
Submitted:
04 October 2025
Posted:
08 October 2025
You are already at the latest version
Abstract
The review discussed the fundamental characteristics of grape phenolic compounds, focusing on their chemical properties and significance in winemaking. Given the high-er phenolic content in red wines, particular attention is devoted to attributes such as pigmentation, co-pigmentation, bitterness, and interactions with proteins—key con-tributors to the perception of astringency. Phenolic compounds undergo substantial qualitative and quantitative transformations during grape ripening, which affect both their inherent properties and their extractability from solid components such as grape skins and seeds. The review also examined extractability dynamics and provides a concise overview of the analytical methods used to assess phenolic compounds in grapes and wine.
Keywords:
grape phenolic compounds
; grape maturity
; grape seeds and skins
; phenolic extractability
1. Introduction
Although phenolic compounds constitute a relatively minor fraction of the grape and, consequently, of wine composition—with concentrations rarely exceeding 0.3–0.4% in red wines and 0.03–0.04% in white wines—they play a vital role in the winemaking process. Phenolic compounds influence wine color, organoleptic characteristics such as bitterness and astringency, and overall structural complexity. Their antioxidant properties are well-documented and are the subject of extensive research. As a result, phenolic compounds are considered a critical component of red wine quality and also significantly contribute to the sensory attributes of white wines. The aim of this review was to summarize the current knowledge about phenolic compounds in grapes, their influence on wine quality and methods for phenolic compounds determination.
2. Phenolic Compounds in Grapevines: Structure, Reactivity, and Role in Plant Development
Vine polyphenols are classified as secondary plant metabolites, primarily functioning as intermediates or end products of the shikimate pathway [1]. Although simple phenolic structures, such as those illustrated in Figure 1, are not found directly in grapes, their molecular configuration helps to explain many phenolic properties. The charge delocalization of aromatic rings, coupled with the inductive effect of sp2-hybridized carbon atoms at position 1 (Figure 1a), weakens the O–H bond, conferring phenolic compounds with more acidic character than typical alcohols. Additionally, the two electron pairs on the oxygen atom of the hydroxyl group render phenolic compounds more reactive than benzene and hydroxyls.
Orbital overlap between high electron-density oxygen atoms and the relatively electron-deficient aromatic rings enables partial electron transfer and increases reactivity at three specific positions on the aromatic ring. The presence of an additional hydroxyl group in the meta-position (Figure 1a) further enhances this reactivity, while another in the ortho-position (Figure 1b) activates both this site and the other oxygen atoms on the ring.
The high chemical reactivity of phenolic bonds may explain their biological roles in plant development, including grape vines. These compounds are primarily concentrated in the outer layers of the plant or fruit, but significant quantities are also present in the roots. This distribution, along with their ability to bind with other molecules, suggests that phenolic compounds may function as transport agents for monosaccharides and other compounds, or as regulators of osmotic pressure [2].
Phenolic compounds are also recognized for their potent antioxidant activity, which likely contributes to plant defense mechanisms, either directly or indirectly in response to biotic or abiotic stressors [3,4,5,6]. Grapes exposed to sunlight or subjected to fungicidal treatment often show elevated phenolic content [7]. In addition, due to their pigmentation properties, phenolic compounds may influence ecological interactions—either attracting animals for pollination and seed dispersal or, conversely, deterring them [8,9].
All phenolic compounds exhibit peaks of absorption in the ultraviolet (UV) spectrum, predominantly in the UV-B region (280–320 nm) and, to a lesser extent, in the UV-A range (320–400 nm). This characteristic, along with their localization in epidermal tissue, supports their proposed function as natural UV filters for plants. The practice of exposing grape bunches to sunlight is well-established in some viticulture regions to enhance grape color. Such exposure increases total phenolic content and anthocyanin concentration and also promotes the accumulation of terpenes and norisoprenoids [10], compounds that affect wine color, taste, and aroma. Balanced vine vigor, combined with partial leaf removal and controlled sunlight exposure after veraison, significantly enhances phenolic accumulation. However, in hotter wine-growing regions, this practice must be carefully timed, as early exposure may lead to adverse effects.
3. Evolution of Phenolic Compounds During Grape Ripening
3.1. Non-Flavonoid Phenolics
Of the two primary groups of non-flavonoid phenolic compounds—hydroxycinnamic acids and hydroxybenzoic acids—the former is more prevalent in grapes and wines. The main hydroxycinnamic acids present in grapes include p-coumaric, caffeic, and ferulic acid [11,12]. These acids exist as both trans and cis isomers, with the trans form being more abundant. The amount of cis isomers is more influenced by cultivation conditions. Caffeic acid is the predominant compound of hydroxycinnamic acids, followed by coumaric acid. They typically occur as esters with tartaric acids (Figure 2). In Vitis vinifera cultivars, coumaric esters are between 3% and 33% of the caffeic ester content, a range thought to reflect cultivar-specific characteristics [12]. Ferulic and p-coumaric acids bind to cell wall polysaccharides, and these bonds—along with the formation of diferulic acid—function as cross-links between pentose chains [13,14,15]. Although hydroxycinnamic acids and their esters occur at higher concentrations in grape skins, they are also present in the pulp. During grape ripening, hydroxycinnamates are thought to serve as precursors for other phenolic compounds. Therefore, their concentration remains relatively stable throughout the maturation phase.
Quantitative analysis has identified coutaric, caftaric, and fertaric acids in red wines in the following concentration ranges (mg/L): 2.3-27.1, 4.0-77.4, and 0.7-6.5, respectively [16]. For the same samples, free coumaric, caffeic, and ferulic acids were found in ranges of 0.6–19.3, 3.1–70.7, and 0.0–2.9 mg/L. The ratios of tartaric esters to free acids ranged from 1.86 for caffeic acid to 2.36 for coumaric acid and 2.85 for ferulic acid.
In white wines, concentrations of coutaric and caftaric acids and their respective non-esterified forms ranged from 2.66–5.74, 14.71–43.53, 0.62–0.71, and 1.57–3.39 mg/L [17]. In these wines, the ester-to-non-ester ratios for coumaric and caffeic acids ranged from 5.22 to 9.26 and from 9.36 to 13.43, respectively.
Although hydroxycinnamic acids concentration in white wines is typically lower, they can represent up to 15% of total phenolics, compared with less than 5% in red wines. Consequently, they may have a more pronounced influence on the taste profile of white wines. These compounds contribute to bitterness and mild acidity, exert limited effects on astringency, and play important roles in color stabilization and oxidation reactions [18]
In addition to hydroxycinnamates, small amounts of gallic and ellagic acids—both belonging to the hydroxybenzoic acid group—are present in grapes, with higher concentrations typically found in oak-aged wines.
Another important non-flavonoid phenolic is resveratrol, a member of the stilbene group. Although present in grapes and wine at relatively low concentrations [19,20], resveratrol has attracted significant research interest because of its potential cardioprotective and anticarcinogenic properties. Notably, its levels differ between organic and conventionally produced wines [21]
3.2. Flavonoid Phenolics
3.2.1. Flavan-3-ols
The structure of flavan-3-ols (also known as flavanols or catechins) allows for the existence of geometric isomers and enantiomers, which contribute to the structural diversity of this group. The major flavan-3-ol compounds found in grapes are shown in Figure 3. Flavan-3-ols are significantly different from other flavonoids in that they do not occur in glycoside form. These compounds are found in the solid parts of the grape, with more than 50% of total catechins in the seeds, followed by the stalks and, to a lesser extent, the skins [22,23,24,25].
The total flavan-3-ol content is largely cultivar-specific. In some grape varieties, concentrations of approximately 230 mg/kg have been reported [25], whereas in more extractive cultivars, the total content may be several times higher. A variation in the distribution of individual flavan-3-ols is also observed: (−)-epigallocatechin is typically absent from grape seeds, (−)-epicatechin predominates in seeds and stalks, while (+)-catechin is more abundant in grape skins.
Although flavan-3-ol accumulation is, in the most cases, genetically determined, it is also influenced by climatic conditions and viticultural practices [23,26,27]. For example, leaf removal reduces monomeric and dimeric phenolics in seeds (by ~3% and 21%, respectively, across two vintages), while simultaneously increasing their concentrations in skins (by ~6% and 14%) [28].
Significant changes in flavan-3-ol content are observed during grape ripening. In various cultivars, catechin levels at veraison are 3–7 times higher than in fully ripe grapes [29]. In Cabernet Sauvignon, a reduction of about 90% in seed flavan-3-ol content has been reported. This decline in flavan-3-ol content across seeds, skins, and stalks has been confirmed in multiple studies [22,23,30,31,32]. In most grape varieties, the most pronounced decline in catechin content occurs during the first weeks after veraison. Along with the overall decline, the relative proportions of individual flavan-3-ols also shift. In Cabernet Sauvignon seeds, the ratios of (+)-catechin (C), (−)-epicatechin (eC), and (−)-epicatechin gallate (eCG)—expressed as percentages of total seed flavan-3-ols—shift from 65:27:8 at veraison to 48:51:1 at full ripening [30]. Similar trends are observed in other Vitis viniffera varieties like Touriga Nacional, where the ratio changes from 32:48:20 to 31:54:15 and for Touriga Francesa, where the ratio shifts from 44:40:16 to 42:53:5 between veraison and ripening [23]. Comparable changes are also seen in grape stems. When the combined content of C and eC is normalized to 100%, the C:eC ratio in Castelão Francês decreases from 83:17 at veraison to 65:35 after 65 days. In Touriga Francesa, the ratio changes from 90:10 to 80:20 over the same period [31].
3.2.2. Proanthocyanidins
Proanthocyanidins (also known as condensed tannins, or simply tannins) are oligomers and polymers of flavan-3-ols. The subunits are linked through C4–C8 (Figure 4B) or C4–C6 bonds. The nature of these linkages allows for the formation of stereoisomers [33,34].
Grape proanthocyanidins are classified into two groups: procyanidins and prodelphinidins. The first group, which includes catechin, epicatechin, and epicatechin gallate, occurs in grape skins, seeds, and stems. In contrast, prodelphinidins contain epigallocatechin and are present only in skins and stems [26,27,30,35,36,37].
Because of the diversity in bond types, stereochemistry, flavan-3-ol subunits, and degrees of polymerization, grape and wine procyanidins comprise a wide range of individual compounds. The main procyanidins identified in grapes—B1, B2, B3, and B4—are shown in Figure 4A. Other forms, such as galates and B5–B8-type dimers (featuring C4–C6 bonds), have also been identified. Up to 14 dimers, 11 trimers, and several tetramers have been reported in grape seeds [36].
Among these, procyanidin B1 predominates in the skins of several white and red grape varieties [25,31,38], whereas procyanidin B2 is the main dimer in seeds. Procyanidin B2 has also been identified as the primary procyanidin in both seeds and skins of Merlot and Cabernet Sauvignon cultivars [29]. During grape ripening, the concentration of dimeric procyanidins in seeds, skins, and stems decreases significantly. In some varieties, this reduction reaches 50–60% compared with concentrations at veraison, with the most pronounced decline occurring in the first weeks post-veraison. The decrease mainly affects procyanidins B1, B2, B3, B4, and B2-gallate [22,23,29,32].
Grape ripening is accompanied by a general decrease in both flavan-3-ols and procyanidins, as well as a shift in their relative proportions. For example, in Cabernet Sauvignon, the proportion of seed-derived catechins decreased from 77% to 41% of total phenolic compounds between veraison and ripening. Over the same period, the relative proportion of procyanidins increased from 23% to 59% [26,30]. Other studies have examined the distribution of phenolics by polymer length. In one of them, it was reported that at grape maturity, monomeric catechins, oligomeric procyanidins (dimers to pentamers), and polymeric procyanidins accounted for a ratio of 11:34:55 [35] whereas in another grape variety the same proportions were 50:44:6 [25].
