Submitted:
12 October 2025
Posted:
14 October 2025
You are already at the latest version
Abstract
In vitro germplasm conservation provides an alternative method for preserving plant species that are vulnerable to natural hazards or for which in situ conservation is costly and challenging to manage. This conservation approach includes two strategies: medium-term conservation, where plants are maintained through the slow growth of explants, facilitated by the gradual release of nutrients and low-temperature storage. The second approach involves long-term preservation, achieved through cryopreservation in liquid nitrogen. Cryopreservation allows for the storage of pollen, calli, somatic, and zygotic embryos. Significant progress has been made in cryopreservation, which was initially limited to cold-tolerant species. New techniques focus on conserving sensitive species, such as oil palm, through rapid dehydration and vitrification procedures using various plant materials, particularly polyembryoids and zygotic embryos. Initial incubation of somatic embryoids in high sucrose concentrations (0.3–0.5 M) enhances their viability, while further dehydration of propagules extends their shelf life. Various loading materials, including 10% (w/v) DMSO and 0.7 M sucrose, along with vitrification solutions like PSV2, have been evaluated with minimal adverse effects on propagule viability. Molecular analyses of the cryopreserved embryos indicate that conservation induces the expression of genes related to reactive oxygen species in somatic embryoids compared to zygotic embryos, likely due to their sensitivity to osmotic stress. This review aims to analyze the importance and challenges associated with various in vitro and cryopreservation methods. It provides an overview of the current advances in cryopreservation technology for oil palm. Moreover, the review offers an outlook on optimizing cryopreservation protocols and addressing challenges to overcome genotype specificity by understanding the mechanisms of stress tolerance in cryopreserved species.
Keywords:
1. Introduction
3. Recalcitrant and Intermediate Nature of Palm Species
4. In Vitro Conservation and Plant Tissue Culture
4.1. Factors Affecting In Vitro Conservation
5. Cryopreservation
5.1. Preparation of Propagules or Explants for Cryopreservation
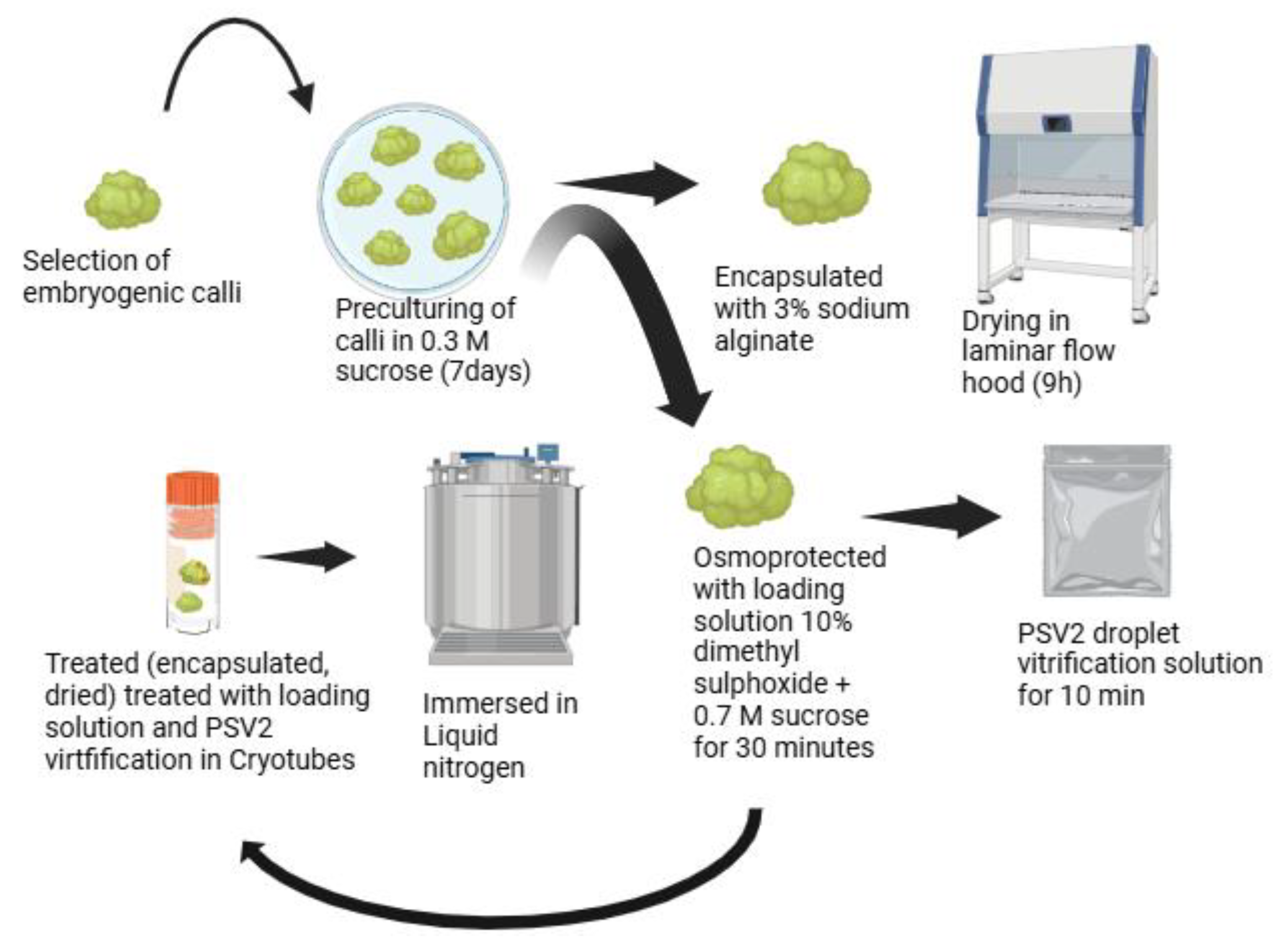
5.1. Pre-Culturing of Somatic Embryoids
5.2. Encapsulation-Dehydration
5.3. Vitrification
5.4. Droplet Vitrification
6. Genetic Modification of the Propagule
7. Problems with Cryopreservation and the Way Forward
8. Outlook
9. Conclusion
Author Contributions
