Submitted:
19 October 2025
Posted:
21 October 2025
You are already at the latest version
Abstract
The visual system's complexity and richness are fundamental to human experience. Underscoring this complexity is a network of diverse cells working in concert to generate coherent visual experience. Despite years of progress in understanding the functional significance of this heterogeneity, many aspects of the intricate workings of the visual system remain enigmatic. This review focuses on the various classes of retinal ganglion cells (RGCs), their receptive fields (RFs), and their role in conscious vision, particularly within the primate visual system. By examining the anatomy and physiology of RGCs and their central projections, particularly to the dorsal lateral geniculate nucleus (LGN), this paper aims to elucidate the contributions of the different RGC types to visual perception. The visual system's complexity and richness are fundamental to human experience. Retinal ganglion cells (RGCs), a diverse population of neurons that form the output layer of the retina, encode distinct aspects of visual information provided to the brain, thereby supporting the translation of the visual scene at the retina to visual perception in the cortex. This review explores the anatomical and functional diversity of RGC types, emphasizing their receptive field properties and central projections, particularly to the dorsal lateral geniculate nucleus (LGN). The contributions of different RGC classes that support parallel processing of spatial resolution, color, motion, and luminance are discussed, along with non-image-forming functions. Special attention is given to low-density, koniocellular-projecting RGCs, whose varied roles are only beginning to be understood. By integrating recent advances in anatomy, physiology, and connectivity, this review underscores how a comprehensive understanding of RGC diversity is critical for unraveling visual processing and informing future approaches to visual restoration and prosthetic development.
Keywords:
retinal ganglion cells
; lateral geniculate nucleus
; primate visual system
; visual perception and restoration
1. Introduction
Sighted individuals benefit from constant access to color, form, texture, and motion information provided by our eyes. However, in the United States alone, 12 million people over the age of 40 suffer from visual impairment (CDC, 2022). The World Health Organization reports over 2.2 billion people globally with eye and vision problems (WHO, 2022). Remarkable bioengineering progress has been made to help restore vision to the visually impaired, including corneal implants (reviewed in Ludwig et al., 2018), STEM cell therapy (reviewed in Singh et al., 2020), genetic engineering (reviewed in Blackshaw & Sanes, 2021), and visual prostheses (reviewed in Mirochnik & Pezaris, 2019). All these techniques rely on deep understanding of visual physiology. Therefore, this review will focus on the classes of retinal ganglion cells (RGC) which contribute to conscious vision, their receptive fields, functional properties, and central projections to identify hypotheses concerning their functional visual contributions. Underlying this effort is our motivation that a greater understanding of RGC diversity can inform better and more comprehensive visual restoration techniques. This review aims to update a 2004 chapter by Dacey (Dacey, 2004) concerning the same topic.
Figure 1.
Retinothalamic projections in the primate retina.
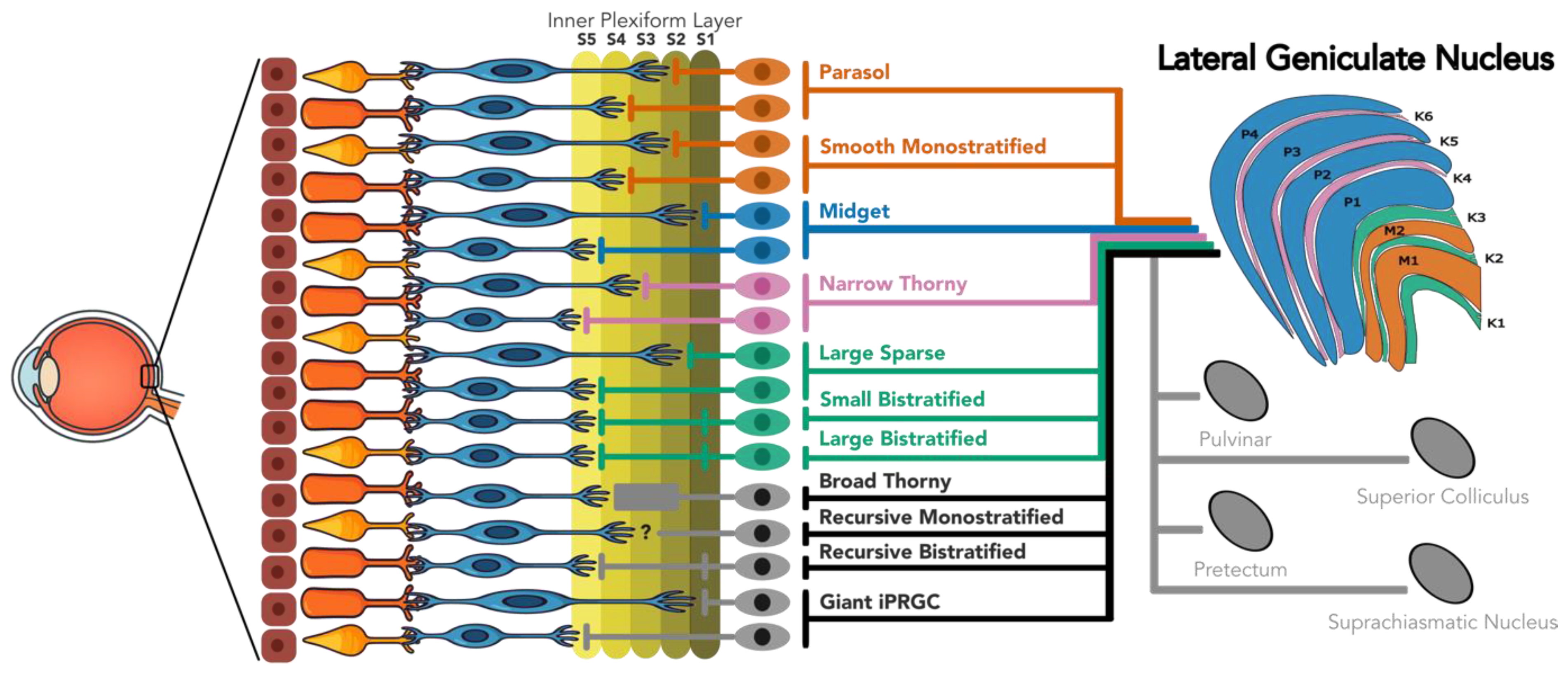
The focus of this review will be the primate visual system, with occasional explicitly mentioned comparisons to other animal models, such as mice, rabbits, or cats. Anatomical and physiological similarities between visual systems in the higher primates makes this research the best source of information to guide vision restoration in humans. Most vision research with primates uses Rhesus macaques, as their visual systems closely resemble that of humans. Recent research by Yan et al. (2020) found that over 90% of human retinal cells correspond transcriptomically to those identified in macaque (Yan et al., 2020). Psychophysics results comparing human and macaque vision have come to similar conclusions (reviewed in Horwitz, 2015; Jacobs, 1993; Lee, 1996). A model with recent interest in vision research is the marmoset, a New-World monkey with a visual system similar to humans, but slightly simpler and more anatomically accessible than in the macaque (Solomon & Rosa, 2014). Marmoset and macaque research will be explicitly differentiated, where appropriate, with the distinction becoming most relevant in the koniocellular-projecting RGC types.
As the primary gatekeeper of conscious vision and the central target for almost every RGC type, the LGN will play an important role in this review. Not only do most RGCs project to the LGN, but most RGC types have no other central target (Dacey, 2004). However, non-LGN retinal targets will be discussed as many of them may play small roles in conscious vision (Ahmadlou et al., 2018; Bridge et al., 2016; Warner et al., 2015; B. J. White et al., 2017). These centers include the superior colliculus, the pulvinar, the suprachiasmatic nucleus, and other sub-cortical nuclei. However, RGC types which show no evidence of LGN projection will not be discussed, as it is assumed that their roles are largely modulatory.
2. Background
Anatomically and physiologically classifying RGC types
Understanding the functional significance of RGC morphologies first requires a brief account of the broader organization of the visual system. Phototransduction for visual experience starts at the outermost retinal layer where photoreceptors – rods and cones – capture photons and convert their energy into electrical signals. Those electrical signals are passed through bipolar cells to retinal ganglion cells (RGC) in the retinal inner plexiform layer (IPL) which then project to the brain via the optic nerve (Wässle, 2004). However, the retina serves a much greater function than merely to transduce and relay visual information. Information passed through the retina undergoes significant processing through parallel information streams (Schiller, 2010). In addition to four distinct types of photoreceptors (one rod type and three cone types), 11 types of bipolar cells (Grünert & Martin, 2020) and as many as 17-22 morphologically distinct RGC types have been described in the primate retina (Dacey, 2004; Field & Chichilnisky, 2007; Yamada et al., 2005). Studies examining molecular clusters of RGC types (via transcriptomic analysis) have shown similar numbers of clusters (Peng et al., 2019; Shekhar & Sanes, 2021; Yan et al., 2020; Zhang et al., 2024). Architecturally perpendicular to the photoreceptor-bipolar-RGC flow of information are two distinct types of horizontal cells (Wässle, Boycott, et al., 1989) and as many as 30-60 morphological types of amacrine cells (Kolb et al., 1992; Mariani, 1990) which integrate and modulate signaling between bipolar cells and RGCs.
The receptive field (RF) of RGCs is the stimulus-to-response relationship between spatiotemporal patterns of light entering the retina and RGC spikes leaving the retina (Wienbar & Schwartz, 2018). Most RGC RFs can be classified in terms of size (i.e., area of coverage across the visual field), polarity of light response (on, off, on-off, suppressed-by-contrast) (Sanes & Masland, 2015), and kinetics of light response (e.g., transient or sustained) (Gouras, 1968). The properties of RGC RFs go far beyond just these three classifications, including chromatic preference, direction selectivity, orientation selectivity, and contrast sensitivity (Baden et al., 2016; Wienbar & Schwartz, 2018). The manifold RGC types and their varied RF properties form the basis of the rich visual world we experience.
Two anatomical features of different RGC types reveal important clues as to RF properties and their functional significance: stratification depth, dendritic tiling radius, and density. The first is depth of stratification in the IPL. As proposed by Cajal (1972), the IPL can be divided into five distinct strata. Further, these strata can be functionally grouped into two sublaminae, with strata 1 and 2 making up sublamina a and strata 3-5 making up sublamina b. Each sublamina contains a distinct population of synapses; on-center pathways tend to synapse in the inner IPL (sublamina b) while off-center pathways tend to synapse in the outer IPL (sublamina a) (Dacey et al., 2000; Famiglietti et al., 1977; Kolb, 1995; Nelson et al., 1978). Further, different types of bipolar cells with different response characteristics also stratify in different IPL layers (B. B. Boycott & Wassle, 1991; Haverkamp et al., 2003). RGCs dendrites co-stratify with bipolar cells within the same IPL stratum, giving each RGC type a unique response characteristic. The second anatomical feature of different RGC types is the size or radius of the dendritic field. While depth of stratification can reveal information about light response polarity dendritic field size can reveal information about receptive field size (Grünert & Martin, 2020).
2.1. LGN – the Primary Central Projection for Conscious Vision
The vast majority of RGCs project to the lateral geniculate nucleus (LGN), the gatekeeper of conscious vision (Dacey, 2004; Perry et al., 1984). Located on the posterior lateral margin of the thalamus, most central-vision RGCs make a dominant connection with a single thalamic relay neurons projecting to V1 – although many RGCs also make weak connections to retinotopically surrounding LGN neurons, proposed to increased spatial discrimination of each receptive field (Usrey et al., 1999). In the periphery, these connections exhibit a higher degree of convergence, with multiple RGCs synapsing onto single LGN neurons (Callaway, 2005). However, just as the retina is much more than a simple transducer, the LGN is much more than a simple relay system from RGCs to striate pyramidal neurons (Kastner et al., 2006; Pregowska et al., 2019). Inhibitory interneurons make up almost 9% of LGN neurons (Braak & Bachmann, 1985) and help refine LGN RFs (Halnes et al., 2011). In conjunction with local lateral inhibition, the LGN receives feedback signals from V1, the thalamic reticular nucleus, and various brainstem structures such as the pulvinar and superior colliculus (Ghodrati et al., 2017). It is estimated that as many as 95% of LGN input fibers are extraretinal and that, perhaps surprisingly, this striking majority has a primarily modulatory role on the flow of information from the retina to the cortex (e.g., Briggs & Usrey, 2011; Gale & Murphy, 2018; Guillery & Sherman, 2002; Zeater et al., 2019).
Retinal input is mapped onto six distinct layers (although see Hickey & Guillery, 1979), each containing a specific population of cells. The bottom two layers of the LGN are composed of magnocellular neurons, while the top four layers contain parvocellular neurons. The four parvocellular layers are sometimes described as two layer with four leaflets because although caudal cross-sectional views of the LGN show four distinct segments, rostrally these segments fuse pairwise into two layers (Kaas et al., 1978; Malpeli et al., 1996). Magnocellular cells (M cells) have large, achromatic RFs which are highly sensitive to luminous contrast, or differences in brightness between two adjacent areas in a visual scene. Parvocellular cells (P cells) have smaller, spectrally opponent RFs, defined as being excited by one range of wavelengths and inhibited by another. In addition, M cells tend to respond faster and preferentially to higher temporal frequencies while P cells respond more slowly, and to lower temporal frequencies. In addition to M and P cells, koniocellular cells (K cells) can be found between the M and P layers, and in some cases, intercalated in bridges within M and P layers. These cells tend to have large RFs and demonstrate some spectral opponency (dominantly blue-on), however display a wide diversity of RF properties – a point to which we will return (Kaplan, 2003; Martin et al., 1997). For review of the current state of LGN research, we direct readers to a trio of recent publications (Casanova & Chalupa, 2023; Ghodrati et al., 2017; Stacy & Van Hooser, 2022).
2.2. LGN – the Diverse Properties of K Cells
Koniocellular layers receive input from many different RGC populations (Dacey, 2004). Consequently, K cells have been shown to express highly heterogenous RF properties (Solomon, 2021). These include blue-on, blue-off, suppressed-by-contrast, M-like, orientation-selective, and direction-selective RFs. Even blue-on RFs are both spatially and temporally different from blue-off RFs (Pietersen et al., 2014; Tailby et al., 2008). This functional asymmetry contrasts with M and P pathways where on and off RFs are organized with similar spatiotemporal responses and differ only in polarity. Furthermore, some K cells may not have any external RF at all, i.e., are not visually activated. A study by Klein et al. (2016) found that less than 50% of macaque K cells had any visually activated RF. This result has been supported by other groups using marmosets and owl monkeys (A. J. White et al., 2001; X. Xu et al., 2001).
Genetic analysis also supports a hetererogeneity in the K population of LGN. Single-nucleus sequencing suggests three to five separate subtypes in macaque and human (Bakken et al., 2021; S. H. Sun et al., 2024). Work in tree shrew suggests a similar subdivision into at least two molecularly distinct channels (Sciaccotta et al., 2025).
Beyond diverse RF and genetic properties, K cells also show exceptional anatomical diversity. K cells as a class project to different layers of V1 (Casagrande et al., 2007; Chatterjee & Callaway, 2003; Hendry & Yoshioka, 1994), demonstrate different degrees of axonal arborization in V1 (Casagrande et al., 2007), express different biomarkers (Bakken et al., 2021; Baldicano et al., 2022; Solomon, 2021), and some form direct extrastriatal projections while others do not (Atapour et al., 2022; Yu et al., 2018). Because of the variety in these anatomical features, and the strong linkages between RGC morphology and function, the argument can be made that K cells are best considered as a collection of distinct populations of functionally differentiated cell types.
Based on current understanding of anatomical and physiological differences, studies have broadly identified two different categories of K cells, here referred to as vK (ventral koniocellular) and dK (dorsal koniocellular) neurons (Bakken et al., 2021; Casagrande et al., 2007; Eiber, Rahman, et al., 2018). vK cells express the biomarker PENK (proenkephalin), in addition to K specific markers such as CAMK2A and PRKCG, and are found enriched in the two K layers directly adjacent to the M layers – K1 and K2 (Bakken et al., 2021; Murray et al., 2008). The most common receptive field type in the vK layers are large and show high contrast sensitivity, and sensitivity to high temporal frequencies (Solomon et al., 2002; A. J. White et al., 2001; X. Xu et al., 2001). In marmosets, M-like, orientation selective, and direction selective RFs have been recorded from vK layers (Cheong et al., 2013; Eiber, Rahman, et al., 2018; A. J. White et al., 2001). Marmoset vK cells also demonstrate extrastriatal projections, primarily to area MT (Bakken et al., 2021; Eiber, Rahman, et al., 2018; Murray et al., 2008). These properties have led some researchers to suggest that the vK layers form a part of a phylogenetically old, primitive motion pathway which has been largely replaced by the M layers throughout primate evolution (Percival et al., 2014), but remains as a subcortical path to sharpen saccadic response times to highly salient stimuli.
dK (dorsal K) cells are found in the remaining K layers adjacent to P layers (K3–6) and express only the K-specific CAMK2A and PRKCG biomarkers (Bakken et al., 2021; Murray et al., 2008). In marmosets, S-cone carrying (blue-on and blue-off) RFs as well as suppressed-by-contrast, orientation selective, and direction selective RFs have all been identified in these layers (Cheong et al., 2013; Eiber, Pietersen, et al., 2018; Tailby et al., 2007; A. J. White et al., 2001).
3. Magnocellular Projecting RGCs
As mentioned above, the LGN consists of six layers that can be functionally partitioned into three cell types. M cells make up the two ventral layers of the six. In common with the P layers, M layers receive dominant input from one high-density type of RGC: the parasol RGC (Dacey, 2004; Ma et al., 2023). However, new research has suggested that another low-density RGC type, the smooth monostratified RGC, also projects to the M layers, showing similar response characteristics to parasol cells (Crook et al., 2008).
3.1. Parasol RGCs
Parasol RGCs are the second most prevalent RGC type in the primate retina and are implicated in low spatial resolution vision. Parasol RGCs project primarily to the M layers of LGN. Making up ~6% of all RGC in the fovea and ~10% of all RGCs in the periphery, parasol cells show larger dendritic spread than midget cells but significantly smaller tiling than some of the other low density wide-field RGC types (Dacey, 2004; Ma et al., 2023). As opposed to midget cells, which make private line cone connections in the fovea, parasol cells contact multiple L- and M-cones indiscriminately. This feature eliminates spectral opponency in parasol cells, but makes parasol cells exceptionally sensitive to even small changes in luminosity (Crook et al., 2014; Dacey & Petersen, 1992; Kaplan & Shapley, 1982; Soto et al., 2020). There is some division across the scientific community with respect to whether parasol cells also indiscriminately contact S-cones (Calkins & Sterling, 2007). Some studies have found evidence of S-cone input to the M pathway (e.g., Chatterjee & Callaway, 2002) while other studies have found no evidence of S-cone input and attributed Chaterjee and Callaway's result to inadvertent stimulation of M and L cones. (e.g., Sun et al., 2006). In addition to high luminance contrast sensitivity, parasol cells are also highly sensitive to high temporal frequencies (Crook et al., 2008, 2014; B. B. Lee, 1996b). The axons of parasol cells are larger than most other RGC axons (Walsh et al., 2000), facilitating higher conduction velocity due to lower internal resistance and increased membrane surface area (Hu & Jonas, 2014). This feature also facilitates a highly transient response to light – corresponding to parasol cells’ sensitivity to high temporal frequencies (Crook et al., 2008).