Ripening also alters the proportion of extension and terminal units in seed procyanidins. For example, in Cabernet Sauvignon, the degree of galloylation (DG)—measured as the proportion of epicatechin gallate extension units—decreased from 38% to 18%, while the mean degree of polymerization (mDP) declined from 8.29 to 5.63 [26,30]. Other studies report smaller changes: in Carmenère, DG decreased from 29.7% to 27.5%, and in Cabernet Sauvignon, from 26.8% to 16.3%, while mDP declined from 3.8 to 3.2 and from 4.3 to 2.7, respectively [39]. In contrast, grape skin procyanidins show the opposite trend, with mDP increasing during ripening. For example, Kennedy et al. reported an increase in skin mDP from 15 to 30 [26,30]. However, in the study of Obreque-Slier et al. [39] two different trends were observed for mDP of Carmenere and Cabernet sauvignon. mDP values in Carmenere decreased from 4.9 to 3.8 while mDP in Cabernet Sauvignon increased from 4.1 to 7.1. In both cultivars, DG in skin procyanidins increased from 7.8 to 12.5 in Carmenère and from 7.5 to 19.0 in Cabernet Sauvignon, respectively [39]. Another study reported a notably lower DG of 3.95 in ripened Merlot grape skins [27]. At full ripeness, the distribution of procyanidins by polymer size in grape skins has been reported as 2% monomers, 8% oligomers (mDP = 2–5), and 90% polymers (mDP > 10) [25].
Given that seed-derived tannins are more astringent than those from skins, DG appears to influence perceived astringency more strongly than mDP. Changes in tannin composition during ripening, reflected by protein precipitation assays and shifts in the seed-to-skin tannin ratio, further confirm these dynamics [40]. For the Melnik 55 grape variety, the concentration of tannins extractable in a model wine solution (12% v/v ethanol) decreased nearly threefold by harvest, dropping to 35–50 mg/L compared to 150–160 mg/L detected four weeks prior to full ripeness [41].
3.2.3. Flavonols
Flavonols are flavonoid phenolic compounds that occur in grapes primarily in the form of glycosides. In contrast to flavan-3-ols and procyanidins, flavonols are predominantly localized in grape skins, with only negligible amounts detected in seeds [42]. They are also found in stems and leaves, where quercetin glycosides are the most common representatives. This group of monomeric phenolics is found at relatively low concentrations in wine, and scientific interest in them arises less from their technological role in winemaking than from their antioxidant capacity and free radical-scavenging activity in grapes and grape-derived products [43,44,45,46,47,48].
3.2.4. Anthocyanins
The term “anthocyanins” refers to the glycosylated forms of plant pigments (Figure 5), whereas the corresponding aglycones are known as “anthocyanidins”. Due to their chemical instability, anthocyanidins are rarely found in free form in plant tissues and fruits [49].
The specific structure of an anthocyanin—defined by the type of anthocyanidin, the sugar moiety, and any additional substituents attached to the sugar— accounts for the wide diversity the wide diversity of anthocyanins observed in nature [50]. The color of anthocyanins, attributed to their conjugated double bond systems, is primarily affected by pH, but is also influenced by substituents on the B-ring (R1 and R2 in Figure 5). Hydroxylation induces a bathochromic shift toward longer wavelengths, resulting in a bluish hue. Methoxylation produces a hyperchromic effect accompanied by a shift toward shorter wavelengths, thereby enhancing the red hue [49]. Other structural modifications, such as acylation or co-pigmentation (communalation), affect not only color expression but also color stability under varying environmental conditions [42,51,52,53,54].
In grape varieties of Vitis vinifera, five main anthocyanins are typically found: delphinidin, cyanidin, malvidin, peonidin, and petunidin. Among them, malvidin is the most abundant anthocyanin in grape skins. Depending on the cultivar, anthocyanins may also include acyl substituents such as acetic, p-coumaric, or caffeic acid, as well as other acid derivatives [55,56,57,58]. The total anthocyanin content in grapes can vary widely—from approximately 30 to 750 mg per 100 g of ripe berries—depending on the variety and viticultural conditions [56,58,59].
The accumulation of anthocyanins in grape skins is the first visible indicator of ripening. During this stage, anthocyanin levels increase until they plateau at full fruit maturity [60]. Some studies report compositional changes during this period: in Syrah, for example, a decrease in the less stable delphinidin has been observed, accompanied by an increase in the more stable malvidin [61]. Other studies note a steady rise in glycosylated anthocyanins, with a temporary decline in acylated forms before ripening, followed by a slight decrease in total anthocyanins just before maturity and a renewed increase thereafter [62]. Overall, anthocyanin concentration is influenced not only by genetic factors [63] but also by environmental variables such as sun exposure [64] and diurnal temperature variation [65]. Viticultural practices are also significant: irrigation often leads to reduced anthocyanin levels [66,67]. However, late irrigation—applied shortly before ripening—has been shown not to cause significant reductions, making it a suitable strategy for vineyards in hot climates [67]. Additional interventions such as leaf removal, shoot thinning, or their combination can likewise strongly influence anthocyanin content and the overall phenolic profile of grapes [68].
4. Phenolic Compounds and Wine Quality
4.1. Pigmentation and Co-Pigmentation
Although phenolic compounds rarely exceed 13–15% of the dry extract in red wines and around 10% in white wines, they exert a disproportionately large influence on wine quality owing to their reactivity and chemical properties. Phenolics readily undergo oxidation, condensation, and protein interactions, making them integral to wine clarification, undesirable browning or pinking in white wines, and, most notably, to the color, taste, and indirectly the aroma of wine. When wine color is discussed, the primary focus is usually on red wine pigmentation.
The intensity and hue of red wine color are key sensory characteristics and are often the first quality perceived by consumers. In young red wines, color is largely due to free anthocyanins extracted from grape skins, whereas in aged wines it is shaped by oligomeric and polymeric pigments formed during vinification and aging [69,70,71,72]. Once extracted into the wine, anthocyanins undergo a series of chemical transformations, including enzymatic and non-enzymatic oxidation, which may negatively affect color stability. [73,74,75]. The concentration of free anthocyanins begins to decline as early as the end of alcoholic fermentation [76], likely due to adsorption onto grape solids or yeast cells, as well as enzymatic transformation by yeast-derived enzymes [77,78].
Anthocyanins also undergo condensation reactions with flavan-3-ols and procyanidins, either directly or via acetaldehyde bridges [79,80,81]. Some studies suggest that the formation of polymeric pigments may begin immediately after grape crushing, with evidence indicating that oligomeric and polymeric pigments can be extracted from grape skins in a model wine solution, even in the absence of acetaldehyde [41].
According to several authors, the initial step in the formation of more stable oligomeric and polymeric pigments is the development of co-pigmentation structures [82]. Co-pigmentation occurs when the planar (flat) portion of the anthocyanin molecule is stacked or overlapped by another compound known as the copigment [54]. Early theories proposed hydrogen bonding between the pigment and copigment [79,83], but later studies suggest that the interaction is driven primarily by hydrophobic forces and π–π stacking, resulting in a noncovalent complex [84]. Although copigments are typically colorless, their association with anthocyanins increases color intensity (a hyperchromic effect). In addition, a bathochromic shift occurs—that is, the absorption maximum of the anthocyanin–copigment complex shifts to longer wavelengths, thereby enhancing the blue hue of the wine [85,86,87].
Figure 6.
Structural transformations of flavylium ion.
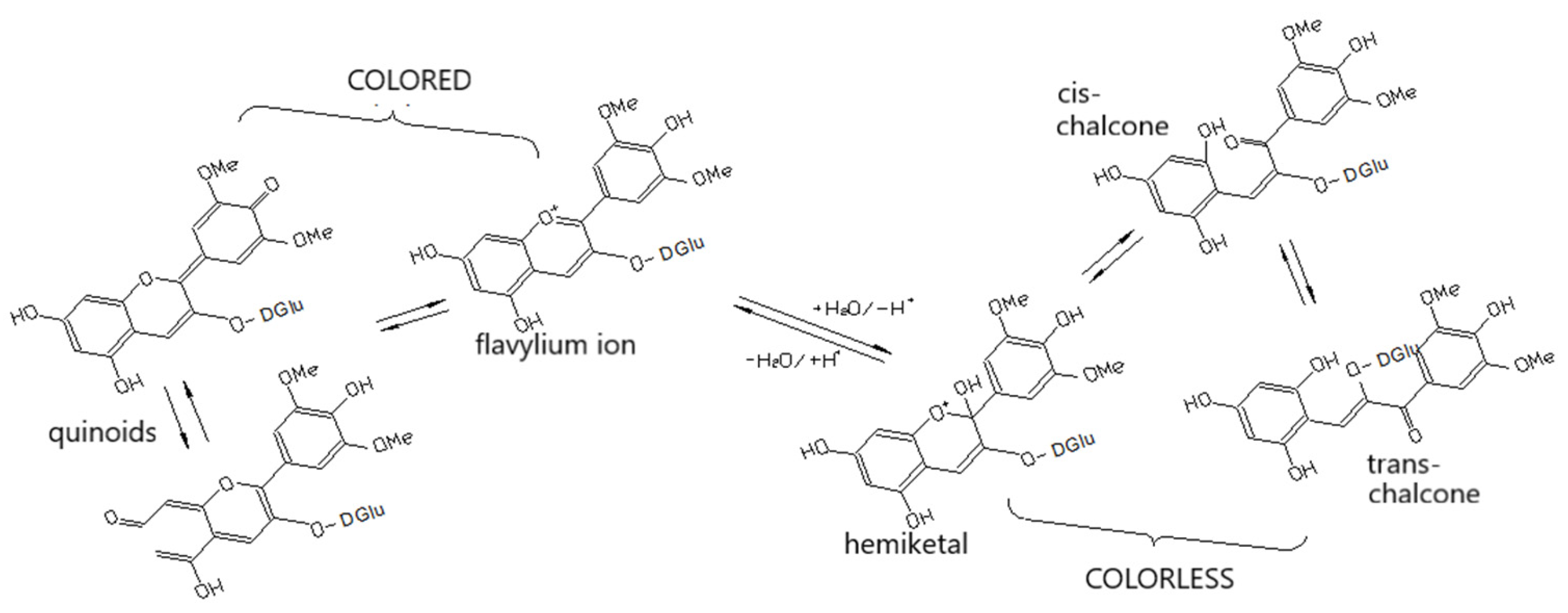
The potential to enhance color through co-pigmentation with colorless phenolics has been the subject of extensive research [82,89,90]. These studies have examined not only the role of anthocyanins themselves [56,85,86,88] but also the influence of environmental factors. In general, the copigmentation effect increases with the degree of anthocyanin methoxylation and glycosylation [88]. Additionally, acylated anthocyanins can engage in both intermolecular and intramolecular co-pigmentation, providing greater potential for the formation of stable pigment complexes [87]. This may explain why wines with similar anthocyanin concentrations can exhibit markedly different color intensities.
Significant differences in anthocyanin content and profile are observed not only between grape varieties [42,52] but also between clones of the same variety [55]. For example, in cultivars such as Pinot Noir and Sangiovese, both the lack of acylated anthocyanins and the low concentration of copigments may explain why deeply colored grapes sometimes yield pale wines [87].
Some research has examined the effect of the pigment-to-copigment ratio, showing a linear increase in copigmentation with rising copigment concentration [85,86,90,91,92,93]. This may explain why wines fermented in the presence of wood chips or ground dried stems exhibit greater color intensity compared with the same wines fermented in stainless steel tanks.
Despite the high concentration of proanthocyanidins in wine, their role as copigments remains relatively underexplored. Some studies [87,94] emphasize the need for further research into copigmentation across different grape varieties and winemaking conditions, particularly during alcoholic fermentation and aging.
Estimates suggest that 30–50% of the color in young red wines is attributable to copigmentation [87,95]. However, the low stability of co-pigmentation structures means that this effect diminishes over time during wine storage and processing. Some authors even argue that copigmentation may account for more than 50% of color in young red wines, and that insufficient copigment concentrations may be a major reason why many wines display low color intensity.
