
Submitted:
08 October 2025
Posted:
13 October 2025
You are already at the latest version
Abstract
The fish liver, as the main detoxification organ, is highly susceptible to xenobiotic exposure, often resulting in various hepatopathies. The cytochrome P450 system plays a central role in xenobiotic metabolism, with cytochrome P450 reductase (CYPOR) supplying the electrons required for CYP enzyme activity. This study aimed to evaluate the relationship between the ecological state of a reservoir and fish health, including CYPOR levels, through hematological, bacteriological, and histological analyses. Samples of water and fish were collected from 12 littoral sites of Lake Ladoga. Forty fish from the carp (Cyprinidae) and perch (Percidae) families were examined. Hematological smears were stained with azure eosin; bacteriological cultures were grown on multiple media; liver samples were stained with hematoxylin and eosin and Sudan III. CYPOR levels in liver homogenates were measured by immunohistochemistry. Physical and hydrochemical analyses indicated a high pollution level in the littoral zones. Isolated bacterial species were non-pathogenic but exhibited broad antibiotic resistance. Hematological evaluation revealed erythrocyte vacuolization and anisocytosis. Histological analysis showed marked fatty degeneration in hepatocytes, indicating toxic damage. CYPOR concentrations ranged from 0.3–0.4 ng/ml in healthy fish to 5–6 ng/ml in exposed specimens, confirming environmental influence on enzyme activity. These findings support CYPOR as a sensitive biomarker of environmental stress and demonstrate differences between controlled and natural conditions in aquatic organisms.
Keywords:
fish
; liver
; CYPOR
; ecology
1. Introduction
The fish liver, being the primary detoxification organ, is the first to be negatively affected by xenobiotics, leading to the development of various forms of hepatopathies [1,2,3,4]. The lipophilic properties of many antibiotics, which allow them to enter the bodies of aquatic organisms through the gills and digestive tract, facilitate rapid transmembrane transfer of the medicines and their accumulation in tissues, particularly in the liver. Agbabiaka L. A., et al. indicate in their work that the concentration of antibiotics in hepatocytes can exceed their concentration in the surrounding aquatic environment by 100-1000 times due to bioaccumulation [5].
Hepatopathies in fish can be caused by various factors, including toxic substances, infections, and genetic predispositions [6,7]. The cytochrome P450 system plays a key role in the metabolism of xenobiotics and endogenous compounds, and CYPOR provides the electrons necessary for the functioning of CYP enzymes [8,9]. Disruptions in the functioning of these systems can lead to the development of hepatopathies.
At the molecular level, the primary detoxification of antimicrobial pharmaceuticals in fish liver is carried out by the cytochrome P450, or CYP, system. The predominant isoforms in fish are CYP1A, CYP2B, and CYP3A [10]. The cytochrome P450 system includes numerous isoenzymes that participate in the biotransformation of various substances. CYPOR is a flavoprotein that transfers electrons from NADPH to CYP, which is necessary for the catalytic activity of the latter. Disruptions in the functioning of CYP and CYPOR can lead to the following consequences:
- Toxin accumulation. Insufficient CYP activity can lead to a slower metabolism of toxic substances, contributing to their accumulation in the liver and the development of hepatopathies.
- Oxidative stress: CYPOR and CYP are involved in the generation of reactive oxygen species (ROS). Disruptions in their function can lead to increased ROS formation, which causes oxidative stress and liver cell damage.
- Inflammatory processes: Metabolic products formed with the participation of CYP can cause inflammatory reactions that contribute to the development of hepatopathies.
Furthermore, the activity of cytochrome isoforms increases significantly with exposure to antibiotics. The metabolism of most antimicrobial drugs is accompanied by the formation of reactive intermediates capable of inducing oxidative stress [10].
Research shows that exposure to various toxicants, such as polycyclic aromatic hydrocarbons (PAHs), polychlorinated biphenyls (PCBs), and pesticides, can lead to changes in the expression and activity of CYP and CYPOR in fish.
Liu et al., in experiments on common carp (Cyprinus carpio), showed that exposure to subtherapeutic doses of oxytetracycline leads to a significant increase in the concentration of malondialdehyde and a decrease in the activity of superoxide dismutase, catalase, and glutathione peroxidase in the liver, which indicates a disturbance in the pro- and antioxidant balance [11]. Reactive oxygen species formed as a result of antibiotic metabolism cause lipid peroxidation of hepatocyte membranes, which is accompanied by a violation of selective permeability, damage to membrane proteins, and the release of intracellular enzymes, such as ALT and AST, into the bloodstream, which is confirmed by the results presented in the works of Chen H., et al., 2015 and Allameh A., et al., 2023 [12,13].
Based on the study by Larina S.N. et al., 2009 on the hepatocyte cell lines of the silver crucian carp (Carassius auratus), it can be assumed that antimicrobial drugs are capable of modulating gene expression in fish hepatocytes through interaction with nuclear receptors such as the pregnane X receptor (PXR), the constitutive androstane receptor (CAR), and the aryl hydrocarbon receptor (AhR). Rodríguez -Mozaz S et al., 2016 found that sulfonamides induce CYP1A gene expression through AhR activation, leading to increased formation of toxic metabolites and increased oxidative stress. In the works of Ambili T. R., et al., 2013 it was noted that macrolide antibiotics stimulate the production of proinflammatory cytokines (IL-1β, TNF-α) by Kupffer cells, which increases the influx of neutrophils and lymphocytes into the liver of fish, and also activate the NF-κB and AP-1 signaling pathways in hepatocytes, enhancing the expression of inflammatory response genes [14,15,16,17].
Long-term exposure to antimicrobials activates programmed cell death processes in the fish liver. According to Paramonov, S.G., et al., 2022 chronic exposure to tetracyclines in carp leads to activation of both the external (via Fas receptors) and internal (mitochondrial) apoptotic pathways [18].
This induces hypermethylation of the promoter regions of antioxidant defense genes, leading to a decrease in their expression and an increase in the sensitivity of hepatocytes to oxidative stress. Ding L., et al., 2017 showed that antimicrobials can modify histone acetylation patterns in the nuclei of fish hepatocytes, affecting the transcriptional activity of genes involved in xenobiotic metabolism [19].
Long-term exposure to antimicrobials also leads to the development of liver fibrosis in fish. In the works of Au-Yeung C. et al., 2025 and Andreeva N.L. et al., 2017, it has been shown that prolonged exposure of carp to sublethal concentrations of chloramphenicol activates Ito cells, which transform into myofibroblasts and synthesize extracellular matrix components, primarily collagen types I and III [20,21].
Considering the above, the goal was set to evaluate codependence between the ecological state of a reservoir and its coastal zone and fish health and its CYPOR levels by hematological, bacteriological, and histological analyses.
The main objectives of the study:
- To study the ecological state of the reservoir and coastal zone to assess the fish habitat.
- To conduct hematological studies of the fish to assess their clinical status.
- To conduct a bacteriological and histological examination of the fish in order to assess their clinical status.
- To determine CYPOR levels and draw conclusions about the impact of the habitat on them.
2. Materials and Methods
The studies were conducted at the Department of Pharmacology and Toxicology of the Federal State Budgetary Educational Institution of Higher Education “St. Petersburg State University of Veterinary Medicine” (St. Petersburg, Russia) and at the Laboratory of Innovative Recombinant Proteomics Preparations of the Experimental Pharmacology and Modeling of Living Systems Department of the Federal State Budgetary Scientific Institution “All-Russian Veterinary Research Institute of Pathology, Pharmacology and Therapy” (Voronezh, Russia).
For the CYPOR study, 40 fish specimens from the carp (Cyprinidae) and perch (Percidae) families were selected using a blind randomization method. For the main study, aquatic organisms were collected from Ladoga Lake, specifically from its shallow southern littoral region, which is most vulnerable to toxic impacts (Figure 1, Table 1). A total of 1,360 specimens of carp and perch fish caught in the Shlisselburg, Volkhov, and Svir Bays were examined. Ladoga Lake water is characterized by low mineralization, a hydrocarbonate-calcium composition, and a slight excess of sulfate ions compared to chloride ions. An assessment of the physical and hydrochemical parameters of water in three areas of Ladoga Lake was carried out using the Aqua Troll 500 multiparameter probe (In-Situ, Fort Collins, USA). We studied the following parameters: temperature (T,℃), mineralization (Mn), concentration of dissolved oxygen (O₂), electrical conductivity (EC), oxidation-reduction potential (ORP), and hydrogen index (pH).
Fish were studied in the littoral zones and adjacent waters of three bays (Shlisselburg, Volkhov, and Svirskaya). For analysis, we selected 10 adult specimens of each species (bream (Abramis brama), perch (Perca flavescens), roach (Rutilus rutilus), and pike-perch (Sander lucioperca)) from waters nearby each research station.
Completely healthy specimens of carp (Cyprinidae) and perch (Percidae) families, maintained under standardized, stable conditions, were used as control specimens. The fish were kept in 150-200 L glass aquariums at a stocking density of 1.0-1.5 kg/m³, with constant aeration and mechanical-biological filtration. A natural light regime was established (10 hours of fluorescent lighting and 14 hours of darkness). The water temperature was maintained at 20-24 °C. The fish were fed a balanced diet at a daily feeding rate of 3% of their biomass, divided into three feeding times (7:00, 12:00, and 17:00) to prevent exposure to additional stressors. To maintain water quality, 30% of the aquarium water volume was changed daily. Uneaten food and feces were pumped out daily. In case of animal death, microbiological and molecular genetic analysis of pathological material from dead animals was performed.
Fish manipulations were carried out in accordance with the bioethical guidelines described in the Guide for the Care and Use of Laboratory Animals (National Research Council, 2011).
2.1. Hematological Examination
Peripheral blood samples from common carp were collected using the method of E.A. Petrova [22] from the caudal vein at the point of intersection of the line running from the posterior edge of the anal fin. At the beginning, the scales were separated, and the puncture site was treated with a 70% alcohol solution. After the puncture, the syringe plunger was pulled back, slowly advancing the needle at a 45° angle toward the head until it touched the spinal column. The inclined needle was rotated until blood appeared in the syringe cannula. Blood was collected, the needle was removed, and the puncture site was treated with an antiseptic.
The prepared samples were stained according to the standard method [23,24]. After drying for 4 minutes, the smears were fixed in Nikiforov’s solution (1:1, methyl alcohol: diethyl ether) for 2 minutes, then stained with methylene blue eosin according to May-Grünwald for 4 minutes. The slides were then washed and dried, followed by staining with azure eosin according to Romanovsky-Giemsa for 10 minutes. The resulting preparations were again washed in containers with distilled water and dried for 30 minutes.
2.2. Bacteriological and Histological Studies
To determine the qualitative and quantitative composition of the microbiota of the studied hydrobandages, cultures were performed on standard media for the accumulation and selection of microorganisms (meat-peptone agar, Endo agar, mannitol salt agar, blood agar (meat-peptone agar with 5% sheep red blood cells), Kitt-Tarozzi, Wilson-Blair, Blaurock, and MRS (for the isolation of lactobacilli)). To isolate facultative anaerobic microorganisms, incubation of the cultures was carried out in a thermostat at 37 °C under aerobic conditions. To isolate anaerobic flora, an anaerobic jar was used. To detect microscopic fungi, samples were placed in a thermostat at 28 °C. After incubation for 18-72 hours, colonies of each microorganism species were counted. The cultural, morphological, and biochemical properties of the isolated microorganisms were studied using standard methods. The species of bacteria were determined using the reference book “Microbiological Diagnostics of Animal Diseases” (2005) [25].
To conduct morphological studies, samples of liver and muscle tissue of the common roach were taken and fixed in a 10% neutral formalin solution. The tissue samples were then dehydrated in ethyl alcohol solutions of increasing concentration and embedded in paraffin. Sections of 3-5 µm thickness were sliced from paraffin blocks, stained with Carazzi hematoxylin and an aqueous-alcoholic solution of eosin, and examined under a light microscope.
For further tissue analysis, carp liver samples were collected and histological preparations were made. These were prepared by freezing the native tissue and then staining with Sudan III and scarlet. This staining method is used to highlight neutral lipids. The resulting sections are first soaked in 50-70% ethanol for 2-3 minutes. Then, they are kept for 15-30 minutes in a pre-filtered saturated alcohol solution (70% ethanol) of the combined dye. The sections are then rinsed in 50-70% ethanol. Rinsing is done in distilled water. The sections are then soaked in hematoxylin solution for 3-5 minutes, followed by rinsing in tap water. Finally, the sections are embedded in glycerogel.
Histological examination of the aquatic animal liver was performed using digital microscopy using a Carl Zeiss Axioscope light microscope (Carl Zeiss Meditec, Jena, Germany). Photos of histological sections were taken using a Levenhuk microscope camera (Levenhuk, Moscow, Russia) and ToupView software (ToupTek, Zhejiang, China).
Histomorphometric parameters of the slides were assessed using OMERO v5.6 (University of Dundee & Open Microscopy Environment, Dundee, United Kingdom), QuPath v0.5.1 [26], and Orbit Image Analysis v3.64 image processing software [27]. Data conversion and visualization were performed using the publicly available cluster v2.1.4 package.
2.3. Studies to Determine CYPOR Concentrations in Liver Homogenates
To study CYPOR levels, selected liver samples were placed in a special tissue homogenization suspension medium (pH 7.4; Tris-HCl buffer supplemented with 0.25 M sucrose and a small amount of ethylenediaminetetraacetic acid and sodium salt) to bind divalent metal ions and glutathione to prevent lipid peroxidation.
Next, using a Stegler S10 homogenizer (Stegler, China), homogenates were obtained and analyzed using the StatFax 3200 enzyme immunoassay method (Awareness technology, Palm City, USA) using the Cytochrome P450 Reductase (CPR) Activity Assay Kit (Elabscience, Houston, USA) according to the instructions for the kit [28].
2.4. Statistical Analysis
The statistical analysis included calculation of the mean (M), median, standard deviation (SD), standard error (SE), and coefficient of variation. The standard deviation (SD) was calculated using the formula:
where xi – i index; — arithmetic mean; n – number of samples.
The coefficient of variation (CV) was calculated using the formula:
where SD – standard deviation; — arithmetic mean.
The standard error (SE) was calculated using the formula:
where SD – standard deviation; n – number of samples.
Post hoc comparisons were performed using the Student’s t-test for independent samples, taking into account the Bonferroni correction. Values for which p < 0.05 were considered significantly different.
