Submitted:
10 October 2025
Posted:
10 October 2025
You are already at the latest version
Abstract
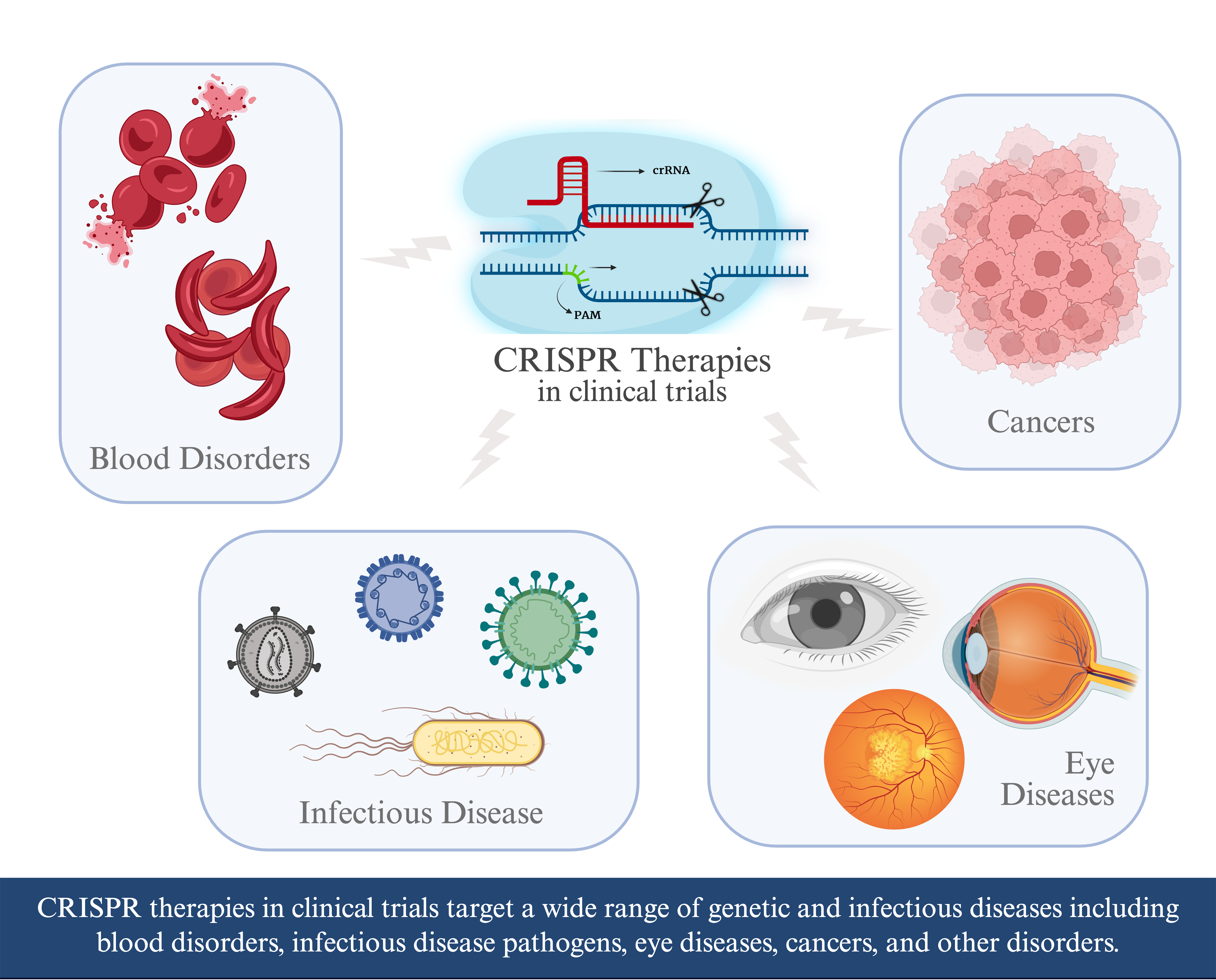
CRISPR-Cas9 is a gene editing tool used extensively in biological research that is now making its way into clinical therapies. With the first CRISPR therapy obtaining approval by the United States’ Food and Drug Administration (FDA) in late 2023, we look at clinical trials of emerging therapies involving CRISPR-Cas9, currently the most prevalent CRISPR-based tool in these trials. A CRISPR-based therapy is currently approved for treatment of both sickle-cell anemia and transfusion-dependent β-thalassemia but clinical trials for CRISPR-based therapeutics include a much broader range of targets. CRISPR-Cas9 is being explored to treat cancer, infectious disease, and more. This review highlights CRISPR-Cas9 clinical trials registered at clinicaltrials.gov as of 12/31/2024.
Keywords:
CRISPR-Cas9
; gene editing
; therapeutics
; clinical trials
; cancer
1. Introduction
Precise gene editing has been made possible by co-opting an adaptive immune system first identified in bacteria. “Clustered Regularly Interspaced Short Palindromic Repeats” (“CRISPR”) genomic regions store bits of foreign DNA, allowing the organism to swiftly recognize and respond if any of these foreign invaders return. The Cas9 endonuclease, guided by RNA transcribed from the CRISPR array, finds and cleaves the foreign genetic material, removing the threat (Yoshizumi et al. 2018). This RNA-targeted endonuclease system enabled development of a precise, programmable gene editing tool that holds immense promise for treating intractable diseases (Abbott 2016). Other CRISPR enzymes, such as Cas12a, are making their way into clinical trials. While Cas12a provides benefits over Cas9 in some situations, such as staggered doublestrand breaks that leaves overhangs leading to more consistent repair, and a motif recognition that works better in AT-rich sequence, it can also yield more off-target effects (Zetsche et al. 2015). Because the vast majority of CRISPR-based therapies currently in trials utilize the Cas9 enzyme, here we focus on CRISPR-Cas9-based therapies.