As early as 1981, an increase in the color of a red dessert wine was observed following the addition of white grape skin extract. Similar color-enhancing effects have been reported after adding enological tannins to young red wines [96,97,98]. However, the organoleptic impact of enological tannins varies depending on their origin, and their use does not always improve the sensory quality of the wine.
4.2. Influence of the Phenolic Compounds on the Wine Taste
Bitterness and astringency are fundamental to the sensory perception of red wines. Bitterness is defined as a gustatory sensation perceived through taste receptors, while astringency is considered a tactile sensation—commonly described as drying, tightening, or puckering of the oral epithelium—typically caused by compounds such as alum or tannins [98,99,100].
In wine, astringency is mainly associated with phenolic compounds, particularly those that are capable of precipitating salivary proteins and thereby reducing their lubricating properties [99,101]. The type of phenolic compound and its molecular size strongly influence whether the sensation is perceived as bitterness or astringency.
Several studies have shown that monomeric anthocyanins contribute little to taste perception [57], and neither monoglycosides nor their acylated forms significantly affect bitterness or astringency [99]. In contrast, flavanols and monomeric flavonols are generally more bitter than astringent [102], while procyanidins are chiefly responsible for astringency. Their ability to interact with proteins increases with their mean degree of polymerization (mDP) [103,104,105] and degree of galloylation (DG) [106,107]. These characteristics explain the increased astringency and harshness often observed in wines made from under-ripe grapes, which typically contain highly polymerized and galloylated procyanidins [100,108].
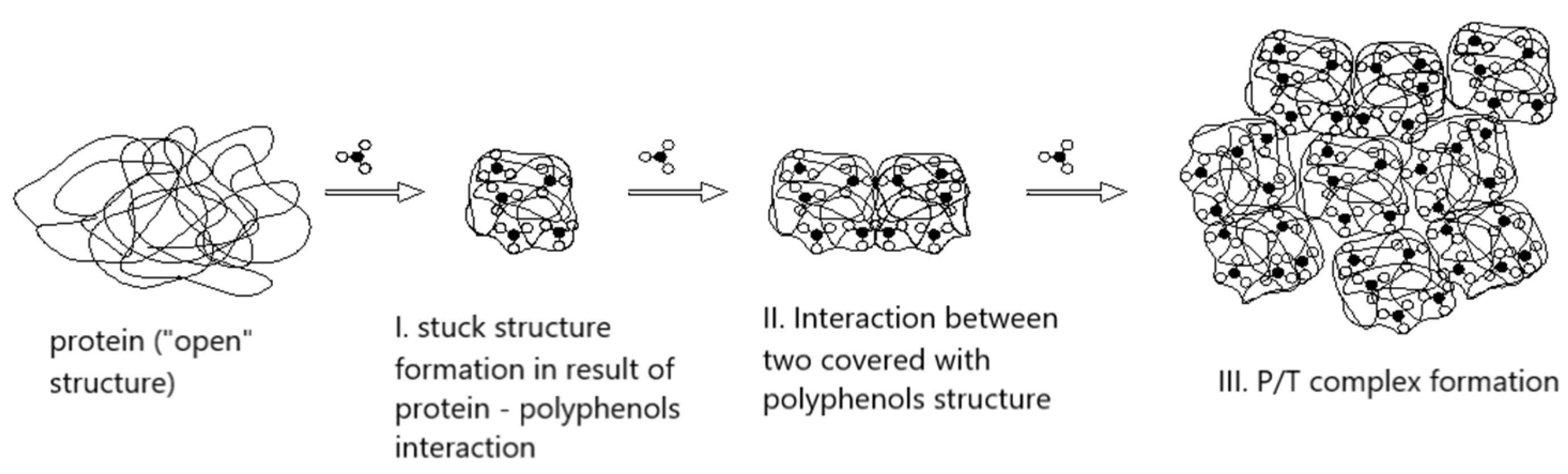
A study of 21 red wines—including Zinfandel, Pinot Noir, Merlot, Cabernet Sauvignon, Grenache, and Barbera—reported monomeric phenolic concentrations (expressed as epicatechin equivalents) ranging from 297.6 to 957.1 mg/L, while polymeric phenolics ranged from 901.3 to 2974.4 mg/L [94]. The polymeric-to-monomeric ratio varied from 1.9 to 9.2, with typical values around 3.9 for Merlot and 4.5 for Cabernet Sauvignon. The higher abundance of polymeric phenolics, togeher with typical wine procyanidins having an mDP of 5.81 to 12.33 [108], explains why astringency tends to dominate over bitterness in red wines. This predominance has made astringency a major focus of both sensory and chemical research. Astringency exhibits a delayed onset, typically requiring approximately 15–20 seconds to fully develop [100,109]. This is thought to be caused by the interaction of polyphenols with basic salivary proline-rich proteins (PRPs). The model proposed by E. Jőbstl [109] describes three stages (see Figure 7):
1. Initial interaction between phenolics and open, flexible protein structures, leading to the formation of compact protein–tannin complexes.
2. Further aggregation of these complexes through mutual binding.
3. Formation of larger cross-linked networks, ultimately leading to precipitation, which underlies the astringent mouthfeel.
Interactions between procyanidins and proteins are mediated by both hydrogen bonding [110] and hydrophobic interactions [111], with experimental evidence supporting both mechanisms [112]. The aromatic rings and C–H backbone of the pyran ring in procyanidins provide multiple hydrophobic binding sites, further stabilized by hydrogen bonds formed via o-dihydroxyphenol groups.
The ability of tannins to bind proteins depends on several factors, including protein molecular weight, structure, and amino acid composition [104,110,113,114,115]. Proteins with high proline content, hydrophobicity, and a flexible, open structure generally exhibit the strongest affinity for tannin binding [103,106,116,117]. Human saliva contains proline-rich proteins (PRPs), which constitute approximately 70% of the total protein content, along with glycosylated PRPs and α-amylase (both glycosylated and non-glycosylated forms) [104,105]. PRPs typically contain 28–40% proline, and together with glutamine and glycine, these three amino acids account for 75–80% of their structure. The aromatic ring of procyanidins is believed to interact with proline residues via flat “face-to-face” stacking, forming a stable protein–tannin complex [103].
Using nephelometric assays, De Freitas and Mateus [106,117] investigated interactions between grape seed procyanidins and three model proteins: PRPs, α-amylase, and bovine serum albumin (BSA). Their results indicated that BSA does not interact with monomeric flavan-3-ols, likely due to its globular structure and poor similarity to salivary proteins, making it a less suitable model despite its widespread use. Nevertheless, Herbertson et al. [40,118] developed a widely accepted method using BSA precipitation to estimate tannin concentration in wine. However, this method excludes dimers and trimers, which exhibit limited interaction with PRPs and minimal sensory impact, even in model wine solutions [40,94,99].
Higher molecular weight procyanidins are more prone to interact with PRPs, which may function as a natural protective mechanism against excessive astringency. Once bound, these procyanidins cannot further interact with other salivary glycoproteins, such as mucins, which have higher molecular mass and more effective lubricating properties [114]. Consequently, assessing wine astringency using BSA may underestimate the astringency potential, since phenolics that strongly bind PRPs show weak affinity for BSA.
Studies have reported that procyanidins with an mDP up to 3.5 account for 14.9–38.7% of procyanidins extracted from grape seeds of Castelão, Syrah, Touriga Nacional, and Cabernet Sauvignon, and 22.4–44.3% of the same fractions in wines made from these varieties [119].
The critical role of bitterness—and particularly astringency—in the sensory profile of red wines underscores the need for more objective and accurate evaluation methods, particularly those assessing procyanidin structure, including mDP and DG. Given their practical relevance, determination of these parameters should aim to be simple, fast, and cost-effective in terms of laboratory reagents and equipment.
5. Extractability of Grape Phenolic Compounds
Given their significant impact on wine quality and sensory characteristics, the extraction and transfer of phenolic compounds from grape solids into the wine is a key focus of many winemaking practices. In red winemaking, the primary phenolic compounds are extracted from grape skins, with a smaller proportion derived from grape seeds. Grapes are typically destemmed prior to crushing, which minimizes phenolic contributions from stems. However, stem residues (ranging from 0.05% to 0.1% of crushed grapes, depending on the crushing method) may still be present and contribute to phenolic extraction. Phenolics from grape stems may also be introduced when whole bunches are used or when dried stems are added—usually at levels of 1–3%—to enhance the mouthfeel of the final wine [120].
Nonetheless, the use of non-destemmed grapes can increase methoxypyrazine concentrations, contributing to green or vegetal notes in wine. In white winemaking, skin contact is typically brief, and the extraction of skin phenolics becomes relevant primarily when maceration techniques are applied, particularly in the production of wines from aromatic, non-pigmented grape varieties.
Although grape seeds contain higher concentrations of tannins, phenolics from grape skins generally dominate the final phenolic composition of the wine [121]. Longer maceration periods, however, increase the contribution of seed-derived phenolics [122,123]. The higher extractability of skin phenolics is attributed to their location within the vacuolar fluid and their association with the vacuolar membrane or cell wall of relatively fragile epidermal cells [36]. In contrast, grape seeds contain monomeric flavan-3-ols mainly in the outer soft coat, while procyanidins are located deeper, in the hard pigmented inner layer [124], although this structural configuration may change during prolonged fermentation [125].
Because of their direct influence on wine color and taste, the extractability of anthocyanins and procyanidins is frequently studied. A wide range of factors influence their extraction, including:
- Sulfur dioxide additions [128]
Anthocyanin extraction typically increases during the early stages of fermentation, peaking when alcohol content reaches 3–6% vol. Thereafter, their concentration begins to decline [76,134,136], while tannin extraction from skins and seeds continues [137]. This pattern does not apply uniformly across all anthocyanin types [132]; in some grape varieties, color intensity and anthocyanin content continue to rise with extended maceration [131].
Studies using model wine solutions indicate that anthocyanin extractability is 2–3 times higher in ripe grapes than in unripe ones. In ripe grapes, the extracted percentage of anthocyanins increases with alcohol concentration, reaching optimal extraction at around 9.0% v/v in unripe grapes [41].
The decline in anthocyanin concentration observed toward the end of fermentation is attributed to adsorption onto solids and interactions with tannins [57]. A study using labeled malvidin-3-glucoside showed that approximately 50% of extracted anthocyanins become associated with grape solids, forming polymeric pigments that are key to wine aging and stability [138]. These reactions influence the equilibrium between compounds remaining in the solids and those dissolved in the wine [87]. As anthocyanins and tannins bind, their free concentrations decrease, promoting further extraction of phenolics from the grape matrix [57,132].
This synergy between anthocyanins and tannins has led to exploration of practices that introduce additional phenolic sources, such as grape stems [25,132], grape seeds [137], enological tannins [140], or drained white grape pomace during red fermentation [87]. While these approaches can enhance color stability, they may also increase astringency, requiring careful calibration.
Beyond technological practices, the genetic predisposition of grape varieties plays a fundamental role in phenolic accumulation and extractability. The degree of polymerization (mDP) and degree of galloylation (DG) of procyanidins in skins, seeds, and stems affect not only their ability to form stable pigment complexes and bind proteins, but also their solubility and extractability. Possible interactions between tannins and cell wall polysaccharides during ripening may further modulate extractability [27].
The pectin matrix—comprising homogalacturonans (HGs), rhamnogalacturonans (RGs), and arabinogalacturonans (AGs)—along with cellulose and hemicellulose, accounts for around 90% of plant cell walls. Their degradation during fermentation significantly impacts the release of anthocyanins and tannins [141]. Once extracted, these compounds may be re-adsorbed by grape solids or yeast cells. HGs, especially those with low methylation, show strong affinity for anthocyanins, while RGs preferentially bind tannins [142]. Enzymatic treatments can aid in pectin solubilization, minimizing phenolic losses and improving stability [141,143].