3. Results
3.1. Ecological Status
The littoral zone of Lake Ladoga’s southern waters is a spawning and nursery ground for the lake’s main fish species. The dominant negative impacts are numerous anthropogenic factors, including both organic and inorganic toxicants. Previous studies have shown that concentrations of high-molecular-weight compounds, particularly polychlorinated biphenyls, significantly exceeded MACs in these waters. [29] The concentration of organochlorine compounds α- and γ-HCH in almost every point of Lake Ladoga and its tributaries exceeds the maximum permissible level. Lindane (γ-HCH), a hazardous xenobiotic, has also been widely distributed throughout Lake Ladoga.
Heavy metals and assessment of physical and hydrochemical parameters of water in three areas of Lake Ladoga were carried out in the summer period from 2022 to 2025, and the results are presented in Table 2.
The mineralization index ranged from 0.030 to 0.122 (station V1). Dissolved oxygen levels ranged from 9.5 to 11.7 mg/L, with a lakewide average of 10.4 mg/L. The oxygen saturation percentage remained high, averaging 102.2%. The oxidation-reduction potential (ORP) ranged from 43 to 156 mV, with a monthly average of 96.5, and the pH varied from 7.4 to 8.4, with a monthly average of 7.6. The obtained results are consistent with the data reflected in the report of the Committee on Natural Resources of the Leningrad Region for the period under study.
The fish study was conducted in the littoral zones and adjacent waters of three bays (Shlisselburg, Volkhov, and Svir). For analysis, we selected 10 specimens of each species (bream, perch, roach, pike-perch) of sexually mature age from each research station living in the surveyed waters.
Upon external examination, the majority of the fish specimens studied in the southern part of the lake had their body surface covered with a thin layer of transparent mucus. The mucus had no unusual odors or cloudy color. When assessing the gills, their appearance was taken into account: some specimens, especially in the Volkhov Bay of Lake Ladoga, showed anemia and hemorrhages of the gill tissue. If profuse hemorrhaging was present in the gills, a musty and sour odor was noted. The presence of mechanical damage and scale coverage was also assessed. In most cases, the scales were intact and tightly attached to the body. There were specimens of fish with mechanical damage to the body, and bruises were noted. The corneal transparency and the position of the eyeball relative to the orbital level were assessed. Freshly caught fish had clear corneas and convex eyes. The abdomen was normal in most cases. The fish specimens also had a sunken, pale pink anus. The fish were firm, with a shiny, cross-sectional color characteristic of each species. The odor was determined by a cooking test and was consistent with fresh fish. Dangerous zoonotic helminths were not detected in the muscle tissue of fish.
Some fish showed poor body condition, soft tissue lesions in the fins, and other changes. This spring study revealed that across all surveyed waters, the lesions in fish were consistent and associated with hemodynamic disturbances and foci of degeneration in the liver.
Liver changes were associated with foci of degeneration and changes in organ color, usually localized, and changes in consistency. Gallbladder filling, changes in color, and consistency of the contents were noted. In the heart, anemia and muscle flaccidity were noted. The kidneys were enlarged, filled with blood, edematous, and sometimes with the formation of stones in the renal tubules. In the gastrointestinal tract, swelling of the mucous membrane, areas of hyperemia, and accumulation of mucous contents were noted. Vascular injection was observed in the gonads. In the swim bladder, blood-filled vessels and rare diffuse hemorrhages were observed. Changes in the brain were associated with vascular injection and sometimes with pinpoint hemorrhages. It’s also important to note that the fish were in good condition outside the littoral zone.
A summer fish study revealed a virtually identical pattern of intoxication, both in percentages and in the severity of the pathological process. Fish with mild injuries predominated, and the pattern of gill tissue damage was milder. It was also noted that older fish and those living near the bottom were more severely affected. Using bleak as an example, it was shown that pelagic fish were less affected, and the intoxication was mild.
Fish caught at monitoring stations showed mild intoxication. During the autumn study of Shlisselburg Bay, the condition of the fish was satisfactory. The pattern of intoxication was milder, with fish showing mild symptoms predominating. While in the spring, fish with poor body condition and external signs of intoxication were observed, such cases were isolated in the fall. Thus, signs of intoxication in the spring were associated with wintering and the increase in toxins in the form of pollutants entering the water via surface runoff and precipitation. Table 3 presents the pathological examination data for fish from the Volkhov Bay of Ladoga Lake.
In this area, fish were collected both in the littoral zone and beyond. As can be seen from Table 3, various aquatic fish species were analyzed in various seasons.
Thus, the southern waters of Lake Ladoga, and especially the shallow littoral zone, judging by the state of fish, are the most polluted waters. The highest level of pollution was recorded in the Volkhov Bay of the lake. Pathological manifestations of chronic intoxication in fish from the southern part of the lake are shown in Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8 and Figure 9.
A pathomorphological study of various fish species in the southern part of Lake Ladoga showed that the development and manifestation of pathological changes is characteristic of a chronic process.
The most pronounced lesions were detected in the gill apparatus. Histological preparations show changes in the architecture of the epithelial and proper gill plates. Damage to the blood vessel walls was visible in necrotic areas. Necrosis with epithelial tissue disintegration was observed, or fusion of secondary lamellae to form solid layers was observed in the area of the gill arches.
Changes in the liver were associated with the accumulation of pigmented lesions and lymphocytes among hepatocytes, which were sometimes vacuolated, often with minor pigmented nuclei, a sign of degenerative changes in the organ. Clusters of pigment cells and lymphocytes were also found in the liver tissue. Fish with hepatocyte discomplexation, proliferation of connective tissue, and accumulation of fibroblasts around the bile ducts and blood vessels were noted.
Focal and diffuse hemorrhages and edema of the connecting glomeruli and convoluted tubules were found in the kidneys of the fish. Vacuolar granular dystrophy was noted in the epithelium of the renal tubules, which was altered, i.e., the epithelium was altered, with numerous vacuoles and granularity within the cells, which causes hemodynamic and mechanical filtration impairment.
In the hematopoietic tissue of the kidney, a large number of hemosiderin granules are observed, and in the connective tissue of varying degrees of decay, it is overfilled with protein lumps. Vacuolar, often granular, degeneration of the epithelial cells of the urinary tubules was also observed. Vacuolization of the epithelium of individual convoluted tubules was observed in fish from both Volkhov and Svir Bays. The most pronounced protein degeneration of the convoluted tubules of the kidney is observed in fish from the Volkhov Bay.
Fish brain tissue is less reactive than tissue in other organs and is capable of recovery. The germinal tissue of fish turned out to be more resistant to toxicological effects, which is an indication of a high degree of reliability in the development and functioning of the reproductive system.
Pathological examinations of the water bodies of the Lake Ladoga basin confirm this, while damage to other organs of the fish was obvious and clearly visible. Histomorphological changes in the internal organs and gill tissue of fish from the southern waters of Lake Ladoga are shown in Figure 10, Figure 11, Figure 12, Figure 13 and Figure 14.
Studies have shown that the identified histomorphological abnormalities in the internal organs of fish and gill tissue in the southern waters of Lake Ladoga are widespread and are clear evidence of the impact of toxicological factors on the ichthyofauna of this water area, which is confirmed by the results of pathological and histological studies and the presence of visual manifestations of chronic intoxication.
3.2. Bacteriological Examination
During the bacteriological examination of pathological material isolated from fish, Aeromonas spp. and Enterococcus spp., which belong to opportunistic microflora, were isolated (Table 4).
Aeromonas spp. were sensitive to four drugs, including chloramphenicol, gentamicin, polymyxin, and norfloxacin (100%). Enterococcus spp. were sensitive to five drugs, including ampicillin, amoxicillin, chloramphenicol, gentamicin, and doxycycline (100%).
Aeromonas spp. were detected in all fish studied, indicating their widespread distribution in this ecosystem. Bacteria of this genus are typical inhabitants of freshwater bodies, but some species can cause furunculosis, ulcers, or septicemia in fish under certain conditions.
Enterococcus spp. were detected in 55.6% of samples, primarily in intestinal contents. This is normal, as these bacteria are part of the normal intestinal microflora of many animals. However, the high level of antibiotic resistance in strains isolated from wild fauna is alarming.
Staphylococcus spp. were detected in 55.6% of samples, primarily on the gills. Staphylococci rarely cause disease in fish but can be an indicator of external contamination.
The conducted study of water samples revealed favorable microbiological characteristics, indicating the suitability of the reservoir for fisheries activities. The results demonstrate compliance with the sanitary and epidemiological standards established by Guidelines 4.2.1884-04.
3.3. Hematological Examination
Microscopic analysis of peripheral blood smears from the fish studied (Figure 15) revealed significant ultrastructural abnormalities in the erythrocytes. In the studied cell population, the percentage of cytoplasmic vacuolization was 45.35%, indicating impaired degradation and recycling of cellular components. Vacuoles were diffusely distributed due to dysfunction of the lysosomal apparatus and the accumulation of lipid inclusions. Vacuolization of nuclei was noted, which is characteristic of impaired nuclear-cytoplasmic transport. The coefficient of variation in erythrocyte diameter was 1.2, significantly exceeding the threshold value, indicating the presence of anisocytosis. Asynchrony of erythroid maturation may be a consequence of induced disruption of membrane structures and mitochondrial activity of erythroblasts against the background of contamination of the reservoir with antimicrobial drugs.
When assessing the cellular diversity of the peripheral blood of the studied fish, it is important to note that the high proportion of phagocytic cells, which have a high migration capacity and actively absorb not only bacteria but also the products of cell and tissue decay (profile metamyelocytes and monocytes) in the leukogram, exceeded reference values, which reflects the intensification of the elimination of cellular destruction products induced by toxicants. The presence of lymphopenia combined with neutrophilia in blood smears is a hematological marker of chronic intoxication of aquatic organisms and indicates prolonged toxic exposure to aquatic contaminants.
3.4. Histological Examination
Histological examination of the liver of healthy fish (Figure 16 A, B) revealed the presence of classical organ architecture. Hepatocytes are characterized by a typical polygonal shape with distinct intercellular boundaries, and the cytoplasmic component is homogeneous. Liver cell nuclei have a spherical configuration with a uniform distribution of chromatin. The presence of binucleated cells indicates the regenerative activity of the organ. The cytoplasm shows moderate granularity. Sinusoidal spaces are clearly differentiated between the hepatic trabeculae. Clearly structured portal triads are visible in the peripheral areas of the liver lobules.
Histological examination of the liver in the studied fish (Figure 16 C, D) revealed significant disorganization of the classical trabecular pattern of the liver parenchyma with disruption of the radial orientation of hepatocytes relative to the central veins. Marked disintegration of tissue architecture is observed, accompanied by disruption of the normal histotopographic relationships between cellular elements. Hepatocytes exhibit a number of pathological changes, manifested by an increase in cellular volume, a decrease in the optical density of the cytoplasm, and an indistinct cytolemma, indicating a disruption of osmotic homeostasis and intracellular fluid accumulation. In the centrilobular regions of hepatocytes, pronounced degeneration is observed with total vacuolization of the cytoplasm, accounting for 56.4% of the cell area. Lipofuscin inclusions are found in the cytoplasm of some cells, a sign of chronic liver tissue damage due to antimicrobial contamination. The presence of binucleated cells indicates the regenerative activity of the organ. The nuclear apparatus of liver cells undergoes significant changes, including a shift of nuclei to the periphery of the cells due to compression by cytoplasmic inclusions, as well as chromatin heterogeneity with areas of condensation.
To confirm the results of staining with hematoxylin Carazzi and aqueous-alcoholic eosin solution, a histological examination was performed using specific staining with Sudan III and scarlet red. Compared to healthy fish (Figure 17A), the livers of experimental aquatic organisms (Figure 17B) exhibited pronounced pathomorphological changes in the liver parenchyma consistent with fatty degeneration due to toxic damage, as evidenced by the abundance of red-orange staining. Discomplexation of the liver beams and disruption of the trabecular structure and boundaries between cells are noted, which is a consequence of hepatocyte hypertrophy due to excessive accumulation of lipids. An increase in the karyocytoplasmic ratio, nuclear hyperchromia, and signs of karyopyknosis in some cells are observed, indicating the initial stages of necrobiotic changes. According to research by Kudersky, L.A., 2013, large lipid vacuoles form in the cytoplasm of hepatocytes during aquatic contamination, which is clearly visible in the presented specimen. Such changes are accompanied by disruption of the metabolic and detoxifying functions of the liver, which significantly reduces the fish’s resistance to adverse environmental factors and can lead to their death [29].
The presented three-dimensional graphical images of the chromatic dispersion coefficient of lipid infiltration clearly demonstrate the structural differences between normal liver tissue from healthy fish and the liver with fatty degeneration of the studied aquatic organisms (Figure 18). Figure 18A shows a predominantly uniform profile with dominant green staining, consistent with the normal histoarchitecture of healthy liver tissue. Figure 18B demonstrates a significant predominance of orange-red coloration and the formation of numerous tall, irregularly shaped peaks. According to the research of Zahran E., et al., 2025, who used similar visualization methods, the intensity of red coloration correlates with the degree of lipid accumulation [30].
Quantitative analysis of the presented surfaces indicates a significant increase in peak height and density in the experimental fish, which, according to Pramanik S. and Biswas J.K., 2024, reflects the disorganization of the beam structure of the liver and an increase in hepatocyte volume due to the accumulation of lipid vacuoles. Increased surface relief also indicates inflammatory changes, often associated with fatty degeneration, which is confirmed in the study of Rana S.V.S., 2024 [31,32].
The presented heat map of the chromatic dispersion coefficient distribution of lipid infiltration (Figure 19) complements and refines the data described above, providing a two-dimensional topographic visualization of the pathological process. Healthy fish (Figure 19A) exhibit a predominantly uniform green background, with blue markers representing lipid droplets present in small quantities in several discrete foci. In the studied fish (Figure 19B), a significant increase in the number of blue markers diffusely distributed over the entire area of the tissue section is observed compared to healthy aquatic organisms, indicating extensive fatty infiltration.
Quantitative assessment suggests that the liver lesion area in the experimental fish constitutes more than 80% of the tissue, which, according to the classification of Pramanik S. and Biswas J.K., 2024, corresponds to a severe degree of steatosis [31].
CYPOR Examination
In healthy aquatic organisms, CYPOR levels ranged from 0.3 to 0.4 ng/ml, while in the test specimens, they ranged from 5 to 6 ng/ml, confirming the influence of environmental factors on CYPOR levels. This significant (15–20-fold) increase in CYPOR activity/expression in the experimental aquatic organisms indicates a strong induction of this enzyme by the studied factors. Since CYPOR is an obligate electron donor for the cytochrome P450 system, such induction can significantly affect the metabolism of xenobiotics and endogenous compounds, potentially altering their biotransformation rate, toxicity, or pharmacological effect.
