SlMlo1 knockout mutant lines that demonstrated complete resistance to the fungal disease powdery mildew [73].
Another gene discovered in Arabidopsis thaliana mutants resistant to powdery mildew through screening was named “powdery mildew resistance” (PMR). These mutants demonstrated resistance to G. cichoracearum and G. orontii [88]. Santillán Martínez et al. used the ortholog of the PMR gene in tomato to test the resistance of new mutant lines to Oidium neolycopersici, a powdery mildew pathogen. They used a CRISPR-Cas9 construct containing four single-guide RNAs (sgRNAs) targeting the tomato PMR4 gene to increase the likelihood of large deletions in mutants. After selection based on PCR and sequencing of transformants, five different mutational events were identified, including deletions ranging from 4 to 900 bp, a 1 bp insertion, and an 892 bp inversion. All of these mutants showed reduced susceptibility to O. neolycopersici based on visual assessment of disease symptoms and quantitative assessment of relative fungal biomass [74].
Previous studies on potatoes have shown that disruption of DND1 (Defense No Death 1) function increases plant resistance to various pathogens, such as powdery mildew (PM) O. neolycopersici, but this comes at the cost of negative effects on overall plant health and viability, such as the expression of a severe dwarf phenotype, autonecrosis, and reduced male fertility [89].
To explore the possibility of minimizing the negative effects of the DND1 mutation while simultaneously increasing disease resistance, Li et al. developed a CRISPR-Cas9 construct with four single-target RNAs targeting three exons of SlDND1 (Solyc02g088560.4.1), which was introduced into the tomato cultivar Moneymaker (MM) via transformation with Agrobacterium tumefaciens. Three T1 lines (named E1, E3, and E4) were crossed with MM and then self-pollinated to produce TF2 families. All TF2 plants in the homozygous dnd1/dnd1 state showed a reduction in PM symptoms compared to heterozygous (DND1/dnd1) and wild-type (DND1/DND1) plants. Two complete knockout mutants (E1 and E4), encoding truncated DND1 proteins, showed clear phenotypes of dwarfism and autonecrosis, while the E3 mutant, carrying a deletion of 3 amino acids, showed normal height growth with fewer autonecrotic spots. Analysis of the three-dimensional structures of both the reference and mutant proteins revealed significant conformational changes in the protein derived from E3, which could potentially affect its function. The dnd1/dnd1 TF2 (TV181848-9, E3) was subjected to whole-genome sequencing using Illumina technology, which confirmed the absence of off-target mutations in selected regions of the genome. In addition, no traces of the Cas9 gene were found, indicating its elimination by segregation [75].
Another group, Li et al., also used the SlPMR4 gene in their study. A CRISPR–Cas9 vector containing four single-stranded RNA guides (sgRNA: sgRNA1, sgRNA6, sgRNA7, and sgRNA8) targeting the same number of SlPMR4 regions, was introduced by transformation using A. tumefaciens into two widely grown Italian tomato varieties: San Marzano (SM) and Oxheart (OX). Thirty-five plants (26 SM and 9 OX) were selected and analyzed to identify CRISPR-Cas9-induced mutations. Different sgRNAs caused mutations with a frequency ranging from 22.1 to 100% and, depending on the type, precise insertions (sgRNA6) or deletions (sgRNA7, sgRNA1, and sgRNA8). sgRNA7 induced a −7 bp deletion in seven SM genotypes in the homozygous state, while sgRNA8 resulted in fifteen SM genotypes with a biallelic mutation (−7 bp and −2 bp). Selected edited lines were inoculated with P. infestans, and four of them, completely knocked out at the PMR4 locus, showed a reduction in disease symptoms (decrease in susceptibility from 55 to 80%) compared to control plants. Four SM lines were sequenced using Illumina whole-genome sequencing for deeper characterization without showing any signs of mutations in candidate off-target regions. Results demonstrated for the first time a reduction in susceptibility to Phytophtora infestans in pmr4 tomato mutants, confirming the role of PMR4 knockout in providing broad-spectrum protection against pathogens [76].
MicroRNAs (miRNAs) are small non-coding RNAs 20–24 nucleotides (n.p.) in length that are processed from MIRNA genes transcribed by RNA polymerase II. In plants, the primary transcript (pri-miRNA) of the MIRNA gene is processed by the DICER-LIKE1 (DCL1) enzyme, similar to RNase III, into a hairpin structure, a miRNA precursor or pre-miRNA, which is further cleaved by DCL1 to form a miRNA/miRNA* duplex from the stem region of the hairpin. The double-stranded miRNA/miRNA* is assembled into a RNA-induced silencing complex (RISC) [90]. miR482 is a conserved and extensive family of microRNAs that plays a key role in regulating plant defense mechanisms by targeting transcripts with leucine-rich nucleotide-binding site (NBS-LRR) motifs. NBS-LRR genes are involved in plant immunity by mediating the recognition of pathogen effectors [91]. Hong et al. 2021 obtained two transgenic plants with simultaneous suppression of miR482b and miR482c and one transgenic line with suppression of only miR482b. Compared to wild-type plants, disease symptoms in the three transgenic plants were reduced upon infection, accompanied by increased expression of their common target genes with nucleotide binding sites and leucine-rich repeats and decreased levels of reactive oxygen species. In addition, simultaneous suppression of miR482b and miR482c was more effective than suppression of only miR482b in tomatoes. It was also found that knockout of miR482b and miR482c can cause disruption of the expression of other miRNAs, indicating cross-regulation between miRNAs. The authors' study demonstrated that simultaneous editing of miR482b and miR482c using CRISPR-Cas9 is an effective strategy for creating pathogen-resistant tomatoes, and that cross-regulation between miRNAs may reveal a new mechanism of interaction between tomatoes and P. infestans [77].
It has been demonstrated that inactivation of a single gene, called Downy Mildew Resistance 6 (DMR6), confers resistance to several pathogens in A. thaliana [92]. This gene is specifically activated during pathogen infection, and mutations in the dmr6 gene lead to increased salicylic acid levels. Using the CRISPR-Cas9 system, Thomazella et al. created tomato plants with small deletions in the SlDMR6-1 gene, which lead to gene inactivation. The authors showed that the resulting mutants have no significant adverse effects on growth and development under greenhouse conditions and demonstrate resistance to various pathogens, including P. syringae, P. capsici, and Xanthomonas spp [78].
Xylem sap protein 10 (XSP10) and salicylic acid methyltransferase (SlSAMT) are two putative negative regulatory genes associated with Fusarium wilt in tomatoes. Tomato tolerance to Fusarium wilt can be developed by targeting these susceptible (S) genes. XSP10 is a non-specific lipid transfer protein (LTP) with a molecular weight of 10 kDa, containing a motif of 8 cysteine residues that forms intramolecular disulfide bonds in tomatoes. According to research, the XSP10 gene acts as a compatibility factor for Fol, enhancing Fol colonization in the roots of tomato plants and promoting the development of disease symptoms in plants. In addition, it transports essential lipids from the plasma membrane to pathogens, increasing susceptibility to disease and its progression [93]. SAMT enzymes regulate salicylic acid (SA) homeostasis in plants by catalyzing the conversion of SA to methyl salicylate (MeSA) with S-adenosyl-L-methionine (SAM) as a methyl donor. The conversion of endogenous SA to MeSA reduces the host's defense against multiple pathogen attacks [94]. F. oxysporum f. sp. lycopersici is a fungal pathogen that attacks plant roots and ranks fifth among the most destructive fungal infections of tomatoes [95]. The fungus's hyphae penetrate and colonize the apoplastic spaces, surrounding the stele and blocking the xylem vessels, which leads to stunted growth, leaf chlorosis, progressive wilting, and cell death [96]. In a study conducted by Debbarma et al. two S genes (XSP10 and SAMT) were analyzed using CRISPR-Cas9 editing of one gene (XSP10 and SlSAMT separately) and two genes (XSP10 and SlSAMT simultaneously). Before proceeding to the creation of stable lines, the efficiency of sgRNA-Cas9 complex editing was first verified by transforming individual protoplast cells. Lines with CRISPR editing of the double genes XSP10 and SlSAMT in the GE1 generation showed strong phenotypic tolerance to Fusarium wilt disease compared to lines with single gene editing. Overall, genetics studies in transient and stable tomato lines showed that XSP10 and SlSAMT function together as negative regulators, providing genetic tolerance to Fusarium wilt disease [79]. In earlier work by Gaona et al. the group screened a Micro-Tom EMS tomato population and found a mutant that showed reduced susceptibility to both necrotrophic fungi. The authors reported a mutation in the tomato PUB17 gene as the cause of reduced susceptibility in this mutant (Gaona et al. 2017). The role of another gene in this family, PUB21, as a susceptibility factor to both necrotrophic fungi was confirmed in transformants with RNAi suppression and CRISPR mutations. The results show that SlPUB21 plays a key role in the susceptibility of tomato plants to the necrotrophic pathogens B. cinerea and A. solani. The SlPUB21 mutation provides a broad spectrum of resistance to these pathogens without causing serious pleiotropic effects. The double mutant pub17/pub21 in tomatoes showed a higher level of resistance, but also exhibited reduced plant size [97].