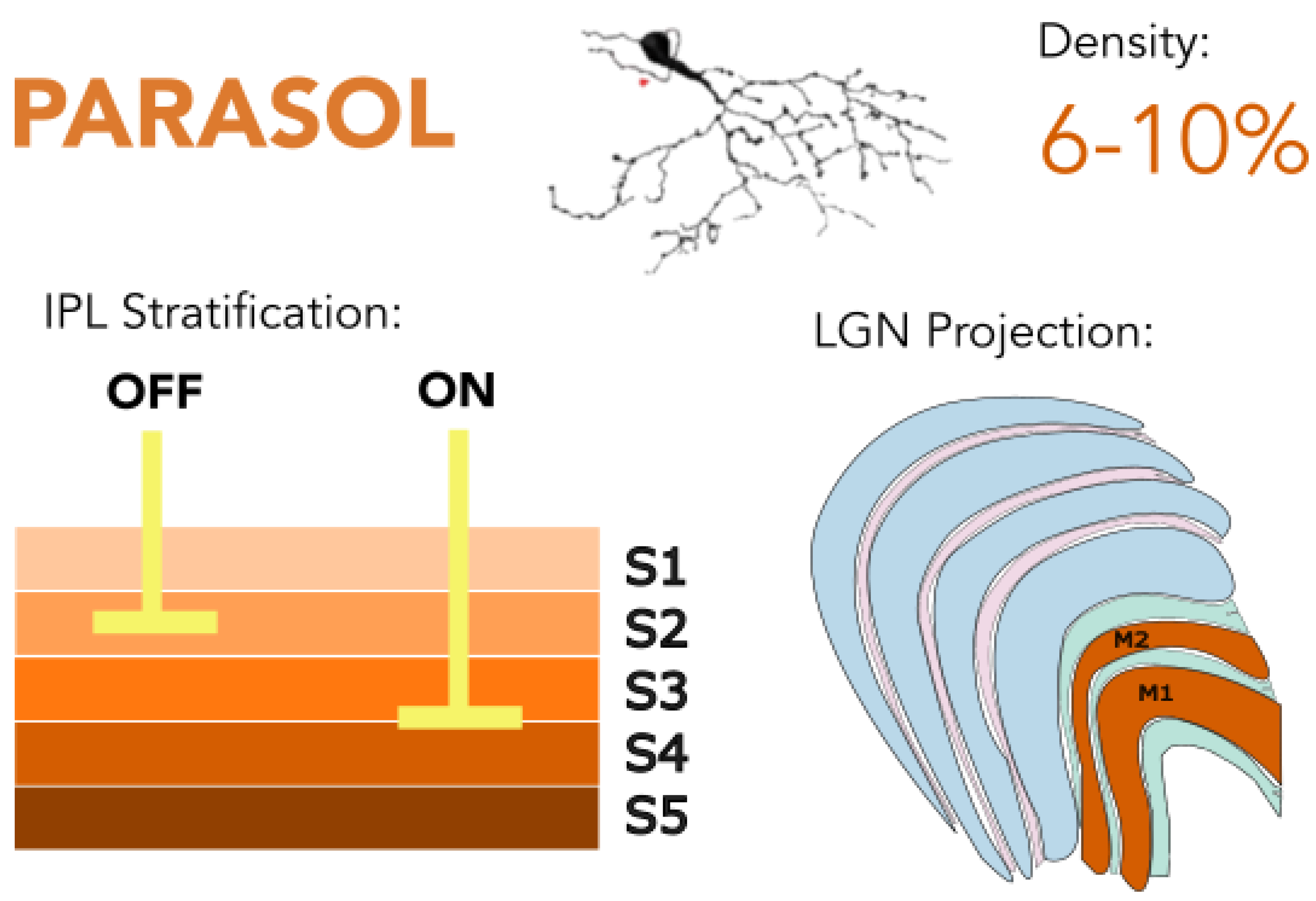
Figure 2.
Parasol retinal ganglion cells (RGCs) stratify in IPL layers S2 and S3-4 and project predominantly to the magnocellular layers of the lateral geniculate nucleus (LGN). They are specialized for detecting high temporal frequencies and luminance contrast, supporting fast, low-resolution visual processing and contributing to motion perception. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.
Figure 2.
Parasol retinal ganglion cells (RGCs) stratify in IPL layers S2 and S3-4 and project predominantly to the magnocellular layers of the lateral geniculate nucleus (LGN). They are specialized for detecting high temporal frequencies and luminance contrast, supporting fast, low-resolution visual processing and contributing to motion perception. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.
Some suggest that parasol cells, due to their response characteristics and their association with the M pathway, are motion sensitive and contribute to the initial stage of motion perception (Appleby & Manookin, 2020; Chichilnisky & Kalmar, 2003; Manookin et al., 2018). This idea has been challenged by lesion studies, finding that lesioning of the magnocellular pathway did not affect motion perception (Merigan et al., 1991; Schiller & Logothetis, 1990). However, those challenges must be considered along with the obvious limitation that lesioning the magnocellular pathway makes visual detection of any stimulus, moving or not, more difficult.
To better understand motion sensitivity in parasol cells, some researchers have focused on the properties of bipolar input to parasol cells. Manookin et al. (2018) found that parasol cells, but not midget cells, showed sensitivity to motion. The authors of that study suggested that the motion sensitivity arose at the level of the bipolar cells, rather than the RGCs. Off parasol cells receive input from diffuse bipolar cell type DB3 (Jacoby et al., 2000) which, in addition to signaling parasol cells, exhibit gap junctions to neighboring DB3 cells (Jacoby & Marshak, 2000). Therefore, according to Manookin et al. (2018), when photons stimulate a cone and that signal is passed to a DB3 cell, part of this electrical current spreads to neighboring DB3 cells. The passage of current across DB3 gap junctions is small enough so as not to elicit a conscious visual phosphene but large enough to increase depolarization of neighboring DB3 cells, giving a stronger response to stimulation during that brief period. Thus, depolarization in one DB3 cell biases neighbor bipolar cells to exhibit a greater response to stimulation.
Unlike midget bipolar cells or S-on bipolar cells, diffuse bipolar cells contact many different RGC types (B. B. Boycott & Wassle, 1991). Thus, the motion sensitivity phenomenon should be present in other low-density RGC types which make synapses with DB3 cells. We consider two points concerning this observation. First, small bistratified cells (another RGC type, discussed later) receive their L+M-off response input from DB3 cells but exhibit no motion sensitivity. This could stand as evidence against the DB3 theory, suggesting that parasol cells’ motion sensitivity is instead associated with their amacrine cell connections. However, concurrent S-on bipolar cell input may confound the motion sensitivity effect in small bipolar RGCs. Second, only off parasol cells make contacts with DB3 (Haverkamp, 2010). On parasol cells make contacts with diffuse bipolar type 5 cells (DB5) and it is currently unclear based on available data if DB5 cells express homologous gap junctions (Jacoby & Marshak, 2000). Thus, it is unclear whether recorded parasol motion sensitivity is only present in the off system. More research targeted at motion sensitivity within DB3 cells would help to clarify the significance of these mechanisms.
In addition to retinal anatomy and physiology, the connections to central structures made by parasol cell also imply a role in motion detection. Parasol cells display branched axons with projections to the M layers of LGN and to the superior colliculus (Crook et al., 2008). The superior colliculus is an integrative thalamic center, using visual, auditory, and somatosensory information to orient the eyes and the head in the environment (Meredith & Stein, 1986). Selectivity for local motion has been found in the superior colliculus, with parasol cells found to contribute (Appleby et al., 2021). Although not specific to parasol cells, a recent anatomical study in macaque found that all RGCs projecting to the superior colliculus had collateral projections to the LGN (Zheng et al., 2024), implying a commonality of input to the two areas, and suggesting that any parasol-mediated information available in the midbrain is provided to the cortical stream as well.
In short, parasol cells have been the attention of much research, but further questions concerning their functionality remain. Most of these questions concern the specific contributions of parasol cells to conscious vision. For example, and as we will discuss later, how does the motion sensitivity of parasol cells differ or support other low-density motion selective pathways? What are the precise roles of parasol and midget cells each in achromatic spatial vision? These questions can help to guide future research into the functional role of parasol cells.
3.2. Smooth Monostratified RGCs
Smooth monostratified RGCs very closely resemble parasol cells but have smooth dendrites. Additionally, they show smaller cell bodies, lower density, and larger tiling radius (Crook et al., 2008). However, like parasol cells, they form inner-on and outer-off populations, and they narrowly stratify in similar IPL strata, implying similar diffuse bipolar input (DB2 or 3 for outer, DB4 or 5 for inner; Grünert & Martin, 2021). Together the on and off populations make up between 2 and 3% of all RGCs at 8 mm of eccentricity (Crook et al., 2008).
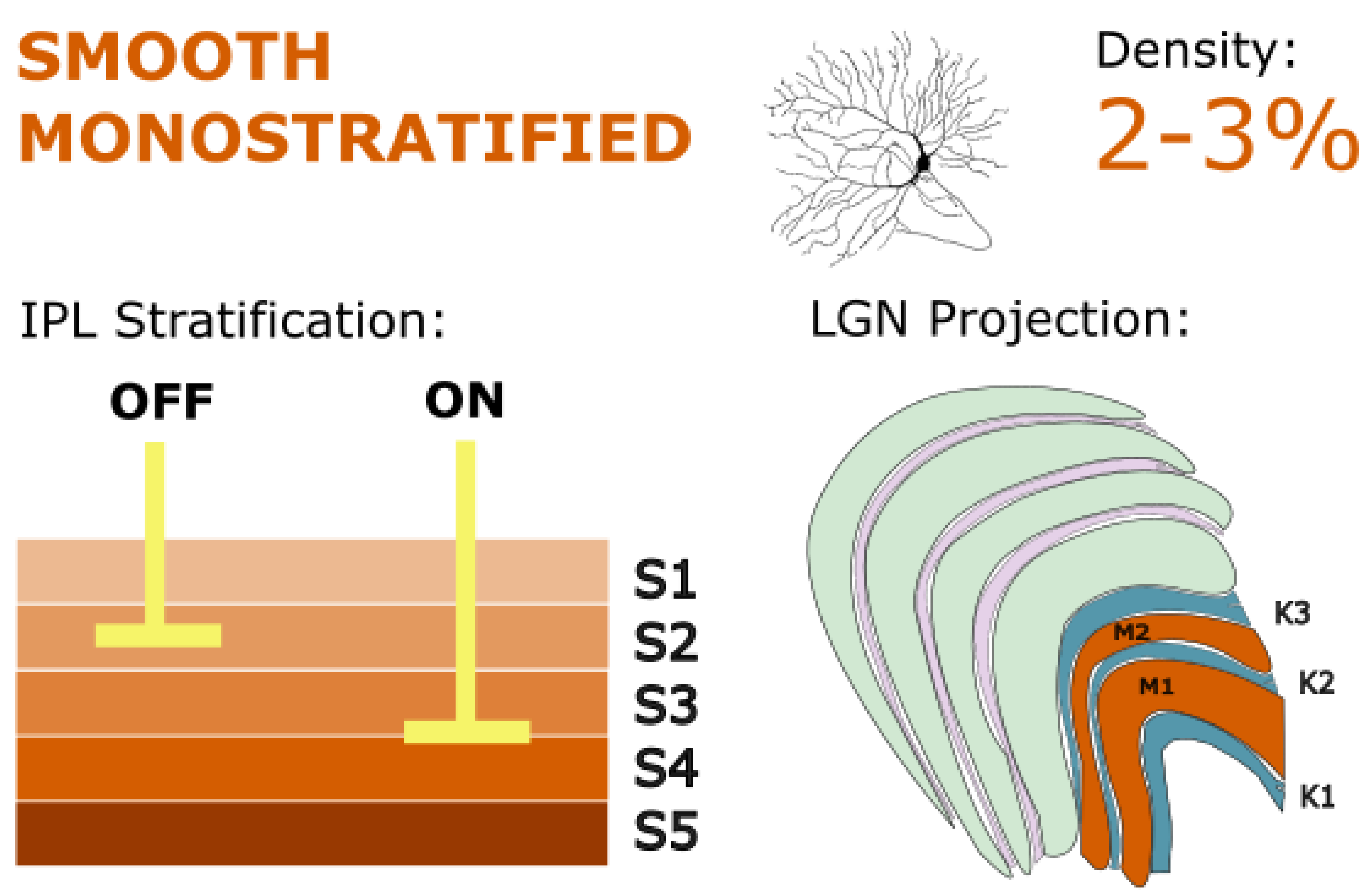
Figure 3.
Smooth monostratified RGCs stratify in IPL layers S2 and S3-4, resembling parasol cells in their magnocellular LGN projections, but also targeting ventral koniocellular layers. They exhibit nonlinear spatial summation and may play a key role in global motion detection and coarse spatial processing. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.
Figure 3.
Smooth monostratified RGCs stratify in IPL layers S2 and S3-4, resembling parasol cells in their magnocellular LGN projections, but also targeting ventral koniocellular layers. They exhibit nonlinear spatial summation and may play a key role in global motion detection and coarse spatial processing. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.
Smooth monostratified RGCs are functionally similar to parasol cells (Crook et al., 2008). They sum input from L and M cones with no measurable contribution from S-cones. They show fast spike discharge rates with high contrast and temporal sensitivity. In fact, the striking similarities between smooth monostratified RGCs and parasol cells have even led some to suggest that these cells must also terminate in the LGN M layers (Kaplan, 2003). Synaptic-resolution mapping revealed the circuit origins of the smooth monostratified distinctive receptive field structure (Patterson et al., 2024), supporting support their role in a parallel M-like stream distinct from parasol cells.
Other research has identified important features of smooth monostratified RGC RFs with distinct functional significance separate from parasol cells. Smooth monostratified RGCs exhibit irregularly shaped RFs with distinct hotspots. Activation in one of these hotspot areas increases RGC response, whereas activation between hotspots decreases response. It is proposed that these hotspot areas arise from a dendritic spiking mechanism (Rhoades et al., 2019). This nonlinear spatial summation is fundamental to RGC computation and allows RGCs to produce responses to spatial patterns that are smaller than the RF, while still sampling a large portion of the retina (Rhoades et al., 2019). Functionally, this property of nonlinear summation connects smooth monostratified RGCs to another type of mammalian RGC, the Y cell, which was first identified in cats and also show nonlinear achromatic spatial summation (Enroth-Cugell & Robson, 1966; Rosenberg & Talebi, 2009). In Y cells, as well as in smooth monostratified RGCs, this nonlinear summation suggests a role for these cell types in motion-detection-related spatial computation.
Smooth monostratified RGCs have been shown to project to LGN (Crook et al., 2008; Dacey, 2004; Percival et al., 2013). Some have suggested the M layers as the primary thalamic target (Kaplan, 2003). Others have proposed that smooth monostratified RGCs project to the koniocellular layers (Crook et al., 2008). Intuitively, given the response characteristics of vK cells, as well as their extrastriatal projections to MT, one might expect smooth monostratified RGCs to target the vK layers. If smooth monostratified RGCs were to project to vK layers, it is possible that they could be the source of the M-like RFs identified in vK layers (Eiber, Rahman, et al., 2018; Solomon, 2021). However, analyses of K-layer projecting RGC types have found smooth monostratified RGCs in dK layer K3 in marmosets as well (Percival et al., 2013). Further research on the specific targets of smooth monostratified RGCs’ LGN projections can help to elucidate this anatomical and thus functional question.
In addition to LGN projections, smooth monostratified RGCs – like parasol cells – show projections to the superior colliculus (Crook et al., 2008; Grünert et al., 2021). Based on the similarity of response characteristics between parasol and smooth monostratified RGCs, it is predicted that these two cell types have similar functions in the superior colliculus. However, there are also distinct differences between the functional contributions of parasol cells and smooth monostratified RGCs in the superior colliculus. For example, smooth monostratified RGCs contribute to global motion detection (e.g., movement of the entire visual field) whereas parasol cells contribute to both global and local motion detection (Appleby et al., 2021). Importantly to conscious vision, the smooth-monostratified-specific sensitivity to global motion in the superior colliculus may also be represented in LGN, forming the basis for the conscious differentiation between local and global motion. This may be one way that functionally similar but different spatial resolution RFs may strengthen the ability of the visual system to encode motion, as predicted by Crook et al. (Crook et al., 2008).
4. Parvocellular Projecting RGCs
Parvocellular cells are the largest population of cells in LGN, making up the four dorsal layers of LGN. Despite outnumbering all other cell types in LGN and being the major contributor of central vision, like the M layers, P layers receive almost exclusive input from one RGC type: the midget cell.
4.1. Midget RGCs
Midget RGCs are the most prevalent RGC type in the primate retina and are implicated in both the high spatial resolution and chromaticity of central vision. Making up 90% of all RGC in the fovea, these RGCs show a dendritic spread as few as 5 microns (Dacey, 2004). In the central few degrees of retina, midget cells receive input from just a single cone via a dedicated midget bipolar cell, forming a basis for high-resolution central vision. As retinal eccentricity increases, the size of the midget cell dendrites also increases, increasing 10-fold only 2-6 mm from the fovea (Dacey, 1993b).
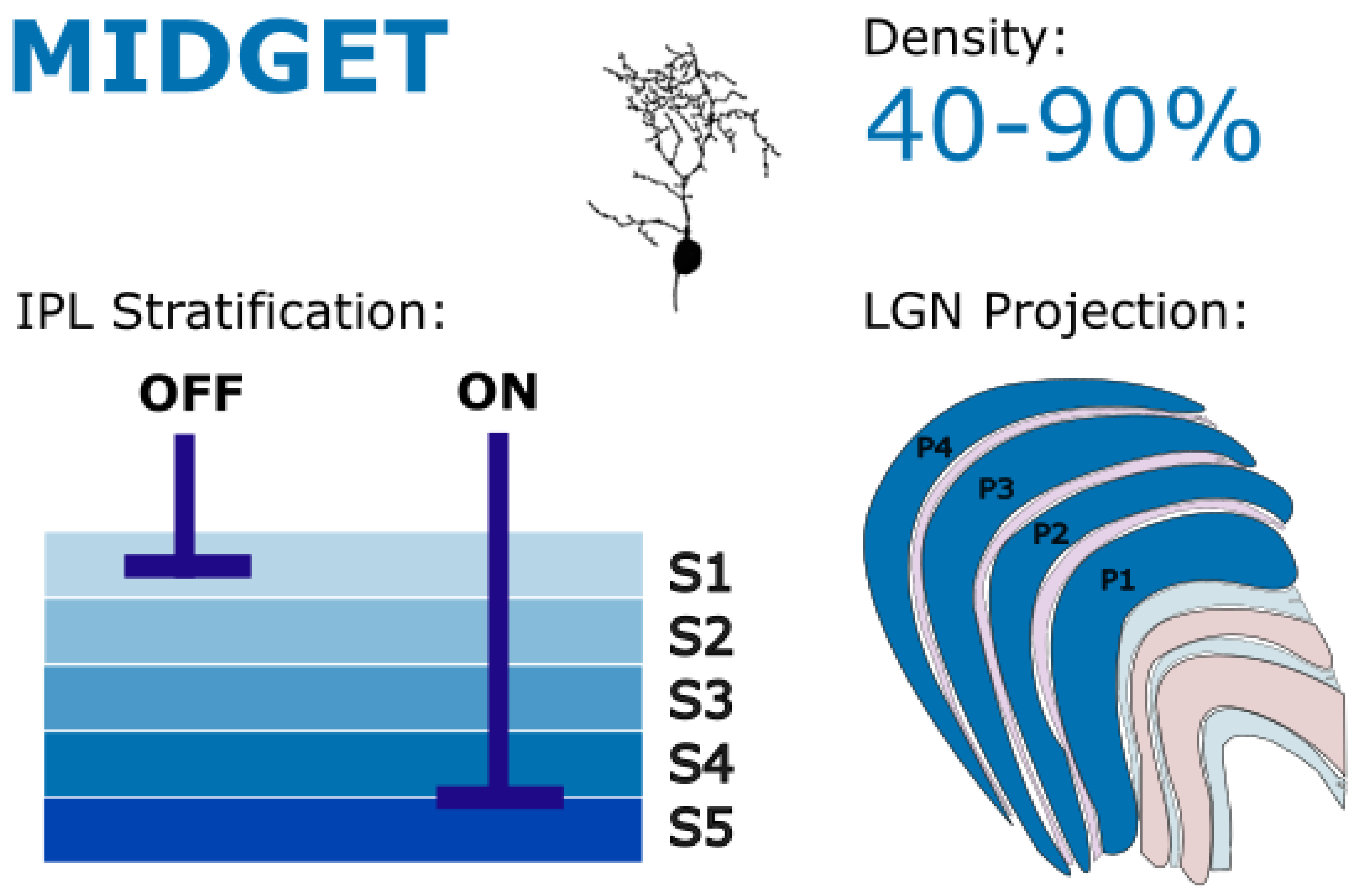
Figure 4.
