Submitted:
08 October 2025
Posted:
09 October 2025
You are already at the latest version
Abstract
Development of maize varieties with enhanced tolerance to drought stress has become a high-priority goal for maize breeding programs worldwide. In order to assess the variability of root and shoot traits in response to drought at an early vegetative stage, a set of 32 maize single-cross hybrids available on the Croatian market was grown under polyethylene glycol 8000 induced drought stress and well-watered control treatments. The induced drought stress negatively affected the growth and development of the maize seedlings. Significant differences among the hybrids, water treatments, and their interaction were found for the four directly measured traits (root and shoot length, and root and shoot fresh weights), as well as for the majority of four derived traits (root and shoot dry matter content, root-to-shoot length ratio, and root fresh weight to shoot fresh weight ratio). In the studied set of hybrids, heritabilities for the four directly measured traits were generally high, and the correlations among traits were strong under both water treatments, indicating their suitability for assessment of drought tolerance at the seedling stage.
Keywords:
maize
; root and shoot traits
; early vegetative stage
; drought resistance
; PEG
1. Introduction
Drought, a period without significant rainfall or a condition where water is limited, has been identified as the most common abiotic stress limiting crop performance worldwide in terms of growth and productivity [1]. It is also considered the most significant environmental factor affecting the productivity of maize (Zea mays L.) [2]. In terms of production quantity, maize is the most important cereal in the world [3], and, together with wheat and rice, provides at least 30% of food calories in developing countries [4]. The demand for maize is increasing as a source of food, oil and biofuel for the growing world population [5], with estimates suggesting that by 2050 the demand for maize in developing countries will almost double the current demand. During its life cycle, which lasts depending on the genotype from 80 to 150 days, maize requires 500–800 mm of water [6], which, according to Sah et al. [7], is near or below the critical level for achieving a good yield. Since 15% to 20% of maize grain yield is lost annually due to drought, with the prospect of further increases as droughts become more frequent and severe [4], developing maize varieties with enhanced tolerance to drought stress and higher water use efficiency has become a high-priority goal for breeding programs, both in the private and public sectors. Maize is most sensitive to water stress during pollination and the early grain-filling period [8,9], but drought is a multidimensional stress, affecting plants at various organizational levels over space and time, with complex and unpredictable physiological responses [4]. Denmead and Shaw [10] found that drought during the vegetative growth stage reduced yield by 25%, while Edmeades et al. [11], according to Akinwale et al. [8], stated that drought at the seedling and early vegetative stage may result in reduced crop establishment, zero yield, or even complete crop failure. Duvick [12] and Duvick and Cassman [13] showed that the change in yield potential on a per plant basis from older to newer hybrids was not significant when measured on non-stressed plants grown at very low plant density, but that the newer hybrids outperformed the older ones at higher plant densities. Imposing higher plant densities induces stress responses in maize and reduces yield on a per plant basis, but this is compensated by the increased number of plants per unit area, resulting in higher net yield [9]. Therefore, establishing high plant density at the seedling and early vegetative stage after germination is a prerequisite for successful maize production.
The seedling stage of maize begins immediately after emergence and continues until the 5-leaf stage, which occurs around 14 days after emergence. This stage is also referred to as the early vegetative stage, during which the plants are highly sensitive to environmental stress such as drought, which at this time can result in total crop failure [14]. Although earlier studies reported that seedling drought tolerance mechanisms are independent of drought responses during the flowering period [15], information on the response of maize genotypes to drought at the seedling and early vegetative stages is essential for achieving rapid progress in selection for drought tolerance in maize [8].
Under field conditions, it is not possible to accurately determine the effects of drought on root development [16]. In that regard, the application of polyethylene glycol (PEG) under controlled laboratory conditions has been used as an alternative to field trials for assessing maize drought stress resistance at germination and early vegetative stages [17,18,19,20,21,22,23,24,25,26]. PEG is considered an ideal regulator of water potential because its molecules, due to their high molecular weight (6000 and above), cannot penetrate cell walls [27]. This allows for precise assessment of the condition of a plant exposed to a specific osmotic potential [28]. These properties have made it the most widely used osmoticum to mimic decreases in soil water potential [29]. The use of PEG is suitable for screening large numbers of genotypes for drought tolerance at early stages [23] and can serve as a selection criterion for improving crop resistance to drought [20]. Phenotyping young seedlings under controlled conditions is a useful approach to identify candidate drought-tolerant genotypes [19], and can reduce, at least in part, the labor-intensive selection required under field conditions [30].
The aim of the present study was to investigate the variability of maize hybrids available on the Croatian market for root and shoot traits under PEG-induced drought stress at an early vegetative growth stage.
2. Materials and Methods
2.1. Plant Material and Experimental Design
A total of 32 single-cross maize hybrids available on the Croatian market were used as plant material. The hybrids belong to different developers and FAO groups, but their commercial names were deliberately omitted (Table S1). The hybrid seeds were sown in quartz sand in plastic containers with lids, and each container was watered with the same amount of water. The containers were covered for two days to prevent moisture loss and enhance germination, after which the lids were removed. Seeds were germinated in a growth chamber at 22 °C with a 16-hour day and 8-hour night photoperiod.
On the seventh day after sowing, seedlings, mostly at the one-leaf stage, were carefully separated and transplanted into plastic tubes with a diameter of 5 cm and a length of 40 cm, filled with vermiculite. Each tube had a piece of white water-permeable, polypropylene nonwoven agrotextile (BR Garden, Nottingham, UK) secured at the bottom with a rubber band to prevent vermiculite leakage. Before transplanting, 60 ml of water was added to each tube, followed by an additional 10 ml the next day and again on the second day after planting. The tubes were placed in trays filled with either half-strength Hoagland solution (0.8 g/L water, Phyto Technology Laboratories, Shawnee Mission, US) (control) or the same Hoagland solution supplemented with 6% polyethylene glycol (PEG) 8000 (Acros Organics, Geel, Belgium) to simulate drought conditions. The osmotic pressure of the PEG solution prepared in this way was -0.064 MPa (at 22 °C), calculated according to the formula of Michel [31]. For each tube, 300 ml of solution was added to the tray (determined sufficient in preliminary tests), and no further solution was added afterward. Through capillary action, the solution reached the top of the tubes. Plants were grown in a growth chamber at 22 °C, 60% relative humidity, under white light (fluorescent tubes), with a 16-hour day / 8-hour night photoperiod for 26 days (Figure 1). The experiment was set up as a completely randomized block design with three replicates (five plants/replicate) within each treatment (control and drought).