Tomato plants with CRISPR-Cas9-induced Phospholipase C2 knockouts are more resistant to Botrytis cinerea than wild-type plants. Perk et al. obtained SlPLC2 knockout tomato lines with reduced ROS (reactive oxygen species) production upon exposure to B. cinerea. Since this fungus requires ROS-induced cell death for reproduction, SlPLC2 knockout plants showed increased resistance with fewer necrotic areas and reduced pathogen reproduction. Thus, using CRISPR-Cas9 genome editing technology, we obtained tomato lines with SlPLC2 loss of function that were more resistant to B. cinerea [81]. Phosphoinositide-specific phospholipase C (PI-PLC) is a signaling enzyme that hydrolyzes membrane phosphoinositides to form lipids and lipid derivatives that act as secondary messengers. In plants, PI-PLCs are involved in various physiological processes, including immunity. It has previously been shown that tomato SlPLC2 is involved in susceptibility to the necrotrophic fungus Botrytis cinerea [81].
Ortega-Salazar et al. simultaneously knocked out two pectin-degrading enzymes, polygalacturonase (SlPG2a) and pectate lyase (SlPL), which play a key role in softening tomato fruits. It was found that SlPG2a and SlPL enzymes act additively, significantly affecting fruit firmness and shelf life, with double CRISPR knockout (CRISPR PGPL line) outperforming wild-type fruits. In addition, compared to wild-type fruits, CRISPR double knockout (PGPL) fruits showed improved or unchanged quality indicators such as sugar-to-acid ratio, aromatic volatiles, and skin color [82]. Fruit with a single CRISPR PL gene knockout is significantly less susceptible to fungal infections caused by B. cinerea compared to WT fruit [98]. The authors inoculated fruits of all tomato lines obtained in the study with B. cinerea and monitored disease development throughout the post-harvest storage period, which showed a more resistant phenotype in CRISPR PL and CRISPR PGPL line compared to the wild type [82].
The Walls Are Thin 1 (WAT1) gene is a disease susceptibility gene first discovered in Arabidopsis and cotton that exhibits susceptibility to Verticillium dahliae, a particularly well-known vascular wilt pathogen [99]. WAT1 encodes an auxin transporter localized in the tonoplast, but its exact role in so-called “vascular immunity” is not yet clear [100]. Hanika et al. identified the tomato WAT1 homolog Solyc04g080940 (SlWAT1) in their study. Temporary and stable suppression of SlWAT1 based on VIGS and RNAi, respectively, did not result in a permanent reduction in tomato susceptibility to V. dahliae. However, CRISPR-Cas9 tomato mutant lines carrying targeted deletions in SlWAT1 showed significantly increased resistance to V. dahliae, as well as to Verticillium alabastrum and Fusarium oxysporum f. sp. lycopersici. Thus, the knockout of the tomato WAT1 gene resulted in a broad spectrum of resistance to various vascular pathogens of tomato [83].
The application of CRISPR-Cas technology to tomatoes has demonstrated impressive success in increasing resistance to major fungal pathogens such as O. neolycopersici, P. infestans, F. oxysporum, B. cinerea and V. dahliae. Through precise editing of susceptibility and defense genes, researchers have created lines with stable disease resistance, increased yield, and no significant growth abnormalities. These achievements demonstrate that CRISPR/Cas is a powerful and sustainable tool for breeding a new generation of tomato varieties that are resistant to fungal diseases.
4.2. Pepper
Pepper (Capsicum annuum L.) is a widely cultivated and important vegetable crop due to its high nutritional value, unique vitamin content, and diverse mineral composition [101]. However, it is susceptible to climate-driven spread of pathogens and vectors [102]. Climate change has exacerbated the problem of powdery mildew, a disease that hinders pepper growth and yield by disrupting photosynthesis and hormone production. Wind and water contribute to the spread of powdery mildew infection [103].
Park et al. used the well-known MLO gene for knockout in C. annuum using previously created guide RNAs targeting the CaMLO2 gene, using RNP for protoplast transformation. This study confirmed the reliability of protoplast isolation and the effectiveness of Cas9/CaMLO2sgRNA1 as a gene editing tool without potential side effects in six commercial hot pepper varieties. Despite the genetic diversity of the six pepper varieties, the study of the genetic structure of CaMLO2 homologs showed that this approach is consistent and reliable for targeting and modifying the gene [84].
Another group had previously demonstrated that pepper protoplasts derived from leaves or callus grown in soil are a useful system for screening effective guide RNAs for CRISPR-Cas9 or CRISPR-Cas12a (Cpf1). This study shows that CRISPR-Cas9 or Cpf1 were delivered as CRISPR/RNP complexes from purified endonucleases mixed with engineered single-guide RNA that can edit the target gene CaMLO2 in two pepper varieties with sequenced complete genomes, C. annuum ‘CM334’ and C. annuum ‘Dempsey’. The engineered guide RNAs (sgRNA for Cas9 or crRNA for Cpf1) are retained for CaMLO2 in both CM334 and Dempsey and cleave CaMLO2 in vitro. CRISPR-Cas9- or /Cpf1-RNP complexes were transfected into freshly isolated protoplasts of CM334 hot pepper and Dempsey sweet pepper using PEG-mediated delivery. Targeted deep sequencing analysis showed that the target gene CaMLO2 was differentially edited in both varieties depending on the CRISPR/RNP applied [85].
Another gene, also an S gene, ERF28, was used for knockout in pepper. This gene was selected by researchers as a gene that is overexpressed during anthracnose infection [104]. Anthracnose, caused by Colletotrichum species, is a major disease of chili peppers leading to significant yield losses before and after harvest in tropical and subtropical regions of the world [105]. Mishra et al. developed a construct with single expression of Cas9, sgRNA driven by the Pol II promoter to modify the CaERF28 gene in the susceptible chili pepper genotype Arka Lohit. Of the 62 T0 transgenic plants, 45 mutant lines induced by CaERF28 (72.5%) were identified. In addition, simultaneous exposure to multiple sites within CaERF28 showed increased mutation efficiency (85.7%). Five homozygous mutants showed increased resistance to anthracnose compared to the wild type, as evidenced by a reduction in spore and fungal growth, as well as induced expression of defense-related genes. In addition, the authors showed that all mutant chili pepper plants were morphologically normal and agronomically similar to the wild type, suggesting that modification of the CaERF28 locus does not interfere with plant development [86].
CRISPR-Cas technology has demonstrated high potential for improving disease resistance in peppers (C. annuum L.). Successful knockout of key susceptibility genes, such as CaMLO2 and CaERF28, has demonstrated increased resistance to major fungal pathogens, such as powdery mildew and anthracnose, without negatively affecting plant growth or yield. These achievements demonstrate the effectiveness and precision of CRISPR/Cas as a tool for creating disease-resistant pepper varieties that contribute to sustainable crop production in a changing climate.
4.3. Eggplant
According to the FAO, eggplant (Solanum melongena L.) is the most important berry crop of the nightshade family after tomatoes, with a production volume of 60 million tons. Since eggplant is a vegetable that contributes to nutrition in large areas of the world, it is extremely important to develop new strategies to increase its disease resistance and, as a result, increase its production in order to cope with the rapid growth of the world's population and the effects of climate change [106].