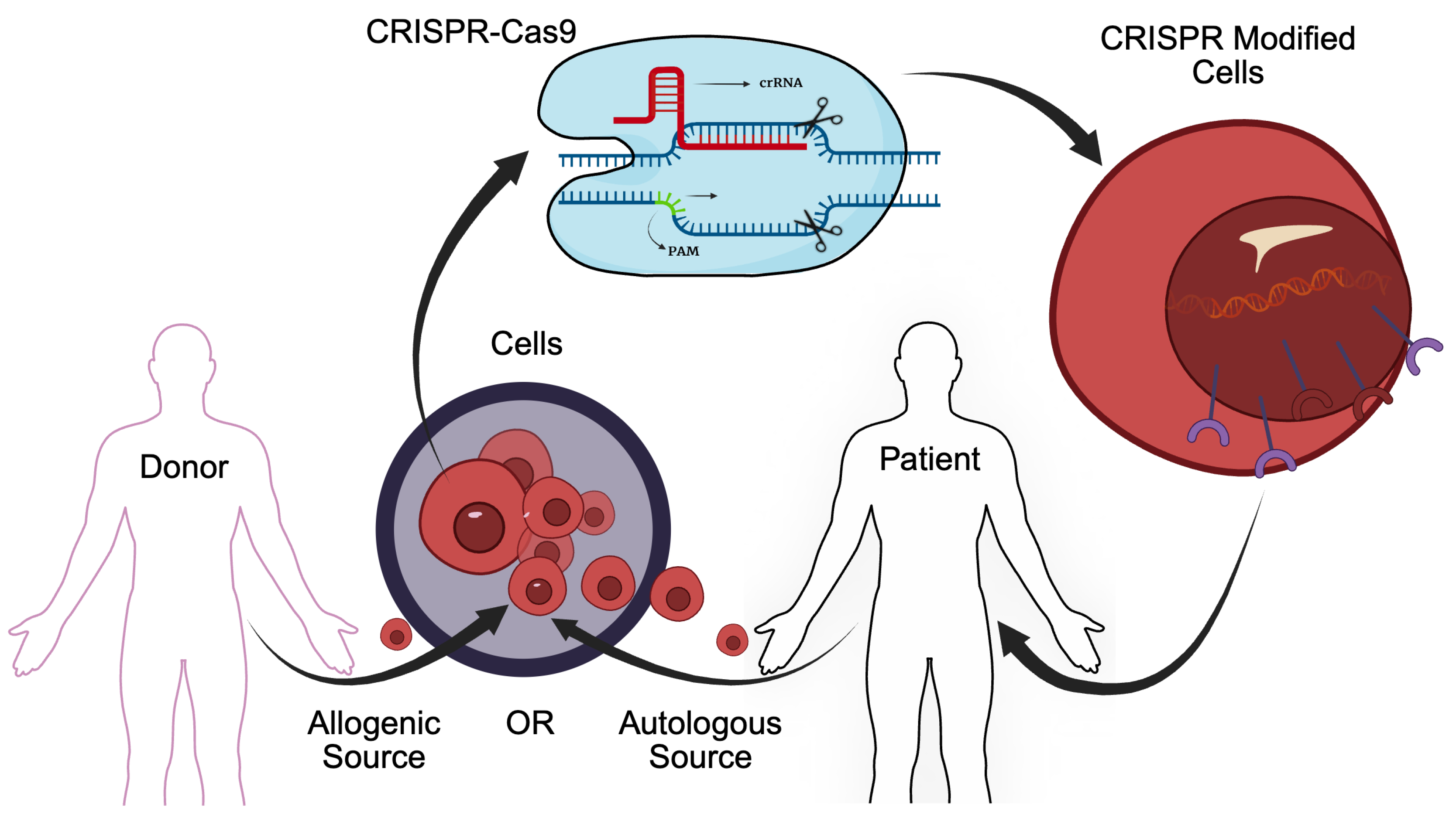
Early CRISPR-Cas9 therapies targeted blood disorders, harvesting CD-34+ hematopoietic stem and progenitor cells (HSPCs) from the patient (autologous) or from a donor (allogenic), modifying cells using CRISPR-Cas9, then (re)introducing the modified cells into the patient (Figure 1). Blood cells derived from the modified HSPCs to quickly become dominant as blood cells turn over. In addition to ex-vivo approaches that modify cells outside of a living organism, in-vivo CRISPR therapies can be injected directly into the patient.
This review explores complete and ongoing CRISPR-Cas9 clinical trials registered at clinicaltrials.gov by 12/31/2024 in any clinical trial phase (Table 1) (National Institutes of Health 2022). Emerging CRISPR-cas9 therapies show immense promise for combating intractable diseases.
2. First Approved CRISPR-Cas9 Therapy
In late 2023, the first CRISPR-Cas9-based gene editing therapy (CASGEVY™) gained FDA approval for sickle-cell disease (SCD) (FDA 2023). SCD is caused by a -globin (HBB) gene mutation, breaking the -subunit of adult hemoglobin (HbA; 2 - and 2 -subunits). This leads to sickle-shaped red blood cells (RBCs), reduced blood flow, and less efficient oxygen delivery. The gene editing does not fix the HBB mutation, but rather increases fetal hemoglobin (HbF) expression (2 - and 2 -subunits), avoiding the mutated -subunit. HbF binds oxygen more strongly and is less likely to cause sickling compared to HbA. To increase HbF, CRISPR-Cas9 breaks the BCL11A gene, which normally represses HbF production. This therapy is more tolerable, effective, and permanent than transfusions and transplants (Singh et al. 2024).
More recently, CASGEVY was approved for transfusion-dependent -thalassemia (TDT), also a result of an HBB mutation that results in insufficient amounts of -globin. CASGEVY treats TDT the same way as SCD, releasing the HbF block, thereby reducing transfusion requirements for TDT patients (Vertex Pharmaceuticals 2024). Ongoing CASGEVY (CTX001) TDT/SCD trials measure long-term success, including engraftment stability, HbF levels, maintenance of transfusion independence, and mitigation of severe vaso-occlusive crises (NCT05356195,NCT03655678,NCT05477563,NCT03745287,NCT05329649, NCT04208529,NCT05951205).
3. Additional CRISPR-Cas9 Therapies In Clinic Trials
Clinically-trialed CRISPR-Cas9 therapies target a broad range of diseases. While only a small portion of clinical trials are successful, the increasing number of therapies and diseases they treat in CRISPR-Cas9-based clinical trials bring hope for treating intractable diseases.