The obtained data suggest that exposure to these factors activates specific signaling pathways (e.g., those mediated by AhR, PXR, or CAR receptors) that regulate the transcription of the POR gene. A significant increase in CYPOR levels may indicate an adaptive organism response aimed at enhancing detoxification but also carries risks of disruption of metabolic homeostasis, activation of procarcinogens, or altered drug efficacy.
Although the observed difference is quantitatively large, further research will help to fully understand its physiological consequences. The key areas:
- identification of specific factors (or their combination) responsible for induction;
- studying changes in the activity of CYPOR-dependent cytochrome P450 isoforms;
- assessment of the functional outcomes of such induction in vivo (e.g., clearance rate of specific substrates, formation of toxic metabolites, oxidative stress).
4. Discussion
The data obtained in the present study demonstrate a dramatic contrast in NADPH-cytochrome P450 reductase (CYPOR) levels between fish maintained under controlled vivarium conditions and fish living in the wild under environmental pressure.
The observed 20-fold increase in CYPOR levels in fish from a natural population compared to their vivarium counterparts (conventionally taken as the baseline level under “ideal” conditions) is not simply a statistically significant difference but rather indicates a profound biochemical restructuring in response to environmental conditions.
CYPOR is an obligate electron donor for the vast majority of microsomal cytochrome P450 (CYP) isoforms, the central system for xenobiotic detoxification and endogenous compound metabolism [8,9]. The observed extreme CYPOR induction in fish under natural conditions distinctly indicates a powerful activation of the entire cytochrome P450 system in response to the complex effects of environmental factors.
This scale of induction (20-fold) goes beyond the scope of normal physiological fluctuations and reflects the chronic adaptive response of the organism to the constant presence in the environment of a wide range of xenobiotics (polycyclic aromatic hydrocarbons (PAHs), polychlorinated biphenyls (PCBs), dioxins, heavy metals, pesticides, etc.), as well as other stressors (hypoxia, temperature fluctuations, changes in salinity), characteristic of real, often anthropogenically disturbed ecosystems. [3,7,10,17,30]
Elevated CYPOR levels are necessary to ensure adequate electron flow to induced CYP isoforms (such as CYP1A, which is known to increase sharply in response to AhR-binding pollutants). [10,14] This allows fish to metabolize and eliminate toxic substances more efficiently, increasing their chances of survival in polluted environments.
Paradoxically, increased CYP system activity mediated by CYPOR induction also carries significant risks. Many xenobiotics are procarcinogens or protoxicants, requiring CYP biotransformation into active forms to exert their toxicity [8,10,14,32]. Consequently, high CYPOR activity can contribute to the increased formation of reactive intermediate metabolites (epoxides, quinones, and free radicals) that cause DNA damage, oxidative stress, cell necrosis, and apoptosis, potentially leading to decreased population viability, the development of pathologies (including neoplasms), and reproductive dysfunction [13,29,31].
The CYP system, controlled by CYPOR, is involved in the biosynthesis and catabolism of steroid hormones, bile acids, fat-soluble vitamins, and signaling molecules. Extreme CYPOR induction can disrupt the delicate balance of these endogenous processes, leading to endocrine dysfunctions that are particularly critical for fish growth, development, reproduction, and behavior.
Maintaining high levels of expression and activity of such a large protein as CYPOR, as well as associated CYP isoforms, requires significant energy resources from the body. This can lead to a redistribution of energy from growth and reproduction to detoxification, a classic trade-off under stress.
Such pronounced CYPOR induction is highly likely mediated by the activation of specific nuclear receptors sensitive to xenobiotics. These receptors, when bound to pollutants, act as transcription factors, directly regulating the expression of the POR gene (encoding CYPOR) and the genes of many CYP isoforms.
5. Conclusions
Our results strongly support the idea that CYPOR itself is a highly sensitive biomarker of the complex impacts of environmental stress on aquatic organisms. Its induction reflects not only the presence of individual pollutants but also the overall “metabolic load” on the organism under real-world conditions. Moreover, being an obligatory component of the system, the CYPOR level can serve as an integral indicator of its overall metabolic capacity, potentially even more stable or informative than the levels of individual, highly inducible CYP isoforms, which are subject to more complex regulation.
Our study data illustrate the fundamental difference between the metabolic status of organisms under strictly controlled laboratory conditions and in the real environment. This highlights the importance of conducting research on natural populations to adequately assess environmental risks.
The twenty-fold difference in CYPOR levels is a worrying indicator of the intensity of environmental pressure experienced by the studied fish population in their natural environment. This suggests that the fish are functioning in a state of constant physiological stress, devoting significant resources to counteracting pollution.
- While our study clearly established the presence of large-scale CYPOR induction, further research is required to fully understand the consequences:
- Identification of the specific pollutants (or combinations thereof) that contribute primarily to the observed induction.
- Concurrent measurement of the activity of key CYP isoforms (especially CYP1A and CYP3A) to assess the functional consequences of elevated CYPOR levels.
- Study of the temporal dynamics of CYPOR induction and its reversibility with decreasing pollution levels.
Author Contributions
Conceptualization, V.P.; methodology, O.P.; software, E.S.; validation, V.P., O.P. and E.M.; formal analysis, E.M.; investigation, E.S. and A.R.; resources, E.M. and A.R..; data curation, V.P..; writing—original draft preparation, V.P., O.P. and E.M.; writing—review and editing, V.P.; visualization, E.S.; supervision, V.P.; project administration, V.P.; funding acquisition, V.P.
Funding
The study supported by the grant of the Russian Science Foundation No. 24-76-10011, https://rscf.ru/project/24-76-10011/.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committees at the Laboratory of Innovative Recombinant Proteomics Preparations of the Experimental Pharmacology and Modeling of Living Systems Department of the Federal State Budgetary Scientific Institution “All-Russian Veterinary Research Institute of Pathology, Pharmacology and Therapy”, Federal State Budgetary Educational Institution of Higher Education “Saint-Petersburg State University of Veterinary Medicine”.
Data Availability Statement
Data is contained within the article. The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors extend their sincere thanks to the members of Laboratory of Innovative Recombinant Proteomics Preparations of the Experimental Pharmacology and Modeling of Living Systems Department of the Federal State Budgetary Scientific Institution “All-Russian Veterinary Research Institute of Pathology, Pharmacology and Therapy” and Federal State Budgetary Educational Institution of Higher Education “Saint-Petersburg State University of Veterinary Medicine” for their help and support.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| CYPOR—cytochrome P450 reductase; |
| CYP—cytochrome P450; |
| NADPH—reduced nicotinamide adenine dinucleotide phosphate; |
| ROS—reactive oxygen species; |
| PAHs—polycyclic aromatic hydrocarbons; |
| PCBs—polychlorinated biphenyls; |
| ALT—Alanine Aminotransferase; |
| AST—aspartate aminotransferase; |
| PXR—pregnane X receptor; |
| CAR—constitutive androstane receptor; |
| AhR—aryl hydrocarbon receptor; |
| Mn—mineralization; |
| EC—electrical conductivity; |
| ORP—oxidation-reduction potential; |
| M—mean; |
| SD—standard deviation; |
| SE—standard error; |
| CV—coefficient of variation; |
| MAC—Maximum Allowable Concentration. |
References
- Popova, O.; Ponamarev, V. The prevalence of hepatopathy in productive animals and aquaculture objects. In Proceedings of the International Conference on Ensuring Sustainable Development: Ecology, Energy, Earth Science and Agriculture (AEES 2023), Moscow, Russia, 21–22 December 2023; EDP Sciences: Les Ulis, France, 2024; p. 4017. [Google Scholar] [CrossRef]
- Feist, S.W.; Stentiford, G.D.; Kent, M.L.; Ribeiro Santos, A.; Lorance, P. Histopathological assessment of liver and gonad pathology in continental slope fish from the northeast Atlantic Ocean. Marine Environmental Research 2015; 106, 42–50. [CrossRef]
- Fuentes-Lopez, K.; Olivero-Verbel, J.; Caballero-Gallardo, K. Presence of Nematodes, Mercury Concentrations, and Liver Pathology in Carnivorous Freshwater Fish from La Mojana, Sucre, Colombia: Assessing Fish Health and Potential Human Health Risks. Archives of Environmental Contamination and Toxicology 2025, 88, 189–209. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.A.; Aktar, M.N.A.S.; Halim, K.A.; Hasanuzzaman, K.M.; Islam, Md.A. Health and disease status of cultured Gulsha (Mystus cavasius) at Mymensingh region of Bangladesh. Research in Agriculture, Livestock and Fisheries 2020, 7, 139–151. [Google Scholar] [CrossRef]
- Agbabiaka, L.A.; Onwuzuruigbo, F.O.; Jimoh, O.A. Threat to fish food safety in Nigeria: Role of antimicrobial usage and resistance in aquaculture. Aquaculture Reports 2025, 40, 102643. [Google Scholar] [CrossRef]
- Liang, X.; Zha, J.; Martyniuk, C.J.; Wang, Z.; Zhao, J. Histopathological and proteomic responses in male Chinese rare minnow (Gobiocypris rarus) indicate hepatotoxicity following benzotriazole exposure. Environmental Pollution 2017, 229, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Walter, G.L.; Jones, P.D.; Giesy, J.P. Pathologic alterations in adult rainbow trout, Oncorhynchus mykiss, exposed to dietary 2,3,7,8-tetrachlorodibenzo-p-dioxin. Aquatic Toxicology 2000, 50, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Ponamarev, V. Methods for determining cytochrome P450 reductase in biological fluids of animals. BIO Web of Conferences 2025, 160, 01030. [Google Scholar] [CrossRef]
- Ponamarev, V.S.; Popova, O.S.; Ukrainskaya, O.A. The role of the cytochrome system in the biotransformation of xenobiotics and drugs (review). Agrarian Science of the Euro-North-East 2025, 26, 21–39. [Google Scholar] [CrossRef]
- Gulyaeva, L.F.; Grishanova, A.Y.; Gromova, O.A.; Slynko, N.M.; Vavilin, V.A.; Lyakhovich, V.V. Microsomal monooxygenase system of living organisms in environmental biomonitoring. Analytical Review, Series “Ecology” 1994, 100. (Siberian Branch of the Russian Academy of Sciences).
- Liu, Y.; Zhang, J.; Gao, B.; Feng, S. Combined effects of two antibiotic contaminants on Microcystis aeruginosa. Journal of Hazardous Materials 2014, 279, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, S.; Xu, X.R.; Liu, S.S.; Zhou, G.J.; Sun, K.F.; Zhao, J.L.; Ying, G.G. Antibiotics in typical marine aquaculture farms surrounding Hailing Island, South China: occurrence, bioaccumulation and human dietary exposure. Marine Pollution Bulletin 2015, 90(1–2), 181–187. [CrossRef]
- Allameh, A.; Niayesh-Mehr, R.; Aliarab, A.; Sebastiani, G.; Pantopoulos, K. Oxidative Stress in Liver Pathophysiology and Disease. Antioxidants 2023, 12, 1653. [Google Scholar] [CrossRef] [PubMed]
- Larina, S.N.; Ignatiev, I.V.; Chebyshev, N.V.; Kukes, V.G. Nuclear receptors and xenobiotic metabolism. Antibiotics and Chemotherapy 2009, 54(3–4), 42–48. https://www.antibiotics-chemotherapy.ru/jour/article/view/221 (accessed 07 October 2025).
- Rodríguez-Mozaz, S.; Huerta, B.; Barceló, D. Bioaccumulation of emerging contaminants in aquatic biota: patterns of pharmaceuticals in Mediterranean river networks. In Emerging Contaminants in River Ecosystems: Occurrence and Effects under Multiple Stress Conditions; Elsevier: Amsterdam, Netherlands, 2016; pp. 121–141.
- Ambili, T.R.; Saravanan, M.; Ramesh, M.; Abhijith, D.B.; Poopal, R.K. Toxicological effects of the antibiotic oxytetracycline to an Indian major carp Labeo rohita. Archives of environmental contamination and toxicology 2013, 64, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.H.D. Combined Toxicity of Microplastics and Antimicrobials on Animals: A Review. Antibiotics 2025, 14, 896. [Google Scholar] [CrossRef] [PubMed]
- Paramonov, S.G.; Zelikova, D.D.; Sklyarova, L.V.; Alkhutova, I.M. Environmental risks from micropollution of the environment with tetracycline. Pharmacy Formulas 2022, 4, 76–88. [Google Scholar] [CrossRef]
- Ding, L.; Zang, L.; Zhang, Y.; Zhang, Y.; Wang, X.; Ai, W.; Ding, N.; Wang, H. Joint toxicity of fluoroquinolone and tetracycline antibiotics to zebrafish (Danio rerio) based on biochemical biomarkers and histopathological observation. The Journal of toxicological sciences 2017, 42, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Au-Yeung, C.; Tsui, Y.-L.; Choi, M.-H.; Chan, K.-W.; Wong, S.-N.; Ling, Y.-K.; Lam, C.-M.; Lam, K.-L.; Mo, W.-Y. Antibiotic Abuse in Ornamental Fish: An Overlooked Reservoir for Antibiotic Resistance. Microorganisms 2025, 13, 937. [Google Scholar] [CrossRef]
- Andreeva, N.L.; Lunegov, A.M.; Baryshev, V.A.; Popova, O.S.; Kuznetsova, E.V. Pharmacology in aquaculture: Study guide. St. Petersburg State Academy of Veterinary Medicine: St. Petersburg, Russia, 2017, 76 p.
- Petrova, E.A. Algorithm for taking blood from different animal species. Krasnoyarsk State Agrarian University: Krasnoyarsk, Russia, 2024, 115 p.
- Sula, E.; Aliko, V.; Pagano, M.; Faggio, C. Digital light microscopy as a tool in toxicological evaluation of fish erythrocyte morphological abnormalities. Microscopy research and technique 2020, 83, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Canedo, A.; de Jesus, L.W.O.; Bailão, E.F.L.C.; Rocha, T.L. Micronucleus test and nuclear abnormality assay in zebrafish (Danio rerio): Past, present, and future trends. Environmental pollution (Barking, Essex : 1987) 2021, 290, 118019. [Google Scholar] [CrossRef] [PubMed]
- Skorodumov, D.I.; Subbotin, V.V. Microbiological diagnostics of bacterial diseases of animals. Izograf: Moscow, Russia, 2005, 656 p.