Funding
Conflicts of Interest
References
- Murphy, D.J.; Goggin, K.; Paterson, R.R.M. Oil palm in the 2020s and beyond: Challenges and Solutions. CABI Agric. Biosci. 2021, 2, 39. [Google Scholar] [CrossRef]
- Bakoumé, C. Genetic Diversity, Erosion, and Conservation in Oil Palm (Elaeis guineensis Jacq.). In Genetic Diversity and Erosion in Plants; Ahuja, M.R., Jain, S.M., Eds.; Sustainable Development and Biodiversity; Springer International Publishing: Cham, 2016; ISBN 978-3-319-25953-6. [Google Scholar]
- Myint, K.A.; Amiruddin, M.D.; Rafii, M.Y.; Abd Samad, M.Y.; Ramlee, S.I.; Yaakub, Z.; Oladosu, Y. Genetic Diversity and Selection Criteria of MPOB-Senegal Oil Palm (Elaeis guineensis Jacq.) Germplasm by Quantitative Traits. Ind. Crops Prod. 2019, 139, 111558. [Google Scholar] [CrossRef]
- Norziha, A.; Zamri, Z.; Zulkifli, Y.; Fadila, A.M.; Marhalil, M. Selection Criteria of MPOB-Angola Germplasm Collection for Yield Improvement of the Oil Palm. Oil Crop Sci. 2024, 9, 20–28. [Google Scholar] [CrossRef]
- Tupaz-Vera, A.; Ayala-Diaz, I.; Barrera, C.F.; Romero, H.M. Genetic Gains for Obtaining Improved Progenies of Oil Palm in Colombia. Euphytica 2023, 219, 38. [Google Scholar] [CrossRef]
- Kushairi, A.; Rajanaidu, N.; Mohd Din, A. Mining the Germplasm. In Proceedings of the ISOPB seminar on the progress of oil palm breeding and selection, Medan, Sumatra, Indonesia; 2003; pp. 6–9. [Google Scholar]
- Rajanaidu, N.; Ainul, M.M. Conservation of Oil Palm and Coconut Genetic Resources. In Conservation of Tropical Plant Species; Normah, M.N., Chin, H.F., Reed, B.M., Eds.; Springer New York: New York, NY, 2013; ISBN 978-1-4614-3775-8. [Google Scholar]
- Gan, S.T.; Teo, C.J.; Manirasa, S.; Wong, W.C.; Wong, C.K. Assessment of Genetic Diversity and Population Structure of Oil Palm (Elaeis guineensis Jacq.) Field Genebank: A Step towards Molecular-Assisted Germplasm Conservation. PLoS One 2021, 16, e0255418. [Google Scholar] [CrossRef] [PubMed]
- Fadila, A.M.; Norziha, A.; Mohd Din, A.; Rajanaidu, N.; Kushairi, A. Evaluation of Bunch Index in MPOB Oil Palm (Elaeis guineensis Jacq.) Germplasm Collection. J. Oil Palm Res. 2016, 28, 442–451. [Google Scholar] [CrossRef]
- Kushairi, A.; Rajanaidu, N.; Mohd Din, A.; Isa, Z.A.; Noh, A.; Junaidah, J. PS5: Breeding Populations Selected for Thin Shell Teneras. MPOB Inf. Ser. 2003. [Google Scholar]
- Hayati, A.; Wickneswari, R.; Maizura, I.; Rajanaidu, N. Genetic Diversity of Oil Palm (Elaeis guineensis Jacq.) Germplasm Collections from Africa: Implications for Improvement and Conservation of Genetic Resources. Theor. Appl. Genet. 2004, 108, 1274–1284. [Google Scholar] [CrossRef]