Hemoglobinopathies
Further clinical trials are underway for hemoglobinopathies (including SCD and TDT), diseases that result in too little hemoglobin, compromising oxygen delivery. BRL-101’s treatment of SCD/TDT also targets the BCL11A gene, disrupting BCL11A’s enhancer, thereby lowering transcription and increasing HbF production. With a safety profile similar to that of the required autologous transplantation, BRL-101 enables transfusion independence and increased HBF and HBA levels (NCT06287099,NCT06287086,NCT06300723,NCT05577312) (Fu et al. 2022,Fu et al. 2023). Other hemoglobinopathy clinical trials directly target the HBB mutation. For example, GPH101 edits HSPCs to reverse HBB’s valine to glutamic acid change in -thalassemia (NCT04819841) (Kanter et al. 2021).
Cancer
CRISPR-Cas9 therapies also combat cancers, often using Chimeric Antigen Receptor T cell (CAR-T) therapy, a type of immunotherapy increasingly employing CRISPR’s precision. In CAR-T therapy, T cells from patient (autologous) or donor (allogenic) and edited ex-vivo to recognize and kill cancer cells. A synthetic gene is inserted that encodes a chimeric antigen receptor (CAR), which includes an antigen binding domain targeting cancer cell surface proteins. These cancer cell surface antigens include various Cluster of Differentiation (CD) genes, which create important functional proteins on the surface of white blood cells (Zhang et al. 2017). These CD proteins are targeted by CARs in cancers derived from white blood cells such as leukemias, lymphomas and myelomas.
Other antigen genes currently being targeted include B-cell Maturation Antigen (BCMA), which is essential for B cell maturation, survival, and proliferation and is overly expressed in multiple myeloma, a B cell-derived cancer (NCT04244656) (Rinaldi et al. 2022). The mutation or overexpression of Epidermal Growth Factor Receptor (EFGR) is often seen in cancers, increasing uncontrolled cell growth, making it another important antigen target in some cancers (NCT04976218) (Sasaki et al. 2013). An important target for mesotheliomas, cancers derived from the lining of different organs, is the mesothelin gene. This is an especially attractive target because this gene’s expression is limited to mesothelial cells, a cell type that is dispensible (NCT03545815,NCT03747965,NCT05812326) (Hassan et al. 2016).
CAR-T therapy has been improved by manipulating additional genes beyond the CAR insertion, increasing it’s effectiveness and longevity. Most CAR-T therapies disrupt the T cell receptor (TRAC) gene by inserting the CAR into it, which has the added advantage of ensuring uniform CAR expression. Without the -subunit, the T cell receptor (TCR) is not functional. This increases therapy effectiveness by reducing spontaneous activation and differentiation of the modified T cells, avoiding T cell exhaustion. In addition, the lack of the TCR protein, which normally recognizes foreign material, helps to avoid graft vs host disease (GvHD), opening up CAR-T therapy to allogenic sources, reducing costs and timelines, and standardizing treatment (Eyquem et al. 2017,Lonez and Breman 2024,Terrett et al. 2023).
CRISPR Therapeutics’s allogenic CAR-T therapies are being improved by editing additional genes beyond the CAR insertion and its disruption of TRAC. CTX110 (NCT04035434), targeting CD19+ cancers (B cell leukemias and lymphomas), and CTX130 (NCT04502446 and NCT04438083), targeting CD70+ cancers (T cell lymphomas and renal cell carcinomas), both knockout the 2M gene, an MCHI subunit. The broken MHCI protein prevents donor CAR-T cells from being recognized and destroyed by the patient’s immune system (host vs. graft disease (HvGD) (McGuirk et al. 2022,Terrett et al. 2023).
The next generation versions of these drugs, CTX112 for CD19+ cancers (NCT05643742) and CTX131 (NCT06492304) for CD70+ cancers, improve on their counterparts through additional gene knockouts. Regnase-1 normally tamps down on cytokine secretion and, by extension, the immune system. The Regnase-1 knockout, therefore, keeps the immune response strong. Likewise, Transforming Growth Factor-beta (TGF-) receptor type 2 (TFGBR2) knockouts create a CAR-T cell without a receptor to recognize the (TGF-) produced in the tumor microenvironment that would normally inhibit the T cell. In CTX131, CD70 is also knocked out, preventing fratricide in CD70-targeting CAR-T cells. The improvements are stark. For example, CTX112 is 10X more potent than CTX110 with imrpovements in persistance and anti-tumor effects (Kalaitzidis et al. 2023).
Further genes been identified whose disruption in CAR-T cells can lead to therapeutic improvements (Feng et al. 2024,Moradi et al. 2024). Knockout of the Programmed Cell Death Protein 1 (PDCD-1) gene can help keep anti-tumor activity strong and avoid immune suppression and T cell exhaustion. PDCD-1 creates the PD-1 protein. When PD-1 binds its ligand (PD-L1), it acts as a brake, inactivating T cells. Tumors take advantage by overexpressing PD-L1, allowing them to inactivate immune cell that recognized the cancer, thereby evading the antitumor immune response and leading to T cell exhaustion (Moradi et al. 2024,Munari et al. 2021). Several clinical trials (NCT03545815, NCT03747965, NCT05812326) deploy PDCD-1 knockout CAR-T cells against mesothelin+ breast and other solid tumors. In one case (NCT03747965), the GC008t therapy stabilized disease in 4 patients and acheived tumor shrinkage for 2 patients, though engraftment could be improved (Wang et al. 2020).
One autologous clinical trial (NCT05566223) uses CRISPR-Cas9 technology to knockout the CISH (Cytokine-induced SH2 protein) gene in tumor infiltrating lymphocytes (TILs), a type of T cell that can penetrate solid tumors. CISH limits T cell activation and signaling, so its disruption keeps anti-tumor responses high. This therapy treats non-small cell lung cancer (NSCLC), which accounts for about 85 percent of all diagnosed lung cancers, with lung cancers being the leading cause of cancer-related deaths globally (Gridelli et al. 2015).
