Submitted:
02 October 2025
Posted:
03 October 2025
You are already at the latest version
Abstract
Depression is a leading cause of global disability and is increasingly recognized as a multifactorial disorder characterized by fundamental disruptions in neuroplasticity, including diminished hippocampal neurogenesis, impaired synaptic plasticity, and dysregulated stress-response systems. Given the limited efficacy of conventional pharmacological treatments, lifestyle-based interventions—most notably physical exercise—have gained considerable attention for their antidepressant effects, partly mediated by secreted exerkines. Among these, adiponectin has emerged as a particularly compelling candidate linking metabolic regulation to neuroplasticity and mood. Recent evidence suggests that adiponectin contributes to the antidepressant effects of exercise by modulating hippocampal neurogenesis, neuroinflammation, and brain-derived neurotrophic factor (BDNF) signalling. Despite these advances, the mechanisms by which adiponectin influences depression remain incompletely understood. This review synthesizes current knowledge on adiponectin’s role in depression pathophysiology, with emphasis on its capacity to enhance neuroplasticity and hippocampal neurogenesis, and its potential to mediate exercise-induced antidepressant effects via defined molecular pathways. Building on these insights, we discuss adiponectin’s translational promise as both a predictive biomarker of treatment response and a novel therapeutic target. By integrating preclinical and clinical evidence, this review offers a comprehensive perspective on adiponectin’s involvement in depression while identifying critical gaps to guide future mechanistic research.
Keywords:
metabolism
; adiponectin
; neuroplasticity
; depression
; physical exercise
1. Introduction
Depression is one of the leading causes of global disability and arises from a complex interplay of biological and environmental factors that shape its multifaceted aetiology. Stressful life events and chronic stress are among the primary etiological drivers [1], capable of both initiating and sustaining depressive pathology. These factors do not act in isolation: stress and inflammation interact bidirectionally in a synergistic manner that amplifies their pathological impact [2], disrupting key brain functions such as neurogenesis—the generation of new neurons in the adult brain [3,4].
The neurogenic hypothesis of depression posits that impaired hippocampal neurogenesis represents a core mechanism underlying depressive symptomatology and contributes to the therapeutic efficacy of antidepressant treatments [5]. Supporting this view, clinical studies have shown that patients with depression undergoing antidepressant therapy exhibit increased hippocampal neural progenitor cell counts [6,7]. Yet, despite these advances, the precise mechanisms through which stress and inflammation converge to suppress neurogenesis—and how these processes collectively shape the neurobiology of depression—remain incompletely understood.
At the same time, the prevalence of psychiatric disorders, particularly depression, has risen steadily in recent years. This trend can partly be attributed to improved diagnostic accuracy and heightened social awareness, but it also reflects profound changes in lifestyle patterns [8]. Of particular concern is the global shift toward sedentary behaviours, driven by urbanization, technological advances, and changing occupational demands, which have progressively displaced incidental physical activities such as walking, standing, and manual tasks [9,10,11]. Physical inactivity may carry important neurobiological consequences: exercise has been shown to enhance hippocampal neurogenesis [12], lower systemic and central inflammation [13] and improve emotional regulation [14]. Accordingly, decreased daily movement is now considered a modifiable risk factor for depression and related mental health disorders [15]. Consistently, both longitudinal and cross-sectional studies demonstrate that individuals with depression engage in less physical activity and report higher sedentary behaviour compared to healthy controls [16,17,18], with lower physical activity closely linked to core depressive symptoms such as anhedonia, fatigue, and hopelessness, symptoms that directly impair motivation and energy, thereby reinforcing inactivity and creating a vicious cycle of worsening mood and reduced behavioural activation [19,20].
