
Submitted:
13 October 2025
Posted:
15 October 2025
You are already at the latest version
Abstract
Pancreatic ductal adenocarcinoma (PDAC) is a highly aggressive disease with limited treatment success and strong resistance to therapy. Natural killer (NK) cells are important immune cells that can destroy cancer cells, and their function can be improved through certain micronutrients and dietary supplements. This review explores a "triple NK cell boost" strategy—enhancing NK cell frequency, cancer-killing ability, and ability to reach tumors—using antioxidants and food-derived supplements. These compounds may help reduce the harmful effects of reactive species which can weaken the immune system and support cancer growth. Additionally, we examine the emerging role of nutrient stress and metabolic sensing pathways in shaping NK cell suppression in PDAC. By modulating both redox and nutrient stress responses mechanisms, supplement therapy may help overcome immune evasion and drug resistance in PDAC, offering a promising direction for better prevention and treatment.
Keywords:
pancreatic cancer
; NK cell
; reactive species stress
; nutrient sensing
; antioxidant
; micronutrient
; triple NK cell approach
; treatment resistance
1. Introduction
Pancreatic ductal adenocarcinoma (PDAC) remains one of the most lethal malignancies, largely due to its aggressive nature, high risk of distant metastasis, and resistance to current therapies [1]. Even after surgical resection, most patients experience disease progression [2]. Chemotherapy is currently the most appropriate treatment option for metastatic PDAC, but its effectiveness is limited, and nearly all patients experience treatment failure during or shortly after [3].
The etiology of treatment resistance in PDAC is multifactorial, involving tumor environment (both the tumor microenvironment (TME) and macroenvironment) [4]. Therefore, overcoming resistance requires a multifaceted strategy that targets both these domains.
One major cause of therapy resistance is reactive species stress, which happens when the balance of harmful molecules—like reactive oxygen (ROS), nitrogen (RNS), and sulfur species (RSS)—is disrupted [5]. These reactive species are highly bioactive molecules. Despite their differences, oxidative, nitrosative, and sulfur-based stresses all lead to disruptions in redox homeostasis, impacting key cellular functions. They regulate redox signaling and can also induce oxidative stress by damaging DNA, proteins, and lipids. [6,7,8]. ROS contribute directly to oxidative stress and have been implicated in promoting tumor progression, immune suppression, and chemoresistance in PDAC [6]. RNS, such as nitric oxide (NO), can interfere with cellular signaling pathways and can lead to both direct nitrative damage—such as protein and DNA nitration—and indirectly contribute to oxidative damage by interacting with reactive oxygen species [7]. Meanwhile, RSS, including hydrogen sulfide (H2S), play complex roles in redox regulation and have gained attention as emerging therapeutic targets due to their involvement in maintaining cellular redox balance [8]. Recent research suggests that restoring redox balance through antioxidants and specific dietary supplements may help counteract the effects of ROS, RNS, and RSS [9]. Interestingly, this redox modulation may restore NK cell function, enhance their cancer-killing ability, and improve treatment outcomes [10].
Another important factor of treatment resistance is nutrient stress, where key pathways like mechanistic target of rapamycin (mTOR) and AMP-activated protein kinase (AMPK) become unbalanced, helping cancer cells survive and avoid the immune system. The mTOR pathway as a nutrient sensor is a critical regulator of cell growth, metabolism, and survival, playing a key role in the development and progression of solid tumors, including PDAC [11]. Despite its significance, mTOR inhibition as a monotherapy has shown limited success in clinical trials for PDAC due to the tumor’s ability to adapt and develop resistance mechanisms [12]. However, emerging preclinical data suggest that combining mTOR inhibitors with micronutrients could offer a promising therapeutic strategy [13]. We hypothesized that this combination approach may enhance immune responses and improve the effectiveness of mTOR-targeted therapies, potentially overcoming the limitations of monotherapy in PDAC treatment.
These findings support the development of a “triple NK boost” strategy [14]—enhancing NK cell proliferation, cytotoxicity, and tumor infiltration—by targeting nutrient and redox stress through supplement therapy. This approach offers a novel adjunct to help overcome immune suppression and therapy resistance in PDAC.
2. Reactive Species Stress and PDAC
In PDAC, ROS, RNS, and RSS contribute to cancer cell survival, metastasis, immune evasion, and therapy resistance [6,7,8,9]. ROS promotes tumor progression by activating oncogenic signaling pathways, while RNS, particularly nitric oxide, influence angiogenesis and immune modulation [6,7]. RSS, such as hydrogen sulfide, are involved in regulating the immunity of TME and can have both pro-tumor and anti-tumor effects depending on their levels [10]. Understanding how these reactive species interact in the context of PDAC may lead to novel therapeutic strategies aimed at restoring redox balance and improving treatment efficacy.
2.1. ROS are produced as by-products of cellular metabolism, particularly in cancer cells where oxidative stress is elevated [5]. In PDAC, ROS contribute to the activation of signaling pathways that promote tumor cell survival, proliferation, and metastasis. Elevated ROS levels can activate pathways like NF-kB and HIF-1α, which are involved in cancer cell growth, angiogenesis, and resistance to cell death (apoptosis)[6].
While ROS can promote immune activation under normal conditions, excessive ROS in the PDAC TME can suppress immune responses [6]. PDAC is very hard to treat with chemotherapy or radiation, and ROS may play an important role in this resistance. Chemotherapeutic agents can increase ROS production, but tumor cells often adapt by upregulating antioxidant defenses, allowing them to survive and continue proliferating despite the oxidative damage caused by treatment [6].
2.2. RNS, especially NO, can influence PDAC progression by modifying proteins, lipids, and DNA, leading to nitrosative stress. Nitrosative stress can alter the function of tumor suppressors and activate oncogenic pathways. NO has been shown to promote angiogenesis (formation of new blood vessels) and metastasis in PDAC by activating endothelial NO synthase in tumor-associated vasculature [7]. NO produced by immune cells or tumor cells in the TME can have dual effects on the immune system [9].
Additionally, RNS can contribute to chemotherapy resistance in PDAC by inducing changes in the TME that promote survival signals and enhance resistance to apoptosis. NO can reduce the sensitivity of PDAC cells to treatments like gemcitabine [15].
2.3. RSS, such as H2S, are emerging as important players in the redox balance of cancer cells [10]. In PDAC, H2S is produced by enzymes such as cystathionine γ-lyase and cystathionine β-synthase [8]. These species influence cell proliferation, survival, and migration, and can also affect cancer cell metabolism. At low concentrations, H2S can promote cancer cell growth, but at higher concentrations, it can lead to cell death, indicating a dose-dependent effect [16].
RSS also regulates the TME by modulating inflammatory pathways. In PDAC, H2S can alter the immune response by promoting tumor-associated macrophages that support tumor growth and suppress immune activity [8]. Furthermore, RSS can interact with ROS and RNS to form a complex redox network, influencing the overall oxidative state of the tumor and impacting treatment responses [17].
3. Nutrient Sensing and PDAC
Nutrient sensing is how cells “sense” the levels of important nutrients like glucose, amino acids, fatty acids, and vitamins. Based on what they detect, cells adjust how they grow, use energy, or respond to stress. It helps them stay balanced and healthy, especially when conditions change [18]. Cancer cells in the tumor environment struggle to survive due to lack of nutrients and increased stress. To survive, they use adaptive mechanisms that take over normal nutrient sensing pathways. In PDAC, the enzyme wild-type IDH1 (wtIDH1) helps cells survive by supporting energy production and protecting against damage. A preclinical study shows that drugs made to block mutant IDH1 (like FDA-approved agent ivosidenib) can also block wtIDH1 when magnesium is low, a condition found in tumors. Blocking wtIDH1 under these conditions kills cancer cells and reduces tumor growth, offering a new potential treatment for cancers with wtIDH [19].
While enzymes like wtIDH1 support survival under nutrient stress by producing energy and protecting against damage, the core regulators of nutrient sensing in PDAC are mTOR and AMPK, which will be discussed below as critical targets for metabolic intervention.
3.1. mTOR in PDAC
mTOR is a crucial nutrient sensor that regulates many cellular processes, including metabolism, immune responses, survival, and growth. It functions through two complexes, mTORC1 and mTORC2, controlling processes like protein synthesis, lipid and nucleotide production, and autophagy. When mTOR becomes overactive due to mutations or alterations in its complexes, it can contribute to aging, neurological diseases, and cancer. Studies show that mTOR’s role in metabolism and immune regulation is influenced by long noncoding RNAs (lncRNAs). Targeting mTOR could potentially improve cancer treatment outcomes [20]. It is closely linked to insulin and IGF-1 signaling. Metformin can inhibit this pathway by lowering cellular energy levels, which activates AMPK and other mechanisms, leading to reduced cell growth and potentially promote cancer cell death [11].
Interestingly, the PI3K/AKT pathway, which activates mTOR, is often overactive in PDAC due to loss of PTEN, a natural blocker of this pathway [21].
Drugs like rapamycin and its versions (temsirolimus, everolimus, ridaforolimus) target mTORC1 but not mTORC2 directly. However, blocking only mTORC1 may cause feedback that reactivates cancer growth. New drugs that block both mTORC1 and mTORC2 are more effective in lab studies and have shown promise in clinical trials for other cancers [22].
3.2. AMPK in PDAC
AMPK is a key regulator of metabolism, activated when ATP levels drop. It helps cells balance energy use by promoting processes that generate ATP and inhibiting those that consume it. AMPK is found in many organisms, from plants to animals, and plays a critical role in growth, and autophagy as well. In mammals, it coordinates energy use in tissues such as the liver, muscle, and fat. Recent research has uncovered new ways AMPK influences cellular functions. Due to its beneficial effects in conditions like cancer, there is growing interest in developing AMPK activators for therapeutic use [23].
AMPK is activated by changes in energy levels and stress signals, and while it was initially thought to be a tumor suppressor, it’s now known that cancer cells use AMPK to adapt to stress. AMPK inhibitors are being explored as potential cancer therapies, with some experimental inhibitors showing promise in preclinical studies. A preclinical study from 2024 focuses on repurposing the PAK4 inhibitor PF-3758309 as an AMPK inhibitor for treating PDAC, showing effectiveness in early models and offering a potential new approach for this aggressive cancer [24].
4. NK Cells and PDAC
NK cells are effector innate lymphoid cells originating from progenitor cells in the bone marrow. In humans, NK cells constitute approximately 1% of all immune cells, 2% of the total lymphocyte population and 10% of the total number of peripheral lymphocytes [25].
NK cells are important for controlling tumor growth in PDAC, but their ability to fight the cancer is weakened by the TME. In PDAC, NK cells have reduced ability to kill cancer cells, which is linked to disease progression. The TME releases factors like TGF-β and IL-10 that suppress NK cell activity and prevent them from recognizing and attacking cancer cells. NK cells also struggle to move to the tumor site. However, studies show that NK cell function can be restored through treatments like ex vivo activation. While NK cells are crucial in the fight against PDAC, more research is needed to understand how they interact with the TME and improve treatments [26].
NK cells can detect stressed cells without needing antigen- specific priming or MHC-I expression, enabling them to target a wide range of tumor cells [27]. Their activity is not dependent on specific tumor antigens, making them effective against various cancers, including those with low mutation loads [28]. NK cells can destroy cancer cells directly or enhance T cell responses through cytokine and chemokine production [29].
Numerous studies have highlighted the role of NK cells in preventing the initiation, progression of cancers [30] and treatment resistance [31]. Simultaneously, other research has emphasized the role of supplement therapy in augmenting NK cell activity [32] and cancer management [33]. Therefore, we hypothesized that some supplements could affect PDAC by influencing NK cell function. Although NK cell infusions are generally safe and do not cause significant graft-versus-host disease [34], the use of supplements to enhance NK cell function appears to be even safer.
While numerous preclinical and clinical studies have demonstrated that antioxidant vitamins and minerals, whether used alone or in combination, can enhance the growth-inhibitory effects of X-irradiation and chemotherapy on tumor [35,36] some physicians worry about potential interference with the efficacy of cancer treatments [37]. Clinical trials have yet to validate or invalidate this hypothesis.
This review aims to explore the current understanding of how various vitamins and minerals impact NK cell activity focusing on mechanistic insight, especially their effect on targeting oxidative stress and nutrient sensing. We will explore three key mechanisms through which the imbalance of these nutrients affects NK cells: 1. reduced NK cell cytotoxicity, 2. decreased NK cell frequency, and 3. diminished NK cell tumor-TME infiltration. Additionally, we will discuss the potential role of micronutrients in preventing cancer and overcoming treatment resistance.
5. NK Cell and Oxidative Stress
ROS, RNS, and RSS can all impact the function of NK cells in different ways. While moderate levels of ROS, RNS, and RSS can help activate NK cells to fight cancer, high levels of these species cause oxidative, nitrosative, and sulfur-related stress, which can damage the NK cells and reduce their ability to perform essential tasks like recognizing and killing tumor cells. ROS can impair NK cell activation and cytotoxicity by damaging cell structures, RNS can alter NK cell receptors and weaken their response to tumors, and RSS can disrupt the redox balance and cellular signaling needed for NK cell function. In excess, these reactive species can lead to immune dysfunction and contribute to cancer progression by reducing the effectiveness of NK cells. Yamasaki et al. discusses the roles of ROS, RNS, and RSS in cellular signaling and their impact on immune responses, including those mediated by NK cells [6,9,10,38].
5.1. NK Cell and ROS
ROS can damage immune cells, including NK cells, and inhibit their ability to recognize and kill tumor cells. This contributes to immune evasion and tumor progression. Studies highlight the critical role of redox-regulating mechanisms in facilitating effective NK cell infiltration and persistence within solid tumors. The ability of NK cells to withstand oxidative stress, particularly through mTOR-dependent thioredoxin activation, underscores the importance of antioxidant capacity in enhancing their antitumor function. This redox resilience appears essential not only for NK cell survival but also for supporting the broader immune landscape within the TME. Therefore, boosting antioxidant systems in NK cells represents a promising strategy to improve tumor infiltration and optimize the clinical efficacy of adoptive NK cell therapies [13,39].
In NK cells, excessive ROS can impair their ability to proliferate, secrete cytokines, and perform cytotoxic functions like killing cancer cells. While moderate ROS levels can activate NK cells and help them respond to infections or tumors, chronic oxidative stress can lead to NK cell dysfunction, reducing their ability to recognize and eliminate cancer cells. High ROS levels can reduce the ability of NK cells to produce cytotoxic molecules (e.g., perforin and granzyme B) that are essential for killing target cells. This limits their effectiveness in eliminating tumor cells. Chia et al. explores the dual roles of ROS in redox signaling and their overproduction leading to cellular damage, which can affect immune cells like NK cells [40].
5.2. NK Cell and NO
Dual immune system affection of NO has been discussed. On one hand, NO can suppress the function of NK cells, macrophages, and other immune cells, weakening the anti-tumor immune response. On the other hand, NO can also activate immune responses in some contexts, making its role complex and context-dependent [9].
High levels of RNS can interfere with NK cell activity. RNS can alter the function of key enzymes involved in NK cell activation and cytotoxicity. RNS can modify the receptors on NK cells (such as the NKG2D receptor), potentially making NK cells less responsive to tumor cells or other pathogens. RNS can promote immune evasion by modifying the TME or directly impacting the signaling pathways that control NK cell function. This may reduce NK cell-mediated tumor surveillance [41]. Andreas et al. also examine how RNS such as NO, influence NK cell function [42].
5.3. NK Cell and RSS
At low to moderate concentrations, RSS may support NK cell activity by acting as signaling molecules that help activate NK cells and regulate immune responses. However, high levels of RSS can lead to toxicity and oxidative damage, which may impair NK cell function, reduce their ability to proliferate, and hinder their ability to fight cancer. RSS can interfere with cellular signaling by modifying sulfur-containing molecules like proteins, altering their activity, and potentially impairing NK cell activation and response. Yang et al. discuss the dual role of hydrogen sulfide as both a signaling molecule that can enhance NK cell function at low levels and its potential toxic effects at high concentrations [43].
6. NK Cell and Nutrient Sensing
NK cells in the TME are influenced by nutrient sensing and metabolism, which are shaped by the availability of nutrients and the cancer cells’ metabolism. Cancer cells’ high rates of glycolysis and lipid metabolism lead to the buildup of harmful byproducts and increased metabolites in the environment. This alters NK cell energy use and disrupts their function. However, the metabolic processes that affect NK cell responses are still not well understood [44]. Here, we focus on the roles of mTOR and AMPK pathways in regulating NK cell metabolism and response.
6.1. NK Cell and mTOR
NK cells are vital for both innate and adaptive immunity, and mTOR plays a crucial role in their development. A preclinical study examines how mTORC1 and mTORC2 affect NK cell development. Deleting Rptor (mTORC1) or Rictor (mTORC2) in mice results in distinct NK cell phenotype defects at different stages. In Rptor-deficient mice, the transition from CD27+CD11b− to CD27+CD11b+ NK cells are impaired, while in Rictor-deficient mice, the final maturation from CD27+CD11b+ to CD27−CD11b+ NK cells is disrupted. The reduced expression of T-bet in Rictor-deficient mice is linked to maturation defects due to impaired mTORC2-AktS473-FoxO1 signaling. This highlights the distinct roles of mTORC1 and mTORC2 in regulating NK cell development and phenotype [45].
The PI3K/AKT/mTOR pathway plays a major role in PDAC progression, and targeted therapies are promising. Many studies showed that in PDAC, NK cell activity is reduced, and inhibiting PI3K in NK cells weakens their ability to fight tumors [46].
6.2. NK cell and AMPK
AMPK is an enzyme that helps regulate the cell’s energy balance. It gets activated when energy levels are low, and it helps the cell conserve energy by promoting energy-producing processes and slowing down energy-consuming ones. AMPK is important for controlling metabolism, growth, and responding to stress [23,24].
Energy metabolism also plays a key role in shaping the TME. AMPK helps boost the anti-tumor function of NK cells, which are key immune cells that attack cancer. In the tumor environment, NK cells often lose their ability to kill tumors due to metabolic changes, like a lack of nutrients and buildup of harmful byproducts such as lactate. One major issue is the loss of a membrane component called sphingomyelin (SM), caused by disrupted serine metabolism, which prevents NK cells from recognizing tumor cells. AMPK can fix this by regulating serine metabolism and blocking SM loss. It also supports NK cell survival, movement into tumor tissues, and their ability to destroy cancer cells. By restoring NK cell function, AMPK may help stop tumors from escaping the immune system and improve the success of immunotherapy [20,23,24].
7. Micronutrient-NK Cell Interaction
Micronutrients, including vitamins and minerals, are essential in small amounts for maintaining proper bodily function. In this section, we explore how micronutrients interact with redox stress and nutrient sensing, and how they influence tumor progression and resistance—specifically through the triple suppression of NK cell functions: proliferation, cytotoxicity, and tumor infiltration [14]. This discussion also underscores the significant potential for future mechanistic studies to further elucidate these relationships.
7.1. Vitamins
Vitamins are essential micronutrients that the human body cannot synthesize in adequate quantities and therefore must be obtained through the diet. The fat-soluble vitamins (A, D, E, K) are crucial for many different processes including immunity. All other vitamins are water-soluble [47,48].
7.1.1. A Vitamin
The term vitamin A encompasses various groups of fat-soluble compounds, such as retinol, retinal, retinoic acid (RA), and carotenoids, which act as precursors to vitamin A [47]. Vitamin A, found in foods like liver, fish oils, and dairy products, is important for cell growth and differentiation. Its active form, RA, regulates processes like development and cell death [48].