CRISPR-Cas9 is also being used to altering CAR-T cells in order to make concurrent treatment with a monoclonal antibodies possible. One autologous therapy (NCT05662904) treats ALL by inactivating the CD33 gene in the patient’s HSPCs to make them immune to the CD33-specific antibody-drug conjugate Gemtuzumab-ozogamicin (GO), allowing for escalating doses of GO to be administered. (Godwin et al. 2017) In another study, PBLTT52CAR19 targets CD19+ pediatric B cell acute lymphoblastic leukemia (ALL) (NCT04557436). In addition to disrupting the TRAC gene, the CD52 gene was also disrupted, allowing the concurrent use of Alemtuzumab (Drugs.com 2024), an anti-CD52 monoclonal therapy. Four out of six patients infused with the CAR-T cells showed CAR-T cell proliferation, achieved remission, and then received allogenic stem cell transplantation for a more permenant therapy. (Ottaviano et al. 2022)
CRISPR-cas9 editing can also introduce safety switches into cancer therapies to avoid serious side effects, including Cytokinin Release Syndrome (CRS) and immune cell-associated neurotoxicity syndrome (ICANS) (Xiao et al. 2021). CT125A is an autologous CAR-T cell therapy that targets CD5+ hematologic malignancies, including T cell-derived leukemias and lymphomas (NCT04767308). The endogenous CD5 gene was disrupted using CRISPR-Cas9 to avoid fratricide. A safety switch was added to the CAR-T cells by editing a truncated epidermal growth factor receptor (tEGFR) into the genome. The resulting receptor, though not functional, was still recognized by Cetuximab, a monoclonal antibody therapy that targets the receptor, killing the CAR-T cells when administered to the patient (Chidharla et al. 2023). Clinical outcomes were both positive and negative. All three patients achieved at least partial remission but one patient (complete remission) died of sepsis and multi-organ dysfunction. The other two patients achieved partial remission but one patient subsequently relapsed. As expected, the therapy caused CRS, which was reversed following administration of cetuximab, allowing for this toxic treatment to be limited in time, though CAR-T cells were not completely eliminated. Nevertheless, this study showed that saftey switches can be viable strategies for limiting patient exposure to therapies with dangerous side effects (Lin et al. 2024).
Improvements upon CARs are being tested. One of these is STAR (Synthetic TCR and Antigen Receptor) T cell therapy. STAR-T therapy uses a construct that mimics TCR, increasing sensitivity to the cancer-presented antigens, which is especially important in solid tumors with low antigen density (Huang et al. 2024). Two related studies (NCT05631912:autologous and NCT06321289: allogenic) are trialing CD19-targeting STAR-T therapy for B cell non-Hodgkin’s lymphoma. Additional CRISPR-Cas9 editing knocked out TRAC, PDCD-1, human leukocyte antigen (HLA)-A/B, and Class II Transactivator (CIITA) genes to strengthen the intervention. In addition to reducing the immune suppression, delaying T cell exhaustion, and increasing anti-tumor activity with the TRAC and PDCD-1 knockouts, knockouts of HLA-A/B and CIITA, which are subunits of MHCI and MHCII proteins, respectivly, reduce the recognition of allogenic STAR-T cells as foreign, thereby reducing the risk of GvHD.
This review describes some of many promising CRISPR-Cas9-based cancer therapies and strategies. The number of antigen targets is expanding, additional constructs are improving on CARs, and therapies are becoming more sophisticated with additional gene edits to improve longevity and safety and keep immune and anti-tumor functions high.
Infectious Disease
CRISPR-Cas9 can also be utilized to fight infectious disease, either by targeting host or pathogen genes. Two clinical trials explore unique methods to treat Acquired Immunodeficiency Syndrome (AIDs), caused by human immuodeficiency virus I (HIV-1). These therapies target the host CCR5 (CC chemokine receptor 5) gene, which is one of the co-receptors that HIV-1 uses to enter the host’s CD4+ lymphocytes, thereby destroying a critical part of the host’s immune function. A frameshifting 32-nt deletion in CCR5 occurs naturally in a small proportion of the human population. This CCR5-32 mutation, when homozygous, prevents HIV-1 from entering the cell, allowing infected individuals (termed "HIV controllers") to live with the virus (Carrington et al. 1997,Oppermann 2004).
One allogenic study (NCT03164135) used CRISPR-Cas9 to modify donor HSPCs, ablating the CCR5 receptor to make the immune cells resistant to HIV-1. This study was designed for HIV patients who also had a hematologic malignancy that required stem cell transplantation, creating an opportunity to simultaneously test CCR5 ablation with minimal additional risk to the patient. One HIV-positive patient in this study had ALL. Transplantation and long-term engraftment was achieved, however, CCR5 was disrupted in only 5 percent of lymphocytes (Xu et al. 2019).
Another AIDS therapy, EBT-101, uses CRISPR-Cas9 to disrupt the HIV-1 genome in aviremic patients (patients that have latent infections without detectable blood virus levels (NCT05144386). Initial results met safety benchmarks and temporarily suppressed viral reservoirs (Johnson 2024).
Other viruses are also being targeted with CRISPR-Cas9 therapies. Persistent human papillomavirus (HPV) infection is the major cause of cervical cancer. The viral E6 and E7 oncoproteins inactivate host tumor suppressor genes promoting uncontrolled cell growth (Narisawa-Saito and Kiyono 2007). Although small interfering RNA targeting of these oncogenes may temporarily inhibit HPV, it does not destroy the viral genes (Hu et al. 2015). Administration of a transcription activator-like effector nucleases (TALENs) or a CRISPR-Cas9 E6/E7-targeting plasmid in a gel reduced E6/E7 DNA and expression, initiated cell death, and prevented tumor growth (NCT03057912) (Hu and Team 2025).
SARS-CoV-2 is a virus causing COVID-19, and is targeted in a study that uses CRISPR-Cas9 to ablate the host PDCD1 and ACE2 receptor genes in CD8+ virus-reactive memory T cells (NCT04990557). PDCD-1 was knocked out because its upregulation during COVID-19 infection, even in patients with mild symptoms, promotes T-cell exhaustion. Knocking out the ACE2 receptor removes SARS-CoV-2’s main entry path into the modified T cell (Scialo et al. 2020).