Efforts to understand the biological basis of exercise’s antidepressant effects have identified a wide range of molecular mediators, collectively termed exerkines. Among them, adiponectin has emerged as a particularly intriguing candidate linking physical activity to enhanced neurobiological resilience against depression [21]. Secreted predominantly by adipose tissue and elevated during exercise, adiponectin has been shown to directly influence the central nervous system (CNS) by promoting hippocampal neurogenesis [22] and modulating inflammatory pathways [23]. By intersecting metabolic regulation, brain plasticity, and inflammation—three key processes implicated in depression—this adipokine represents a pivotal mediator of exercise’s neuroprotective and antidepressant effects [24].
Despite increasing interest in adiponectin’s role, significant gaps remain in our mechanistic understanding of how it contributes to exercise-induced resilience against depression. The aim of this review is therefore to synthesize current evidence on adiponectin, clarifying its role as a biological link between physical activity and improved brain health outcomes. Specifically, we highlight its capacity to enhance neuroplasticity and hippocampal neurogenesis, discuss the molecular pathways through which it acts, and explore its potential as a therapeutic target. By integrating findings from preclinical, clinical, and experimental studies, this review provides a comprehensive perspective on adiponectin’s contribution to the neurobiology of depression, with particular emphasis on its mechanistic role in mediating the antidepressant effects of exercise.
2. Neuroplasticity Dysregulation in Depression
Neuroplasticity represents a fundamental property of the brain, encompassing its ability to adapt both structurally and functionally in response to internal and external stimuli [3]. This multifaceted term includes different processes, from neurogenesis [25] and synaptogenesis [26] to activity-dependent processes like long-term potentiation (LTP) [27], and synaptic remodelling [28]. Collectively, these mechanisms allow the brain to dynamically reshape neuronal circuits and to strengthen or weaken connections in accordance with experience and environmental demands [29,30].
The neuroplasticity hypothesis of depression, which has gained considerable empirical support, posits that impairments in these processes arise from a complex interplay between environmental stressors, such as chronic stress or early-life adversity, and intrinsic factors, including genetic vulnerability, which together compromise the brain’s adaptive capacity [31,32]. The resulting neuroplastic changes occur across multiple biological levels and are context dependent: while in some situations they may promote adaptive responses and resilience, in others—particularly when dysregulated—they contribute to the emergence and persistence of core depressive symptoms, including anhedonia, cognitive impairment, and negative affective states [1].
Structurally, neuroimaging and postmortem studies have consistently revealed alterations in brain regions involved in cognition and emotional regulation, most notably a reduction in hippocampal volume, which is widely regarded as a robust biomarker of depression severity, with the degree of atrophy correlating with the number of depressive episodes and illness chronicity [33]. Interestingly, neuronal loss in the hippocampus has been estimated at approximately 20–35%, while in other regions, such as the prefrontal cortex, volumetric and cellular alterations appear to be even more pronounced [34,35]. These morphological findings may reflect disruptions in adult hippocampal neurogenesis (AHN)—the process by which neural stem cells (NSCs) in the subgranular zone (SGZ) of the dentate gyrus (DG) differentiate into mature neurons, thereby supporting hippocampal plasticity and cognitive flexibility [36].
Impairments in AHN limit the brain’s ability to form new connections, adapt to external challenges, and recover from stress-related insults, with mounting evidence suggesting that both stress and depression markedly suppress this process, thereby contributing to the emotional and cognitive dysfunctions observed in patients [37]. For example, rodent studies show that corticosterone administration, a well-established model of depressive-like behaviour, reduces BrdU-positive hippocampal cells in the DG [38], and decreases BDNF levels, thereby impairing AHN through a neuronal autophagy-dependent mechanism, ultimately leading to depressive-like phenotypes [39]. Importantly, AHN is a dynamic and treatment-responsive process: antidepressants, electroconvulsive therapy [40], transcranial magnetic stimulation [41], and exercise [42] have all been shown to enhance hippocampal neurogenesis, underscoring its value as a therapeutic target.