6. Methods for Phenolic Compounds Determination
Accurate assessment of phenolics is crucial for understanding extractability and wine composition. The most widely used method for determining total phenolics is the Folin–Ciocalteu (FC) assay, first proposed by Singleton and Rossi [144]. This colorimetric method detects unoxidized phenolics based on their reducing capacity. Although inexpensive and widely accepted, it requires manual labor and is not ideal for high-throughput winery use, although automation has been developed [145].
Another method is the Somers method, based on UV absorbance at 280 nm, which captures both oxidized and unoxidized phenolics [70,71,146]. While rapid, this method lacks specificity, as non-phenolic compounds also absorb at this wavelength.
For detailed qualitative and quantitative profiling, HPLC remains the most effective method. It enables identification of individual phenolics using different stationary phases, mobile phases, and detectors [147,148]. However, since procyanidins represent 50–90% of grape phenolics, detailed identification of minor components may offer limited practical value.
Techniques such as acid-catalyzed degradation of procyanidins in the presence of thiol reagents have been developed to determine terminal and extension units, mDP, and DG [149,150]. Although these techniques require specialized equipment and trained personnel, they provide highly valuable data for winemaking.
One practical method, especially relevant for evaluating astringency, is the Adams–Harbertson BSA assay, which precipitates tannins using bovine serum albumin (BSA) [40,118]. Despite BSA’s limitations as a model for salivary proteins, this method provides a direct link between phenolic structure and sensory impact. Additionally, it allows quantification of pigments resistant to SO2 bleaching, such as oligomeric and polymeric pigments.
7. Conclusions
The extraction of grape phenolic compounds is influenced by multiple factors, including grape variety, maturity, and vinification techniques. The diversity and complexity of phenolics, along with the relative impact of these variables, make comprehensive analysis a challenging task. From a winemaking perspective, however, it is crucial to monitor not only the overall quantity and composition of phenolics but also their evolution during ripening, particularly in the critical period from veraison to harvest.
Although numerous analytical techniques are available, many remain time-consuming, costly, or require specialized equipment and expertise. Consequently, there is an increasing need for rapid, accessible, and reliable methods capable of providing general phenolic characterization as well as insights into their key sensory contributions—namely, color, astringency, and bitterness.
Recent advances in spectroscopic techniques, such as FT-NIR and mid-infrared spectroscopy, offer promising opportunities for rapid, non-destructive phenolic profiling. Combined with AI-driven data analysis, these tools may soon enable winemakers to make real-time decisions regarding optimal harvest timing and the most appropriate winemaking strategies, tailored to the grape’s phenolic maturity and the desired wine style.
Author Contributions
Conceptualization, N.S and T.Y.; writing—original draft preparation, N.S. and. S.T.; writing—review and editing, V.S and T.Y.; supervision, G.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Data Availability Statement
Data derived from public domain resources
Conflicts of Interest
The authors declare no conflicts of interest
References
- Elshafie, H.S.; Camele, I.; Mohamed, A.A. A Comprehensive Review on the Biological, Agricultural and Pharmaceutical Properties of Secondary Metabolites Based-Plant Origin. International Journal of Molecular Science 2023, 24, 3266. [Google Scholar] [CrossRef]
- Saini, N. , Anmol A., Kumar S., Wani A., Bakshi M., Dhiman Z.. Exploring phenolic compounds as natural stress alleviators in plants—a comprehensive review. Physiological and Molecular Plant Pathology 2024, 133, 102383. [Google Scholar] [CrossRef]
- Aherne, S.A.; O’Brein, N.M. Dietary flavonols: chemistry, food content and metabolism. Nutrition 2002, 18, 75–81. [Google Scholar] [CrossRef]
- Bravo, L. Polyphenols: chemistry, dietary sources, metabolism and nutritional significance. Nutrition Reviews. 1998, 56, 317–333. [Google Scholar] [CrossRef]
- Harbrone, J.B.; Williams, C.A. Advances in flavonoids research since 1992. Phytochemistry. 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Robards, K.; Prenzle, P.D.; Tucker, G.; Swatsitang, P.; Glover, W. ; Phenolic compounds and their role in oxidative processes in fruits. Food Chemistry. 1999, 66, 401–436. [Google Scholar] [CrossRef]
- Taiz L.; Zeiger E.; Stress Physiology, Plant Phisiology. 3th ed.Sinauer Associates Publisher, Sunderland, Massachusetts, 2003, pp.591.
- Delgado-Vargas, F.; Jimenez-Aparcio, A.R.; Paredes-Lopez, O. Natural pigments: carotenoids, anthocyanins and betalains—characteristics, biosynthesis, processing and stability. Critical Reviews in Food Science and Nutrition. 2000, 40, 173–289. [Google Scholar] [CrossRef]
- Stinzing, F.C.; Carle, R. ; Functional properties of anthocyanins and betalains in plants, food, and in human nutrition. Trends in Food Science and Technology. 2004, 15, 19–38. [Google Scholar] [CrossRef]
- Songs, J.; Smart, R.; Wang, H.; Dambergs, B.; Sparrow, A.; Qian, M.C. Effect of grape bunch sunlight exposure and UV radiation on phenolics and volatile composition of Vitis vinifera L. cv. Pinot noir wine. Food Chemistry. 2015, 173, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Zaya, J.; Trousdale, E.; Salgues, M. Caftaric acid in grapes and conversion into a reaction product during processing. Vitis. 1984, 23, 113–120. [Google Scholar]
- Singleton, V.L.; Zaya, J.; Trousdale, E. Compositional changes in ripening grapes: cafftaric and coutaric acids. Vitis. 1986, 25, 107–117. [Google Scholar]
- Goulao, L. , Fernandes J., Lopes P., Amancio S. Tacking the cell wall of the grape berry. The Biochemistry of the Grape Berry. 2012, 172–193. [Google Scholar]
- Clifford, M. Chlorogenic acids and other cinamates—nature, occurrence, dietary burden, absorption and metabolism. Journal of the Science of Food and Agriculture. 2000, 80, 1033–1043. [Google Scholar] [CrossRef]
- Šikuten, I. , Štambuk P. , Andabaka Ž., Tomaz I., Markovic Z., Stupic D.,Maletic E., Kontic J., Preiner D. Grapevine as a rich source of polyphenolic compounds. Molecules. 2020, 25, 5604. [Google Scholar] [PubMed]
- Buiarelli, F.; Coccioli, F.; Merolle, M.; Jasionawska, R.; Terracciano, A. Identification of hydroxycinnamic acid-tartaric acid esters in wine by HPLC—tandem mass spectrometry. Food Chemistry. 2010, 123, 827–833. [Google Scholar] [CrossRef]
- Mitic, M.; Obradovic, M.; Grabovac, Z.; Pavlovic, A. Antioxidant Capacities and phenolic levels of different varieties of Serbian white wines. Molecules 2010, 15, 2016–2027. [Google Scholar] [CrossRef]
- Darias-Martin, J.; Martin-Luis, B.; Carrilo-Lopez, M.; Lamuela-Raventos, R.; Diaz-Romero, C.; Boulton, R. Effect of caffeic acid on the color of red wine. Journal of Agricultural and Food Chemistry. 2002, 50, 2062–2067. [Google Scholar] [CrossRef]
- Adrean, M.; Jeandet, P.; Breuil, A.; Levite, D.; Debord, S.; Bessis, R. Assay of resveratrol and derivative stilbenes in wines by direct injection high performance liquid chromatography. American Journal of Enology and Viticulture. 2000, 51, 37–41. [Google Scholar] [CrossRef]
- Haygarov, V.; Yoncheva, T.; Dimitrov, D. Study of resveratrol content in grapes and wine of the varieties Storgozia, Kaylashki Rubin, Trapezitsa, Rubin, Bouquet and Pinot noir. Journal of Mountain Agriculture on the Balkans. 2017, 20, 300–311. [Google Scholar]
- Tintunen, S.; Lehtonen, P. Distinguishing organic wines from normal wines on the basis of concentration of phenolic compounds and spectral data. European Food Research Technology. 2001, 212, 90–394. [Google Scholar] [CrossRef]
- Jordão, A.; Ricardo-da-Silva, J.; Laureano, O. Evolution of catechins and oligomeric procyanidins during grape maturation of Castelão francês and Touriga Francesa. American Journal of Enology and Viticulture. 2001, 52, 230–234. [Google Scholar] [CrossRef]
- Mateus, N.; Marques, S.; Gonçalves, A.; Machado, J.; De Freitas, V. Proanthocyanidin composition of red Vitis Vinifera varieties from the Douro valley during ripening: influence of cultivation altitude. American Journal of Enology and Viticulture. 2001, 52, 230–234. [Google Scholar] [CrossRef]
- Monagas, M. , Gomez-Cordoves C., Bartolome B., Laureano Ol, Ricardo da Silva J. Monomeric, oligomeric, and polymeric flavan-3-ol composition of wines and grapes from Vitis vinifera L. cv. Graciano, Tempranillo and Cabernet Sauvignon. Journal of Agricultural and Food Chemistry 2003, 51, 6475–6481. [Google Scholar] [CrossRef]
- Sun, B.; Pinto, T.; Leandro, M.; Ricardo-da-Silva, J.; Spranger, M. Transfer of catechins and proanthocyanidins from solid parts of the grape cluster into wine. American Journal of Enology and Viticulture. 1999, 50, 179–183. [Google Scholar] [CrossRef]
- Kennedy, J.; Troup, G.; Pilbrow, J.; Hutton, D.; Hewitt, D.; Humter, C.; Ristic, R.; Iland, P.; Jones, G. Development of seed polyphenols in berries from vitis vinifera cv. Shiraz. Australian Journal of Grape and Wine Research. 2000, 6, 244–254. [Google Scholar] [CrossRef]
- Kennedy, J. , Hayasaka Y.; Vidal S.; Waters E.; Jones E. Composition of grape skin proanthocyanidins at different stage of berry development. Journal of Agricultural and Food Chemistry. 2001, 49, 5348–5355. [Google Scholar] [CrossRef] [PubMed]
- Blancquaert, E.; Oberholster, A.; Ricardo-da-Silva, J.; Deloire, A. Grape flavonoid evolution and composition under altered light and temperature conditions in Cabernet Sauvignone (Vitis vinifere L.). Front Plant Science. 2019, 8, 1062. [Google Scholar] [CrossRef] [PubMed]