- Bankhead, P.; Loughrey, M. B.; Fernández, J. A.; Dombrowski, Y.; McArt, D. G.; Dunne, P. D.; McQuaid, S.; Gray, R. T.; Murray, L. J.; Coleman, H. G. , et al. QuPath: Open source software for digital pathology image analysis. Scientific reports 2017, 7, 16878. [Google Scholar] [CrossRef] [PubMed]
- Stritt, M.; Stalder, A.K.; Vezzali, E. Orbit Image Analysis: An open-source whole-slide image analysis tool. PLoS Computational Biology 2020, 16, e1007313. [Google Scholar] [CrossRef] [PubMed]
- Elabscience®. Cytochrome P450 Reductase (CPR) Activity Assay Kit Manual. No. E-BC-K808-M, p. 16. https://www.elabscience.com/p/cytochrome-p450-reductase-cpr-activity-assay-kit--e-bc-k808-m?srsltid=AfmBOopI_JLEBko6qm6MqBJlsC7K5d_tokVjeglzfoxDDnHfxX3TkR5k (accessed 7 October 2025).
- Kudersky, L.A. Selected works: Research in ichthyology, fisheries and related disciplines. Vol. 3. KMK: Moscow, Russia, 2013, 526 p.
- Zahran, E.; Mamdouh, A.Z.; Elbahnaswy, S.; El-Son, M.M.A.; Risha, E.; ElSayed, A.; El Barbary, M.I.; El Sebaei, M.G. The impact of heavy metal pollution: bioaccumulation, oxidative stress, and histopathological alterations in fish across diverse habitats. Aquacult Int 2025, 33, 371. [Google Scholar] [CrossRef]
- Pramanik, S.; Biswas, J.K. Histopathological fingerprints and biochemical changes as multi-stress biomarkers in fish confronting concurrent pollution and parasitization. iScience 2024, 27, 109457. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.V.S. Biomarkers of trace element toxicity in fish: A new paradigm in environmental health risk assessment. In book: Fish Species in Environmental Risk Assessment Strategies. Elsevier: Amsterdam, Netherlands, 2024, 6-31. [CrossRef]
Figure 1.
Geography of the research stations.
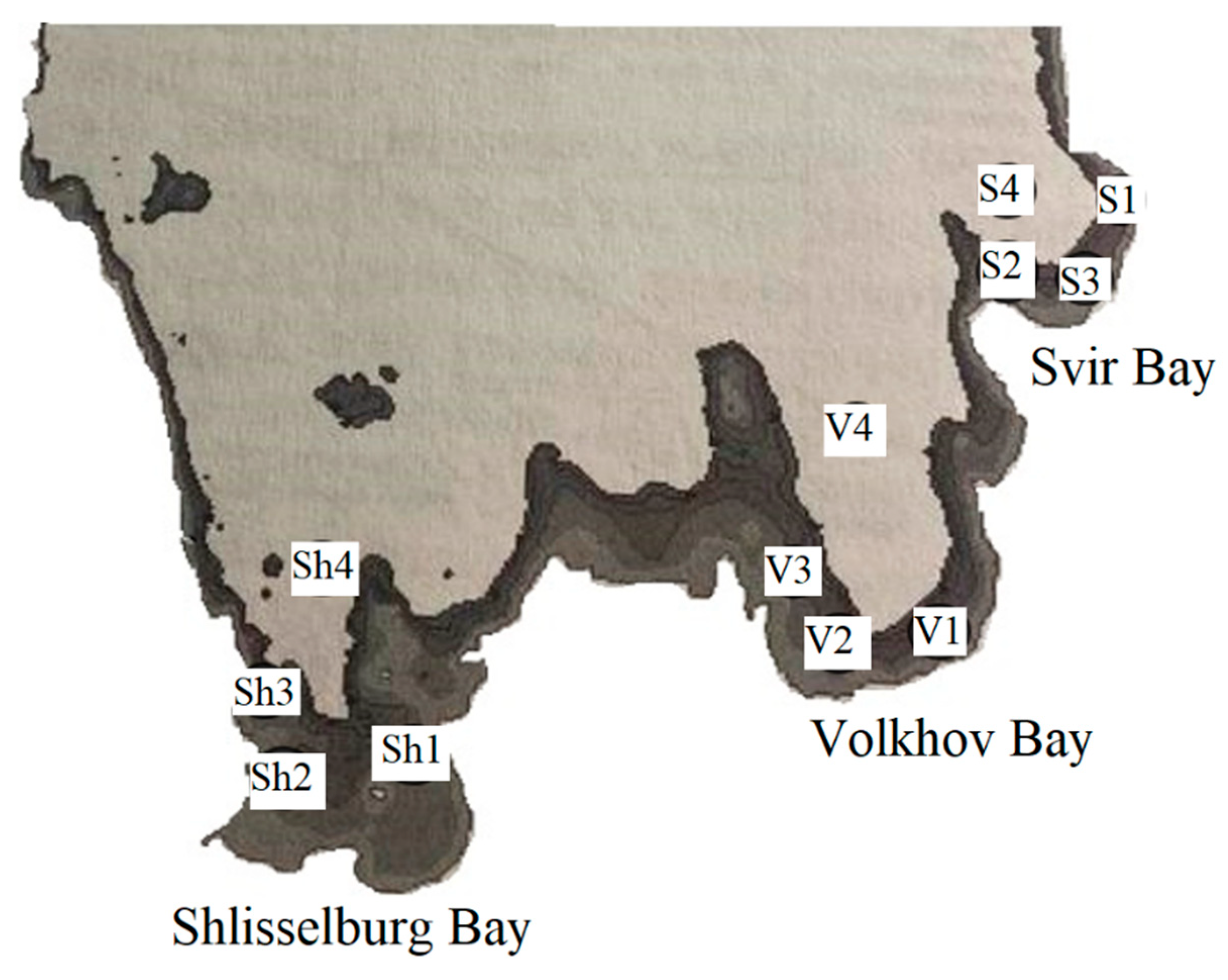
Figure 2.
Anemia of fish gill tissue.

Figure 3.
Pathology of fish gills: uneven coloration, soft tissue damage.

Figure 4.
Pathology of fish gills: uneven coloration, regeneration of tissue necrosis.
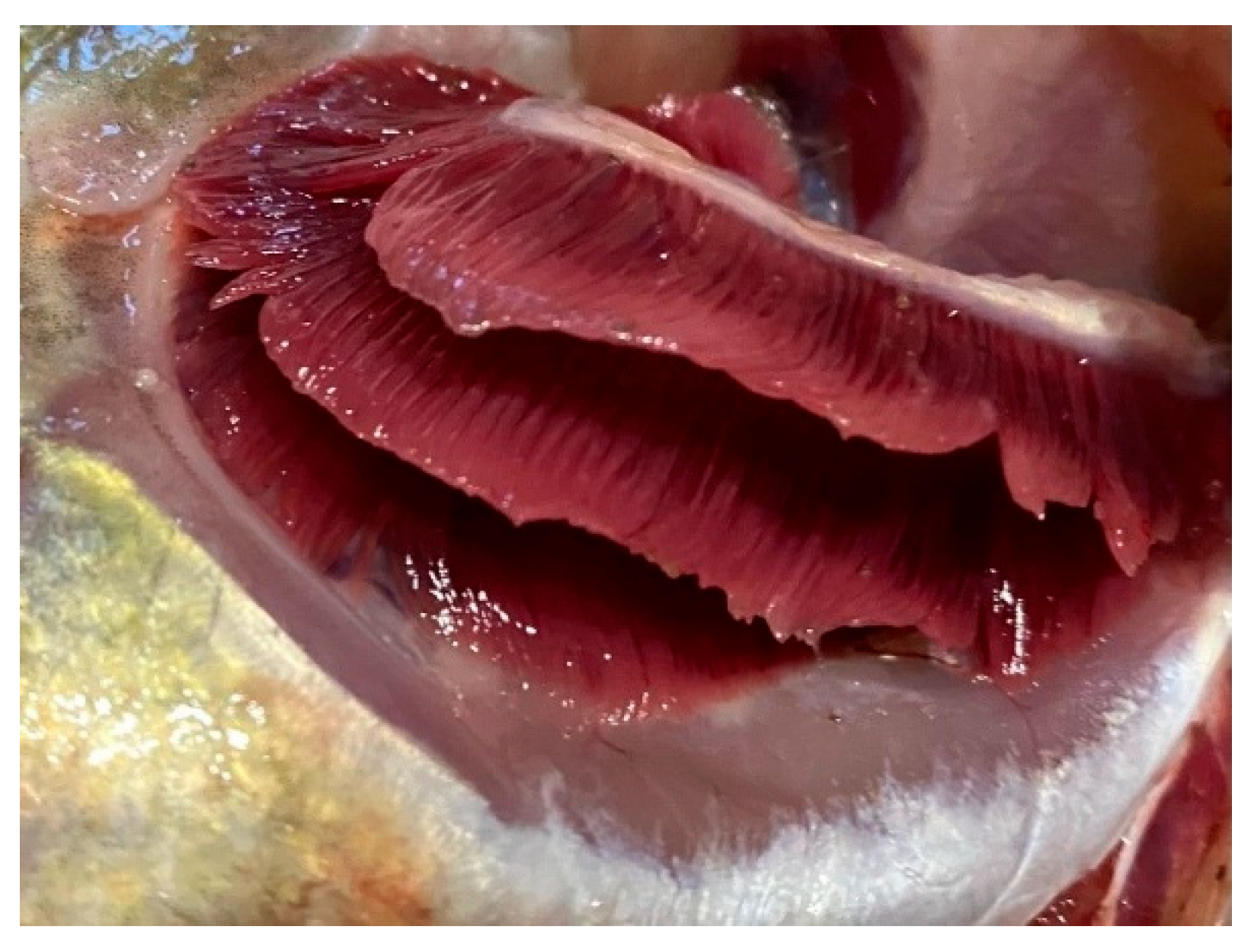
Figure 5.
Bream liver in normal physiological condition.

Figure 6.
Fish liver pathology: liver degeneration.
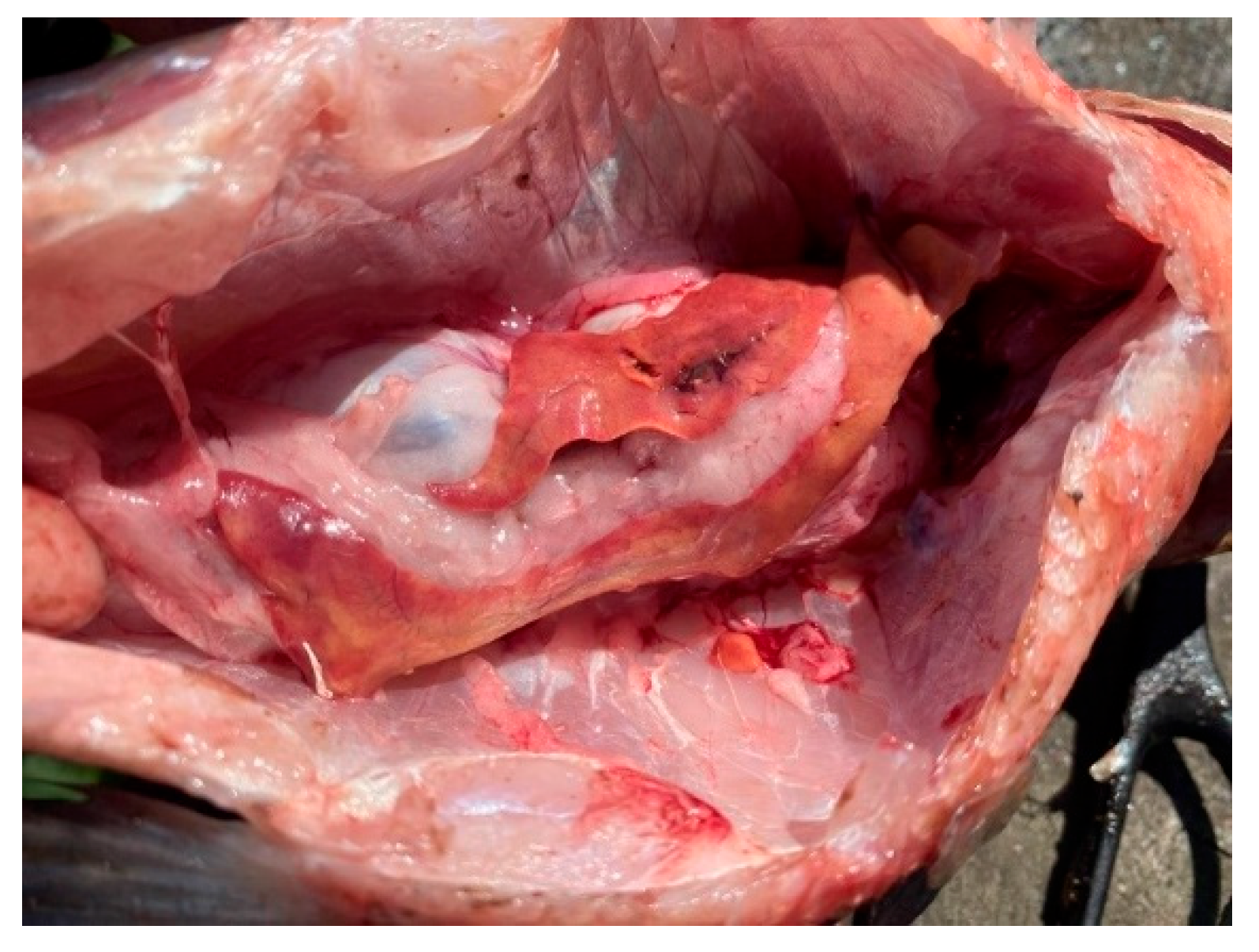
Figure 7.
Fish liver pathology: hyperemia of the fish liver.