Midget RGCs stratify in IPL layers S1 (off) and S4-5 (on), with projections to the parvocellular layers of the LGN. They are critical for high-acuity central vision and mediate red-green color opponency, supporting both fine spatial detail and chromatic discrimination. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.
Figure 4.
Midget RGCs stratify in IPL layers S1 (off) and S4-5 (on), with projections to the parvocellular layers of the LGN. They are critical for high-acuity central vision and mediate red-green color opponency, supporting both fine spatial detail and chromatic discrimination. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.
Midget RGCs are classified into two distinct subpopulations. Both subpopulations are narrowly unistratified, however one population stratifies in the on sublamina of the IPL at stratum 1, while the other stratifies in the off sublamina of the IPL at stratum 4/5 (Dacey, 1993). These stratifications near the IPL borders indicate high specificity for on- or on-center responses in each respective population, with little overlap (Watanabe & Rodieck, 1989). These opposite-polarity responses create push-pull parallel information streams that help to increase visual acuity. In the central retina, one cone photoreceptor connects to two midget bipolar cells (one on and one off) which then feed into their respective on or off midget RGC, creating spatial coincidence between on and off pathways. On midget RGCs receive input from both long-wavelength (L; reddish) and medium-wavelength (M; greenish) cones as well as input from rod photoreceptors in the periphery. Off midget RGCs receive input from the same photoreceptors as their on counterpart but additionally receive input from short-wavelength cones (S; blueish; Schiller & Tehovnik, 2015).
Molecularly, midget cells (along with parasol cells) were matched as the ortholog of sustained alpha and transient alpha cells, respectively, in non-primate mammals (Hahn et al., 2023). This indicates a likely similarity in functionality between these primate and non-primate cell types which is supported by electrophysiological and anatomic studies as well (Dacey & Brace, 1992; Dacey & Petersen, 1992).
Despite much research, inferring the functional contribution of midget RGCs from their anatomy and physiology may be complicated. Midget cells appear uniquely suited for two different, and seemingly unrelated, conscious visual phenomena: spectral and spatial opponency. Like the RFs of all RGCs, midget cell RFs have an opponent center-surround organization, comparing the input from one central cone with the group of cones in close proximity to the central cone (Kuffler, 1953). At the level of the fovea, private-line communication between cones and midget RGCs, and opponent center-surround RF, appear to serve the high spatial resolution present in central vision. In addition, the ability of midget RGCs to compare a single, spectrally sensitive center cone with other spectrally sensitive cones in the RF surround makes midget cells uniquely suited for color vision (Martin et al., 2011; Schiller et al., 1990). Physiologically, midget cells have been found to indiscriminately respond to both chromatic and achromatic edges (Wiesel & Hubel, 1966), leading some to believe that midget cells encode both color and spatial resolution (Lennie & D’Zmura, 1988). To support this claim, it has been repeatedly shown that parvocellular specific lesioning and pathology impair both chromatic and fine, achromatic contrast sensitivity (Al-Hashmi et al., 2011; Merigan et al., 1991; Schiller et al., 1990). However, from an information processing standpoint, it has been challenging to reconcile these two functionalities. The neural unit of communication, action potential, is discrete. Thus, information regarding chromatic and achromatic contrast from midget RGCs should be indistinguishable by parvocellular neurons (Patterson et al., 2019). However, from experience we know that achromatic contrast and equiluminant chromatic contrast are differentiable; you can tell the difference between a red/green edge and a black/white edge.
This seeming paradox of encoding has been the subject of tremendous discussion across the scientific community and will not be the focus of this review. For a more in-depth review of midget cell color and spatial encoding, we direct the reader to Patterson et al.’s 2019 review (Patterson et al., 2019).
5. Koniocellular Projecting RGCs
The K cells are found intercalated between the M and P layers. Despite being the numerically smallest population of cells in most primates, the K layers receive the widest ranges in diversity of RGC input to LGN, and thus their description will occupy much of this report. Many of the RGC types which project to the K layers are low density cells, tiling broadly and sparsely across the retina. This makes them more difficult to identify and study compared to midget and parasol cells which are significantly higher density. However, as techniques of identifying RGCs from K layers have improved, our understanding of the vast diversity of K-projecting RGC types has likewise increased.
5.1. Small Bistratified RGCs
The small bistratified ganglion cell was the first non-M, non-P retinal ganglion cell to be identified in the primate retina when retrograde injections from the LGN revealed a population of retinal cells with bistratified dendrites in the IPL (Rodieck, 1991). Making up between 1% and 10% of retinal ganglion cells depending on eccentricity (Dacey, 1993a), the small bistratified RGC has a medium tiling radius and bistratifies narrowly near the IPL borders in both stratum 1 and stratum 4/5 – comparable to on and off midget cells (Dacey, 1993a, 2004). Dendrite bistratification in both on and off sublayers makes this cell an on/off type, responding to both the onset and offset of light. In the on sublamina, small bistratified ganglion cells contact S-on bipolar cells. In the off sublamina, small bistratified cells contact diffuse bipolar cell type 3 (DB3) carrying mixed L-cone and M-cone signals (Marshak & Mills, 2014). Thus, the small bistratified cell has a spectral preference, responding specifically to both the onset of short-wavelength light and the offset of long-wavelength light (Neitz & Neitz, 2017). This cell type was initially shown to project to the extreme borders of parvocellular layers 3 and 4, where it was presumed to have a role in color vision (Dacey, 1993a; Michael, 1988; Rodieck, 1991). As axon tracing, intracellular injection, and LGN anatomy has become more refined, the true target of small bistratified ganglion cells in both macaques and marmosets has been shown to be the dK layers, specifically K3 and K4 (Hendry & Reid, 2000; Roy et al., 2009; Szmajda et al., 2008).
Figure 5.
Small bistratified RGCs bistratify narrowly in IPL layers S1 and S4-5 and project to dorsal koniocellular layers of the LGN. These cells encode blue-on/yellow-off chromatic opponency, forming the basis of the blue-yellow axis in color vision. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.
Figure 5.
Small bistratified RGCs bistratify narrowly in IPL layers S1 and S4-5 and project to dorsal koniocellular layers of the LGN. These cells encode blue-on/yellow-off chromatic opponency, forming the basis of the blue-yellow axis in color vision. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.

The identification and physiological classification of small bistratified RGCs has paved the way for a new hypothesis concerning color vision, suggesting that red and green chromatic opponency is carried through midget pathways while blue and yellow chromatic opponency is carried through a separate pathway entirely, starting at the small bistratified RGC.
This small bistratified hypothesis of color vision suggests that small bistratified RGCs play an important role in the blue/yellow axis of conscious color vision (Dacey, 2004; Szmajda et al., 2008; Thoreson & Dacey, 2019) while midget cell spectral opponency creates the red/green axis. The distinct RF characteristics of small bistratified RGCs, as well as their projections to LGN, makes them well suited to form the basis of blue/yellow color vision. In addition, their density matches the density of S cones in most retinal locations (Calkins et al., 1998) supporting a mechanism for the matching between psychophysical blue-yellow grating discrimination and S-cone density (Mullen & Kingdom, 2002). Proponents for the small bistratified theory of color coding argue that red/green and blue/yellow color circuits remain segregated in the LGN through P and K channels, respectively. These color signals anatomically and functionally terminate in distinct V1 layers – 4C for red/green and 2/3B/4A for blue/yellow – and are combined in the cortex (Chatterjee & Callaway, 2003; Jayakumar et al., 2013).
5.2. Large Bistratified RGCs
Large bistratified RGCs make up just over 2.5% of all RGCs at approximately 8 mm from the fovea (Wässle, Grünert, et al., 1989). Although this cell type has not yet been classified physiologically, many of its functional properties can be postulated from anatomical and connectomic information. Anatomically, large bipolar retinal ganglion cells resemble small bistratified RGCs, but with broader tiling and sparser dendritic spread (Dacey, 2004). Large bistratified RGCs co-bistratify narrowly with small bistratified RGCs at the borders of the IPL in the on and off sublaminae – stratum 1 and stratum 4/5 – implying response characteristics that are similar to small bistratified RGCs (S-on/L+M-off) corresponding with their similar stratification depth. Broad tiling across the retina contributes to their lower density as well as giving these cells a larger RF than the small bistratified RGCs, however as large bistratified cells approach the fovea, the tiling size significantly shrinks. This may imply a greater central density, as exhibited by midget cells, however density data from the fovea or central retina are not yet available.
Figure 6.
Large bistratified RGCs stratify in IPL layers S1 and S4-5 and project to dorsal koniocellular layers of the LGN and the inferior pulvinar. They are thought to contribute to blue-yellow color processing and may modulate visuomotor functions through subcortical projections. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.
Figure 6.
Large bistratified RGCs stratify in IPL layers S1 and S4-5 and project to dorsal koniocellular layers of the LGN and the inferior pulvinar. They are thought to contribute to blue-yellow color processing and may modulate visuomotor functions through subcortical projections. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.
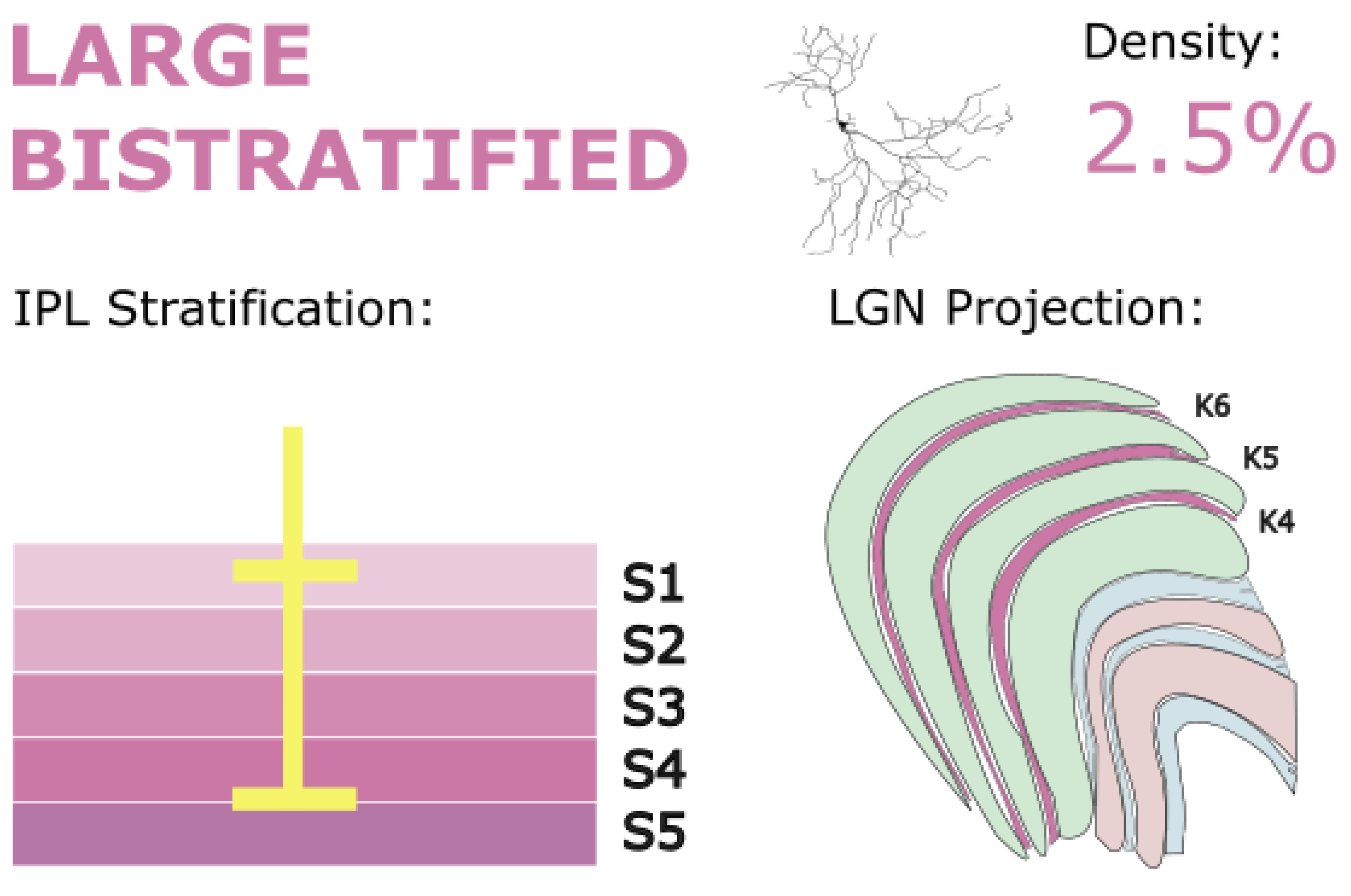
Studies in both marmosets and macaques have found that large bistratified RGCs project to the dK layers of LGN, specifically K3, where they might play a similar role to small bistratified RGCs in chromatic processing (Dacey et al., 2003; Dacey & Packer, 2003; Percival et al., 2013). However, another study found that large bistratified cells did not make up a significant portion of K3 input (Szmajda et al., 2008), but those authors attributed some of this result to methodological constraints. In this same study, however, other K3-projecting populations such as small bistratified and large sparse RGCs could be identified, even despite these experimental limitations, indicating that large bistratified RGCs likely only make up a small portion of K3 efferents. This observation agrees with other large bistratified RGC tracer studies such as Percival et al. (2013) who found only a few large bistratified axons from multiple K3 tracer injections.
If it is true that large bistratified cells increase in density as they approach the fovea, these cells may have the appropriate criteria to form the basis for an S-on response to support blue/yellow central color vision. However, the fact that that these cells are difficult to isolate from K layer injections, whereas small bistratified cells are much readily identifiable due to K3 efferent density, suggests that this speculation may not be correct.
Large bistratified RGCs also project to the pulvinar, which may reveal more about their functionality. According to a study by Grünert et al. (2021), large bistratified RGCs made up around 25% of retinal input to the inferior pulvinar. The pulvinar, like the superior colliculus, is largely a modulatory entity (Kaas & Lyon, 2007). The pulvinar receives input from the visual cortex along with input from other cortical areas, the retina, and the superior colliculus, and sends the processed combination back to the cortex where the pulvinar output plays a modulatory activity. Specifically, the inferior pulvinar projections modulate activity in areas associated with the dorsal stream of visual processing, such as MT and MST (Baldwin et al., 2017). Functional reviews of the pulvinar have focused on visual salience, in which pulvinar cells signal the importance of stimuli in different pulvinar receptive fields (Grieve et al., 2000; Robinson & Petersen, 1992). The pulvinar cells form a retinotopic receptive field map, much like the LGN. Pulvinar receptive fields show heterogeneity in response characteristics, coinciding with diverse pulvinar inputs (retina, cortex, superior colliculus). As some pulvinar receptive fields have been shown to be color specific (Benevento & Miller, 1981), there may be a potential role for large bistratified cell signals in the pulvinar.
Overall, the fact that large bistratified RGCs project to the LGN implicates them in conscious vision, however the stronger pulvinar projection suggests a primary role in non-conscious control of movement and attention (Neitz & Neitz, 2017). Experiments which directly and selectively stimulate large bistratified RGCs in alert participants can help to define the conscious contribution of large bistratified RGCs. Perhaps the lower density and broader tiling, yet functional similarity to small bistratified RGCs, indicates that these two populations form a coordinated dichotomy in spatial resolution like the achromatic spatial resolutions of midget versus parasol cells. This arrangement may improve the ability to encode chromatic spatial scale. Further research on these lines of reasoning can benefit the functional understanding of this cell type.
5.3. Narrow Thorny RGCs
Narrow thorny RGCs are a population of low density RGCs which, in total, make up a similar percentage of all RGCs as large bistratified RGCs (~2.5% at 8 mm from the fovea; Wässle et al., 1989). The tiling radius of narrow thorny RGCs, like for most RGC types, increases with eccentricity (Chandra et al., 2017; Masri, Percival, et al., 2019), ranging from 150 microns – somewhat close to the tiling radius of parasol cells (Dacey & Petersen, 1992) – to as large as 500 microns. Like midget and parasol cells, narrow thorny cells comprise two separate populations, an inner-IPL-stratifying population and an outer-IPL-stratifying population (Masri, Percival, et al., 2019). The inner-stratifying narrow thorny cells have dendrites near the center of the IPL in stratum 3 while the dendrites of outer-stratifying cells are found near the border of the IPL in stratum 5 (Yamada et al., 2005). A recent study by Lee et al. (2024) found that these two cell populations could be differentiated by their expression of the Foxp1 (outer) and Foxp2 (inner) genes. Interestingly, the dendrites from both populations fall within the inner-on IPL sublamina. Although physiological recording has not yet been done, both populations would be expected to exhibit an on response. This is likely true for the stratum-5-stratifying narrow thorny RGC population which contact diffuse bipolar type 6 (DB6) cells (Percival et al., 2014), presumed to have an on-center RF based on their morphology (S. C. S. Lee et al., 2004). Since the stratum-3-stratifying narrow thorny RGCs are found more centrally in the IPL, their light polarity response is more difficult to predict. These cells stratify just above the level of diffuse bipolar DB3 cells, which carry an off response, but whether they actually contact DB3 cells has yet to be determined (Masri, Percival, et al., 2019). If it is true that both cell populations show on responses, the functional significance of two, separate but parallel on pathways needs additional study.
Figure 7.
Narrow thorny RGCs stratify in IPL layers S3 and S5 and project primarily to ventral koniocellular layers of the LGN. They are implicated in motion detection circuits and may underlie blindsight phenomena via extrastriate pathways. Golgi stain image of cell from (Percival et al., 2014), with minor modifications, used under CC BY-NC-SA 4.0.
Figure 7.
Narrow thorny RGCs stratify in IPL layers S3 and S5 and project primarily to ventral koniocellular layers of the LGN. They are implicated in motion detection circuits and may underlie blindsight phenomena via extrastriate pathways. Golgi stain image of cell from (Percival et al., 2014), with minor modifications, used under CC BY-NC-SA 4.0.
