
Submitted:
20 October 2025
Posted:
20 October 2025
You are already at the latest version
Abstract
A plant viral vector engineered from an in vivo infectious clone of zucchini yellow mosaic virus (Potyvirus cucurbitaflavitesselati; ZYMV) was used to express the human insulin in planta. The proinsulin gene was in frame inserted between the P1 and HC-Pro genes of the ZYMV vector. Six histidine residues and a NIa protease cleavage site were added at the C-terminal region of the inserts to facilitate detection and process of free form of the ex-pressed insulin, respectively. Also, N-terminal chloroplast signal peptide and C-terminal KDEL sequence was used to target insulin into chloroplast and endoplasmic reticulum, respectively. The infectious activity of the recombinant vectors was approved by rubbing the plasmid on Chenopodium quinoa and observing local lesions. A single local lesion was inoculated on zucchini squash leaves and these plants were sampled 5, 10, 15 and 20 days after inoculation. Analysis of inoculated plants by enzyme-linked immunosorbent assay (ELISA) and western blot indicated that insulin protein was expressed in Squash leaves. RT-PCR result showed the recombinant vector was remarkably stable in squash for seven serial passages and after 30 days. Also, using KDEL retention signal increased ex-pression level of recombinant protein significantly. This procedure provides a convenient and fast way for production of large quantities of insulin in planta.
Keywords:
zucchini yellow mosaic virus
; transient expression
; recombinant protein
1. Introduction
The growing needs in every field demand mass production of the desired product with minimal inputs. Likewise, the increasing demand for various pharmaceuticals has brought us to switch to a more efficient system. The increasing global incidence of diabetes mellitus necessitates the development of efficient and cost-effective methods for producing insulin, a critical therapeutic protein. Traditional methods of insulin production, relying on bacterial or yeast fermentation, have faced challenges related to cost, scalability, and potential immunogenicity [1,2]. Plant-based expression systems offer a promising alternative, providing a scalable, low-cost platform for the production of biopharmaceuticals, including therapeutic proteins like insulin [3,4]. Plants have been cultivated for ages to fulfill our basic needs majorly. But with the progress in genomics, proteomics, and metabolomics, plants are now being used as factories for the production of desired proteins, pharmaceuticals, vaccines, industrial enzymes, etc. They are the most cost-effective systems for the production of recombinant proteins. Various factors are considered before choosing a system, such as a cost-effectiveness, product safety, storage conditions, purification process, environmental concerns, and acceptance by the public. Unlike mammals, plants have a natural barrier against the ingression of a mammalian pathogen. Moreover, plant infecting viruses cannot infect during plant cell cultures by providing restrictions via plasmodesmata that restrict the route of virus movement [5]. Molecular Pharming, a term that appears in the literature in the 1980s to refer to the production of high-value compounds in transgenic animals, is now mainly used to refer to the production of pharmaceutically important and commercially valuable proteins in plants (Fischer and Emans 2000). Plant cells combine the advantage of a full posttranslational modification potential with simple growth requirements and basically unlimited scalability of whole plants in the field. Over the following years, a vast variety of pharmaceutical proteins were expressed in different plant species (with tobacco remaining the main production host) and targeted to different organelles or organs [6]. Plants and plant cells are versatile production systems, also allowing targeting of the recombinant proteins produced to different organs or subcellular compartments, thus allowing improved protection against proteolysis [7]. The use of plant-based transient expression for insulin production has been investigated, though challenges remain regarding protein accumulation, stability, and efficient downstream processing. Specifically, the proper folding and post-translational modification of human insulin, which is crucial for its biological activity, must be carefully considered within the plant system [8]. Furthermore, targeted subcellular expression of recombinant proteins can also influence protein accumulation levels and glycosylation patterns, which is particularly important for therapeutics [9]. Plant virus-based expression systems, particularly those employing transient expression, present several advantages for rapid and high-level recombinant protein production. These systems bypass the need for stable plant transformation, leading to faster production timelines and reduced regulatory hurdles [10]. Furthermore, viral vectors can be engineered to target specific subcellular compartments, allowing for protein accumulation in locations that optimize stability, processing, and potentially purification [11,12]. The use of plant viral vectors for the transient expression of heterologous proteins offers a useful tool for the large-scale production of proteins of pharmaceutical and industrial importance. The plant viral vectors provide a fast and efficient method for production of specific proteins in eukaryotic plant cells. Zucchini yellow mosaic virus (ZYMV; Potyvirus cucurbitaflavitesselati) causes one of the most devastating diseases in cucurbits including cucumber, squash, melon and water-melon. Similar to all potyviruses, the ZYMV genome consists of a positive single strand RNA molecule of 9.6 kb encapsidated in a filamentous particle. Viral RNA is translated into a large polyprotein that is proteolytically processed to 8–9 functional proteins by three virus-encoded proteases: P1, HC-Pro, and NIa [13]. The P1 and HC-Pro proteases are located at the N-terminal region of the polyprotein and catalyze auto proteolytic cleavages at their own C-termini. The NIa protease is responsible for cis and trans proteolytic cleavages of the remaining polyprotein [14].