Beyond neurogenesis, synaptic plasticity processes such as LTP and long-term depression (LTD) are also fundamental for hippocampal function, particularly in enabling memory formation, learning, and the integration of newly generated neurons into existing circuits [43] and in supporting the adaptive integration of new neurons into existing circuits [44,45]. LTP, a central mechanism of synaptic strengthening, is triggered by high-frequency stimulation and relies on the coordinated activity of AMPA and NMDA glutamate receptors. While AMPA receptors facilitate sodium influx and depolarization of the postsynaptic membrane, NMDA receptors—upon relief of the magnesium block—permit calcium entry, which in turn activates intracellular signalling cascades involving kinases such as CaMKII and PKC. These cascades promote AMPA receptor phosphorylation and trafficking to the postsynaptic membrane, thereby strengthening synaptic efficacy [46]. Conversely, LTD provides an essential counterbalance by selectively weakening synaptic connections through AMPA receptor internalization, a process triggered by modest NMDA-mediated calcium influx and subsequent activation of protein phosphatases [47]. This bidirectional plasticity maintains network stability, prevents synaptic saturation, and supports adaptive circuit refinement [48].
In depression and chronic stress, however, these mechanisms are significantly disrupted, with evidence from both preclinical and clinical studies pointing to reduced LTP alongside enhanced LTD, particularly within hippocampal circuits [49,50,51]. Such an imbalance contributes to impaired communication between neurons, ultimately manifesting as deficits in memory consolidation, attention, and executive functioning, as well as in motivational processes tied to reward-related circuitry [45,52,53]. Specifically, reductions in LTP have been linked to the cognitive dysfunction frequently observed in depression [54] while excessive LTD, especially within mesocorticolimbic pathways, may underlie anhedonia and reduced motivation.
The persistence of synaptic strengthening depends not only on early-phase LTP but also on late-phase LTP (L-LTP), which requires the activation of transcriptional and translational pathways such as PKA and MAPK. These cascades enable the synthesis of proteins necessary for structural synaptic modifications, including dendritic spine growth and synaptogenesis, thereby supporting long-term memory consolidation and circuit stabilization [55,56]. Disruptions in these processes impair the maintenance of long-lasting synaptic modifications, leading to reduced brain adaptability and compromised recovery during depression [33], with impairments in L-LTP-related signalling pathways linked not only to cognitive deficits observed in depression but also to the severity of depressive symptoms [54]. A central contributor to these impairments is the disruption of the BDNF - Tropomyosin receptor kinase B (TrkB) signalling pathway: under normal conditions BDNF binds to its receptor TrkB, activating signalling cascades such as MAPK, phospholipase C (PLC) and phosphatidylinositol-3 kinase (PI3K), along with its downstream effector mammalian target of rapamycin (mTOR). This activation directly promotes CREB phosphorylation, which in turn enhances proliferation and differentiation of neural progenitor cells [57].
BDNF signalling is one of the most crucial pathways, not only in promoting synaptic protein translation and dendritic spine growth, but also in modulating neurogenesis [1,58], in particular in the hippocampus [1]. In patients affected by depressive symptoms as well as in the context of exposure to chronic stress, plenty of evidence indicate that BDNF signalling is profoundly dysregulated, which in turn compromise synaptic plasticity and destabilize of neuronal circuits [39,44,59]. Recent meta-analyses and systematic reviews confirm that BDNF levels are significantly lower, both in the blood and in the CNS, in patients with depression compared to controls; moreover, lower peripheral BDNF levels have been associated with more severe symptoms [59,60,61,62,63]. Crucially, rather than being a static vulnerability factor or trait marker, BDNF is considered as a dynamic "state marker" of depression, reflecting current disease activity and treatment responsiveness, as consistently supported by evidence showing that interventions which effectively reduce depressive symptoms often lead to increased BDNF levels [61,62].
Odaira and colleagues (2019) [64] demonstrated that activation of AMP-activated protein kinase (AMPK) promotes hippocampal neurogenesis in a manner comparable to the effects of antidepressants such as selective serotonin reuptake inhibitors (SSRIs). Conversely, loss of AMPK function impairs CREB phosphorylation, resulting in reduced BDNF expression and diminished cell viability, thereby underscoring the pivotal role of this kinase in sustaining neurogenic processes [65].