2.2. Trait Measurements
Twenty-six days after transplanting plants (including roots) were removed from the tubes, and the roots were carefully separated from the vermiculite using a wire mesh. Taproot length (RL) and shoot length (SL) were measured for each plant. Roots were thoroughly rinsed under tap water to remove all vermiculite and dried with paper towels. Root fresh weight (RFW) and shoot fresh weight (SFW) were measured on pooled samples of five plants per replicate. The shoot and root samples were then placed in paper bags and dried in an oven for 48 hours at 70 °C, after which root dry weight and shoot dry weight were determined. Root dry matter (RDM) and shoot dry matter (SDM) contents were calculated from the fresh and dry weights of root and shoot, and expressed as a percentage of RFW and SFW, respectively. The two derived traits, root-to-shoot length ratio (RL/SL) and root-to-shoot fresh weight ratio (RFW/SFW), were calculated from the measured traits.
2.3. Statistical Analysis
Analysis of variance (ANOVA) across treatments (control and drought) and within treatments was performed using the GLM procedure of SAS/STAT [32]. Variance components for the traits were determined by equating the observed mean squares from the within-treatment ANOVA to their expectations and resolving for the desired variance components. Broad-sense heritability (h2) was calculated using the following formula: h2= 2G / (σ2G + σ2ε/r), where 2G and σ2ε are the genotypic and error variances, respectively, and r is the number of replicates [33]. Pearson correlation coefficients between traits were calculated using PROC CORR from the SAS/STAT [32]. Correlogram figures between the pairs of traits under both experimental conditions, respectively, were obtained by using the corrplot() function in R version 4.4.2 [34] following the procedure described by Soetewey [35].
3. Results
3.1. Analysis of Variance
Analysis of variance across water treatments (control and drought) revealed a significant effect of genotype (G) for all studied traits (Table 1). Similarly, the effect of water treatment (T) was significant for all traits except the root-to-shoot length ratio (RL/SL).

Table 1.
Analysis of variance (ANOVA) for eight seedling traits of 32 maize hybrids grown under two water treatments (control and drought).
Table 1.
Analysis of variance (ANOVA) for eight seedling traits of 32 maize hybrids grown under two water treatments (control and drought).
| Trait | Treatment | ANOVA across Treatments | ANOVA by Treatment | |||
| Genotype (G) | Treatment (T) | G × T | Genotype | h2 | ||
| RL | Control | ** | ** | ** | ** | 0.61 |
| Drought | ** | 0.83 | ||||
| SL | Control | ** | ** | ** | ** | 0.88 |
| Drought | ** | 0.82 | ||||
| RFW | Control | ** | ** | ** | ** | 0.85 |
| Drought | ** | 0.81 | ||||
| SFW | Control | ** | ** | ** | ** | 0.91 |
| Drought | ** | 0.80 | ||||
| RDM | Control | ** | ** | NS | ** | 0.54 |
| Drought | ** | 0.41 | ||||
| SDM | Control | ** | ** | ** | ** | 0.75 |
| Drought | ** | 0.80 | ||||
| RL/SL | Control | ** | NS | ** | ** | 0.77 |
| Drought | ** | 0.81 | ||||
| RFW/SFW | Control | ** | ** | ** | ** | 0.59 |
| Drought | ** | 0.83 | ||||
*, ** F test of corresponding mean squares significant at the 0.05, and 0.01 probability levels, respectively. NS F test of corresponding mean squares not significant. Traits: root length (RL), shoot length (SL), root fresh weight (RFW), shoot fresh weight (SFW), root dry matter (RDM), shoot dry matter (SDM), root-to-shoot length ratio (RL/SL), root-to-shoot fresh weight ratio (RFW/SFW). h2 broad-sense heritability.
The G × T interaction was significant for all traits except root dry matter content (RDM). ANOVA by treatment showed a significant effect of genotype for all traits under both control and drought conditions. Broad sense heritability estimates were similar under both treatments, except for RL and the RFW/SFW ratio, for which heritability was much higher under drought than under control conditions (0.83 vs. 0.61 and 0.83 vs. 0.59, respectively). Under the control treatment, heritability ranged from 0.54 (RDM) to 0.91 (SFW). Under drought conditions, heritability ranged from 0.41 (RDM) to 0.83 (RL and RFW/SFW ratio). Higher heritability estimates were generally observed for shoot traits than for root traits.
3.2. Effect of Drought on Trait Means
Means of eight seedling traits varied widely among the 32 maize genotypes under both control and drought conditions (Table 2, Table S2).
Table 2.
Means and descriptive statistics for eight seedling traits in 32 maize genotypes under control and drought conditions.
Table 2.
Means and descriptive statistics for eight seedling traits in 32 maize genotypes under control and drought conditions.