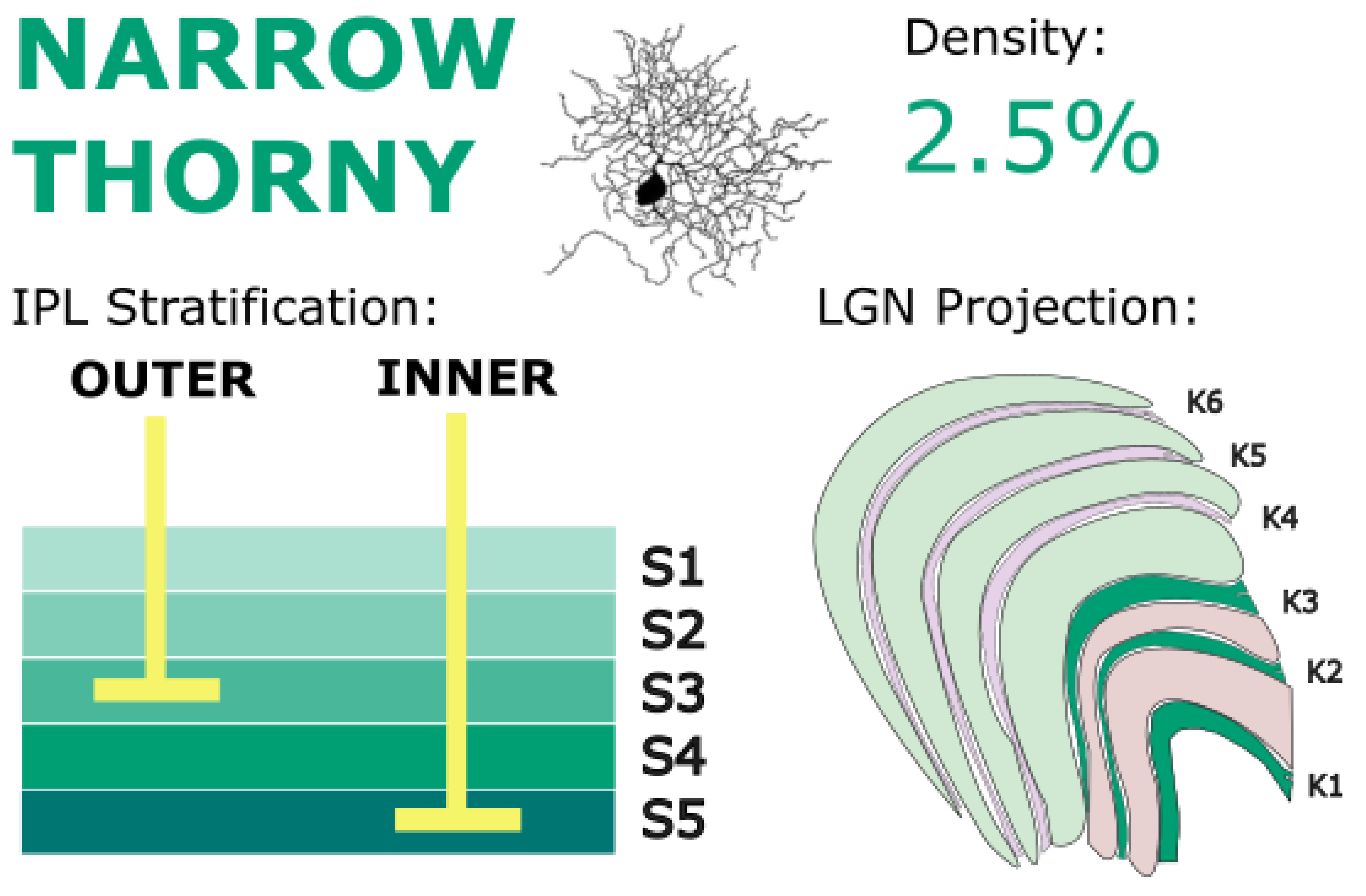
Narrow thorny RGCs have been identified projecting to the LGN (Dacey et al., 2003). Specifically, narrow thorny cells were preferentially labeled when retrograde tracer injections were made in marmoset vK layer K1 (Percival et al., 2014). Some studies have also found very minor projections to the dK layers, specifically layer K3 (Percival et al., 2013), but these are certainly not the dominant LGN projection of narrow thorny RGCs. Thus, given the response characteristics of the vK layers, narrow thorny cells may play a role in motion detection. In fact, when it was discovered that narrow thorny cells preferentially target K1, Percival et al. (2014) speculated that the diffuse bipolar DB6 – narrow thorny RGC – vK pathway formed the basis for the blindsight phenomenon where patients with lesions, injury, or other dysfunction of V1 retain the ability to detect and appraise visual motion despite no conscious perception (Cowey, 2010; Yu et al., 2013). The extrastriatal projections of LGN have been heavily implicated in blindsight (Ajina & Bridge, 2018; Isa & Yoshida, 2021; Schmid et al., 2010), and the discovery of this RGC type which preferentially targets the vK layers gives a retinal substrate for this pathway. This discovery highlights an important point. Although narrow thorny RGCs do target LGN, their role in conscious vision may be minimal, instead showing a greater role in the modulation of conscious vision with respect to motion perception.
Despite having LGN projections, research has shown that the LGN is not the major projection target of narrow thorny RGCs (Chandra et al., 2017). This observation comes from the mismatch in measured cell density when retinal immunohistochemistry is used versus when retrograde tracer injections from the LGN are used. In addition to the thalamic projections, narrow thorny RGCs send axons to the superior colliculus where make up 17% of retinal input (Grünert et al., 2021). The colliculus as the primary target of narrow thorny RGCs, would support the hypothesis that narrow thorny RGCs have an important role in motion detection, as the superior colliculus is important for motor planning and visuomotor control (Chen et al., 2019; Ito & Feldheim, 2018). The relative strength of the collicular versus thalamic projections also supports the speculation that narrow thorny RGCs have only a minor role in conscious vision, if any.
5.4. Broad Thorny RGCs
Broad thorny RGCs are a separate population of low density, thorny RGCs. These cells show morphology similar to the narrow thorny RGC type but differ in their stratification depth (Dacey, 2004). Broad thorny RGCs are about as dense as either the on or off narrow thorny RGCs alone (~1.2% at 8 mm from the fovea; Wässle et al., 1989) and display tiling radius almost identical to narrow thorny and large bistratified RGCs (Chandra et al., 2017; Masri, Percival, et al., 2019): from 69 microns to almost 500 microns. Unlike narrow thorny RGCs, broad thorny RGCs represent only a single population, stratifying broadly near the center of the IPL, continuously from stratum 2 to stratum 4 (Yamada et al., 2005). Consistent with its broad stratification in both the on and off sublaminae, the cell shows a distinct on/off-center light response (Puller et al., 2015) with a strong inhibitory surround. Unlike small bistratified RGCs, which exhibit spectrally sensitive on/off-center RFs, broad thorny RGCs are likely spectrally insensitive (Calkins & Sterling, 2007). Much like other achromatic RGCs (such as narrow thorny and parasol cells), broad thorny RGCs have been linked with diffuse bipolar cell types, including DB2, DB3, and DB3a, all of which stratify in the outer IPL and have an off response (Bordt, Patterson, Girresch, Perez, Tseng, Anderson, Mazzaferri, Kuchenbecker, Gonzales-Rojas, et al., 2021; Calkins & Sterling, 2007; Masri et al., 2016; Puthussery et al., 2014). The coexisting on response has been less well characterized. Bordt et al. (2021) reported that the majority of bipolar input came from on type bipolar cells, but specific on bipolar types were not defined. Some previous studies have hypothesized that on input could derive from diffuse bipolar DB4 or DB5 cells, both of which are achromatic and have been associated with on parasol cells (B. Boycott & Wassle, 1999; Percival et al., 2011). It is also likely that a large portion of the response characteristics are shaped by amacrine cells, which make up 98% of inputs (Bordt, Patterson, Girresch, Perez, Tseng, Anderson, Mazzaferri, Kuchenbecker, Gonzales-Rojas, et al., 2021). Although there is evidence that the proportion of bipolar cell input compared to amacrine cell input increases with proximity to the fovea (Calkins & Sterling, 1996, 2007), the bipolar input to broad thorny RGCs remains lower than for any other characterized primate RGC. This unusual position has important functional implications. Xu et al. (Y. Xu et al., 2008) have proposed that low bipolar input implies low sensitivity to luminosity and visual stimuli. As mentioned above, midget and parasol RGC levels of bipolar input have helped to explain various functional capacities of those cell classes: midget cells receive fewer bipolar inputs and are less sensitive to small changes in luminosity while parasol cells receive more bipolar inputs and are more sensitive to those changes. Broad thorny RGCs’ low proportion of bipolar input suggests that they may require higher luminous contrast to fire compared to other RGC types. At the same time, broad thorny RGCs broad tiling and broad stratification through three IPL strata give them greater dendritic surface area and, thus, more synaptic inputs than other RGC types (Percival et al., 2011). This additional input may help to counteract the reduced luminous sensitivity from a low bipolar to amacrine synapse ratio.
Figure 8.
Broad thorny RGCs stratify diffusely across IPL layers S2 to S4 and project to koniocellular layers of the LGN, with additional inputs to the superior colliculus and pulvinar. These cells are involved in detecting small moving objects and likely support catch-up saccades and motion tracking during smooth pursuit. Golgi stain image of cell from (Moritoh et al., 2013), with minor modifications. Copyright: © 2013 Moritoh et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Figure 8.
Broad thorny RGCs stratify diffusely across IPL layers S2 to S4 and project to koniocellular layers of the LGN, with additional inputs to the superior colliculus and pulvinar. These cells are involved in detecting small moving objects and likely support catch-up saccades and motion tracking during smooth pursuit. Golgi stain image of cell from (Moritoh et al., 2013), with minor modifications. Copyright: © 2013 Moritoh et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.

Detailed circuit reconstructions in macaque retina show that broad thorny RGCs receive a distinctive mix of synaptic inputs from bipolar and amacrine cells (Bordt, Patterson, Girresch, Perez, Tseng, Anderson, Mazzaferri, Kuchenbecker, Gonzales-Rojas, et al., 2021). This wiring supports their putative role in transmitting a specialized K-pathway signal to LGN. Such detailed circuit reconstructions remain rare for K-projecting cell types, and further comparative work across primate species will be essential for linking morphology to functional streams.
Unlike many other low density RGC types, the response characteristics of the broad thorny RGC have been studied in detail, primarily in macaque models. Multiple studies have found that broad thorny cells respond to small moving stimuli, increasing in firing rate transiently when that stimuli enters or leaves their receptive field (Bordt, Patterson, Girresch, Perez, Tseng, Anderson, Mazzaferri, Kuchenbecker, Gonzales-Rojas, et al., 2021; Puller et al., 2013, 2015). However, notably these cells were relatively unresponsive to either a static or moving background by itself and showed no direction selectivity (Bordt, Patterson, Girresch, Perez, Tseng, Anderson, Mazzaferri, Kuchenbecker, Gonzales-Rojas, et al., 2021; Puller et al., 2015). Broad thorny RGCs show striking anatomical and physiological similarities to rabbit local edge detector RGCs and mouse W3 ganglion cells, both of which have been suggested as important in signaling tracking errors during smooth pursuit eye movements (Puller et al., 2013). When the eye is accurately tracking an object with smooth pursuit, the object stays directly in the center of a broad thorny receptive field, and thus, tracked stimuli will appear static to the cell while the background moves behind. Both stimuli – a static object and a dynamic background – elicit little to no response from broad thorny RGCs. However, if tracking falters, the tracked object will begin to drift from broad thorny RGC receptive fields, initiating a strong response and signaling a need for a catch-up saccade to re-center the tracked object.
In addition to this proposed contribution to visuomotor control via catch-up saccades, the projections from broad thorny RGCs suggest a role in conscious vision as well. Broad thorny RGC projections have been recorded in the LGN, specifically in the koniocellular layers of both macaques and marmosets (Dacey et al., 2003; Percival et al., 2011; Rodieck, 1991; Szmajda et al., 2008), likely targeting the vK layers, and K1 in particular (Percival et al., 2011; Szmajda et al., 2008). This placement appears consistent given broad thorny RGC response characteristics (Bordt, Patterson, Girresch, Perez, Tseng, Anderson, Mazzaferri, Kuchenbecker, Gonzales-Rojas, et al., 2021; Puller et al., 2013, 2015) and similar vK response characteristics (A. J. White et al., 2001). Particularly, Eiber et al. (2018) identified on-off, non-direction selective K-cell responses preferentially isolated to the vK layers: It is likely that broad thorny RGCs form the basis for this response (Solomon, 2021).
Outside of LGN, broad thorny RGCs have also been found retrogradely labeled from both the superior colliculus (24% of retinal input) and the inferior pulvinar (Dacey et al., 2003; Grünert et al., 2021; Rodieck & Watanabe, 1993). Like narrow thorny RGCs, this implicates broad thorny RGCs in visuomotor function and motor planning, consistent with their physiologically proposed functionality signaling catch-up saccades in smooth pursuit. In fact, broad thorny input is reported to form the foundational basis for local motion sensitivity in the superior colliculus (Appleby et al., 2021). Given Eiber et al.’s (Eiber, Rahman, et al., 2018) hypothesis that a broad thorny RGC – vK pathway may play an important role in dorsal stream processing, strong input to the inferior pulvinar appears consistent given the inferior pulvinar’s association with the dorsal stream. Consistent with the functional roles mentioned above based on RF characteristics, the anatomical evidence also suggests that broad thorny RGCs could help direct visual attention to moving objects and facilitate tracking during smooth pursuit.
Although current research has emphasized the visuomotor role of broad thorny RGCs, their role in conscious vision remains poorly defined. Given their projections to the LGN, it is likely that they have some effect on conscious vision, however that role may be modulatory, perhaps like large bistratified RGCs or narrow thorny RGCs. Their morphological similarity to narrow thorny RGC may suggest that they play a similar role in subconscious blindsight or modulating motion perception, with a different light polarity response. These hypotheses should be the target of future study.
5.5. Recursive RGCs
Recursive RGCs have also been suggested to play a role in direction-selective motion perception, like the thorny RGCs (Dacey, 2004). These cells exhibit moderate branching and have secondary dendrites which curve back towards the cell body, whence their name. Recursive cell have dendritic fields with a high degree of overlap between individual cells (Detwiler et al., 2019) and form two separate subpopulations - recursive monostratified and recursive bistratified. However, conflicting information exists in the literature regarding the specific morphology and functionality of each of these RGC subtypes. This lack of consensus suggests that recursive RGCs exist in more than just two distinct populations. Alternatively, it may reflect species-specific morphologies, as differences in recursive RGS are known to exist between New World (marmoset) and Old World (macaque) primates (Nasir-Ahmad et al., 2021, 2022) but, most research on recursive RGCs has been done in marmoset models. Therefore, further studies are needed to identify and enumeratethese different populations, their functionalities, and any species-specific characteristics.
Figure 9.
Recursive monostratified RGCs stratify narrowly in the on sublamina of the IPL, but there are conflicting reports as to which strata. They project to koniocellular layers of the LGN. They are hypothesized to play a role in direction-selective motion processing and contribute to visuomotor responses. Golgi stain image of cell from (Moritoh et al., 2013), with minor modifications. Copyright: © 2013 Moritoh et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Figure 9.
Recursive monostratified RGCs stratify narrowly in the on sublamina of the IPL, but there are conflicting reports as to which strata. They project to koniocellular layers of the LGN. They are hypothesized to play a role in direction-selective motion processing and contribute to visuomotor responses. Golgi stain image of cell from (Moritoh et al., 2013), with minor modifications. Copyright: © 2013 Moritoh et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
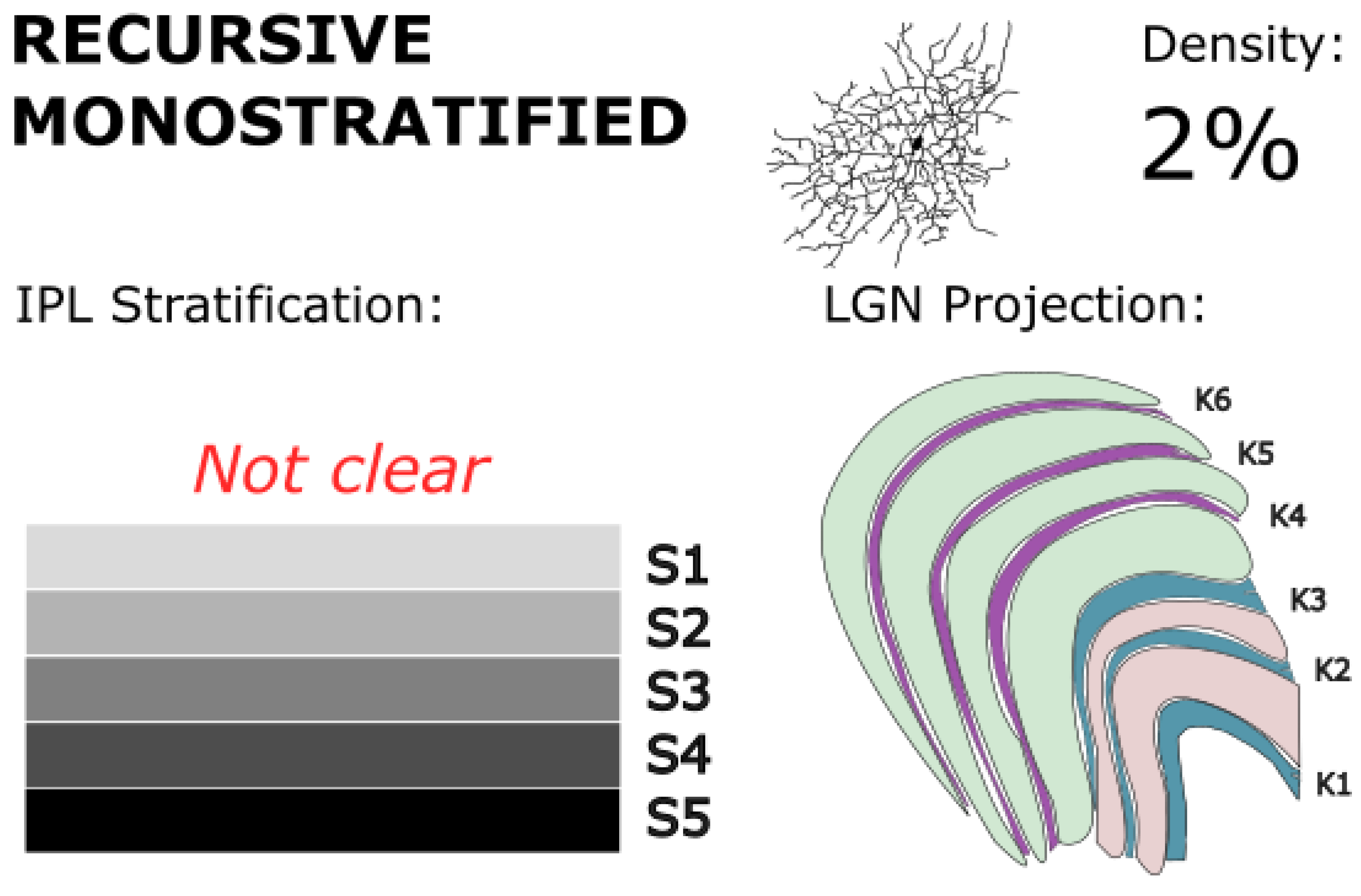
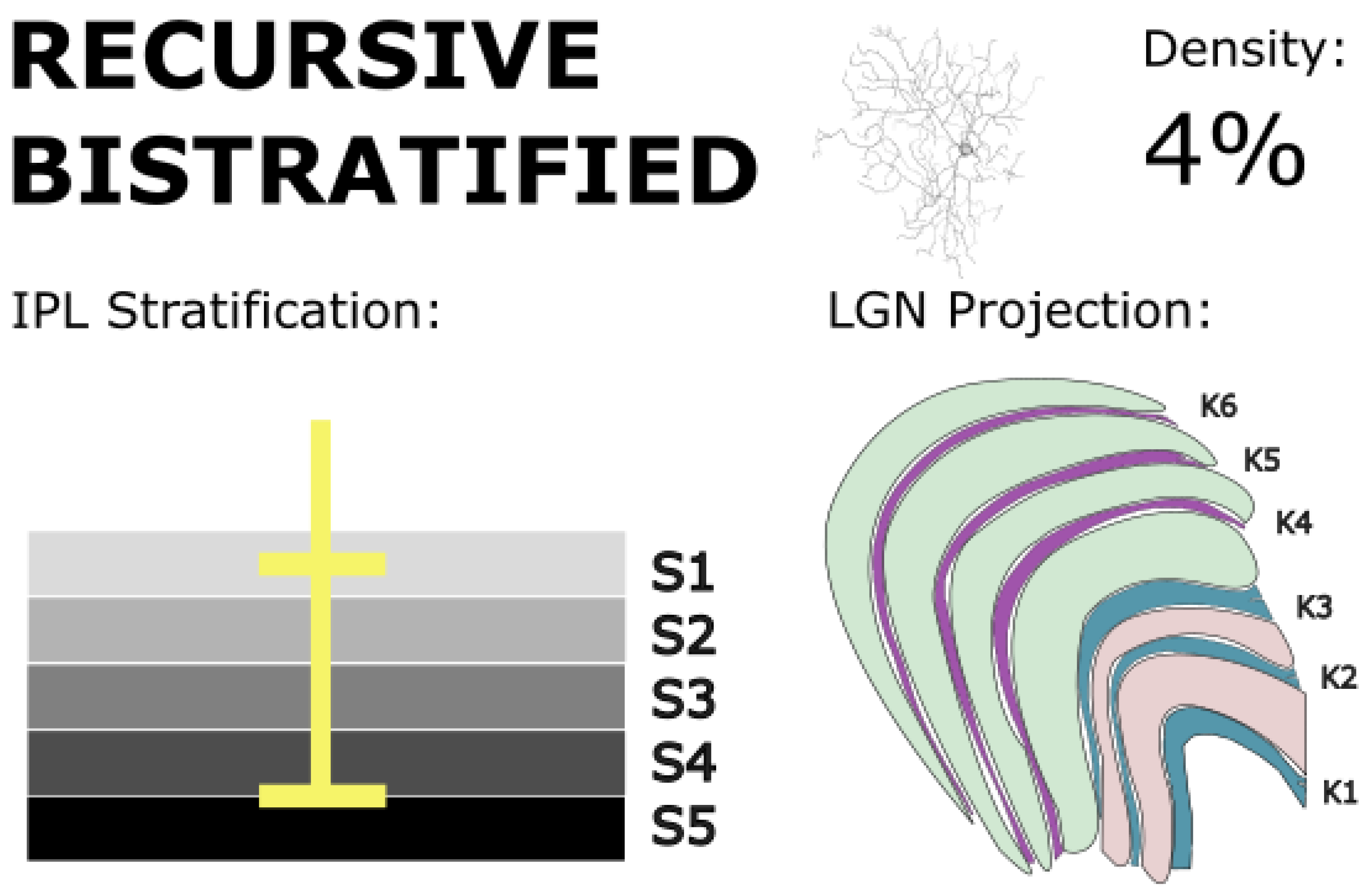
Figure 10.