In this study, a plant viral vector engineered from an in vivo infectious clone of ZYMV was used to express the human insulin in planta. This paper investigates the transient expression of human insulin in squash plants using ZYMV vector. We focus on the optimization of expression through different subcellular targeting strategies to enhance the accumulation of properly folded and potentially biologically active human insulin in planta. The study aims to contribute to the development of more efficient and cost-effective plant-based insulin production platforms by optimizing both expression levels and protein localization.
2. Results
2.1. Infectivity, Stability and Confirmation of the ZYMV Recombinants
Chlorotic local lesions similar to those induced by ZYMV wild type developed on inoculated leaves of quinoa 7–9 days after inoculation with the chimeric constructs (Figure 1-A). One week after inoculation, systemic symptoms of yellow mosaic were noticed on all squash plants. ZYMV recombinants in the infected plants (Figure 1-B) were confirmed by RT-PCR and serological assays. Also, INS expression and using signal peptides changed symptoms. The plants inoculated by ZINS showed leaf deformation and yellowing; inoculating of the ZChlINS changed leaf symptoms to severe mosaic with dark green part and thickened similar to those caused by ZChlINSKDEL. The ZINSKDEL symptom was yellowing with a few green islands.
The DNA fragments with about 493 bp length corresponding to the inserted INS ORF was amplified till the flowering stage about 30 days after initial inoculating and also after every successive trans-inoculation of active viral progeny on new host individuals by RT-PCR and western blotting analyses. RT-PCR result showed the recombinant vector was remarkably stable in squash for six serial passages (Figure 2). Sequenced RT-PCR product sampled from 6th passage didn’t show any change in amino acid sequence (data not shown).
2.2. Detection of ZYMV-Expressed INS
ZYMV-expressed INS containing a histidine tag was identified by immunoblotting using the His-MAb (Figure 3). Protein of 25 KDa was present in the crude extracts of zucchini squash. No serological reactions were detected from extracts of ZYMV wild type inoculated squash plants. The highest level of ZYMV-expressed INS was shown by ELISA by the treatment ZINSKDEL and at 15 and 10 dpi, respectively (Figure 3).
2.3. Purification of ZYMV-Expressed INS from the Infected Squash Plants
Using the affinity column chromatography, INS expressed by the ZYMV recombinants were purified from infected squash plants. After purification, samples were collected and the presence of expressed INS was monitored by SDS–PAGE coupled with immunoblotting. The 25 kDa protein corresponding to the ZYMV-expressed INS obtained in the soluble fractions was confirmed by immunoblotting using the His-Mab (Figure 4). The expressed INS ranging from 1 to 2 mg was purified from every 100 g tissues of squash plants inoculated with ZYMV recombinants as determined by the comparison with BSA in SDS-PAGE and by the software Spot Density of AlphaInnotech IS2000 (data not shown). Also, using KDEL retention signal increased expression level of recombinant protein and the highest and the lowest level of the INS expression was observed in the treatment inoculated by the ZINSKDEL and ZChlINS vectors, respectively.
3. Discussion
Virus-based vectors have been widely employed for heterologous protein expression and functional genomic studies in plants [15,16,17,18]. A diverse range of viruses, including tobamoviruses, potexviruses, potyviruses, necroviruses, hordeiviruses, criniviruses, comoviruses, geminiviruses, and caulimoviruses, have been successfully engineered into viral vectors for use in both model plants and economically important crops [11,17,19]. Among these, potyviruses have emerged as particularly versatile platforms for heterologous protein expression in plants [17,20,21,22]. Previous studies have identified three primary cleavage sites within potyviruses—the N-terminus of the P1 domain, the P1/HC-Pro junction, and the NIb/CP junction—as suitable insertion sites for foreign genes [23,24]. In this study, we engineered the P1/HC-Pro junction of Zucchini yellow mosaic virus (ZYMV) as a cloning site, flanked by P1 and NIa-Pro cleavage sequences, to facilitate the insertion of genes of interest (Figure 5).
The ZYMV-based vector demonstrated efficient expression of foreign genes throughout the plant, including upper leaves, stems, roots, and flowers (Figure 1). The foreign proteins remained stable and accumulated abundantly during viral replication and systemic spread in squash (Figure 1). Notably, the proinsulin gene remained detectable in squash plants even after six serial passages via mechanical inoculation (Figure 2), highlighting the stability and utility of this vector for foreign protein expression. Furthermore, the incorporation of the KDEL endoplasmic reticulum (ER) retention signal markedly increased proinsulin production at 15 days post-inoculation (Figure 3). These results highlight the critical role of signal peptide selection in optimizing recombinant protein expression systems. The enhanced accumulation of recombinant protein observed with the KDEL signal is likely attributable to its ability to direct proinsulin to the ER, thereby shielding it from N-terminal degradation and promoting its stability. This finding aligns with previous studies demonstrating the efficacy of ER retention signals in improving recombinant protein yields and underscores their utility in biotechnological applications [25,26].