Emerging evidence suggests that inflammation may exacerbate disruptions in neurotrophic signalling pathways, thereby contributing to the synaptic and structural abnormalities observed in depression. Particularly in the prefrontal cortex and hippocampus, glutamate imbalance is exacerbated by elevated proinflammatory cytokine levels and impaired astrocyte function, which raise excitotoxicity and excessive synaptic pruning [66,67,68,69]. In addition, microglial cells, crucial for their immune role, become activated after chronic stress, releasing pro-inflammatory cytokines [70]. Together, these maladaptive mechanisms reinforce dysfunctional neural circuits and contribute to the emotional and cognitive symptoms of depression by affecting the ability of the brain to adapt to environmental stimuli.
Taken together, these findings underscore that neuroplasticity disruptions—involving neurogenesis, synaptic plasticity, BDNF signalling, energy-sensing pathways, and inflammation—undermine the functional integrity of neural circuits central to cognition, emotional regulation, and stress resilience. Such alterations not only constitute core mechanisms of depression but also correlate with illness severity and chronicity, thereby limiting therapeutic responsiveness and highlighting neuroplasticity as a crucial target for novel interventions [71].
3. Adiponectin: From Peripheral Hormone to Brain Modulator
Adiponectin, an adipokine that plays a pivotal role in sustaining systemic metabolic homeostasis, represents one of the most abundant peptide hormones secreted by white adipose tissue (WAT), particularly in response to hormonal cues and nutritional status [72], both of which exert a profound influence on its production and secretion. Lifestyle interventions such as caloric restriction and physical activity have been shown to increase circulating levels of adiponectin by 18–48%, whereas obesity, and especially the accumulation of visceral adiposity, markedly suppresses its expression through a variety of interrelated mechanisms [73]. At the cellular level, the secretory capacity of adipocytes depends on their maturation state and is tightly regulated by microenvironmental factors such as hypoxia, nutrient availability, and the prevailing inflammatory milieu [74]. The biological activity of adiponectin is further dictated by its multimeric organization, with high-molecular-weight (HMW) isoforms displaying superior metabolic efficacy, primarily by enhancing insulin sensitivity and conferring cardiovascular protection [75]. Nevertheless, these bioactive isoforms remain particularly vulnerable to disruption under conditions of endoplasmic reticulum (ER) stress and oxidative stress, both of which are accentuated in states of metabolic dysfunction [76,77,78].
Functionally, adiponectin exerts its pleiotropic effects through two distinct receptors (AdipoRs), which, despite their structural similarities, display tissue-specific expression patterns and preferentially activate distinct signalling pathways. AdipoR1 primarily engages AMPK to regulate energy homeostasis and mitochondrial function, whereas AdipoR2 predominantly signals through peroxisome proliferator-activated receptor alpha (PPARα), modulating lipid metabolism and exerting anti-inflammatory effects [79]. These complementary pathways converge to produce insulin-sensitizing and anti-inflammatory effects in key metabolic organs: in the liver, adiponectin suppresses gluconeogenesis while promoting fatty acid oxidation [80,81], whereas in skeletal muscle, it enhances glucose uptake and β-oxidation [82]. Notably, these metabolic benefits are further potentiated by adiponectin’s immunomodulatory capacity, which attenuates the production of pro-inflammatory cytokines while simultaneously enhancing anti-inflammatory mediators such as IL-10 [83,84,85,86]. From a pharmacological perspective, the adiponectin pathway demonstrates substantial therapeutic potential, as exemplified by thiazolidinediones (TZDs), PPARγ agonists widely used in the management of type 2 diabetes. TZDs increase circulating adiponectin levels by promoting both its transcription and secretion, thereby improving insulin sensitivity and alleviating metabolic dysfunction [87,88].