Among the S genes, DMR6 encodes an enzyme involved in the degradation of salicylic acid (SA), and its inactivation in other Solanaceae species has been shown to increase SA levels and confer tolerance to a wide range of pathogens, as described earlier. Ferrero et al. identified two orthologs of this gene in the eggplant genome, namely SmDMR6–1 and SmDMR6–2, for increased resistance to the oomycete pathogens Phytophthora capsici and Phytophthora infestans. In the Black Beauty variety, only the expression of SmDMR6–1 was significantly increased upon infection with two oomycetes, P. infestans and P. capsici, indicating its involvement in regulating the plant's response to biotic stresses. The authors demonstrated the knockout of the SmDMR6–1 gene using CRISPR-Cas9 technology in eggplants. Regenerated T0 plants were screened using Sanger sequencing, one of them was selected and self-pollinated to obtain T1 and then T2 plants. The mutant lines were tested for pathogenicity, which revealed increased tolerance to infection by P. infestans and P. capsici compared to unedited plants [87].
Knockout of the SmDMR6–1 gene in eggplant (S. melongena L.) using CRISPR-Cas9 successfully increased resistance to late blight, a major oomycete pathogen affecting crop yield. This demonstrates the potential of genome editing to enhance eggplant disease resistance without compromising plant growth. Such achievements represent an effective strategy for creating sustainable varieties that contribute to sustainable production and global food security in the context of climate change.
5. Challenges, Future Perspectives and Conclusions
Recent studies have clearly shown that CRISPR/Cas technology has become a key tool for precision genome editing and creating fungal disease-resistant varieties of major crops. The application of this approach has made it possible to successfully modify susceptibility genes in wheat, rice, barley, soybeans, tomatoes, peppers, and eggplants, providing resistance to fungal pathogens such as Fusarium graminearum, Fusarium oxysporum, Parastagonospora nodorum, Blumeria graminis f. sp. tritici, Puccinia striiformis f. sp. tritici, Puccinia triticina Erikss, Magnaporthe oryzae, Erysiphe diffusa, Phytophthora sojae, Oidium neolycopersici, Phytophthora infestans, Phytophthora capsici, Botrytis cinerea, Verticillium dahliae, Leveillula taurica and Colletotrichum truncatum. CRISPR-Cas technology provides high accuracy, efficiency, and reproducibility without affecting plant growth and yield, making it a powerful and environmentally safe tool for sustainable agriculture. Further development of areas related to S- and R-gene editing, the introduction of plasmid-free systems, multiplex editing, and CRISPRi regulatory platforms will open up new opportunities for accelerated breeding of crops with complex disease resistance and adaptation to changing climatic conditions.
Currently, the fastest and most reliable way to obtain genetically modified plants with the desired disease resistance phenotype is to use CRISPR-Cas technology. There are many studies demonstrating the high potential of CRISPR-Cas in obtaining new varieties in just 10 months [72]. CRISPR-Cas9 has been successfully used to combat various fungal diseases in many agricultural crops, demonstrating its potential to accelerate research in biotechnology and agronomy. CRISPR-Cas technology is mainly used for gene knockout in loss-of-function studies [107]. Given the large number of S-genes and their conservative nature, it allows the creation of resistant varieties for a wide range of diseases, as well as the discovery of new S-genes from one crop and their transfer to another. For example, MLO genes, originally discovered in A. thaliana, and the ortholog of this gene in tomatoes have demonstrated resistance to powdery mildew [108]. The same has happened with the PMR gene, which was also originally discovered in Arabidopsis and is used in many different crops to increase their resistance [88].
Beyond gene knockouts, CRISPR-Cas possesses diverse variants such as base editing and transcription activation/inhibition, RNAi and more [109,110]. However, despite its broad capabilities, the use of other types of CRISPR in fungal disease resistance remains quite limited. Most of the research focuses on susceptibility gene knockouts. Other editing approaches either constitute a relatively small minority of studies or are entirely dedicated to basic research [111]. Gene knock-in studies represent another avenue for further research. This technique involves inserting specific DNA sequences into precise locations via the HDR pathway. This process requires conditions such as the target sequence presented as donor DNA and molecules that trigger HDR efficacy. Non-pathogenic factors (Avr) and effector proteins are secreted by plant pathogens into plant cells during initial infection. These effectors suppress plant defense mechanisms, facilitating pathogen establishment. However, plants have evolved defense mechanisms by acquiring resistance proteins encoded by host resistance genes (R genes), which confer marked resistance against specific pathogen strains [112]. For example, R genes such as RPS5 (resistance to Pseudomonas syringae) and Avr gene AVRPphB can enhance resistance to fungal diseases when introduced into host organisms [113]. Even when already present, these genes can be overexpressed using transcription-regulating Cas proteins.
The most widespread method of utilizing the Cas9 cassette in plants is Agrobacterium-mediated transformation using a plasmid containing the Cas9 protein and the sgRNA sequence [114]. After transformation, Agrobacterium inserts the CRISPR-Cas9 cassette into the plant genome, but this can lead to various undesirable consequences such as permanent expression of the Cas9 protein or Cas9 protein off-target effects [115]. In addition, the presence of the Cas9 gene in the plant genome raises ethical concerns.
Author Contributions
Designing and reviewing Section 1, Section 2 and Section 5, Z.S.; Designing and reviewing Section 3, Section 4 and Section 5, R.K.; reviewing Section 2, D.D.; reviewing Section 3, A.D.; reviewing Section 4, M.S.; reviewing the manuscript prior to submission, K.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research and the APC were funded by the Science Committee of the Ministry of Science and Higher Education of the Republic of Kazakhstan (Grant No. BR21882269).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We extend our heartfelt appreciation to the Science Committee of the Ministry of Science and Higher Education of the Republic of Kazakhstan for financial support. Their insightful comments and suggestions have significantly enhanced the quality and rigor of this review. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of this review, in the collection of data, analyses, or interpretation of data, in the writing of the manuscript, or in the decision to publish this review.
Abbreviations
The following abbreviations are used in this manuscript:
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats |
| Cas | CRISPR-associated protein |
| FHB | Fusarium head blight |
| BSMV | Barley mosaic virus |
| VIGS | Virus-induced gene silencing |
| SNB | Septoria nodorum blotch |
| MLO | Powdery mildew locus O |
| CRISPRi | CRISPR interference |
| BYDV | Barley yellow dwarf virus |
| FAO | Food and Agriculture Organization of the United Nations |
| DMR6 | Downy mildew resistance 6 |
| DND1 | Defense no death 1 |
| DCL1 | Dicer-like1 |
| NBS-LRR | Nucleotide-binding site |
| LRR | Leucine-rich repeat |
| XSP10 | Xylem sap protein 10 |
| SlSAMT | Salicylic acid methyltransferase |
| LTP | Lipid transfer protein |
| SA | Salicylic acid |
| MeSA | Methyl salicylate |
| SAM | S-adenosyl-L-methionine |
| SlPG2a | Polygalacturonase |
| SlPL | Pectate lyase |
References
- The United Nations. World population projected to reach 9.8 billion in 2050, and 11.2 billion in 2100. Available online: URL Available online: https://www.un.org/en/desa/world-population-projected-reach-98-billion-2050-and-112-billion-2100. (accessed on 2 October 2025).
- Food and Agriculture Organization. Plant Production and Protection. Available online: URL Available online: https://www.fao.org/plant-production-protection/about/en. (accessed on 2 October 2025).
- Mikaberidze, A.; Gokhale, C.S.; Bargués-Ribera, M.; Verma, P. The Cost of Fungicide Resistance Evolution in Multi-Field Plant Epidemics. PLOS Sustainability and Transformation 2025, 4. [CrossRef]
- Wang, N.; Sundin, G.W.; De La Fuente, L.; Cubero, J.; Tatineni, S.; Brewer, M.T.; Zeng, Q.; Bock, C.H.; Cunniffe, N.J.; Wang, C.; et al. Key Challenges in Plant Pathology in the Next Decade. Phytopathology 2024, 114, 837–842.
- Różewicz, M.; Wyzińska, M.; Grabiński, J. The Most Important Fungal Diseases of Cereals—Problems and Possible Solutions. Agronomy 2021, 11. [CrossRef]
- Sbai, H.; Hajib, A.; Msairi, S.; Amalich, S.; Bouyahya, A.; Lee, L.-H.; Wen Goh, K.; Tabyaoui, M.; Harhar, H. Fungal Infections of Legume Crops: Challenges and Management Approaches. Journal of Agriculture and Food Research 2024, 18, 101447. [CrossRef]
- Tripathi, A.N.; Maurya, S.; Pandey, K.K.; Behera, T.K. Global Scenario of Vegetable Fungal Diseases. Vegetable Science 2024, 51, 54–65. [CrossRef]
- El Afandi, G.; Irfan, M. Pesticides Risk Assessment Review: Status, Modeling Approaches, and Future Perspectives. Agronomy 2024, 14.