- De Freitas, V.; Glories, Y.; Monique, A. Development changes of procyanidins in grape of red Vitis vinifera varieties and their composition in respective wines. American Journal of Viticulture and Enology. 2000, 51, 397–403. [Google Scholar] [CrossRef]
- Kennedy, J.; Matthews, M.; Waterhouse, A. Changes in grape seed polyphenols during fruit ripening. Phytochemistry. 2000, 55, 77–85. [Google Scholar] [CrossRef]
- Jordão, A.; Ricardo-da-Silva, J.; Laureano, O. Evolution of proanthocyanidins in bunch stems during berry development (Vitis vinifera L.). Vitis. 2001, 40, 17–22. [Google Scholar]
- Ojeda, H.; Andary, C.; Kraeva, E.; Carbonneau, A.; Deloire, A. Influence of pre- and postveraison water deficit on synthesis and concentration of skin phenolic compounds during berry growth of vitis vinifera cv. Shiraz. American Journal of Enology and Viticulture. 2002, 53, 261–267. [Google Scholar]
- Ricardo-da-Silva, J.; Rigaud, J.; Cheynier, V.; Cheminat, A.; Moutounet, M. Procyanidin dimers and trimers from grape seeds. Phytochemistry. 1991, 30, 1259–1264. [Google Scholar] [CrossRef]
- Padilla-Gonzalez, G. , Grosskorf E., Sadgrove N., Simmonds M. Chemical diversity of flavan-3-ols in grape seeds: modulating factors and quality requirements. Plants 2022, 11, 809. [Google Scholar] [CrossRef]
- Prieur, C.; Rigaud, J.; Cheynier, V.; Moutonet, M. Oligomeric and polymeric procyanidins from grape seeds. Phytochemistry 1994, 36, 781–784. [Google Scholar] [CrossRef]
- Souquet, J.; Cheynier, V.; Brossaud, F.; Moutounet, M. Polymeric procyanidins from grape skin. Phytochemistry 1996, 43, 509–512. [Google Scholar] [CrossRef]
- Yu, J. , Ahmedna. Functional component of grape pomace: their composition, biological properties and potential applications. Food Science and Technologies 2013, 48, 221–237. [Google Scholar]
- De Freitas, V.; Glories, Y. Concentration and compositional changes of procyanidins in grape seeds and skins of white vitis vinifera varieties. Journal of the Scienceof Food and Agriculture. 1998, 79, 1601–1606. [Google Scholar] [CrossRef]
- Obreque-Slier, E.; Pena-Neira, A.; Lopez-Solis, R.; Zamora-Marin, F.; Ricardo-da-Silva, J.; Laureno, O. Comparative study of the phenolic composition of seeds and skins from Carmenere and Cabernet Sauvignon grape varieties (Vitis vinifera L.) during ripening. Agricultural and Food Chemistry. 2010, 58, 3591–3599. [Google Scholar] [CrossRef]
- Harbertson, J.; Kennedy, J.; Adams, D. Tannin in skins and seeds of Cabernet sauvignon, Syrah and Pinot noir berries during ripening. American Journal of Enology and Viticulture 2002, 53, 54–59. [Google Scholar] [CrossRef]
- Stoyanov, N.; Mitev, P.; Galabova, M.; Tagareva, S. Phenolic compounds extractability from Melnik 55 grape solid parts during grape maturity. Bio Web of Conference. 2023, 58, 01016. [Google Scholar] [CrossRef]
- Kammerer, D.; Claus, A.; Carle, R.; Schieber, A. Polyphenol screening of pomace from red and white grape varieties (Vitis vinifera) by HPLC-DAD-MS/MS. Journal of Agricultural and Food Chemistry. 2004, 52, 4360–4367. [Google Scholar] [CrossRef]
- Burns, J.; Cardner, P.; O’Neil, J.; Crawford, S.; Morecroft, I.; McPhail, D.; Lister, C.; Matthews, D.; Maclean, M.; Lean, M.; Duthie, G.; Croizier, A. Relationshipamong antioxidant activity, vasodilation capacity and phenolic content of red wines. , Journal of Agricultural and Food Chemistry. 2000, 48, 220–230. [Google Scholar] [CrossRef]
- Kilmartin, P.; Zou, H.; Waterhouse, A. A cyclic voltammetry method suitable for characterizing antioxidant properties of wine and wine phenolics. Journal of Agricultural and Food Chemistry. 2001, 49, 1957–1965. [Google Scholar] [CrossRef]
- Frankel, E.; Waterhouse, A.; Teissedre, P. Principal phenolic phitochemicals in selected California wines and their antioxidant activity in inhibiting oxidation of human low-density lipoproteins. Journal of Agricultural and Food Chemistry. 1995, 43, 890–894. [Google Scholar] [CrossRef]
- Sanchez-Moreno, C.; Larrauri, C.; Saura-Calixto, F. Free radical scavenging capacity and inhibition of lipid oxidation of wines, grape juices and related polyphenolic constituents. Food Research International 1999, 32, 407–412. [Google Scholar] [CrossRef]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Orgiu, F.; Frigerio, G.; Regazzoni, L.; Sousa, L.; Bavaresco, L.; Bosso, A.; Aldini, G.; Restani, P. Phenolic profile and antioxidant activity of different grape (Vitis vinifera L.) varieties. BIO Web of Conference. 2019, 12, 04005. [Google Scholar] [CrossRef]
- Radoeva, R.; Yankova, I.; Enchev, B.; Karsheva, M.; Ivanova, E.; Iliev, I. Poluphenols of grape pomace from local Bulgarian variety Mavrud. Antioxidant and antitumor effect against breast cancer., Journal of Chemical Technology and Metallurgy. 2022, 57, 508–521. [Google Scholar]
- He, F. , Mu L., Yan G.L., Liang N., Pan Q., Wang J., Reeves M., Duan C. Biosynthesis of anthocyanins and their regulation in colored grapes. Molecules. 2010, 15, 9057–9091. [Google Scholar] [CrossRef] [PubMed]
- Harbrone, J.; Williams, C. Anthocyanidins and other flavonoids. Natural Products Report. 2004, 4, 539–573. [Google Scholar]
- Yoncheva, T.; Kostov, G.; Spasov, H. Content of total phenolic compounds, anthocyanins and spectral characteristics of Gamza red wines depending on the alcohol fermentation conditions. Bulgarian Journal of Agricultural Science. 2023, 29, 159–170. [Google Scholar]
- George, F.; Figueiredo, P.; Toki, K.; Tatsuzawa, F.; Saito, N.; Brouillard, R. Influence of trans-cis isomerization of coumaric acid substituents on colour variance and stabilization in anthocyanins. Phitochemistry. 2001, 57, 791–795. [Google Scholar] [CrossRef]
- Favre, G. , Hermosin-Gutierrez I., Piccardo D., Gomez-Alonso S., Gonzalez-Neves G.. Selectivity of pigments extraction from grapes and their partial retention in the pomace during red-winemaking. Food Chemistry. 2019, 277, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Malien-Aubert, C.; Dangles, O.; Amiot, M. Color stability of commercial anthocyanin -based extracts in relation to the phenolic composition. Protective effect by intra- and intermolecular copigmentation. Journal of Agricultural and Food Chemistry. 2001, 49, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.; Mullen, W.; Landrault, N.; Teissedre, P.; Lean, M.; Crozier, A. Variation in the profile and content of anthocyanins in wines made from Cabernet sauvignon and hybrid grapes. Journal of Agricultural and Food Chemistry. 2002, 50, 4096–4102. [Google Scholar] [CrossRef]
- Mazza, G. Anthocyanins in grape and grape products. Critecal Reviews in Food Science and Nutrition. 1995, 35, 341–371. [Google Scholar] [CrossRef]
- Singleton, V.; Trousdale, E. Anthocyanin-tannin interaction expalaining differences in polymeric phenols between white and red wines. American Journal of Enology and Viticulture. 1992, 43, 63–70. [Google Scholar] [CrossRef]
- Benmeziane, F. , Cadot Y. , Djamai R., Djermoun L.Determination of major anthocyanin pigments and flavonols in red grape skin of some table grape varieties (Vitis vinifer sp.) by high-performance liquid chromatography—photodiode array detection (HPLC-DAD). Oeno One 2016, 50, 125–135. [Google Scholar]
- Romero-Cascales, I.; Ortega, A.; Lopez-Roca, J.; Fernandez, J.; Gomez-Plaza, E. ; Differences in anthocyanins extractability from grapes to wines according to variety. American Journal of Enology and Viticulture. 2005, 56, 212–219. [Google Scholar] [CrossRef]
- Kennedy, J.; Matthews, M.; Waterhous, A. Effect of maturity and vine water status on grape skin and wine flavonoids. American Journal of Enology and Viticulture. 2002, 53, 268–274. [Google Scholar] [CrossRef]
- Roggero, J.; Coen, S.; Ragonnet, B. High performance liquid chromatography survey on changes in pigment content in ripening prapes of Syrah. An approach to anthocyanin metabolism. American Journal of Enology and Viticulture. 1986, 37, 77–83. [Google Scholar] [CrossRef]
- Gonzalez-San Jose, M.; Barron, L.; Diez, C. Evolution of anthocyanins during maturation of Tempranillo grape variety (Vitis vinifera) using polynominal regression models. Journal of the Science of Food and Agriculture. 1990, 51, 337–343. [Google Scholar] [CrossRef]
- Costa, E.; Cosme, F.; Jordão, A.; Mendes-Faia, A. Anthocyanins profile and antioxidant activity from 24 grape varieties cultivated in two Portuguese wine region. Journal International des Science de la Vigne et du Vin. 2014, 48. [Google Scholar] [CrossRef]
- Chorti, E. , Guidoni S., Ferrandino A., Novello V. Effect of different cluster sunlight exposure levels on ripening and anthocyanin accumulation in Nebbiolo grape. American Journal of Enology and Viticulture 2010, 61, 23–30. [Google Scholar] [CrossRef]
- Yan, Y. , Song C., Falginella L., Castellarin S. Day temperature has a stronger effect than night temperature on anthocyanin and flavonol accumulation in Merlot (Vitis vinifera L.) grape during ripening. Frontiers in Plant Science 2020, 11, 1095. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Moreno, A.; Perez-Alvarez, E.; Lopez-Urrea, R.; Paladinez-Quezada, D.; Moreno-Olivares, J.; Intrigliolo, D.; Gil-Munoz, R. Effect of deficit irrigation with saline water on wine color and polyphenolic composition of Vitis vinifera L. cv. Monastrell. Scientia Horticulturae. 2021, 283, 110085. [Google Scholar] [CrossRef]
- Theocharis, S.; Nikolau, N.; Zioziou, E.; Kyraleou, M.; Kallithraka, S.; Kotseridis, Y.; Koundouras, S. Effect of post-veraison irrigation on the phenolic composition of Vitis vinifera L., cv. Xinomavaro grapes. Oeno One. International Viticulture and Enology Society. 2021, 55, 173–189. [Google Scholar]