Numerous studies suggest a correlation between vitamin A and cancer prevention [49]. Retinoids are being studied for their potential to prevent cancer by inhibiting tumor growth and promoting cell differentiation [48]. All-trans-retinoic acid (ATRA) is a vitamin A derivative used to regulate cell growth and differentiation. It is notably effective in treating acute promyelocytic leukemia and is being studied for its potential in other cancers [50], especially solid tumors in preclinical and clinical settings [51]. However, some studies indicate that high-dose beta-carotene supplements may be associated with an increased risk of lung cancer, especially in smokers [52]. Therefore, it is important to avoid excessive vitamin A intake and ensure that only a balanced amount of vitamin A is recommended. Notably, large randomized controlled trials such as the Alpha-Tocopherol Beta-Carotene (ATBC) Cancer Prevention Study and the Beta-Carotene and Retinol Efficacy Trial (CARET) have shown that high-dose supplementation of beta-carotene and retinyl palmitate increased the incidence of lung cancer and overall mortality among smokers and asbestos-exposed individuals [52a,52b]. These findings highlight the need for clinical caution when recommending vitamin A or its derivatives, particularly in high-risk populations. Supplementation beyond recommended dietary levels should be avoided unless clinically indicated and closely monitored.
In a mouse model of breast cancer, researchers utilized a combination of cryo-thermal therapy and ATRA to target myeloid-derived suppressor cells. This combination significantly improved long-term survival rates in mice through various mechanisms, including the enhancement of NK cells [53]. Many studies have demonstrated that saffron and natural carotenoids possess anti- tumor effects [54]. Oral beta-carotene, a plant-derived inactive form of vitamin A, has been shown to enhance NK cell cytotoxicity in human subjects [55]. According to two studies, the frequency of lymphoid cells with surface markers for NK cells was higher in peripheral blood mononuclear cells from individuals who received beta-carotene supplementation [56,57].
Vitamin A boosts NK cell cytotoxicity and IFN-γ production, while its deficiency lowers them. RA, the active form of vitamin A, enhances NK cell function by increasing RAE-1 expression. In cancer therapy, 13cRA raises lymphocyte and NK cell counts. Retinoic acid also plays a dual role in regulating NK cells in inflammatory diseases, showing promise for treatment [58].
Vitamin A also regulates ROS production in immune cells, including NK cells. By modulating the redox environment, vitamin A helps maintain NK cell function, supporting their ability to fight cancer and infections [59].
Vitamin A, through its effects on immune cells, can also help prevent nitrative stress by regulating NOS expression and activity, which controls the levels of RNS in tissues. Retinoic acid has been shown to enhance the production of NO in certain immune cells like macrophages, which can boost the cytotoxic activity of NK cells against tumors. However, excessive NO can lead to nitrosative stress, so vitamin A’s role in balancing NO production is crucial for maintaining immune cell function without inducing damage [60]. RA has been shown to modulate redox signaling pathways, which can influence the production of reactive sulfur species like H2S. RA may influence enzymes like cystathionine γ-lyase, which are involved in H2S production, thus affecting the overall sulfur-related redox balance in immune cells [61]. Vitamin A’s antioxidant properties may also help mitigate the potential toxicity of elevated levels of RSS in cells, preventing damage to immune cells, including NK cells [17,61].
In addition to redox stress, nutrient sensing pathways like mTOR can affect tumor progression including the function of cancer stem cells (CSCs). A preclinical study tested ATRA to force CSCs to become less harmful, especially when combined with mTOR inhibitors. ATRA reduced CSC traits effects like growth, and movement, that were stronger with mTOR blockers. ATRA worked through the ERK1/2 pathway, not mTOR. These findings suggest combining ATRA with mTOR inhibitors may improve cancer treatment [13].
Vitamin A is specifically studied in PDAC and a meta-analysis of 11 studies (2,705 cases) found that higher dietary vitamin A intake may reduce PDAC risk (RR = 0.839, 95% CI: 0.712–0.988) with low heterogeneity and no publication bias. The association was stronger in case-control studies and European populations [62].
RA plays a significant role in regulating PDAC progression [48]. RA impacts cancer cell growth and differentiation by interacting with retinoic acid receptors (RARs) and retinoid X receptors (RXRs). RAR-β is crucial for maintaining a non-malignant phenotype, and its loss is linked to tumor progression. RA also influences intracellular signaling pathways, such as protein kinase C, and reduces PDAC cell adhesion to the basement membrane, limiting metastasis. Additionally, RA affects cancer-associated fibroblasts (CAFs), inhibiting their activity and migration, further reducing tumor spread. Preclinical studies support the potential of RA in preventing recurrence and metastasis in PDAC [63]. Clinical trials have shown some promise, such as a phase I study with RA and HDAC inhibitors resulting in prolonged stable disease [64]. However, other studies have not shown significant improvements in response rates, and more research is needed to better understand the therapeutic potential of vitamin A in PDAC [65]. ATRA has also been repurposed in combination with gemcitabine-nab-paclitaxel chemotherapy in the treatment of patients with PDAC [66].
Vitamin A and its active derivatives, particularly retinoic acid, play a multifaceted role in cancer prevention and therapy by regulating cell differentiation, redox balance, and immune function. Evidence supports its potential to enhance natural killer cell activity and modulate TME, contributing to improved therapeutic outcomes. However, maintaining an optimal balance is essential, as excessive intake or inappropriate supplementation may pose health risks and counteract its benefits.
7.1.2. D Vitamin
Vitamin D is a fat-soluble micronutrient that includes ergocalciferol (D2) and cholecalciferol (D3), essential for bone health and calcium balance [67]. It is made in the skin through sunlight and found in foods like fish, egg yolks, and fortified dairy. Its active form, calcitriol, helps regulate calcium and phosphate levels, keeping bones strong and supporting cell functions. Research suggests that getting enough vitamin D may lower the risk of osteoporosis and some cancers by boosting immunity and controlling cell growth [68].
In a secondary analysis of a randomized trial involving 25,871 patients, vitamin D3 supplementation was found to effectively reduce the risk of advanced cancer in adults without a baseline cancer diagnosis, with the greatest benefit for those with a normal body mass index [69]. Meta-analyses reveal that while observational studies link low vitamin D levels to an increased risk of cancer, randomized trials indicate that vitamin D supplementation lowers overall cancer mortality without affecting cancer incidence [70]. This may suggest that vitamin D might help reduce resistance to cancer treatments or slow disease progression.
Vitamin D, particularly its active form 1,25-dihydroxyvitamin D (1,25D), helps regulate the immune system and may influence NK cell activity. However, its effects are controversial. Some studies show that 1,25D boosts NK cell cytotoxicity, increases certain receptors like NKp30, NKp44, NKG2D, and raises levels of markers involved in killing target cells. Other studies report the opposite—reduced NK cell activity, lower stimulatory receptor levels (like CD69), and decreased immune signals (like IFN-γ and TNFα). More research is needed to clarify these conflicting results [67,71]. A preclinical study confirmed that dietary vitamin D supplementation can enhance innate immunity by boosting NK cell cytotoxicity in normal-weight mice. The study attributed this effect to changes in the splenic NK cell population [72]. Another study demonstrated that vitamin D enhances the antibody-dependent cytotoxicity (ADCC) of NK cells [67]. And finally, Yu et al. highlighted the significance of balanced vitamin D for the proper function of NK cells [73].
1,25 D also plays a key role in reducing intracellular oxidative stress. It acts as a potent antioxidant, supporting mitochondrial function and protecting against oxidative damage. Adequate vitamin D levels help lower systemic inflammation and prevent DNA, protein, and lipid oxidation [74]. Ravid et al. showed that calcitriol, another active form of vitamin D, boosts the cancer-fighting effects of immune signals and certain drugs by increasing oxidative stress in tumor cells. This effect depends on ROS and may also help NK cells work more effectively against cancer [75]. In overactive or inflammatory conditions, too much IFN-γ and oxidative stress can damage NK cells and tissues. Vitamin D improves redox balance by reducing oxidative stress-related pathways like TLRs and IFN-γ signaling. This lowers ROS production, protecting NK cells from dysfunction. A healthier redox state enhances NK cell efficiency and cytotoxicity. Overall, vitamin D supports precise, less inflammatory immune responses [71,76].
In addition to its role in managing oxidative stress, 1,25 D has been shown to influence key cellular pathways involved in nutrient sensing and growth regulation. Interestingly, one 2021 study shows that 1,25 D affects the mTOR pathway in osteosarcoma cells by regulating DNA Damage-Inducible Transcript 4 (DDIT4/REDD1), an mTORC1 inhibitor. After treatment, DDIT4/REDD moves to the cytoplasm, which suggests 1,25 D helps control mTORC1 activity, influencing cell growth and metabolism. This highlights the potential of 1,25 D in regulating cancer growth through mTOR [77].
Another study suggests that vitamin D3 may promote autophagy and apoptosis in gastric cancer cells by activating the p53/AMPK/mTOR pathway, which helps inhibit cancer cell growth. These findings reveal that the interaction between mTOR and p53 may enhance tumor suppression, potentially by also activating NK cells, which play a key role in immune surveillance and targeting cancer cells for destruction [78].
Building on these findings, both preclinical and clinical studies have explored how vitamin D might help PDAC patients. Vitamin D and its analogues, like MART-10 and 1-α,25(OH)2D3, show potential by regulating key cellular processes such as the cell cycle, energy metabolism, and tumor progression [79]. Vitamin D activates the vitamin D receptor, leading to antiproliferation, promoting differentiation, and encouraging apoptosis. It induces cell cycle arrest through upregulation of cyclin-dependent kinase inhibitors (p21 and p27) and downregulation of cyclins, while also promoting AMPK activation, which further inhibits cancer cell growth [80]. Vitamin D also blocks the PI3K/Akt pathway, reducing cell migration and invasion, particularly by suppressing epithelial-to-mesenchymal transition (EMT) [78]. Preclinical studies have shown that vitamin D analogues effectively inhibit PDAC cell growth and metastasis, especially when combined with AMPK activation [80]. However, clinical trials have had mixed results, with some showing slight improvement in progression-free survival but no objective responses [81]. Epidemiological studies suggest that low vitamin D levels may be linked to a higher risk of PDAC, though the results are inconsistent. Genetic variants in vitamin D-related genes may also influence cancer risk, highlighting the need for more research. Despite encouraging preclinical findings, the clinical efficacy of vitamin D analogues in PDAC treatment remains unclear and require further studies before they can be widely used [58,79,80,81].
Vitamin D plays a vital role in maintaining immune balance, redox homeostasis, and cell growth regulation, contributing to both cancer prevention and therapy. Its ability to modulate NK cell activity, oxidative stress, and key pathways such as mTOR and PI3K/Akt underscores its therapeutic potential, particularly in pancreatic and other solid tumors. However, inconsistent clinical outcomes highlight the need for further research to clarify optimal dosing, mechanisms of action, and its true efficacy in cancer treatment.
7.1.3. E Vitamin
Vitamin E is another fat-soluble vitamin that comprises eight natural forms, including α, β, δ, and γ isoforms of both tocopherols and tocotrienols [82]. Alfa- Tocopherol has long been regarded as the primary form of Vitamin E in dietary supplements due to its higher presence in tissues and greater efficacy compared to other forms of Vitamin E. However, γ-Tocopherol has demonstrated a notable ability to neutralize reactive nitrogen species and inhibit cyclooxygenase, which results in a more potent anti-inflammatory and anti-cancer effect than α-Tocopherol [83].
In 1982, alpha-tocopheryl succinate (alpha-TS) was identified as the most effective vitamin E form for inducing cancer cell differentiation, inhibiting proliferation, and promoting apoptosis. Studies over the past two decades confirm its efficacy in cancer cells while sparing normal cells [84]. Based on existing literature, vitamin E—when consumed through a diet or supplements rich in γ- and δ-tocopherols—may have cancer-preventive properties. However, high-dose supplementation with α-tocopherol does not appear to offer the same benefits, though it does not diminish the effectiveness of cancer treatments [85].
Interestingly research involving mice and in vitro human NK cell systems has demonstrated that vitamin E can boost NK cell activity [86].
In mice, a daily dose of 100 mg of α-tocopherol greatly boosted NK cell activity. In lab studies, α-tocopherol increased NK cell tumor-fighting power at concentrations of 0.5, 1.0, and 2.0 mg/ml, while tocotrienol achieved similar results at ten times lower doses. The palmvitee blend, palm-oil-vitamin E concentrate, was most effective with 12 µg of α- tocopherol and 24 µg of tocotrienol [87]. A pilot study from 2007 indicated that vitamin E can enhance NK cell function in patients with colorectal cancer and suggested its potential as an adjuvant in cancer immunotherapy [88].
Vitamin E, comprising tocopherols and tocotrienols, acts as an antioxidant and supports anticancer responses by reducing ROS. Studies consistently show that vitamin E, particularly α-tocopherol, enhances NK cell activity. In aging mice, α-tocopherol restored NK cell cytotoxicity, and similar effects were observed in colorectal cancer patients, cattle, and AIDS-infected mice, where NK cell activity and IFN-γ levels improved. Despite these findings, the molecular mechanisms behind vitamin E’s impact on NK cells remain unclear, warranting further research to explore its role in cancer treatment and prevention [58].
Vitamin E is a powerful antioxidant that easily enters cells, neutralizing harmful molecules and may play a role in managing cancer progression [89]. Sandhu et al. showed that dietary vitamin E reduces mutation frequency in tumor cells, suggesting it helps protect against genetic damage. Vitamin E may work by acting as an antioxidant and controlling neutrophil infiltration. These results support vitamin E’s potential to reduce mutations caused by both spontaneous and NO-induced factors. More research is needed to understand its role in cancer prevention [90]. While excessive oxidative stress can lead to DNA damage and cancer development, many clinical randomized controlled studies have shown that high vitamin E can be harmful, even increasing all-cause mortality in some cases [91].
Notably, large randomized controlled trials such as the Selenium and Vitamin E Cancer Prevention Trial (SELECT) demonstrated that high-dose α-tocopherol supplementation (400 IU/day) increased the risk of prostate cancer in healthy men, without providing overall cancer-preventive benefits [92a]. These findings emphasize the need for clinical caution when recommending vitamin E supplementation. High-dose or long-term supplementation should be avoided unless clearly indicated, as it may increase mortality and specific cancer risks. Dietary intake through natural food sources remains the preferred and safest means of maintaining adequate vitamin E levels.
Vitamin E succinate (VES) activates autophagy in gastric cancer cells by inhibiting the mTOR pathway. It does this by activating AMPK, which reduces mTOR activity and its targets, P70S6K and 4EBP-1. This inhibition of mTOR is key to VES-induced autophagy, with AMPK playing an important role in regulating this process [92].
Vitamin E, particularly its tocotrienol isoforms, demonstrates promising antitumor effects in PDAC through several mechanisms, including inhibition of NF-κB activity, regulation of the cell cycle, apoptosis induction, and modulation of the Ras–Raf–MEK–ERK signaling pathway [93]. Among the tocotrienols, δ- and γ-tocotrienol show the most consistent inhibition of PDAC cell growth and survival in vitro and in vivo, with δ-tocotrienol being the most potent. Furthermore, it sensitizes the PDAC cell to gemcitabine [94]. These tocotrienols suppress NF-κB activity, induce cell cycle arrest by increasing the tumor suppressor protein p27Kip1, and promote apoptosis through mitochondrial signaling and caspase activation. Additionally, δ-tocotrienol has shown potential in enhancing the effects of gemcitabine, a chemotherapy drug. However, the effects of vitamin E succinate (VES) are less clear, with some studies indicating minimal impact on cell growth inhibition. Clinical trials are still in the early stages, with a phase I trial on δ-tocotrienol showing no significant toxicity at high doses. Despite promising preclinical findings, further clinical studies are necessary to establish the efficacy of vitamin E, particularly tocotrienols, as a viable adjunctive treatment for PDAC [48].
Vitamin E exhibits strong antioxidant, anti-inflammatory, and anticancer properties, primarily through its tocopherol and tocotrienol isoforms. Evidence suggests that it enhances NK cell cytotoxicity, reduces oxidative damage, and modulates key signaling pathways such as mTOR and NF-κB, contributing to tumor suppression and improved immune function. While tocotrienols, especially the δ and γ forms, show promising results in pancreatic and other cancers, inconsistencies in clinical outcomes and risks linked to high-dose supplementation highlight the need for further investigation.
7.1.4. Vitamin K (VK)
VK is the last fat-soluble vitamin in this review and is essential for blood clotting, bone health, and heart function. Blackberries, blueberries, green fruits like kiwi, leafy greens and cereal grains are rich in VK. It exists mainly as VK-1 (from leafy greens) and VK-2 (from animal products and fermented foods). VK-1 supports clotting, while VK-2 helps with calcium regulation in bones and arteries. VK-2 (especially its specific subtype MK-7) stays longer in the body and may be more effective for bone and cardiovascular health. Both forms are better absorbed with dietary fat [95].
Emerging research also suggests that VK, particularly VK-2, may have potential anti-cancer effects by influencing cell growth and apoptosis. VK exhibits potential anticancer effects in various cancers, including PDAC and prostate cancers, by leveraging its anti-inflammatory and antioxidant properties to prevent cellular senescence and inhibit metastasis, thus improving cancer prognosis [96]. It also suppresses cancer cell proliferation, growth, and differentiation through mechanisms involving c-myc and c-fos gene induction, regulation of Bcl-2 and p21 genes, and angiogenesis inhibition. This highlights VK’s potential as an adjuvant therapy, either alone or in combination with traditional chemotherapy or other vitamins, for enhanced cancer treatment outcomes [95,96].
Cellular senescence protects against cancer in younger people but can help tumors grow in older individuals through the senescence-associated secretory phenotype (SASP). NK cells help remove cancerous and senescent cells, but their function declines with age. Vitamin K might help maintain NK cell activity and improve immune function. By supporting NK cells, vitamin K could reduce the effects of cellular senescence on cancer growth. This could help strengthen the immune system in older people, potentially lowering cancer risk [97]. Textor et al. showed that reducing c-Myc or N-Myc lowers B7-H6 levels on tumor cells like melanoma, PDAC, and neuroblastoma. This weakens NKp30, an important receptor that activates NK cells. VK deficiency can also reduce Myc levels, leading to weaker NK cell responses. So, VK may help support NK cell activity by keeping Myc and B7-H6 levels up [98].
Additionally, NK cell activity depends on genes like Bcl-2, c-fos, and p21. Bcl-2 keeps NK cells alive, c-fos helps activate immune responses, and p21 controls their growth. VK can boost the expression of these genes by supporting key signaling pathways. So, enough VK may help NK cells stay active, survive longer, and function better [99,100,101].
One study found that VK-2 triggers apoptosis in bladder cancer cells by affecting the mitochondria and activating certain pathways (JNK and p38 MAPK). Inhibiting these pathways stopped the cell death process. Furthermore, ROS played a role in this process, and blocking ROS with an antioxidant, prevented the apoptosis [102,103].
VK-2 triggers autophagic cell death [99,100,101] in bladder cancer cells through AMPK activation during metabolic stress [104].
VK has shown potential as an adjunctive agent in PDAC treatment through mechanisms primarily involving apoptosis, Ras–Raf–MEK–ERK, and JNK MAPK signaling pathways which all affect NK cell activities. VK-1, VK-2, and VK-3 induce apoptosis in PDAC cells via caspase-dependent pathways, involving caspase-3 activation, and through interactions with wild-type p53, ROS, and intracellular calcium. VK also influences the MEK-ERK pathway, where high concentrations of VK-1 and sorafenib lead to apoptosis via inhibition of phospho-MEK and phospho-ERK, while low concentrations induce apoptosis through the extrinsic pathway [105,106]. Additionally, VK-1 and sorafenib together activate the JNK/c-Jun pathway, upregulating FasL-mediated apoptosis (combining VK-1 with sorafenib in PANC-1, PL-5, and MIA PaCa-2 cells enhanced apoptosis by inhibiting phospho-MEK/ERK and activating the JNK/c-Jun–FasL extrinsic death pathway). Menadione (VK-3) quickly turns on caspase-3, cuts PARP, and activates ERK and JNK in PDAC and acinar cells; adding L-cysteine blocks ERK but not JNK, showing that ROS drive the ERK response. This matters for NK cells because they also use ROS-dependent ERK activation to release their killing granules. VK-3 can help weaken cancer cells and make them easier for NK cells to kill.