- Islam, T.; Danishuddin; Tamanna, N.T.; Matin, M.N.; Barai, H.R.; Haque, M.A. Resistance Mechanisms of Plant Pathogenic Fungi to Fungicide, Environmental Impacts of Fungicides, and Sustainable Solutions. Plants 2024, 13.
- Morgounov, A.; Akin, B.; Demir, L.; Keser, M.; Kokhmetova, A.; Martynov, S.; Orhan, Ş.; Özdemir, F.; Özseven, ̄zzet; Sapakhova, Z.; et al. Yield Gain Due to Fungicide Application in Varieties of Winter Wheat (Triticum Aestivum) Resistant and Susceptible to Leaf Rust. Crop Pasture Sci 2015, 66, 649–659. [CrossRef]
- Madenova, A.; Sapakhova, Z.; Bakirov, S.; Galymbek, K.; Yernazarova, G.; Kokhmetova, A.; Keishilov, Z. Screening of Wheat Genotypes for the Presence of Common Bunt Resistance Genes. Saudi J Biol Sci 2021, 28, 2816–2823. [CrossRef]
- Sun, L.; Lai, M.; Ghouri, F.; Nawaz, M.A.; Ali, F.; Baloch, F.S.; Nadeem, M.A.; Aasim, M.; Shahid, M.Q. Modern Plant Breeding Techniques in Crop Improvement and Genetic Diversity: From Molecular Markers and Gene Editing to Artificial Intelligence—A Critical Review. Plants 2024, 13.
- Gao, M.; Hao, Z.; Ning, Y.; He, Z. Revisiting Growth–Defence Trade-Offs and Breeding Strategies in Crops. Plant Biotechnol J 2024, 22, 1198–1205.
- Ahmad, S.; Wei, X.; Sheng, Z.; Hu, P.; Tang, S. CRISPR/Cas9 for Development of Disease Resistance in Plants: Recent Progress, Limitations and Future Prospects. Brief Funct Genomics 2018, 19, 26–39.
- Ma, L.; Kong, F.; Sun, K.; Wang, T.; Guo, T. From Classical Radiation to Modern Radiation: Past, Present, and Future of Radiation Mutation Breeding. Front Public Health 2021, 9.
- Permyakova, N. V.; Deineko, E. V. Crop Improvement: Comparison of Transgenesis and Gene Editing. Horticulturae 2024, 10.
- Ahmad, S.; Rana, I.A.; Folta, K.M.; Lorenzo, C.D.; Khan, S.H. Editorial: Gene Editing to Achieve Zero Hunger. Front Genome Ed 2025, 7. [CrossRef]
- Waites, J.; Achary, V.M.M.; Syombua, E.D.; Hearne, S.J.; Bandyopadhyay, A. CRISPR-Mediated Genome Editing of Wheat for Enhancing Disease Resistance. Front Genome Ed 2025, 7.
- Lau, C.-H.; Tin, C.; Suh, Y. CRISPR-Based Strategies for Targeted Transgene Knock-in and Gene Correction. Fac Rev 2020, 9. [CrossRef]
- Sapakhova, Z.; Kanat, R.; Choi, K.; Daurov, D.; Daurova, A.; Zhambakin, K.; Shamekova, M. CRISPR-Cas Gene Editing Technology in Potato. Int J Mol Sci 2025, 26. [CrossRef]
- Rynjah, D.; Sandhanam, K.; Bhattacharjee, B.; Deka, B.; Newar, A.; Kalita, T.; Nath, J.; Ahmed, A.B.; Sahu, R.K.; Das, T. CRISPR/Cas9 Gene Editing Systems for Enhancing Secondary Metabolite Biosynthesis via Reproductive Tissue Modification. Discover Plants 2025, 2, 245. [CrossRef]
- Kokhmetova, A.; Kremneva, O.; Volkova, G.; Atishova, M.; Sapakhova, Z. Evaluation of Wheat Cultivars Growing in Kazakhstan and Russia for Resistance to Tan Spot. Journal of Plant Pathology 2017, 99, 161–167. [CrossRef]
- Kokhmetova, A.M.; Ali, S.; Sapakhova, Z.; Atishova, M.N. Identification of Genotypes-Carriers of Resistance to Tan Spot Ptr ToxA and Ptr ToxB of Pyrenophora Tritici-Repentis in Common Wheat Collection. Vavilovskii Zhurnal Genet Selektsii 2018, 22, 978–986. [CrossRef]
- Madenova, A.; Kokhmetova, A.; Sapakhova, Z.; Galymbek, K.; Keishilov, Z.; Akan, K.; Yesserkenov, A. Effect of Common Bunt [Tilletia Caries (DC) Tul] Infection on Agronomic Traits and Resistance of Wheat Entries. Research on Crops 2020, 21, 791–797. [CrossRef]
- Taj, M.; Sajjad, M.; Li, M.; Yasmeen, A.; Mubarik, M.S.; Kaniganti, S.; He, C. Potential Targets for CRISPR/Cas Knockdowns to Enhance Genetic Resistance Against Some Diseases in Wheat (Triticum Aestivum L.). Front Genet 2022, 13. [CrossRef]
- Mu, K.; Ren, X.; Yang, H.; Zhang, T.; Yan, W.; Yuan, F.; Wu, J.; Kang, Z.; Han, D.; Deng, R.; et al. CRISPR-Cas12a-Based Diagnostics of Wheat Fungal Diseases. J Agric Food Chem 2022, 70, 7240–7247. [CrossRef]
- Su, Z.; Bernardo, A.; Tian, B.; Chen, H.; Wang, S.; Ma, H.; Cai, S.; Liu, D.; Zhang, D.; Li, T.; et al. A Deletion Mutation in TaHRC Confers Fhb1 Resistance to Fusarium Head Blight in Wheat. Nat Genet 2019, 51, 1099–1105. [CrossRef]
- Chen, H.; Su, Z.; Tian, B.; Liu, Y.; Pang, Y.; Kavetskyi, V.; Trick, H.N.; Bai, G. Development and Optimization of a Barley Stripe Mosaic Virus-Mediated Gene Editing System to Improve Fusarium Head Blight Resistance in Wheat. Plant Biotechnol J 2022, 20, 1018–1020. [CrossRef]
- Karmacharya, A.; Li, D.; Leng, Y.; Shi, G.; Liu, Z.; Yang, S.; Du, Y.; Dai, W.; Zhong, S. Targeting Disease Susceptibility Genes in Wheat Through Wide Hybridization with Maize Expressing Cas9 and Guide RNA. Molecular Plant-Microbe Interactions 2023, 36, 554–557. [CrossRef]
- Brauer, E.K.; Balcerzak, M.; Rocheleau, H.; Leung, W.; Schernthaner, J.; Subramaniam, R.; Ouellet, T.; Genome Editing of a Deoxynivalenol-Induced Transcription Factor Confers Resistance to Fusarium graminearum in Wheat. Mol Plant Microbe Interact. 2020, 33, 553-560. [CrossRef]
- Jones, D.A.B.; Rybak, K.; Hossain, M.; Bertazzoni, S.; Williams, A.; Tan, K.C.; Phan, H.T.T.; Hane, J.K. Repeat-Induced Point Mutations Driving Parastagonospora Nodorum Genomic Diversity Are Balanced by Selection against Non-Synonymous Mutations. Commun Biol 2024, 7. [CrossRef]
- Poddar, S.; Tanaka, J.; Running, K.L.D.; Kariyawasam, G.K.; Faris, J.D.; Friesen, T.L.; Cho, M.J.; Cate, J.H.D.; Staskawicz, B. Optimization of Highly Efficient Exogenous-DNA-Free Cas9-Ribonucleoprotein Mediated Gene Editing in Disease Susceptibility Loci in Wheat (Triticum Aestivum L.). Front Plant Sci 2023, 13. [CrossRef]
- Khan, H.; McDonald, M.C.; Williams, S.J.; Solomon, P.S. Assessing the Efficacy of CRISPR/Cas9 Genome Editing in the Wheat Pathogen Parastagonspora Nodorum. Fungal Biol Biotechnol 2020, 7. [CrossRef]
- Zou, S.; Xu, Y.; Li, Q.; Wei, Y.; Zhang, Y.; Tang, D. Wheat Powdery Mildew Resistance: From Gene Identification to Immunity Deployment. Front Plant Sci 2023, 14.