- Ithnin, M.; Teh, C.-K.; Ratnam, W. Genetic Diversity of Elaeis Oleifera (HBK) Cortes Populations Using Cross Species SSRs: Implication’s for Germplasm Utilization and Conservation. BMC Genet. 2017, 18, 37. [Google Scholar] [CrossRef]
- Bakoumé, C.; Wickneswari, R.; Siju, S.; Rajanaidu, N.; Kushairi, A.; Billotte, N. Genetic Diversity of the World’s Largest Oil Palm (Elaeis guineensis Jacq.) Field Genebank Accessions Using Microsatellite Markers. Genet. Resour. Crop Evol. 2015, 62, 349–360. [Google Scholar] [CrossRef]
- Engelmann, F. Importance of Desiccation for the Cryopreservation of Recalcitrant Seed and Vegetatively Propagated Species. 1997.
- Nguyen, V.-A.; Nguyen, P.T.; Le, M.-A.; Bazrafshan, A.; Sisunandar, S.; Kalaipandian, S.; Adkins, S.W.; Nguyen, Q.T. A Practical Framework for the Cryopreservation of Palm Species. Vitro Cell. Dev. Biol. - Plant 2023, 59, 425–445. [Google Scholar] [CrossRef]
- Imarhiagbe, O.; Osazee, J.O.; Aiwansoba, R.O.; Shittu, O.H. In Vitro Germplasm Collection and Storage: A Review. In Vitro 2016, 1. [Google Scholar]
- Arif, M.; Ilyas, S.; Widajati, E.; Qadir, A.; Suprianto, E.; Palupi, E.R. Physical, Biochemical, and Physiological Changes on Oil Palm (Elaeis guineensis L. In Jacq) Seeds throughout Short-Term Storage. In Proceedings of the IOP Conference Series: Earth and Environmental Science; IOP Publishing, 2024; Vol. 1308; p. 012012. [Google Scholar]
- Gomes, H.T.; Machado, L.G.; Bartos, P.M.C.; Balzon, T.A.; Da Silva Costa, F.H.; Azevedo, V.C.R.; Cardoso, I.M.D.A.S.; De Souza, A.L.X.; Inglis, P.W.; Scherwinski-Pereira, J.E. Somatic Embryogenesis in Oil Palm from Immature Leaves with Emphasis on Leaf Position, Sequential Callus Re-Collection, Use of Temporary Immersion System, and Assessment of Genetic and Epigenetic Fidelity of the Resulting Clones. Plant Cell Tissue Organ Cult. PCTOC 2024, 156, 32. [Google Scholar] [CrossRef]
- Sparjanbabu, D.S.; Prathapani, N.K.; Krishna, M.S.R.; Ramajayam, D.; Susanthi, B. Differential Response of Oil Palm (Elaeis guineensis Jacq.) Genotypes on Somatic Embryogenesis and Plantlet Regeneration from Zygotic Embryo. J Appl Biol Biotechnol 2023, 11, 139–143. [Google Scholar] [CrossRef]
- Jayanthi, M.; Murali Mohan, N.; Mandal, P.K. Direct Somatic Embryogenesis and Plantlet Regeneration in Oil Palm. J. Plant Biochem. Biotechnol. 2011, 20, 249–251. [Google Scholar] [CrossRef]
- Romyanon, K.; Mosaleeyanon, K.; Kirdmanee, C. Direct-Shoot Organogenesis as an Alternative Protocol for in Vitro Regeneration of Oil Palm (Elaeis guineensis Jacq.). Sci. Hortic. 2015, 195, 1–7. [Google Scholar] [CrossRef]