| Trait | Treatment | Absolute units | Change (% of Control) | |||||
| Mean | Min | Max | CV (%) | Mean | Min | Max | ||
| RL | Control | 443.81 | 346.40 | 517.00 | 6.9 | −33 | −7 | −61 |
| Drought | 296.12 | 112.40 | 448.00 | 27.6 | ||||
| SL | Control | 445.17 | 307.60 | 571.00 | 15.4 | −34 | −21 | −46 |
| Drought | 292.50 | 155.00 | 421.40 | 15.4 | ||||
| RFW | Control | 16.01 | 7.83 | 33.48 | 26.5 | −53 | −37 | −68 |
| Drought | 7.61 | 4.66 | 13.60 | 25.6 | ||||
| SFW | Control | 15.98 | 7.24 | 29.24 | 28.8 | −66 | −47 | −76 |
| Drought | 5.37 | 2.33 | 10.88 | 30.7 | ||||
| RDM | Control | 8.91 | 5.12 | 13.79 | 16.3 | 120 | 70 | 165 |
| Drought | 19.47 | 12.97 | 27.99 | 16.3 | ||||
| SDM | Control | 8.83 | 7.09 | 10.39 | 7.5 | 63 | 40 | 97 |
| Drought | 14.34 | 10.34 | 19.37 | 14.5 | ||||
| RL/SL | Control | 1.02 | 0.76 | 1.50 | 15.3 | 0 | −37 | 42 |
| Drought | 1.01 | 0.42 | 1.54 | 23.7 | ||||
| RFW/SFW | Control | 1.02 | 0.73 | 1.42 | 16.4 | 42 | 10 | 93 |
| Drought | 1.45 | 0.96 | 2.03 | 15.3 | ||||
Traits: root length (RL), shoot length (SL), root fresh weight (RFW), shoot fresh weight (SFW), root dry matter (RDM), shoot dry matter (SDM), root-to-shoot length ratio (RL/SL), root-to-shoot fresh weight ratio (RFW/SFW).
Under drought conditions RL and SL were reduced by 33% and 34%, respectively, compared to the control, with a wide range of reduction among the genotypes (from 7% to 61% for RL and from 21% to 46% for SL). The mean reductions in plant fresh weight were even higher, reaching 52% for RFW and 66% for SFW. Variation in reduction among the genotypes for these two traits was also wide, ranging from 37% to 68% for RFW and from 47% to 76% for SFW. A contrasting genotypic response to drought was observed for the RL/SL ratio, with the relative change in individual genotypes ranging from −37% to +42%, resulting in no difference in means between the control and drought conditions for this trait. Mean RDM content increased under drought conditions by 120%, and all genotypes showed a consistent and substantial increase in RDM content under drought conditions, ranging from 70% to 165%. For SDM, genotypes also showed a consistent and strong increase in values under drought conditions resulting in large differences in trait means between the control and drought conditions (Table 2). The lowest coefficient of variation (CV) under control conditions was found for RL (5.2%), and under drought conditions for SDM (12.1%), while the highest CV under both conditions was observed for SFW (27.4% under control and 25.7% under drought). For RL, SDM, and RL/SL higher CVs were observed under drought conditions, while for the remaining five traits, CVs under control and drought conditions were of similar magnitude.
Means of eight traits in 32 maize genotypes under control and drought conditions are shown in Figure 2 and Figure 3 and in Table S2.
The three hybrids with the longest root under control conditions were 23, 11 and 14, while under drought conditions the hybrids with the longest root were 30, 3 and 23 (Figure 2). All hybrids showed a reduction in root length under drought conditions with the smallest reductions observed in hybrids 3, 30 and 6 (6.8%, 10.8% and 16.3%, respectively). The correlation between the two treatments for root length (RL) was weak to moderate and positive (0.38*). A strong and positive correlation between the two treatments was observed for shoot length (SL), 0.71**. All hybrids exhibited a reduction in SL under drought conditions, ranging from 20.8% to 46.3%. Three hybrids, 8, 1 and 7, had the longest shoots under both control and drought conditions, while the hybrids with the smallest reduction in SL between the two treatments were 30, 13 and 17. Hybrids 1, 8 and 31 had the highest root fresh weight (RFW) under control conditions, and hybrids 8, 3, 30 and 23 had the highest RFW under drought conditions. Hybrid 8, which had high RFW values under both control and drought conditions, was also among the hybrids with the smallest reduction in RFW between the two conditions (41.7%). A decrease in shoot fresh weight (SFW) under drought conditions, ranging from 46.9% to 76.3%, was observed for all studied hybrids. Hybrids 1 and 8 had the highest SFW under both control and drought conditions. A strong and positive correlation was found between the two treatments for SFW (0.76**).
A substantial increase in RDM and SDM contents was observed for all hybrids under drought conditions, with a greater increase in DM content in roots (ranging from 59.8% to 165.3%) than in shoots (ranging from 40.5% to 96.8%) (Figure 3). Hybrids 28 and 22 were among those with the highest RDM (23.1% and 22.8%) and SDM (17.6% and 17.21%) contents under drought conditions, as well as among those with the greatest increase in RDM and SDM contents between the control and drought conditions. The correlation between the two treatments was moderate and positive (0.47**) for RDM and strong and positive (0.67**) for SDM. The correlation between the two treatments for the RL to SL ratio was weak and positive (0.35*) and for this trait, hybrids showed contrasting responses, some hybrids reduced, while others increased the value of this ratio under drought conditions. Hybrids showing the highest increase in RL to SL ratio under drought conditions were 10, 6, 23, 3 and 31 (22.2%, 29.2%, 31.2%, 36.5% and 41.8%, respectively). A moderate and positive correlation between the two treatments was observed for the RFW to SFW ratio (0.46**). An increase in this ratio under drought conditions, ranging from 10.4% to 93.4%, was observed for all studied hybrids. The two hybrids with the highest relative increase of this ratio under drought conditions were 3 (93.4%) and 23 (81.4%).