- Li, S.; Lin, D.; Zhang, Y.; Deng, M.; Chen, Y.; Lv, B.; Li, B.; Lei, Y.; Wang, Y.; Zhao, L.; et al. Genome-Edited Powdery Mildew Resistance in Wheat without Growth Penalties. Nature 2022, 602, 455–460. [CrossRef]
- Zhang, Y.; Bai, Y.; Wu, G.; Zou, S.; Chen, Y.; Gao, C.; Tang, D. Simultaneous Modification of Three Homoeologs of TaEDR1 by Genome Editing Enhances Powdery Mildew Resistance in Wheat. Plant Journal 2017, 91, 714–724. [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.L. Simultaneous Editing of Three Homoeoalleles in Hexaploid Bread Wheat Confers Heritable Resistance to Powdery Mildew. Nat Biotechnol 2014, 32, 947–951. [CrossRef]
- Kokhmetova, A.; Rathan, N.D.; Sehgal, D.; Malysheva, A.; Kumarbayeva, M.; Nurzhuma, M.; Bolatbekova, A.; Krishnappa, G.; Gultyaeva, E.; Kokhmetova, A.; et al. QTL Mapping for Seedling and Adult Plant Resistance to Stripe and Leaf Rust in Two Winter Wheat Populations. Front Genet 2023, 14. [CrossRef]
- Ochilov, B.O.; Turakulov, K.S.; Meliev, S.K.; Melikuziev, F.A.; Aytenov, I.S.; Murodova, S.M.; Khalillaeva, G.O.; Chinikulov, B.K.; Azimova, L.A.; Urinov, A.M.; et al. Development of Yellow Rust-Resistant and High-Yielding Bread Wheat (Triticum Aestivum L.) Lines Using Marker-Assisted Backcrossing Strategies. Int J Mol Sci 2025, 26. [CrossRef]
- Malysheva, A.; Kokhmetova, A.; Urazaliev, R.; Kumarbayeva, M.; Keishilov, Z.; Nurzhuma, M.; Bolatbekova, A.; Kokhmetova, A. Phenotyping and Identification of Molecular Markers Associated with Leaf Rust Resistance in the Wheat Germplasm from Kazakhstan, CIMMYT and ICARDA. Plants 2023, 12. [CrossRef]
- Liu, S.; Zhang, F.; Su, J.; Fang, A.; Tian, B.; Yu, Y.; Bi, C.; Ma, D.; Xiao, S.; Yang, Y. CRISPR-Targeted Mutagenesis of Mitogen-Activated Protein Kinase Phosphatase 1 Improves Both Immunity and Yield in Wheat. Plant Biotechnol J 2024, 22, 1929–1941. [CrossRef]
- Wang, N.; Tang, C.; Fan, X.; He, M.; Gan, P.; Zhang, S.; Hu, Z.; Wang, X.; Yan, T.; Shu, W.; et al. Inactivation of a Wheat Protein Kinase Gene Confers Broad-Spectrum Resistance to Rust Fungi. Cell 2022, 185, 2961-2974.e19. [CrossRef]
- Liu, S.; Liu, H.; Guo, M.; Pan, Y.; Hao, C.; Hou, J.; Yan, L.; Zhang, X.; Chen, X.; Li, T. Knockout of GRAIN WIDTH2 Has a Dual Effect on Enhancing Leaf Rust Resistance and Increasing Grain Weight in Wheat. Plant Biotechnol J 2024, 22, 2007–2009. [CrossRef]
- Conde, S.; Catarino, S.; Ferreira, S.; Temudo, M.P.; Monteiro, F. Rice Pests and Diseases Around the World: Literature-Based Assessment with Emphasis on Africa and Asia. Agriculture (Switzerland) 2025, 15.
- Foster, A.J.; Martin-Urdiroz, M.; Yan, X.; Wright, S.; Soanes, D.M.; Talbot, N.J. CRISPR-Cas9 Ribonucleoprotein-Mediated Co-Editing and Counterselection in the Rice Blast Fungus 2018.
- Wang, F.; Wang, C.; Liu, P.; Lei, C.; Hao, W.; Gao, Y.; Liu, Y.G.; Zhao, K. Enhanced Rice Blast Resistance by CRISPR/ Cas9-Targeted Mutagenesis of the ERF Transcription Factor Gene OsERF922. PLoS One 2016, 11. [CrossRef]
- Ma, J.; Chen, J.; Wang, M.; Ren, Y.; Wang, S.; Lei, C.; Cheng, Z.; Sodmergen Disruption of OsSEC3A Increases the Content of Salicylic Acid and Induces Plant Defense Responses in Rice. J Exp Bot 2018, 69, 1051–1064. [CrossRef]
- Wang, R.J.; Zhao, J.; Bhadauria, V.; Peng, Y.L. Efficient Gene Editing with an Arg-TRNA Promoter-Driven CRISPR/Cas9 in the Rice Blast Fungus Pyricularia Oryzae. Phytopathology Research 2024, 6. [CrossRef]
- Wang, F-Q.; Fan, F-J.; Li, W-Q.; Zhu, J-Y.; Wang, J.; Zhong, W.-G.; Yang, J. Knock-out Efficiency Analysis of Pi21 Gene Using CRISPR/Cas9 in Rice. Chinese Journal ofF Rice Science 2016, 30, 469-478. [CrossRef]
- Xu, P.; Wang, H.; Tu, R.; Liu, Q.; Wu, W.; Fu, X.; Cao, L.; Shen, X. Orientation improvement of blast resistance in rice via CRISPR/Cas9 system. Chinese Journal of Rice Science 2019, 33, 313-322. [CrossRef]
- Li, S.; Shen, L.; Hu, P.; Liu, Q.; Zhu, X.; Qian, Q.; Wang, K.; Wang, Y. Developing Disease-Resistant Thermosensitive Male Sterile Rice by Multiplex Gene Editing. J Integr Plant Biol 2019, 61, 1201–1205. [CrossRef]
- Yang, J.; Fang, Y.; Wu, H.; Zhao, N.; Guo, X.; Mackon, E.; Peng, H.; Huang, S.; He, Y.; Qin, B.; et al. Improvement of Resistance to Rice Blast and Bacterial Leaf Streak by CRISPR/Cas9-Mediated Mutagenesis of Pi21 and OsSULTR3;6 in Rice (Oryza Sativa L.). Front Plant Sci 2023, 14. [CrossRef]
- Zhou, Y.; Xu, S.; Jiang, N.; Zhao, X.; Bai, Z.; Liu, J.; Yao, W.; Tang, Q.; Xiao, G.; Lv, C.; et al. Engineering of Rice Varieties with Enhanced Resistances to Both Blast and Bacterial Blight Diseases via CRISPR/Cas9. Plant Biotechnol J 2022, 20, 876–885. [CrossRef]
- Peng, Q.; Li, J.; Xu, H.; Zhang, X.; Zhang, D.; Zhu, S. Editing Pi21 Gene to Improve the Blast Resistance of Cultivated Rice dalixiang by Using CRISPR/Cas9 System. Molecular Plant Breeding 2022. https://kns.cnki.net/kcms/detail/46.1068.S.20220221.1756.019.htm.
- Wu, X.; Li, J-L.; Ceng, Q.-H.; Zhang, D.-S.; Xu, H.-F.; Jiang, X.; Song, Li.; Peng, Q.; Zhu, S.-S. Improvement of Blast Resistance of Rice Variety Dalixiang by CRISPR/Cas9 Gene Editing Technology. Seed 2021, 40, 50-55. [CrossRef]
- Zhu, Z.; Yin, J.; Chern, M.; Zhu, X.; Yang, C.; He, K.; Liu, Y.; He, M.; Wang, J.; Song, L.; et al. New Insights into Bsr-D1-Mediated Broad-Spectrum Resistance to Rice Blast. Mol Plant Pathol 2020, 21, 951–960. [CrossRef]
- Zhang, Y.; Zheng, L.; Xie, K. CRISPR/DCas9-Mediated Gene Silencing in Two Plant Fungal Pathogens. mSphere 2023, 8. [CrossRef]
- Ren, B.; Yan, F.; Kuang, Y.; Li, N.; Zhang, D.; Zhou, X.; Lin, H.; Zhou, H. Improved Base Editor for Efficiently Inducing Genetic Variations in Rice with CRISPR/Cas9-Guided Hyperactive HAID Mutant. Mol Plant 2018, 11, 623–626.