Contrary to our expectations, the chloroplast signal peptide did not enhance recombinant proinsulin accumulation; instead, it resulted in lower protein levels compared to the ZINS treatment (Figure 3 and Figure 4). This outcome emphasizes the critical role of signal peptide selection in optimizing recombinant protein expression systems. Furthermore, while the chloroplast signal peptide alone proved less effective, the combination of both signals (ZChlINSKDEL) produced results comparable to ZINSKDEL, albeit with distinct symptom profiles. These findings collectively highlight the importance of carefully selecting and evaluating signal peptides to achieve optimal recombinant protein expression and stability.
The ZYMV-based vector also represents a valuable tool for functional genomics studies. In this work, we utilized the vector to overexpress proinsulin in squash, demonstrating its potential for characterizing viral genes and advancing research on viral pathogenicity. Plants offer distinct advantages as bioreactors, including cost-effectiveness, scalability, and environmental sustainability [6,27]. Viral-based expression systems further expedite protein production without the need for genetic transformation, making them particularly attractive for rapid protein synthesis. Additionally, plants can produce a wide range of proteins with diverse biological activities, including pharmaceuticals, antioxidants, antimicrobials, antibodies, and insecticidal proteins [6,28,29]. The high accumulation of recombinant proteins in ZYMV-infected plants highlights the potential of this system for producing valuable proteins and secondary metabolites in squash.
Despite these advantages, the utility of some viral vectors remains limited due to severe symptom induction and low infection rates. In this study, we demonstrated that ZYMV can serve as a highly efficient expression vector for recombinant protein production. Moreover, we achieved successful proinsulin expression throughout the plant, paving the way for further investigations into the functional roles of genes involved in plant-pathogen interactions, such as anti-nematode, antifungal, or antibacterial proteins. Future studies could also explore the application of ZYMV-based vectors for evaluating virulence genes and symptom determinants. Additionally, advanced vector systems, including virus-induced gene silencing (VIGS), virus-mediated genome editing, and miRNA expression, could be developed to expand the utility of viral vectors in plant research.
In summary, this study establishes the ZYMV-based vector as a robust platform for gene expression, functional genomics, and pathogenicity studies in squash. The findings provide significant insights into the infectivity, stability, and expression of recombinant proteins in a plant-based system. Chlorotic local lesions on inoculated quinoa leaves and systemic yellow mosaic symptoms in squash plants confirmed the pathogenicity of the engineered virus while maintaining its functionality as a gene expression vector. RT-PCR and serological assays confirmed the stability of ZYMV recombinants over six serial passages and 30 days (Figure 2), with consistent proinsulin expression observed throughout this period.
The market for therapeutic and recombinant proteins continues to grow, and plant-based expression systems offer unique advantages, including low production costs, high scalability, and enhanced safety due to the absence of human pathogens and endotoxins [27]. While plant systems are less advanced than microbial and mammalian platforms in terms of regulatory approval and good manufacturing practices, companies like Medicago Inc. and Icon Genetics have made significant strides in this area [30]. For instance, Medicago Inc. recently demonstrated the potential of transient plant systems for producing vaccines against SARS-CoV-2 and influenza, with consistent immunogenicity observed in clinical trials [31,32]. These advancements underscore the potential of plant-based systems for industrial and therapeutic applications.
Finally, our work highlights the potential of ZYMV as an innovative vector for recombinant protein expression in squash, offering a valuable platform for both basic research and biotechnological applications. Future studies should focus on optimizing vector design, exploring additional signal peptides, and expanding the range of expressible proteins to fully realize the potential of plant-based viral vectors.
4. Materials and Methods
4.1. Plant Materials and DNA Sources
Quinoa (Chenopodium quinoa L.) and squash (Cucurbita pepo L.) plants were grown in a controlled greenhouse at 25 ºC and were used as local lesion and systemic hosts, respectively. A previously constructed infectious plant virus clone, p35SZYMVGFPhis3, contains the full-length cDNA of a Taiwan isolate of ZYMV, specifically the TW-TN3 strain provided by Lin et al. (2002) was used as an expression vector. This clone is driven by the cauliflower mosaic virus (CaMV) 35S promoter including a green fluorescent protein (GFP) reporter gene located either between the P1 and HC-Pro genes [33]. It was engineered as an in vivo viral vector to express the proinsulin in squash plants.
The proinsulin gene (INS) was in frame inserted between the P1 and HC-Pro genes of the ZYMV vector. Six histidine residues and a NIa protease cleavage site were added at the C-terminal region of the inserts to facilitate detection and process of free form of the expressed insulin, respectively. Also, N-terminal chloroplast signal peptide and C-terminal KDEL sequence were used to target insulin into chloroplast and endoplasmic reticulum, respectively. Appropriate primers for adding signal peptides were designed with SphI and KpnI restriction sites in the forward and reverse primer, respectively (Table 1). The resultant clones were designated as ZINS, ZChlINS (harboring chloroplast signal peptide at N-terminal), ZINSKDEL (with SEKDEL signal peptide at C-terminal), and ZChlINSKDEL (containing both chloroplast and SEKDEL signal peptides at N- and C-terminal, respectively) (Figure 5, Table 2).