3.3. Correlation Between Traits
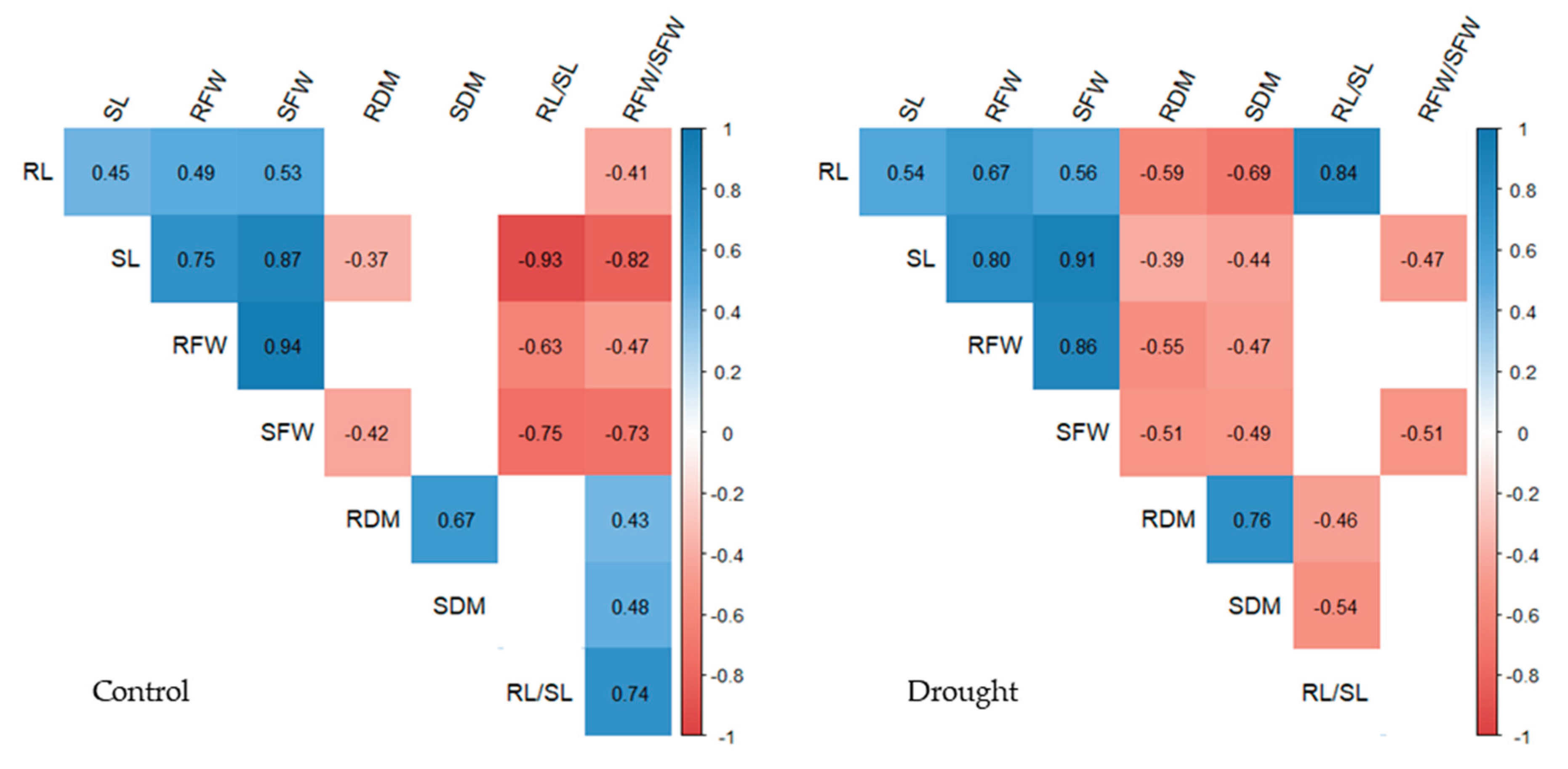
The correlations between the pairs of traits under control and drought conditions are presented in Figure 4. The direction of correlation coefficients between the pairs of traits detected simultaneously under control and drought conditions remained the same. The positive correlation coefficients were of similar magnitude under both conditions, but the two negative correlation coefficients, between SL and RFW/SFW, and SFW and RFW/SFW, were of higher magnitude under control conditions compared to drought conditions (−0.82 vs. −0.47 and −0.73 vs. −0.51, respectively).
While a significant negative correlation between SDM and most other traits (except RDM, which was positive under both treatments) was detected only under drought conditions, the correlation between RFW/SFW and the remaining traits was significant predominantly under control conditions. The correlation between RL/SL and other traits showed inconsistent behavior, those significant under control conditions were not detected under drought conditions, and vice versa.
4. Discussion
In the present study, the effect of PEG-induced drought on seedling traits was examined in 32 commercial maize hybrids. The effect of genotype (G) was significant for all traits, while the effect of water treatment (T) was significant for all traits except root-to-shoot length ratio (RL/SL). The G × T interaction was significant for all traits except RDM. A significant effect of genotype as well as water treatment has previously been reported for maize seedling traits in various germplasm, including inbreds [14,20,21], hybrids [8,22,23,24], and local varieties/landraces [16,17,18,25,26]. The significant G × T interaction for all traits except RDM indicates that hybrids showed distinct responses when exposed to drought stress and may reflect the existing genetic variability for drought tolerance in the studied germplasm. The response of the studied hybrids to simulated drought stress was typical, resulting in decreased values of the four directly measured traits: RL, SL, RFW and SFW. According to Álvarez-Iglesias et al. [23], mild and moderate stress conditions typically reduce shoot growth more than root growth in maize seedlings. Maintenance of main root growth is an adaptive mechanism for optimizing soil water uptake [36], enabling roots to penetrate deeper into the soil and increasing the likelihood of finding water sources. In the set of maize hybrids studied here, root and shoot length were affected to a similar extent, with hybrid no. 30 standing out for maintaining both root and shoot growth under drought conditions. As mentioned earlier, a typical response under drought conditions is an increase of root-to-shoot length ratio [16], which, according to Queiroz et al. [22], indicates that shoot development was indeed more affected by drought than root development. The hybrids which in the present study expressed an increase of RL/SL (hybrids no. 31, 3, 23) under drought conditions were, together with hybrid no. 30, among those with the longest RL and the smallest relative RL reduction under drought. The similar magnitude of RL and SL reduction in this study might be explained by the relatively mild drought stress applied, which may also account for the non-significant difference observed between the two conditions for the root-to-shoot length ratio. Nevertheless, a milder effect of drought was observed for RFW than for SFW (-53% vs. -66% decrease under drought stress, respectively), resulting in an increased RFW/SFW ratio. According to Álvarez-Iglesias et al. [23], under drought conditions, initial growth of both main and secondary roots is necessary, as they increase the surface area for water uptake and could ensure subsequent water supply to the main root before water deficit becomes severe. In this regard, hybrid 30 ranked second in terms of the smallest relative reduction of RFW and SFW under drought conditions and it was the only hybrid to maintain consistently good relative performance under induced drought stress.