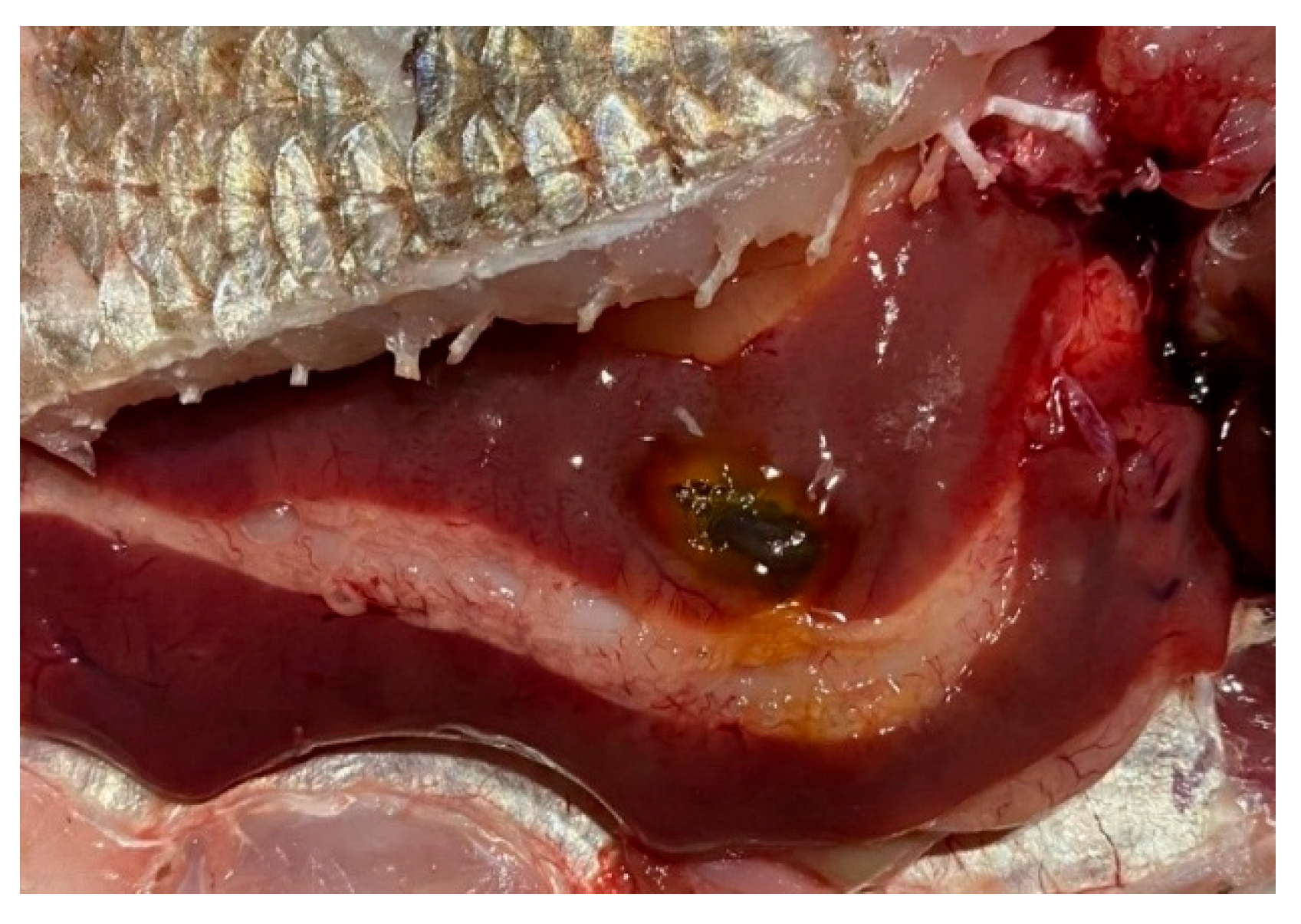
Figure 8.
Pathology of fish liver: degeneration and deformation of the organ.
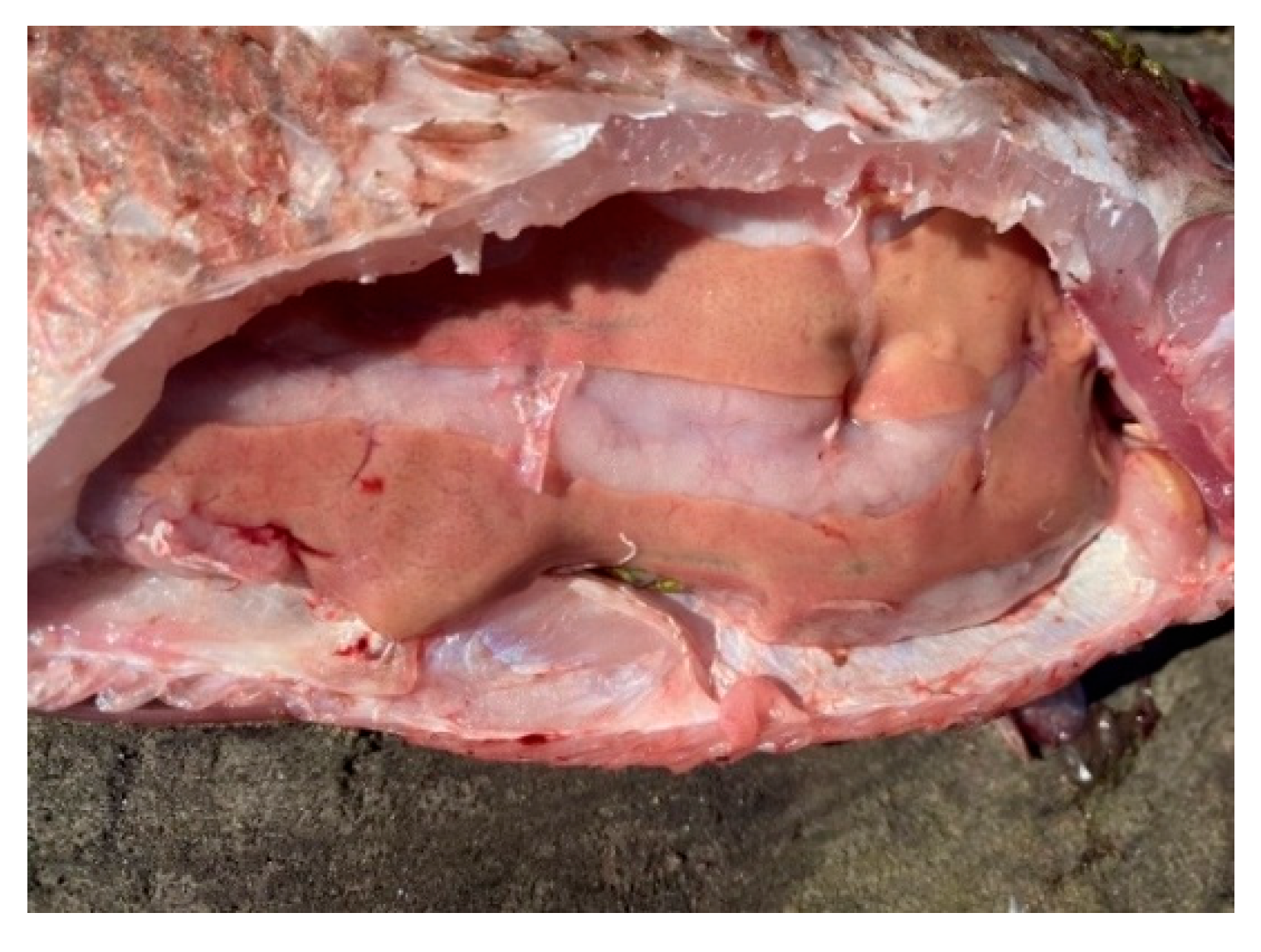
Figure 9.
Pathology of fish liver: foci of degeneration.
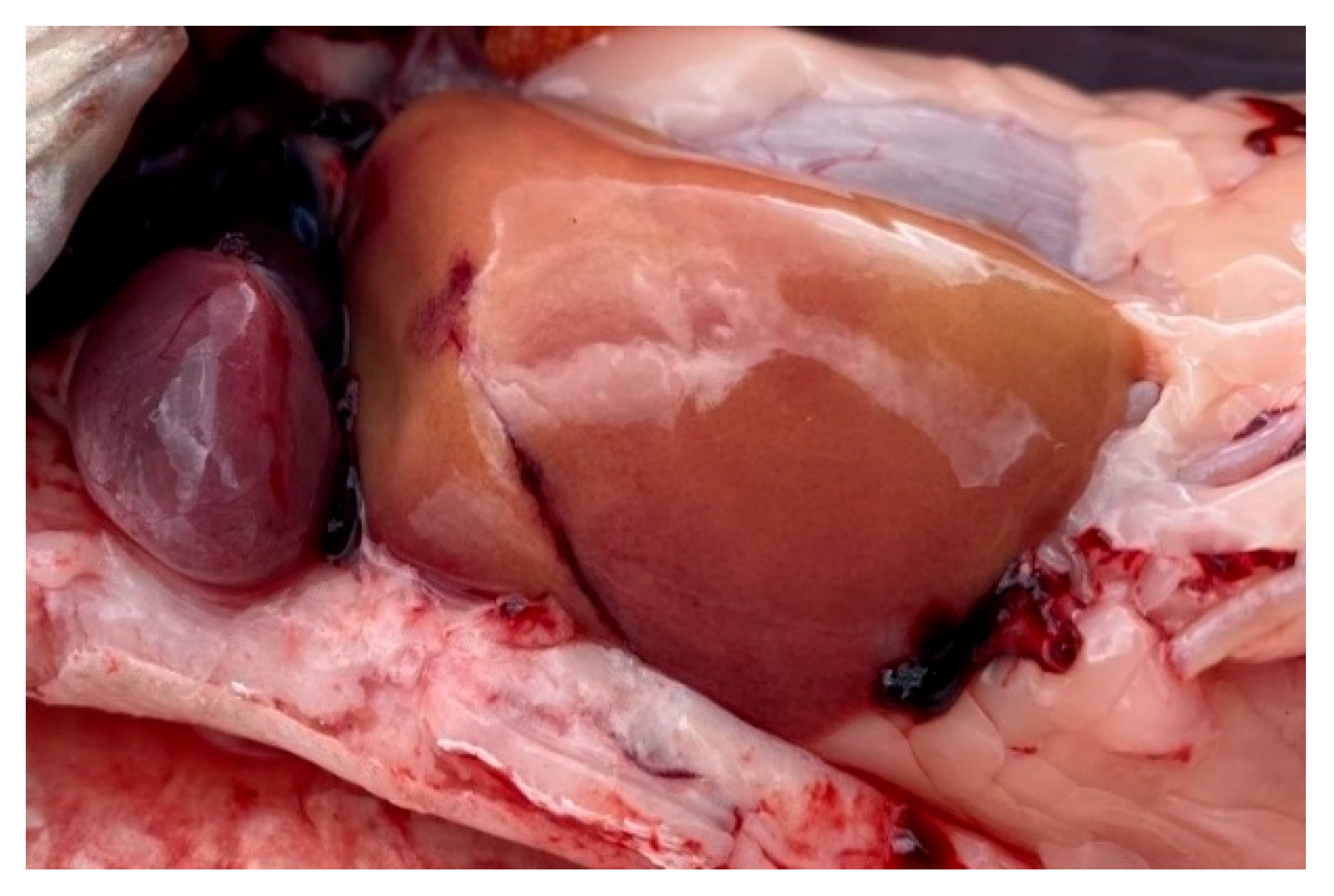
Figure 10.
A fragment of the bream gills from the southern part of Ladoga Lake with preserved architecture. 1 — petals (primary lamellae); 2 — petals (secondary lamellae); 3 — capillaries. Stained with iron hematoxylin according to Heidenhain, magnification: x200.
Figure 10.
A fragment of the bream gills from the southern part of Ladoga Lake with preserved architecture. 1 — petals (primary lamellae); 2 — petals (secondary lamellae); 3 — capillaries. Stained with iron hematoxylin according to Heidenhain, magnification: x200.
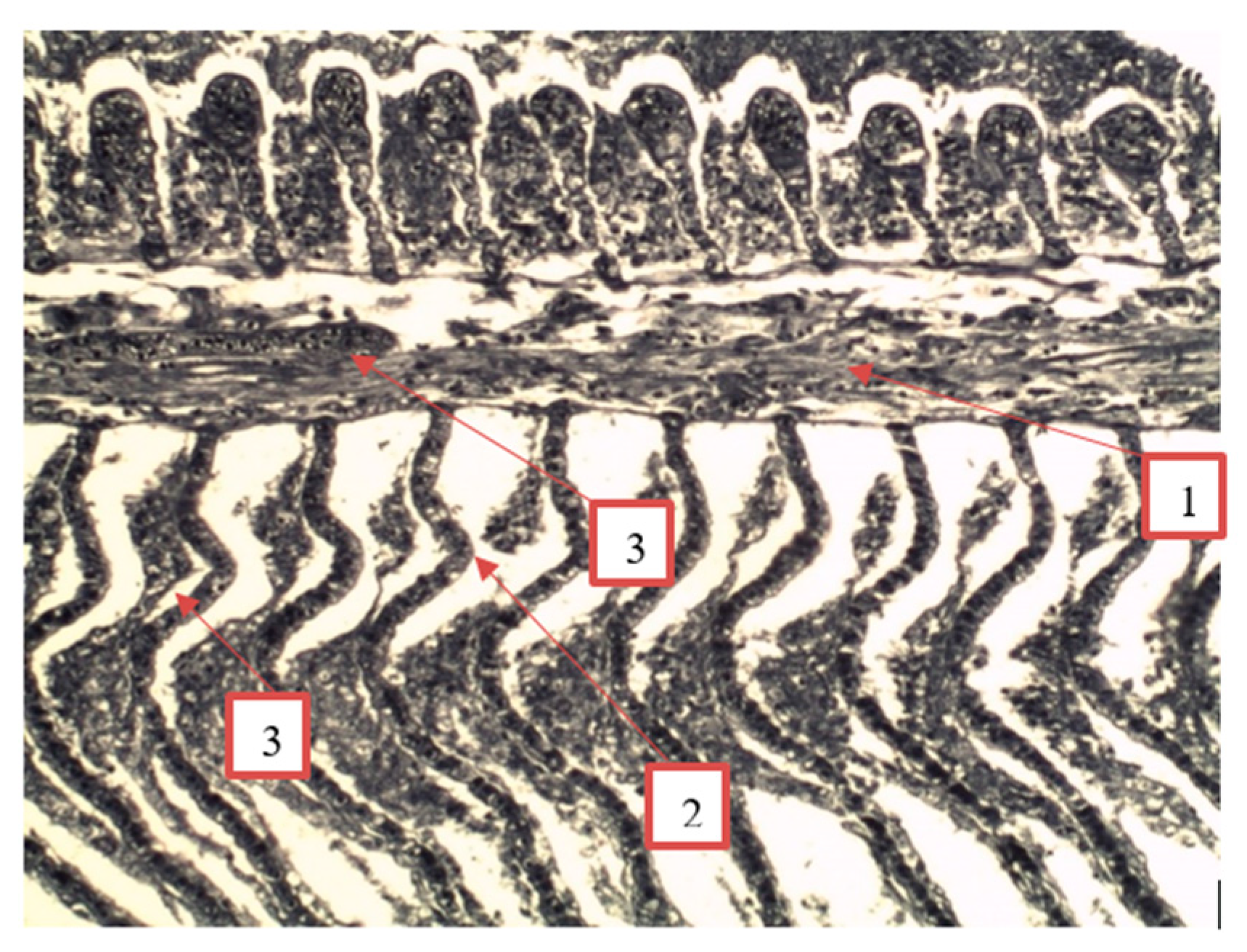
Figure 11.