4.2. Polymerase Chain Reaction
(PCR) was performed in a total 50 μl final volume, using 2mM of each deoxyribonucleotide triphosphate (dNTPs), 0.4µM of each primer, 1mM Mg2+, and 0.5 unit of KOD plus DNA polymerase enzyme. Thermocycler was programmed for one cycle at 94 °C for 5 min, followed by 35 cycles at 94 °C for 30 second; 55 °C for 30 second; 68 °C for 40 second and one cycle at 68 °C for 3 min as a final extension. The resulted band was cleaned up using the DNA clean up kit (GeneMark).
4.3. Digestion and Cloning of INS to ZYMV Vector
Individual INS ORFs were directly inserted into the cloning site between the P1 and HC-Pro genes of the ZYMV vector (named ZINS) as shown in Figure 1. The purified INS ORF was double digested by SphI/KpnI enzyme (Invitrogen). The same restriction enzymes were also used to remove the GFP ORF from the ZYMV vector p35SZYMVGFPhis3, then digested products were run on 0.8% agarose gel and the resulted band was purified using the agarose gel DNA extraction kit (GeneMark). The extracted vector and insert were used in the ligation reaction in a ratio 1:8 respectively for 30 min. The ligation reaction product was transformed in Escherichia coli XL1 using heat shock method [34]. Resulted colony was screened by with colony PCR using INS (pINS-SN) and ZYMV (mz1155) specific forward and reverse primers that amplify a DNA fragment with 601-639 bp length (depends on the signal type). Two positive colonies were selected for plasmid purification.
4.4. Infectivity Assay of Chimeric Constructs and RT-PCR Confirmation
The infectious activity of the recombinant vectors was approved by rubbing the plasmid on quinoa and observing local lesions. Plasmids of the individual chimeric constructs were isolated by the mini-prep plasmid purification kit (GeneMark), and mechanically rubbed with a cotton stick on leaves (10µg in 10µl per leaf) of local lesion host quinoa previously dusted with 600 mesh carborundum. Individual lesions were mechanically transferred to the systemic host plant zucchini squash at the stage of two-cotyledon and these plants were sampled 5, 10, 15 and 20 days after inoculation. Total RNAs were extracted from the infected squash leaves using the TRIzol isolation system (Invitrogene). Specific primer pairs pINS-SN and mINS-KN (Table 2) were used to amplify the inserted INS ORF by RT-PCR. Plants inoculated by ZYMV wild type (WT) was used as negative control. Amplified DNA fragments were analyzed by electrophoresis in 1% agarose gels to confirm the presence of the insert.
4.5. Stability Assay of Recombinants in the Host Plants
The stability of ZYMV containing INS (ZINS) was assessed by successive passages of recombinant from infected plant tissues. Recombinant viruses in the infected squash were mechanically transferred to healthy plants at 10 days intervals. After several passages, the leaf tissues collected from the inoculated plants were analyzed by western blotting and RT-PCR with appropriate antisera and primer pairs, respectively. To further monitor the possible deletion of INS ORF from ZINS progeny-infected hosts, the amplified DNA fragment was sequenced and the translated amino acid sequence was analyzed by the program of BioEdit version 7.0.0 [17].
4.6. Immunoassay Tests (ELISA, Western Blot)
Immunoblotting [35] and indirect enzyme-linked immunosorbent assays (ELISA) [36] were utilized to quantitatively detect the proinsulin expressed in the infected squash plants at 5, 10, 15, and 20 days post-inoculation (dpi), respectively. For immunoblotting, total proteins were extracted from leaf tissues infected with recombinant viruses. For this purpose, 0.5 g of the leaf tissue was frozen in liquid nitrogen, then grinded into a fine powder. Next, 2 mL of dissociation buffer was added, which contained 100 mM Tris-HCl (pH 7.2), 2% β-mercaptoethanol, 10% sucrose, 0.005% bromophenol blue, and 10 mM EDTA. The mixture was then denatured by boiling for five minutes. After a brief centrifugation, the supernatant was separated using a 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS–PAGE) and subsequently transferred to nitrocellulose membranes which were blocked with 5% (w/v) non-fat dry milk powder in the PBST buffer. In indirect ELISA, polystyrene microtitration plates (Greiner Bio-One, Frickenhausen, Germany) were coated with leaf crude extracts diluted 100-fold in a coating buffer consisting of 50 mM sodium carbonate, pH 9.6, and containing 0.01% sodium azide. In both immunological assays, we used a monoclonal antibody targeting the histidine tag (His-MAb from Amersham Pharmacia Biotech, Buckinghamshire, England). The antibody was diluted to 1:5000 to detect the histidine-tagged INS. The alkaline phosphatase (AP)-conjugated goat anti-mouse and goat anti-rabbit immunoglobulin G (IgG) from Jackson ImmunoResearch Laboratories (West Grove, PA) were used at a dilution of 1:5000 to react with the monoclonal antibody. The reactions were visualized by adding a chromogenic substrate consisting of nitro-blue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate para toluidine salt in a solution of 100 mM NaCl, 5 mM MgCl2, and 100 mM Tris–HCl (pH 9.5) for immunoblotting. For indirect ELISA, the substrate solution was prepared by dissolving AP substrate tablets (Sigma 104; Sigma-Aldrich Fine Chemicals, Milwaukee, WI) in a substrate buffer containing 9.7% diethanolamine and 0.02% sodium azide (pH 9.8) in a final concentration of 0.5 mg/mL. The absorbance at 405 nm was measured for 1 hour after adding the substrate using an EL×800 universal microplate reader (Bio-Tek Instruments, Winooski, VT, USA). The samples were considered positive if there was clear development of color and the absorbance readings were at least three times higher than those of healthy samples. ELISA absorbance data were subjected to a completely randomized design with four replicates, and the means were normalized using the Duncan test. The relevant statistical analyses and histograms were generated using MicrosoftTM Excel® software.