CRISPR-Cas9 therapies also target bacterial pathogens. One study uses CRISPR-Cas9 to disrupt virulence and -lactam antibiotic resistance genes in Enterobacteriaceae genomes (NCT05850871).
Eye diseases
CRISPR-Cas9 therapies work well for eye diseases because they can be injected directly into relevant eye tissue. In Intraocular Hypertensive Primary Open Angle Glaucoma (POAC), increased intraocular pressure damages the optic nerve, leading to blindness (Quigley et al. 1983). Dominant mutations in the cytosketetal myocilin (MYOC) gene, which is expressed in the trabecular meshwork where intraocular pressure is regulated, can cause POAC. The therapy (BD113) is delivered in a virus-like particle (VLP) by eye injection. It aims to knockdown or knockout the mutated MYOC gene, reducing the amount of mutated protein (NCT06465537).
Another VLP therapy (BD111) is injected into the cornea to treat recalcitrant herpes stromal keratitis, which can cause infectious blindness (NCT04560790). The therapy uses CRISPR-Cas9 to disrupt the herpes simplex virus type 1 (HSV-1) genome. No HSV-1 was detected follow-ups, averaging 18 months (Wei et al. 2023).
Reinitis pigmentosa results in rod cell loss, leading to night blindness, and the gradual loss of cone cells, leading to tunnel vision or blindness. The therapy, ZVS203e, is administered by subretinal injection and fixes a causal rhodopsin (RHO) gene mutation to create a functional protein that is activated under low light conditions (NCT05805007) (Nathans and Hogness 1984,National Library of Medicine 2025).
Another CRISPR-Cas9 therapy (EDIT-101) targets Leber Congenital Amaurosis 10 (LCA10) (NCT03872479). A homozygous mutation in the centrosomal protein 290 (CEP290) gene (or a heterozygous compound mutation) causes retinal degeneration leading to blindness or severe vision loss at birth or shortly thereafter (den Hollander et al. 2006). The mutation causes an additional splice site that forms a cryptic (additional) exon. Initial clinical trial results established safety and 75% of particpants showed measureably improved vision.
Other Conditions
Hemophilia B a bleeding disorder caused by a mutated coagulation Factor IX (FIX) gene that results in insufficient FIX (Kurachi and Kurachi 2000). CRISPR-Cas9-based therapies insert wildtype FIX gene into liver and B cells, enabling clotting factor production (NCT06379789,NCT06611436).
Hereditary Angioedema (HAE) results in debilitating or fatal swelling under the skin. Treatments target kallikrein, a protease encoded by the KLB1 gene, which causes swelling when overproduced in blood plasma (Banerji et al. 2017,Longhurst et al. 2022). NTLA-2002 is a CRISPR-Cas9-based therapy that disrupts KLB1 in liver cells, reducing plasma kallikrein levels. (Longhurst et al. 2022). Initial results established safety and showed a reduction in plasma kallikrein levels (NCT05120830,NCT06634420).
4. Discussion
CRISPR is a rapidly developing gene editing-tool revolutionizing research and clinical applications. The recent FDA approval of the first CRISPR-based therapy and the number of CRISPR-Cas9-based therapies in the clinical trial pipeline, promise transformative therapies on the horizon. Indeed, a recent example highlights the power of CRISPR-Cas9-based therapies. In May 2025, a research team supported by the National Institutes of Health developed and successfully delivered a personalized CRISPR-Cas9-based therapy to treat an infant born with a mutation in the carbamoyl-phosphate synthetase 1 (CPS1) gene. This meant he could not break down byproducts of protein metabolism within the liver, leading to ammonia toxicity. The team designed a patient-specific base-editing therapy to correct the mutation and administered it via lipid nanoparticles. The infant received two doses at ages 7 and 8 months. The patient tolerated high dietary protein even while cutting the dose of the initial nitrogen-scavenger drug in half with no severe adverse effects (Musunuru et al. 2025). Customized CRISPR-based therapies are expensive and not widely accessible. But autologous strategies that work across many patients with the same or similar diseases, and allogenic therapies that use donor cells to provide “off-the-shelf” solutions bring hope for treating currently intractable conditions.

Table 1.
CRISPR-Cas9 clinical trials. All study numbers in bold are outlined within the review.