Endocrine regulation further modulates adiponectin dynamics, with oestrogens shown to upregulate its expression, which may partly explain the higher plasma concentrations typically observed in females [89]. Conversely, chronic low-grade inflammation, characterized by elevated pro-inflammatory cytokines such as tumour necrosis factor alpha (TNF-α) and interleukin-6 (IL-6), suppresses adiponectin gene expression via activation of nuclear factor kB (NF-κB), thereby exacerbating metabolic dysfunction and promoting insulin resistance [84,85].
Beyond its well-established role in metabolic regulation, adiponectin has also emerged as a critical player in neuropsychiatric disorders. Unlike many peripheral metabolic hormones, adiponectin can cross the blood-brain barrier (BBB) and exert direct effects on CNS, particularly within the hippocampus [90,91], a capability confirmed by its detection in the cerebrospinal fluid (CSF) of both rodents [38] and humans [92], although its concentration in human CSF is approximately 1,000-fold lower than in serum [93]. These findings support the notion that adiponectin functions as a key molecular messenger, linking peripheral metabolic cues to central neurobiological adaptations. In the brain, AdipoRs are widely expressed in regions critical for mood regulation, neuroplasticity, and energy homeostasis, including the hippocampus, cerebral cortex, hypothalamus, pituitary gland, and brainstem [94]. Rodent studies examining hippocampal synaptosomes indicate that AdipoR1 is present in both presynaptic and postsynaptic compartments, whereas AdipoR2 is predominantly presynaptic, suggesting potential roles for these receptors in synaptic function, learning, and memory [95]. Moreover, AdipoRs are also expressed in glial cells such as astrocytes and microglia, indicating that adiponectin may influence neuroinflammatory and immune mechanisms relevant to depression [94].
Clinical evidence demonstrates an inverse relationship between circulating adiponectin levels and depression, with lower concentrations associated not only with disease presence but also with greater symptom severity, suggesting a potential causal role for this adipokine in depressive pathophysiology [96,97,98,99]. This association is further supported by longitudinal studies in patients treated with interferon alpha (IFN-α) for hepatitis C virus (HCV), in which the emergence of depressive symptoms during treatment correlated with progressively rising adiponectin levels, likely representing a compensatory response to treatment-induced inflammation, mediated by the hormone’s anti-inflammatory properties [100].
Preclinical studies provide mechanistic insight into adiponectin’s neurobehavioral effects, demonstrating its capacity to modulate anxiety-like behaviours in specific brain regions [101]. Disruption of adiponectin signalling in the CNS has been associated with neurobehavioral dysfunctions. For example, our recent study [102] showed that prenatal stress reduced AdipoR1 and AdipoR2 expression in both liver and ventral hippocampus, coinciding with social deficits and elevated inflammation. Importantly, adiponectin exerts potent anti-inflammatory effects within the CNS by downregulating microglial activation and suppressing pro-inflammatory cytokines, including TNF-α and IL-6, thereby creating a more favourable environment that facilitates synaptic plasticity and cognitive performance [103,104,105].
In addition to its link with depressive and anxiety symptoms, adiponectin may also serve as a predictor of pharmacological treatment response [106]. Low baseline plasma adiponectin levels have been associated with a rapid clinical response to ketamine, with lower initial concentrations correlating with greater treatment efficacy [107]. Similarly, in treatment-resistant depression, clinical improvements following interventions such as electroconvulsive therapy are accompanied by increases in adiponectin levels. Moreover, baseline adiponectin concentrations have been shown to correlate with BDNF levels, suggesting that both biomarkers may dynamically reflect depressive state. Collectively, these findings indicate that adiponectin and BDNF fluctuate together in relation to symptom severity, highlighting the central role of neuroplasticity in depression [108], as will be further discussed in the following section.