But too much ROS could also suppress NK cells directly—more research is needed to understand that balance. VK-3 is synthetic and generates ROS, but it can be toxic to the liver and is not approved for human use in most countries [106,107,108].
Although preclinical studies support these findings, there are no clinical trials evaluating VK’s role in PDAC, and further studies are needed before VK can be recommended for clinical use in treatment [48,106,107].
Vitamin K, particularly VK-2, demonstrates promising anticancer properties through its ability to regulate apoptosis, oxidative stress, and key signaling pathways such as Ras–Raf–MEK–ERK and JNK MAPK. It may enhance NK cell survival and cytotoxic function by modulating genes like Bcl-2, c-fos, and p21, as well as maintaining Myc-dependent activation signals. While preclinical studies suggest potential benefits for cancer prevention and PDAC therapy, clinical validation is still lacking, underscoring the need for further investigation into its safety, mechanisms, and therapeutic applications.
7.1.5. B Vitamin
B vitamins, a group of eight water-soluble vitamins, are essential for all living cells, from bacteria to humans. They play crucial roles in energy production, DNA synthesis, brain function, and maintaining healthy skin and blood cells. Other B vitamins include B1 (thiamin), which helps convert carbohydrates into energy and is found in whole grains, pork, and legumes; B2 (riboflavin), important for energy metabolism and antioxidant function, found in dairy products, eggs, and leafy greens; B3 (niacin), involved in DNA repair and metabolism, found in meat, poultry, fish, and peanuts; B5 (pantothenic acid), which supports hormone and neurotransmitter production, found in avocados, mushrooms, and whole grains; and B7 (biotin), essential for fat and carbohydrate metabolism, commonly found in eggs, nuts, and seeds [108,109].
Recent evidence highlights the role of B vitamins in regulating immune cells, which may explain the inconsistent findings of studies investigating the link between B vitamins and cancer development [110,111]. A meta-analysis study found that low intake of folate, the natural form of vitamin B9 (folic acid or folate), combined with methylenetetrahydrofolate reductase (MTHFR C677T) polymorphisms, is linked to a higher risk of breast cancer [112].
Sawaengsri et al. demonstrated that high folic acid intake reduces NK cell cytotoxicity in aged mice [113]. A clinical study suggested vitamin B12 (cobalamin) as an immunomodulator for cellular immunity and confirmed its crucial role for NK cell cytotoxicity [114]. In aged rats, B12 deficiency led to reduced NK cells activity and a decrease in certain B lymphocyte subsets. However, supplementing with B12 can encourage NK cell expansion, but excessive unmetabolized folic acid might impair NK cell activity [109,111,115]. Therefore, it’s important to emphasize that only a balanced intake of B vitamins is both safe and beneficial for preventing cancer progression and treatment resistance [116]. B3 increases CD62L expression, aiding NK cell recruitment, while B6 deficiency does not affect NK cell activity despite reducing T cell cytotoxicity. B7 deficiency lowers NK cell cytotoxicity in Crohn’s disease patients, and B9 deficiency correlates with reduced NK cell toxicity, though high doses of folic acid show no direct effect. Vitamin B12 improves NK cell activity in anemia patients and prevents NK cell decline in aged rats. Despite these findings, the mechanisms behind these interactions remain unclear and need further research [58].
B vitamins can influence cancer by helping protect DNA and reducing harmful oxidative stress. Folate and B12 supplements have been shown to lower ROS levels and prevent DNA damage in animal cancer models, whereas low folate can let ROS accumulate and promote tumor growth. B1 and its more bioavailable form, benfotiamine, reduce oxidative stress and NF-κB–driven inflammation, slowing down tumor progression in preclinical studies. Clinically, higher blood B1 levels have been linked to an increased risk of breast cancer, while higher B3 levels are associated with a lower risk [115,116,117].
Ayuk and Abrahamse (2019) explored how mTOR signaling affects cancer and the use of photodynamic therapy (PDT) in treatment. PDT uses a photosensitizer activated by light to produce ROS that kill cancer cells. Its effectiveness depends on factors like oxygen, light, and the type of photosensitizer, such as vitamin B12. PDT can block the mTOR pathway and combining it with mTOR inhibitors can improve cancer treatment. B vitamins like B12 may also help enhance PDT by influencing the mTOR pathway [118].
Nutrient competition in the TME affects tumor growth and immune response. High vitamin B6 (pyridoxine) demand by PDAC cells impairs NK cell function. Supplementing vitamin B6 while inhibiting its metabolism boosts NK cell activity and reduces tumor growth in PDAC models. Two clinical trials have directly tested the addition of a B-vitamin derivative—folinic acid (leucovorin)—to chemotherapy in PDAC. In gemcitabine-refractory advanced PDAC (JapicCTI-111554), adding oral leucovorin (25 mg twice daily) to S-1 significantly prolonged median PFS (3.9 vs. 2.8 months) and increased the disease control rate compared with S-1 alone. In a multicenter Phase II trial of FOLFIRINOX (5-FU, leucovorin, irinotecan, oxaliplatin) involving 46 patients with advanced disease, the regimen achieved a 31% objective response rate, median PFS of 6.4 months, and median OS of 11.1 months, albeit with higher grade 3–4 neutropenia [119,120,121].
B vitamins play vital roles in energy metabolism, DNA repair, and immune regulation, with growing evidence linking their balance to cancer prevention and treatment outcomes. They influence NK cell activity, redox balance, and pathways like mTOR, highlighting their potential in enhancing immune responses and improving therapeutic efficacy. However, both deficiencies and excessive intake can disrupt immune function or promote tumor growth, emphasizing the importance of maintaining optimal B vitamin levels for effective cancer management.
7.1.6. C vitamin
Vitamin C (ascorbic acid), another member of the water-soluble vitamin group, plays a vital role in collagen synthesis, immune function, iron absorption, and antioxidant protection against free radicals. Unlike most animals, humans cannot produce Vitamin C endogenously, making dietary intake essential. It is abundant in fresh fruits and vegetables, particularly citrus fruits like oranges, lemons, and grapefruits, as well as kiwi, strawberries, bell peppers, broccoli, Brussels sprouts, and tomatoes. Regular intake of Vitamin C is important to support tissue repair, wound healing, and overall cellular health [122].
Vitamin C plays a significant role in cancer prevention, progression, and treatment, both for its general health benefits and its ability to act as a pro-oxidant at high doses [123]. Additionally, ascorbate has been reported to act as an adjuvant, enhancing the anti-tumor effects of cancer treatments like radiotherapy, radio-chemotherapy, chemotherapy, immunotherapy, or even monotherapy. It promotes tumor cell death by generating ROS and inducing ferroptosis [122,123,124].
Vitamin C enhances NK cell activity and supports immune function. Ascorbic acid stimulates the growth of NK cell populations (proliferation) in cultures, making it beneficial for NK cell therapy [125,126,127].
A preclinical study demonstrated that low-dose Vitamin C promotes demethyla- tion of the Killer Immunoglobulin-like Receptor (KIR) promoter and enhances KIR expression, indicative of NK cell maturation [128]. High doses of Vitamin C have been shown to increase the secretion of perforin and granzyme B by activated NK cells in a mouse model [127].
Vitamin C enhances cytokine expression by promoting the activity of Ten-Eleven Translocation (TET) enzymes, which modify DNA methylation and impact gene expression related to immune responses, including modulating NK cell function. This leads to the activation of the IFN-γ/JAK2/STAT1 signaling pathway, which increases tumor-infiltrating lymphocytes (TILs) and improves the efficacy of immuno-checkpoint therapy [125]. Since 1980, the influence of vitamin C on NK cell activity has been investigated [128]. Although the results are controversial, it can be concluded that normalizing vitamin C levels favors NK cell activity, while high doses can decrease this effect [32].Early studies showed that AA (ascorbic acid) inhibited NK cell activity in a dose- and time-dependent way but did not affect activation by IFN-γ, IL-2, or effector-target binding. Later research found that oral AA increased NK cell activity, likely through protein kinase C (PKC) activation. Recent studies confirmed AA’s role in boosting NK cell cytotoxicity, aiding NK cell maturation, and enhancing activity during H1N1 infection with red ginseng. Further research is needed to fully understand its potential [58].
Vitamin C, particularly in high intravenous doses, has been explored in treating PDAC. At pharmacologic concentrations, it creates ROS that selectively target and kill cancer cells while sparing normal tissue. This effect is due to the formation of hydrogen peroxide, which damages tumor cells. Additionally, vitamin C depletes ATP levels in cancer cells by interfering with DNA repair and glycolysis, leading to cell death. Vitamin C also induces autophagy, a process that contributes to tumor cell destruction. At high doses, like in IV treatments, it can create hydrogen peroxide that kills cancer cells without harming normal ones. This makes vitamin C a promising option in cancer therapy that targets oxidative stress. [48,124,129,130,131].
A large retrospective cohort study in the U.S. found that individuals without a history of vitamin C prescriptions had a higher incidence of PDAC across all age groups and regions. Specifically, the incidence was 0.88% in those without vitamin C prescriptions compared to 0.48% in those with such prescriptions, indicating a statistically significant difference [132].
Another study shows that pharmacological vitamin C (VC) blocks the mTOR pathway by increasing ROS, leading to the degradation of Rictor through FBXW7. HMOX1 is crucial for this process. Removing FBXW7 or HMOX1 stops VC from affecting mTOR, cell growth, and autophagy. In vivo, VC reduces tumor growth by inhibiting mTOR, but HMOX1 deficiency can reverse this effect. These results suggest that vitamin C could be a potential cancer treatment by targeting the mTOR pathway [130].
Preclinical studies have demonstrated that intravenous vitamin C decreases PDAC cell survival, inhibits tumor growth, and enhances the efficacy of chemotherapy. Clinical trials have shown promising results when vitamin C is combined with chemotherapy drugs like gemcitabine, with some patients showing tumor size reduction and improved performance status. A meta-analysis of 17 studies with 4,827 PDAC cases found that higher vitamin C intake was linked to a lower risk of developing PDAC, with a relative risk of 0.705, indicating a protective effect [48,129,131,132,133].A review looking at 23 clinical trials examined the use of high-dose intravenous vitamin C (IVC ≥15 g) in cancer treatment. It found that IVC, when combined with chemotherapy or radiotherapy, offered the greatest improvements in quality of life and added anti-tumor effects, compared to using it alone. Promising results were seen in patients with advanced PDAC, breast, and ovarian cancers. While IVC is not curative, it may enhance supportive care, and further studies are needed to assess its impact on survival [134]. Several active clinical trials are ongoing to further explore the role of vitamin C as an adjunctive treatment for PDAC, particularly in combination with chemotherapy and radiation. Although current results are promising, larger trials are needed to fully confirm the effectiveness of vitamin C in this context. A randomized trial hypothesized that adding high-dose intravenous vitamin C to gemcitabine and nab-paclitaxel would increase survival in patients with metastatic PDAC [135]. A phase I/IIa study tested high-dose IVC combined with gemcitabine for PDAC. IVC kills cancer cells by depleting energy and affecting cell movement and metastasis. It also helped reduce cancer spread and was safe for patients. IVC did not interfere with gemcitabine, showing potential to extend survival with low toxicity, suggesting it could be a helpful treatment for advanced PDAC [136].
Vitamin C serves as a potent antioxidant and immunomodulator, supporting NK cell proliferation, cytotoxicity, and cytokine production while influencing key signaling pathways such as IFN-γ/JAK2/STAT1 and mTOR. At pharmacological doses, it acts as a selective pro-oxidant, generating ROS that promote cancer cell death and enhance the efficacy of chemotherapy and immunotherapy. Although clinical and preclinical findings show promising outcomes—particularly in pancreatic cancer—further large-scale studies are needed to establish optimal dosing, safety, and long-term therapeutic benefits.
7.2. Minerals
Minerals are inorganic elements required by the body in small amounts to perform a wide range of physiological functions, including enzyme activity, bone health, nerve signaling, and immune support [137]. Mineral therapy, which involves the use of essential minerals, has shown potential as an adjuvant in cancer therapy due to its effects on immune function, including the activity of NK cells [138].
7.2.1. Calcium
Calcium is an essential mineral, with 99% stored in bones and teeth, and the remaining 1% circulating in the blood as calcium ions (Ca2+). These ions are crucial for muscle function, nerve signaling, and blood clotting. Proper calcium regulation is key for overall health [139].
Emerging research suggests that calcium, particularly its ion form, may play a role in cancer prevention by influencing cellular signaling, apoptosis (programmed cell death), and tumor growth regulation. Intracellular Ca2+ levels affect both cancer cell growth and death. Researchers developed pH-sensitive sodium hyaluronate-modified calcium peroxide nanoparticles (SH-CaO2 NPs) that induce calcium overload specifically in tumors, effectively killing cancer cells while sparing normal cells and aiding in tumor imaging and control [140]. Thus, calcium overload in this method does not interfere with cancer treatment.
High Ca2+ is also essential for NK cells to effectively target and kill cancer cells [141]. NK cells deficient in STIM1/ORAI1 activity show severely impaired calcium entry and are unable to kill target cells, highlighting the importance of this complex for NK cell function [142]. Calcium overload can damage and kill cells.
Calcium signaling intersects with ROS in cancer, forming a complex network that influences tumor progression and survival. Intracellular Ca2+ fluxes regulate oxidative metabolism and ROS production, while oxidative stress can alter calcium homeostasis, leading to tumor-promoting or tumor-suppressive outcomes depending on the cellular context. This bidirectional interplay is crucial in various cellular processes, including apoptosis and proliferation. [143,144,145].
Changes in calcium levels in ovarian cancer cells are linked to the mTORC1 signaling pathway and fatty acid production. TRPV4 regulates this process through the calcium-mTOR/SREBP1 pathway, which helps promote ovarian cancer progression [146].
Ca2+ play a key role in the growth and spread of PDAC by affecting cell signaling. Problems with calcium signaling are linked to tumor growth and resistance to treatment in PDAC. Research shows that targeting calcium pathways, like using calcium channel blockers, might make treatments like gemcitabine more effective. This suggests calcium could be a useful target for improving cancer treatment [147,148,149]. A meta-analysis of observational studies examined the association between calcium intake and PDAC risk. The analysis included eight studies and found an inverse association between total calcium intake (dietary and supplement) and PDAC risk (relative risk = 0.83, 95% CI: 0.72–0.97), suggesting that higher calcium intake may reduce the risk of developing PDAC [150]. Additionally, a Phase II randomized study investigated the combination of BAX2398 with 5-fluorouracil and calcium levofolinate in Japanese patients with metastatic PDAC that had progressed after prior gemcitabine-based therapy. This trial aimed to assess the efficacy of incorporating calcium levofolinate into the treatment regimen to enhance therapeutic outcomes (NCT02697058).
Calcium plays a multifaceted role in both cellular homeostasis and cancer regulation, influencing signaling pathways, apoptosis, and tumor progression. It is essential for NK cell cytotoxicity and interacts closely with ROS and mTOR signaling, affecting both cancer cell survival and immune responses. Evidence from preclinical and clinical studies suggests that maintaining adequate calcium levels may lower pancreatic cancer risk and improve treatment efficacy, though further research is needed to refine its therapeutic applications and clarify its complex mechanisms
7.2.2. Iron
Iron is another critical element for the body that helps carry oxygen in the blood and supports energy production, immune function, and overall health. It is a key part of hemoglobin, which transports oxygen to the body’s tissues. Iron is found in foods like red meat, poultry, beans, spinach, and fortified cereals. The body stores it in the liver and spleen. Having the right amount of iron is important, as too little can cause anemia, while too much can harm organs [151].
Its imbalances affect tumor development and treatment. Recent research highlights iron metabolism disorders in tumors, its role in ferroptosis and ferritinophagy, and new iron-based cancer therapies [152].
Additionally, iron can affect the proliferation and activity of immune cells. In a preclinical study, ferumoxytol, an iron replacement product, was combined with NK cells, inducing ferroptosis in cancer cells and enhancing NK cell cytotoxicity [152]. Both iron deficiency and overload can negatively impact NK cell function and overall immune response [153].
Several studies explore the dual role of iron and ROS in cancer cells. While they are essential for cellular functions, their imbalance can contribute to carcinogenesis. The therapeutic strategies targeting iron metabolism and redox regulation in cancer treatment are also discussed in many papers. Furthermore, the mechanisms of iron homeostasis and its regulation of ROS in tumors are explored significantly. The controversial roles of excess iron and iron deficiency in tumor progression and options for iron-related treatments are mentioned [154,155].
Iron chelators inhibit mTORC1 signaling by activating the AMPK pathway and partially involving HIF-1/REDD1 and Bnip3 pathways. This effect is not related to ROS, copper chelation, or PP2A activation. Iron is necessary for mTORC1 activation, and its chelation disrupts this process through multiple mechanisms [156].
Interestingly, iron plays an important role in PDAC, affecting both the development of the disease and patient health. Some studies suggest higher iron levels may increase the risk of developing PDAC, although results are mixed. Patients with PDAC often experience anemia and iron deficiency, which can make treatment more difficult. Cancer cells also need more iron to grow, so regulating iron levels could be a potential treatment strategy. Excess iron has been linked to increased cancer cell spread and resistance to treatment. An article discusses how ferritinophagy, a process that releases iron from ferritin, supports PDA C cell survival and resistance to therapy. Another study suggests that iron chelation could reduce metastasis by inhibiting epithelial-mesenchymal transition (EMT) and regulating NDRG1. EMT plays a key role in cancer cell migration and treatment resistance, allowing cells to evade therapies. By managing iron levels and influencing EMT, iron chelation may decrease cancer spread and improve treatment effectiveness, offering a potential strategy for overcoming resistance in cancers like PDAC. These findings highlight the complex role of iron in PDAC and the importance of managing iron levels in patients [157,158,159,160].
Iron is essential for oxygen transport, energy metabolism, and immune function, but its imbalance can strongly influence cancer development and progression. It regulates key processes such as ferroptosis, ROS production, and mTOR signaling, affecting both tumor growth and NK cell cytotoxicity. While iron overload can promote metastasis and treatment resistance, controlled modulation of iron—through chelation or metabolic targeting—shows promise in reducing pancreatic cancer progression and improving therapeutic outcomes.
7.2.3. Magnesium
Magnesium is an important mineral that helps with energy production, muscle function, and nerve signaling. It also supports healthy bones, blood sugar levels, and a normal heart rhythm. Magnesium can be found in foods like leafy greens, nuts, seeds, whole grains, and legumes. Many people don’t get enough magnesium, which can lead to problems like muscle cramps and fatigue. Keeping magnesium levels balanced is important for good health [161].
Magnesium deficiency is linked to a higher risk of certain cancer. A case-control study indicated that higher magnesium consumption was linked to a reduced risk of breast cancer [162] but understanding its role in cancer development is difficult due to factors like genotype, nutrient interactions, and inflammation [161]. Hypomagnesemia must be addressed in cancer management. However, managing hypermagnesemia is also crucial for patients at high risk. Therefore, monitoring magnesium levels during supplement therapy is essential to avoid adverse effects on cancer treatment.
Magnesium, involved in various biochemical reactions in the body, is also important for maintaining immune system balance. A deficiency in magnesium may reduce NK cell effectiveness [161].
Research shows that low magnesium levels can reduce the activity of NK cells, which are responsible for attacking infected or cancerous cells. Specifically, a lack of magnesium can affect the NKG2D receptor on NK cells, which is crucial for their activation. Studies have shown that supplementing magnesium can restore NK cell activity and improve immune responses. This highlights the importance of keeping magnesium levels balanced for proper immune function [163].
A study found that higher levels of magnesium inside cells help protect mitochondria from damage caused by ROS, like hydrogen peroxide. This shows that magnesium can help reduce oxidative stress. Research on biodegradable magnesium materials showed they can reduce harmful levels of ROS and RNS in immune cells. This suggests magnesium may help control inflammation and protect cells [164,165].