A fragment of bream gills from the southern part of Ladoga Lake with damaged blood vessels of the lamellae. 1 – lobes (primary lamellae); 2 – lobes (secondary lamellae); 3 – damaged capillaries. Stained with iron hematoxylin according to Heidenhain, magnification x200.
Figure 11.
A fragment of bream gills from the southern part of Ladoga Lake with damaged blood vessels of the lamellae. 1 – lobes (primary lamellae); 2 – lobes (secondary lamellae); 3 – damaged capillaries. Stained with iron hematoxylin according to Heidenhain, magnification x200.
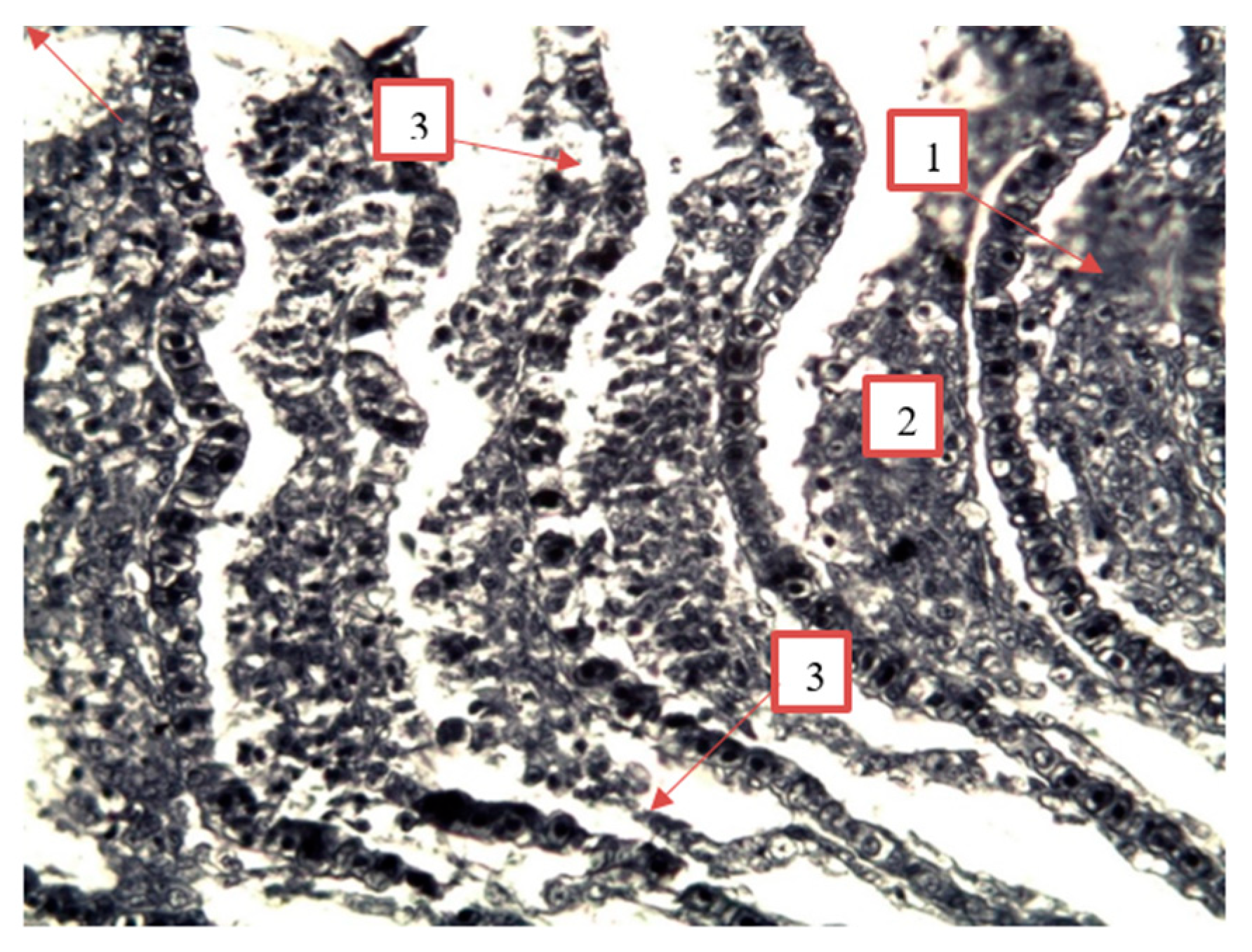
Figure 12.
Granular dystrophy of the epithelial cells of the renal tubules of roach from the southern part of Ladoga Lake. 1 — urinary collecting tube; 2 — proximal convoluted tubule; 3 — vascular glomerulus; 4 — distal convoluted tubule. Stained with iron hematoxylin according to Heidenhain, magnification: x200.
Figure 12.
Granular dystrophy of the epithelial cells of the renal tubules of roach from the southern part of Ladoga Lake. 1 — urinary collecting tube; 2 — proximal convoluted tubule; 3 — vascular glomerulus; 4 — distal convoluted tubule. Stained with iron hematoxylin according to Heidenhain, magnification: x200.
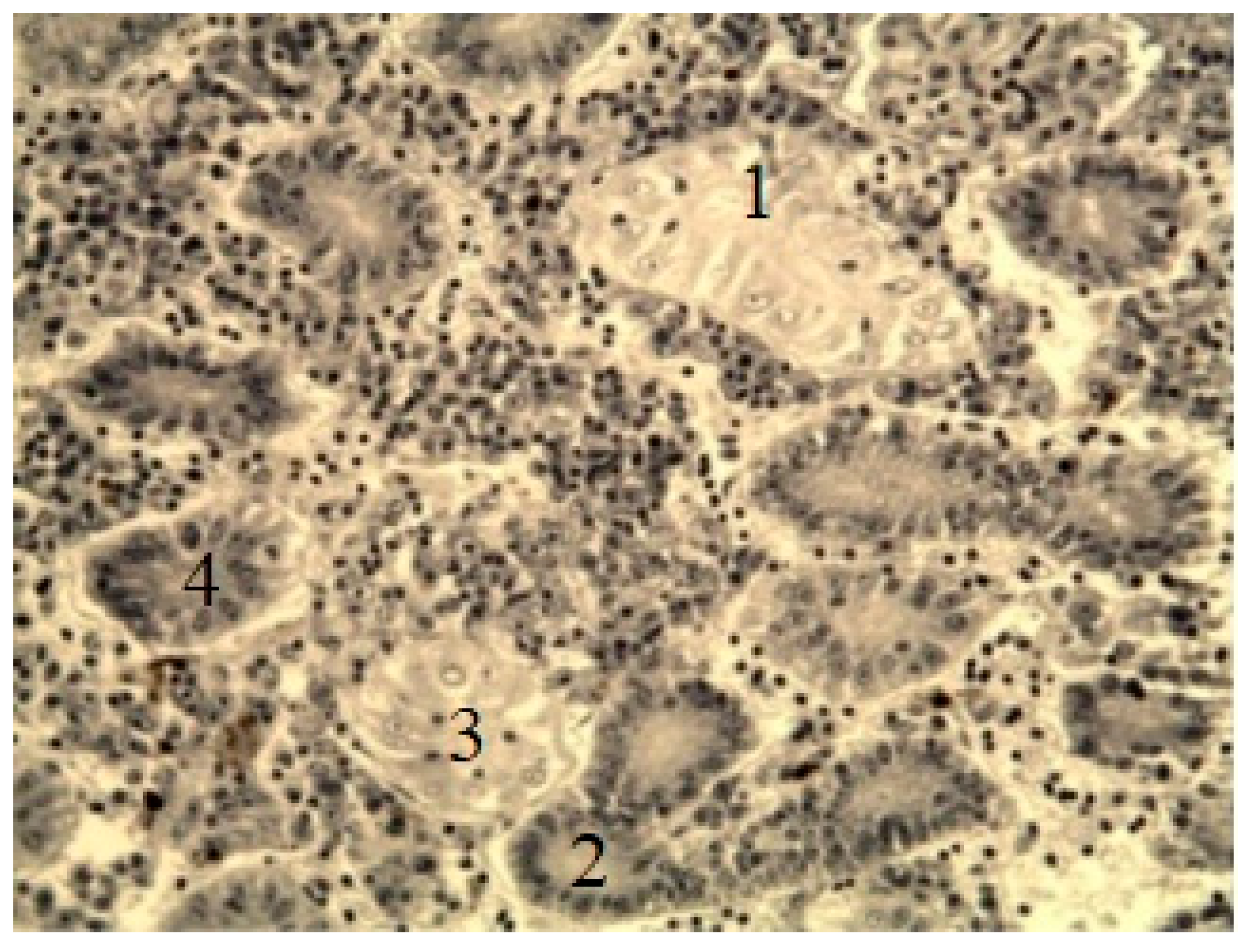
Figure 13.
A fragment of the liver of bream (A) and perch (B) from the southern part of Ladoga Lake. A) 1 — hepatocyte nuclei; 2 — central vein (secondary lamellae); B) 1 — blood vessel; 2 — accumulation of macrophages containing melanin by Heidenhain’s hematoxylin, magnification: x200.
Figure 13.
A fragment of the liver of bream (A) and perch (B) from the southern part of Ladoga Lake. A) 1 — hepatocyte nuclei; 2 — central vein (secondary lamellae); B) 1 — blood vessel; 2 — accumulation of macrophages containing melanin by Heidenhain’s hematoxylin, magnification: x200.
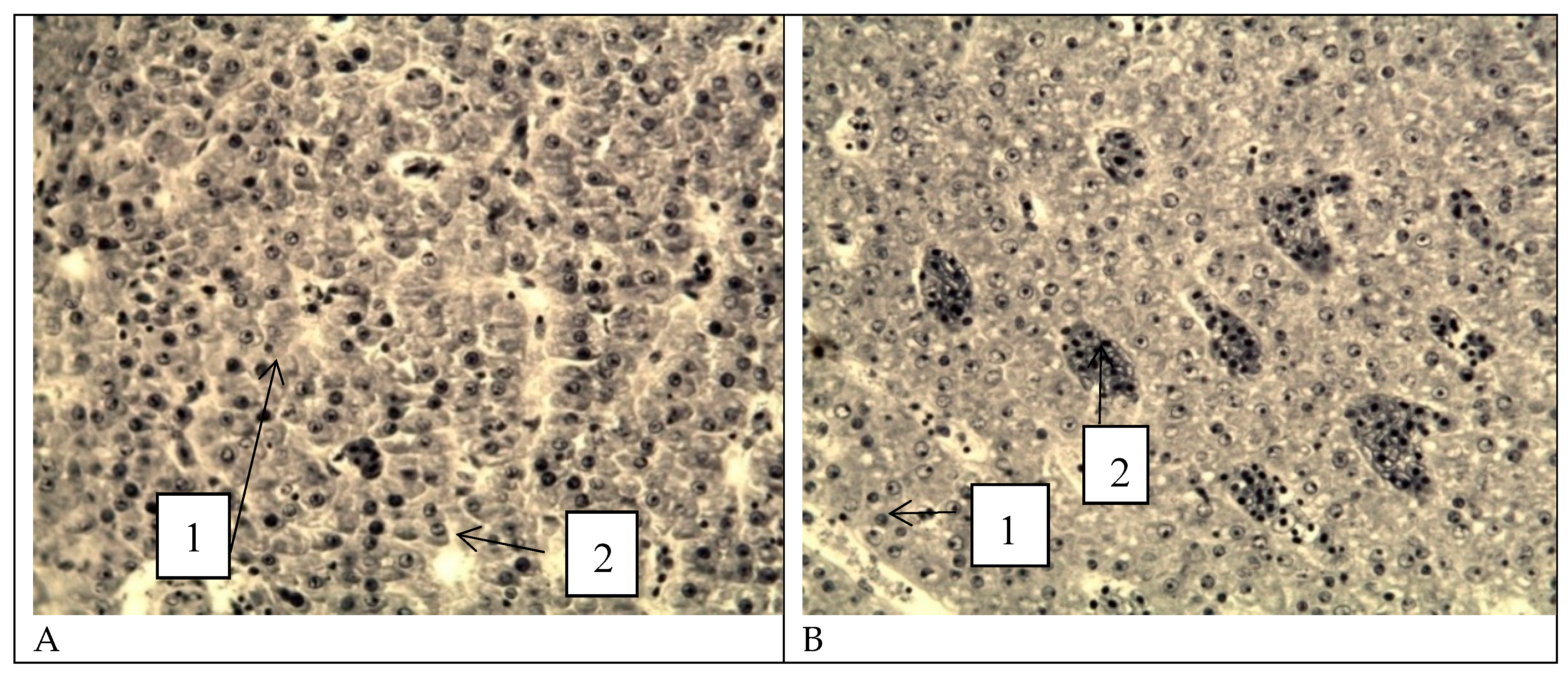
Figure 14.
Vacuolization of older oocytes during vitellogenesis in perch from the southern part of Ladoga Lake. Iron hematoxylin staining according to Heidenhain, magnification: x200.
Figure 14.
Vacuolization of older oocytes during vitellogenesis in perch from the southern part of Ladoga Lake. Iron hematoxylin staining according to Heidenhain, magnification: x200.