- Martínez-Moreno, F.; Solís, I.; Igartua, E. Barley History and Breeding in Spain. Agriculture (Switzerland) 2024, 14.
- Çelik Oğuz, A.; Karakaya, A. Genetic Diversity of Barley Foliar Fungal Pathogens. Agronomy 2021, 11.
- Poursafar, A.; Leng, Y.; Zhong, S. Development of a CRISPR/Cas9-Mediated Gene Knockout Method for Functional Genomics of the Barley Spot Blotch Pathogen Bipolaris Sorokiniana . PhytoFrontiersTM 2024, 4, 682–689. [CrossRef]
- Ge, C.; Moolhuijzen, P.; Hickey, L.; Wentzel, E.; Deng, W.; Dinglasan, E.G.; Ellwood, S.R. Physiological Changes in Barley Mlo-11 Powdery Mildew Resistance Conditioned by Tandem Repeat Copy Number. Int J Mol Sci 2020, 21, 1–18. [CrossRef]
- Mishra, R.; Tripathi, M.K.; Sikarwar, R.S.; Singh, Y.; Tripathi, N. Soybean (Glycine Max L. Merrill): A Multipurpose Legume Shaping Our World. Plant Cell Biotechnol Mol Biol 2024, 25, 17–37. [CrossRef]
- McCaghey, M.; Willbur, J.; Smith, D.L.; Kabbage, M. The Complexity of the Sclerotinia Sclerotiorum Pathosystem in Soybean: Virulence Factors, Resistance Mechanisms, and Their Exploitation to Control Sclerotinia Stem Rot. Trop Plant Pathol 2019, 44, 12–22.
- Bui, T.P.; Le, H.; Ta, D.T.; Nguyen, C.X.; Le, N.T.; Tran, T.T.; Van Nguyen, P.; Stacey, G.; Stacey, M.G.; Pham, N.B.; et al. Enhancing Powdery Mildew Resistance in Soybean by Targeted Mutation of MLO Genes Using the CRISPR/Cas9 System. BMC Plant Biol 2023, 23. [CrossRef]
- Dou, D.; Kale, S.D.; Liu, T.; Tang, Q.; Wang, X.; Arredondo, F.D.; Basnayake, S.; Whisson, S.; Drenth, A.; Maclean, D.; et al. Different Domains of Phytophthora Sojae Effector Avr4/6 Are Recognized by Soybean Resistance Genes Rps4 and Rps6. / 425 MPMI 2010, 23, 425–435. [CrossRef]
- Fang, Y.; Tyler, B.M. Efficient Disruption and Replacement of an Effector Gene in the Oomycete Phytophthora Sojae Using CRISPR/Cas9. Mol Plant Pathol 2016, 17, 127–139. [CrossRef]
- Pavan, S.; Jacobsen, E.; Visser, R.G.F.; Bai, Y. Loss of Susceptibility as a Novel Breeding Strategy for Durable and Broad-Spectrum Resistance. Molecular Breeding 2010, 25, 1–12.
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol Plant Pathol 2012, 13, 414–430.
- Bü, R.; Hollricher, K. The Barley Mlo Gene: A Novel Control Element of Plant Pathogen Resistance; 1997; Vol. 88;.
- Acevedo-Garcia, J.; Kusch, S.; Panstruga, R. Magical Mystery Tour: MLO Proteins in Plant Immunity and Beyond. Journal of Physiology 2014, 204, 273–281.
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid Generation of a Transgene-Free Powdery Mildew Resistant Tomato by Genome Deletion. Sci Rep 2017, 7. [CrossRef]
- Pramanik, D.; Shelake, R.M.; Park, J.; Kim, M.J.; Hwang, I.; Park, Y.; Kim, J.Y. CRISPR/Cas9-Mediated Generation of Pathogen-Resistant Tomato against Tomato Yellow Leaf Curl Virus and Powdery Mildew. Int J Mol Sci 2021, 22, 1–18. [CrossRef]
- Santillán Martínez, M.I.; Bracuto, V.; Koseoglou, E.; Appiano, M.; Jacobsen, E.; Visser, R.G.F.; Wolters, A.M.A.; Bai, Y. CRISPR/Cas9-Targeted Mutagenesis of the Tomato Susceptibility Gene PMR4 for Resistance against Powdery Mildew. BMC Plant Biol 2020, 20. [CrossRef]
- Li, R.; Cui, L.; Martina, M.; Bracuto, V.; Meijer-Dekens, F.; Wolters, A.M.A.; Moglia, A.; Bai, Y.; Acquadro, A. Less Is More: CRISPR/Cas9-Based Mutations in DND1 Gene Enhance Tomato Resistance to Powdery Mildew with Low Fitness Costs. BMC Plant Biol 2024, 24. [CrossRef]
- Li, R.; Maioli, A.; Yan, Z.; Bai, Y.; Valentino, D.; Milani, A.M.; Pompili, V.; Comino, C.; Lanteri, S.; Moglia, A.; et al. CRISPR/Cas9-Based Knock-Out of the PMR4 Gene Reduces Susceptibility to Late Blight in Two Tomato Cultivars. Int J Mol Sci 2022, 23. [CrossRef]
- Hong, Y.; Meng, J.; He, X.; Zhang, Y.; Liu, Y.; Zhang, C.; Qi, H.; Luan, Y. Editing Mir482b and Mir482c Simultaneously by Crispr/Cas9 Enhanced Tomato Resistance to Phytophthora Infestans. Phytopathology 2021, 111, 1008–1016. [CrossRef]
- Paula de Toledo Thomazella, D.; Brail, Q.; Dahlbeck, D.; Staskawicz, B. CRISPR-Cas9 Mediated Mutagenesis of a DMR6 Ortholog in Tomato Confers Broad-Spectrum Disease Resistance 2016.
- Debbarma, J.; Saikia, B.; Singha, D.L.; Das, D.; Keot, A.K.; Maharana, J.; Velmurugan, N.; Arunkumar, K.P.; Reddy, P.S.; Chikkaputtaiah, C. CRISPR/Cas9-Mediated Mutation in XSP10 and SlSAMT Genes Impart Genetic Tolerance to Fusarium Wilt Disease of Tomato (Solanum Lycopersicum L.). Genes (Basel) 2023, 14. [CrossRef]
- Ramirez Gaona, M.; van Tuinen, A.; Schipper, D.; Kano, A.; Wolters, P.J.; Visser, R.G.F.; van Kan, J.A.L.; Wolters, A.M.A.; Bai, Y. Mutation of PUB17 in Tomato Leads to Reduced Susceptibility to Necrotrophic Fungi. Plant Biotechnol J 2023, 21, 2157–2159. [CrossRef]
- Perk, E.A.; D’Ambrosio, J.M.; Cerrudo, I.; Robuschi, L.; Juárez, M.; Vélez, P.; Mary, V.; Theumer, M.; Segretin, M.E.; Laxalt, A.M. Phosphoinositide-Specific Phospholipase C 2 (SlPLC2) Facilitates Vesicle Formation and Modulates Immune Signaling in Tomato Phytophthora Infestans Interactions 2025.