4. Linking Exercise, Adiponectin, and Neuroplasticity
4.1. How Exercise Enhances Adiponectin Levels: Cellular and Molecular Mechanisms
When the body undergoes physical exercise, a cascade of cellular and molecular changes is initiated, establishing exercise as one of the most effective non-pharmacological strategies for promoting brain health [21]. Among the myriad bioactive compounds released in response to physical activity, collectively termed exerkines [109], adiponectin has garnered particular attention due to its central role at the intersection of metabolic regulation, neuroplasticity, and mood modulation [110].
Exercise, particularly aerobic and endurance training, consistently elevates circulating adiponectin levels, especially following long-term interventions [111]. However, findings regarding the effects of acute exercise on adiponectin are less consistent: some studies report modest but significant increases in total adiponectin shortly after intense aerobic sessions [112,113,114], while the majority report no change or even slight decreases [113,115,116]. Preclinical studies have also demonstrated that exercise can lead to a four-fold increase in adiponectin levels in mice subjected to chronic corticosterone treatment [38].
Interestingly, adiponectin behaves differently from many other adipokines, with its circulating levels rising in response to physical activity despite being inversely correlated with body fat [110]. While part of this exercise-induced increase may be mediated by weight loss, elevated adiponectin levels have been observed following physical activity in healthy individuals, people with obesity, and in various rodent models, suggesting that exercise can modulate adiponectin levels independently of changes in adiposity [117].
The precise mechanisms through which exercise enhances adiponectin secretion remain incompletely understood, although several hypotheses have emerged from both rodent and human studies [103,118,119]. Central to these mechanisms is adipose tissue, comprising brown adipose tissue (BAT), which is limited in distribution and primarily involved in thermogenesis [120], and WAT, which serves both as an energy reservoir and as an active endocrine organ.
As illustrated in Figure 1, during exercise, increased energy demand triggers lipolysis in WAT, mobilizing stored triacylglycerols to provide fatty acids to working muscles and other tissues [121]. This activation of WAT is accompanied by functional adaptations, including enhanced mitochondrial activity and increased glucose uptake through the translocation of glucose transporter type 4 (GLUT4) [122], which together contribute to improved systemic insulin sensitivity and energy homeostasis [123]. Consequently, circulating insulin levels decrease, fostering a more insulin-sensitive and less inflammatory microenvironment within adipose tissue, thereby favouring the expression of the ADIPOQ gene that encodes adiponectin [124,125]. Moreover, insulin itself directly stimulates ADIPOQ expression, particularly in insulin-sensitive individuals, as demonstrated by increased ADIPOQ mRNA following acute insulin administration in subcutaneous adipose tissue [126]. Thus, exercise-induced improvements in insulin sensitivity substantially contribute to adiponectin upregulation.
Within adipocytes, ADIPOQ transcription is regulated by pathways involving PPARγ and AMPK, both of which are activated by physical activity [127,128,129] and by reductions in pro-inflammatory signalling [130]. PPARγ, a nuclear receptor essential for adipocyte differentiation and lipid metabolism, acts as a principal transcriptional regulator of ADIPOQ [131]. Upon binding free fatty acids released by lipolysis [132], PPARγ heterodimerizes with the retinoid X receptor (RXR) and binds to PPAR response elements (PPREs) in the ADIPOQ promoter, robustly enhancing its transcription [133]. Pharmacological activation of PPARγ has been shown to increase circulating adiponectin, particularly the metabolically protective HMW isoform, in both mice and humans, through both direct upregulation of ADIPOQ transcription and enhanced expression of molecular chaperones necessary for adiponectin multimerization and secretion [131,134,135]. Animal studies further corroborate this link, demonstrating that PPARγ upregulation following weight loss correlates with both increased adiponectin and improved insulin sensitivity [136].
Simultaneously, AMPK, a key cellular energy sensor, is activated during exercise in response to an increased AMP/ATP ratio driven by heightened energy demand and rapid ATP consumption required for muscle contraction and other cellular activities [137]. Once activated, AMPK enhances PPARγ activity and may also upregulate the expression of PPARG, the gene encoding PPARγ [138], thereby further stimulating adiponectin production. This crosstalk between AMPK and PPARγ establishes a mechanistic link between exercise and adiponectin upregulation [103].