- Torres, N.; Martinez-Luscher, J.; Porte, E.; Yu, R.; Kurtural, S. Impact of leaf removal and shoot thinning on cumulative daily light intensity and thermal time and their cascading effect of grapevine (Vitis vinifera L.) berry and wine chemistry in warm climates. Food Chemistry. 2021, 343, 128447. [Google Scholar] [CrossRef]
- Saucier, C.; Little, D.; Glories, Y. First evidence of acetaldehyde-flavonol condensation products in red wine. American Journal of Enology and Viticulture. 1997, 48, 370–373. [Google Scholar] [CrossRef]
- Somers, T. The polymeric nature of wine pigments. Phytochemistry. 1971, 10, 2175–2186. [Google Scholar] [CrossRef]
- Somers, T. Interactions of color composition in young red wines. Vitis. 1978, 17, 161–167. [Google Scholar]
- Somers, T.; Evans, M. Evolution of red wines. An assessment of the role of acetaldehyde. Vitis. 1986, 25, 31–39. [Google Scholar]
- Caestellari, M.; Arfelli, G.; Riponi, C.; Amati, A. Evolution of phenolic compounds in red winemaking as affected by must oxidation. American Journal of Enology and Viticulture. 1998, 49, 91–94. [Google Scholar] [CrossRef]
- Rio Segade S,. Torchio F., Giacosa S., Aimonino D., Gay P., Lambri M., Dordoni R., Gerbi V., Rolle L. Impact os several pre-treatments on the extraction of phenolic compounds in winegrape varieties with different anthocyanin profiles and skin mechanical properties. Journal of Agricultural and Food Chemistry 2014, 62, 8437–8451. [CrossRef] [PubMed]
- Yokotsuka, K.; Singleton, V. Effects of seed tannins on enzymatic decolorization of wine pigment in the presence of oxidizable phenols. American Journal of Enology and Viticulture. 2001, 52, 93–100. [Google Scholar] [CrossRef]
- Gil-Munoz, R. , Gomez-Plaza E., Martinez A., Lopez-Roca. Evolution of phenolic compounds during wine fermentation and post-fermentation: influence of grape temperature. Journal of Food Composition and Analysis 1999, 12, 259–272. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Dellacassa, E.; Carrau, F. Yeast interaction with anthocyanins during red wine fermentation. American Journal of Enology and Viticulture. 2005, 56, 104–109. [Google Scholar] [CrossRef]
- Wenzel, K. Die selection einer hefemutante zur verminderung der farstoffverluste wahreng der rotweingarung. Vitis. 1989, 28, 111–120. [Google Scholar]
- Escott, C. , Morata A., Zamora F., Loira I., Manuel del Fresno J., Saurez-Lepe J. Study of the interaction of anthocyanins with phenolic aldehydes in a model wine solution. ACS Omega 2018, 3, 15575–15581. [Google Scholar] [CrossRef]
- Marquez, A. , Serratosa M., Merida J. Pyranoanthocyanin derived pigments in wine: Structure and formation during winemaking. Journal of Chemistry 2013, 2013, 713028. [Google Scholar] [CrossRef]
- Sheridan, M.; Elias, R. Reaction of acetaldehyde with wine flavonoids in the presence of sulfur dioxide. Journal of the Agricultural and Food Chemistry. 2016, 3, 8615–8624. [Google Scholar] [CrossRef]
- Liao, H. , Cai Y., Haslam E. Polyphenol interactions. Anthocyanins: Copigmentation and colour changes in red wines. Journal of the Science of Food and Agriculture. 1992, 59, 299–305. [Google Scholar] [CrossRef]
- Asen, S.; Stewart, R.; Norris, K. Co-pigmentation of anthocyanins in plant tissues and its effect on color. Phytochemistry. 1972, 11, 1139–1145. [Google Scholar] [CrossRef]
- Zhao, C. , Yu Y., Chen Z., Wen Z., Wei F., Zheng Q., Wang C., Xiao X. Stability-increasing effects of anthocyanin glycosyl acylation. Food Chemistry 2017, 214, 119–128. [Google Scholar] [CrossRef]
- Davies, A.; Mazza, G. Copigmentation of simple and acylated anthocyanins with colorless phenolic compounds. Journal of Agricultural and Food Chemistry. 1993, 41, 716–720. [Google Scholar] [CrossRef]
- Mazza, G.; Brouillard, R. The mechanism of co-pigmentation of anthocyanins in aqueous solution. Phytochemistry. 1990, 29, 1097–1102. [Google Scholar] [CrossRef]
- Boulton, R. The copigmentation of anthocyanins and its role in the color of red wine: A critical reviews. American Journal of Enology and Viticulture. 2001, 52, 67–87. [Google Scholar] [CrossRef]
- Mirabel, M.; Saucier, C.; Guerra, C.; Glorie, Y. Copigmentation in model wine solutions: Occurrence and relation to wine ageing. American Journal of Enology and Viticulture. 1999, 50, 211–218. [Google Scholar] [CrossRef]
- Haslam, E. In vino veritas. Oligomeric procyanidins and the ageing of red wines. Phytochemistry. 1980, 19, 2577–2582. [Google Scholar]
- Chen, L.; Hrazdina, G. Structural properties of anthocyanins-flavonoid complex formation and its role in plant copor. Phytochemistry. 1981, 20, 297–302. [Google Scholar] [CrossRef]
- Baranac, J.; Petranovic, N.; Dimitric-Marcovic, J. Spectrophotometric study of anthocyanin copigmentation reactions. Journal of Agricultural and Food Chemistry. 1996, 44, 1333–1336. [Google Scholar] [CrossRef]
- Markovic, J.; Petranovic, N.; Baranac, J. A spectrophotometric study of the copigmentation of malvin with caffeic and ferulic acid. Journal of Agricultural and Food Chemistry. 2000, 48, 5530–5536. [Google Scholar] [CrossRef]
- Baranac, J.; Petranovic, N.; Dimitric-Marcovic, J. Spectrophotometric study of anthocyanin copigmentation reactions 2 Malvin and the nonglycosidez flavone quercetin. Journal of Agricultural and Food Chemistry. 1997, 45, 1694–1697. [Google Scholar] [CrossRef]
- De Beer, D. , Harbertson J.; Kilmartin P.; Roginsky V.; Barsukova T.; Adams D.; Waterhouse A. Phenolics: A comparison of diverse analytical methods. American Journal of Enology and Viticulture. 2004, 55, 389–400. [Google Scholar] [CrossRef]
- Sacchi, K.; Bisson, L.; Adams, D. A review of the effect of winemaking techniques on phenolic extraction in red wines. American Journal of Enology and Viticulture. 2005, 56, 197–206. [Google Scholar] [CrossRef]
- Lempereur, V.; Blateyron-Pic, L.; Labarde, B.; Saucier, C.; Kelebek, H.; Glories, Y. Groupe national de travail sur les tanins oenologiques: premier resultats. Review Frances DOenologique. 2002, 196, 23–29. [Google Scholar]
- Naves, A.; Spranger, M.; Zhao, Y.; Leandro, M.; Sun, B. Effect of addition of commercial grape seed tannins on phenolic composition, chromatic characteristics and antioxidant activity of red wine. Journal of Agriculture and Food Chemistry. 2010, 58, 11775–11782. [Google Scholar] [CrossRef]
- Arnold, R.; Noble, A. Bitternes and astringencyof grape seed phenolics in a model wine solution. American Journal of Enology and Viticulture. 1978, 29, 150–152. [Google Scholar] [CrossRef]
- Vidal, S.; Francis, L.; Noble, A.; Kwiatowski, M.; Cheynier, V.; Waters, E. Taste and mouth-feel properties of different type of tannin-like polyphenolic compounds and anthocyanins in wine. Analitica Chimica Acta. 2004, 513, 57–65. [Google Scholar] [CrossRef]
- Brossaud, F.; Cheynier, V.; Noble, A. Bitterness and astringency of grape and wine polyphenols. Australian Journal of Grape and Wine Research. 2001, 7, 33–39. [Google Scholar] [CrossRef]
- Smith, A.; June, H.; Nobble, A. Effects of viscosity on the bitterness and astringency of grape seed tannin. Food Quality and Preference. 1996, 7, 161–166. [Google Scholar] [CrossRef]
- Peleg, H.; Gacon, K.; Schlich, P.; Noble, A. Bitterness and astringency of flavan-3-ol monomers, dimmers and trimers. Joutnal of the Science of Food and Agriculture. 1999, 79, 1123–1128. [Google Scholar] [CrossRef]
- Baxter, N.; Lilley, T.; Haslam, E.; Williamson, M. Multiple interactions between polyphenols and a salivary proline-rich protein repeat result in complexation and precipitation. Biochemistry. 1997, 36, 5566–5577. [Google Scholar] [CrossRef]
- Luck, G.; Liao, H.; Murray, N.; Grimmer, H.; Warminski, E.; Williamson, M.; Lilley, T.; Haslam, E. Polyphenols astringency and prolin-rich proteins. Phytochemistry. 1994, 37, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Sarni-Manchado, P.; Gheynier, V.; Moutonet, M. Interaction of grape seed tannins with salivary proteins. Journal of Agricultural and Food Chemistry. 1999, 47, 42–47. [Google Scholar] [CrossRef]
- De Freitas, V.; Mateus, N. Nephelometric study of salivary protein-tannin aggregates. Journal of the Science of Food and Agriculture. 2001, 82, 113–119. [Google Scholar] [CrossRef]
- Ricardo-da-Silva, J.; Cheynier, V.; Souquet, J.M.; Moutonet, M.; Cabanis, J.C.; Bourzeix, M. Interaction of grape seed procyanidins with various proteins in relation to wine finning. Journal of the Science of Food and Agriculture. 1991, 57, 111–125. [Google Scholar] [CrossRef]
- Maury, G.; Sarni-Manchado, P.; Lefebvre, S.; Cheynier, V.; Moutonet, M. Influence of fining with different molecular weight gelatin on procyanidin composition and precipitation of wines. American Journal of Enology and Viticulture. 2001, 52, 140–145. [Google Scholar] [CrossRef]
- Jőbstl, E.; O’Connell, J.; Fairclough, P.; Williamson, M. Molecular model for astringency produced by polyphenol/protein interactions. Biomacromolecules 2004, 5, 942–949. [Google Scholar] [CrossRef]
- Hagerman, A.; Butler, L. The scecificity of proanthocyanidin-protein interactions. The Journal of Biological Chemistry. 1981, 259, 4494–4497. [Google Scholar] [CrossRef]
- Oh, H.; Hoff, J.; Armstrong, G.; Haff, L. Hydrophobic interaction in tannin-protein complex. Journal of Agricultural and Food Chemistry. 1980, 28, 394–398. [Google Scholar] [CrossRef]
- Artz, W.; Bishop, P.; Dunker, K.; Schanus, E.; Swanson, B. Interaction of synthetic proanthocyanidin dimmer and trimer with bovine serum albumin and purified bean globulin fraction G-1. Journal of Agricultural and Food Chemistry. 1987, 35, 417–421. [Google Scholar] [CrossRef]
- Kawamoto, H.; Nakatsudo, F.; Murakani, K. Stoichometric studies of tannin-protein coprecipitation. Phytochemistry. 1996, 41, 1427–1431. [Google Scholar] [CrossRef]
- Sarni-Manchado, P.; Deleris, A.; Avallone, A.; Cheynier, V.; Moutonet, M. Analysis and characterization of wine condensed tannins precipitated by proteins used as fining agent in Enology. American Journal of Enology and Viticulture. 1999, 50, 81–86. [Google Scholar] [CrossRef]