Under drought stress, all hybrids showed a strong increase in RDM and SDM content, with average increases of 120% and 63%, respectively. The very high average increase in RDM under drought conditions, which ranged from 70% to 165%, implies a similar reaction of the studied hybrids to drought stress for this trait and may explain the non-significant G × T interaction observed for this trait. The increase in DM in roots and shoots may indicate that another potentially valuable stress tolerance mechanism has been activated in the studied germplasm: the adjustment of osmotic potential achieved by active accumulation of solutes in cells [4,37]. This accumulation enables the retention of water during episodes of low external water potential, limiting turgor loss and damage from cell shrinkage. Under more prolonged or severe moisture deficit, these solutes are also implicated in the stabilization of various macromolecular structures [4].
The calculated trait heritability values under control and drought conditions were of similar magnitude and generally high. In Badr et al. [19], the broad-sense trait heritability coefficients calculated under control and 10% PEG drought stress treatment, which is comparable to the PEG treatment applied in the present study, were also of similar magnitude. High broad-sense heritability coefficients for seedling traits such as germination percentage, root and shoot length, and fresh and dry weight were also found in the studies of Mustamu et al. [18], who investigated PEG-induced drought stress at the early seedling stage of local Indonesian maize varieties, as well as in the study of Khan et al. [38], who investigated drought tolerance of 40 Pakistani maize inbreds at seedling and maturity stages. The results from these studies, as well as from the present study, indicate that selection of maize-tolerant genotypes may be based on the high h2 seedling traits such as shoot length and shoot and root fresh and dry weight.
In the present study, strong positive correlations between control and drought conditions were found for SL, RFW, and SFW (0.71**, 0.68** and 0.76**, respectively). These traits also exhibited high heritabilities under both conditions. Between the same three directly measured traits, including RL, and between RDM and SDM, positive and moderate to strong correlations were observed under both control and drought conditions (Figure 4). Adhikari et al. [39] reported similar correlations between RL and RFW and SFW under both control and drought conditions, while Badr et al. [19] found correlations between the same traits under control and 10% PEG treatment to be generally weaker, but slightly stronger under drought stress conditions. Conversely, Khan et al. [38] observed a higher number of strong and significant correlations among RL, SL, RFW, and SFW under control than under drought conditions. In the present study, correlations between directly measured and derived traits, as well as among derived traits, showed contrasting patterns and were specific to the designated water stress treatment. Therefore, using the four easily and directly measured traits: RL, SL, RFW and SFW for assessing drought tolerance at the maize seedling stage appears appropriate.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Code, FAO group and breeders of 32 maize single-cross hybrids used as experimental material; Table S2: Hybrid mean root length (RL), shoot length (SL), root fresh weight (RFW), shoot fresh weight (SFW), root dry matter (RDM), shoot dry matter (SDM), root-to-shoot length ratio (RL/SL), and root fresh weight to shoot fresh weight ratio (RFW/SFW) values under control and drought conditions.
Author Contributions
Conceptualization, I.P. S.K.; methodology, S.K. and H.Š.; software, M.B. and H.Š.; validation, I.P. and H.Š.; formal analysis, S.K., M.B. and H.Š.; investigation, S.K. and A.L.; resources, I.P.; data curation, S.K. and H.Š.; writing—original draft preparation, M.B and H.Š.; writing—review and editing, I.P., S.K. and A.L.; visualization, S.K., M.B and H.Š.; supervision, I.P. and H.Š.; project administration, I.P.; funding acquisition, I.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Environmental Protection and Energy Efficiency Fund with the support of the Croatian Science Foundation of the Republic of Croatia (project AGRODROUGHT-ADAPT, PKP-2016-06-8290) and by the project PK.1.1.10.0008 (Research and Development of Plant Genetic Resources for Sustainable Agriculture), Centre of Excellence for Biodiversity and Molecular Plant Breeding (CoE CroP-Bio-Div), Zagreb, Croatia.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Deribe, H. Review on Effects of Drought Stress on Maize Growth, Yield and Its Management Strategies, Communications in Soil Science and Plant Analysis 2025, 56:1, 123–143. [CrossRef]
- Yin, X.; Olesen, J. E.; Wang, M.; Kersebaum, K.-C.; H. Chen, H.; Baby, S.; Öztürk, I.; Chen. F. Adapting maize production to drought in the northeast farming region of China. The European Journal of Agronomy 2016, 77, 47–58. [CrossRef]
- FAOSTAT. Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database. Available online: https://www.fao.org/faostat/en/#data (accessed on 15th of September 2025).
- Maazou, A.-R.S.; Tu, J.L.; Qiu, J.; Liu, Z.Z. Breeding for Drought Tolerance in Maize (Zea mays L.). American Journal of Plant Sciences 2016, 7, 1858–1870. [Google Scholar] [CrossRef]
- Rasheed, A.; Jie, H.; Ali, B.; He, P.; Zhao, L.; Ma, Y.; Xing, H.; Qari, S.H.; Hassan, M.U.; Hamid, M.R.; et al. Breeding Drought-Tolerant Maize (Zea mays) Using Molecular Breeding Tools: Recent Advancements and Future Prospective. Agronomy 2023, 13, 1459. [Google Scholar] [CrossRef]
- Critchley, W.; Siegert, K. Water Harvesting: A Manual for the Design and Construction of Water Harvesting Schemes for Plant Production. FAO, Rim, Italy, 1991.
- Sah, R.P.; Chakraborty, M. : Prasad, K.; Pandit, M.; Tudu, V.K.; Chakravarty, M.K.; Narayan, S.C.; Rana, M.; Moharana, D. Impact of water deficit stress in maize: Phenology and yield components. Sci. Rep. 2022, 10, 2944. [Google Scholar] [CrossRef]
- Akinwale, R.O.; Awosanmi, F.E.; Ogunniyi, O.; Fadoju, A.O. Determinants of drought tolerance at seedling stage in early and extra-early maize hybrids. Maydica 2017, 62, M4. [Google Scholar]
- Bruce, W.B.; Edmeades, G.O.; Barker, T.C. Molecular and physiological approaches to maize improvement for drought resistance. J. Exp. Bot. 2002, 53, 13–25. [Google Scholar] [PubMed]
- Denmead, O. T.; Shaw, R. H. The effects of soil moisture stress at different stages of growth on the development and yield of corn. Agron. J. 1960, 52, 272–274. [Google Scholar] [CrossRef]
- Edmeades, G.O.; Lafitte, H.R.; Bolanos, J.; Chapman, S.C.; Bänziger, M.; Deutsch, J.A. Developing maize that tolerates drought or low nitrogen conditions. In: Stress tolerance breeding: maize that resists insects, drought, low nitrogen, and acid soils. Edmeades, G.O., Deutsch, J.A., Eds.; 1994, pp. 21–84.