| Disease Category | Intervention | Study Numbers |
|---|---|---|
| Hemoglobinopathies | CTX001 | NCT05356195, NCT03655678, NCT05477563, NCT03745287, NCT05329649, NCT04208529, NCT05951205, NCT06287099 |
| BRL-101 | NCT06287086, NCT06300723, NCT05577312 | |
| ET-01 | NCT04925206 | |
| Plerixafor + Busulfan + Gene-modified CD34+ Cells | NCT06506461 | |
| CRISPR-SCD001 | NCT04774536 | |
| nula-cel Drug Product | NCT04819841 | |
| OTQ923 | NCT04443907 | |
| Hematologic Malignancies | CTX131 | NCT06492304, NCT04502446 |
| CTX110 | NCT04035434 | |
| Universal Dual Specificity CD19 and CD20 or CD22 CAR-T Cells | NCT03398967 | |
| CTX112 | NCT05643742 | |
| UCART019 | NCT03166878 | |
| CT125A cells + Cyclophosphamide + fludarabine | NCT04767308 | |
| PBLTT52CAR19 | NCT04557436 | |
| Donor-derived CD34+ HSC with CRISPR/Cas9-mediated CD33 deletion + emtuzumab Ozogamicin | NCT05662904 | |
| REGV131 + LNP1265 | NCT06379789 | |
| BE-101 | NCT06611436 | |
| NTLA-2002 + Normal Saline IV Administration | NCT06634420 | |
| Biological NTLA-2002 + Normal Saline IV Administration | NCT05120830 | |
| CTX120 | NCT04244656 | |
| Solid Tumor | Anti-mesothelin CAR-T cells | NCT03545815 |
| Mesothelin-directed CAR-T cells | NCT03747965 | |
| TGFR-KO CAR-EGFR T Cells | NCT04976218 | |
| MT027 cells suspension | NCT06726564 | |
| Autologous CD19-STAR-T cell + Fludarabine + Cyclophosphamide | NCT05631912 | |
| Allogeneic CD19-STAR T cell + Fludarabine + Cyclophosphamide | NCT06321289 | |
| TRAC and Power3 Genes Knock-out Allogeneic CD19-targeting CAR-T cell (ATHENA CAR-T) + Fludarabine + Cyclophosphamide | NCT06014073 | |
| CTX131 | NCT05795595 | |
| Fludarabine + Cyclophosphamide + CISH Inactivated TIL + Aldesleukin + Pembrolizumab | NCT05566223 | |
| Transcatheter arterial chemoembolization|BIOLOGICAL: PD-1 knockout engineered T cells | NCT04417764 | |
| Fludarabine + Cyclophosphamide + Interleukin-2 | NCT03044743 | |
| Cyclophosphamide + PD-1 Knockout T Cells | NCT02793856 | |
| MT027 cells suspension | NCT06742593, NCT06737146 | |
| Cyclophosphamide + Fludarabine + Tumor-Infiltrating Lymphocytes (TIL) + Aldesleukin | NCT04426669 | |
| PD-1 Knockout T Cells | NCT03081715 | |
| AJMUC1- PD-1 gene knockout anti-MUC1 CAR-T cells | NCT05812326 | |
| Infectious Disease | CAZ/AVI plus Aztreonam + Conventional treatment | NCT05850871 |
| CCR5 gene modification | NCT03164135 | |
| EBT-101 | NCT05144386 | |
| TALEN + CRISPR/Cas9 | NCT03057912 | |
| PD-1 and ACE2 Knockout T Cells + PD-1 and ACE2 Knockout T Cells + PD-1 and ACE2 Knockout T Cells | NCT04990557 | |
| Ophthalmic Disorders | BD113vVLP | NCT06465537 |
| BD111 Adult single group Dose | NCT04560790 | |
| ZVS203e | NCT05805007 | |
| EDIT-101 | NCT03872479 | |
| Other Conditions | VCTX211 | NCT05565248 |
| VCTX210A unit | NCT05210530 | |
| NTLA-2001 | NCT04601051 | |
| Conclusive genetic testing + Genotype-phenotype correlation for personalized diagnosis + Personalized study of variants of uncertain clinical significance (VUS) through functional studies on 3D organ-on-a-chip | NCT06325072 |
Figure 1.
CRISPR-Cas9 therapies often involve removing cells from the patient’s body (autologous therapies) or obtaining cells from a donor (allogenic therapies), applying the therapy to alter the target gene, and (re)introducing the modified cells into the patient. The figure illustrates CAR-T therapy, often used in cancer but other CRISP-Cas9 therapies that edit cells outside of the body have a similar workflow.
Figure 1.
CRISPR-Cas9 therapies often involve removing cells from the patient’s body (autologous therapies) or obtaining cells from a donor (allogenic therapies), applying the therapy to alter the target gene, and (re)introducing the modified cells into the patient. The figure illustrates CAR-T therapy, often used in cancer but other CRISP-Cas9 therapies that edit cells outside of the body have a similar workflow.
Author Contributions
AB was responsible for identifying and researching the CRISPR-Cas9 clinical trials and wrote the manuscript. JM conceived of the idea, mentored AB, and edited the manuscript.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Abbott, Alison. 2016. A crispr vision. Nature 532(7600), 432–434.
- Banerji, A.; Busse, P.; Shennak, M.; Lumry, W.; Davis-Lorton, M.; Wedner, H.J.; Jacobs, J.; Baker, J.; Bernstein, J.A.; Lockey, R.; et al. Inhibiting Plasma Kallikrein for Hereditary Angioedema Prophylaxis. New Engl. J. Med. 2017, 376, 717–728. [CrossRef]
- Carrington, M.; Kissner, T.; Gerrard, B.; Ivanov, S.; O'Brien, S.J.; Dean, M. Novel Alleles of the Chemokine-Receptor Gene CCR5. Am. J. Hum. Genet. 1997, 61, 1261–1267. [CrossRef]
- Chidharla, Anusha, Meghana Parsi, and Anup Kasi. 2023, May. Cetuximab. StatPearls [Internet]. https://www.ncbi.nlm.nih.gov/books/NBK459293/.
- den Hollander, A.I.; Koenekoop, R.K.; Yzer, S.; Lopez, I.; Arends, M.L.; Voesenek, K.E.; Zonneveld, M.N.; Strom, T.M.; Meitinger, T.; Brunner, H.G.; et al. Mutations in the CEP290 (NPHP6) Gene Are a Frequent Cause of Leber Congenital Amaurosis. Am. J. Hum. Genet. 2006, 79, 556–561. [CrossRef]
- Drugs.com. 2024. Alemtuzumab (multiple sclerosis) (monograph). https://www.drugs.com/monograph/alemtuzumab-multiple-sclerosis.html.
- Eyquem, J.; Mansilla-Soto, J.; Giavridis, T.; van der Stegen, S.J.C.; Hamieh, M.; Cunanan, K.M.; Odak, A.; Gönen, M.; Sadelain, M. Targeting a CAR to the TRAC locus with CRISPR/Cas9 enhances tumour rejection. Nature 2017, 543, 113–117. [CrossRef]
- FDA. 2023. FDA approves first gene therapies to treat patients with sickle cell disease. https://www.fda.gov/news-events/press-announcements/fda-approves-first-gene-therapies-treat-patients-sickle-cell-disease.
- Feng, X.; Li, Z.; Liu, Y.; Chen, D.; Zhou, Z. CRISPR/Cas9 technology for advancements in cancer immunotherapy: from uncovering regulatory mechanisms to therapeutic applications. Exp. Hematol. Oncol. 2024, 13, 1–19. [CrossRef]
- Fu, Bin, Jiaoyang Liao, Shuanghong Chen, Wei Li, Qiudao Wang, Jian Hu, Fei Yang, Shenlin Hsiao, Yanhong Jiang, Liren Wang, et al. 2022. Crispr–cas9-mediated gene editing of the bcl11a enhancer for pediatric β0/β0 transfusion-dependent β-thalassemia. Nature medicine 28(8), 1573–1580.