Recursive bistratified RGCs bistratify at the IPL borders in S1 and S4/5, and project to koniocellular layers of the LGN and the superior colliculus. These cells exhibit direction-selective responses and may support motion detection and tracking behaviors. Golgi stain image of cell from (Kim et al., 2022), with minor modifications, used under CC BY 4.0.
Figure 10.
Recursive bistratified RGCs bistratify at the IPL borders in S1 and S4/5, and project to koniocellular layers of the LGN and the superior colliculus. These cells exhibit direction-selective responses and may support motion detection and tracking behaviors. Golgi stain image of cell from (Kim et al., 2022), with minor modifications, used under CC BY 4.0.
Little is known concerning the functional differences between recursive monostratified RGCs and recursive bistratified RGCs, however each group displays distinct morphologies implying unique functional niches. Recursive monostratified RGCs stratify narrowly in both marmosets and macaques, typically only in the on sublamina of the IPL (Crook et al., 2014; Dacey, 2004; Kim et al., 2021; Masri, Percival, et al., 2019). This architecture contrasts with other monostratified cell populations (e.g., midget, parasol, narrow thorny) which can be found in both on and off populations. In marmosets specifically, some researchers have reported the stratification of this cell type as being near the center of the IPL (stratum 3 or 4; Percival et al., 2013), while others have reported the stratification of this cell type as being at the inner edge of the IPL (stratum 5; Masri et al., 2019). Recursive monostratified RGCs make up just under 2% of RGCs at 8 mm of eccentricity (Wässle, Grünert, et al., 1989). Only one group has identified any example of recursive monostratified RGCs in marmosets which stratify in the off sublamina, but the proportion of off recursive monostratified RGCs is unknown (Percival et al., 2013).
The population of recursive monostratified RGCs identified by Masri et al. (Masri, Percival, et al., 2019), at the inner edge of the IPL, has been speculated to contact diffuse bipolar DB6 cells, which demonstrate a spectrally insensitive on response, contacting all three cone types (Jusuf et al., 2004; S. C. S. Lee et al., 2004). The population of recursive monostratified RGCs identified by Percival et al. (2013) may derive input from diffuse bipolar DB5 cells (Grünert & Martin, 2021). However, in both cases, the exact circuitry has not been defined.
Recursive bistratified RGCs have dendrites in both the on and off sublaminae and bistratify near the IPL border (Masri, Percival, et al., 2019). At 8 mm of eccentricity, these cells make up over 4% of total RGCs (Wässle, Grünert, et al., 1989), which is a relatively high percentage compared to other low-density RGC types. Both bistratified and monostratified recursive RGCs have similar tiling radius to many other low-density RGCs (e.g. large bistratified; Masri et al., 2019; Percival et al., 2013). However, the exact bipolar inputs to recursive bistratified RGCs have not been determined, although it has been proposed that these cells potentially make connections with diffuse bipolar cell types DB2, DB3a, and DB5 (Grünert & Martin, 2021; Masri et al., 2016).
Functionally, both bistratified and monostratified recursive RGCs have been proposed to play a role in direction selectivity at the retinal level (Dacey, 2004; Detwiler et al., 2019; Grünert & Martin, 2021). This conclusion is drawn from the striking morphological similarities between recursive cell types in primates and direction-selective RGCs (DSRGCs) identified in rabbit retinas (Vaney et al., 2012), along with similarities in cell markers that suggest a functional equivalence (Dhande et al., 2019; Nasir-Ahmad et al., 2021, 2022). Indeed, initial recordings from recursive bistratified RGCs confirm that recursive cells show a direction-selective response at the level of the retina (Detwiler et al., 2019).
The response characteristics of recursive RGCs can be contrasted with the broad thorny RGCs response properties – identified in macaques – which have also been implicated in motion perception (Puller et al., 2015). Unlike broad thorny RGCs, recursive bistratified RGCs show strong spiking output to motion, even background motion. However, within the recursive RGC group, there is little research on the similarities or differences in response properties between monostratified and bistratified RGCs. Any specific response differences between recursive mono- and bistratified RGCs and their functional significances should be further explored.
It is widely agreed that directional selectivity in DSRGCs must arise from temporal and spatial information that is shared between multiple cells (Percival et al., 2019; Tang et al., 2022). Thus, communication via amacrine signalling is functionally necessary, like with broad thorny RGCs. Recursive bistratified RGCs in macaques have been associated specifically with amacrine cell type A1 (Dacey et al., 2019), which has a transient on-off receptive field with a direction-selective response (Davenport et al., 2007; Kim et al., 2021). In addition to potentially helping generate direction selectivity within the recursive bistratified RGC, A1 responses may be important in inhibiting motion perception during saccadic eye movement (Roska & Werblin, 2003).
Recursive monostratified RGCs have been identified with minor projections to the LGN in both marmosets and macaques, suggesting a contribution to conscious motion perception (Dacey, 2004; Kim et al., 2021; Percival et al., 2013). Like most low density RGC types, these cells project preferentially to the K layers. Given their response characteristics, it would be expected that these projections terminate in the vK layers, however layer-specific identification of these projections has not yet been done. Interestingly, previous research has shown that almost 20% of LGN cells exhibit direction-selective responses, and the degree of direction selectivity does not vary significantly by cell class (Kim et al., 2022; X. Xu et al., 2002). While it is unlikely that all direction-selectivity across M, P, and K cell classes comes from recursive cells due to their low density, it is possible that the effects of direction-selective amacrine cells are broader than previously thought. A1 cells have been shown to form a significant portion of presynaptic input to off parasol cells and midget cells, which may be the source of the broad, cell-class-nonspecific direction selective response characteristics in the LGN (Bordt et al., 2006; Kim et al., 2021).
Mouse on-off DSRGCs, the equivalent of recursive bistratified RGCs, have been shown to project to the LGN (Huberman et al., 2009). One study identified recursive bistratified RGC projections in the LGN in primates, but no other studies have replicated these findings, suggesting that if recursive bistratified RGCs do show LGN projections in primates, they are only minor in nature (Kim et al., 2021). Recursive bistratified and monostratified RGCs also show projections to the superior colliculus (Dacey, 2004; Detwiler et al., 2019; Grünert et al., 2021; Masri, Percival, et al., 2019; Rodieck & Watanabe, 1993), where direction sensitivity may be important for orienting the head and eyes to track moving objects. Recursive cells, in conjunction with signals from broad thorny RGCs, may help form the basis of effective smooth pursuit and visuomotor tracking. In addition, one study identified a single recursive bistratified RGC projection to the inferior pulvinar, which, like LGN, may also be a minor target for this subclass (Grünert et al., 2021).
Further research is needed to understand the precise behavioral or psychophysical function of recursive cells in conscious vision. Nonetheless, it is likely that temporally specific cortical summation plays a crucial role in direction selectivity and motion perception.
5.6. Sparse Monostratified RGCs (Large Sparse RGCs)
Large sparse RGCs are a poorly studied, low-density population of RGCs, comprising only 3% of RGC types at 8 mm of eccentricity (Wässle, Grünert, et al., 1989). These cells have a larger tiling radius than any of the RGC types presented in this review so far, averaging 428 microns (Dhande et al., 2019). Like midget and parasol cells, large sparse RGCs comprise two separate populations, with an inner-IPL-stratifying on population and an outer-IPL-stratifying-off population (Grünert & Martin, 2021). The on cells stratify near the inner border of the IPL in stratum 4/5, while the off cells stratify near the outer border in stratum 1/2. It has been proposed that the on population of large sparse RGCs receives input from diffuse bipolar DB6 cells (Percival et al., 2011), and that they may contact S-on bipolar cells (Patterson et al., 2020). However, early recordings from these cells showed an S-off response (Peterson et al., 2003). One hypothesis is that S-on signals to large sparse cells are inverted by a specific amacrine cell type to form an S-off response. Large sparse cells have been confirmed to receive amacrine input (Yamada et al., 2005), and a blue-inverting amacrine cell type has been identified in ground squirrels (Li & Chen, 2011). This idea of a blue-inverting amacrine cell has also been proposed for other cell types, such as giant intrinsically photosensitive RGCs in primates (Mills et al., 2014; Patterson et al., 2020, 2022), but a specific amacrine cell type that inverts S-cone signals at large sparse RGCs remains undiscovered.
Figure 11.
Large sparse RGCs stratify in IPL layers S1/2 (off) and S4/5 (on), with projections to dorsal koniocellular layers of the LGN. Their broad dendritic fields and low density suggest a role in integrating S-cone signals over wide retinal areas for chromatic and luminance processing. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.
Figure 11.
Large sparse RGCs stratify in IPL layers S1/2 (off) and S4/5 (on), with projections to dorsal koniocellular layers of the LGN. Their broad dendritic fields and low density suggest a role in integrating S-cone signals over wide retinal areas for chromatic and luminance processing. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.
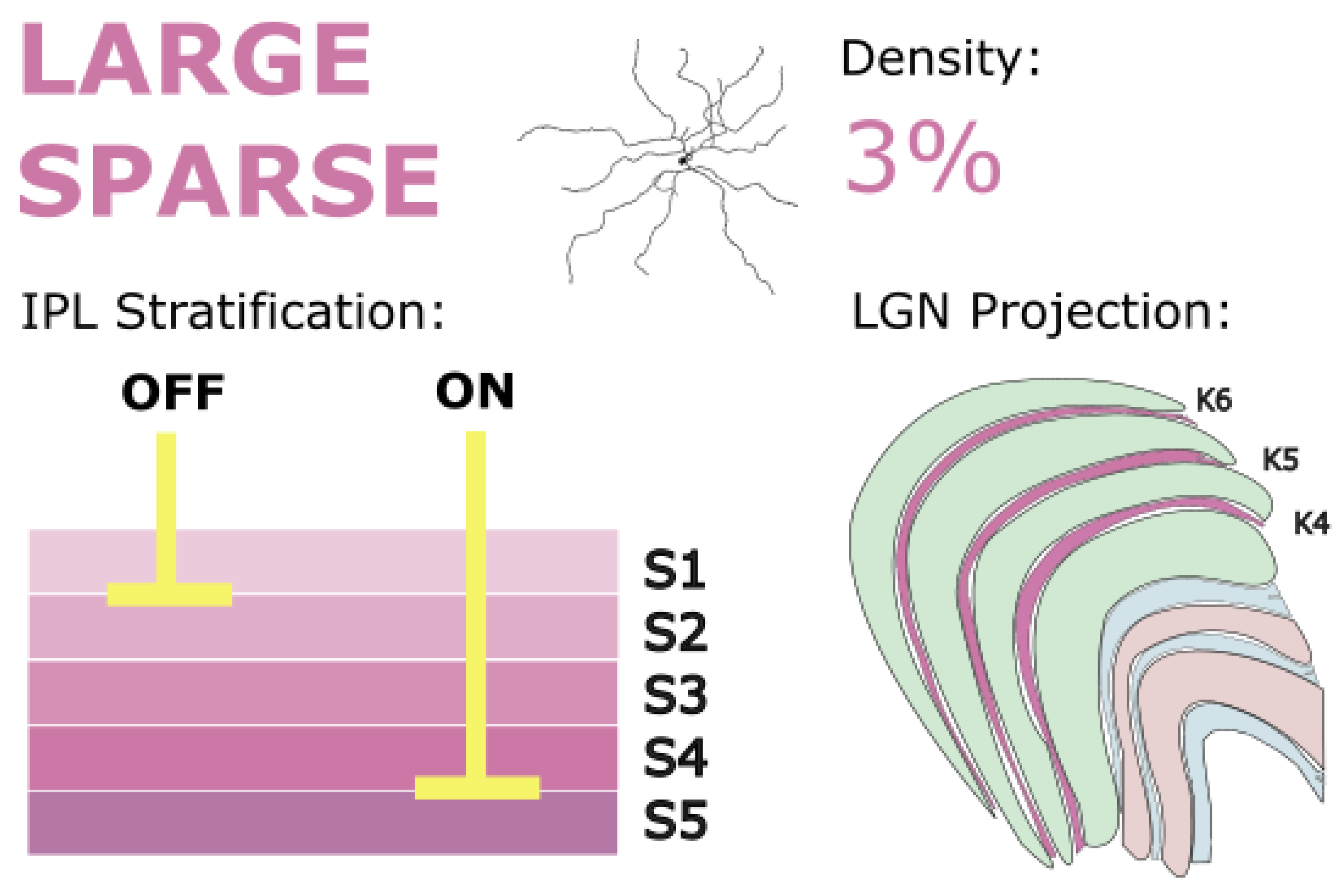
Large sparse RGCs project to LGN (Dacey, 2004; Dacey et al., 2003; Szmajda et al., 2008) and target dK layers, which is consistent with their role in carrying S-cone signals (Szmajda et al., 2006). Szmajda et al. (2008) found that large sparse RGCs occupy neighboring positions to small bistratified RGCs on a hierarchical tree, suggesting that these S-cone-carrying cells form a distinct morphological group. However, the tiling and dendritic spread of large sparse RGCs implies a functional dissimilarity between the two classes. Large sparse RGCs have sparse dendrites with few synapses; their broad tiling suggests that they might be suited for sampling holistic elements of the visual scene. Nonetheless, the acknowledgement of morphological similarities between small bistratified RGCs and large sparse RGCs raises the important point that many uncertainties concerning the contribution of small and large bistratified RGCs to conscious vision may also apply to large sparse RGCs.
5.7. Giant, Melanopsin-Containing, Intrinsically Photosensitive RGCs
Giant, melanopsin-containing, intrinsically photosensitive RGCs (giant iPRGCs) are the last low-density, LGN-projecting RGC type covered in this review. Giant iPRGCs display enormous tiling radius of up to 1 mm in the peripheral retina (Dacey et al., 2005) and exceptionally low density – from 1% (Wässle, Grünert, et al., 1989) to as low as 0.2% of all RGCs (Dacey et al., 2005). These cells comprise many distinct morphological subtypes which will not be discussed in detail for this review (Schmidt et al., 2011); here we will concentrate on purely functional distinctions. iPRGCs are monostratified and form two separate populations, with on and off populations stratifying on the very edges of the IPL, firmly in stratum 1 and stratum 5 (Grünert & Martin, 2021). These cells peak in density around the fovea, but their dendrites never enter the fovea, forming a depression called the foveal pit (Dacey, 2004; Schmidt et al., 2011). It is likely that this foveal pit exists in order to ensure that these cells with giant RFs, who sample luminosity broadly across the retina, don’t interfere with high acuity foveal vision (Berson et al., 2010; Dacey et al., 2005).
Figure 12.
Giant intrinsically photosensitive RGCs (iPRGCs) stratify in IPL layers S1 and S5 and project to koniocellular layers of the LGN as well as non-image-forming centers like the suprachiasmatic nucleus and pretectal olivary nucleus. They integrate intrinsic melanopsin-driven photosensitivity with extrinsic photoreceptor input to regulate circadian rhythms, and pupil reflexes, with a possible modulatory role in conscious vision. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.
Figure 12.
Giant intrinsically photosensitive RGCs (iPRGCs) stratify in IPL layers S1 and S5 and project to koniocellular layers of the LGN as well as non-image-forming centers like the suprachiasmatic nucleus and pretectal olivary nucleus. They integrate intrinsic melanopsin-driven photosensitivity with extrinsic photoreceptor input to regulate circadian rhythms, and pupil reflexes, with a possible modulatory role in conscious vision. Golgi stain image of cell from (Masri, Lee, et al., 2019), with minor modifications, used under CC BY-NC-ND 4.0.

What makes giant iPRGCs unique among retinal ganlion cells is that they show photosensitivity in the absence of any cone or rod connections. For iPRGCs, photosensitivity instead comes from the photopigment menanopsin that is separate from the opsins expressed within code and rod photoreceptors. Melanopsin is only distantly related to other visual pigments and shares more in common with invertebrate photopigments (Hankins et al., 2008). In addition to having intrinsic photosensitivity, giant iPRGCs also form connections with bipolar and amacrine cells (Grünert et al., 2011; Jusuf et al., 2007; Liao et al., 2016; Nasir-Ahmad et al., 2019; Patterson et al., 2020). Inner-on iPRGCs receive input from diffuse bipolar DB6 cells (Grünert et al., 2011) as well as S-on bipolar cells (Patterson et al., 2020). Physiologically, this S-on bipolar input is manifested as an S-off response (Dacey et al., 2005), again implying the presence of an S-cone-inverting amacrine pathway (Patterson et al., 2020, 2022). Given stratification depth, it is possible that outer-off iPRGCs contact diffuse bipolar DB1 cells (Grünert & Martin, 2021), however these connections are not as well defined. Broadly, iPRGCs have been shown to display connections with rod photoreceptors (Güler et al., 2008; Hannibal et al., 2017). Receiving input from both external photoreceptors and intrinsic melanopsin provides giant iPRGCs with a unique dual response propertiy. They both fire transiently with a fast onset from external input, as well as tonically with a substantially slower onset from melanopsin activation (Barrionuevo & Cao, 2016; Drouyer et al., 2007). It is thought that combination of visual opsin and melanopsin input helps these cells better detect general irradiance (Dacey, 2004; Walmsley et al., 2015), fueling their roles in entraining the circadiam rhythim though a projection to the suprachiasmatic nucleus, and in forming the light-based pupillary reflex through a projection to the pretectal olivary nucleus (Dacey, 2004; Gooley et al., 2001; Peterson et al., 2003; Sollars et al., 2003). These functions are integral parts of the visual system but are non-image forming functions of giant iPRGCs; however, they may also play a role in image-forming functions and conscious vision through their projections to LGN.