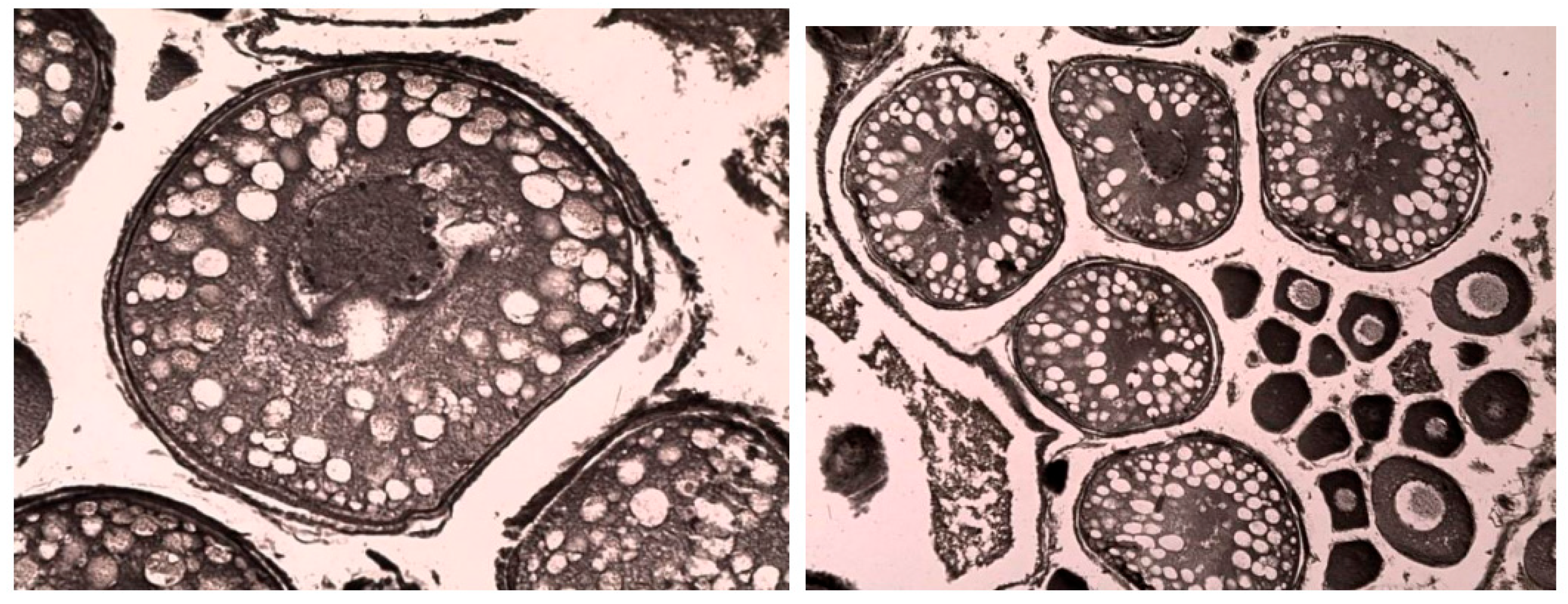
Figure 15.
Microphotograph of a peripheral blood smear. 1 - vacuolation of the erythrocyte cytoplasm, 2 - vacuolation of the erythrocyte nucleus, 3 - anisocytosis of erythrocytes. Stained according to Romanovsky. Magnification x1000.
Figure 15.
Microphotograph of a peripheral blood smear. 1 - vacuolation of the erythrocyte cytoplasm, 2 - vacuolation of the erythrocyte nucleus, 3 - anisocytosis of erythrocytes. Stained according to Romanovsky. Magnification x1000.
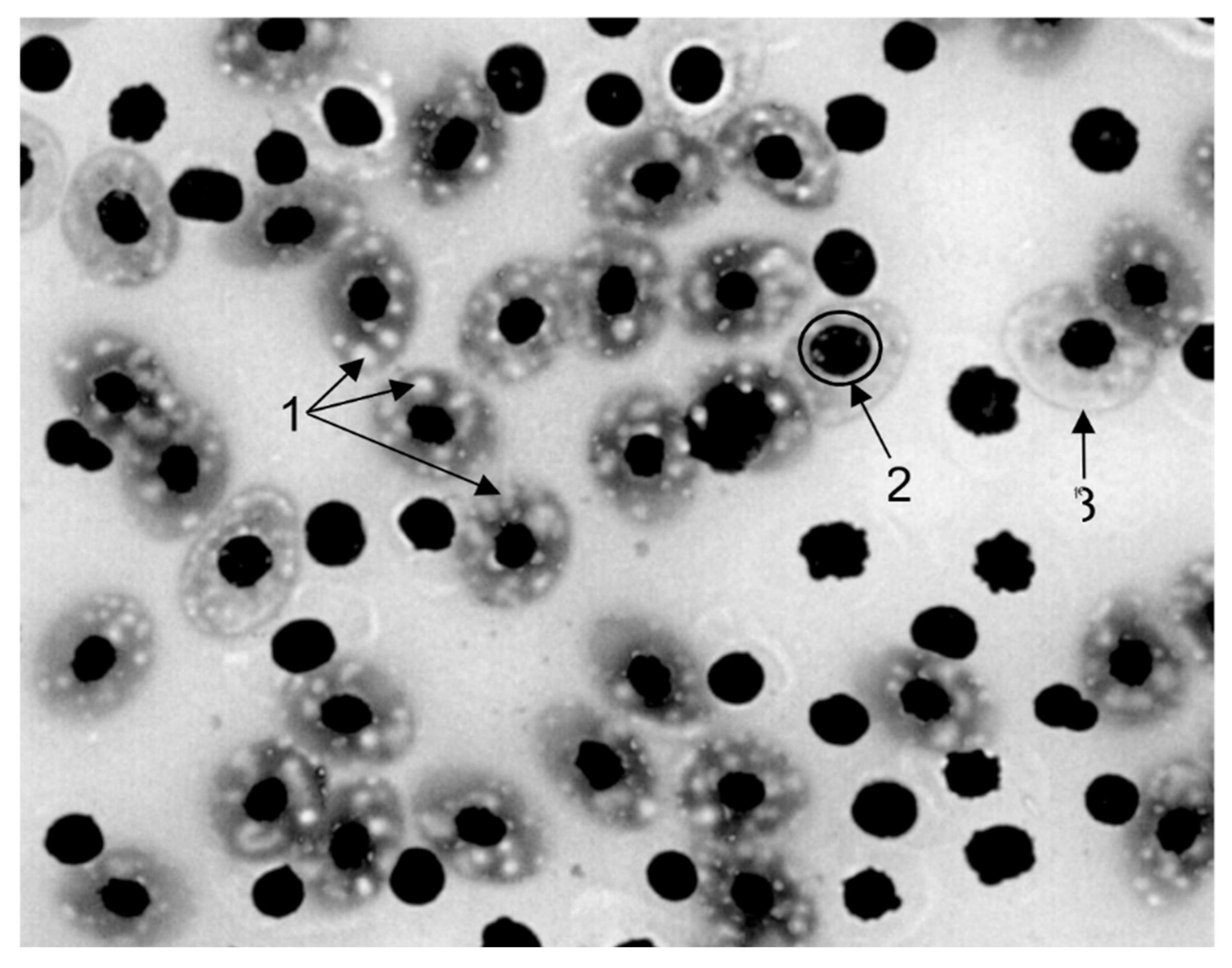
Figure 16.
Micrographs of liver tissue. A, B - healthy fish, C, D - studied fish. 1 - Vacuolization of the hepatocyte cytoplasm. 2 - Binucleated cells. 3 - Shift of nuclei to the periphery of hepatocytes. Stained with hematoxylin, Carazzi, and eosin. Magnification x1000.
Figure 16.
Micrographs of liver tissue. A, B - healthy fish, C, D - studied fish. 1 - Vacuolization of the hepatocyte cytoplasm. 2 - Binucleated cells. 3 - Shift of nuclei to the periphery of hepatocytes. Stained with hematoxylin, Carazzi, and eosin. Magnification x1000.
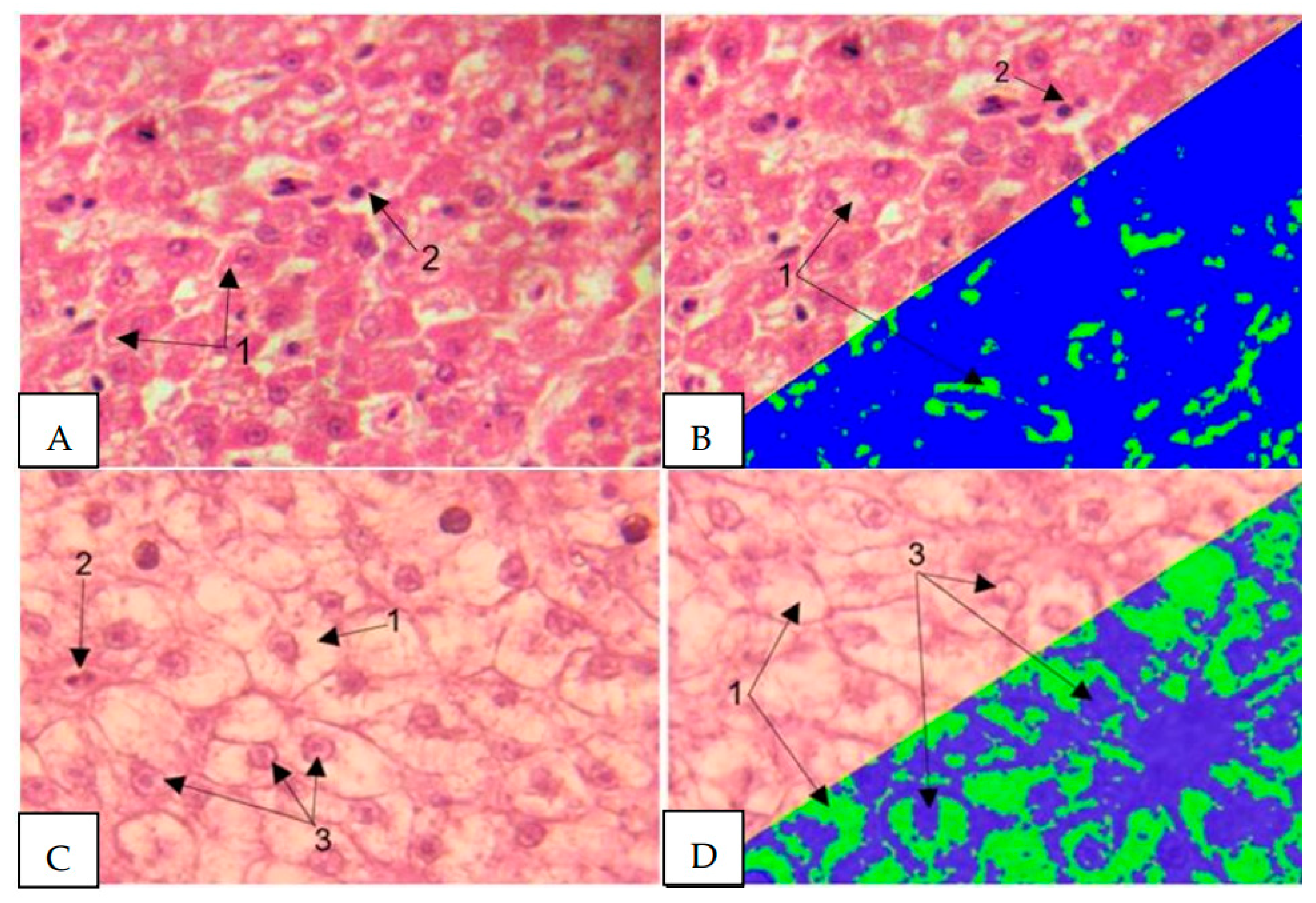
Figure 17.
Micrographs of common carp (Cyprinus carpio) liver tissue. A – healthy specimens. B – specimens under study. Stained with Sudan III and scarlet red, magnification x320.
Figure 17.
Micrographs of common carp (Cyprinus carpio) liver tissue. A – healthy specimens. B – specimens under study. Stained with Sudan III and scarlet red, magnification x320.

Figure 18.
Three-dimensional mapping of the chromatic dispersion coefficient of lipid infiltration in the liver of healthy (A) and experimental (B) fish.
Figure 18.
Three-dimensional mapping of the chromatic dispersion coefficient of lipid infiltration in the liver of healthy (A) and experimental (B) fish.
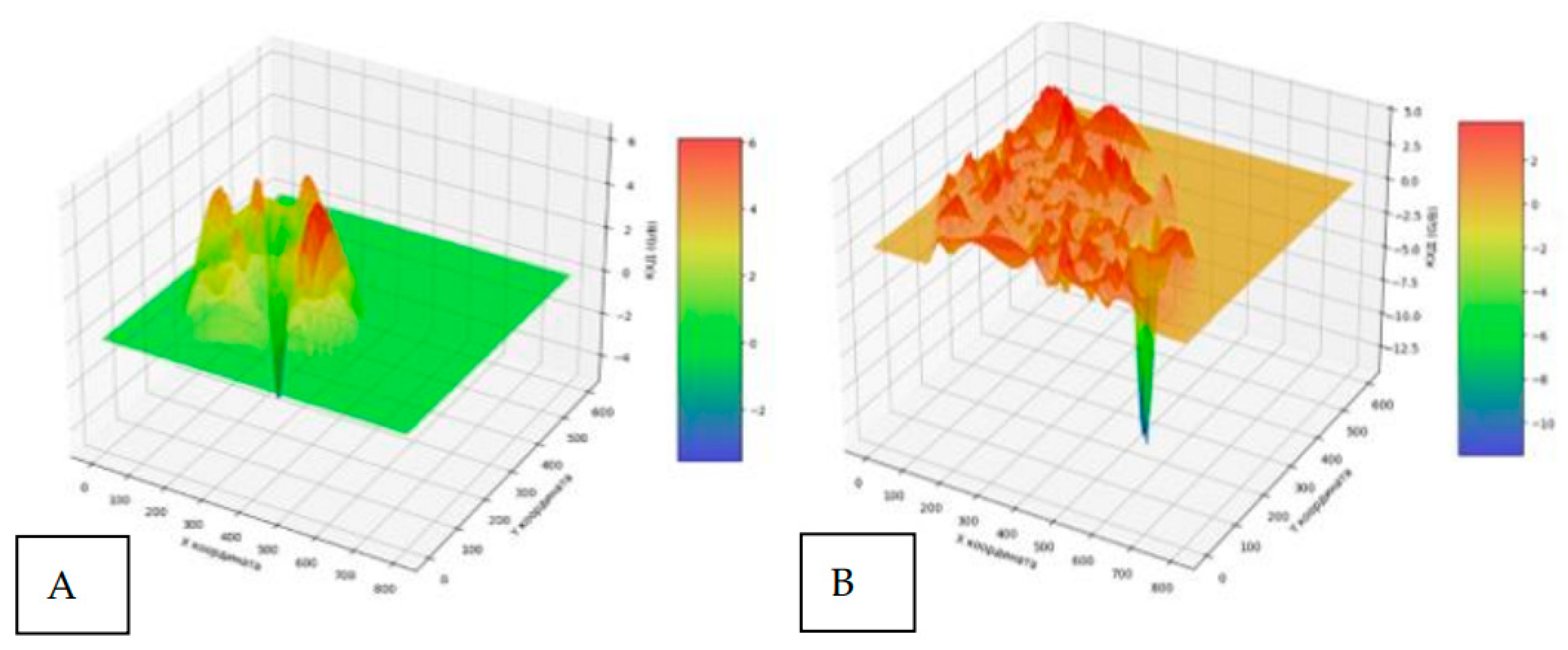
Figure 19.