- Romyanon, K.; Watana, K.; Pichakum, A.; Mosaleeyanon, K.; Kirdmanee, C. Adjustment of Medium Composition and Iso-Osmotic Potential in Direct-Shoot Organogenesis Produces True-to-Type Oil Palm (Elaeis guineensis Jacq.) Plantlets. Hortic. Environ. Biotechnol. 2017, 58, 601–612. [Google Scholar] [CrossRef]
- Chauhan, R.; Singh, V.; Quraishi, A. In Vitro Conservation Through Slow-Growth Storage. In Synthetic Seeds; Faisal, M., Alatar, A.A., Eds.; Springer International Publishing: Cham, 2019; ISBN 978-3-030-24630-3. [Google Scholar]
- Engelmann, F. Intérêt de La Cryoconservation Des Organes Végetax: Cas Des Embryons Somatiques de Palmier à Huile (Elaeis guineensis Jacq.). Int. J. Refrig. 1990, 13, 26–30. [Google Scholar] [CrossRef]
- Engelmann, F. Use of Biotechnologies for the Conservation of Plant Biodiversity. Vitro Cell. Dev. Biol. - Plant 2011, 47, 5–16. [Google Scholar] [CrossRef]
- El-Bahr, M.K.; Abd EL-Hamid, A.; Matter, M.A.; Shaltout, A.; Bekheet, S.A.; El-Ashry, A.A. In Vitro Conservation of Embryogenic Cultures of Date Palm Using Osmotic Mediated Growth Agents. J. Genet. Eng. Biotechnol. 2016, 14, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Camillo, J.; Scherwinski-Pereira, J.E. In Vitro Maintenance, under Slow-Growth Conditions, of Oil Palm Germplasm Obtained by Embryo Rescue. Pesqui. Agropecuária Bras. 2015, 50, 426–429. [Google Scholar] [CrossRef]
- Palanyandy, S.R.; Gantait, S.; Subramaniam, S.; Sinniah, U.R. Cryopreservation of Oil Palm (Elaeis guineensis Jacq.) Polyembryoids via Encapsulation–Desiccation. 3 Biotech 2020, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Spinoso-Castillo, J.L.; Pérez-Sato, J.A.; Schettino-Salomón, S.S.; Bello-Bello, J.J. An Alternative Method for Medium-Term in Vitro Conservation of Different Plant Species through Gibberellin Inhibitors. Vitro Cell. Dev. Biol. - Plant. [CrossRef]
- Hassan, M.M. In Vitro Conservation of Date Palm Somatic Embryos Using Growth-Retardant Conditions. In Date Palm Biotechnology Protocols Volume II; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Methods in Molecular Biology; Springer New York: New York, NY, 2017; ISBN 978-1-4939-7158-9. [Google Scholar]
- Konan, E.K.; Durand-Gasselin, T.; Kouadio, Y.J.; Niamké, A.C.; Dumet, D.; Duval, Y.; Rival, A.; Engelmann, F. Field Development of Oil Palms (Eloeis guineensis JACQ.) Originating from Cryopreserved Stabilized Polyembryonic Cultures. CryoLetters 2007, 28, 377–386. [Google Scholar] [PubMed]
- Prakash, K.; Kumar, K.S.; Chaudhury, R. Cryopreservation of Kernel and Zygotic Embryos of Oil Palm (Elaeis guineensis Jacq.). 2019.