- Fu, B.; Zhang, X.; Wang, L.; Liao, J.; Chen, S.; Zheng, B.; Li, W.; Wang, F.; Li, D.; Liu, M.; et al. S271: AN UPDATED FOLLOW-UP OF BRL-101, CRISPR-CAS9-MEDIATED GENE EDITING OF THE BCL11A ENHANCER FOR TRANSFUSION-DEPENDENT BETA-THALASSE. HemaSphere 2023, 7, e406095b. [CrossRef]
- Godwin, C.D.; Gale, R.P.; Walter, R.B. Gemtuzumab ozogamicin in acute myeloid leukemia. Leukemia 2017, 31, 1855–1868. [CrossRef]
- Gridelli, Cesare, Antonio Rossi, David P Carbone, Juliana Guarize, Niki Karachaliou, Tony Mok, Francesco Petrella, Lorenzo Spaggiari, and Rafael Rosell. 2015. Non-small-cell lung cancer. Nature reviews Disease primers 1(1), 1–16.
- Hassan, R.; Thomas, A.; Alewine, C.; Le, D.T.; Jaffee, E.M.; Pastan, I. Mesothelin Immunotherapy for Cancer: Ready for Prime Time? J. Clin. Oncol. 2016, 34, 4171–4179. [CrossRef]
- Hu, Zheng, Wencheng Ding, Da Zhu, Lan Yu, Xiaohui Jiang, Xiaoli Wang, Changlin Zhang, Liming Wang, Teng Ji, Dan Liu, Dan He, Xi Xia, Tao Zhu, Juncheng Wei, Peng Wu, Changyu Wang, Ling Xi, Qinglei Gao, Gang Chen, Rong Liu, Kezhen Li, Shuang Li, Shixuan Wang, Jianfeng Zhou, Ding Ma,, and Hui Wang. 2015. Talen-mediated targeting of hpv oncogenes ameliorates hpv-related cervical malignancy. The Journal of clinical investigation 125(1), 425–436.
- Hu, Zheng and CRISPR Medicine News Editorial Team. 2025, May. A safety and efficacy study of transcription activator-like effector nucleases and crispr/cas9 in the treatment of hpv-related cervical intraepithelial neoplasia i (nct03057912). CRISPR Medicine News. Summarizes trial design, methods (CRISPR & TALEN targeting HPV E6/E7), plasmid-gel delivery, enrollment and status.
- Huang, D.; Li, Y.; Rui, W.; Sun, K.; Zhou, Z.; Lv, X.; Yu, L.; Chen, J.; Zhou, J.; Liu, V.; et al. TCR-mimicking STAR conveys superior sensitivity over CAR in targeting tumors with low-density neoantigens. Cell Rep. 2024, 43. [CrossRef]
- Johnson, Victoria. 2024, May. Crispr-editing ebt-101 therapy safe, temporarily suppresses hiv infection.
- Kalaitzidis, Demetrios, Mohammed Ghonime, Robert Chain, Nivedita Jaishankar, Davis Settipane, Zinkal Padalia, Lauren Zakas, Meghna Kuppuraju, Paul Tetteh, Mary-Lee Dequeant, et al. 2023. 274 development of ctx112 a next generation allogeneic multiplexed crispr-edited cart cell therapy with disruptions of the tgfbr2 and regnase-1 genes for improved manufacturing and potency. Journal for ImmunoTherapy of Cancer. [CrossRef]
- Kanter, J.; DiPersio, J.F.; Leavey, P.; Shyr, D.C.; A Thompson, A.; Porteus, M.H.; Intondi, A.; Lahiri, P.; Dever, D.P.; Petrusich, A.; et al. Cedar Trial in Progress: A First in Human, Phase 1/2 Study of the Correction of a Single Nucleotide Mutation in Autologous HSCs (GPH101) to Convert HbS to HbA for Treating Severe SCD. Blood 2021, 138, 1864–1864. [CrossRef]
- Kurachi, S.; Deyashiki, Y.; Takeshita, J.; Kurachi, K. Genetic Mechanisms of Age Regulation of Human Blood Coagulation Factor IX. Science 1999, 285, 739–743. [CrossRef]
- Lin, H.; Cheng, J.; Zhu, L.; Zeng, Y.; Dai, Z.; Zhang, Y.; Zhu, X.; Mu, W. Anti-CD5 CAR-T cells with a tEGFR safety switch exhibit potent toxicity control. Blood Cancer J. 2024, 14, 1–5. [CrossRef]
- Lonez, C.; Breman, E. Allogeneic CAR-T Therapy Technologies: Has the Promise Been Met?. Cells 2024, 13, 146. [CrossRef]
- Longhurst, H.; Fijen, L.; Lindsay, K.; Butler, J.; Golden, A.; Maag, D.; Xu, Y.; Cohn, D. IN VIVO CRISPR/CAS9 EDITING OF KLKB1 IN PATIENTS WITH HEREDITARY ANGIOEDEMA: A FIRST-IN-HUMAN STUDY. Ann. Allergy, Asthma Immunol. 2022, 129, S10–S11. [CrossRef]
- McGuirk, J.P.; Tam, C.S.; Kröger, N.; Riedell, P.A.; Murthy, H.S.; Ho, P.J.; Maakaron, J.E.; Waller, E.K.; Awan, F.T.; Shaughnessy, P.J.; et al. CTX110 Allogeneic CRISPR-Cas9-Engineered CAR T Cells in Patients (Pts) with Relapsed or Refractory (R/R) Large B-Cell Lymphoma (LBCL): Results from the Phase 1 Dose Escalation Carbon Study. Blood 2022, 140, 10303–10306. [CrossRef]
- Moradi, V.; Khodabandehloo, E.; Alidadi, M.; Omidkhoda, A.; Ahmadbeigi, N. Progress and pitfalls of gene editing technology in CAR-T cell therapy: a state-of-the-art review. Front. Oncol. 2024, 14, 1388475. [CrossRef]
- Munari, E.; Mariotti, F.R.; Quatrini, L.; Bertoglio, P.; Tumino, N.; Vacca, P.; Eccher, A.; Ciompi, F.; Brunelli, M.; Martignoni, G.; et al. PD-1/PD-L1 in Cancer: Pathophysiological, Diagnostic and Therapeutic Aspects. Int. J. Mol. Sci. 2021, 22, 5123. [CrossRef]
- Musunuru, K.; Grandinette, S.A.; Wang, X.; Hudson, T.R.; Briseno, K.; Berry, A.M.; Hacker, J.L.; Hsu, A.; Silverstein, R.A.; Hille, L.T.; et al. Patient-Specific In Vivo Gene Editing to Treat a Rare Genetic Disease. New Engl. J. Med. 2025, 392, 2235–2243. [CrossRef]
- Narisawa-Saito, M.; Kiyono, T. Basic mechanisms of high-risk human papillomavirus-induced carcinogenesis: Roles of E6 and E7 proteins. Cancer Sci. 2007, 98, 1505–1511. [CrossRef]
- Nathans, J.; Hogness, D.S. Isolation and nucleotide sequence of the gene encoding human rhodopsin. Proc. Natl. Acad. Sci. USA 1984, 81, 4851–4855. [CrossRef]
- National Institutes of Health. 2022. The basics. https://www.nih.gov/health-information/nih-clinical-research-trials-you/basics.