Mg2+ promotes tumor cell death by activating the AKT/mTOR and Bax signaling pathways. These pathways contribute to apoptosis, helping to control tumor growth [166].
A preclinical study found that a magnesium transporter protein called SLC41A1 was lower in PDAC tissues and cells. When researchers increased this protein in cancer cells and tested them in mice, the tumors grew much smaller. This suggests that magnesium transport may help slow down PDAC growth and could be a useful target for future treatments [166]. Additionally, research has shown that magnesium intake may be linked to a lower risk of PDAC. In a study of more than 66,000 men and women aged 50 to 76, it was found that for every 100-milligram decrease in daily magnesium consumption, the risk of developing PDAC increased by 24%. Furthermore, those whose magnesium intake was less than 75% of the recommended daily allowance (RDA) had a 76% higher risk compared to individuals meeting the RDA. These results suggest that maintaining adequate magnesium levels might help in preventing PDAC. However, it’s important to note that more research is needed to fully understand the connection between magnesium and PDAC risk [167]. A large prospective study examined dietary intake of calcium, magnesium, and phosphorus among participants. Findings indicated that higher total magnesium intake was associated with a reduced risk of PDAC. Specifically, participants in the highest quartile of magnesium intake had a hazard ratio (HR) of 0.61 (95% confidence interval [CI]: 0.37–1.00) compared to those in the lowest quartile, with a significant trend observed (p-trend = 0.023). These results suggest that increased magnesium consumption may be linked to a lower risk of developing PDAC [167]. Currently, there are no completed clinical trials specifically evaluating magnesium supplementation as a treatment for PDAC. However, a Phase 3 clinical trial (NCT01362582) investigated the effects of magnesium sulfate in patients with PDAC. This trial was terminated and did not yield conclusive results. In a prospective cohort study involving U.S. male health professionals, researchers investigated the relationship between magnesium intake and PDAC risk over 20 years. While no overall association was found between magnesium intake and PDAC risk, a significant inverse relationship was observed among overweight individuals (body mass index ≥25 kg/m2). In this subgroup, higher magnesium intake was associated with a reduced risk of PDAC (relative risk [RR] = 0.67; 95% CI: 0.46–0.99; p-trend = 0.04). This finding suggests that magnesium may play a protective role against PDAC in overweight populations [168].
Magnesium plays a vital role in maintaining cellular energy balance, immune regulation, and oxidative stress control. Adequate magnesium levels support NK cell activation through NKG2D receptor function and help protect mitochondria from ROS-induced damage, contributing to both immune efficiency and cancer prevention. Evidence suggests that sufficient magnesium intake may reduce the risk of pancreatic cancer and inhibit tumor growth through mTOR and apoptotic pathways, though more clinical research is needed to confirm its therapeutic potential and define optimal supplementation strategies.
7.2.4. Zinc, Copper, and Cadmium
Zinc, copper, and cadmium are three essential metals that play crucial roles in various biological processes. However, their concentrations must be tightly regulated to prevent toxicity. Metallothioneins (MTs) are small, cysteine-rich proteins that bind to these metals, aiding in their transport, storage, and detoxification. Zinc is vital for enzyme function, immune system regulation, and DNA synthesis. Copper is essential for electron transport, iron metabolism, and antioxidant defense. While cadmium is toxic in excess, MTs can isolate it, mitigating its harmful effects. By binding to these metals, MTs ensure their safe management within the body, maintaining a balance that is critical for health. Foods that are rich in zinc include meat, shellfish, legumes, and seeds, while copper can be found in foods like shellfish, nuts, seeds, and organ meats. Cadmium, although toxic, is found in small amounts in foods such as shellfish, liver, and certain cereals [169,170].
Elevated MT levels have been found in various cancers, suggesting they play a role in tumor growth. MTs help protect cancer cells by managing zinc and copper levels, reducing oxidative stress, and potentially contributing to drug resistance. However, while MTs can reduce the harmful effects of cadmium, long-term exposure to cadmium has been linked to an increased cancer risk. This highlights the complex relationship between these metals, MTs, and cancer development. It is now clear that the reactivity and coordination dynamics of MTs with Zn2+ and Cu+ align with the biological needs for regulating the binding and delivery of these metal ions within cells. A comprehensive review of expression of MT isoforms in various types of cancers highlights the application of MTs in cancer management [171,172,173,174].
Zinc is essential for the development and function of immune cells, including NK cells. Zinc deficiency can impair NKA and increase susceptibility to infections and cancer. Many preclinical studies have concluded that zinc deficiency leads to decreased NKA in rats [175,176,177,178]. Elevated MT expression has been linked to enhanced NK cell activity, indicating a role in cancer immune defense. However, some compounds that increase MT levels in NK cells can reduce zinc levels inside the cells, weakening their anti-tumor function. Additionally, MT overexpression in tumor cells is associated with stronger anti-cancer immune responses, including increased NK cell activity. These findings highlight the complex relationship between MTs and NK cells, suggesting that adjusting MT expression could influence NK cell activity in cancer therapy [179,180].
Zinc helps important antioxidant enzymes, like superoxide dismutase (SOD), that protect the body by neutralizing harmful molecules called ROS. When zinc is low, the body’s defenses weaken, leading to more oxidative stress, which can damage DNA and lead to cancer. Zinc also affects the levels of other reactive molecules like RNS and RSS, which are involved in inflammation and cell signaling processes that often go wrong in cancer. Cancer cells often have abnormal zinc levels, which can help them grow and resist treatment. Keeping zinc levels balanced may help protect against cancer and improve treatment results [181,182].
ZIP7 is a protein that controls zinc release from inside cells. This release activates key signaling pathways like MAPK, mTOR, and PI3K-AKT, which help cells survive and grow. These pathways are often overactive in cancer. Phosphorylation of specific sites on ZIP7 is essential for its activation, which was confirmed through mutagenesis studies [183].
Copper helps PDAC grow by activating the mTOR signaling pathway. A protein called CTR1, which brings copper into cells, is found at high levels in PDAC. When copper is reduced—either by blocking CTR1 or using a copper-removing drug—it slows down the mTOR pathway, which controls cancer cell growth and blood vessel formation. This makes the cancer grow more slowly. Combining copper reduction with an mTOR-blocking drug like rapamycin works even better, suggesting that targeting copper could help improve cancer treatment [184].
Another study shows that even low levels of cadmium can harm liver cells by causing fat buildup, scarring, and faster cancer cell growth. It does this by increasing inflammation and activating the Notch and AKT/mTOR pathways linked to liver disease and cancer [185].
Researchers found that levels of a specific MT isoform (MT1G) were lower in PDAC cells that resisted gemcitabine. When they increased MT1G, the cancer cells became less stem-like and responded better to treatment. MT1G worked by blocking a pathway (NF-κB/activin A) linked to cancer growth and drug resistance. This suggests MT1G may help slow cancer and make treatments more effective [186]. Another study looked at MT levels in 75 PDAC samples. Conversely, it found that tumors with high MT were more aggressive, had worse cell structure, and were more likely to spread. This means high MT expression in PDAC is linked to higher tumor grade and poor prognosis, indicating a more aggressive cancer. Additionally, MT expression is associated with metastasis and poorer histological grading in PDAC [180].
A study from 2022 looked at zinc levels in PDAC patients after surgery (pancreaticoduodenectomy). It found that 17.8% of patients had zinc deficiency even though they were given 50 mg of zinc daily. Patients with lower zinc levels after surgery had worse outcomes, suggesting that the usual zinc dose might not be enough and higher amounts could be needed during treatment [187]. Another study looked at how chemotherapy before surgery affects zinc levels in PDAC patients. It found that treatments, including gemcitabine, significantly lowered zinc levels. This suggests that checking and possibly supplementing zinc may be important during chemotherapy [188].
Interestingly taste changes (dysgeusia) can be linked to oxidative stress, particularly in cancer patients undergoing treatments like chemotherapy or radiotherapy. A 2024 clinical trial investigated zinc supplementation in patients with unresectable PDAC experiencing taste disorders (dysgeusia) during chemotherapy. The study found that administering zinc acetate hydrate significantly increased serum zinc levels and improved taste perception over a 12-week period. This suggests that zinc supplementation may help maintain nutritional status and quality of life in PDAC patients undergoing treatment [189].
Zinc, copper, and cadmium are essential metals regulated by metallothioneins (MTs), which control their storage, detoxification, and cellular effects. Zinc supports NK cell function, antioxidant defenses, and DNA protection, while copper can promote tumor growth via mTOR signaling, and cadmium is toxic even at low levels. Altered MT expression and metal imbalances influence cancer progression, treatment response, and patient outcomes, highlighting the importance of monitoring and managing these metals in cancer care.
7.2.5. Iodine
Iodine is another trace mineral needed for thyroid hormone production, which helps regulate metabolism and growth. The main sources of iodine are iodized salt, seafood (fish, shrimp, seaweed), dairy products, and some plant foods grown in iodine-rich soil. In areas with iodine deficiency, adding iodine to salt has been a successful way to prevent related health problems. Getting enough iodine is important for good thyroid function and overall health [190].
Iodine acts as an antioxidant and can prevent cancer, but excess intake may increase the risk of papillary thyroid cancer. Managing iodine deficiency is crucial for reducing cancer risk. Using alternative sources besides iodized salt and regularly monitoring urine iodine levels are recommended [191]. It is important to highlight that iodine toxicity can cause gastrointestinal and cognitive issues and may lead to thyroid papillary cancer. Therefore, regular monitoring of iodine levels is essential to avoid toxicity in cancer patients [192].
Iodine also boosts the innate immune system’s ability to combat both bacterial and viral infections [193]. Some studies suggest that high iodine levels can alter NK cell function, while other research shows that radioiodine therapy can boost NK cell activity against tumor cells, offering potential cancer treatment benefits. These findings highlight the complex relationship between iodine and NK cells, with implications for both immune health and cancer therapy [194,195,196].
Iodine helps protect the body from damage by harmful molecules like ROS, acting as an antioxidant. But in some conditions, it can also help create reactive iodine species, which fight germs but may also affect the body’s redox balance. So, iodine can both protect cells and play a role in creating reactive molecules, depending on the situation [197,198].
Too much iodine may contribute to papillary thyroid cancer (PTC) by activating the AKT/mTOR cell growth pathway. Moderate iodine levels increased PTC cell growth, while high levels slowed it down. Blocking this pathway reduced cell growth and movement, suggesting iodine plays a role in thyroid cancer development [199].
Iodine deficiency (ID) activates thyroid blood flow and VEGF expression through the mTOR pathway, with AMPK acting to limit this effect. In mice, rapamycin blocked ID effects, while metformin (which activates AMPK) reduced them. In mice lacking AMPK, ID effects lasted longer. mTOR is needed for early ID-induced changes, while AMPK helps control them [200].
Iodine is also being explored in PDAC diagnosis and treatment. Research shows that iodine uptake in PDAC can be measured using CT imaging, helping to understand tumor characteristics and treatment responses. Additionally, iodine-125 (^125I) seeds are being used in some treatments for PDAC, where they are implanted into the tumor to deliver targeted radiation, potentially reducing pain and slowing tumor growth. Some experimental treatments also use iodine-131 (^131I) combined with biogels to focus radiation directly on the tumor while protecting healthy tissue. These studies suggest that iodine could play an important role in both diagnosing and treating PDAC [201,202,203].
Iodine is essential for thyroid hormone production, metabolism, and growth, with sources including iodized salt, seafood, and dairy. It acts as an antioxidant, supports NK cell function, and may influence cancer risk, though excess iodine can contribute to papillary thyroid cancer via the AKT/mTOR pathway. Iodine is also being explored in PDAC for imaging and targeted radiotherapy using ^125I or ^131I, highlighting its potential in diagnosis and treatment.
7.2.6. Selenium
Selenium is an essential trace element important for human health due to its role in antioxidant defense, immune function, and thyroid hormone metabolism. It is found in a variety of foods such as Brazil nuts, seafood, meat, poultry, eggs, grains, and dairy, although the selenium content in plant-based foods depends on soil levels. Maintaining an adequate selenium status is important, as both deficiency and excess can lead to health issues. A recent review highlights that proper selenium intake is linked to reduced risk of chronic diseases and improved overall health outcomes [204].
Selenium is an essential micronutrient associated with cancer prevention, primarily due to its role in selenoproteins that protect against oxidative damage and DNA injury. Nonetheless, oxidative stress can both support and inhibit tumor growth. Selenoproteins also play roles in both tumor promotion and suppression [205]. Lobb et al. investigated how methylseleninic acid (MSA) impacts chemotherapy and radiation therapy on both cancerous and normal human blood cells. At high doses, MSA harmed normal cells and increased radiation damage, while boosting the effectiveness of cancer treatments on malignant cells [206].
Research shows that selenium supplements can boost NK cell function and help them kill cancer cells more effectively. Some forms of selenium, like selenium nanoparticles, have been shown to improve the immune system’s ability to attack tumors. However, both too little and too much selenium can reduce NK cell function, so it’s important to keep selenium levels balanced for the best immune support [207,208].
Selenium supplementation in mice significantly boosted IL-2 and IFN-γ production, underscoring the importance of adequate selenium intake for maintaining balanced immune responses and NKA [209,210].
Selenium helps protect cells by being part of enzymes that reduce harmful molecules like ROS and RNS. While it supports immune function and prevents damage at normal levels, too much selenium can produce ROS and harm cancer cells. Selenium nanoparticles are being researched for their potential to treat cancer [211,212].
Selenium helps reduce cancer risk by activating AMPK, which lowers COX-2 levels and stops cell growth. ROS triggers AMPK activation, making AMPK an important target for cancer prevention [213].
Selenium-rich foods and selenium nanoparticles both effectively inhibit the PI3K/AKT/mTOR signaling pathway in cancers. This action is mainly mediated by selenium-containing proteins [214].
Selenium may play a helpful role in fighting PDAC. Some studies have found that certain selenium compounds, like dibenzyl diselenide (DBDS), can trigger a special type of cancer cell death called ferroptosis, helping slow tumor growth. Other research suggests that combining selenium with chemotherapy drugs like gemcitabine can make the treatment more effective. Interestingly, many people with PDAC have low selenium levels—up to 90% in some studies—showing how important this nutrient might be. These findings suggest selenium could support PDAC treatment, though more research is still needed. A 2023 study found that selenium nanoparticles made with Prunella vulgaris helped fight PDAC, showing promise for selenium-based treatments. Another study showed that selenium can boost the cancer-fighting effects of juglone, suggesting they may work well together [215,216,217,218].
Selenium is a vital trace element important for antioxidant defense, immune function, and thyroid metabolism, found in foods like Brazil nuts, seafood, meat, and grains. It supports NK cell activity and may reduce cancer risk through selenoproteins that protect against oxidative damage and regulate pathways like PI3K/AKT/mTOR. Selenium compounds and nanoparticles show promise in enhancing chemotherapy efficacy and inducing cancer cell death, including in PDAC, highlighting its potential as an adjunctive cancer therapy.
7.2.7. Phosphorus
Phosphorus is an essential mineral that supports many important functions in the body. It helps form bones and teeth, plays a key role in energy production, and is part of DNA and RNA. Phosphorus is found in foods like meat, dairy, fish, nuts, seeds, and whole grains. Having the right amount of phosphorus is important for good health, as both too little and too much can cause problems [219].
Hypophosphatemia (serum phosphorus <2.5 mg/dL), frequently seen in cancer patients, particularly those with advanced stages. It can lead to severe complications and impact their prognosis. This condition can drastically impair tissue oxygenation and disrupt immune function [220]. Conversely high phosphorus levels reduce vitamin D to lower phosphate absorption and lead to tumor initiation [221]. Therefore, maintaining a balanced intake of phosphorus while avoiding both deficiency and toxicity is crucial for optimal cancer management.
Phosphorus is important for the proper functioning of NK cells. It helps regulate NK cell development, activation, and movement through signaling pathways that involve phosphorus-containing molecules. Studies show that phosphorus-based compounds can boost NK cell activity, enhancing the immune response against cancer and infections. Additionally, phosphorus affects the movement of NK cells to areas of the body where they are needed, such as infection sites or tumors. This shows how phosphorus supports NK cell functions and overall immune health [222,223,224,225,226].
The 2017 study by Sies highlights how cancer cells adapt to oxidative stress by increasing NADPH levels through pathways like AMPK activation and the pentose phosphate pathway (PPP), helping maintain redox balance and support rapid cell growth. While phosphorus is crucial for cellular functions, its role in modulating ROS, RNS, and RSS in cancer needs further exploration to understand its full impact on tumor biology and treatment. A 2021 study shows that elevated phosphate levels can increase mitochondrial potential and ROS production, contributing to oxidative stress and vascular issues. These studies emphasize the complex relationship between phosphorus levels and oxidative stress in cancer, indicating that both low and high phosphorus levels affect health outcomes [227,228].
Hypophosphatemia is common in cancer patients treated with mTOR inhibitors. The incidence of levels <2 mg/dL was 13% with temsirolimus, 4% with everolimus, and 21.7% with ridaforolimus [229].
A 2022 study demonstrated that mice fed a low-phosphate diet followed by ethanol ingestion exhibited increased susceptibility to alcohol-induced pancreatitis. This was attributed to decreased mitochondrial function and cellular ATP levels in pancreatic acinar cells, suggesting that low phosphate levels can exacerbate pancreatic inflammation [230]
Research has shown that patients undergoing liver and pancreas resections often experience postoperative hypophosphatemia. This condition is associated with increased serum levels of nicotinamide phosphoribosyltransferase (NAMPT), a phosphaturic factor that downregulates phosphate transporters in the kidneys, leading to phosphate wasting [231] Low phosphorus levels may weaken NK cell function, possibly allowing pancreatic cancer cells to escape immune detection. More research is needed to understand how phosphorus affects PDAC and if it can help improve treatment by boosting NK cell activity [226].
Phosphorus, specifically phosphorus-32 (^32P), has been studied as a potential treatment for PDAC. In one approach, ^32P microparticles are injected directly into the tumor, and when combined with chemotherapy, they may help slow tumor growth. A device called OncoSil™ delivers ^32P directly to pancreatic tumors, offering targeted treatment. While these treatments show promise, more research is needed to fully assess their effectiveness and safety in treating PDAC [232].
Further studies are needed to clarify any potential connections between phosphorus, NK cells, and PDAC.
Phosphorus is an essential mineral involved in bone formation, energy production, and nucleic acid synthesis, found in foods like meat, dairy, and whole grains. It supports NK cell development, activation, and migration, enhancing immune responses against cancer. Both low and high phosphorus levels can disrupt redox balance and cellular function, impacting cancer progression. Experimental therapies using phosphorus-32 (^32P) show potential in PDAC treatment, though further research is needed.
Figure 1.
Integrated Profiling of NK Cells, Redox Status, and Micronutrients in Blood and Tumor Tissue. Overview of a combined testing approach using blood samples and tumor biopsies to study cancer patients. NK Cell Profiling: Measures the number and activity of natural killer (NK) cells in the blood and inside the tumor. Redox Marker Testing: Checks for signs of oxidative stress using markers like malondialdehyde (MDA), 8-hydroxy-2′-deoxyguanosine(8-OHdG), and advanced glycation end products (AGEs). Nutrient Testing: Measures levels of important micronutrients like zinc, copper, and selenium that support immune and antioxidant function. This triple profiling helps build a more complete picture of a patient’s immune health, stress response, and nutritional status to guide treatment decisions.
Figure 1.
Integrated Profiling of NK Cells, Redox Status, and Micronutrients in Blood and Tumor Tissue. Overview of a combined testing approach using blood samples and tumor biopsies to study cancer patients. NK Cell Profiling: Measures the number and activity of natural killer (NK) cells in the blood and inside the tumor. Redox Marker Testing: Checks for signs of oxidative stress using markers like malondialdehyde (MDA), 8-hydroxy-2′-deoxyguanosine(8-OHdG), and advanced glycation end products (AGEs). Nutrient Testing: Measures levels of important micronutrients like zinc, copper, and selenium that support immune and antioxidant function. This triple profiling helps build a more complete picture of a patient’s immune health, stress response, and nutritional status to guide treatment decisions.