Giant iPRGCs’ projections have been found in LGN after retrograde injections to the K layers (Dacey et al., 2003; Rodieck & Watanabe, 1993). This result seems reasonable given that these retinal cells carry S-cone signals via S-on bipolar input, moreover, given the S-cone signals, one might expect that these projections target the dK layers in particular. But given the broad consensus concerning their role in non-image forming vision, the idea that giant iPRGCs have a role to play in image-forming vision has been controversial. One suggestion, by Dacey et al. (2005), is that these cells’ S-off response form the basis for the S-off/L+M-on response to mirror small bistratified RGCs role for S-on/L+M-off color vision. Others have argued against this hypothesis (Neitz & Neitz, 2017). Given the extremely large RFs of melanopsin-containing iPRGCs, it seems unlikely that these cells can mirror small bistratified RGCs and inform conscious color vision in a robust way. Additionally, from an information processing standpoint, the achromatic input from melanopsin may be difficult to separate from the chromatic input from external photoreceptors, precluding the ability of giant iPRGCs to function chromatically, although the different temporal profiles of the two responses may provide a basis for this separation.
Indeed, many reports support a role of giant iPRGCs in conscious vision. First, there have been studies done of melanopsin-containing iPRGC system in mice, which shows fundamental similarities to that of primates (Schmidt et al., 2011). Ecker and colleagues (2010) found that mice with knocked out cone and rod opsins, and thus with iPRGCs as their only functional photoreceptors, were able to discriminate high contrast gratings from uniform grey stimuli, although it is not known if this ability was from genuinely conscious vision or nonconscious blindsight-like vision. To help answer that question, Zaidi and colleagues (2007) found that humans lacking rods and cones still show conscious visual awareness. The participant in this study reported the presence of conscious percepts described as, “brightness.” However, the visual percepts exhibited very slow onset (upwards of 10 seconds), mirroring the response properties of melanopsin (Schmidt et al., 2008).
Despite these apparent limitations, melanopsin-driven responses have been shown to influence image-forming vision in primates, including potential contributions to color perception. Human ERG and VEP recordings show melanopsin–cone interactions in cortex (Adhikari et al., 2024), and psychophysical studies point to a possible role of ipRGCs in color discrimination under natural viewing conditions (Barrionuevo et al., 2024).
These experiments help reveal the functionality of giant iPRGCs in conscious vision. It is likely that this role is primarily modulatory, providing a precise signal of illuminance which is superimposed over other visually activated RFs from other RGCs (Liu et al., 2023; Schmidt et al., 2011). Like with irradiance information projected to the suprachiasmatic nucleus and principal olivary nucleus (PON) for non-image forming vision, the contribution of external photoreceptor input could help to refine this illuminance signal as well as maintain color constancy of the broad scene by reporting the spectrum of the illuminance (Thoreson & Dacey, 2019). More specifically, it’s likely that melanopsins help RGCs which show low contrast sensitivity (such as midget cells) to encode high acuity vision with greater luminance nuance and support adaptation in high luminance conditions (Barrionuevo & Cao, 2014; Brown et al., 2010). This hypothesis is supported with a physiological study finding that melanopsin helps sets the tonic firing rate of around 40% of all light responsive RFs in the LGN (Brown et al., 2010). It is also supported by studies which found that melanopsin-driven responses enhance the perception of brightness (Brown et al., 2012), and amplify cone-driven signals in cortex (Adhikari et al., 2024). These findings together with the ones discussed earlier, underscore the integrative role of giant iPRGCs in modulating visual perception, highlighting their contribution to luminance encoding and the maintenance of visual stability across varying lighting conditions, and suggesting that there may be merely a modulatory rather than a direct role in conscious image perception.
6. Summary and Conclusions
Significant progress has been made in characterizing the diversity of retinal ganglion cells (RGCs) in the primate visual system and their targets that are primarily in the lateral geniculate nucleus (LGN) and other subcortical nuclei. The different RGC types, from high-density to low-density, contribute to distinct aspects of visual processing, including high-acuity spatial vision, color discrimination, motion sensitivity, and luminance modulation. Understanding how the myriad of these parallel pathways are integrated and within and outside of the classical M/P/K framework has deepened our knowledge of the functional organization of the visual system and its remarkable capacity to encode complex and dynamic aspects of the visual scene.
Clinically, this expanding basic knowledge of RGC diversity has direct implications for the diagnosis and treatment of retinal diseases and the development of technologies for visual restoration. By identifying the vulnerabilities and resilience of different RGC types, it may be possible to develop targeted neuroprotective pharmacological strategies to preserve or restore function in affected pathways. Additionally, the success of emerging therapies—including optogenetic approaches, gene therapy, and both retinal and central visual prostheses—depends on a precise understanding of RGC circuitry. By selectively engaging specific RGC classes and tailoring stimulation strategies to their physiological properties, these interventions hold the potential to restore functional, and perhaps even naturalistic vision in patients with advanced photoreceptor degeneration.
In conclusion, a comprehensive, detailed cataloging of RGC classes, their anatomical and functional diversity, and how their outputs are integrated into the activity of central targets, is not only critical for advancing basic neuroscience but holds transformative potential for the treatment of blinding diseases and the development of visual prostheses. Much work has been done since the original Dacey review (2004) on which this report is based, but much remains for a full understanding the retina and the functional basis of conscious vision.
Figure 13.
The diversity of LGN-projecting RGC types.

Acknowledgments
The authors would like to acknowledge Dr. Paul R. Martin for helpful comments. Funding has been provided by: the National Eye Institute under aware EY029322; the NIMA Foundation; the William M. Wood Foundation, Bank of America, Trustee; Peter and Yavi Pezaris, Micheael Gersh, and Don Good.
Conflicts of Interest
The authors declare no competing interests.
References
- Adhikari, P.; Uprety, S.; Feigl, B.; Zele, A.J. Melanopsin-mediated amplification of cone signals in the human visual cortex. Proc. R. Soc. B: Biol. Sci. 2024, 291, 20232708. [Google Scholar] [CrossRef]
- Ahmadlou, M.; Zweifel, L.S.; Heimel, J.A. Functional modulation of primary visual cortex by the superior colliculus in the mouse. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Ajina, S.; Bridge, H. Blindsight relies on a functional connection between hMT+ and the lateral geniculate nucleus, not the pulvinar. PLOS Biol. 2018, 16, e2005769. [Google Scholar] [CrossRef] [PubMed]
- Al-Hashmi, A.M.; Kramer, D.J.; Mullen, K.T. Human vision with a lesion of the parvocellular pathway: an optic neuritis model for selective contrast sensitivity deficits with severe loss of midget ganglion cell function. Exp. Brain Res. 2011, 215, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Appleby, T.R.; Manookin, M.B. Selectivity to approaching motion in retinal inputs to the dorsal visual pathway. eLife 2020, 9. [Google Scholar] [CrossRef]
- Appleby, T. R. , Rieke, F., & Manookin, M. B. (). Separate pathways for processing local and global motion in the primate retina. Investigative Ophthalmology & Visual Science 2021, 62, 3039–3039. [Google Scholar]
- Atapour, N.; Worthy, K.H.; Rosa, M.G.P. Remodeling of lateral geniculate nucleus projections to extrastriate area MT following long-term lesions of striate cortex. Proc. Natl. Acad. Sci. 2022, 119. [Google Scholar] [CrossRef] [PubMed]
- Baden, T.; Berens, P.; Franke, K.; Rosón, M.R.; Bethge, M.; Euler, T. The functional diversity of retinal ganglion cells in the mouse. Nature 2016, 529, 345–350. [Google Scholar] [CrossRef]
- E Bakken, T.; van Velthoven, C.T.; Menon, V.; Hodge, R.D.; Yao, Z.; Nguyen, T.N.; Graybuck, L.T.; Horwitz, G.D.; Bertagnolli, D.; Goldy, J.; et al. Single-cell and single-nucleus RNA-seq uncovers shared and distinct axes of variation in dorsal LGN neurons in mice, non-human primates, and humans. eLife 2021, 10. [Google Scholar] [CrossRef]
- Baldicano, A.K.; Nasir-Ahmad, S.; Novelli, M.; Lee, S.C.; Do, M.T.H.; Martin, P.R.; Grünert, U. Retinal ganglion cells expressing CaM kinase II in human and nonhuman primates. J. Comp. Neurol. 2022, 530, 1470–1493. [Google Scholar] [CrossRef]
- Baldwin, M.K.; Balaram, P.; Kaas, J.H. The evolution and functions of nuclei of the visual pulvinar in primates. J. Comp. Neurol. 2017, 525, 3207–3226. [Google Scholar] [CrossRef]
- Barrionuevo, P.A.; Cao, D. Contributions of rhodopsin, cone opsins, and melanopsin to postreceptoral pathways inferred from natural image statistics. J. Opt. Soc. Am. A 2014, 31, A131–A139. [Google Scholar] [CrossRef]
- Barrionuevo, P.A.; Cao, D. Luminance and chromatic signals interact differently with melanopsin activation to control the pupil light response. J. Vis. 2016, 16, 29–29. [Google Scholar] [CrossRef]
- Barrionuevo, P.A.; Salinas, M.L.S.; Fanchini, J.M. Are ipRGCs involved in human color vision? Hints from physiology, psychophysics, and natural image statistics. Vis. Res. 2024, 217, 108378. [Google Scholar] [CrossRef]
- Benevento, L.; Miller, J. Visual responses of single neurons in the caudal lateral pulvinar of the macaque monkey. J. Neurosci. 1981, 1, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Berson, D.M.; Castrucci, A.M.; Provencio, I. Morphology and mosaics of melanopsin-expressing retinal ganglion cell types in mice. J. Comp. Neurol. 2010, 518, 2405–2422. [Google Scholar] [CrossRef] [PubMed]
- Blackshaw, S.; Sanes, J.R. Turning lead into gold: reprogramming retinal cells to cure blindness. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Bordt, A.S.; Hoshi, H.; Yamada, E.S.; Perryman-Stout, W.C.; Marshak, D.W. Synaptic input to OFF parasol ganglion cells in macaque retina. J. Comp. Neurol. 2006, 498, 46–57. [Google Scholar] [CrossRef]
- Bordt, A.S.; Patterson, S.S.; Girresch, R.J.; Perez, D.; Tseng, L.; Anderson, J.R.; Mazzaferri, M.A.; Kuchenbecker, J.A.; Gonzales-Rojas, R.; Roland, A.; et al. Synaptic inputs to broad thorny ganglion cells in macaque retina. J. Comp. Neurol. 2021, 529, 3098–3111. [Google Scholar] [CrossRef]
- Bordt, A.S.; Patterson, S.S.; Girresch, R.J.; Perez, D.; Tseng, L.; Anderson, J.R.; Mazzaferri, M.A.; Kuchenbecker, J.A.; Gonzales-Rojas, R.; Roland, A.; et al. Synaptic inputs to broad thorny ganglion cells in macaque retina. J. Comp. Neurol. 2021, 529, 3098–3111. [Google Scholar] [CrossRef]
- Boycott, B.B.; Wässle, H. Morphological Classification of Bipolar Cells of the Primate Retina. Eur. J. Neurosci. 1991, 3, 1069–1088. [Google Scholar] [CrossRef] [PubMed]
- Boycott, B.; Wässle, H. Parallel processing in the mammalian retina: the Proctor Lecture. . 1999, 40, 1313–27. [Google Scholar] [PubMed]
- Braak, H.; Bachmann, A. The percentage of projection neurons and interneurons in the human lateral geniculate-nucleus. Human Neurobiology 1985, 4, 91–95. [Google Scholar]
- Bridge, H.; Leopold, D.A.; Bourne, J.A. Adaptive Pulvinar Circuitry Supports Visual Cognition. Trends Cogn. Sci. 2016, 20, 146–157. [Google Scholar] [CrossRef]
- Briggs, F.; Usrey, W.M. Corticogeniculate feedback and visual processing in the primate. J. Physiol. 2010, 589, 33–40. [Google Scholar] [CrossRef]
- Brown, T.M.; Gias, C.; Hatori, M.; Keding, S.R.; Semo, M.; Coffey, P.J.; Gigg, J.; Piggins, H.D.; Panda, S.; Lucas, R.J. Melanopsin Contributions to Irradiance Coding in the Thalamo-Cortical Visual System. PLOS Biol. 2010, 8, e1000558. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.M.; Tsujimura, S.-I.; Allen, A.E.; Wynne, J.; Bedford, R.; Vickery, G.; Vugler, A.; Lucas, R.J. Melanopsin-Based Brightness Discrimination in Mice and Humans. Curr. Biol. 2012, 22, 1134–1141. [Google Scholar] [CrossRef]
- Calkins, D.J.; Sterling, P. Absence of spectrally specific lateral inputs to midget ganglion cells in primate retina. Nature 1996, 381, 613–615. [Google Scholar] [CrossRef]
- Calkins, D.J.; Sterling, P. Microcircuitry for Two Types of Achromatic Ganglion Cell in Primate Fovea. J. Neurosci. 2007, 27, 2646–2653. [Google Scholar] [CrossRef]
- Calkins, D.J.; Tsukamoto, Y.; Sterling, P. Microcircuitry and Mosaic of a Blue–Yellow Ganglion Cell in the Primate Retina. J. Neurosci. 1998, 18, 3373–3385. [Google Scholar] [CrossRef]
- Callaway, E.M. Structure and function of parallel pathways in the primate early visual system. J. Physiol. 2005, 566, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Casagrande, V.; Yazar, F.; Jones, K.; Ding, Y. The Morphology of the Koniocellular Axon Pathway in the Macaque Monkey. Cereb. Cortex 2007, 17, 2334–2345. [Google Scholar] [CrossRef] [PubMed]
- Casanova, C.; Chalupa, L.M. The dorsal lateral geniculate nucleus and the pulvinar as essential partners for visual cortical functions. Front. Neurosci. 2023, 17, 1258393. [Google Scholar] [CrossRef] [PubMed]
- CDC. (2022, December 19). Fast Facts of Common Eye Disorders | CDC. https://www.cdc.gov/visionhealth/basics/ced/fastfacts.
- Chandra, A.J.; Lee, S.C.S.; Grünert, U. Thorny ganglion cells in marmoset retina: Morphological and neurochemical characterization with antibodies against calretinin. J. Comp. Neurol. 2017, 525, 3962–3974. [Google Scholar] [CrossRef]
- Chatterjee, S.; Callaway, E.M. S Cone Contributions to the Magnocellular Visual Pathway in Macaque Monkey. Neuron 2002, 35, 1135–1146. [Google Scholar] [CrossRef]
- Chatterjee, S.; Callaway, E.M. Parallel colour-opponent pathways to primary visual cortex. Nature 2003, 426, 668–671. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Hoffmann, K.-P.; Distler, C.; Hafed, Z.M. The Foveal Visual Representation of the Primate Superior Colliculus. Curr. Biol. 2019, 29, 2109–2119.e7. [Google Scholar] [CrossRef]
- Cheong, S.K.; Tailby, C.; Solomon, S.G.; Martin, P.R. Cortical-Like Receptive Fields in the Lateral Geniculate Nucleus of Marmoset Monkeys. J. Neurosci. 2013, 33, 6864–6876. [Google Scholar] [CrossRef]
- Chichilnisky, E.J.; Kalmar, R.S. Temporal Resolution of Ensemble Visual Motion Signals in Primate Retina. J. Neurosci. 2003, 23, 6681–6689. [Google Scholar] [CrossRef]
- Cowey, A. Visual System: How Does Blindsight Arise? Curr. Biol. 2010, 20, R702–R704. [Google Scholar] [CrossRef]
- Crook, J.D.; Packer, O.S.; Dacey, D.M. A synaptic signature for ON- and OFF-center parasol ganglion cells of the primate retina. Vis. Neurosci. 2013, 31, 57–84. [Google Scholar] [CrossRef]
- Crook, J.D.; Peterson, B.B.; Packer, O.S.; Robinson, F.R.; Troy, J.B.; Dacey, D.M. Y-Cell Receptive Field and Collicular Projection of Parasol Ganglion Cells in Macaque Monkey Retina. J. Neurosci. 2008, 28, 11277–11291. [Google Scholar] [CrossRef]
- Dacey, D.M. Morphology of a small-field bistratified ganglion cell type in the macaque and human retina. Vis. Neurosci. 1993, 10, 1081–1098. [Google Scholar] [CrossRef]
- Dacey, D. The mosaic of midget ganglion cells in the human retina. J. Neurosci. 1993, 13, 5334–5355. [Google Scholar] [CrossRef] [PubMed]
- Dacey, D. M. (2004). Origins of Perception: Retinal Ganglion Cell Diversity and the Creation of Parallel Visual Pathways. In M. S. Gazzaniga (Ed.), The Cognitive Neurosciences Iii (p. 281). MIT Press.