- Ortega-Salazar, I.; Crum, D.; Sbodio, A.O.; Sugiyama, Y.; Adaskaveg, A.; Wang, D.; Seymour, G.B.; Li, X.; Wang, S.C.; Blanco-Ulate, B. Double CRISPR Knockout of Pectin Degrading Enzymes Improves Tomato Shelf-Life While Ensuring Fruit Quality. Plants People Planet 2024, 6, 330–340. [CrossRef]
- Hanika, K.; Schipper, D.; Chinnappa, S.; Oortwijn, M.; Schouten, H.J.; Thomma, B.P.H.J.; Bai, Y. Impairment of Tomato WAT1 Enhances Resistance to Vascular Wilt Fungi Despite Severe Growth Defects. Front Plant Sci 2021, 12. [CrossRef]
- Park, J.H.; Kim, H. Harnessing CRISPR/Cas9 for Enhanced Disease Resistance in Hot Peppers: A Comparative Study on CaMLO2-Gene-Editing Efficiency across Six Cultivars. Int J Mol Sci 2023, 24. [CrossRef]
- Kim, H.; Choi, J.; Won, K.H. A Stable DNA-Free Screening System for CRISPR/RNPs-Mediated Gene Editing in Hot and Sweet Cultivars of Capsicum Annuum. BMC Plant Biol 2020, 20. [CrossRef]
- Mishra, R.; Mohanty, J.N.; Mahanty, B.; Joshi, R.K. A Single Transcript CRISPR/Cas9 Mediated Mutagenesis of CaERF28 Confers Anthracnose Resistance in Chilli Pepper (Capsicum Annuum L.). Planta 2021, 254. [CrossRef]
- Ferrero, M.; Valentino, D.; Milani, A.M.; Comino, C.; Lanteri, S.; Acquadro, A.; Moglia, A. Enhancing Tolerance to Phytophthora Spp. in Eggplant through DMR6–1 CRISPR/Cas9 Knockout. Plant Stress 2024, 14. [CrossRef]
- Vogel, J.; Somerville, S. Isolation and Characterization of Powdery Mildew-Resistant Arabidopsis Mutants; PNAS, 1999; Vol. 97;.
- Sun, K.; Wolters, A.M.A.; Loonen, A.E.H.M.; Huibers, R.P.; van der Vlugt, R.; Goverse, A.; Jacobsen, E.; Visser, R.G.F.; Bai, Y. Down-Regulation of Arabidopsis DND1 Orthologs in Potato and Tomato Leads to Broad-Spectrum Resistance to Late Blight and Powdery Mildew. Transgenic Res 2016, 25, 123–138. [CrossRef]
- Zhu, Q.H.; Fan, L.; Liu, Y.; Xu, H.; Llewellyn, D.; Wilson, I. MiR482 Regulation of NBS-LRR Defense Genes during Fungal Pathogen Infection in Cotton. PLoS One 2013, 8. [CrossRef]
- McHale, L.; Tan, X.; Koehl, P.; Michelmore, R.W. Plant NBS-LRR Proteins: Adaptable Guards. Genome Biol 2006, 7.
- Zeilmaker, T.; Ludwig, N.R.; Elberse, J.; Seidl, M.F.; Berke, L.; Van Doorn, A.; Schuurink, R.C.; Snel, B.; Van Den Ackerveken, G. Downy Mildew Resistant 6 and DMR6-like Oxygenase 1 Are Partially Redundant but Distinct Suppressors of Immunity in Arabidopsis. Plant Journal 2015, 81, 210–222. [CrossRef]
- Krasikov, V.; Dekker, H.L.; Rep, M.; Takken, F.L.W. The Tomato Xylem Sap Protein XSP10 Is Required for Full Susceptibility to Fusarium Wilt Disease. J Exp Bot 2011, 62, 963–973. [CrossRef]
- Koo, Y.J.; Kim, M.A.; Kim, E.H.; Song, J.T.; Jung, C.; Moon, J.-K.; Kim, J.-H.; Seo, H.S.; Song, S.I.; Kim, J.-K.; Lee, J.S.; Cheong, J.-J.; Choi, Y.D. Overexpression of Salicylic Acid Carboxyl Methyltransferase Reduces Salicylic Acid-Mediated Pathogen Resistance in Arabidopsis thaliana. Plant Mol Biol 2007, 64, 1–15. doi.org/10.1007/s11103-006-9123-x.
- Gordon, T.R. Fusarium Oxysporum and the Fusarium Wilt Syndrome. Annu Rev Phytopathol 2025, 15. [CrossRef]
- Joshi, R. A Review of Fusarium Oxysporum on Its Plant Interaction and Industrial Use. Journal of Medicinal Plants Studies 2018, 6, 112–115. [CrossRef]
- Ramírez Gaona, M.; van Tuinen, A.; Schipper, D.; Ramos Peregrina, Á.; Visser, R.G.F.; van Kan, J.A.L.; Bai, Y.; Wolters, A.-M.A. Mutation of PUB21 in Tomato Leads to Reduced Susceptibility to Necrotrophic Fungi. BMC Plant Biol 2025, 25, 1038. [CrossRef]
- Silva, C.J.; Van Den Abeele, C.; Ortega-Salazar, I.; Papin, V.; Adaskaveg, J.A.; Wang, D.; Casteel, C.L.; Seymour, G.B.; Blanco-Ulate, B. Host Susceptibility Factors Render Ripe Tomato Fruit Vulnerable to Fungal Disease despite Active Immune Responses. J Exp Bot 2021, 72, 2696–2709. [CrossRef]
- Denancé, N.; Ranocha, P.; Oria, N.; Barlet, X.; Rivière, M.P.; Yadeta, K.A.; Hoffmann, L.; Perreau, F.; Clément, G.; Maia-Grondard, A.; et al. Arabidopsis Wat1 (Walls Are Thin1)-Mediated Resistance to the Bacterial Vascular Pathogen, Ralstonia Solanacearum, Is Accompanied by Cross-Regulation of Salicylic Acid and Tryptophan Metabolism. Plant Journal 2013, 73, 225–239. [CrossRef]
- Ranocha, P.; Dima, O.; Nagy, R.; Felten, J.; Corratge-Faillie, C.; Novak, O.; Morreel, K.; Lacombe, B.; Martinez, Y.; Pfrunder, S.; et al. Arabidopsis WAT1 Is a Vacuolar Auxin Transport Facilitator Required for Auxin Homoeostasis. Nat Commun 2013, 4. [CrossRef]
- Cui, L.; van den Munckhof, M.C.; Bai, Y.; Voorrips, R.E. Resistance to Anthracnose Rot Disease in Capsicum. Agronomy 2023, 13. [CrossRef]
- Sanati, S.; Razavi, B.M.; Hosseinzadeh, H. A Review of the Effects of Capsicum Annuum L. And Its Constituent, Capsaicin, in Metabolic Syndrome. Iran J Basic Med Sci 2018, 21, 439–448.
- Mieslerová, B.; Cook, R.T.A.; Wheater, C.P.; Lebeda, A. Ecology of Powdery Mildews – Influence of Abiotic Factors on their Development and Epidemiology. Critical Reviews in Plant Sciences 2022, 41, 365–390. [CrossRef]
- Mishra, R.; Rout, E.; Joshi, R.K. Identification of Resistant Sources Against Anthracnose Disease Caused by Colletotrichum Truncatum and Colletotrichum Gloeosporioides in Capsicum Annuum L. Proceedings of the National Academy of Sciences India Section B - Biological Sciences 2019, 89, 517–524. [CrossRef]
- Oo, M.M.; Oh, S.-K. Chilli Anthracnose (Colletotrichum Spp.) Disease and Its Management Approach. Korean Journal of Agricultural Science 2016, 43, 153–162. [CrossRef]
- Taher, D.; Solberg, S.; Prohens, J.; Chou, Y.Y.; Rakha, M.; Wu, T.H. World Vegetable Center Eggplant Collection: Origin, Composition, Seed Dissemination and Utilization in Breeding. Front Plant Sci 2017, 8. [CrossRef]
- Saini, H.; Thakur, R.; Gill, R.; Tyagi, K.; Goswami, M. CRISPR/Cas9-Gene Editing Approaches in Plant Breeding. GM Crops Food 2023, 14, 1–17. [CrossRef]
- Kusch, S.; Panstruga, R. Mlo-Based Resistance: An Apparently Universal “Weapon” to Defeat Powdery Mildew Disease. Molecular Plant-Microbe Interactions 2017, 30, 179–189. [CrossRef]
- Liu, Z.; Dong, H.; Cui, Y.; Cong, L.; Zhang, D. Application of Different Types of CRISPR/Cas-Based Systems in Bacteria. Microb Cell Fact 2020, 19.