4.6. Purification of the Expressed INS
The affinity chromatography described [37] was modified for purification of the ZYMV-expressed INS from infected plants. Fifteen days after inoculation with the ZYMV recombinants, 100 g infected zucchini squash leaves were harvested and ground in 300 ml of buffer A [50mM Tris–HCl, pH 8.0, 15mM MgCl2, 10mM KCl, 20% (v/v) glycerol, 0.05% β-mercaptoethanol, and 0.1mM phenylmethylsulphonyl fluoride (PMSF)] in a blender. Extracts were clarified by centrifugation at 20000×g for 10 min, and supernatant filtered through Miracloth (Calbiochem, La Jolla, CA). The filtrate, treated with 2% Triton X-100 at 4 ◦C for 30 min, and approximately 1ml of Ni2+-NTA resin (Ni-NTA, Invitrogen), pre-equilibrated in buffer B [50mM Tris–HCl, pH 8.2, 15mM MgCl2, 20% (v/v) glycerol, 0.05% β-mercaptoethanol, and 0.1mM PMSF], was added. The mixture was gently stirred for 1 h at 4 ◦C and loaded onto a column. After allowing the resin to settle, the unbound material was collected and the resin was washed with equal volume of buffer B. The proteins bound to the resin were eluted with 6 ml of buffer B containing 250mM imidazole in three steps. Equal volumes (5 µl for each well) of each fraction obtained from each purification step were analyzed using His-MAbs in immunoblotting. The amounts of purified proteins were estimated by the comparison with bovine serum albumin (BSA) in SDS–PAGE, and estimated by the software Spot Density of AlphaInnotech IS2000 (Alpha Innotech Corporation).
5. Conclusions
The use of plants as hosts for the overexpression of foreign proteins is still a developing field. Many choices are available that affect the rates of transcription, translation, and protein accumulation based on the type of expression desired. The present study was performed in order to investigate whether insulin protein could be successfully expressed and targeted transiently in a plant-virus expression system. This procedure provides a convenient and fast way for production of large quantities of insulin in planta. The system also has a potential for production of other proteins of interest in cucurbits to use as immunogen to produce antiserum or use for other purposes.
In conclusion, plant-based expression systems for biologics represent both significant progress and ongoing challenges. Success in this field requires a deep understanding of molecular processes, cellular environments, and regulatory frameworks. With the integration of diverse perspectives and advanced technologies, plant-based biologics are poised to play a transformative role in medicine, industry, and agriculture. As the potential of these systems continues to unfold, they are set to become an indispensable tool in shaping the future of biologics production.
References
- Walsh, G.; Walsh, E. Biopharmaceutical benchmarks 2022. Nat. Biotechnol. 2022, 40, 1722–1760.
- Baeshen, N.A.; Baeshen, M.N.; Sheikh, A.; Bora, R.S.; Ahmed, M.M.M.; Ramadan, H.A.; Saini, K.S.; Redwan, E.M. Cell factories for insulin production. Microb. Cell Factories 2014, 13, 1–9.
- Jadhav, R.R.; Khare, D. Green biotherapeutics: overcoming challenges in plant-based expression platforms. Plant Biotechnol. Rep. 2024, 1–22.
- Buyel, J.F. Plant molecular farming–integration and exploitation of side streams to achieve sustainable biomanufacturing. Front. Plant Sci. 2019, 9, 1893.
- Sethi, L.; Kumari, K.; Dey, N. Engineering of plants for efficient production of therapeutics. Mol. Biotechnol. 2021, 63, 1125–1137.
- Gaikwad, B.; Wagh, N.; Lakkakula, J. Current scenario of recombinant proteins: extraction, purification, concentration, and storage. In Fundamentals of Recombinant Protein Production, Purification and Characterization; Elsevier: 2025; pp. 173–189.
- Rozov, S.M.; Deineko, E.V. Increasing the efficiency of the accumulation of recombinant proteins in plant cells: the role of transport signal peptides. Plants 2022, 11, 2561.