- National Library of Medicine. 2025. Rho gene: Medlineplus genetics. https://medlineplus.gov/genetics/gene/rho/#conditions.
- Oppermann, M. Chemokine receptor CCR5: insights into structure, function, and regulation. Cell. Signal. 2004, 16, 1201–1210. [CrossRef]
- Ottaviano, G.; Georgiadis, C.; Gkazi, S.A.; Syed, F.; Zhan, H.; Etuk, A.; Preece, R.; Chu, J.; Kubat, A.; Adams, S.; et al. Phase 1 clinical trial of CRISPR-engineered CAR19 universal T cells for treatment of children with refractory B cell leukemia. Sci. Transl. Med. 2022, 14, eabq3010. [CrossRef]
- Quigley, H.A.; Hohman, R.M.; Addicks, E.M.; Massof, R.W.; Green, W.R. Morphologic Changes in the Lamina Cribrosa Correlated with Neural Loss in Open-Angle Glaucoma. Arch. Ophthalmol. 1983, 95, 673–691. [CrossRef]
- Rinaldi, I.; Muthalib, A.; Edina, B.C.; Wiyono, L.; Winston, K. Role of Anti-B-Cell Maturation Antigen (BCMA) in the Management of Multiple Myeloma. Cancers 2022, 14, 3507. [CrossRef]
- Sasaki, T.; Hiroki, K.; Yamashita, Y. The Role of Epidermal Growth Factor Receptor in Cancer Metastasis and Microenvironment. BioMed Res. Int. 2013, 2013, 1–8. [CrossRef]
- Scialo, F.; Daniele, A.; Amato, F.; Pastore, L.; Matera, M.G.; Cazzola, M.; Castaldo, G.; Bianco, A. ACE2: The Major Cell Entry Receptor for SARS-CoV-2. Lung 2020, 198, 867–877. [CrossRef]
- Singh, A.M.; Irfan, H.M.; Fatima, E.M.; Nazir, Z.M.; Verma, A.M.; Akilimali, A. Revolutionary breakthrough: FDA approves CASGEVY, the first CRISPR/Cas9 gene therapy for sickle cell disease. Ann. Med. Surg. 2024, 86, 4555–4559. [CrossRef]
- Terrett, JA, D Kalaitzidis, ML Dequeant, S Karnik, M Ghonime, C Guo, et al. 2023. Ctx112 and ctx131: next-generation crispr/cas9-engineered allogeneic (allo) car t cells incorporating novel edits that increase potency and efficacy in the treatment of lymphoid and solid tumors. Cancer Res 83, 7_Suppl.
- Vertex Pharmaceuticals. 2024. Vertex Announces US FDA Approval of CASGEVY™ (exagamglogene autotemcel) for the Treatment of Transfusion-Dependent Beta Thalassemia. https://investors.vrtx.com/news-releases/news-release-details/vertex-announces-us-fda-approval-casgevytm-exagamglogene.
- Wang, Z.; Chen, M.; Zhang, Y.; Liu, Y.; Yang, Q.; Nie, J.; Shen, L.; Jiang, P.; He, J.; Ye, X.; et al. Phase I study of CRISPR-engineered CAR-T cells with PD-1 inactivation in treating mesothelin-positive solid tumors.. J. Clin. Oncol. 2020, 38, 3038–3038. [CrossRef]
- Wei, A.; Yin, D.; Zhai, Z.; Ling, S.; Le, H.; Tian, L.; Xu, J.; Paludan, S.R.; Cai, Y.; Hong, J. In vivo CRISPR gene editing in patients with herpetic stromal keratitis. Mol. Ther. 2023, 31, 3163–3175. [CrossRef]
- Xiao, X.; Huang, S.; Chen, S.; Wang, Y.; Sun, Q.; Xu, X.; Li, Y. Mechanisms of cytokine release syndrome and neurotoxicity of CAR T-cell therapy and associated prevention and management strategies. J. Exp. Clin. Cancer Res. 2021, 40, 1–23. [CrossRef]
- Xu, L.; Wang, J.; Liu, Y.; Xie, L.; Su, B.; Mou, D.; Wang, L.; Liu, T.; Wang, X.; Zhang, B.; et al. CRISPR-Edited Stem Cells in a Patient with HIV and Acute Lymphocytic Leukemia. N. Engl. J. Med. 2019, 381, 1240–1247. [CrossRef]
- Ishino, Y.; Krupovic, M.; Forterre, P. History of CRISPR-Cas from Encounter with a Mysterious Repeated Sequence to Genome Editing Technology. J. Bacteriol. 2018, 200, e00580-17. [CrossRef]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [CrossRef]
- Zhang, Cheng, Jun Liu, Jiang F Zhong, and Xi Zhang. 2017. Engineering car-t cells. Biomarker research 5(1), 22.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