- Duvick, D.N. What is yield? In: Developing drought and low-N tolerant maize. Edmeades, G.O., Bänziger, M., Mickelson, H.R., Peña-Valdivia, C.B., Eds., CIMMYT: El Batan, Mexico, 1997, pp. 332–335.
- Duvick, D. N.; Cassman, K. G. Post-green revolution trends in yield potential of temperate maize in the North-Central United States, Crop Sci, 1999, 39, 1622–1630.
- Adewale, S.A.; Akinwale, R.O.; Fakorede, M.A.B.; Badu-Apraku, B. Genetic analysis of drought-adaptive traits at seedling stage in early-maturing maize inbred lines and field performance under stress conditions. Euphytica. [CrossRef]
- Meeks, M.; Murray, S.C.; Hague, S.; Hays, D. Measuring Maize Seedling Drought Response in Search of Tolerant Germplasm. Agronomy 2013, 3, 135–147. [Google Scholar] [CrossRef]
- Djemel, A.; Álvarez-Iglesias, L.; Pedrol, N.; López-Malvar, A.; Ordás, A.; Revila, P. Identification of drought tolerant populations at multi-stage growth phases in temperate maize germplasm. Euphytica 2018, 214. [Google Scholar] [CrossRef]
- Mustamu, N.E.; Tampubolon, K.; Alridiwirsah, A.; Basyuni, M. Preliminary Identification of Local Maize Under Drought Stress By PEG-6000. BIO Web Conf. 2023, 69, 01018. [Google Scholar] [CrossRef]
- Mustamu, N.E.; Tampubolon, K.; Alridiwirsah, A.; Basyuni, M. , AL-Taey D. K.A., Janabi, H.J.K., Mehdizadeh, M. Drought stress induced by polyethylene glycol (PEG) in local maize at the early seedling stage, Heliyon 2023, 9, e20209. [Google Scholar] [CrossRef]
- Badr, A.; El-Shazly, H.H.; Tarawneh, R.A.; Börner, A. Screening for Drought Tolerance in Maize (Zea mays L.) Germplasm Using Germination and Seedling Traits under Simulated Drought Conditions. Plants 2020, 9, 565. [Google Scholar] [CrossRef]
- Magar, M.M.; Atit Parajuli, A.; Sah, B.P.; Shrestha, J.; Sakha, B.M.; Koirala, K.B.; Dhital, S.P. (2019). Effect of PEG Induced Drought Stress on Germination and Seedling Traits of Maize (Zea mays L.) Lines. Turkish Journal of Agricultural and Natural Sciences 2019, 6, 196–205. [Google Scholar]
- Ul Islam, N.; Ali, G.; Dar, Z.A.; Maqbool, S.; Khulbe, R.K.; Bhat, A. (2019). Effect of PEG Induced Drought Stress on Maize (Zea mays L.) Inbreds. Plant Archives 2019, 19, 1677–1681. [Google Scholar]
- Queiroz, M.; da Silva Oliveira, C.E.; Steiner, F.; Zuffo, A.M.; Zoz, T.; Vendruscolo, E.P.; Silva Mennes, V.; Mello, B.F.F.R.; Cabral, R.C.; Menis, F.T. Drought Stresses on Seed Germination and Early Growth of Maize and Sorghum. Journal of Agricultural Science 2019, 11, 310–310. [Google Scholar] [CrossRef]
- Álvarez-Iglesias, L.; de la Roza-Delgado, B.; Reigosa, M.J.; Revilla, P.; Pedrol, N. A simple, fast and accurate screening method to estimate maize (Zea mays L.) tolerance to drought at early stages. Maydica 2017, 62, M34. [Google Scholar]
- Partheeban, C.; Chandrasekhar, C.N.; Jeyakumar, P. : Ravikesavan, R. ; Gnanam, R. Effect of PEG Induced Drought Stress on Seed Germination and Seedling Characters of Maize (Zea mays L.) Genotypes, Int. J. Curr. Microbiol. App. Sci. 2017, 6, 1095–1104. [Google Scholar] [CrossRef]
- Khodarahmpour, Z. Effect of drought stress induced by polyethylene glycol (PEG) on germination indices in corn (Zea mays L.) hybrids. African Journal of Biotechnology 2011, 10, 18222–18227. [Google Scholar] [CrossRef]
- Khayatnezhad, M.; Gholamin, R.; Jamaati-e-Somarin, S.; Zabihi-e-Mahmoodabad, R. Effects of PEG Stress on Corn Cultivars (Zea mays L.) at Germination Stage. World Applied Sciences Journal 2010, 11, 504–506. [Google Scholar]
- Zhang, L.J.; Fan, J.J.; Ruan, Y.Y. Application of polyethylene glycol in the study of plant osmotic stress physiology. Plant Physiol. Commun. 2004, 40, 361–368. [Google Scholar]
- Guan, Z.; Wang, L.; Duan, L.; Zhou, Z.; Zhang, F.; Wang, Y. Effects of PEG simulated drought stress on seed germination of Abutilon theophrasti medicus. Seed 2022, 41, 66–70. [Google Scholar] [CrossRef]
- Wang, X.; Li, X.; Zhao, W.; Hou, X.; Dong, S. Current views of drought research: experimental methods, adaptation mechanisms and regulatory strategies. Front. Plant Sci. 2024, 15, 1371895. [Google Scholar] [CrossRef]
- Avramova, V.; Nagel, K.A.; AbdElgawad, H.; Bustos, D.; DuPlessis, M.; Fiorani, F.; Beemster, G.T.S. Screening for drought tolerance of maize hybrids by multi-scale analysis of root and shoot traits at the seedling stage. J. Exp. Bot. 2016, 67, 2453–2466. [Google Scholar] [CrossRef] [PubMed]
- Michel, B.E. Evaluation of the water potentials of solutions of polyethylene glycol 8000. Plant Physiology 1983, 72, 66–70. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute, Statistical Analysis Software (SAS) User’s Guide Version 9.4. SAS Institute Inc., Cary, NC, USA, 2016.