- Suranthran, P.; Gantait, S.; Sinniah, U.R.; Subramaniam, S.; Alwee, S.S.R.S.; Roowi, S.H. Effect of Loading and Vitrification Solutions on Survival of Cryopreserved Oil Palm Polyembryoids. Plant Growth Regul. 2012, 66, 101–109. [Google Scholar] [CrossRef]
- Wei, Q.; Shi, P.; Khan, F.S.; Htwe, Y.M.; Zhang, D.; Li, Z.; Wei, X.; Yu, Q.; Zhou, K.; Wang, Y. Cryopreservation and Cryotolerance Mechanism in Zygotic Embryo and Embryogenic Callus of Oil Palm. Forests 2023, 14, 966. [Google Scholar] [CrossRef]
- Gantait, S.; Sinniah, U.R.; Suranthran, P.; Palanyandy, S.R.; Subramaniam, S. Improved Cryopreservation of Oil Palm (Elaeis guineensis Jacq.) Polyembryoids Using Droplet Vitrification Approach and Assessment of Genetic Fidelity. Protoplasma 2015, 252, 89–101. [Google Scholar] [CrossRef]
- Beulé, T.; Ilbert, P.; Adeoti, K.; Durand-Gasselin, T.; Dumet, D.; Engelmann, F.; Morcillo, F. Recovery of Oil Palm (Elaeis guineensis Jacq.) Somatic Embryos Cryostored for 20 Years. CryoLetters 2018, 39, 60–66. [Google Scholar]
- Suranthran, P.; Gantait, S.; Sinniah, U.R. Water Content Significantly Influences Post-Cryopreservation Survival of Air-Desiccated Oil Palm (Elaeis guineensis Jacq.) Zygotic Embryos: A Thermal and Ultrastructural Study. Ind. Crops Prod. 2023, 204, 117343. [Google Scholar] [CrossRef]
- Tandon, R.; Chaudhury, R.; Shivanna, K.R. Cryopreservation of Oil Palm Pollen. Curr. Sci. 2007, 92, 182–183. [Google Scholar]
- Kartha, K.K. Meristem Culture and Germplasm Preservation. In Cryopreservation of plant cells and organs; Crc Press, 2024; pp. 115–134.
- Norziha, A.; Marhalil, M.; Fadila, A.M.; Zulkifli, Y.; Maizura, I.; Mohd Din, A.; Rajanaidu, N.; Kushairi, A. Long-Term Storage of Oil Palm Germplasm Zygotic Embryo Using Cryopreservation. J. Oil Palm Res. 2017, 29, 541–547. [Google Scholar]
- Nishizawa, S.; Sakai, A.; Amano, Y.; Matsuzawa, T. Cryopreservation of Asparagus (Asparagus Officinalis L.) Embryogenic Suspension Cells and Subsequent Plant Regeneration by Vitrification. Plant Sci. 1993, 91, 67–73. [Google Scholar] [CrossRef]
- Matsumoto, T.; Sakai, A.; Yamada, K. Cryopreservation of in Vitro-Grown Apical Meristems of Wasabi (Wasabia Japonica) by Vitrification and Subsequent High Plant Regeneration. Plant Cell Rep. 1994, 13. [Google Scholar] [CrossRef]
- Matsumoto, T.; Sakai, A.; Yamada, K. Cryopreservation of in Vitro-Grown Apical Meristems of Lily by Vitrification. Plant Cell Tissue Organ Cult. 1995, 41, 237–241. [Google Scholar] [CrossRef]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of Nucellar Cells of Navel Orange (Citrus Sinensis Osb. Var. Brasiliensis Tanaka) by Vitrification. Plant Cell Rep. 1990, 9. [Google Scholar] [CrossRef]
- Uragami, A.; Sakai, A.; Nagai, M.; Takahashi, T. Cryopreservation by vitrification of cultured cells and somatic embryos from mesophyll tissue of asparagus Acta Hortic. 1990, 109–116. [CrossRef]
- Nishizawa, S.; Sakai, A.; Amano, Y.; Matsuzawa, T. Cryopreservation of Asparagus (Asparagus Officinalis L.) Embryogenic Cells and Subsequent Plant Regeneration by a Simple Freezing Method. 1992.