- Dacey, D.M.; Brace, S. A coupled network for parasol but not midget ganglion cells in the primate retina. Vis. Neurosci. 1992, 9, 279–290. [Google Scholar] [CrossRef]
- Dacey, D. M. , Kim, Y. J., Packer, O. S., & Detwiler, P. B. ON-OFF direction selective ganglion cells in macaque monkey retina are tracer-coupled to an ON-OFF direction selective amacrine cell type. Investigative Ophthalmology & Visual Science 2019, 60, 5280–5280. [Google Scholar]
- Dacey, D.M.; Liao, H.-W.; Peterson, B.B.; Robinson, F.R.; Smith, V.C.; Pokorny, J.; Yau, K.-W.; Gamlin, P.D. Melanopsin-expressing ganglion cells in primate retina signal colour and irradiance and project to the LGN. Nature 2005, 433, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Dacey, D.M.; Packer, O.S. Colour coding in the primate retina: diverse cell types and cone-specific circuitry. Curr. Opin. Neurobiol. 2003, 13, 421–427. [Google Scholar] [CrossRef]
- Dacey, D.; Packer, O.S.; Diller, L.; Brainard, D.; Peterson, B.; Lee, B. Center surround receptive field structure of cone bipolar cells in primate retina. Vis. Res. 2000, 40, 1801–1811. [Google Scholar] [CrossRef]
- Dacey, D.M.; Petersen, M.R. Dendritic field size and morphology of midget and parasol ganglion cells of the human retina. Proc. Natl. Acad. Sci. 1992, 89, 9666–9670. [Google Scholar] [CrossRef]
- Dacey, D.M.; Peterson, B.B.; Robinson, F.R.; Gamlin, P.D. Fireworks in the Primate Retina. Neuron 2003, 37, 15–27. [Google Scholar] [CrossRef]
- Davenport, C.M.; Detwiler, P.B.; Dacey, D.M. Functional polarity of dendrites and axons of primate A1 amacrine cells. Vis. Neurosci. 2007, 24, 449–457. [Google Scholar] [CrossRef]
- Detwiler, P. B. , Crook, J., Packer, O., Robinson, F., & Dacey, D. M. The recursive bistratified ganglion cell type of the macaque monkey retina is ON-OFF direction selective. Investigative Ophthalmology & Visual Science 2019, 60, 3884–3884. [Google Scholar]
- Dhande, O.S.; Stafford, B.K.; Franke, K.; El-Danaf, R.; Percival, K.A.; Phan, A.H.; Li, P.; Hansen, B.J.; Nguyen, P.L.; Berens, P.; et al. Molecular Fingerprinting of On–Off Direction-Selective Retinal Ganglion Cells Across Species and Relevance to Primate Visual Circuits. J. Neurosci. 2018, 39, 78–95. [Google Scholar] [CrossRef]
- Drouyer, E.; Rieux, C.; Hut, R.A.; Cooper, H.M. Responses of Suprachiasmatic Nucleus Neurons to Light and Dark Adaptation: Relative Contributions of Melanopsin and Rod–Cone Inputs. J. Neurosci. 2007, 27, 9623–9631. [Google Scholar] [CrossRef]
- Ecker, J.L.; Dumitrescu, O.N.; Wong, K.Y.; Alam, N.M.; Chen, S.-K.; LeGates, T.; Renna, J.M.; Prusky, G.T.; Berson, D.M.; Hattar, S. Melanopsin-Expressing Retinal Ganglion-Cell Photoreceptors: Cellular Diversity and Role in Pattern Vision. Neuron 2010, 67, 49–60. [Google Scholar] [CrossRef]
- Eiber, C.; Pietersen, A.; Zeater, N.; Solomon, S.; Martin, P. Chromatic summation and receptive field properties of blue-on and blue-off cells in marmoset lateral geniculate nucleus. Vis. Res. 2018, 151, 41–52. [Google Scholar] [CrossRef]
- Eiber, C.D.; Rahman, A.S.; Pietersen, A.N.; Zeater, N.; Dreher, B.; Solomon, S.G.; Martin, P.R. Receptive Field Properties of Koniocellular On/Off Neurons in the Lateral Geniculate Nucleus of Marmoset Monkeys. J. Neurosci. 2018, 38, 10384–10398. [Google Scholar] [CrossRef]
- Enroth-Cugell, C.; Robson, J.G. The contrast sensitivity of retinal ganglion cells of the cat. J. Physiol. 1966, 187, 517–552. [Google Scholar] [CrossRef] [PubMed]
- Famiglietti, E.V.; Kaneko, A.; Tachibana, M. Neuronal Architecture of On and Off Pathways to Ganglion Cells in Carp Retina. Science 1977, 198, 1267–1269. [Google Scholar] [CrossRef]
- Field, G.; Chichilnisky, E. Information Processing in the Primate Retina: Circuitry and Coding. Annu. Rev. Neurosci. 2007, 30, 1–30. [Google Scholar] [CrossRef]
- Gale, S.D.; Murphy, G.J. Distinct cell types in the superficial superior colliculus project to the dorsal lateral geniculate and lateral posterior thalamic nuclei. J. Neurophysiol. 2018, 120, 1286–1292. [Google Scholar] [CrossRef]
- Ghodrati, M.; Khaligh-Razavi, S.-M.; Lehky, S.R. Towards building a more complex view of the lateral geniculate nucleus: Recent advances in understanding its role. Prog. Neurobiol. 2017, 156, 214–255. [Google Scholar] [CrossRef]
- Gooley, J.J.; Lu, J.; Chou, T.C.; Scammell, T.E.; Saper, C.B. Melanopsin in cells of origin of the retinohypothalamic tract. Nat. Neurosci. 2001, 4, 1165–1165. [Google Scholar] [CrossRef]
- Gouras, P. Identification of cone mechanisms in monkey ganglion cells. J. Physiol. 1968, 199, 533–547. [Google Scholar] [CrossRef]
- Grieve, K.L.; Acuña, C.; Cudeiro, J. The primate pulvinar nuclei: vision and action. Trends Neurosci. 2000, 23, 35–39. [Google Scholar] [CrossRef]
- Grünert, U.; Jusuf, P.R.; Lee, S.C.; Nguyen, D.T. Bipolar input to melanopsin containing ganglion cells in primate retina. Vis. Neurosci. 2010, 28, 39–50. [Google Scholar] [CrossRef]
- Grünert, U.; Lee, S.C.S.; Kwan, W.C.; Mundinano, I.-C.; Bourne, J.A.; Martin, P.R. Retinal ganglion cells projecting to superior colliculus and pulvinar in marmoset. Anat. Embryol. 2021, 226, 2745–2762. [Google Scholar] [CrossRef]
- Grünert, U.; Martin, P.R. Cell types and cell circuits in human and non-human primate retina. Prog. Retin. Eye Res. 2020, 78, 100844. [Google Scholar] [CrossRef]
- Grünert, U.; Martin, P.R. Morphology, Molecular Characterization, and Connections of Ganglion Cells in Primate Retina. Annu. Rev. Vis. Sci. 2021, 7, 73–103. [Google Scholar] [CrossRef]
- Guillery, R.; Sherman, S. Thalamic Relay Functions and Their Role in Corticocortical Communication. Neuron 2002, 33, 163–175. [Google Scholar] [CrossRef]
- Güler, A.D.; Ecker, J.L.; Lall, G.S.; Haq, S.; Altimus, C.M.; Liao, H.-W.; Barnard, A.R.; Cahill, H.; Badea, T.C.; Zhao, H.; et al. Melanopsin cells are the principal conduits for rod–cone input to non-image-forming vision. Nature 2008, 453, 102–105. [Google Scholar] [CrossRef]
- Hahn, J.; Monavarfeshani, A.; Qiao, M.; Kao, A.H.; Kölsch, Y.; Kumar, A.; Kunze, V.P.; Rasys, A.M.; Richardson, R.; Wekselblatt, J.B.; et al. Evolution of neuronal cell classes and types in the vertebrate retina. Nature 2023, 624, 415–424. [Google Scholar] [CrossRef]
- Halnes, G.; Augustinaite, S.; Heggelund, P.; Einevoll, G.T.; Migliore, M. A Multi-Compartment Model for Interneurons in the Dorsal Lateral Geniculate Nucleus. PLOS Comput. Biol. 2011, 7, e1002160. [Google Scholar] [CrossRef]
- Hankins, M.W.; Peirson, S.N.; Foster, R.G. Melanopsin: an exciting photopigment. Trends Neurosci. 2008, 31, 27–36. [Google Scholar] [CrossRef]
- Hannibal, J.; Christiansen, A.T.; Heegaard, S.; Fahrenkrug, J.; Kiilgaard, J.F. Melanopsin expressing human retinal ganglion cells: Subtypes, distribution, and intraretinal connectivity. J. Comp. Neurol. 2017, 525, 1934–1961. [Google Scholar] [CrossRef]
- Haverkamp, S. (2010). Morphology of Interneurons: Bipolar Cells. In Encyclopedia of the Eye (pp. 65–73). Elsevier. [CrossRef]
- Haverkamp, S.; Haeseleer, F.; Hendrickson, A. A comparison of immunocytochemical markers to identify bipolar cell types in human and monkey retina. Vis. Neurosci. 2003, 20, 589–600. [Google Scholar] [CrossRef]
- Hendry, S.H.C.; Reid, R.C. The Koniocellular Pathway in Primate Vision. Annu. Rev. Neurosci. 2000, 23, 127–153. [Google Scholar] [CrossRef]
- Hendry, S.H.C.; Yoshioka, T. A Neurochemically Distinct Third Channel in the Macaque Dorsal Lateral Geniculate Nucleus. Science 1994, 264, 575–577. [Google Scholar] [CrossRef]
- Hickey, T.L.; Guillery, R.W. Variability of laminar patterns in the human lateral geniculate nucleus. J. Comp. Neurol. 1979, 183, 221–246. [Google Scholar] [CrossRef]
- Horwitz, G. What studies of macaque monkeys have told us about human color vision. Neuroscience 2015, 296, 110–115. [Google Scholar] [CrossRef]
- Hu, H.; Jonas, P. A supercritical density of Na+ channels ensures fast signaling in GABAergic interneuron axons. Nat. Neurosci. 2014, 17, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Huberman, A.D.; Wei, W.; Elstrott, J.; Stafford, B.K.; Feller, M.B.; Barres, B.A. Genetic Identification of an On-Off Direction- Selective Retinal Ganglion Cell Subtype Reveals a Layer-Specific Subcortical Map of Posterior Motion. Neuron 2009, 62, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Isa, T.; Yoshida, M. Neural Mechanism of Blindsight in a Macaque Model. Neuroscience 2021, 469, 138–161. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Feldheim, D.A. The Mouse Superior Colliculus: An Emerging Model for Studying Circuit Formation and Function. Front. Neural Circuits 2018, 12, 10. [Google Scholar] [CrossRef]
- Jacobs, G.H. THE DISTRIBUTION AND NATURE OF COLOUR VISION AMONG THE MAMMALS. Biol. Rev. 1993, 68, 413–471. [Google Scholar] [CrossRef]
- Jacoby, R.A.; Marshak, D.W. Synaptic connections of DB3 diffuse bipolar cell axons in macaque retina. J. Comp. Neurol. 2000, 416, 19–29. [Google Scholar] [CrossRef]
- Jacoby, R.A.; Wiechmann, A.F.; Amara, S.G.; Leighton, B.H.; Marshak, D.W. Diffuse bipolar cells provide input to OFF parasol ganglion cells in the macaque retina. J. Comp. Neurol. 2000, 416, 6–18. [Google Scholar] [CrossRef]
- Jayakumar, J.; Dreher, B.; Vidyasagar, T.R. Tracking blue cone signals in the primate brain. Clin. Exp. Optom. 2013, 96, 259–266. [Google Scholar] [CrossRef]
- Jusuf, P.R.; Lee, S.C.; Grünert, U. Synaptic connectivity of the diffuse bipolar cell type DB6 in the inner plexiform layer of primate retina. J. Comp. Neurol. 2004, 469, 494–506. [Google Scholar] [CrossRef]
- Jusuf, P.R.; Lee, S.C.S.; Hannibal, J.; Grünert, U. Characterization and synaptic connectivity of melanopsin-containing ganglion cells in the primate retina. Eur. J. Neurosci. 2007, 26, 2906–2921. [Google Scholar] [CrossRef] [PubMed]
- Kaas, J.H.; Huerta, M.F.; Weber, J.T.; Harting, J.K. Patterns of retinal terminations and laminar organization of the lateral geniculate nucleus of primates. J. Comp. Neurol. 1978, 182, 517–553. [Google Scholar] [CrossRef] [PubMed]
- Kaas, J.H.; Lyon, D.C. Pulvinar contributions to the dorsal and ventral streams of visual processing in primates. Brain Res. Rev. 2007, 55, 285–296. [Google Scholar] [CrossRef]
- Kaplan, E. (2003). The M, P, and K Pathways of the Primate Visual System. In L. M. Chalupa & J. S. Werner (Eds.), The Visual Neurosciences, 2-vol. Set (pp. 481–493). The MIT Press. [CrossRef]
- Kaplan, E.; Shapley, R.M. X and Y cells in the lateral geniculate nucleus of macaque monkeys. J. Physiol. 1982, 330, 125–143. [Google Scholar] [CrossRef]
- Kastner, S. , Schneider, K. A., & Wunderlich, K. (2006). Beyond a relay nucleus: Neuroimaging views on the human LGN. In Progress in Brain Research (Vol. 155, pp. 125–143). Elsevier. [CrossRef]
- Kim, Y.J.; Peterson, B.B.; Crook, J.D.; Joo, H.R.; Wu, J.; Puller, C.; Robinson, F.R.; Gamlin, P.D.; Yau, K.-W.; Viana, F.; et al. Origins of direction selectivity in the primate retina. Nat. Commun. 2022, 13, 1–20. [Google Scholar] [CrossRef]
- Kim, Y.J.; Peterson, B.B.; Crook, J.D.; Joo, H.R.; Wu, J.; Puller, C.; Robinson, F.R.; Gamlin, P.D.; Yau, K.-W.; Viana, F.; et al. Origins of direction selectivity in the primate retina. Nat. Commun. 2022, 13, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Evrard, H.C.; Shapcott, K.A.; Haverkamp, S.; Logothetis, N.K.; Schmid, M.C. Cell-Targeted Optogenetics and Electrical Microstimulation Reveal the Primate Koniocellular Projection to Supra-granular Visual Cortex. Neuron 2016, 90, 143–151. [Google Scholar] [CrossRef]
- Kolb, H. (1995). Roles of Amacrine Cells. In H. Kolb, E. Fernandez, & R. Nelson (Eds.), Webvision: The Organization of the Retina and Visual System. University of Utah Health Sciences Center. http://www.ncbi.nlm.nih.gov/books/NBK11539/.
- Kolb, H.; Linberg, K.A.; Fisher, S.K. Neurons of the human retina: A Golgi study. J. Comp. Neurol. 1992, 318, 147–187. [Google Scholar] [CrossRef]
- Kuffler, S.W. Discharge patterns and functional organization of mammalian retina. J. Neurophysiol. 1953, 16, 37–68. [Google Scholar] [CrossRef]
- Lee, B.B. Receptive field structure in the primate retina. Vis. Res. 1996, 36, 631–644. [Google Scholar] [CrossRef]
- Lee, B.B. Receptive field structure in the primate retina. Vis. Res. 1996, 36, 631–644. [Google Scholar] [CrossRef]
- Lee, S.C.; Jusuf, P.R.; Grünert, U. S-cone connections of the diffuse bipolar cell type DB6 in macaque monkey retina. J. Comp. Neurol. 2004, 474, 353–363. [Google Scholar] [CrossRef]
- Lee, S.C.S.; Wei, A.J.; Martin, P.R.; Grünert, U. Thorny and Tufted Retinal Ganglion Cells Express the Transcription Factor Forkhead Proteins Foxp1 and Foxp2 in Marmoset (Callithrix jacchus). J. Comp. Neurol. 2024, 532, e25663. [Google Scholar] [CrossRef]
- Lennie, P.; Dzmura, M. Mechanisms of color-vision. Critical Reviews in Neurobiology 1988, 3, 333–400. [Google Scholar] [PubMed]
- Li, W. , & Chen, S. The Origin of the S-Off Signal in a Mammalian Retina. Investigative Ophthalmology & Visual Science 2011, 52, 3027–3027. [Google Scholar]
- Liao, H.; Ren, X.; Peterson, B.B.; Marshak, D.W.; Yau, K.; Gamlin, P.D.; Dacey, D.M. Melanopsin-expressing ganglion cells on macaque and human retinas form two morphologically distinct populations. J. Comp. Neurol. 2016, 524, 2845–2872. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Milner, E.S.; Peng, Y.-R.; Blume, H.A.; Brown, M.C.; Bryman, G.S.; Emanuel, A.J.; Morquette, P.; Viet, N.-M.; Sanes, J.R.; et al. Encoding of environmental illumination by primate melanopsin neurons. Science 2023, 379, 376–381. [Google Scholar] [CrossRef]
- Ludwig, P.E.; Huff, T.J.; Zuniga, J.M. The potential role of bioengineering and three-dimensional printing in curing global corneal blindness. J. Tissue Eng. 2018, 9, 2041731418769863. [Google Scholar] [CrossRef]
- Ma, I.C.; Nasir-Ahmad, S.; Lee, S.C.; Grünert, U.; Martin, P.R. Contribution of parasol-magnocellular pathway ganglion cells to foveal retina in macaque monkey. Vis. Res. 2022, 202, 108154. [Google Scholar] [CrossRef]
- Malpeli, J. G. , Lee, D., & Baker, F. H. Laminar and retinotopic organization of the macaque lateral geniculate nucleus: Magnocellular and parvocellular magnification functions. The Journal of Comparative Neurology 1996, 375, 363–377. [Google Scholar] [CrossRef]
- Manookin, M.B.; Patterson, S.S.; Linehan, C.M. Neural Mechanisms Mediating Motion Sensitivity in Parasol Ganglion Cells of the Primate Retina. Neuron 2018, 97, 1327–1340.e4. [Google Scholar] [CrossRef]
- Mariani, A.P. Amacrine cells of the rhesus monkey retina. J. Comp. Neurol. 1990, 301, 382–400. [Google Scholar] [CrossRef]
- Marshak, D.W.; Mills, S.L. Short-wavelength cone-opponent retinal ganglion cells in mammals. Vis. Neurosci. 2014, 31, 165–175. [Google Scholar] [CrossRef]
- Martin, P.R.; Blessing, E.M.; Buzás, P.; Szmajda, B.A.; Forte, J.D. Transmission of colour and acuity signals by parvocellular cells in marmoset monkeys. J. Physiol. 2011, 589, 2795–2812. [Google Scholar] [CrossRef]
- Martin, P.R.; White, A.J.R.; Goodchild, A.K.; Wilder, H.D.; Sefton, A.E. Evidence that Blue-on Cells are Part of the Third Geniculocortical Pathway in Primates. Eur. J. Neurosci. 1997, 9, 1536–1541. [Google Scholar] [CrossRef] [PubMed]
- Masri, R.A.; Lee, S.C.S.; Madigan, M.C.; Grünert, U. Particle-Mediated Gene Transfection and Organotypic Culture of Postmortem Human Retina. Transl. Vis. Sci. Technol. 2019, 8, 7–7. [Google Scholar] [CrossRef] [PubMed]
- Masri, R.A.; Percival, K.A.; Koizumi, A.; Martin, P.R.; Grünert, U. Connectivity between the OFF bipolar type DB3a and six types of ganglion cell in the marmoset retina. J. Comp. Neurol. 2015, 524, 1839–1858. [Google Scholar] [CrossRef] [PubMed]
- Masri, R.A.; Percival, K.A.; Koizumi, A.; Martin, P.R.; Grünert, U. Survey of retinal ganglion cell morphology in marmoset. J. Comp. Neurol. 2017, 527, 236–258. [Google Scholar] [CrossRef]
- Meredith, M.A.; Stein, B.E. Visual, auditory, and somatosensory convergence on cells in superior colliculus results in multisensory integration. J. Neurophysiol. 1986, 56, 640–662. [Google Scholar] [CrossRef]
- Merigan, W.; Katz, L.; Maunsell, J. The effects of parvocellular lateral geniculate lesions on the acuity and contrast sensitivity of macaque monkeys. J. Neurosci. 1991, 11, 994–1001. [Google Scholar] [CrossRef]
- Michael, C.R. Retinal afferent arborization patterns, dendritic field orientations, and the segregation of function in the lateral geniculate nucleus of the monkey. Proc. Natl. Acad. Sci. 1988, 85, 4914–4918. [Google Scholar] [CrossRef]
- Mills, S.L.; Tian, L.-M.; Hoshi, H.; Whitaker, C.M.; Massey, S.C. Three Distinct Blue-Green Color Pathways in a Mammalian Retina. J. Neurosci. 2014, 34, 1760–1768. [Google Scholar] [CrossRef]
- Mirochnik, R.M.; Pezaris, J.S. Contemporary approaches to visual prostheses. Mil. Med Res. 2019, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Moritoh, S.; Komatsu, Y.; Yamamori, T.; Koizumi, A. Diversity of Retinal Ganglion Cells Identified by Transient GFP Transfection in Organotypic Tissue Culture of Adult Marmoset Monkey Retina. PLOS ONE 2013, 8, e54667. [Google Scholar] [CrossRef] [PubMed]
- Mullen, K.T.; Kingdom, F.A. Differential distributions of red–green and blue–yellow cone opponency across the visual field. Vis. Neurosci. 2002, 19, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Murray, K.D.; Rubin, C.M.; Jones, E.G.; Chalupa, L.M. Molecular Correlates of Laminar Differences in the Macaque Dorsal Lateral Geniculate Nucleus. J. Neurosci. 2008, 28, 12010–12022. [Google Scholar] [CrossRef]
- Nasir-Ahmad, S.; Lee, S.C.; Martin, P.R.; Grünert, U. Melanopsin-expressing ganglion cells in human retina: Morphology, distribution, and synaptic connections. J. Comp. Neurol. 2017, 527, 312–327. [Google Scholar] [CrossRef]
- Nasir-Ahmad, S.; Lee, S.C.S.; Martin, P.R.; Grünert, U. Identification of retinal ganglion cell types expressing the transcription factor Satb2 in three primate species. J. Comp. Neurol. 2021, 529, 2727–2749. [Google Scholar] [CrossRef]
- Nasir-Ahmad, S.; Vanstone, K.A.; Novelli, M.; Lee, S.C.S.; Do, M.T.H.; Martin, P.R.; Grünert, U. Satb1 expression in retinal ganglion cells of marmosets, macaques, and humans. J. Comp. Neurol. 2021, 530, 923–940. [Google Scholar] [CrossRef]
- Neitz, J.; Neitz, M. Evolution of the circuitry for conscious color vision in primates. Eye 2016, 31, 286–300. [Google Scholar] [CrossRef]
- Nelson, R.; Famiglietti, E.V.; Kolb, H. Intracellular staining reveals different levels of stratification for on- and off-center ganglion cells in cat retina. J. Neurophysiol. 1978, 41, 472–483. [Google Scholar] [CrossRef] [PubMed]
- Patterson, S.S.; Girresch, R.J.; Mazzaferri, M.A.; Bordt, A.S.; Piñon-Teal, W.L.; Jesse, B.D.; Perera, D.-C.W.; Schlepphorst, M.A.; Kuchenbecker, J.A.; Chuang, A.Z.; et al. Synaptic Origins of the Complex Receptive Field Structure in Primate Smooth Monostratified Retinal Ganglion Cells. eneuro 2024, 11. [Google Scholar] [CrossRef]
- Patterson, S.S.; Mazzaferri, M.A.; Bordt, A.S.; Chang, J.; Neitz, M.; Neitz, J. Another Blue-ON ganglion cell in the primate retina. Curr. Biol. 2020, 30, R1409–R1410. [Google Scholar] [CrossRef]
- Patterson, S.S.; Neitz, M.; Neitz, J. Reconciling Color Vision Models With Midget Ganglion Cell Receptive Fields. Front. Neurosci. 2019, 13, 865. [Google Scholar] [CrossRef]
- Patterson, S.S.; Neitz, M.; Neitz, J. S-cone circuits in the primate retina for non-image-forming vision. Semin. Cell Dev. Biol. 2021, 126, 66–70. [Google Scholar] [CrossRef]
- Peng, Y.-R.; Shekhar, K.; Yan, W.; Herrmann, D.; Sappington, A.; Bryman, G.S.; van Zyl, T.; Do, M.T.H.; Regev, A.; Sanes, J.R. Molecular Classification and Comparative Taxonomics of Foveal and Peripheral Cells in Primate Retina. Cell 2019, 176, 1222–1237.e22. [Google Scholar] [CrossRef]
- Percival, K.A.; Koizumi, A.; Masri, R.A.; Buzás, P.; Martin, P.R.; Grünert, U. Identification of a Pathway from the Retina to Koniocellular Layer K1 in the Lateral Geniculate Nucleus of Marmoset. J. Neurosci. 2014, 34, 3821–3825. [Google Scholar] [CrossRef]
- Percival, K.A.; Martin, P.R.; Grünert, U. Synaptic inputs to two types of koniocellular pathway ganglion cells in marmoset retina. J. Comp. Neurol. 2010, 519, 2135–2153. [Google Scholar] [CrossRef]
- Percival, K.A.; Martin, P.R.; Grünert, U. Organisation of koniocellular-projecting ganglion cells and diffuse bipolar cells in the primate fovea. Eur. J. Neurosci. 2013, 37, 1072–1089. [Google Scholar] [CrossRef] [PubMed]
- Percival, K.A.; Venkataramani, S.; Smith, R.G.; Taylor, W.R. Directional excitatory input to direction-selective ganglion cells in the rabbit retina. J. Comp. Neurol. 2017, 527, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Perry, V.; Oehler, R.; Cowey, A. Retinal ganglion cells that project to the dorsal lateral geniculate nucleus in the macaque monkey. Neuroscience 1984, 12, 1101–1123. [Google Scholar] [CrossRef]
- Peterson, B. B. , Liao, H., Dacey, D. M., Yau, K., Gamlin, P. D., Robinson, F. R., & Marshak, D. W. Functional architecture of the photoreceptive ganglion cell in primate retina: Morphology, mosaic organization and central targets of melanopsin immunostained cells. Investigative Ophthalmology & Visual Science 2003, 44, 5182–5182. [Google Scholar]
- Pietersen, A.N.J.; Cheong, S.K.; Solomon, S.G.; Tailby, C.; Martin, P.R. Temporal response properties of koniocellular (blue-on and blue-off) cells in marmoset lateral geniculate nucleus. J. Neurophysiol. 2014, 112, 1421–1438. [Google Scholar] [CrossRef]
- Pregowska, A.; Casti, A.; Kaplan, E.; Wajnryb, E.; Szczepanski, J. Information processing in the LGN: a comparison of neural codes and cell types. Biol. Cybern. 2019, 113, 453–464. [Google Scholar] [CrossRef]
- Puller, C.; Manookin, M.B.; Neitz, J.; Rieke, F.; Neitz, M. Broad Thorny Ganglion Cells: A Candidate for Visual Pursuit Error Signaling in the Primate Retina. J. Neurosci. 2015, 35, 5397–5408. [Google Scholar] [CrossRef]
- Puller, C. , Manookin, M., Neitz, M., Rieke, F., & Neitz, J. (). Response properties of broad thorny ganglion cells in the primate retina. Investigative Ophthalmology & Visual Science 2013, 54, 1295–1295. [Google Scholar]
- Puthussery, T.; Percival, K.A.; Venkataramani, S.; Gayet-Primo, J.; Grünert, U.; Taylor, W.R. Kainate Receptors Mediate Synaptic Input to Transient and Sustained OFF Visual Pathways in Primate Retina. J. Neurosci. 2014, 34, 7611–7621. [Google Scholar] [CrossRef]
- Ramón y Cajal, S. (1972). The structure of the retina. C. C. Thomas.