- Yokotsuka, K.; Singleton, V. Interactive precipitation between phenolic fractions and peptides in wine-like model solutions:turbidity, partical size and residual content as influenced by pH, temperature and peptide concentration. American Journal of Enology and Viticulture. 1995, 46, 329–338. [Google Scholar] [CrossRef]
- Asquith, T.; Uhlig, J.; Mehansho, H.; Putman, L.; Carlson, D.; Butler, L. Binding of condensed tannins to salivary prolin-rich glycoproteins: the role of carbohydrate. Journal of Agricultural and Food Chemistry. 1987, 35, 331–334. [Google Scholar] [CrossRef]
- De Freitas, V.; Mateus, N. Structural features of procyanidin interactions with salivary proteins. Journal of Agricultural and Food Chemistry. 2001, 49, 940–945. [Google Scholar] [CrossRef]
- Harbertson, J.; Picciotto, E.; Adams, D. Measurement of pigments in grape berry extracts and wines using protein precipitation assay combined with bisulfite bleaching. American Journal of Enology and Viticulture. 2003, 54, 301–306. [Google Scholar] [CrossRef]
- Cosme, F.; Ricardo-Da-Silva, J.; Laureano, O. Tannin profiles of Vitis vinifera L. cv. Red grapes growing in Lisboa and from their monovaraietal wines. Food Chemistry. 2009, 112, 197–204. [Google Scholar] [CrossRef]
- Wimalasiri, P.; Olejar, K.; Harrison, R.; Hider, R.; Tian, B. Whole bunch fermentation and the use of grape stems: effect on phenolic and volatile aroma composition of Vitis vinifera cv. Pinot noir wine. Australian Journal of Grape and Wine Research. 2021, 28, 395–406. [Google Scholar] [CrossRef]
- Kennedy, J.; Robinson, S.; Walker, M. Grape and wine tannins: production, perfection, perception. Practical Winery and Vineyard 2007, 57–67. [Google Scholar]
- Kovac, V.; Alonso, E.; Revilla, E. The effect of adding supplementary quantity of seeds during fermentation on the phenolic composition of wines. American Journal of Enology and Viticulture. 1995, 46, 363–367. [Google Scholar] [CrossRef]
- Lee, J.; Kennedy, J.; Devlin, C.; Redhead, M.; Rennaker, C. Effect of early seed removal during fermentation on proanthocyanidin extraction in red wine: A commercial production example. Food Chemistry. 2008, 1270–1273. [Google Scholar] [CrossRef]
- Thorngate, J.; Singleton, V. Localization of procyanidins in grape seeds. American Journal of Enology and viticulture. 1994, 45, 259–262. [Google Scholar] [CrossRef]
- Gillispie, E.; Miller, K.; McElrone, A.; Block, D.; Rippner, D. Red wine fermentation alters grape seed morphology and internal porosity. American Journal of Enology and Viticulture. 2023, 74, 0740030. [Google Scholar] [CrossRef]
- Girard, B.; Kopp, T.; Reynolds, A.; Gliff, M. Influence of vinification treatments on aroma constituents and sensory descriptors of Pinot noir wines. American Journal of Enology and Viticulture. 1997, 48, 198–206. [Google Scholar] [CrossRef]
- Reynolds, A.; Cliff, M.; Girard, B.; Kopp, T. Influence of fermentation temperature on composition and sensory properties of Semillion and Shiraz wines. American Journal of Enology and Viticulture. 2001, 52, 235–240. [Google Scholar] [CrossRef]
- Bakker, J.; Bridle, P.; Bellworthy, S.; Garcia-Viguera, C.; Reader, H.; Watkins, S. Effect of sulfur dioxide and must extraction on colour, phenolic composition and sensory quality of red table wine. Journal of the Science of Food and Agriculture. 1998, 78, 297–307. [Google Scholar] [CrossRef]
- Rolle, L.; Torchio, F.; Ferrandino, A.; Guidoni, S. Influence of wine-grape skin hardness on the kinetics of anthocyanin extraction. International Journal of Food Properties 2012, 15, 249–261. [Google Scholar] [CrossRef]
- Budić-Leto, I.; Lovric, T.; Pezo, I.; Kljusuric, J. Study of dynamics of polyphenol extraction during traditional and advanced maceration processes of the Babic grape variety. Food Technology and Biotechnology. 2005, 43, 47–53. [Google Scholar]
- Gomez-Plaza, E.; Gil-Munoz, R.; Lopez-Roca, J.; Martinez-Cutillas, A.; Fernandez, J. Phenolic compounds and color stability of red wines: Effect of skin maceration time. American Journal of Enology and Viticulture. 2001, 52, 266–270. [Google Scholar] [CrossRef]
- Sims, C.; Baters, R. Effect of skin fermentation time on the phenols, anthocyanins, ellagic acid sediment, and sensory characteristics of a red vitis rotundifolia wine. American Journal of Enology and Viticulture. 1994, 45, 56–62. [Google Scholar] [CrossRef]
- Yokotsuka, K.; Sato, M.; Ueno, N.; Singleton, V. Colour and sensory characteristics of Merlot red wines caused by prolonged pomace contact. Journal of Wine Research. 2001, 11, 7–18. [Google Scholar] [CrossRef]
- Scudamore-Smith, P.; Hooper, R.; McLaren, E. Color and phenolic changes of Cabernet sauvignon wine made by simultaneous yeast/bacterial fermentation and extended pomace contact. American Journal of Enology and Viticulture. 1990, 41, 57–67. [Google Scholar] [CrossRef]
- Stoyanov, N.; Mitev, P.; Tagareva, S.; Kemilev, S. Influence of the process of cold maceration and non-Saccharomyces yeast application in red winemaking. Journal of Mountain Agriculture on the Balkans. 2017, 20. [Google Scholar]
- Gao, L.; Girard, B.; Mazza, G.; Reynolds, A. Changes in anthocyanins and color characteristics of Pinot noir wines during different vinification processes. Journal of Agricultural and Food Chemistry. 1997, 45, 2003–2008. [Google Scholar] [CrossRef]
- Xia, N. , Liu A., Qi M., Zhang H., Huang Y., He F., Duan C., Pan Q. Enhancing the color and astringency of red wines through white grape seeds addition: repurposing wine production byproducts. Food Chemistry 2024, 23, 101700. [Google Scholar]
- Zimman, A.; Waterhouse, A. Incorporation of malvidiv-3-glucoside into high molecular weight polyphenols during fermentation and wine ageing. American Juornal of Enology and Viticulture. 2004, 55, 139–146. [Google Scholar] [CrossRef]
- Kantz, K.; Singleton, V. Isolation and determination of polymeric polyphenols using Sephadex LH-20 and analysis of grape tissue extracts. American Journal of Enology and Viticulture. 1990, 41, 223–228. [Google Scholar] [CrossRef]
- Mateus, N.; Pinto, R.; Ruao, P.; De Freitas, V. Influence of the addition of grape seed procyanidins to Port wines in the resulting reactivity with human salivary proteins. Food Chemistry. 2004, 84, 195–200. [Google Scholar] [CrossRef]
- Garrido-Banuelos, G.; Buica, A.; Du Toit, W. Relationship between anthocyanins, proanthocyanidins, and cell wall polysaccharides in grape and red wines. A current state-of-art review. Critical Reviews in Food Science and Nutrition. 2022, 62, 7743–7759. [Google Scholar] [CrossRef]
- Boulet, J.; Vernhet, A.; Poncet-Legrand, C.; Cheynier, V.; Doco, T. Exploring the role of grape cell wall and yeast polysaccharides in the extraction and stabilization of anthocyanins and tannins in red wines. OENO One. 2024, 58, 7793. [Google Scholar]
- Espejo, F. Role of commercial enzymes in wine production: a critical review of recent research. Journal of the Food Science and Technology. 2020, 58, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.; Rossi, J. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. American Journal of Enology and Viticulture. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Lee, E.; Nomura, N.; Patil, B.; Yoo, K. Measurement of total phenolic content in wine using an automatic Folin-Ciocalteu assay method. International Journal of Food Science and Technology. 2014, 49, 2364–2372. [Google Scholar] [CrossRef]
- Somers, T.; Evans, M. Spectral evaluation of young red wines: Anthocyanin equilibria, total phenolics, free and molecular SO2, “chemical age”. Journal of the Science of Food and Agriculture. 1977, 28, 279–287. [Google Scholar] [CrossRef]
- Kafkas, N.; Kosar, M.; Oz, A.; Mitchell, A. Advanced analytical methods for phenolics in fruits. Journal of Food Quality. 2018, 1, 1–6. [Google Scholar] [CrossRef]
- Vrhovsek, U.; Mattivi, F.; Waterhouse, A. Analysis of red wine phenolics: Comparison of HPLC and spectrophotometric methods. Vitis. 2001, 40, 87–91. [Google Scholar]
- Labarde, B.; Cheynier, V.; Brossaud, F.; Souquet, J.M.; Moutounet, M. Quantitative fractionation of grape proanthocyanidins according to their degree of polymerization. Journal of Agricultural and Food Chemistry. 1999, 47, 2719–2723. [Google Scholar] [CrossRef]
- Souquet, J.M.; Labarde, B.; Le Guerneve, C.; Cheynie, V.; Moutounet, M. Phenolic composition of grape stems. Journal of Agricultural and Food Chemistry. 2000, 48, 1076–1080. [Google Scholar] [CrossRef]
- Hagerman, A.; Rice, M.; Richard, N. Mecchanism of protein precipitation for two tannins, pentagalloyl glucose and epicatechin16 (4→8) catechin (procyanidin). Journal of Agricultural and Food Chemistry. 1998, 46, 2590–2595. [Google Scholar] [CrossRef]
- Fragoso, S.; Acena, L.; Guasch, J.; Mesters, M.; Busto, O. Quantification of phenolic compouds during red winemaking using FT-MIR spectroscopy and PLS-regression. Journal of Agricultural and Food Chemistry. 2011, 59, 10795–10802. [Google Scholar] [CrossRef] [PubMed]
- Rolle, L.; Torchio, F.; Lorrain, B.; Giacosa, S.; Rio Segade, S.; Cagnasso, E.; Gerbi, V.; Teissedre, P. Rapis methods for the evaluation of total phenol content and extractability in intact grape seeds of Cabernet sauvignon: Instrumental mechanical properties and FT_NIR spectrum. Journal International des Science de la Vigne et du Vin. 2012, 46, 29–40. [Google Scholar]
- Garcia-Hernandez, C.; Salvo-Comino, C.; Martin-Pedrosa, F. ; Garcia-Cabezon, Rodriguez-Mendez M. Analysis of red wines using an electronic tongue and infrared spectroscopy. Correlations with phenolic content and color parameters. LWT-Food Science and Technology. 2020, 118, 108785. [Google Scholar]
Figure 1.
Structure and reactivity of phenolic compounds.
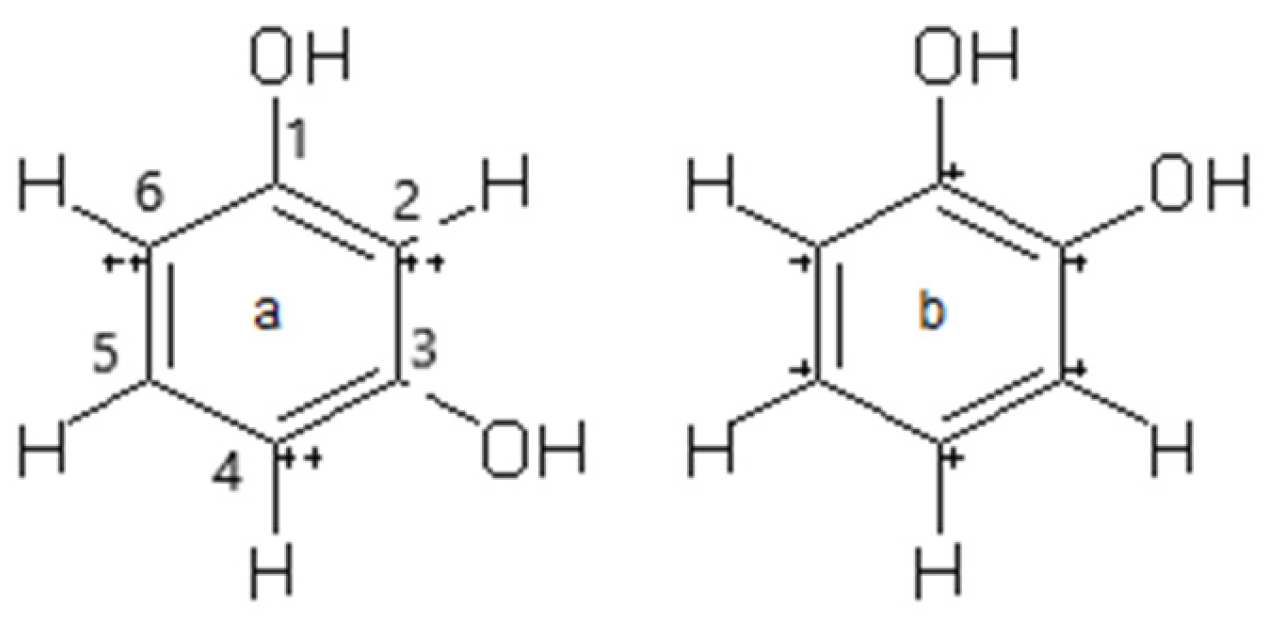
Figure 2.
Hydroxycinnamic esters.
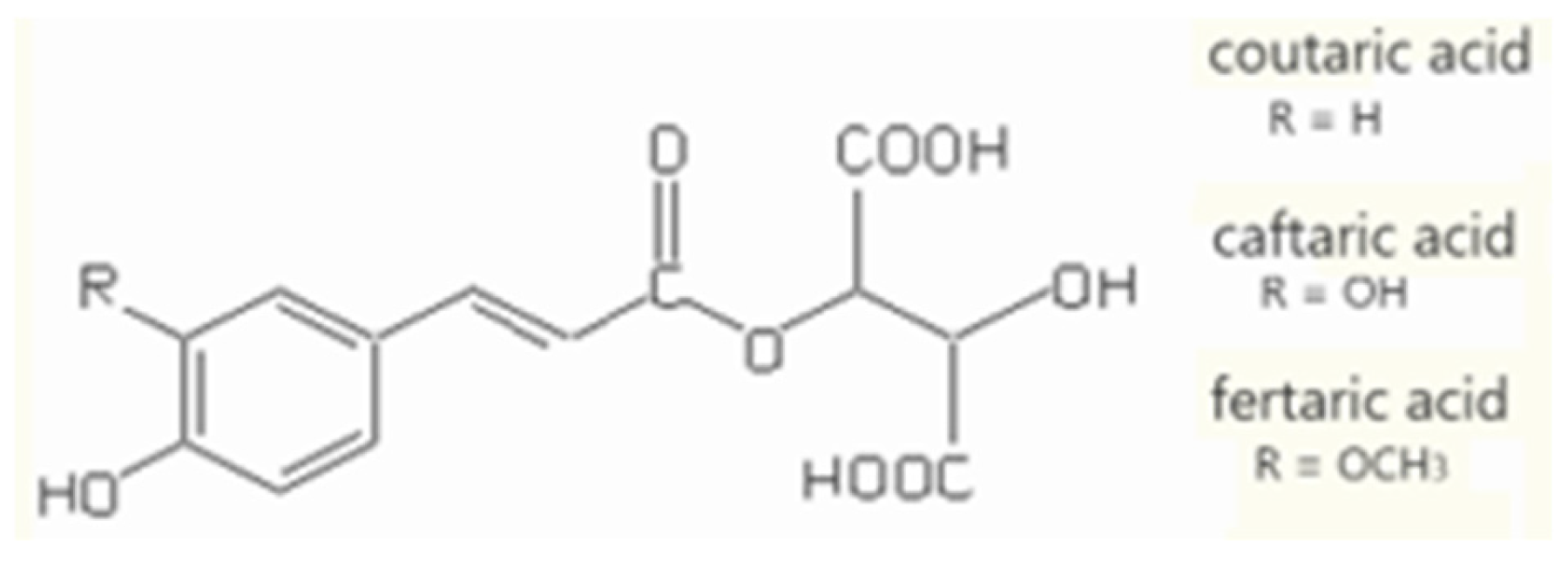
Figure 3.