- Koonin, E. V.; Makarova, K.S.; Zhang, F. Diversity, Classification and Evolution of CRISPR-Cas Systems. Curr Opin Microbiol 2017, 37, 67–78. [CrossRef]
- Huang, J.; Gu, L.; Zhang, Y.; Yan, T.; Kong, G.; Kong, L.; Guo, B.; Qiu, M.; Wang, Y.; Jing, M.; et al. An Oomycete Plant Pathogen Reprograms Host Pre-MRNA Splicing to Subvert Immunity. Nat Commun 2017, 8. [CrossRef]
- Rasul, I.; Zafar, F.; Ali, M.A.; Nadeem, H.; Siddique, M.H.; Shahid, M.; Ashfaq, U.A.; Azeem, F. Genetic Basis for Biotic Stress Resistance in Plants from Solanaceae Family: A Review. Int J Agric Biol 2019, 22, 178–194.
- Pottinger, S.E.; Innes, R.W. RPS5-Mediated Disease Resistance: Fundamental Insights and Translational Applications. Annu. Rev. Phytopathol. 2020, 58, 139–60. [CrossRef]
- Zhang, Z.; Hua, L.; Gupta, A.; Tricoli, D.; Edwards, K.J.; Yang, B.; Li, W. Development of an Agrobacterium-Delivered CRISPR/Cas9 System for Wheat Genome Editing. Plant Biotechnol J 2019, 17, 1623–1635. [CrossRef]
- Gan, W.C.; Ling, A.P.K. CRISPR/Cas9 in Plant Biotechnology: Applications and Challenges. Biotechnologia 2022, 103, 81–93. [CrossRef]
Figure 1.
Developing plants resistant to fungal diseases. Conventional plant breeding (left panel); Mutagenesis (middle panel); and Genetic engineering and CRISPR-mediated gene editing technology (right panel) for improving fungal disease resistance in plants. In conventional breeding, developing fungal disease–resistant crops typically take 12–14 years (left panel), while marker-assisted selection (MAS) reduces this to 6–8 years (middle panel). Using mutagenesis or genetic engineering tools requires approximately 8–10 years (right panel). By contrast, CRISPR-mediated genome editing can shorten the development time to 4–6 years (right panel).
Figure 1.
Developing plants resistant to fungal diseases. Conventional plant breeding (left panel); Mutagenesis (middle panel); and Genetic engineering and CRISPR-mediated gene editing technology (right panel) for improving fungal disease resistance in plants. In conventional breeding, developing fungal disease–resistant crops typically take 12–14 years (left panel), while marker-assisted selection (MAS) reduces this to 6–8 years (middle panel). Using mutagenesis or genetic engineering tools requires approximately 8–10 years (right panel). By contrast, CRISPR-mediated genome editing can shorten the development time to 4–6 years (right panel).
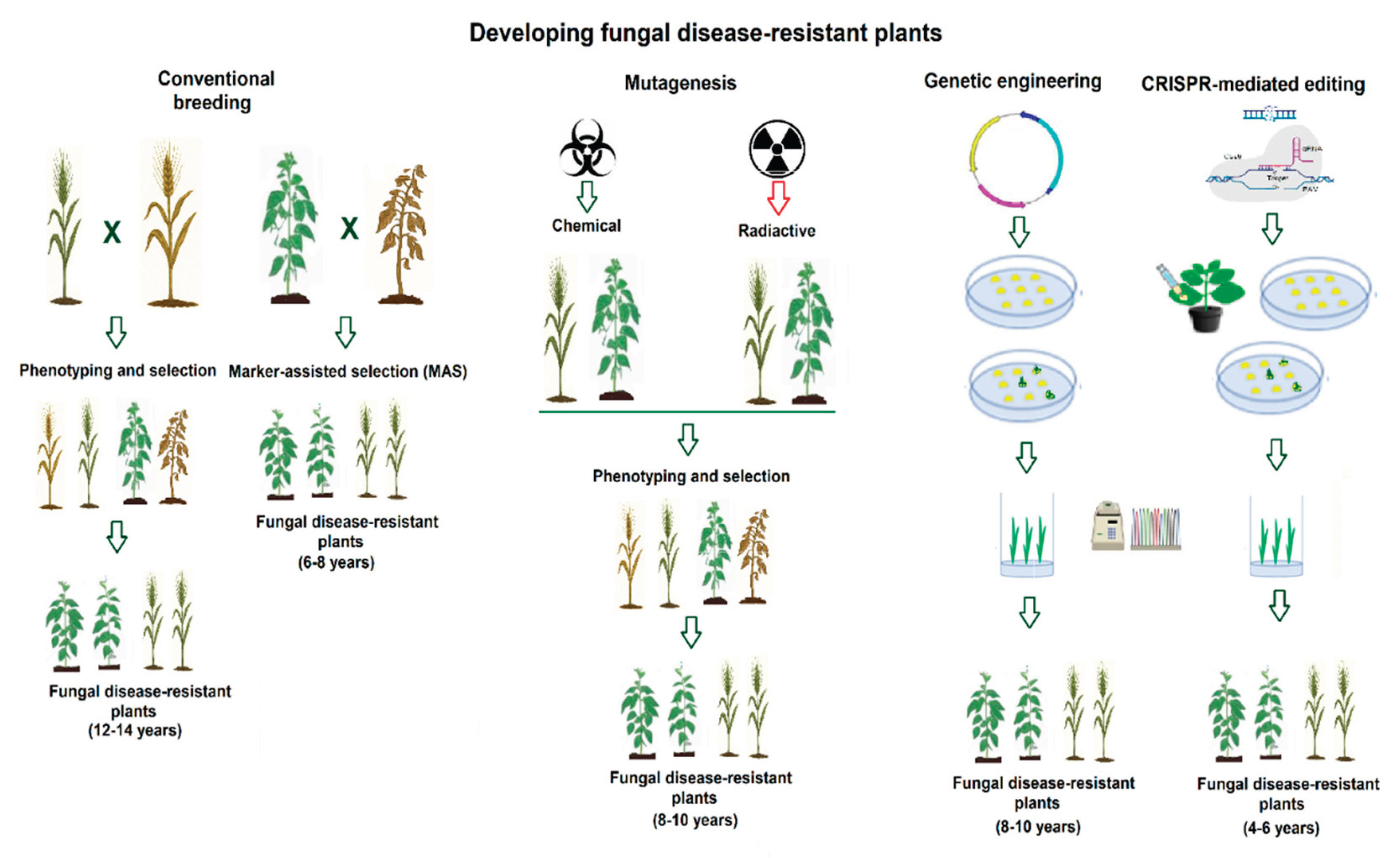
Table 3.
Using CRISPR-Cas technology to improve fungal-disease resistance in vegetables.
| Pathogen | Target Gene | Delivery Method | Type of Editing | References |
| Tomato | ||||
| Oidium neolycopersici | SlMlo1 |
Agrobacterium- mediated transformation |
Knockout | [72] |
| Oidium neolycopersici | SlMlo1, SlPelo |
Agrobacterium- mediated transformation |
Knockout | [73] |
| Oidium neolycopersici | PMR4 |
Agrobacterium- mediated transformation |
Knockout | [74] |
| Oidium neolycopersici | SlDND1 |
Agrobacterium- mediated transformation |
Knockout | [75] |
| Phytophthora infestans | SlPMR4 |
Agrobacterium- mediated transformation |
Knockout | [76] |
| Phytophthora infestans | miR482b, miR482c |
Agrobacterium- mediated transformation |
Knockout | [77] |
| Phytophthora capsici | SlDMR6-1 |
Agrobacterium- mediated transformation |
Knockout | [78] |
| Fusarium oxysporum | XSP10, SlSAMT |
Agrobacterium- mediated transformation |
Knockout | [79] |
| Fusarium oxysporum | SlPUB21, SlPUB17/SlPUB21 |
Agrobacterium- mediated transformation |
Knockout | [80] |
| Botrytis cinerea | SlPLC2 |
Agrobacterium- mediated transformation |
Knockout | [81] |
| Botrytis cinerea | SlPG2a, SlPL |
Agrobacterium- mediated transformation |
Knockout | [82] |
| Verticillium dahliae | SlWAT |
Agrobacterium- mediated transformation |
Knockout | [83] |
| Pepper | ||||
| Leveillula taurica | CaMLO2 | RNP | Knockout | [84] |
| Leveillula taurica | CaMLO2 | RNP | Knockout | [85] |
| Colletotrichum truncatum | CaERF28 | Agrobacterium-mediated transformation | Knockout | [86] |
| Eggplant | ||||
| Phytophthora infestans, Phytophthora capsici | SmDMR6 | Agrobacterium-mediated transformation | Knockout | [87] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