- Hallauer, A.R.; Carena, M.J.; Filho, J.B.M. Quantitative Genetics in Maize Breeding; Springer, New York, USA, 2010.
- R Core Team _R: A Language and Environment for Statistical Computing_, version 4.4.2., R Foundation for Statistical Computing, Vienna, Austria, 2024 https://www.R-project.
- Soetewey, A. Correlation coefficient and correlation test in R, Stats and R. Available online: https://statsandr.com/blog/correlation-coefficient-and-correlation-test-in-r/ (accessed on 16th of September 2025).
- Sharp, R.E.; Davies, W.J. Regulation of growth and development of plants growing with a restricted supply of water. In Plants under Stress. Biochemistry, Physiology and Ecology and Their Application to Plant Improvement, Jones, H.G., Flowers, T.J., Jones, M.B., Eds.; Cambridge University Press, Cambridge, UK, 1989.
- Ribaut, J.M.; Betran, J.; Monneveux, P.; Setter, T. Drought Tolerance in Maize. In Handbook of Maize: Its Biology, Bennetzen, J., Hake, S., Eds., Springer, New York, USA, 2009; pp. 311–344. [CrossRef]
- Khan, N. H.; Ahsan, M.; Naveed, M.; Sadaqat, H. A.; Javed, I. Genetics of drought tolerance at seedling and maturity stages in Zea mays L. Spanish Journal of Agricultural Research 2016, 14, e0705. Spanish Journal of Agricultural Research 2016, 14, e0705. [Google Scholar] [CrossRef]
- Adhikari, B.; K. J., Sa.; Lee, J.K. Drought tolerance screening of maize inbred lines at an early growth stage. Plant Breeding and Biotechnology 2019, 7, 326–339. [Google Scholar] [CrossRef]
Figure 1.
Plant growth under treatments in the growth chamber–second day after transplanting.

Figure 2.
Means for root length (RL), shoot length (SL), root fresh weight (RFW), and shoot fresh weight (SFW) in 32 maize hybrids in control and drought conditions and the Pearson correlation coefficients (r) between the two treatments. *, ** Correlation coefficient significant at the 0.05 and 0.01 probability levels, respectively.
Figure 2.
Means for root length (RL), shoot length (SL), root fresh weight (RFW), and shoot fresh weight (SFW) in 32 maize hybrids in control and drought conditions and the Pearson correlation coefficients (r) between the two treatments. *, ** Correlation coefficient significant at the 0.05 and 0.01 probability levels, respectively.
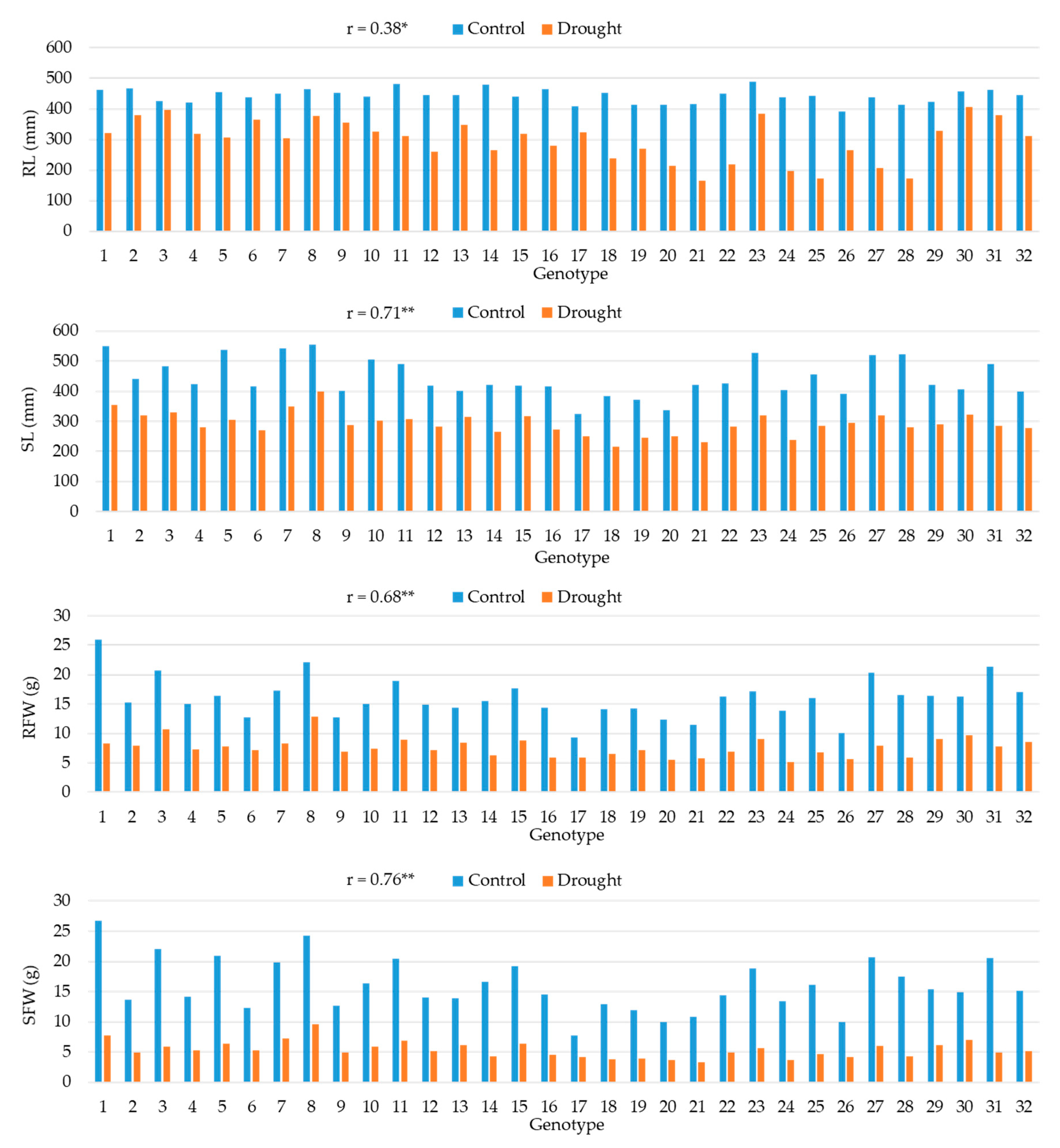
Figure 3.