Heat map of the distribution of the chromatic dispersion coefficient of lipid infiltration of the liver of healthy (A) and experimental (B) fish.
Figure 19.
Heat map of the distribution of the chromatic dispersion coefficient of lipid infiltration of the liver of healthy (A) and experimental (B) fish.
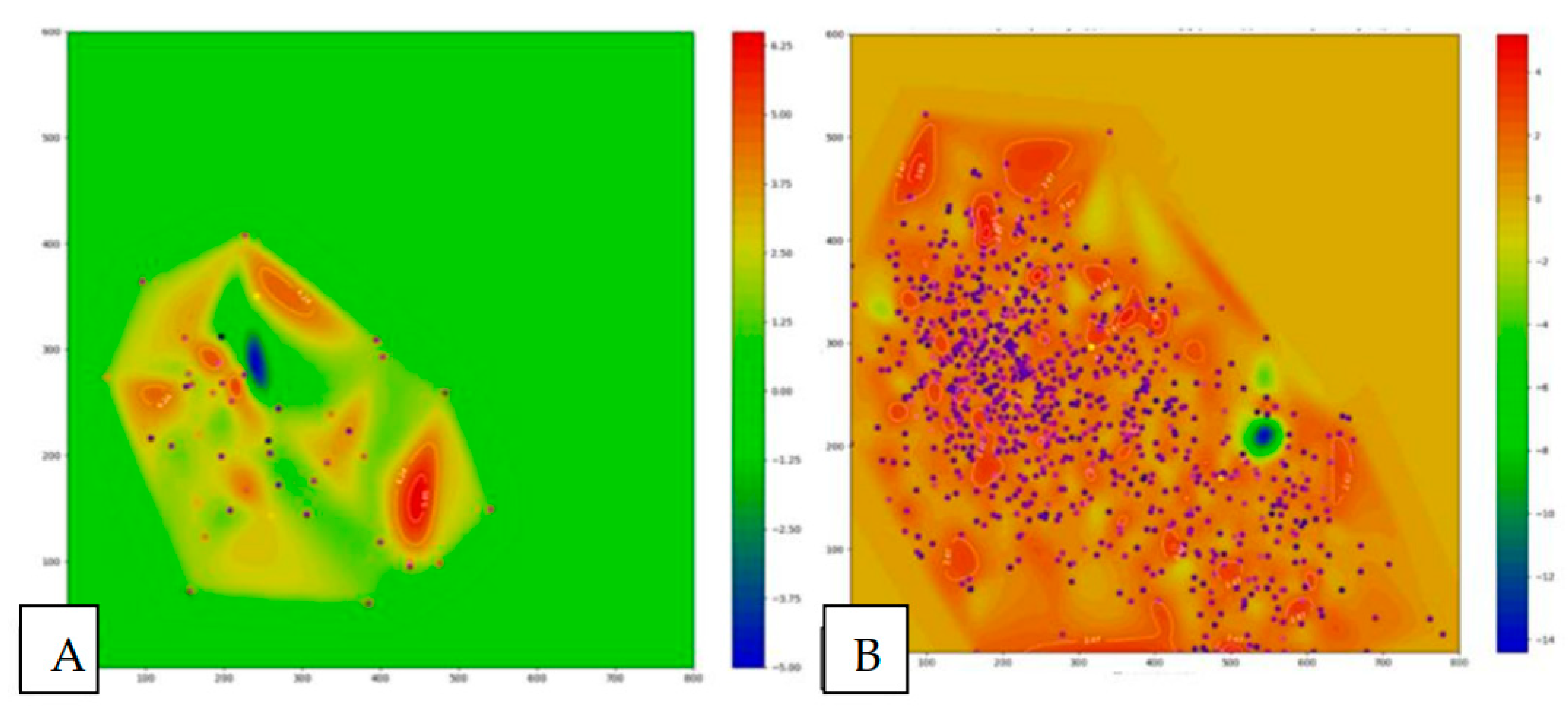

Table 1.
Depth at the research stations.
| Water Area | Shlisselburg Bay | Volkhov Bay | Svir Bay | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Station | Sh1 | Sh2 | Sh3 | Sh4(C*) | V1 | V2 | V3 | V4(C*) | S1 | S2 | S3 | S4(C*) |
| Depth, m | 8 | 3,5 | 5 | 15 | 6 | 6,3 | 6,5 | 10 | 7,1 | 8,3 | 6,4 | 20 |
* C - control stations.
Table 2.
Physical and hydrochemical parameters of water in the southern part of Ladoga Lake.
| Water area | Station | h, m | Horizon | Т,℃ | El, µS/cm | Mn, g/l | О2, mg/l | % sat, (О2) | pH | Eh, mV |
|---|---|---|---|---|---|---|---|---|---|---|
| Volkhov Bay | V1 | 6 | surface | 19,1±0,7 | 162,2±0,5 | 0,103±0,060 | 9,5±0,5 | 103,2±1,3 | 8,2±0,4 | 133,0±1,3 |
| bottom | 19,8±0,6 | 187,3±0,6 | 0,122±0,020 | 8,8±0,3 | 95,5±1,5 | 7,8±0,3 | 80,0±1,2 | |||
| V2 | 6,3 | surface | 19,4±0,2 | 158,9±0,9 | 0,103±0,030 | 9,8±0,7 | 106,2±0,9 | 8,3±0,3 | 43,0±0,9 | |
| bottom | 18,6±0,3 | 171,1±0,7 | 0,115±0,050 | 9,5±0,5 | 101,3±1,5 | 8,1±0,4 | 62,0±0,7 | |||
| V3 | 6,5 | surface | 16,7±0,2 | 177,7±0,5 | 0,067±0,008 | 10,6±0,3 | 110,5±1,6 | 8,1±0,5 | 71,0±1,2 | |
| bottom | 15,8±0,6 | 103,3±0,2 | 0,066±0,003 | 10,2±0,1 | 106,7±1,2 | 8,2±0,5 | 83,0±1,2 | |||
| V4 | 10 | surface | 18,6±0,3 | 102,2±0,9 | 0,114±0,010 | 9,8±0,6 | 103,5±1,4 | 8,0±0,6 | 66,0±1,8 | |
| bottom | 17,4±0,3 | 101,2±0,7 | 0,095±0,001 | 11,0±0,4 | 96,5±1,3 | 7,8±0,2 | 84,0±0,9 | |||
| Svir Bay | S1 | 7,1 | surface | 12,8±0,5 | 94,5±0,8 | 0,061±0,002 | 11,2±0,3 | 105,7±1,7 | 8,0±0,1 | 99,0±0,6 |
| bottom | 12,0±0,85 | 95,9±0,6 | 0,060±0,005 | 11,00±0,1 | 106,6±1,0 | 8,0±0,3 | 112,0±1,0 | |||
| S2 | 8,3 | surface | 12,8±0,6 | 84,3±0,8 | 0,056±0,005 | 11,1±0,4 | 105,5±0,9 | 7,9±0,5 | 90,0±1,3 | |
| bottom | 12,6±0,4 | 83,9±0,7 | 0,055±0,006 | 11,3±0,2 | 105,5±1,5 | 7,9±0,2 | 125,0±1,8 | |||
| S3 | 6,4 | surface | 15,8±0,2 | 71,8±0,6 | 0,032±0,003 | 10,2±0,3 | 102,3±1,8 | 7,7±0,3 | 109,0±1,6 | |
| bottom | 15,5±0,3 | 71,2±0,5 | 0,030±0,006 | 10,3±0,5 | 100,8±1,4 | 7,4±0,2 | 130,0±0,9 | |||
| S4 | 22 | surface | 9,7±0,2 | 92,8±0,7 | 0,060±0,003 | 12,2±0,6 | 105,8±1,9 | 7,4±0,5 | 131,5±1,2 | |
| bottom | 9,5±0,4 | 95,2±0,2 | 0,060±0,002 | 12,3±0,5 | 102,6±1,8 | 7,1±0,4 | 110,0±1,0 | |||
| Shlisselburg Bay | Sh1 | 8 | surface | 16,0±0,4 | 92,5±0,5 | 0,061±0,001 | 10,5±0,1 | 105,8±1,2 | 7,6±0,4 | 84,2±0,3 |
| bottom | 14,5±0,2 | 94,7±0,5 | 0,061±0,001 | 10,4±0,2 | 98,7±1,4 | 7,8±0,4 | 156,2±1,9 | |||
| Sh2 | 3,5 | surface | 14,5±0,1 | 84,2±0,5 | 0,053±0,002 | 10,7±0,2 | 105,5±1,6 | 8,4±0,1 | 70,5±0,2 | |
| bottom | 13,0±0,2 | 87,3±0,4 | 0,053±0,004 | 11,2±0,3 | 105,5±1,2 | 8,0±0,5 | 92,5±0,3 | |||
| Sh3 | 5 | surface | 11,2±0,3 | 91,4±1,2 | 0,055±0,003 | 11,4±0,3 | 103,0±1,6 | 8,4±0,1 | 87,0±0,5 | |
| bottom | 10,2±0,7 | 90,7±0,5 | 0,063±0,005 | 11,5±0,4 | 98,9±1,0 | 7,9±0,1 | 98,3±1,0 | |||
| Sh4 | 15 | surface | 10,3±0,5 | 94,3±1,0 | 0,065±0,003 | 10,6±0,2 | 101,6±1,7 | 7,6±0,7 | 96,0±1,5 | |
| bottom | 10,0±0,4 | 93,5±0,7 | 0,066±0,001 | 10,5±0,9 | 97,7±1,2 | 7,4±0,6 | 95,0±1,7 |
Table 3.
Results of the seasonal study of the southern part of Ladoga Lake.
| Seasons | Fishing stationsб | Fish species | Number of fish examined | Fish condition assessment | ||
|---|---|---|---|---|---|---|
| Number of fish with pathological changes (%) | Pathology severity (points) | Number of fish by points | ||||
| Spring | V1 5 km from the mouth of the Volkhov River |
bream pike-perch roach perch |
10 10 10 10 |
50 50 60 50 |
2-3-4,0 2-3,0 2-3-4,0 2-3,0 |
2-2,0;2-3,0;1-4,0 2-2,0;3-3,0 3-2,0;2-3,0;1-4,0 2-2,0;2-3,0; 1 -4,0 |
| V2 Syassky Pulp and Paper Mill area |
bream pike-perch roach perch |
10 10 10 10 |
60 70 60 50 |
2-3,0 2-3-4,0 2-3,0 2-3,0 |
2-2,0;4-3,0 2-2,0;4-3,0; 1-4,0 2-2,0; 4-3,0 2-2,0;3-3,0 |
|
| V3 5 km to the left of the mouth of the Volkhov River |
bream pike-perch roach perch |
10 10 10 10 |
50 40 50 40 |
2-3,0 2-3,0 2-3,0 2-3,0 |
2-2,0;3-3,0 2-2,0;2-3,0 2-2,0;3-3,0 2-2,0;2-3,0 |
|
| V4 Outside the littoral zone |
bream pike-perch perch smelt |
10 10 10 10 |
40 40 50 30 |
2-3,0 2-3,0 2-3,0 2,0 |
2-2,0;2-3,0 2-2,0;2-3,0 1-2,0;4-3,0 3-2,0 |
|
| Summer | V1 5 km from the mouth of the Volkhov River |
bream pike-perch roach perch |
10 10 10 10 |
50 60 50 50 |
2-3,0 2-3,0 2-3,0 2-3,0 |
2-2,0;3-3,0 4-2,0;2-3,0 3-2,0;2-3,0 2-2,0;3-3,0 |
| V2 Syassky Pulp and Paper Mill area |
bream pike-perch roach perch |
10 10 10 10 |
60 60 50 60 |
2-3-4,0 2-3-4,0 2-3,0 2-3,0 |
2-2,0;3-3,0;1-4,0 2-1,0;4-3,0 2-2,0;3-3,0 2-2,0;4-3,0 |
|
| V3 5 km to the left of the mouth of the Volkhov River |
bream pike-perch roach perch |
10 10 10 10 |
50 40 50 40 |
2-3,0 2-3,0 2-3,0 2-3,0 |
2-2,0;3-3,0 1-2,0;3-3,0 2-2,0;3-3,0 2-2,0;2-3,0 |
|
| V4 Outside the littoral zone |
bream pike perch roach smelt |
10 10 10 10 |
40 30 50 30 |
2-3,0 2-3,0 2-3,0 2,0 |
2-2,0;2-3,0 2-2,0;1-3,0 2-2,0;3-3,0 3-2,0 |
|
| Autumn | V1 5 km from the mouth of the Volkhov River |
bream pike-perch roach perch |
10 10 10 10 |
40 50 40 50 |
2-3,0 2-3,0 2-3,0 2-3,0 |
2-2,0;2-3,0 3-2,0;2-3,0 2-2,0;2-3,0 2-2,0;3-3,0 |
| V2 Syassky Pulp and Paper Mill area |
bream pike-perch roach perch |
10 10 10 10 |
50 60 50 50 |
2-3,0 2-3,0 2-3,0 2-3,0 |
3-2,0;2-3,0 2-2,0;4-3,0 2-2,0;3-3,0 2-2,0;3-3,0 |
|
| V3 5 km to the left of the mouth of the Volkhov River |
bream pike-perch roach perch |
10 10 10 10 |
40 40 30 40 |
2-3,0 2-3,0 2-3,0 2-3,0 |
2-2,0;2-3,0 2-2,0;2-3,0 2-2,0;1-3,0 2-2,0;2-3,0 |
|
| V4 Outside the littoral zone |
bream pike perch roach smelt |
10 10 10 10 |
40 30 40 20 |
2-3,0 2-3,0 2-3,0 2,0 |
3-2,0;1-3,0 2-2,0;1-3,0 2-2,0;2-3,0 2-2,0 |
|
Table 4.
Bacteriological examination of pathological material isolated from fish.
| Pathological material tested | Bacteriological examination results | |
|---|---|---|
| Sample No. 1 | Isolated:
|
|
| Sample No. 2 | Isolated:
|
|
| Sample No. 3 | Isolated:
|
|
| Sample No. 4 | Isolated:
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