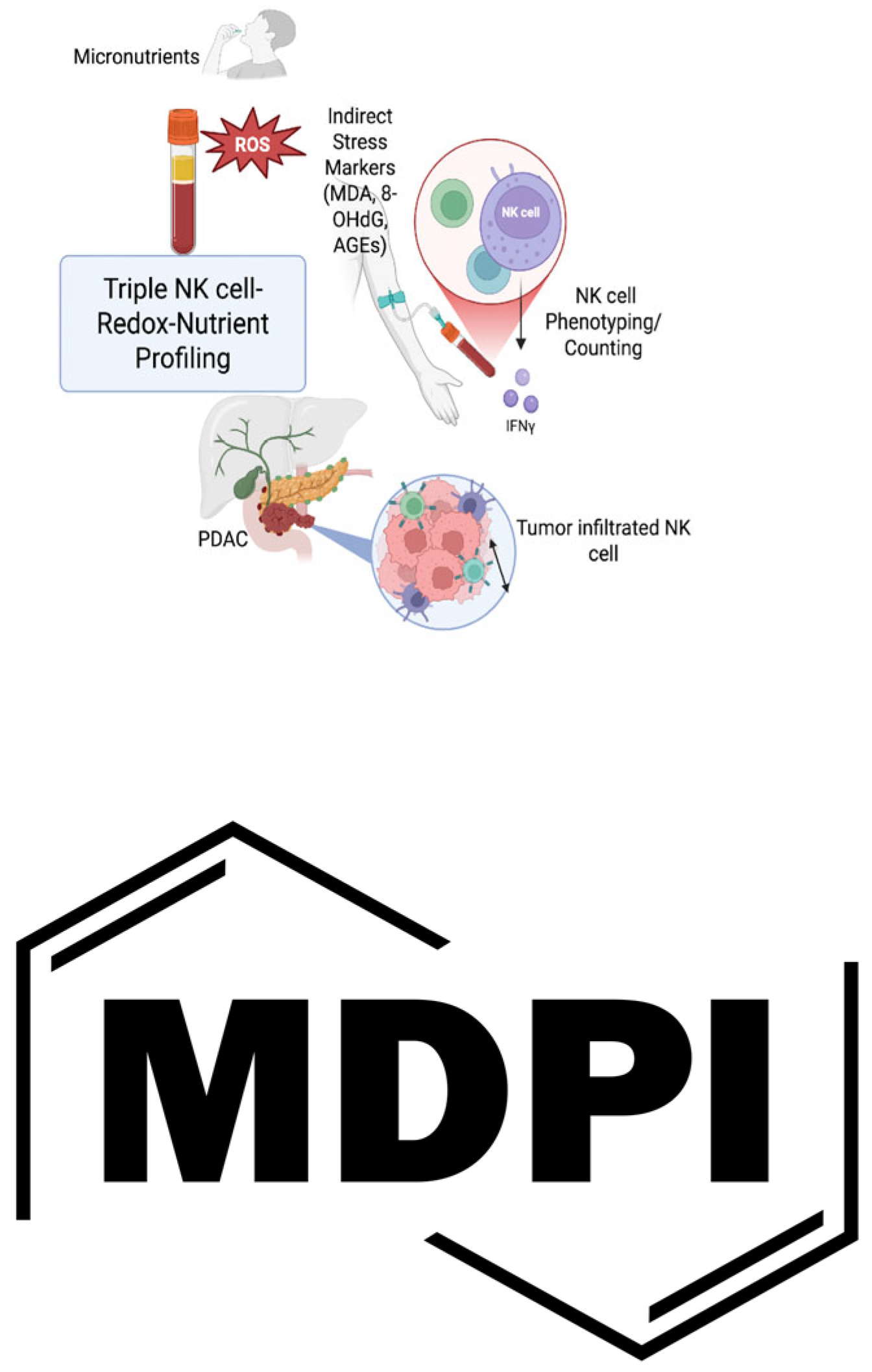
Figure 2.
Interaction Between Reactive Species Stress, Nutrient Stress, PDAC Metabolism, and NK Cell Activity. This figure illustrates the interconnected landscape of pancreatic ductal adenocarcinoma (PDAC) metabolism, nutrient/redox stress, and immune regulation by NK cells. PDAC Cell Metabolism (Nutrient-Deprived Conditions): Tumor cells in PDAC often exist in a nutrient-poor microenvironment, leading to metabolic adaptations that suppress mTOR (mechanistic target of rapamycin) signaling and activate AMPK (AMP-activated protein kinase). This shift promotes autophagy, survival, and stress resistance in PDAC cells. Reactive Species Stress (Oxidative Stress): Due to high metabolic demand and limited antioxidant defenses, PDAC cells produce excessive reactive oxygen and nitrogen species (ROS/RNS). Markers like MDA, 8-OHdG, and AGEs indicate elevated oxidative stress, which may further influence mTOR/AMPK signaling and immune evasion mechanisms. Nutrient Stress and Immune Crosstalk: Limited availability of key micronutrients (e.g., zinc, selenium) impairs both cancer cell metabolism and NK cell function. PDAC-induced nutrient depletion and oxidative stress can downregulate NK cell cytotoxicity by impairing receptor expression, granule release, and mitochondrial integrity. NK Cell Response: Despite the hostile microenvironment, NK cells attempt to target PDAC cells. However, persistent nutrient and oxidative stress in the tumor niche can dampen NK cell metabolism, reduce mTOR activation, and induce NK cell exhaustion or dysfunction. Overall, the figure highlights a vicious cycle in which PDAC-induced metabolic stress not only supports tumor survival via mTOR/AMPK modulation but also creates an immunosuppressive environment that weakens NK cell-mediated anti-tumor responses.
Figure 2.
Interaction Between Reactive Species Stress, Nutrient Stress, PDAC Metabolism, and NK Cell Activity. This figure illustrates the interconnected landscape of pancreatic ductal adenocarcinoma (PDAC) metabolism, nutrient/redox stress, and immune regulation by NK cells. PDAC Cell Metabolism (Nutrient-Deprived Conditions): Tumor cells in PDAC often exist in a nutrient-poor microenvironment, leading to metabolic adaptations that suppress mTOR (mechanistic target of rapamycin) signaling and activate AMPK (AMP-activated protein kinase). This shift promotes autophagy, survival, and stress resistance in PDAC cells. Reactive Species Stress (Oxidative Stress): Due to high metabolic demand and limited antioxidant defenses, PDAC cells produce excessive reactive oxygen and nitrogen species (ROS/RNS). Markers like MDA, 8-OHdG, and AGEs indicate elevated oxidative stress, which may further influence mTOR/AMPK signaling and immune evasion mechanisms. Nutrient Stress and Immune Crosstalk: Limited availability of key micronutrients (e.g., zinc, selenium) impairs both cancer cell metabolism and NK cell function. PDAC-induced nutrient depletion and oxidative stress can downregulate NK cell cytotoxicity by impairing receptor expression, granule release, and mitochondrial integrity. NK Cell Response: Despite the hostile microenvironment, NK cells attempt to target PDAC cells. However, persistent nutrient and oxidative stress in the tumor niche can dampen NK cell metabolism, reduce mTOR activation, and induce NK cell exhaustion or dysfunction. Overall, the figure highlights a vicious cycle in which PDAC-induced metabolic stress not only supports tumor survival via mTOR/AMPK modulation but also creates an immunosuppressive environment that weakens NK cell-mediated anti-tumor responses.


Table 1.
Supplement therapy in Cancer Managemcnet.
| Year, Reference | Cases | Cancer Type | Vitamin, Dosis | Mechanism | Conclusion |
|---|---|---|---|---|---|
| 1986, [Fraker LD] | Athymic mice | Breast Cancer |
Retinol | Increasing the cytotoxicity measurements using a specific cytotoxicity assay | NK cell levels in immunocompetent mice are increased significantly after retinol treatment |
| 1989[prabhala] | In vitro (human PMBC) | Canthaxanthin (vitamin A) | Increased NK cell activation marker | ||
| 1991[Prabhala] | Clinical | Betacaroten (Vitamin A) | Increased NK cell cytotoxicity | ||
| 1991[Watson] | Clinical | Betacaroten (Vitamin A) | Increased NK cell number | ||
| 1992[Garewal] | Clinical | Betacaroten (Vitamin A) | Increased NK cell number | ||
| 1994[Ozturk] |
Preclinical | Zinc | Increased NK cell cytotoxicity | ||
| 1996[Santos] | Clinical | Betacaroten | Increased NKA | ||
| 1997[Adachi] | Clinical (a 16-months boy with Shwachman-Diamons syndrome) | Vit E | Increased NKA | ||
| 1998[Xu] | Preclinical | Zinc | Low Zinc resulted in decreased NKA | ||
| 2006[Troen] | Clinical | Folic Acid | Folate supplement in women with low folate diet, increased NKA, while in folate rich diet decrease the NKA | ||
| 2007[Giacconi] | Clinical (atherosclerotic patients) | Intercellular Magnesium (Mg) and zinc | Decreased level of Mg, zinc leading to decreased NKA | ||
| 2007, [Hanson MG] | 13 Cancer patients | CRC, Duke C and D | Vitamin E, 750 mg/day, for 2 weeks | A slight but consistent increase in NKG2D expression in all CRC patients | Vitamin E can be an adjuvant in cancer immunotherapy |
| 2022, [Lou Y] | Cell line | Breast Cancer | ATRA, 200 mcg/day from day before to 3 day after cryo-thermal therapy | Vitamin A can enhance NK cell cytotoxicity and improve survival | |
| 2022, [Kim KS] | Cell line | Prostate Cancer | Ferumoxytol (iron) | Increase IFNG and NKG2D | Ferumoxytol enhance NK cell cytotoxicity and significant tumor volume regression |
| 2023[christofyllakis] | Clinical | Vitamin D | Increased the expression of over 120 NKA genes | ||
| 2024, [He C] | Cell line | PDAC | B6 supplement | Intracellular glycogen breakdown and energy production | Increase NK cell cytotoxicity and inhibit tumor growth |
| 2024, [Partearroyo] | Preclinical (aged rats) | B12 | Decreased level of B12, leading to decreased NKA |
ATRA: All ; CRC: Colorectal Cancer; IFNG: ; NKG2D: ; PDAC: Pancreatic Ductal Adenocarcinoma;.
This table provides an overview of studies investigating the use of vitamin and nutrient supplements as supportive therapies in cancer treatment. It includes information on cancer types, specific supplements and dosages used, proposed mechanisms of action—such as enhancing immune responses or modulating oxidative stress—and summarized conclusions from each study. A particular focus is placed on the role of supplements in influencing natural killer (NK) cell activity, redox balance, and overall immune modulation in various cancer contexts.
4. Discussion and Conclusions
Recent studies have highlighted the complex role of micronutrients in cancer treatment, particularly in their effect on oxidative stress and NK cell function. NK cells are vital in the body’s defense against cancer, helping to target and eliminate tumor cells. Our previous study shows that NK cell phenotype, including triple NK cell suppression mechanisms, plays an important role in the immune response within the PDAC TME [11].
The use of antioxidants in cancer therapy remains controversial due to their dual role in affecting redox stress and NK cell function. Antioxidants neutralize harmful reactive species like ROS, RNS, and RSS which can cause cell damage and contribute to cancer progression. However, excessive antioxidant supplementation may produce more reactive species, suppress NK cell activity, and promote tumor growth, especially in advanced stages [38,39,40,41,42]. Therefore, careful consideration of antioxidant intake, micronutrient types and their specific isoforms is necessary for an effective cancer treatment.
We propose that measuring both micronutrient levels, redox and nutrient stress markers could provide a clearer understanding of their effects on NK cells. Although directly measuring oxidative species in patients is challenging, relying on indirect markers, such as malondialdehyde (MDA), 8-hydroxy-2’-deoxyguanosine (8-OHdG), and advanced glycation end-products (AGEs), along with verifying circulating NK cell signatures, could help monitor NK cell status and guide treatment responses. This approach offers a non-invasive way to track immune function and optimize cancer treatment strategies.
Furthermore, clinical trials on micronutrients and cancer treatment need to be precise and consider the stage of cancer. For example, interventions in early-stage cancers may differ from those in advanced-stage cancers. It’s also important to consider different micronutrient isoforms, as they may have varied effects on NK cell function. Without this attention to detail, results could be misleading.
Looking ahead, more well-designed studies are needed to investigate how micronutrients can improve NK cell function and cancer treatment outcomes. Research should also explore different micronutrient forms to understand how they influence the immune system and tumor progression.
The interaction between micronutrients and mTOR signaling, as it relates to NK cells, is another important aspect that needs more focus. This is particularly relevant because the effects of micronutrients can be context dependent. For example, when a micronutrient works as an mTORC1 inhibitor in cancer cells, it has a cancer-preventive effect by limiting cell proliferation and survival. However, when the same micronutrient inhibits mTORC1 in NK cells, it can lead to NK cell suppression, which could reduce the immune system’s ability to detect and destroy cancer cells, potentially leading to carcinogenic effects. Therefore, understanding these nuanced interactions is crucial for optimizing cancer therapy strategies that involve micronutrient supplementation.
In conclusion, combining measuring micronutrient and stress markers with NK cell phenotyping could offer a promising approach to enhance cancer therapy. By adjusting treatment based on oxidative stress, micronutrient levels, and NK cell activity, we can improve responses to treatment and help combat cancer resistance. Our findings emphasize the importance of balanced antioxidant use, especially in advanced cancer stages. While antioxidants can support NK cell activity and counter treatment resistance, too much supplementation may have the opposite effect. This highlights the need for targeted, monitored, and personalized cancer therapies with proper micronutrient dosing.
A balanced intake of vitamins and minerals appears to be optimal for supporting NK cell function, maintaining redox homeostasis, and enhancing cancer treatment outcomes. This balance can be achieved through individualized diet plans tailored to each patient’s nutritional needs, ensuring sufficient micronutrient intake while avoiding excess that could impair immune function.
Author Contributions
S.F. contributes to the conceptualization and design of the review content. S.F. wrote the first draft of the manuscript. L.H.J. provide critical feedback and suggestions on the manuscript. Both authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Abbreviations
The following abbreviations are used in this manuscript:
| alpha-TS | alpha-TS alpha-tocopheryl succinate |
| ATRA | All-trans-retinoic acid |
| Ca. | Calcium |
| CRC | Colorectal Cancer |
| CU | Copper |
| IFNG | Interferon Gamma |
| KIR | Killer Immunoglobulin-like Receptor |
| MDSCs | Myeloid-derived suppressor cells |
| Mg | Magnesium |
| MSA | Methylseleninic acid |
| MTHFR | Methylenetetrahydrofolate reductase |
| MTs | Metallothioneins |
| NKA | Natural Killer cell Activity |
| NKG2D | Natural killer group 2, member D |
| NO | Nitric Oxide |
| PBMC | Peripheral blood mononuclear cell |
| PDAC | Pancreatic ductal adenocarcinoma |
| ROS. | Reactive Oxygen |
| TEPA | Tetraethylenepentamine Pentahydrochloride |
| TET | Ten-Eleven Translocation |
| TME | Tumor microenvironment |
| Zn. | Zinc |
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics. CA Cancer J. Clin. 2021, 71, 7–33.
- Wood, L.D.; Canto, M.I.; Jaffee, E.M.; Simeone, D.M. Pancreatic Cancer: Pathogenesis, screening, diagnosis, and treatment. Gastroenterology 2022, 163, 386–402.
- Conroy, T.; Desseigne, F.; Ychou, M.; Bouché, O.; Guimbaud, R.; Bécouarn, Y.; Adenis, A.; Raoul, J.L.; Gourgou-Bourgade, S.; De La Fouchardière, C.; et al. FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N. Engl. J. Med. 2011, 364, 1817–1825.
- Fanijavadi, S.; Jensen, L.H. Dysbiosis–NK Cell Crosstalk in Pancreatic Cancer: Toward a Unified Biomarker Signature for Improved Clinical Outcomes. Int. J. Mol. Sci. 2025, 26, 730. [CrossRef]
- Hayyan, M., Hashim, M. A., & AlNashef, I. M. (2016). Superoxide ion: Generation and chemical implications. Chemical Reviews, 116(5), 3029–3085. [CrossRef]
- Saha, S., et al. (2022). Role of ROS in the development of chemoresistance in pancreatic cancer. Antioxidants, 11(7), 1355.); (Galli & Ruggiero.
- Xu, G., et al. (2023). Nitric oxide in the tumor microenvironment: Implications for therapy in pancreatic cancer. Cancer Letters, 529, 172-181.
- Zhang, X., et al. (2023). Hydrogen sulfide as a potential therapeutic target in pancreatic cancer. Frontiers in Pharmacology, 14, 725908.).
- Jiang, H., et al. (2023). The dual role of nitric oxide in immune modulation in pancreatic cancer. Journal of Immunology, 210(2), 319-328.
- Galli, F., & Ruggiero, A. Hydrogen sulfide and its role in redox signaling in immune cells. Biochimica et Biophysica Acta (BBA)—General Subjects, 2016, 1860(10), 2187-2196].
- Gong J, Robbins LA, Lugea A, Waldron RT, Jeon CY, Pandol SJ. Diabetes, pancreatic cancer, and metformin therapy. Front Physiol (2014) 5:426. 10.3389/fphys.2014.00426.
- Iriana S, Ahmed S, Gong J, Annamalai AA, Tuli R, Hendifar AE. Targeting mTOR in Pancreatic Ductal Adenocarcinoma. Front Oncol. 2016 Apr 25;6:99. [CrossRef] [PubMed] [PubMed Central]
- Friedman MD, Jeevan DS, Tobias M, Murali R and Jhanwar-Uniyal M: Targeting cancer stem cells in glioblastoma multiforme using mTOR inhibitors and the differentiating agent all-trans retinoic acid. Oncol Rep 30: 1645-1650, 2013.
- Fanijavadi, S.; Thomassen, M.; Jensen, L.H. Targeting Triple NK Cell Suppression Mechanisms: A Comprehensive Review of Biomarkers in Pancreatic Cancer Therapy. Int. J. Mol. Sci. 2025, 26, 515. [CrossRef]
- Bhat, S. U., et al. (2022). Therapeutic resistance in pancreatic cancer: Role of RNS and NO in resistance to gemcitabine. Frontiers in Oncology, 12, 746218.
- Li, Y., et al. (2022). Hydrogen sulfide: A novel modulator of redox signaling in pancreatic cancer. Redox Biology, 45, 102053.
- Muller, L., et al. (2023). Reactive sulfur species in cancer and their modulation: Implications for therapy. Antioxidants & Redox Signaling, 39(5), 547-560.).