- Ishikawa, K.; Harata, K.; Mii, M.; Sakai, A.; Yoshimatsu, K.; Shimomura, K. Cryopreservation of Zygotic Embryos of a Japanese Terrestrial Orchid (Bletilla Striata) by Vitrification. Plant Cell Rep. 1997, 16, 754–757. [Google Scholar] [CrossRef]
- Towill, LeighE. Cryopreservation of Isolated Mint Shoot Tips by Vitrification. Plant Cell Rep. 1990, 9. [Google Scholar] [CrossRef]
- Watanabe, K.; Steponkus, P.L. Vitrification of Oryza Sativa L. Cell Suspensions. Cryo-Lett. 1995, 16, 255–262. [Google Scholar]
- John Martin, J.J.; Yarra, R.; Wei, L.; Cao, H. Oil Palm Breeding in the Modern Era: Challenges and Opportunities. Plants 2022, 11, 1395. [Google Scholar] [CrossRef]
- Harding, K. Genetic Integrity of Cryopreserved Plant Cells: A Review. CryoLetters 2004, 25, 3–22. [Google Scholar] [PubMed]
- Palanyandy, S.R.; Gantait, S.; Suranthran, P.; Sinniah, U.R.; Subramaniam, S. Storage of Encapsulated Oil Palm Polyembryoids: Influence of Temperature and Duration. Vitro Cell. Dev. Biol. - Plant 2015, 51, 118–124. [Google Scholar] [CrossRef]
- Al-Qurainy, F.; Khan, S.; Nadeem, M.; Tarroum, M.; Alansi, S.; Al-Ameri, A.A.; Gaafar, A.-R.Z.; Alshameri, A. Assessing Genetic Fidelity in Regenerated Plantlets of Date Palm Cultivars after Cryopreservation. Fresenius Environ. Bull. 2017, 26, 1727–1735. [Google Scholar]
- Adu-Gyamfi, R.; Wetten, A.; Marcelino Rodríguez López, C. Effect of Cryopreservation and Post-Cryopreservation Somatic Embryogenesis on the Epigenetic Fidelity of Cocoa (Theobroma Cacao L.). PLoS One 2016, 11, e0158857. [Google Scholar] [CrossRef]
- Kaity, A.; Ashmore, S.E.; Drew, R.A.; Dulloo, M.E. Assessment of Genetic and Epigenetic Changes Following Cryopreservation in Papaya. Plant Cell Rep. 2008, 27, 1529–1539. [Google Scholar] [CrossRef]
- Rohela, G.K.; Jogam, P.; Saini, P.; Sandhya, D.; Peddaboina, V.; Shekhawat, M.S. Assessing the Genetic Stability of In Vitro Raised Plants. In Commercial Scale Tissue Culture for Horticulture and Plantation Crops; Gupta, S., Chaturvedi, P., Eds.; Springer Nature Singapore: Singapore, 2022; ISBN 978-981-19-0054-9. [Google Scholar]
- Zhang, A.; Li, Y.; Yarra, R.; Li, R.; Cao, H.; Jin, L. Genome-Wide Identification of WUSCHEL-Related Homeobox Gene Family and Their Expression Analysis During Somatic Embryogenesis in Oil Palm (Elaeis guineensis). Trop. Plant Biol. 2022, 15, 55–64. [Google Scholar] [CrossRef]
- Ooi, S.-E.; Choo, C.-N.; Ishak, Z.; Ong-Abdullah, M. A Candidate Auxin-Responsive Expression Marker Gene, EgIAA9, for Somatic Embryogenesis in Oil Palm (Elaeis Guineensis Jacq.). Plant Cell Tissue Organ Cult. PCTOC 2012, 110, 201–212. [Google Scholar] [CrossRef]
- Santos, I.R.; Maximiano, M.R.; Almeida, R.F.; da Cunha, R.N.V.; Lopes, R.; Scherwinski-Pereira, J.E.; Mehta, A. Genotype-Dependent Changes of Gene Expression during Somatic Embryogenesis in Oil Palm Hybrids (Elaeis oleifera x E. guineensis). Plos One 2018, 13, e0209445. [Google Scholar] [CrossRef] [PubMed]
- Sahara, A.; Roberdi, R.; Wiendi, N.M.A.; Liwang, T. Transcriptome Profiling of High and Low Somatic Embryogenesis Rate of Oil Palm (Elaeis guineensis Jacq. Var. Tenera). Front. Plant Sci. 2023, 14, 1142868. [Google Scholar] [CrossRef]