- Lee, J.; Lee, S.-K.; Park, J.-S.; Lee, K.-R. Plant-made pharmaceuticals: exploring studies for the production of recombinant protein in plants and assessing challenges ahead. Plant Biotechnol. Rep. 2023, 17, 53–65.
- Eidenberger, L.; Kogelmann, B.; Steinkellner, H. Plant-based biopharmaceutical engineering. Nat. Rev. Bioeng. 2023, 1, 426–439.
- Mahmood, M.A.; Naqvi, R.Z.; Rahman, S.U.; Amin, I.; Mansoor, S. Plant virus-derived vectors for plant genome engineering. Viruses 2023, 15, 531.
- Song, S.-J.; Diao, H.-P.; Guo, Y.-F.; Hwang, I. Advances in Subcellular Accumulation Design for Recombinant Protein Production in Tobacco. BioDesign Res. 2024, 6, 0047.
- Mardanova, E.S.; Vasyagin, E.A.; Ravin, N.V. Virus-like Particles Produced in Plants: A Promising Platform for Recombinant Vaccine Development. Plants 2024, 13, 3564.
- Wang, D.; Li, G. Biological and molecular characterization of Zucchini yellow mosaic virus isolates infecting melon in Xinjiang, China. Can. J. Plant Pathol. 2017, 39, 48–59.
- Bubici, G.; Navarro, B.; Carluccio, A.V.; Ciuffo, M.; Di Serio, F.; Cillo, F. Genomic sequence variability of an Italian Zucchini yellow mosaic virus isolate. Eur. J. Plant Pathol. 2020, 156, 325–332.
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Viral vectors for the expression of proteins in plants. Curr Opin Biotechnol 2007, 18, 134–141, doi:S0958-1669(07)00033-X [pii].
- Zaidi, S.S.-e.-A.; Mansoor, S. Viral vectors for plant genome engineering. Front. Plant Sci. 2017, 8, 539.
- Yang, Y.-Z.; Xie, L.; Gao, Q.; Nie, Z.-Y.; Zhang, D.-L.; Wang, X.-B.; Han, C.-G.; Wang, Y. A potyvirus provides an efficient viral vector for gene expression and functional studies in Asteraceae plants. Plant Physiol. 2024, 196, 842–855.
- Nassaj Hosseini, S.M.; Shamsbakhsh, M.; Yeh, S.D. Co-Infection of Cucumber Mosaic Virus Could Stabilize Its Recombinant Coat Protein Expressed by Zucchini Yellow Mosaic Virus. Iran. J. Biotechnol. 2025, 23, 92–101.
- Gupta, S.; Mishra, P.; Mishra, P.; Tewari, V.; Pandey, S. Transgenic animals and plants: application and future scope. In Medicinal Biotechnology; Elsevier: 2025; pp. 61–77.
- Hosseini, S.M.N.; Bakhsh, M.S.; Salmanian, A.H.; Ye, S.D. Expression and purification of movement protein of Cucumber green mottle mosaic virus using a plant-virus expression system. 2013.
- Song, E.G.; Ryu, K.H. A pepper mottle virus-based vector enables systemic expression of endoglucanase D in non-transgenic plants. Arch. Virol. 2017, 162, 3717–3726.
- Mei, Y.; Beernink, B.M.; Ellison, E.E.; Konečná, E.; Neelakandan, A.K.; Voytas, D.F.; Whitham, S.A. Protein expression and gene editing in monocots using foxtail mosaic virus vectors. Plant Direct 2019, 3, e00181.
- German-Retana, S.; Candresse, T.; Alias, E.; Delbos, R.-P.; Le Gall, O. Effects of green fluorescent protein or β-glucuronidase tagging on the accumulation and pathogenicity of a resistance-breaking Lettuce mosaic virus isolate in susceptible and resistant lettuce cultivars. Mol. Plant-Microbe Interact. 2000, 13, 316–324.
- Arazi, T.; Slutsky, S.G.; Shiboleth, Y.M.; Wang, Y.; Rubinstein, M.; Barak, S.; Yang, J.; Gal-On, A. Engineering zucchini yellow mosaic potyvirus as a non-pathogenic vector for expression of heterologous proteins in cucurbits. J. Biotechnol. 2001, 87, 67–82.
- Nykiforuk, C.L.; Boothe, J.G.; Murray, E.W.; Keon, R.G.; Goren, H.J.; Markley, N.A.; Moloney, M.M. Transgenic expression and recovery of biologically active recombinant human insulin from Arabidopsis thaliana seeds. Plant Biotechnol. J. 2006, 4, 77–85.
- Boyhan, D.; Daniell, H. Low--cost production of proinsulin in tobacco and lettuce chloroplasts for injectable or oral delivery of functional insulin and C--peptide. Plant Biotechnol. J. 2011, 9, 585–598.
- Jin, C.; Hinterdorfer, P.; Lee, J.H.; Ko, K. Plant production systems for recombinant immunotherapeutic proteins. Plant Biotechnol. Rep. 2025, 19, 1–14.