- Lal MK, Sharma E, Tiwari RK, Devi R, Mishra UN, Thakur R, Gupta R, Dey A, Lal P, Kumar A, Altaf MA, Sahu DN, Kumar R, Singh B, Sahu SK. Nutrient-Mediated Perception and Signalling in Human Metabolism: A Perspective of Nutrigenomics. Int J Mol Sci. 2022 Sep 25;23(19):11305. doi: 10.3390/ijms231911305. [PubMed] [PubMed Central]
- Vaziri-Gohar, A., Cassel, J., Mohammed, F.S. et al. Limited nutrient availability in the tumor microenvironment renders pancreatic tumors sensitive to allosteric IDH1 inhibitors. Nat Cancer 3, 852–865 (2022). [CrossRef]
- Saravia J, Raynor JL, Chapman NM, Lim SA, Chi H. Signaling networks in immunometabolism. Cell Res. 2020 Apr;30(4):328-342. [CrossRef] [PubMed] [PubMed Central]
- Faivre S, Kroemer G, Raymond E. Current development of mTOR inhibitors as anticancer agents. Nat Rev Drug Discov (2006) 5:671–88. [CrossRef]
- Populo H, Lopes JM, Soares P. The mTOR signalling pathway in human cancer. Int J Mol Sci (2012) 13:1886–918. [CrossRef]
- Wang, N., Wang, B., Maswikiti, E.P. et al. AMPK–a key factor in crosstalk between tumor cell energy metabolism and immune microenvironment?. Cell Death Discov. 10, 237 (2024). [CrossRef]
- Schneider, J. Hilbert, F. Genevaux, S. Höfer, L. Krauß, F. Schicktanz, C. T. Contreras, S. Jansari, A. Papargyriou, T. Richter, A. M. Alfayomy, C. Falcomatà, C. Schneeweis, F. Orben, R. Öllinger, F. Wegwitz, A. Boshnakovska, P. Rehling, D. Müller, P. Ströbel, V. Ellenrieder, L. Conradi, E. Hessmann, M. Ghadimi, M. Grade, M. Wirth, K. Steiger, R. Rad, B. Kuster, W. Sippl, M. Reichert, D. Saur, G. Schneider, A Novel AMPK Inhibitor Sensitizes Pancreatic Cancer Cells to Ferroptosis Induction. Adv. Sci. 2024, 11, 2307695. [CrossRef]
- R S, et al. (2023) The total mass, number, and distribution of immune cells in the human body. Proc Natl Acad Sci USA 120.
- Huber, M.; Brehm, C.U.; Gress, T.M.; Buchholz, M.; Alashkar Alhamwe, B.; von Strandmann, E.P.; Slater, E.P.; Bartsch, J.W.; Bauer, C.; Lauth, M. The Immune Microenvironment in Pancreatic Cancer. Int. J. Mol. Sci. 2020, 21, 7307. [CrossRef]
- TJ L, A B, K R (2022) Natural killer cells in antitumor adoptive cell immunotherapy. Nat Rev Cancer 22(10). [CrossRef]
- E V, L R, E NM, et al. (2024) Natural killer cell therapies. Nature 626. URL . [CrossRef]
- J C, F G, et al. (2022a) Natural killer cells: a promising immunotherapy for cancer. J Transl Med 20(1). [CrossRef]
- Ran G, q LY, et al. (2022) Natural killer cell homing and trafficking in tissues and tumors: from biology to application. Sig Transduct Target Ther 7(205). URL . [CrossRef]
- M‘ F, PM A, KY T (2023) Potentiation of natural killer cells to overcome cancer resistance to NK cell-based therapy and to enhance antibody-based immunotherapy. Front Immunol . [CrossRef]
- M G, A R (2018) Effect of Natural Compounds on NK Cell Activation. J Immunol Res . [CrossRef]
- Cuenca-Micó O, Aceves C. Micronutrients and Breast Cancer Progression: A Systematic Review. Nutrients. 2020 Nov 25;12(12):3613. doi: 10.3390/nu12123613.
- N D, F G, et al. (2021) NK Cell Therapy: A Rising Star in Cancer Treatment. Cancers (Basel) 13(16). [CrossRef]
- KN P, A K, et al. (1999) High doses of multiple antioxidant vitamins: essential ingre- dients in improving the efficacy of standard cancer therapy. J Am Coll Nutr 18(1). [CrossRef]
- KN P, WC C, et al. (2002) Pros and cons of antioxidant use during radiation therapy. Cancer Treat Rev 28(2). [CrossRef]
- CB A, GR Z, et al. (2020) Dietary Supplement Use During Chemotherapy and Survival Outcomes of Patients with Breast Cancer Enrolled in a Cooperative Group Clinical Trial (SWOG S0221). J Clin Oncol 38(8). [CrossRef]
- Yamasaki, H., et al. (2024). Spatiotemporal Characteristics Determining the Multifaceted Nature of Reactive Oxygen, Nitrogen, and Sulfur Species in Relation to Proton Homeostasis. Antioxidants & Redox Signaling.
- Yang Y, Neo SY, Chen Z, Cui W, Chen Y, Guo M, Wang Y, Xu H, Kurzay A, Alici E, Holmgren L, Haglund F, Wang K, Lundqvist A. Thioredoxin activity confers resistance against oxidative stress in tumor-infiltrating NK cells. J Clin Invest. 2020 Oct 1;130(10):5508-5522. [CrossRef] [PubMed] [PubMed Central]
- Chia, D., et al. (2025). Navigating the dichotomy of reactive oxygen, nitrogen, and sulfur species: detection strategies and therapeutic interventions. RSC Chemical Biology].
- Mariappan, M. M., & Balasubramanian, K. (2017). Nitrosative stress in immune responses: The role of reactive nitrogen species in NK cell function. Antioxidants & Redox Signaling, 26(11), 595-608., Reference: Ju, R. L., & Wu, F. (2018). Nitrosative stress and the role of nitric oxide in natural killer cell function. Journal of Immunology Research, 2018, Article ID 6234321.
- Andrés, C. M. C., et al. (2022). The Role of Reactive Species on Innate Immunity. Vaccines, 10(10), 1735. [CrossRef]
- Yang, G., & Wu, L. (2015). Hydrogen sulfide and its effects on immune cell function. Frontiers in Immunology, 6, 2. [CrossRef]
- Borde S, Matosevic S. Metabolic adaptation of NK cell activity and behavior in tumors: challenges and therapeutic opportunities. Trends Pharmacol Sci. 2023 Nov;44(11):832-848. [CrossRef] [PubMed]
- Yang C, Tsaih SW, Lemke A, Flister MJ, Thakar MS, Malarkannan S. mTORC1 and mTORC2 differentially promote natural killer cell development. Elife. 2018 May 29;7:e35619. [CrossRef] [PubMed] [PubMed Central]
- Ouissam AJ, Hind C, Sami Aziz B, Said A. Inhibition of the PI3K/AKT/mTOR pathway in pancreatic cancer: is it a worthwhile endeavor? Ther Adv Med Oncol. 2024 Oct 4;16:17588359241284911. [CrossRef] [PubMed] [PubMed Central]
- R.H. P, G.S H, et al. (1991) The effects of 13-cis-retinoic acid and beta-carotene on cellular immunity in humans. Cancer 67. [CrossRef]
- Ashley H Davis-Yadley, Mokenge P Malafa, Vitamins in Pancreatic Cancer: A Review of Underlying Mechanisms and Future Applications, Advances in Nutrition, Volume 6, Issue 6, 2015, Pages 774-802, ISSN 2161-8313. (https://www.sciencedirect.com/science/article/pii/S2161831323001242). [CrossRef]
- N T, D S, et al. (2022) Vitamin A in health care: Suppression of growth and induction of differentiation in cancer cells by vitamin A and its derivatives and their mechanisms of action. Pharmacol Ther . [CrossRef]
- MV G, PN H, et al. (2020) Current Trends in ATRA Delivery for Cancer Therapy. Pharmaceutics pp 12–8. [CrossRef]
- M.A. C, A Z, M. T, et al. (2024) Cellular and micro-environmental responses influencing the antitumor activity of all-trans retinoic acid in breast cancer. Cell Commun Signal 22(127). URL . [CrossRef]
- J Y, Y Z, et al. (2022c) β-Carotene Supplementation and Risk of Cardiovascular Dis- ease: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 14(6). [CrossRef]
- The Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study Group. The effect of vitamin E and beta carotene on the incidence of lung cancer and other cancers in male smokers. N Engl J Med. 1994;330(15):1029–1035. [CrossRef]
- Omenn GS, et al. Effects of a combination of beta carotene and vitamin A on lung cancer and cardiovascular disease. N Engl J Med. 1996;334(18):1150–1155. [CrossRef]
- Y L, P P, et al. (2022) Combining all-trans retinoid acid treatment targeting myeloid- derived suppressive cells with cryo-thermal therapy enhances antitumor immunity in breast cancer. Front Immunol 13. [CrossRef]
- A B, A K, et al. (2014) Saffron and natural carotenoids: Biochemical activities and anti- tumor effects. Biochim Biophys Acta 1854(1). [CrossRef]
- R.H. P, V. M, et al. (1989) Enhancement of the expression of activation markers of human peripheral blood mononuclear cells by in vitro culture with retinoids and carotenoids J Leuk Biol 45. [CrossRef]
- R.R. W, R.H. P, et al. (1991) Effects of β-carotene on lymphocyte subpopulations in elderly humans: evidence for a dose-response relationship. Am J Clin Nutr 53.
- H.S. G, N.M. A, et al. (1992) A preliminary trial of beta carotene in subjects infected with the human immunodeficiency virus. J Nutr 122. [CrossRef]
- Grudzien, Malgorzata, Rapak, Andrzej, Effect of Natural Compounds on NK Cell Activation, Journal of Immunology Research, 2018, 4868417, 11 pages, 2018. [CrossRef]
- Hu, H., et al. (2023). Vitamin A modulates immune cell responses through regulation of reactive oxygen species. Frontiers in Immunology, 14, 947021.
- Müller, L., et al. (2022). Vitamin A and its impact on nitric oxide production in immune cells. International Journal of Molecular Sciences, 23(12), 6924.
- Kolluru, G. K., et al. (2023). Retinoic acid and the regulation of reactive sulfur species in immune function. Redox Biology, 55, 102380.
- Zhang T, Chen H, Qin S, Wang M, Wang X, Zhang X, Liu F, Zhang S. The association between dietary vitamin A intake and pancreatic cancer risk: a meta-analysis of 11 studies. Biosci Rep. 2016 Nov 22;36(6): e00414. [CrossRef] [PubMed] [PubMed Central]
- Guan J, Zhang H, Wen Z, Gu Y, Cheng Y, Sun Y, Zhang T, Jia C, Lu Z, Chen J. Retinoic acid inhibits pancreatic cancer cell migration and EMT through the downregulation of IL-6 in cancer associated fibroblast cells. Cancer Lett. 2014 Apr 1;345(1):132-9. [CrossRef] [PubMed]
- Pili R, Salumbides B, Zhao M, Altiok S, Qian D, Zwiebel J, Carducci MA, Rudek MA. Phase I study of the histone deacetylase inhibitor entinostat in combination with 13-cis retinoic acid in patients with solid tumours. Br J Cancer. 2012 Jan 3;106(1):77-84. [CrossRef] [PubMed] [PubMed Central]
- HM K, B B, et al. (2020) Phase I clinical trial repurposing all-trans retinoic acid as a stromal targeting agent for pancreatic cancer. Nat Commun. 11(1). [CrossRef]
- Kocher HM, Basu B, Froeling FEM, Sarker D, Slater S, Carlin D, deSouza NM, De Paepe KN, Goulart MR, Hughes C, Imrali A, Roberts R, Pawula M, Houghton R, Lawrence C, Yogeswaran Y, Mousa K, Coetzee C, Sasieni P, Prendergast A, Propper DJ. Phase I clinical trial repurposing all-trans retinoic acid as a stromal targeting agent for pancreatic cancer. Nat Commun. 2020 Sep 24;11(1):4841. [CrossRef] [PubMed] [PubMed Central]
- K C, F N, M B, et al. (2023) Vitamin D Enhances Immune Effector Pathways of NK Cells Thus Providing a Mechanistic Explanation for the Increased Effectiveness of Therapeutic Monoclonal Antibodies. Nutrients 15(16). [CrossRef]
- Sheeley MP, Andolino C, Kiesel VA, Teegarden D. Vitamin D regulation of energy metabolism in cancer. Br J Pharmacol. 2022 Jun;179(12):2890-2905. [CrossRef] [PubMed] [PubMed Central]
- PD C, WY C, et al. (2020) Effect of Vitamin D3 Supplements on Development of Advanced Cancer: A Secondary Analysis of the VITAL Randomized Clinical Trial. JAMA Netw Open 3(11). [CrossRef]
- JD S, JE M, R S (2020) Vitamin D and Clinical Cancer Outcomes: A Review of Meta-Analyses. JBMR Plus 5(1). [CrossRef]
- GY L, CY P, et al. (2018) Differential effect of dietary vitamin D supplementation on natural killer cell activity in lean and obese mice. J Nutr Biochem 55. [CrossRef]
- F N, F A (2018) Determination of optimum vitamin D3 levels for NK cell-mediated rituximab- and obinutuzumab-dependent cellular cytotoxicity. Cancer Immunol Immunother 67(11). [CrossRef]
- S Y, MT C (2008) The vitamin D receptor is required for iNKT cell development. Proc Natl Acad Sci U S A 105(13). [CrossRef]
- Mijatović, S.; Savić-Radojević, A.; Plješa-Ercegovac, M.; Simić, T.; Nicoletti, F.; Maksimović-Ivanić, D. The Double-Faced Role of Nitric Oxide and Reactive Oxygen Species in Solid Tumors. Antioxidants 2020, 9, 374. [CrossRef]
- Ravid A, Koren R. The role of reactive oxygen species in the anticancer activity of vitamin D. Recent Results Cancer Res. 2003;164:357-67. [CrossRef] [PubMed]
- Fenercioglu AK. The Anti-Inflammatory Roles of Vitamin D for Improving Human Health. Curr Issues Mol Biol. 2024 Nov 26;46(12):13514-13525. [CrossRef] [PubMed] [PubMed Central]
- Quigley M, Rieger S, Capobianco E, Wang Z, Zhao H, Hewison M, Lisse TS. Vitamin D Modulation of Mitochondrial Oxidative Metabolism and mTOR Enforces Stress Adaptations and Anticancer Responses. JBMR Plus. 2021 Dec 1;6(1):e10572. [CrossRef] [PubMed] [PubMed Central]
- Wang, Yanan and He, Qingmin and Rong, Kang and Zhu, Mingyang and Zhao, Xiaoxiao and Zheng, Pengyuan and Mi, Yang; Vitamin D3 promotes gastric cancer cell autophagy by mediating p53/AMPK/mTOR signaling; Frontiers in Pharmacology; Volume 14—2023, 2024; https://www.frontiersin.org/journals/pharmacology/articles/10.3389/fphar.2023.1338260; ISSN 1663-9812. [CrossRef]
- Moukayed, M.; Grant, W.B. Molecular Link between Vitamin D and Cancer Prevention. Nutrients 2013, 5, 3993-4021. [CrossRef]
- Colston KW, James SY, Ofori-Kuragu EA, Binderup L, Grant AG. Vitamin D receptors and anti-proliferative effects of vitamin D derivatives in human pancreatic carcinoma cells in vivo and in vitro. Br J Cancer. 1997;76(8):1017-20. [CrossRef] [PubMed] [PubMed Central]
- Evans TR, Colston KW, Lofts FJ, Cunningham D, Anthoney DA, Gogas H, de Bono JS, Hamberg KJ, Skov T, Mansi JL. A phase II trial of the vitamin D analogue Seocalcitol (EB1089) in patients with inoperable pancreatic cancer. Br J Cancer. 2002 Mar 4;86(5):680-5. [CrossRef] [PubMed] [PubMed Central]
- A A, AJ K (2019) Vitamin E and its anticancer effects. Crit Rev Food Sci Nutr 59(17). [CrossRef]
- Q J (2019) Natural forms of vitamin E and metabolites-regulation of cancer cell death and underlying mechanisms. IUBMB Life 71(4). [CrossRef]
- KN P, B K, et al. (2003) Alpha-tocopheryl succinate, the most effective form of vitamin E for adjuvant cancer treatment: a review. J Am Coll Nutr 22(2). [CrossRef]
- CS Y, N S, AN K (????) Does vitamin E prevent or promote cancer? Cancer Prev Res (Phila) 5(5). [CrossRef]
- MK A, HS Z, M AW (2000) Vitamin E and beta-carotene affect natural killer cell function. Int J Food Sci Nutr.
- MG H, V O, et al. (2007) A short-term dietary supplementation with high doses of vitamin E increases NK cell cytolytic activity in advanced colorectal cancer patients. Cancer Immunol Immunother 56(7). [CrossRef]
- N A, M M, et al. (1997) Depressed natural killer cell activity due to decreased natural killer cell population in a vitamin E-deficient patient with Shwachman syndrome: reversible natural killer cell abnormality by alpha-tocopherol supplementation. Eur J Pediatr.