Flavan-3-ols.
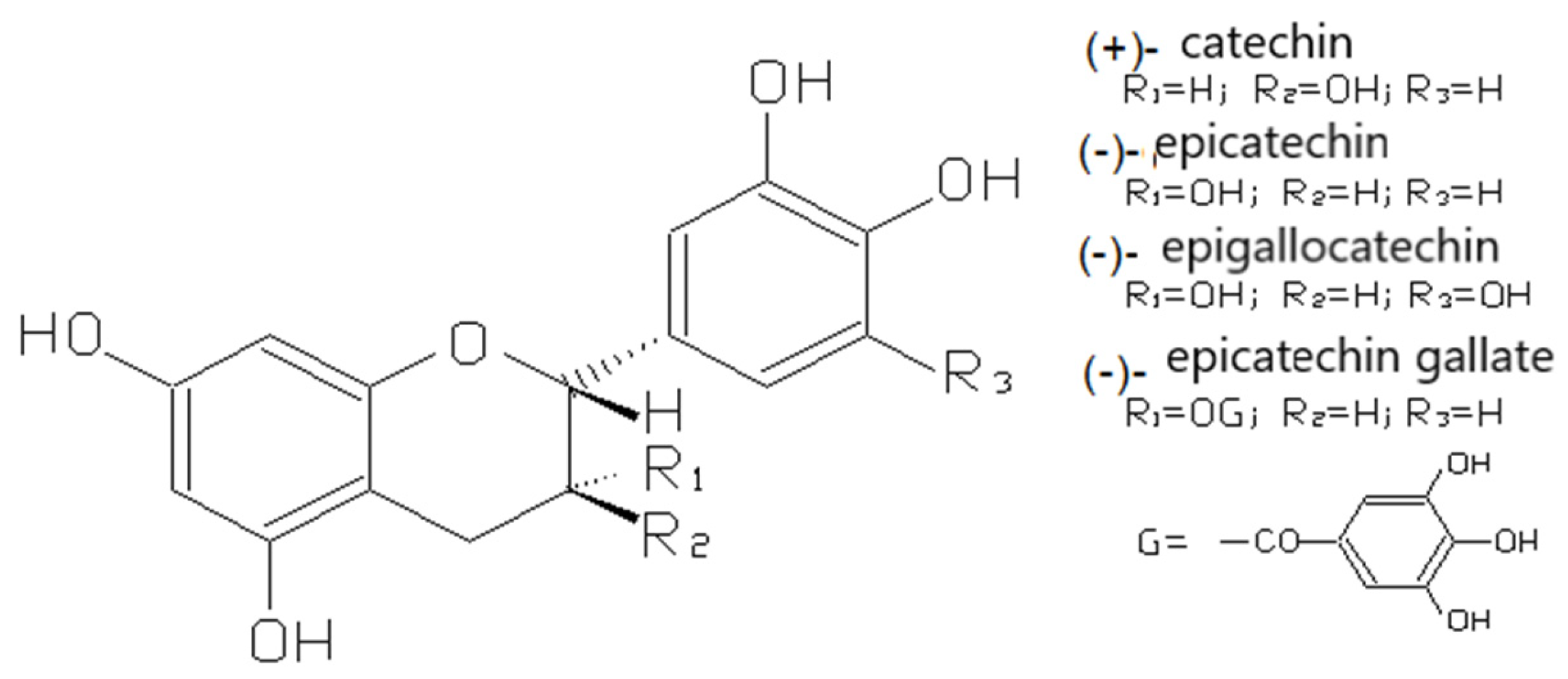
Figure 4.
Proanthocyanidins.
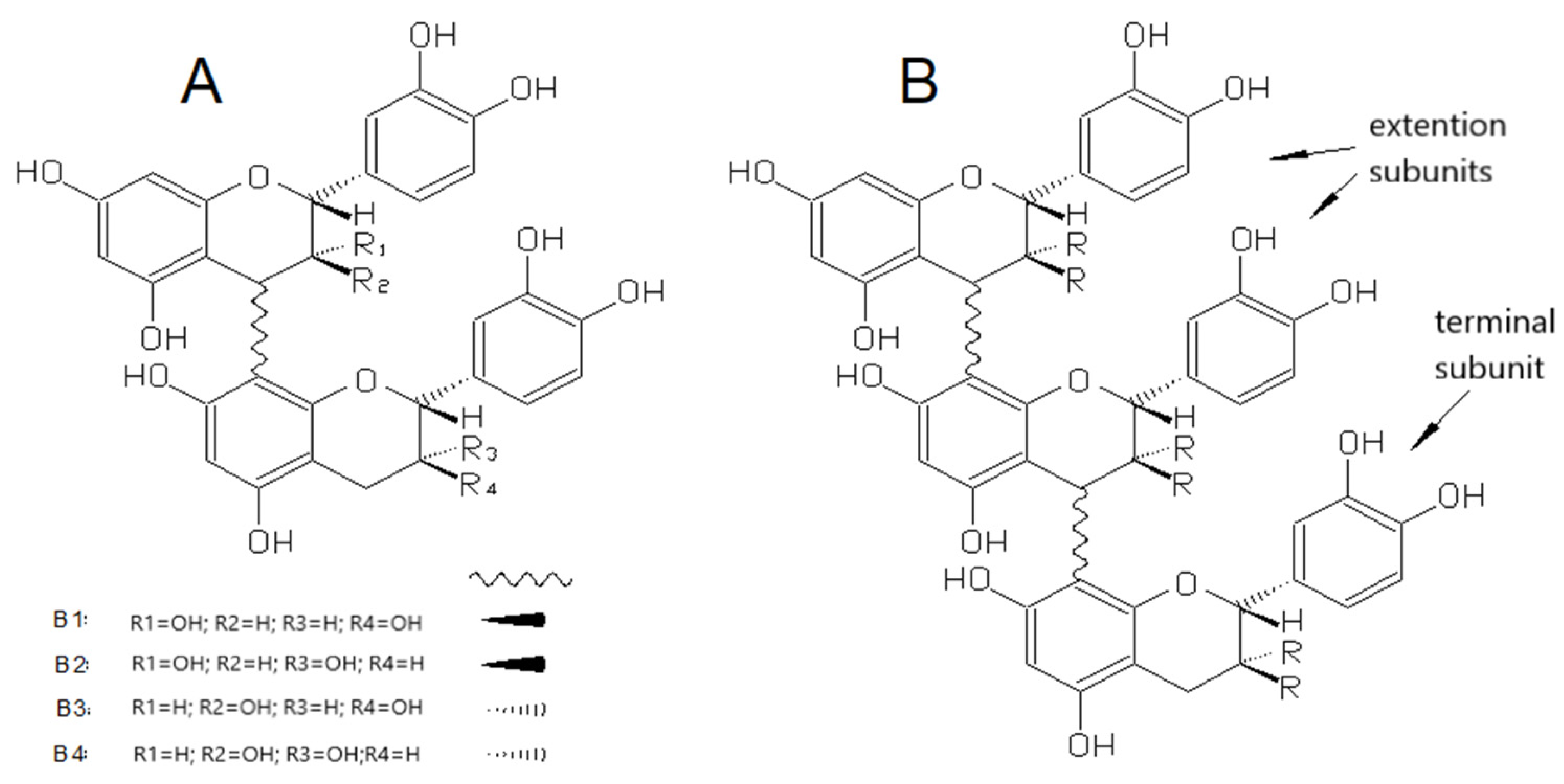
Figure 5.
Anthocyanins.
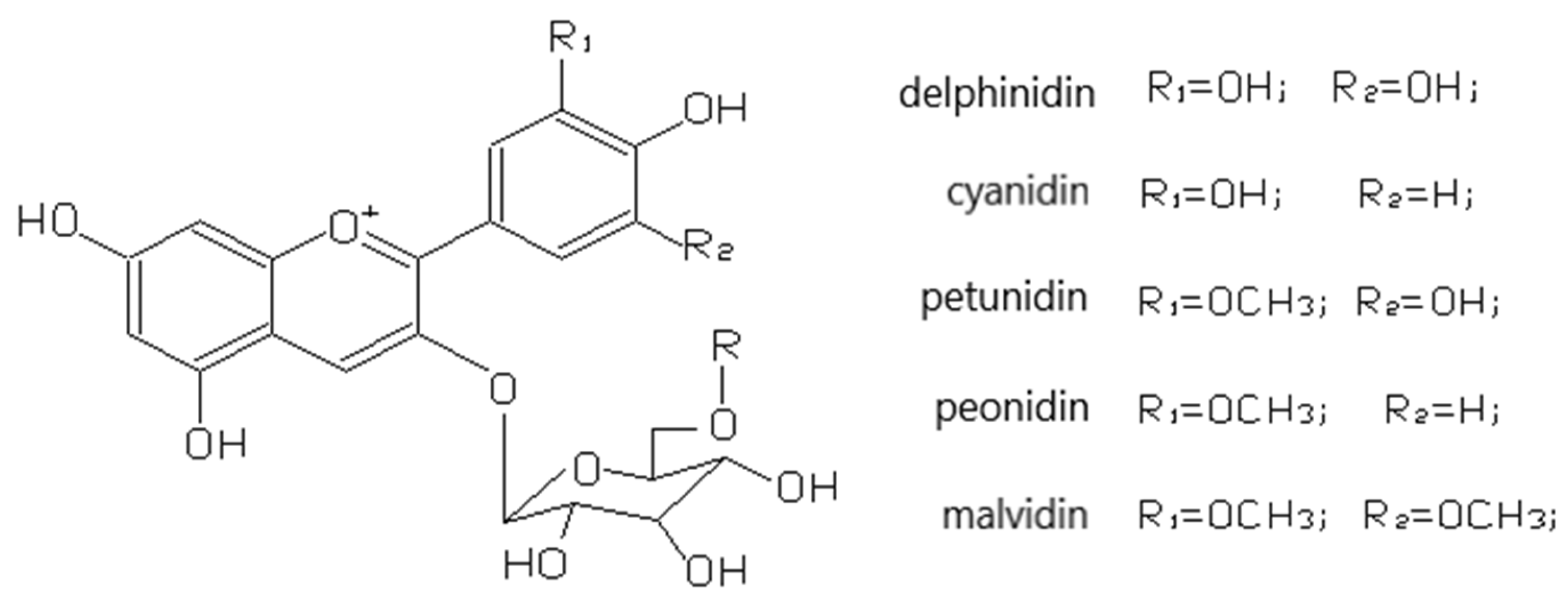
Figure 7.
Protein—Tannin model interaction [109].
Figure 7.
Protein—Tannin model interaction [109].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