- De Araújo Silva-Cardoso, I.M.; Gomes, A.C.M.M.; Scherwinski-Pereira, J.E. Cellular Responses of Oil Palm Genotypes during Somatic Embryogenesis Involve Participation of Procambial Cells, DNA Demethylation, and Auxin Accumulation. Plant Cell Rep. 2022, 41, 1875–1893. [Google Scholar] [CrossRef]
- Song, Y.; John Martin, J.J.; Liu, X.; Li, X.; Hou, M.; Zhang, R.; Xu, W.; Li, W.; Cao, H. Unraveling the Response of Secondary Metabolites to Cold Tolerance in Oil Palm by Integration of Physiology and Metabolomic Analyses. BMC Plant Biol. 2025, 25, 279. [Google Scholar] [CrossRef]
- Li, J.; Yang, Y.; Iqbal, A.; Qadri, R.; Shi, P.; Wang, Y.; Wu, Y.; Fan, H.; Wu, G. Correlation Analysis of Cold-Related Gene Expression with Physiological and Biochemical Indicators under Cold Stress in Oil Palm. PLoS One 2019, 14, e0225768. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, M.; Abdullah, S.N.A.; Aziz, M.A.; Namasivayam, P. A Novel CBF That Regulates Abiotic Stress Response and the Ripening Process in Oil Palm (Elaeis guineensis) Fruits. Tree Genet. Genomes 2015, 11, 56. [Google Scholar] [CrossRef]

| Explant | Survival rate | Preculturing | Loading solution | Factors | Reference |
|---|---|---|---|---|---|
| Somatic embryoids (torpedo stage) | 73% | Preculturing sucrose (0.3– 1 M) |
3% (w/v) sodium alginate and 100 mM CaCl2 | Encapsulated 3% sodium alginate and 100 mM CaCl2 Drying: 9 h, Moisture content 23% |
[28] |
| Somatic embryoids (torpedo stage) | 68% | Sucrose 0.5M for 12 h | [10 % (w/v) dimethyl sulphoxide (DMSO) plus 0.7 M sucrose] 30 minutes | Drying for 5 minutes | [35] |
| Somatic embryoids (clumps) | 33% | Sucrose 0.75 M |
28 surviving clones, 3 lost due to contamination. Regenerated after 20 years | [36] | |
| Somatic embryoids |
45% |
0.5 M 12 hours | Several loading solutions | L5 was the best loading solution while PVS2 gave the highest survival%. Exposure time was 5 minutes for the PVS2 before LN storage | [33] |
| Zygotic embryos | 80% | Moisture 0.16 g DW-1 |
- | Zygotic embryos were put in 1.5 ml cryotubes and then plunged into liquid nitrogen. | [32,37] |
| Code # | Recipe | References |
|---|---|---|
| Loading solution | ||
| L1 | [2.0 M glycerol + 0.4 M sucrose] | [41] |
| L2 | [1.5 M glycerol + 0.4 M sucrose + 5% (w/v) dimethyl sulphoxide (DMSO)] | [42] |
| L3 | [0.5 M glycerol + 0.3 M sucrose + 10% (w/v) DMSO] | [43] |
| L4 | (30% (w/v) glycerol + 15% (w/v) ethylene glycol (EG) + 15% DMSO + 0.4 M sucrose] | [44] |
| L5 | [10% (w/v) DMSO + 0.7 M sucrose] | [35] |
| Plant Vitrification Solutions | ||
| PVS | [22% (w/v) glycerol + 15% (w/v) EG + 15% (w/v) propylene gly-col + 7% (w/v) DMSO + 0.5 M sorbitol | [45] |
| PVS2 | [30% (w/v) glycerol + 15% (w/v) EG + 15% (w/v) DMSO + 0.4 M sucrose | [44] |
| PVS3 | [50% (w/v) glycerol + 50% sucrose | [46] |
| L-solution | [22% (w/v) glycerol + 30% (w/v) EG + 7% (w/v) DMSO + 15% (w/v) sucrose + 10 m CaCl2 ( | [47] |
| Towill solution | EG + 1 M DMSO + 10% PEG | [48] |
| Watanabe solution | [44.5% (w/v) DMSO + 18.7% (w/v) sorbitol for 10 min | [49] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