- Ehizuelen Ebhohimen, I., Stephen Okanlawon, T., Ododo Osagie, A., & Norma Izevbigie, O. (2021). Vitamin E in Human Health and Oxidative Stress Related Diseases. IntechOpen. [CrossRef]
- Jagdeep K. Sandhu, Arsalan S. Haqqani, H. Chaim Birnboim, Effect of Dietary Vitamin E on Spontaneous or Nitric Oxide Donor-Induced Mutations in a Mouse Tumor Model, JNCI: Journal of the National Cancer Institute, Volume 92, Issue 17, 6 September 2000, Pages 1429–1433. [CrossRef]
- Fabio Hecht, Marco Zocchi, Fatemeh Alimohammadi, Isaac S. Harris, Regulation of antioxidants in cancer, Molecular Cell, Volume 84, Issue 1, 2024, Pages 23-33, ISSN 1097-2765. https://www.sciencedirect.com/science/article/pii/S1097276523009176. [CrossRef]
- Yu Y, Hou L, Song H, Xu P, Sun Y, Wu K. Akt/AMPK/mTOR pathway was involved in the autophagy induced by vitamin E succinate in human gastric cancer SGC-7901 cells. Mol Cell Biochem. 2017 Jan;424(1-2):173-183. [CrossRef] [PubMed]
- Klein EA, Thompson IM Jr, Tangen CM, et al. Vitamin E and the Risk of Prostate Cancer: The Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA. 2011;306(14):1549–1556. [CrossRef]
- Husain K, Francois RA, Yamauchi T, Perez M, Sebti SM, Malafa MP. Vitamin E δ-tocotrienol augments the antitumor activity of gemcitabine and suppresses constitutive NF-κB activation in pancreatic cancer. Mol Cancer Ther. 2011 Dec;10(12):2363-72. [CrossRef] [PubMed] [PubMed Central]
- Kunnumakkara AB, Sung B, Ravindran J, Diagaradjane P, Deorukhkar A, Dey S, Koca C, Yadav VR, Tong Z, Gelovani JG, Guha S, Krishnan S, Aggarwal BB. {Gamma}-tocotrienol inhibits pancreatic tumors and sensitizes them to gemcitabine treatment by modulating the inflammatory microenvironment. Cancer Res. 2010 Nov 1;70(21):8695-705. Erratum in: Cancer Res. 2018 Sep 1;78(17):5181. doi: 10.1158/0008-5472.CAN-18-1826. [CrossRef] [PubMed] [PubMed Central]
- Gröber U, Reichrath J, Holick MF, Kisters K. Vitamin K: an old vitamin in a new perspective. Dermatoendocrinol. 2015 Jan 21;6(1):e968490. [CrossRef] [PubMed] [PubMed Central]
- Anqi Chen, Jialu Li, Nianxuan Shen, Haifeng Huang, Qinglei Hang, Vitamin K: New insights related to senescence and cancer metastasis, Biochimica et Biophysica Acta (BBA)—Reviews on Cancer, Volume 1879, Issue 2, 2024, 189057, ISSN 0304-419X, (https://www.sciencedirect.com/science/article/pii/S0304419X23002068) . [CrossRef]
- Gergues Marina , Bari Rafijul , Koppisetti Sharmila , Gosiewska Anna , Kang Lin , Hariri Robert J. Senescence, NK cells, and cancer: navigating the crossroads of aging and diseaseFrontiers in Immunology; Volume 16—2025; https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2025.1565278; ISSN=1664-3224. [CrossRef]
- Textor S, Bossler F, Henrich KO, Gartlgruber M, Pollmann J, Fiegler N, Arnold A, Westermann F, Waldburger N, Breuhahn K, Golfier S, Witzens-Harig M, Cerwenka A. The proto-oncogene Myc drives expression of the NK cell-activating NKp30 ligand B7-H6 in tumor cells. Oncoimmunology. 2016 Jul 28;5(7): e1116674. [CrossRef] [PubMed] [PubMed Central]
- Ponti, C., Gibellini, D., Boin, F., Melloni, E., Manzoli, F.A., Cocco, L., Zauli, G. and Vitale, M. (2002), Role of CREB transcription factor in c-fos activation in natural killer cells. Eur. J. Immunol., 32: 3358-3365. [CrossRef]
- Liu W, Nakamura H, Yamamoto T, Ikeda N, Saito M, Ohno M, Hara N, Imanishi H, Shimomura S, Yamamoto T, Sakai T, Nishiguchi S, Hada T. Vitamin K2 inhibits the proliferation of HepG2 cells by up-regulating the transcription of p21 gene. Hepatol Res. 2007 May;37(5):360-5. [CrossRef] [PubMed]
- Diana Schnoegl, Angela Hiesinger, Nicholas D Huntington, Dagmar Gotthardt, AP-1 transcription factors in cytotoxic lymphocyte development and antitumor immunity, Current Opinion in Immunology, Volume 85, 2023, 102397, ISSN 0952-7915. (https://www.sciencedirect.com/science/article/pii/S0952791523001164). [CrossRef]
- Article Source: Vitamin K2 Induces Mitochondria-Related Apoptosis in Human Bladder Cancer Cells via ROS and JNK/p38 MAPK Signal Pathways Duan F, Yu Y, Guan R, Xu Z, Liang H, et al. (2016) Vitamin K2 Induces Mitochondria-Related Apoptosis in Human Bladder Cancer Cells via ROS and JNK/p38 MAPK Signal Pathways. PLOS ONE 11(8): e0161886. [CrossRef]
- Duan F, Yu Y, Guan R, Xu Z, Liang H, et al. (2016) Vitamin K2 Induces Mitochondria-Related Apoptosis in Human Bladder Cancer Cells via ROS and JNK/p38 MAPK Signal Pathways. PLOS ONE 11(8): e0161886. [CrossRef]
- Duan, F., Mei, C., Yang, L. et al. Vitamin K2 promotes PI3K/AKT/HIF-1α-mediated glycolysis that leads to AMPK-dependent autophagic cell death in bladder cancer cells. Sci Rep 10, 7714 (2020). [CrossRef]
- Showalter SL, Wang Z, Costantino CL, Witkiewicz AK, Yeo CJ, Brody JR, Carr BI. Naturally occurring K vitamins inhibit pancreatic cancer cell survival through a caspase-dependent pathway. J Gastroenterol Hepatol. 2010 Apr;25(4):738-44. [CrossRef] [PubMed]
- Wei G, Wang M, Carr BI. Sorafenib combined vitamin K induces apoptosis in human pancreatic cancer cell lines through RAF/MEK/ERK and c-Jun NH2-terminal kinase pathways. J Cell Physiol. 2010 Jul;224(1):112-9. [CrossRef] [PubMed] [PubMed Central]
- Osada S, Tomita H, Tanaka Y, Tokuyama Y, Tanaka H, Sakashita F, Takahashi T. The utility of vitamin K3 (menadione) against pancreatic cancer. Anticancer Res. 2008 Jan-Feb;28(1A):45-50. [PubMed]
- Hanna M, Jaqua E, Nguyen V, Clay J. B Vitamins: Functions and Uses in Medicine. Perm J. 2022 Jun 29;26(2):89-97. [CrossRef] [PubMed] [PubMed Central]
- AM T, B M, B S, et al. (2006) Unmetabolized folic acid in plasma is associated with reduced natural killer cell cytotoxicity among postmenopausal women. J Nutr 136(1). [CrossRef]
- CT P, DA R, et al. (2020) B Vitamins and Their Role in Immune Regulation and Cancer. Nutrients. Nutrients 12(11). [CrossRef]
- X C, H A, et al. (2019) Association of intake folate and related gene polymorphisms with breast cancer. J Nutr Sci Vitaminol 65.
- Sohn Ji Yun, Kim Eun Young, Jeong Jae-Hwang, Kim Dae Joong, Nam Sang Yoon, Lee Hyun Jik, Lee Beom Jun. Protective effects of folic acid on colon carcinogenesis induced by azoxymethane in mice. J Biomed Transl Res 2021;22(3):111-120. [CrossRef]
- H S, J W, et al. (2016) High folic acid intake reduces natural killer cell cytotoxicity in aged mice. J Nutr Biochem . [CrossRef]
- J T, MHSMMTTTSKHKubota K, et al. (1999) Immunomodulation by vitamin B12: augmentation of CD8+ T lymphocytes and natural killer (NK) cell activity in vitamin B12-deficient patients by methyl-B12 treatment. . Clin Exp Immunol 116(1).
- T P, U’beda N, et al. (2013) Vitamin B (12) and folic acid imbalance modifies NK cyto- toxicity, lymphocytes B and lymphoprolipheration in aged rats. Nutrients. 5(12). [CrossRef]
- Frost, Z.; Bakhit, S.; Amaefuna, C.N.; Powers, R.V.; Ramana, K.V. Recent Advances on the Role of B Vitamins in Cancer Prevention and Progression. Int. J. Mol. Sci. 2025, 26, 1967. [CrossRef]
- Xie, S., Tan, M., Li, H. et al. Study on the correlation between B vitamins and breast cancer. Cancer Cell Int 23, 22 (2023). [CrossRef]
- Ayuk S.M., Abrahamse H. mTOR Signaling Pathway in Cancer Targets Photodynamic Therapy In Vitro. Cells. 2019;8:431. [CrossRef]
- Ueno M, Okusaka T, Omuro Y, Isayama H, Fukutomi A, Ikeda M, Mizuno N, Fukuzawa K, Furukawa M, Iguchi H, Sugimori K, Furuse J, Shimada K, Ioka T, Nakamori S, Baba H, Komatsu Y, Takeuchi M, Hyodo I, Boku N. A randomized phase II study of S-1 plus oral leucovorin versus S-1 monotherapy in patients with gemcitabine-refractory advanced pancreatic cancer. Ann Oncol. 2016 Mar;27(3):502-8. [CrossRef] [PubMed] [PubMed Central]
- C H, D W, SK S, et al. (2024) Vitamin B6 Competition in the Tumor Microenvironment Hampers Antitumor Functions of NK Cells. Cancer Discov. 14(1). [CrossRef]
- Jin-Peng Yong, Xiao-Yan Mu, Chao-Feng Zhou, Ke-Ke Zhang, Jie-Qiong Gao, Zhi-Zhong Guo, Shi-Fan Zhou, Zhen Ma, Radiofrequency ablation of liver metastases in a patient with pancreatic cancer and long-term survival: A case report, World Journal of Clinical Cases, 13, 20, (2025). [CrossRef]
- Carr, A.C.; Maggini, S. Vitamin C and Immune Function. Nutrients 2017, 9, 1211. [CrossRef]
- Wilson MK, Baguley BC, Wall C, Jameson MB, Findlay MP. Review of high-dose intravenous vitamin C as an anticancer agent. Asia Pac J Clin Oncol. 2014 Mar;10(1):22-37. [CrossRef] [PubMed]
- Ngo, B., Van Riper, J.M., Cantley, L.C. et al. Targeting cancer vulnerabilities with high-dose vitamin C. Nat Rev Cancer 19, 271–282 (2019). [CrossRef]
- Huijskens MJ, Walczak M, Sarkar S, Atrafi F, Senden-Gijsbers BL, Tilanus MG, Bos GM, Wieten L, Germeraad WT. Ascorbic acid promotes proliferation of natural killer cell populations in culture systems applicable for natural killer cell therapy. Cytotherapy. 2015 May;17(5):613-20. [CrossRef] [PubMed]
- M V, J F (2021) The Role of Vitamin C in Cancer Prevention and Therapy: A Litera- ture Review. Antioxidants (Basel) 10(12). [CrossRef]
- J GM, S C, et al. (2022b) Ascorbate as a Bioactive Compound in Cancer Ther- apy: The Old Classic Strikes Back. Molecules 27(12). [CrossRef]
- CY W, B Z, et al. (2020) Ascorbic Acid Promotes KIR Demethylation during Early NK Cell Differentiation. J Immunol 205(6). [CrossRef]
- Du J, Martin SM, Levine M, Wagner BA, Buettner GR, Wang SH, Taghiyev AF, Du C, Knudson CM, Cullen JJ. Mechanisms of ascorbate-induced cytotoxicity in pancreatic cancer. Clin Cancer Res. 2010 Jan 15;16(2):509-20. [CrossRef] [PubMed] [PubMed Central]
- Qin S, Wang G, Chen L, Geng H, Zheng Y, Xia C, Wu S, Yao J, Deng L. Pharmacological vitamin C inhibits mTOR signaling and tumor growth by degrading Rictor and inducing HMOX1 expression. PLoS Genet. 2023 Feb 14;19(2):e1010629. [CrossRef] [PubMed] [PubMed Central]
- Espey MG, Chen P, Chalmers B, Drisko J, Sun AY, Levine M, Chen Q. Pharmacologic ascorbate synergizes with gemcitabine in preclinical models of pancreatic cancer. Free Radic Biol Med. 2011 Jun 1;50(11):1610-9. [CrossRef] [PubMed] [PubMed Central]
- Pereira M, Cardeiro M, Frankel L, Greenfield B, Takabe K, Rashid OM. Increased Vitamin C Intake Is Associated With Decreased Pancreatic Cancer Risk. World J Oncol. 2024 Aug;15(4):543-549. [CrossRef] [PubMed] [PubMed Central]
- Fan, H., Kou, J., Han, D. et al. Association between vitamin C intake and the risk of pancreatic cancer: a meta-analysis of observational studies. Sci Rep 5, 13973 (2015). [CrossRef]
- Conte E, Rizzolo E, Flower G, Legacy M, Seely D. Intravenous Vitamin C in Cancer Care: Evidence Review and Practical Guidance for Integrative Oncology Practitioners. CANDJ [Internet]. 2024 Mar. 21 [cited 2025 Apr. 18];31(1):2-18. Available from: https://candjournal.ca/index.php/candj/article/view/149.
- Bodeker KL, Smith BJ, Berg DJ, Chandrasekharan C, Sharif S, Fei N, Vollstedt S, Brown H, Chandler M, Lorack A, McMichael S, Wulfekuhle J, Wagner BA, Buettner GR, Allen BG, Caster JM, Dion B, Kamgar M, Buatti JM, Cullen JJ. A randomized trial of pharmacological ascorbate, gemcitabine, and nab-paclitaxel for metastatic pancreatic cancer. Redox Biol. 2024 Nov;77:103375. [CrossRef] [PubMed] [PubMed Central]
- Polireddy, K., Dong, R., Reed, G. et al. High Dose Parenteral Ascorbate Inhibited Pancreatic Cancer Growth and Metastasis: Mechanisms and a Phase I/IIa study. Sci Rep 7, 17188 (2017). [CrossRef]
- Seyed Mohammad Taghi Gharibzahedi, Seid Mahdi Jafari, The importance of minerals in human nutrition: Bioavailability, food fortification, processing effects and nanoencapsulation, Trends in Food Science & Technology, Volume 62, 2017, Pages 119-132, ISSN 0924-2244, https://www.sciencedirect.com/science/article/pii/S0924224416306203. [CrossRef]
- Janakiram NB, Mohammed A, Madka V, Kumar G, Rao CV. Prevention and treatment of cancers by immune modulating nutrients. Mol Nutr Food Res. 2016 Jun;60(6):1275-94. [CrossRef] [PubMed] [PubMed Central]
- Pravina, P., Sayaji, D. and Avinash, M. (2013) Calcium and Its Role in Human Body. International Journal of Research in Pharmaceutical and Biomedical Sciences, 4, 659-668.
- Zhang M, Song R (2019) Calcium-Overload-Mediated Tumor Therapy by Calcium Peroxide Nanoparticles. CelPress URL . [CrossRef]
- EC S, B Q, M H (2013) Calcium, cancer and killing: the role of calcium in killing cancer cells by cytotoxic T lymphocytes and natural killer cells. Biochim Biophys Acta 1833(7). [CrossRef]
- A MP, SC C, et al. (2011) ORAI1-mediated calcium influx is required for human cytotoxic lymphocyte degranulation and target cell lysis Proc. Natl Acad Sci 108.
- Görlach A, Bertram K, Hudecova S, Krizanova O. Calcium and ROS: A mutual interplay. Redox Biol. 2015 Dec;6:260-271. [CrossRef] [PubMed] [PubMed Central]
- Rizzuto, R., De Stefani, D., Raffaello, A. et al. Mitochondria as sensors and regulators of calcium signalling. Nat Rev Mol Cell Biol 13, 566–578 (2012). [CrossRef]
- Huang Xuan , Reye Gina , Momot Konstantin I. , Blick Tony , Lloyd Thomas , Tilley Wayne D. , Hickey Theresa E. , Snell Cameron E. , Okolicsanyi Rachel K. , Haupt Larisa M. , Ferro Vito , Thompson Erik W. , Hugo Honor J; Heparanase Promotes Syndecan-1 Expression to Mediate Fibrillar Collagen and Mammographic Density in Human Breast Tissue Cultured ex vivo; Frontiers in Cell and Developmental Biology; Volume 8—2020 https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2020.00599; 10.3389/fcell.2020.00599; ISSN=2296-634X.
- Lin, Lan et al.; TRPV4 enhances the synthesis of fatty acids to drive the progression of ovarian cancer through the calcium-mTORC1/SREBP1 signaling pathway; iScience, Volume 26, Issue 11, 108226.
- Bettaieb L, Brulé M, Chomy A, Diedro M, Fruit M, Happernegg E, Heni L, Horochowska A, Housseini M, Klouyovo K, Laratte A, Leroy A, Lewandowski P, Louvieaux J, Moitié A, Tellier R, Titah S, Vanauberg D, Woesteland F, Prevarskaya N, Lehen’kyi V. Ca2+ Signaling and Its Potential Targeting in Pancreatic Ductal Carcinoma. Cancers (Basel). 2021 Jun 21;13(12):3085. [CrossRef] [PubMed] [PubMed Central]
- D.R. Principe,A.F. Aissa,S. Kumar,T.N.D. Pham,P.W. Underwood,R. Nair,R. Ke,B. Rana,J.G. Trevino,H.G. Munshi,E.V. Benevolenskaya,& A. Rana, Calcium channel blockers potentiate gemcitabine chemotherapy in pancreatic cancer, Proc. Natl. Acad. Sci. U.S.A. 119 (18) e2200143119. (2022). [CrossRef]
- Fong ZV, Severs G, Moir J, White S, Qadan M, Tingle S. Calcium channel blockers are associated with improved survival in pancreatic cancer patients undergoing neoadjuvant chemotherapy and resection. HPB (Oxford). 2024 Mar;26(3):418-425. [CrossRef] [PubMed]
- Bahrami A, Mohammadzadeh M, Abdi F, Paydareh A, Khalesi S, Hejazi E. Calcium Intake and the Pancreatic Cancer Risk: A Systematic Review and Meta-Analysis of Observational Studies. Clin Nutr Res. 2024 Oct 30;13(4):284-294. [CrossRef] [PubMed] [PubMed Central]
- Abbaspour N, Hurrell R, Kelishadi R. Review on iron and its importance for human health. J Res Med Sci. 2014 Feb;19(2):164-74. [PubMed] [PubMed Central]
- Q G, L L, S H, et al. (2021) The Role of Iron in Cancer Progression. Front Oncol . [CrossRef]
- Kim K, Choi B, et al. (2022) Enhanced natural killer cell anti-tumor activity with nanoparticles mediated ferroptosis and potential therapeutic application in prostate cancer. J Nanobiotechnol 20(428). URL . [CrossRef]
- An, X., Yu, W., Liu, J. et al. Oxidative cell death in cancer: mechanisms and therapeutic opportunities. Cell Death Dis 15, 556 (2024). [CrossRef]
- Zhou, Q., Meng, Y., Li, D. et al. Ferroptosis in cancer: from molecular mechanisms to therapeutic strategies. Sig Transduct Target Ther 9, 55 (2024). [CrossRef]
- Shang C, Zhou H, Liu W, Shen T, Luo Y, Huang S. Iron chelation inhibits mTORC1 signaling involving activation of AMPK and REDD1/Bnip3 pathways. Oncogene. 2020 Jul;39(29):5201-5213. [CrossRef] [PubMed] [PubMed Central]
- Jain V, Amaravadi RK. Pumping Iron: Ferritinophagy Promotes Survival and Therapy Resistance in Pancreatic Cancer. Cancer Discov. 2022 Sep 2;12(9):2023-2025. [CrossRef] [PubMed] [PubMed Central]
- Julián-Serrano S, Yuan F, Barrett MJ, Pfeiffer RM, Stolzenberg-Solomon RZ. Hemochromatosis, Iron Overload-Related Diseases, and Pancreatic Cancer Risk in the Surveillance, Epidemiology, and End Results (SEER)-Medicare. Cancer Epidemiol Biomarkers Prev. 2021 Nov;30(11):2136-2139. [CrossRef] [PubMed] [PubMed Central]
- Iron and Lipocalin-2 Modulate Cellular Responses in the Tumor Micro-environment of Pancreatic Ductal Adenocarcinoma; Valentina Pita-Grisanti, Andrew W. Dangel, Kristyn Gumpper, Andrea Ludwig, Olivia Ueltschi, Xiaokui Mo, Maciej Pietrzak, Amy Webb, Rosa F. Hwang, Madelyn Traczek, Niharika Badi, Zobeida Cruz-Monserrate; bioRxiv 2020.01.14.907188;. [CrossRef]
- Osmola M, Gierej B, Mleczko-Sanecka K, Jończy A, Ciepiela O, Kraj L, Ziarkiewicz-Wróblewska B, Basak GW. Anemia, Iron Deficiency, and Iron Regulators in Pancreatic Ductal Adenocarcinoma Patients: A Comprehensive Analysis. Curr Oncol. 2023 Aug 18;30(8):7722-7739. [CrossRef] [PubMed] [PubMed Central]
- S A, S K, et al. (2023) A narrative review on the role of magnesium in immune regulation, inflammation, infectious diseases, and cancer. J Health Popul Nutr 42(1). [CrossRef]
- Huang, WQ., Long, WQ., Mo, XF. et al. Direct and indirect associations between dietary magnesium intake and breast cancer risk. Sci Rep 9, 5764 (2019). [CrossRef]
- Chaigne-Delalande B, Li FY, O’Connor GM, Lukacs MJ, Jiang P, Zheng L, Shatzer A, Biancalana M, Pittaluga S, Matthews HF, Jancel TJ, Bleesing JJ, Marsh RA, Kuijpers TW, Nichols KE, Lucas CL, Nagpal S, Mehmet H, Su HC, Cohen JI, Uzel G, Lenardo MJ. Mg2+ regulates cytotoxic functions of NK and CD8 T cells in chronic EBV infection through NKG2D. Science. 2013 Jul 12;341(6142):186-91. [CrossRef] [PubMed] [PubMed Central]
- Fujita, K., Shindo, Y., Katsuta, Y. et al. Intracellular Mg2+ protects mitochondria from oxidative stress in human keratinocytes. Commun Biol 6, 868 (2023). [CrossRef]
- Kwesiga MP, Gillette AA, Razaviamri F, Plank ME, Canull AL, Alesch Z, He W, Lee BP, Guillory RJ 2nd. Biodegradable magnesium materials regulate ROS-RNS balance in pro-inflammatory macrophage environment. Bioact Mater. 2022 Nov 17;23:261-273. [CrossRef] [PubMed] [PubMed Central]
- Xie J, Cheng C, Zhu XY, Shen YH, Song LB, Chen H, Chen Z, Liu LM, Meng ZQ. Magnesium transporter protein solute carrier family 41 member 1 suppresses human pancreatic ductal adenocarcinoma through magnesium-dependent Akt/mTOR inhibition and bax-associated mitochondrial apoptosis. Aging (Albany NY). 2019 May 8; 11:2681-2698 . [CrossRef]
- Dibaba D, Xun P, Yokota K, White E, He K. Magnesium intake and incidence of pancreatic cancer: the VITamins and Lifestyle study. Br J Cancer. 2015 Dec 1;113(11):1615-21. [CrossRef] [PubMed] [PubMed Central]
- Xianpeng Qin, Jing Chen, Guiqing Jia, Zhou Yang,Dietary Factors and Pancreatic Cancer Risk: An Umbrella Review of Meta-Analyses of Prospective Observational Studies,Advances in Nutrition,Volume 14, Issue 3,2023,Pages 451-464,ISSN 2161-8313. (https://www.sciencedirect.com/science/article/pii/S2161831323002636). [CrossRef]
- H.E. Heilmaier, G.A. Drasch, E. Kretschmer, K.H. Summer, Metallothionein, cadmium, copper and zinc levels of human and rat tissues, Toxicology Letters,Volume 38, Issue 3,1987,Pages 205-211,ISSN 0378-4274. (https://www.sciencedirect.com/science/article/pii/0378427487900014. [CrossRef]
- Nordberg M, Nordberg GF. Metallothionein and Cadmium Toxicology-Historical Review and Commentary. Biomolecules. 2022 Feb 24;12(3):360. [CrossRef] [PubMed] [PubMed Central]
- zel A K, et al. (2017) The Functions of Metamorphic Metallothioneins in Zinc and Copper Metabolism. Int J Mol Sci 18(6). [CrossRef]
- M.George Cherian, A. Jayasurya, Boon-Huat Bay, Metallothioneins in human tumors and potential roles in carcinogenesis, Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis, Volume 533, Issues 1–2, 2003, Pages 201-209, ISSN 0027-5107. (https://www.sciencedirect.com/science/article/pii/S0027510703002173). [CrossRef]
- Takahashi, S. Molecular functions of metallothionein and its role in hematological malignancies. J Hematol Oncol 5, 41 (2012). [CrossRef]
- Yang, R.; Roshani, D.; Gao, B.; Li, P.; Shang, N. Metallothionein: A Comprehensive Review of Its Classification, Structure, Biological Functions, and Applications. Antioxidants 2024, 13, 825. [CrossRef]
- G O, D E, et al. (1994) Decreased natural killer (NK) cell activity in zinc-deficient rats. Gen Pharmacol 25(7). [CrossRef]
- H X, F Z, et al. (1998) Effect of zinc on the cellular immunity and NK cell activity in acute heat exposed rats. Wei Sheng Yan Jiu 27(5).