Means for root dry matter (RDM), shoot dry matter (SDM), root-to-shoot length ratio (RL/SL), and root-to-shoot fresh weight ratio (RFW/SFW) in 32 maize hybrids in control and drought conditions and the Pearson correlation coefficients (r) between the two treatments. *, ** Correlation coefficient significant at the 0.05 and 0.01 probability levels, respectively.
Figure 3.
Means for root dry matter (RDM), shoot dry matter (SDM), root-to-shoot length ratio (RL/SL), and root-to-shoot fresh weight ratio (RFW/SFW) in 32 maize hybrids in control and drought conditions and the Pearson correlation coefficients (r) between the two treatments. *, ** Correlation coefficient significant at the 0.05 and 0.01 probability levels, respectively.
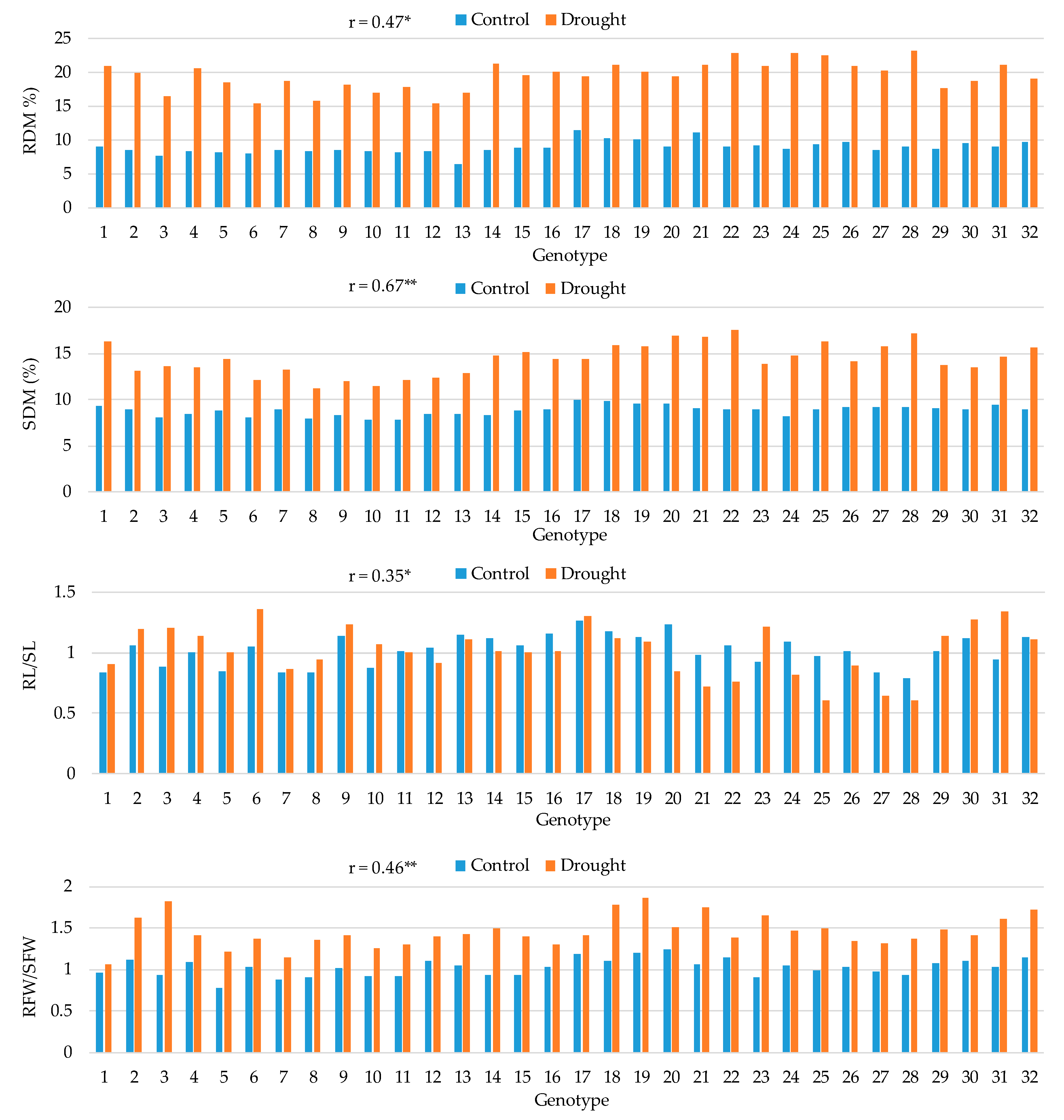
Figure 4.
Correlation coefficients between maize seedling traits in 32 maize genotypes under control and drought conditions. Traits: root length (RL), shoot length (SL), root fresh weight (RFW), shoot fresh weight (SFW), root dry matter (RDM), shoot dry matter (SDM), root-to-shoot length ratio (RL/SL), root-to-shoot fresh weight ratio (RFW/SFW).
Figure 4.
Correlation coefficients between maize seedling traits in 32 maize genotypes under control and drought conditions. Traits: root length (RL), shoot length (SL), root fresh weight (RFW), shoot fresh weight (SFW), root dry matter (RDM), shoot dry matter (SDM), root-to-shoot length ratio (RL/SL), root-to-shoot fresh weight ratio (RFW/SFW).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