Collectively, these coordinated pathways promote ADIPOQ transcription, secretion, and multimerization, contributing to improved neuroplasticity, enhanced metabolic adaptation, and alleviation of depressive symptoms [139,140]. Notably, adiponectin itself can activate AMPK within the CNS, leading to further beneficial effects on neuronal function, which will be discussed in the following section.
4.2. Molecular Mechanisms Underlying Adiponectin’s Effects on Neurogenesis and Neuroplasticity
As previously discussed, numerous studies have identified adiponectin as a key molecular mediator linking physical exercise to neurogenesis and mood regulation [38,91]. For instance, in a diabetic mouse model, voluntary running failed to restore hippocampal neurogenesis in the absence of adiponectin [12], while reduced serum adiponectin levels have been associated with impaired hippocampal neurogenesis [141], highlighting its essential role in mediating exercise-induced neurogenic effects and its broader relevance beyond metabolic regulation. The involvement of adiponectin signalling is further supported by evidence that pharmacological activation via AdipoRon, a selective AdipoRs agonist, alleviates depression- and anxiety-like behaviours and enhances hippocampal plasticity [26,142]. Notably, in vivo studies demonstrate that voluntary running increases hippocampal cell proliferation and neurogenesis only in wild-type mice, whereas adiponectin knockout mice fail to exhibit these antidepressant-like effects. Similarly, in vitro exposure of NSCs to AdipoRon for 48 hours activates key signalling pathways involved in neurogenesis and synaptic plasticity [143], confirming that adiponectin is necessary for both mood-enhancing and neuroplastic benefits [91].
Collectively, these findings position adiponectin as a central player in translating physical activity into cognitive and emotional resilience through mechanisms that converge on hippocampal neurogenesis and plasticity. The question then arises: through which specific molecular pathways are these effects mediated? Mechanistically, adiponectin engages multiple signalling cascades that converge on hippocampal neurogenesis and synaptic plasticity, as illustrated in Figure 2. Upon activation of AdipoR1 and AdipoR2, downstream pathways include (i) metabolic sensors such as the AMPK cascade and its downstream kinases, (ii) survival and anti-apoptotic signalling, (iii) developmental and stem cell fate regulators such as Notch, (iv) neurotrophic factor pathways including STAT3/BDNF, and (v) anti-inflammatory signalling via suppression of NF-κB. Together, these interconnected networks translate metabolic and exercise-related signals into structural and functional adaptations in the hippocampus.
Among these pathways, the AdipoR1-AMPK axis is the most extensively characterized, functioning as a central metabolic sensor linking energy status to neuronal adaptation. Exercise promotes hippocampal plasticity, protects neurons from stress-induced atrophy, enhances dendritic remodelling, and alleviates depressive-like behaviours in wild-type mice via AMPK phosphorylation. In contrast, adiponectin knockout mice fail to activate this pathway, underscoring the critical role of AdipoR1-AMPK signalling in mediating exercise-induced neuronal growth and energy metabolism [24].
Further insights into AMPK’s role in NSC regulation come from in vitro studies. B. Liu et al. demonstrated that AdipoRon treatment significantly increases phosphorylated AMPK expression in NSCs, whereas co-administration of Compound C, a selective AMPK inhibitor, abolishes this effect. AdipoRon also activates the transcription factor CREB, whose phosphorylation is suppressed by Compound C, indicating that adiponectin promotes NSC proliferation via the AMPK/CREB axis [143]. Additional evidence highlights the involvement of AMPK downstream kinases in sustaining neurogenic actions: Whitaker and Cook (2021) [144] reported that adiponectin stimulates adult human NSC proliferation via AMPK-mediated activation of p38MAPK, a kinase with both proliferative and anti-proliferative roles. Inhibition of p38MAPK significantly attenuates adiponectin-induced proliferation, without affecting basal cell growth, while phosphorylation of GSK-3β induced by adiponectin is dependent on p38MAPK activity, enhancing transcription of neurogenesis-related genes [145]. These findings reinforce the AdipoR1-AMPK axis as a critical metabolic translator, converting physical effort into neurogenic benefits.