- Burlec, A.F.; Pecio, Ł.; Mircea, C.; Cioancă, O.; Corciovă, A.; Nicolescu, A.; Oleszek, W.; Hăncianu, M. Chemical profile and antioxidant activity of Zinnia elegans Jacq. fractions. Molecules 2019, 24, 2934.
- Venkataraman, S. Plant molecular pharming and plant-derived compounds towards generation of vaccines and therapeutics against coronaviruses. Vaccines 2022, 10, 1805.
- Moustafa, K.; Makhzoum, A.; Trémouillaux-Guiller, J. Molecular farming on rescue of pharma industry for next generations. Crit. Rev. Biotechnol. 2016, 36, 840–850.
- Ward, B.J.; Gobeil, P.; Séguin, A.; Atkins, J.; Boulay, I.; Charbonneau, P.-Y.; Couture, M.; D’Aoust, M.-A.; Dhaliwall, J.; Finkle, C. Phase 1 randomized trial of a plant-derived virus-like particle vaccine for COVID-19. Nat. Med. 2021, 27, 1071–1078.
- Venkataraman, S.; Khan, I.; Habibi, P.; Le, M.; Lippert, R.; Hefferon, K. Recent advances in expression and purification strategies for plant made vaccines. Front. Plant Sci. 2023, 14, 1273958.
- Hsu, C.H.; Lin, S.S.; Liu, F.L.; Su, W.C.; Yeh, S.D. Oral administration of a mite allergen expressed by zucchini yellow mosaic virus in cucurbit species downregulates allergen-induced airway inflammation and IgE synthesis. J Allergy Clin Immunol 2004, 113, 1079–1085. [CrossRef]
- Sambrook, J.; Russell, D.W. Detection of DNA in agarose gels. Mol. Cloning A Lab. Man. ,(3rd Ed.) Cold Spring Harb. Lab. Press New York 2001, 5–14.
- Gooderham, K. Transfer techniques in protein blotting. Methods Mol Biol 1984, 1, 165–178. [CrossRef]
- Wang, M.; Gonsalves, D. ELISA detection of various tomato spotted wilt virus isolates using specific antisera to structural proteins of the virus. 1990.
- Gal-On, A.; Canto, T.; Palukaitis, P. Characterisation of genetically modified cucumber mosaic virus expressing histidine-tagged 1a and 2a proteins. Arch. Virol. 2000, 145, 37–50.
Figure 1.
Evidence of infection by the appearance of local lesion (part A) and systemic symptoms on the leaves of quinoa and squash plants respectively, by the recombinants ZINS vector. The treatment names are shown in each image below.
Figure 1.
Evidence of infection by the appearance of local lesion (part A) and systemic symptoms on the leaves of quinoa and squash plants respectively, by the recombinants ZINS vector. The treatment names are shown in each image below.

Figure 2.
Examining of stability of ZINS after six passages. A. RT-PCR amplification profile from squash plants infected with ZINS vectors after 1-6 serial passages corresponding to lane 1-6; lane 7: negative control (ZYMV-WT).
Figure 2.
Examining of stability of ZINS after six passages. A. RT-PCR amplification profile from squash plants infected with ZINS vectors after 1-6 serial passages corresponding to lane 1-6; lane 7: negative control (ZYMV-WT).
Figure 3.
ELISA results using His-monoclonal antibody for each treatment over a period of 25 days after inoculation.
Figure 3.
ELISA results using His-monoclonal antibody for each treatment over a period of 25 days after inoculation.