- Si, M., Lang, J. The roles of metallothioneins in carcinogenesis. J Hematol Oncol 11, 107 (2018). [CrossRef]
- ndreia O. Latorre, Beatriz D. Caniceiro, Heidge Fukumasu, Dale R. Gardner, Fabricio M. Lopes, Harry L. Wysochi, Tereza C. da Silva, Mitsue Haraguchi, Fabiana F. Bressan, Silvana L. Górniak, Ptaquiloside reduces NK cell activities by enhancing metallothionein expression, which is prevented by selenium, Toxicology, Volume 304, 2013, Pages 100-108, ISSN 0300-483X. (https://www.sciencedirect.com/science/article/pii/S0300483X12004295). [CrossRef]
- Li K, Zhang Z, Mei Y, Yang Q, Qiao S, Ni C, Yao Y, Li X, Li M, Wei D, Fu W, Guo X, Huang X, Yang H. Metallothionein-1G suppresses pancreatic cancer cell stemness by limiting activin A secretion via NF-κB inhibition. Theranostics. 2021 Jan 1;11(7):3196-3212. [CrossRef] [PubMed] [PubMed Central]
- Ohshio, G., Imamura, T., Okada, N. et al. Immunohistochemical study of metallothionein in pancreatic carcinomas. J Cancer Res Clin Oncol 122, 351–355 (1996). [CrossRef]
- Marreiro DD, Cruz KJ, Morais JB, Beserra JB, Severo JS, de Oliveira AR. Zinc and Oxidative Stress: Current Mechanisms. Antioxidants (Basel). 2017 Mar 29;6(2):24. [CrossRef] [PubMed] [PubMed Central]
- Lee SR. Critical Role of Zinc as Either an Antioxidant or a Prooxidant in Cellular Systems. Oxid Med Cell Longev. 2018 Mar 20;2018:9156285. [CrossRef] [PubMed] [PubMed Central]
- Nimmanon T, Ziliotto S, Morris S, Flanagan L, Taylor KM. Phosphorylation of zinc channel ZIP7 drives MAPK, PI3K and mTOR growth and proliferation signalling. Metallomics. 2017 May 24;9(5):471-481. [CrossRef] [PubMed] [PubMed Central]
- Ruiman Geng, Nengwen Ke, Ziyao Wang, Yu Mou, Bin Xiang, Zhengkun Zhang, Xuxu Ji, Jiaqiong Zou, Dingxue Wang, Zhaoru Yin, Xubao Liu, Fang Xie, Yanan Zhao, Dan Chen, Jingying Dong, Wenbing Wu, Lihong Chen, Huawei Cai, Ji Liu, Copper deprivation enhances the chemosensitivity of pancreatic cancer to rapamycin by mTORC1/2 inhibition, Chemico-Biological Interactions,Volume 382, 2023, 110546, ISSN 0009-2797. (https://www.sciencedirect.com/science/article/pii/S0009279723002132). [CrossRef]
- Niture S, Gadi S, Lin M, Qi Q, Niture SS, Moore JT, Bodnar W, Fernando RA, Levine KE, Kumar D. Cadmium modulates steatosis, fibrosis, and oncogenic signaling in liver cancer cells by activating notch and AKT/mTOR pathways. Environ Toxicol. 2023 Mar;38(4):783-797. [CrossRef] [PubMed]
- Li K, Zhang Z, Mei Y, Yang Q, Qiao S, Ni C, Yao Y, Li X, Li M, Wei D, Fu W, Guo X, Huang X, Yang H. Metallothionein-1G suppresses pancreatic cancer cell stemness by limiting activin A secretion via NF-κB inhibition. Theranostics. 2021 Jan 1;11(7):3196-3212. [CrossRef] [PubMed] [PubMed Central]
- TAKISHITA, CHIE and NAGAKAWA, YUICHI and OSAKABE, HIROAKI and NAKAGAWA, NOBUHIKO and MITSUKA, YUSUKE and MAZAKI, JUNICHI and IWASAKI, KENICHI and ISHIZAKI, TETSUO and KOZONO, SHINGO, Significance of Zinc Replacement Therapy After Pancreaticoduodenectomy, volume 42, 5833-5837, 2022. https://ar.iiarjournals.org/content/42/12/5833}, Anticancer Research. [CrossRef]
- Iseki M, Mizuma M, Shimura M, Kokumai T, Sato H, Kusaka A, Aoki S, Inoue K, Nakayama S, Douchi D, Miura T, Maeda S, Ishida M, Nakagawa K, Kamei T, Unno M. Preoperative Chemotherapy With Gemcitabine for Pancreatic Cancer Causes Zinc Deficiency. Pancreas. 2025 Feb 1;54(2):e75-e81. [CrossRef] [PubMed]
- Seiki Y, Ikezawa K, Watsuji K, Urabe M, Kai Y, Takada R, Yamai T, Mukai K, Nakabori T, Uehara H, Ishibashi M, Ohkawa K. Zinc supplementation for dysgeusia in patients with unresectable pancreatic cancer. Int J Clin Oncol. 2024 Aug;29(8):1173-1181. [CrossRef] [PubMed]
- Gunnarsdottir I, Dahl L. Iodine intake in human nutrition: a systematic literature review. Food Nutr Res. 2012;56. [CrossRef] [PubMed] [PubMed Central]
- M W, and Boral A KZ, et al. (2022) The Impact of Iodine Concentration Disorders on Health and Cancer. Nutrients 14(11). [CrossRef]
- Winder M, Kosztyła Z, Boral A, Kocełak P, Chudek J. The Impact of Iodine Concentration Disorders on Health and Cancer. Nutrients. 2022 May 26;14(11):2209. [CrossRef] [PubMed] [PubMed Central]
- Kawai, T., Akira, S. Innate immune recognition of viral infection. Nat Immunol 7, 131–137 (2006). [CrossRef]
- Xiaoyi Chen, Liegang Liu, Ping Yao, Dong Yu, Liping Hao, Xiufa Sun. Effect of excessive iodine on immune function of lymphocytes and intervention with selenium. Current Medical Science, 2007, 27(18): 422‒425 . [CrossRef]
- Xie S, Wu Z, Zhou L, Liang Y, Wang X, Niu L, Xu K, Chen J, Zhang M. Iodine-125 seed implantation and allogenic natural killer cell immunotherapy for hepatocellular carcinoma after liver transplantation: a case report. Onco Targets Ther. 2018;11:7345-7352. [CrossRef]
- Stanciu, A.E.; Verzia, A.; Stanciu, M.M.; Zamfirescu, A.; Gheorghe, D.C. Analysis of the Correlation between the Radioactive Iodine Activity and Neutrophil-to-Lymphocyte Ratio in Patients with Differentiated Thyroid Cancer. Cancers 2022, 14, 1899. [CrossRef]
- Karbownik-Lewińska M, Stępniak J, Iwan P, Lewiński A. Iodine as a potential endocrine disruptor-a role of oxidative stress. Endocrine. 2022 Nov;78(2):219-240. [CrossRef] [PubMed] [PubMed Central]
- Fuxiong Lu,Reactive oxygen species in cancer, too much or too little?, Medical Hypotheses,mVolume 69, Issue 6, 2007, Pages 1293-1298, ISSN 0306-9877.(https://www.sciencedirect.com/science/article/pii/S0306987707002514). [CrossRef]
- Siyu Liu, Peisen Ding, Xiaomeng Yu et al. Excess iodine promotes papillary thyroid carcinoma through the AKT/mTOR pathway, 19 November 2024, PREPRINT (Version 1) available at Research Square [. [CrossRef]
- J. Craps, V. Joris, B. De Jongh, P. Sonveaux, S. Horman, B. Lengelé, L. Bertrand, M.-C. Many, I. M. Colin, A.-C. Gérard, Involvement of mTOR and Regulation by AMPK in Early Iodine Deficiency-Induced Thyroid Microvascular Activation, Endocrinology, Volume 157, Issue 6, 1 June 2016, Pages 2545–2559. [CrossRef]
- Song, Yinghuia,b; Qi, Yuchenc; Yu, Zhangtaoa; Zhang, Zhihuaa; Li, Yuhanga; Huang, Junkaia; Liu, Sulaia,b,*. Iodine-125 seed implantation combined chemotherapy for metastatic pancreatic adenocarcinoma with primary colon cancer: A case report. Medicine 101(36):p e30349, September 09, 2022. |. [CrossRef]
- Kawamoto, S., Fuld, M. K., Laheru, D., Huang, P., & Fishman, E. K. (2018). Assessment of iodine uptake by pancreatic cancer following chemotherapy using dual-energy CT. Abdominal Radiology, 43(2), 445-456. [CrossRef]
- Yao H, ZhuGe Y, Jin S, Chen S, Zhang H, Zhang D, Chen Z. The efficacy of coaxial percutaneous iodine-125 seed implantation combined with arterial infusion chemotherapy for advanced pancreatic cancer: a randomized clinical trial. Int J Radiat Biol. 2024;100(7):1041-1050. [CrossRef] [PubMed]
- Sun Y, Wang Z, Gong P, Yao W, Ba Q, Wang H. Review on the health-promoting effect of adequate selenium status. Front Nutr. 2023 Mar 16;10:1136458. [CrossRef] [PubMed] [PubMed Central]
- SP S, CS W (2017) Selenoproteins in Tumorigenesis and Cancer Progression. Adv Cancer Res . [CrossRef]
- RJ L, GM J, et al. (2018) The Interaction of Selenium with Chemotherapy and Radiation on Normal and Malignant Human Mononuclear Blood Cells. Int J Mol Sci 19(10). [CrossRef]
- Yu Yang and Ying Liu and Qingxia Yang and Ting Liu, The Application of Selenium Nanoparticles in Immunotherapy,2024, Nano Biomedicine and Engineering, volume 16,3, 345-356; https://www.sciopen.com/article/10.26599/NBE.2024.9290100},.
- Gao S, Li T, Guo Y, Sun C, Xianyu B, Xu H. Selenium-Containing Nanoparticles Combine the NK Cells Mediated Immunotherapy with Radiotherapy and Chemotherapy. Adv Mater. 2020 Mar;32(12):e1907568. [CrossRef] [PubMed]
- Ali Razaghi, Mansour Poorebrahim, Dhifaf Sarhan, Mikael Björnstedt, Selenium stimulates the antitumour immunity: Insights to future research, European Journal of Cancer, Volume 155, 2021, Pages 256-267, ISSN 0959-8049. (https://www.sciencedirect.com/science/article/pii/S0959804921004627). [CrossRef]
- Murugan, D.; Murugesan, V.; Panchapakesan, B.; Rangasamy, L. Nanoparticle Enhancement of Natural Killer (NK) Cell-Based Immunotherapy. Cancers 2022, 14, 5438. [CrossRef]
- Wallenberg M, Misra S, Wasik AM, Marzano C, Björnstedt M, Gandin V, Fernandes AP. Selenium induces a multi-targeted cell death process in addition to ROS formation. J Cell Mol Med. 2014 Apr;18(4):671-84. [CrossRef] [PubMed] [PubMed Central]
- Parvin GhasemiAtefeh MaddahAlireza SalehzadehNasrin ZiamajidiAshkan Kalantary-CharvadehRoghayeh AbbasalipourkabirMaryam Salehzadehet al.Study of Oxidative Stress and Apoptotic Genes in SW480 Cell Lines Treated with Selenium Nanoparticles.J Rep Pharm Sci.13(1):e151064. [CrossRef]
- Hwang JT, Kim YM, Surh YJ, Baik HW, Lee SK, Ha J, Park OJ. Selenium regulates cyclooxygenase-2 and extracellular signal-regulated kinase signaling pathways by activating AMP-activated protein kinase in colon cancer cells. Cancer Res. 2006 Oct 15;66(20):10057-63. [CrossRef] [PubMed]
- Zhu Y, Pu Q, Zhang Q, et al. Selenium-binding protein 1 inhibits malignant progression and induces apoptosis via distinct mechanisms in non-small cell lung cancer. Cancer Med. 2023; 12: 17149-17170. [CrossRef]
- Roberta Noè, Noemi Inglese, Patrizia Romani, Thauan Serafini, Carlotta Paoli, Beatrice Calciolari, Marco Fantuz, Agata Zamborlin, Nicoletta C. Surdo, Vittoria Spada, Martina Spacci, Sara Volta, Maria Laura Ermini, Giulietta Di Benedetto, Valentina Frusca, Claudio Santi, Konstantinos Lefkimmiatis, Sirio Dupont, Valerio Voliani, Luca Sancineto, Alessandro Carrer, Organic Selenium induces ferroptosis in pancreatic cancer cells, Redox Biology, Volume 68, 2023, 102962,ISSN 2213-2317.(https://www.sciencedirect.com/science/article/pii/S2213231723003634). [CrossRef]
- Wooten, D. J., Sinha, I., & Sinha, R. (2022). Selenium Induces Pancreatic Cancer Cell Death Alone and in Combination with Gemcitabine. Biomedicines, 10(1), Article 149. [CrossRef]
- Zhang, Shihai & Wang, Yang & Wang, Meiqin & Jiang, Li & Ma, Xue & Huang, Yong & Liu, Ting & Zheng, Lin & Li, Yongjun. (2024). Construction and anti-pancreatic cancer activity of selenium nanoparticles stabilized by Prunella vulgaris polysaccharide. International Journal of Biological Macromolecules. 278. 134924. 10.1016/j.ijbiomac.2024.134924.
- Kaya DE, Göktürk F, Batırbek F, Arıkoğlu H. Investigation of the Effects of Juglone-Selenium Treatments on Epithelial-Mesenchimal Transition and Migration in BxPC-3 and PANC-1 Human Pancreatic Cancer Cells. Med J Bakirkoy. 2023 Sep;19(3):248-255. [CrossRef]
- Kraft, M.D. (2015), Phosphorus and Calcium. Nutrition in Clinical Practice, 30: 21-33. [CrossRef]
- S A, O M (2021) Hypophosphatemia in cancer patients. Clin Kidney J 14(11). [CrossRef]
- R.B. B (2019) Vitamin D, cancer, and dysregulated phosphate metabolism. Endocrine 65. URL . [CrossRef]
- Kerr WG, Colucci F. Inositol phospholipid signaling and the biology of natural killer cells. J Innate Immun. 2011;3(3):249-57. [CrossRef] [PubMed] [PubMed Central]
- Thirumalai A, Elboughdiri N, Harini K, Girigoswami K, Girigoswami A. Phosphorus-carrying cascade molecules: inner architecture to biomedical applications. Turk J Chem. 2023 Jun 23;47(4):667-688. [CrossRef] [PubMed] [PubMed Central]
- Zhu HY, Da WM. [Regulation of immunity by sphingosine 1-phosphate and its G protein-coupled receptors--review].
- L. He, J. Zhao, H. Li, B. Xie, L. Xu, G. Huang, T. Liu, Z. Gu, T. Chen, Metabolic Reprogramming of NK Cells by Black Phosphorus Quantum Dots Potentiates Cancer Immunotherapy. Adv. Sci. 2023, 10, 2202519. [CrossRef]
- Fiore PF, Di Pace AL, Conti LA, Tumino N, Besi F, Scaglione S, Munari E, Moretta L, Vacca P. Different effects of NK cells and NK-derived soluble factors on cell lines derived from primary or metastatic pancreatic cancers. Cancer Immunol Immunother. 2023 Jun;72(6):1417-1428. [CrossRef] [PubMed] [PubMed Central]
- Hayes JD, Dinkova-Kostova AT, Tew KD. Oxidative Stress in Cancer. Cancer Cell. 2020 Aug 10;38(2):167-197. [CrossRef] [PubMed] [PubMed Central]
- McCarty MF, Lerner A, DiNicolantonio JJ, Iloki-Assanga SB. High Intakes of Bioavailable Phosphate May Promote Systemic Oxidative Stress and Vascular Calcification by Boosting Mitochondrial Membrane Potential-Is Good Magnesium Status an Antidote? Cells. 2021 Jul 9;10(7):1744. [CrossRef] [PubMed] [PubMed Central]
- Soefje SA, Karnad A, Brenner AJ.. Common toxicities of mammalian target of rapamycin inhibitors. Target Oncol 2011; 6: 125–129.
- Lee JP, Darlington K, Henson JB, Kothari D, Niedzwiecki D, Farooq A, Liddle RA. Hypophosphatemia as a Predictor of Clinical Outcomes in Acute Pancreatitis: A Retrospective Study. Pancreas. 2024 Jan 1;53(1):e3-e8. [CrossRef] [PubMed] [PubMed Central]
- Zheng J, Glezerman IG, Sadot E, McNeil A, Zarama C, Gönen M, Creasy J, Pak LM, Balachandran VP, D’Angelica MI, Allen PJ, DeMatteo RP, Kingham PT, Jarnagin WR, Jaimes EA. Hypophosphatemia after Hepatectomy or Pancreatectomy: Role of the Nicotinamide Phosphoribosyltransferase. J Am Coll Surg. 2017 Oct;225(4):488-497e2. [CrossRef] [PubMed] [PubMed Central]
- Peiró-Valgañón V, Guardiola-Arévalo A, López Fernández A, Llorente Herrero E, Martín Fernández-Gallardo MT. A multidisciplinary challenge: Therapy with phosphorus-32 for locally advanced pancreatic cancer. Rev Esp Med Nucl Imagen Mol (Engl Ed). 2023 Nov-Dec;42(6):403-409. [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