- Rhoades, C.E.; Shah, N.P.; Manookin, M.B.; Brackbill, N.; Kling, A.; Goetz, G.; Sher, A.; Litke, A.M.; Chichilnisky, E. Unusual Physiological Properties of Smooth Monostratified Ganglion Cell Types in Primate Retina. Neuron 2019, 103, 658–672.e6. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.L.; Petersen, S.E. The pulvinar and visual salience. Trends Neurosci. 1992, 15, 127–132. [Google Scholar] [CrossRef]
- Rodieck, R. W. (1991). Which Cells Code for Color? In A. Valberg & B. B. Lee (Eds.), From Pigments to Perception: Advances in Understanding Visual Processes (pp. 83–93). Springer US. [CrossRef]
- Rodieck, R.W.; Watanabe, M. Survey of the morphology of macaque retinal ganglion cells that project to the pretectum, superior colliculus, and parvicellular laminae of the lateral geniculate nucleus. J. Comp. Neurol. 1993, 338, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, A.; Talebi, V. The Primate Retina Contains Distinct Types of Y-Like Ganglion Cells: Figure 1. J. Neurosci. 2009, 29, 5048–5050. [Google Scholar] [CrossRef]
- Roska, B.; Werblin, F. Rapid global shifts in natural scenes block spiking in specific ganglion cell types. Nat. Neurosci. 2003, 6, 600–608. [Google Scholar] [CrossRef]
- Roy, S.; Jayakumar, J.; Martin, P.R.; Dreher, B.; Saalmann, Y.B.; Hu, D.; Vidyasagar, T.R. Segregation of short-wavelength-sensitive (S) cone signals in the macaque dorsal lateral geniculate nucleus. Eur. J. Neurosci. 2009, 30, 1517–1526. [Google Scholar] [CrossRef]
- Sanes, J.R.; Masland, R.H. The Types of Retinal Ganglion Cells: Current Status and Implications for Neuronal Classification. Annu. Rev. Neurosci. 2015, 38, 221–246. [Google Scholar] [CrossRef]
- Schiller, P.H. Parallel information processing channels created in the retina. Proc. Natl. Acad. Sci. 2010, 107, 17087–17094. [Google Scholar] [CrossRef] [PubMed]
- Schiller, P.H.; Logothetis, N.K. The color-opponent and broad-band channels of the primate visual system. Trends Neurosci. 1990, 13, 392–398. [Google Scholar] [CrossRef]
- Schiller, P.H.; Logothetis, N.K.; Charles, E.R. Functions of the colour-opponent and broad-band channels of the visual system. Nature 1990, 343, 68–70. [Google Scholar] [CrossRef]
- Schiller, P.H.; Tehovnik, E.J. Vision and the Visual System; Oxford University Press (OUP): Oxford, Oxfordshire, United Kingdom, 2015. [Google Scholar]
- Schmid, M.C.; Mrowka, S.W.; Turchi, J.; Saunders, R.C.; Wilke, M.; Peters, A.J.; Ye, F.Q.; Leopold, D.A. Blindsight depends on the lateral geniculate nucleus. Nature 2010, 466, 373–377. [Google Scholar] [CrossRef]
- Schmidt, T.M.; Do, M.T.H.; Dacey, D.; Lucas, R.; Hattar, S.; Matynia, A. Melanopsin-Positive Intrinsically Photosensitive Retinal Ganglion Cells: From Form to Function. J. Neurosci. 2011, 31, 16094–16101. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.M.; Taniguchi, K.; Kofuji, P. Intrinsic and Extrinsic Light Responses in Melanopsin-Expressing Ganglion Cells During Mouse Development. J. Neurophysiol. 2008, 100, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Sciaccotta, F.; Kipcak, A.; Erisir, A. Morphological and Molecular Distinctions of Parallel Processing Streams Reveal Two Koniocellular Pathways in the Tree Shrew DLGN. eneuro 2025, 12. [Google Scholar] [CrossRef]
- Shekhar, K.; Sanes, J.R. Generating and Using Transcriptomically Based Retinal Cell Atlases. Annu. Rev. Vis. Sci. 2021, 7, 43–72. [Google Scholar] [CrossRef]
- Singh, M.S.; Park, S.S.; Albini, T.A.; Canto-Soler, M.V.; Klassen, H.; MacLaren, R.E.; Takahashi, M.; Nagiel, A.; Schwartz, S.D.; Bharti, K. Retinal stem cell transplantation: Balancing safety and potential. Prog. Retin. Eye Res. 2020, 75, 100779. [Google Scholar] [CrossRef]
- Sollars, P.J.; Smeraski, C.A.; Kaufman, J.D.; Ogilvie, M.D.; Provencio, I.; Pickard, G.E. Melanopsin and non-melanopsin expressing retinal ganglion cells innervate the hypothalamic suprachiasmatic nucleus. Vis. Neurosci. 2003, 20, 601–610. [Google Scholar] [CrossRef]
- Solomon, S. G. (2021). Retinal ganglion cells and the magnocellular, parvocellular, and koniocellular subcortical visual pathways from the eye to the brain. In Handbook of Clinical Neurology (Vol. 178, pp. 31–50). Elsevier. [CrossRef]
- Solomon, S.G.; Rosa, M.G.P. A simpler primate brain: the visual system of the marmoset monkey. Front. Neural Circuits 2014, 8. [Google Scholar] [CrossRef]
- Solomon, S.G.; White, A.J.R.; Martin, P.R. Extraclassical Receptive Field Properties of Parvocellular, Magnocellular, and Koniocellular Cells in the Primate Lateral Geniculate Nucleus. J. Neurosci. 2002, 22, 338–349. [Google Scholar] [CrossRef]
- Soto, F.; Hsiang, J.-C.; Rajagopal, R.; Piggott, K.; Harocopos, G.J.; Couch, S.M.; Custer, P.; Morgan, J.L.; Kerschensteiner, D. Efficient Coding by Midget and Parasol Ganglion Cells in the Human Retina. Neuron 2020, 107, 656–666.e5. [Google Scholar] [CrossRef] [PubMed]
- Stacy, A. K. , & Van Hooser, S. D. Development of Functional Properties in the Early Visual System: New Appreciations of the Roles of Lateral Geniculate Nucleus. Current Topics in Behavioral Neurosciences 2022, 53, 3–35. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Smithson, H.E.; Zaidi, Q.; Lee, B.B. Specificity of Cone Inputs to Macaque Retinal Ganglion Cells. J. Neurophysiol. 2006, 95, 837–849. [Google Scholar] [CrossRef]
- Sun, S. H. , Renshaw, K., & Pezaris, J. S. (2024). Single-cell transcriptomic analysis of macaque LGN neurons reveals novel subpopulations (p. 2024.11.14.623611). bioRxiv. [CrossRef]
- Szmajda, B.A.; Buzás, P.; FitzGibbon, T.; Martin, P.R. Geniculocortical relay of blue-off signals in the primate visual system. Proc. Natl. Acad. Sci. 2006, 103, 19512–19517. [Google Scholar] [CrossRef] [PubMed]
- Szmajda, B.A.; Grünert, U.; Martin, P.R. Retinal ganglion cell inputs to the koniocellular pathway. J. Comp. Neurol. 2008, 510, 251–268. [Google Scholar] [CrossRef]
- Tailby, C.; Solomon, S.G.; Dhruv, N.T.; Majaj, N.J.; Sokol, S.H.; Lennie, P. A New Code for Contrast in the Primate Visual Pathway. J. Neurosci. 2007, 27, 3904–3909. [Google Scholar] [CrossRef] [PubMed]
- Tailby, C.; Solomon, S.G.; Lennie, P. Functional Asymmetries in Visual Pathways Carrying S-Cone Signals in Macaque. J. Neurosci. 2008, 28, 4078–4087. [Google Scholar] [CrossRef]
- Tang, C.; Todo, Y.; Ji, J.; Tang, Z. A novel motion direction detection mechanism based on dendritic computation of direction-selective ganglion cells. Knowledge-Based Syst. 2022, 241. [Google Scholar] [CrossRef]
- Thoreson, W.B.; Dacey, D.M. Diverse Cell Types, Circuits, and Mechanisms for Color Vision in the Vertebrate Retina. Physiol. Rev. 2019, 99, 1527–1573. [Google Scholar] [CrossRef]
- Usrey, W.M.; Reppas, J.B.; Reid, R.C. Specificity and Strength of Retinogeniculate Connections. J. Neurophysiol. 1999, 82, 3527–3540. [Google Scholar] [CrossRef]
- Vaney, D.I.; Sivyer, B.; Taylor, W.R. Direction selectivity in the retina: symmetry and asymmetry in structure and function. Nat. Rev. Neurosci. 2012, 13, 194–208. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, L.; Hanna, L.; Mouland, J.; Martial, F.; West, A.; Smedley, A.R.; A Bechtold, D.; Webb, A.R.; Lucas, R.J.; Brown, T.M. Colour As a Signal for Entraining the Mammalian Circadian Clock. PLOS Biol. 2015, 13, e1002127–e1002127. [Google Scholar] [CrossRef] [PubMed]
- Walsh, N.; Ghosh, K.K.; FitzGibbon, T. Intraretinal axon diameters of a New World primate, the marmoset (Callithrix jacchus). Clin. Exp. Ophthalmol. 2000, 28, 423–430. [Google Scholar] [CrossRef]
- Warner, C.E.; Kwan, W.C.; Wright, D.; Johnston, L.A.; Egan, G.F.; Bourne, J.A. Preservation of Vision by the Pulvinar following Early-Life Primary Visual Cortex Lesions. Curr. Biol. 2015, 25, 424–434. [Google Scholar] [CrossRef]
- Wässle, H. Parallel processing in the mammalian retina. Nat. Rev. Neurosci. 2004, 5, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Wässle, H.; Boycott, B.B.; Röhrenbeck, J. Horizontal Cells in the Monkey Retina: Cone connections and dendritic network. Eur. J. Neurosci. 1989, 1, 421–435. [Google Scholar] [CrossRef]
- Wässle, H.; Grünert, U.; Röhrenbeck, J.; Boycott, B.B. Cortical magnification factor and the ganglion cell density of the primate retina. Nature 1989, 341, 643–646. [Google Scholar] [CrossRef]
- Watanabe, M.; Rodieck, R.W. Parasol and midget ganglion cells of the primate retina. J. Comp. Neurol. 1989, 289, 434–454. [Google Scholar] [CrossRef]
- White, A.J.R.; Solomon, S.G.; Martin, P.R. Spatial properties of koniocellular cells in the lateral geniculate nucleus of the marmoset Callithrix jacchus. J. Physiol. 2001, 533, 519–535. [Google Scholar] [CrossRef]
- White, B.J.; Kan, J.Y.; Levy, R.; Itti, L.; Munoz, D.P. Superior colliculus encodes visual saliency before the primary visual cortex. Proc. Natl. Acad. Sci. 2017, 114, 9451–9456. [Google Scholar] [CrossRef]
- WHO. (2022, October 13). Vision impairment and blindness. https://www.who.int/news-room/fact-sheets/detail/blindness-and-visual-impairment.
- Wienbar, S.; Schwartz, G.W. The dynamic receptive fields of retinal ganglion cells. Prog. Retin. Eye Res. 2018, 67, 102–117. [Google Scholar] [CrossRef]
- Wiesel, T.N.; Hubel, D.H. Spatial and chromatic interactions in the lateral geniculate body of the rhesus monkey. J. Neurophysiol. 1966, 29, 1115–1156. [Google Scholar] [CrossRef]
- Xu, X.; Ichida, J.M.; Allison, J.D.; Boyd, J.D.; Bonds, A.B.; Casagrande, V.A. A comparison of koniocellular, magnocellular and parvocellular receptive field properties in the lateral geniculate nucleus of the owl monkey (Aotus trivirgatus). J. Physiol. 2001, 531, 203–218. [Google Scholar] [CrossRef]
- Xu, X.; Ichida, J.; Shostak, Y.; Bonds, A.; Casagrande, V.A. Are primate lateral geniculate nucleus (LGN) cells really sensitive to orientation or direction? Vis. Neurosci. 2002, 19, 97–108. [Google Scholar] [CrossRef]
- Xu, Y.; Vasudeva, V.; Vardi, N.; Sterling, P.; Freed, M.A. Different types of ganglion cell share a synaptic pattern. J. Comp. Neurol. 2008, 507, 1871–1878. [Google Scholar] [CrossRef] [PubMed]
- Yamada, E.S.; Bordt, A.S.; Marshak, D.W. Wide-field ganglion cells in macaque retinas. Vis. Neurosci. 2005, 22, 383–393. [Google Scholar] [CrossRef]
- Yan, W.; Peng, Y.-R.; van Zyl, T.; Regev, A.; Shekhar, K.; Juric, D.; Sanes, J.R. Cell Atlas of The Human Fovea and Peripheral Retina. Sci. Rep. 2020, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-H.; Atapour, N.; Chaplin, T.A.; Worthy, K.H.; Rosa, M.G. Robust Visual Responses and Normal Retinotopy in Primate Lateral Geniculate Nucleus following Long-term Lesions of Striate Cortex. J. Neurosci. 2018, 38, 3955–3970. [Google Scholar] [CrossRef]
- Yu, H.-H.; Chaplin, T.A.; Egan, G.W.; Reser, D.H.; Worthy, K.H.; Rosa, M.G.P. Visually Evoked Responses in Extrastriate Area MT after Lesions of Striate Cortex in Early Life. J. Neurosci. 2013, 33, 12479–12489. [Google Scholar] [CrossRef]
- Zaidi, F.H.; Hull, J.T.; Peirson, S.N.; Wulff, K.; Aeschbach, D.; Gooley, J.J.; Brainard, G.C.; Gregory-Evans, K.; Rizzo, J.F.; Czeisler, C.A.; et al. Short-Wavelength Light Sensitivity of Circadian, Pupillary, and Visual Awareness in Humans Lacking an Outer Retina. Curr. Biol. 2007, 17, 2122–2128. [Google Scholar] [CrossRef]
- Zeater, N.; Buzás, P.; Dreher, B.; Grünert, U.; Martin, P.R. Projections of three subcortical visual centers to marmoset lateral geniculate nucleus. J. Comp. Neurol. 2018, 527, 535–545. [Google Scholar] [CrossRef]
- Zhang, L.; Cavallini, M.; Wang, J.; Xin, R.; Zhang, Q.; Feng, G.; Sanes, J.R.; Peng, Y.-R. Evolutionary and developmental specialization of foveal cell types in the marmoset. Proc. Natl. Acad. Sci. 2024, 121. [Google Scholar] [CrossRef]
- Zheng, Y.J.; Adams, D.L.; Gentry, T.N.; Dilbeck, M.D.; Economides, J.R.; Horton, J.C. Retinal Input to Macaque Superior Colliculus Derives from Branching Axons Projecting to the Lateral Geniculate Nucleus. J. Neurosci. 2024, 44. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