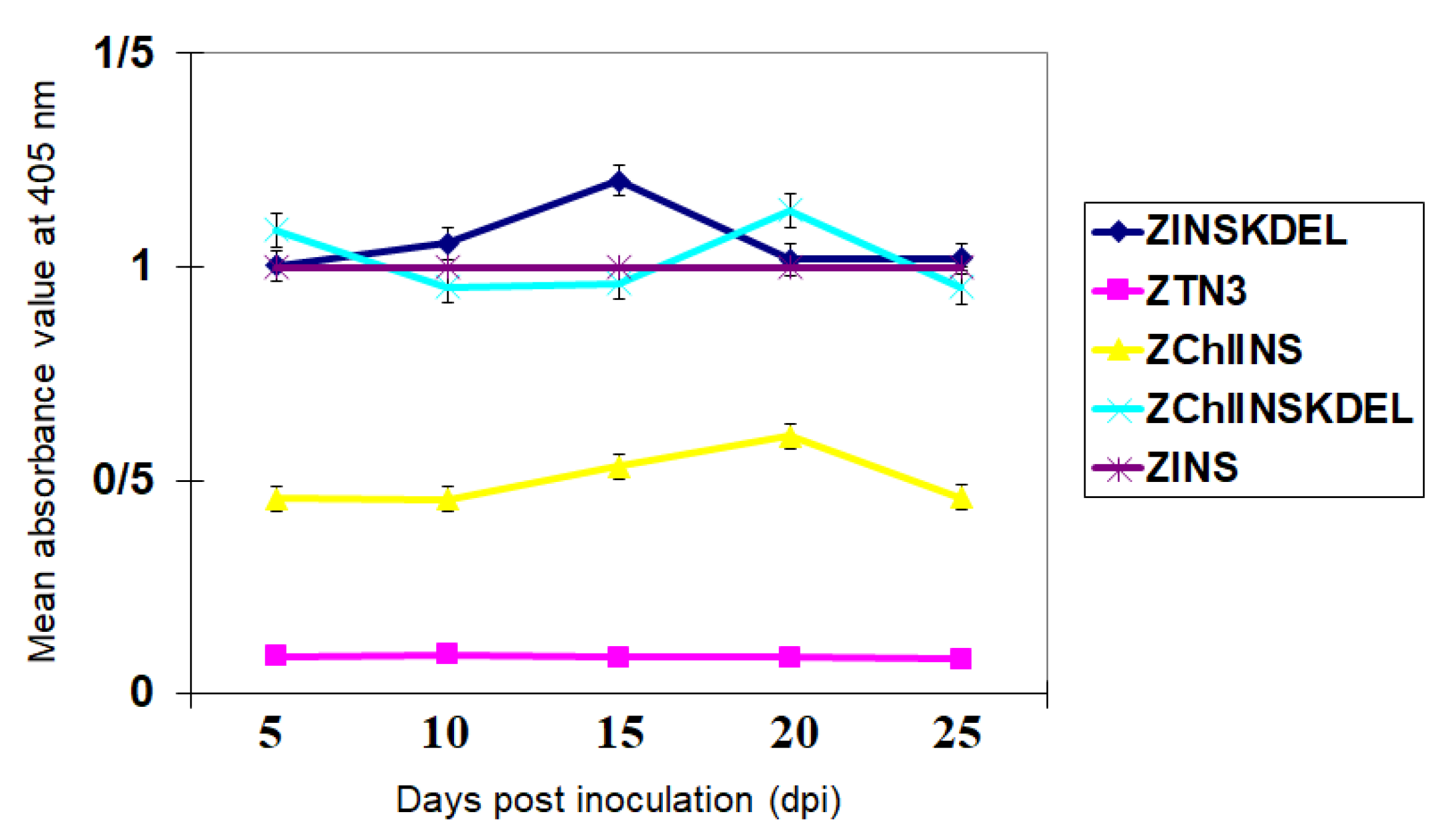
Figure 4.
Immunoassay results using antibody against His-tag for each treatment over a period of 15 days after inoculation.
Figure 4.
Immunoassay results using antibody against His-tag for each treatment over a period of 15 days after inoculation.
Figure 5.
The schematic diagram of the genomic organization of the ZYMV vector along with the enzymatic restriction map.
Figure 5.
The schematic diagram of the genomic organization of the ZYMV vector along with the enzymatic restriction map.

Table 1.
Primers used in this study along with their applications and cleavage sites.
| Manufacturing company | application | Restriction site | Primer length | Sequence (restriction sites have been underlined) | Primer name | ||
| MDBio | To produce complementary DNA | - | 20 | ACTTTGCACACATGATCTGG | mz1155 | ||
| MDBio | To run RT-PCR | - | 22 | AGAAAGTGGTGCAAAAGCAAAC | pZ629 | ||
| Genomics | To amplify Pro-Insulin with SphI restriction site | SphI | 33 | CACCATGGCATGCATGGAAGCGGGATTCAACCA | pINS-SN | ||
| Genomics | To amplify Pro-Insulin with KpnI restriction site | KpnI | TAGCTAGCGGTACCGTTGCAGTAGTTTTCCAG | mINS-KN | |||
| Genomics | To add chloroplast signal peptide | SphI | 48 | ATGCATGCACCTGCGCATTGGACTCTTCCATAGTGGTCAACTTCGCTA | mzp1Chltag | ||
| Genomics | To screen recombinants with chloroplast signal peptide | - | 21 | GGAAGAGTCCAATGCGCAGGT | pchltag | ||
| MDBio | To add and screen SEKDEL signal peptide | NcoI | 47 | TTTTCCATGGAAGACTGGTGAAACACATGTAGCTCATCTTTCTCAGA | mKDELnss |
Table 2.
Abbreviated names of viral clones along with the type of foreign gene and the signal peptide and the tag used.
Table 2.
Abbreviated names of viral clones along with the type of foreign gene and the signal peptide and the tag used.
| Abbreviate name | Foreign gene | Tag type and its position | Signal peptide type and its position |
| ZINS | Proinsulin | His tag at the C-termini | - |
| ZChlINS | Proinsulin | His tag at the C-termini | Chloroplast signal peptide at the N-termini |
| ZChlINSKDEL | Proinsulin | His tag at the C-termini | Chloroplast signal peptide at the N-termini and SEKDEL at the C-termini |
| ZINSKDEL | Proinsulin | His tag at the C-termini | SEKDEL at the C-termini |
| ZTN3 | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