Adiponectin also promotes NSC survival and anti-apoptotic signalling. Under stressful conditions, such as high glucose exposure, adiponectin downregulates apoptotic regulators including p21, p53, and c-Myc, while upregulating neurogenic factors, thereby shifting the balance from cell death toward regeneration [146]. Blockade of AdipoR1 abolishes this protective effect, resulting in increased apoptosis, confirming that receptor engagement is essential for maintaining NSC viability under metabolic stress.
In addition to survival signalling, adiponectin modulates developmental regulators of stem cell fate, with the Notch pathway emerging as a key mediator [147]. Early studies demonstrated that a homolog of adiponectin upregulates ADAM10 and ADAM17, metalloproteinases essential for Notch receptor activation, in the hippocampus [148]. Subsequent research showed that physical exercise increases expression of Notch1/2 receptors, ADAM10, and downstream targets in wild-type mice, whereas this effect is absent in adiponectin knockout mice, indicating that adiponectin is necessary for exercise-induced Notch activation [142]. This activation is further regulated via PPARα signalling, emphasizing the role of the adiponectin-Notch axis in mediating neurogenesis, cognitive function, and the protective effects of exercise against chronic stress-induced impairments in hippocampal plasticity.
Beyond neurogenesis and survival, adiponectin influences broader neuroplasticity processes, modulating synaptic structure and hippocampal circuit remodelling. Direct brain infusion of adiponectin increases dendritic spine density and arborization in hippocampal granule neurons, key indicators of synaptic efficacy [22]. Bloemer et al. (2019) [95] demonstrated that AdipoRs signalling in hippocampal synapses is reduced in adiponectin knockout mice, resulting in deficits in memory and cognitive tasks. In these mice, reductions in LTP correlate with decreased glutamatergic receptor subunits in the hippocampus, supporting the role of adiponectin in synaptic strengthening. Similarly, Pousti et al. (2018) [104] showed that adiponectin enhances presynaptic release probability and induces chemical LTP in the dentate gyrus, further confirming its role in synaptic plasticity.
Additional pathways contribute to adiponectin’s neuroprotective effects, most notably the BDNF pathway, which is critical for neuronal survival, synaptic plasticity, and hippocampal function [149]. While exercise independently enhances BDNF signalling [11], adiponectin may facilitate this effect via STAT3-mediated upregulation of BDNF in astrocytes. Inhibition of either STAT3 or BDNF abolishes the neural and behavioural benefits, highlighting the essential role of the adiponectin-STAT3-BDNF axis in mediating neuroplasticity [150]. In corticosterone-treated depressive-like mice, AdipoRon restores hippocampal expression of neurotrophic genes, including BDNF, VEGFα, IGF1, and NGF, while activating AMPK signalling, emphasizing its targeted role in restoring stress-induced impairments rather than broadly enhancing growth [151].
Inflammation represents another critical dimension in adiponectin-mediated neuroplasticity. Physical exercise reverses stress-induced changes in hippocampal microglia and neuroinflammation, exerting antidepressant effects [70]. Voluntary running promotes an anti-inflammatory microglial phenotype, suppressing neuroinflammation in the hippocampus, and these effects are dependent on the adiponectin/AdipoR1 pathway [103].
According to Chabry et al. (2015) [152], adiponectin levels increase following voluntary exercise, promoting a microglial anti-inflammatory state that reduces neuroinflammation as well as anxiety- and depressive-like behaviours. A subsequent study from the same group further demonstrated that this effect involves AMPK phosphorylation, which is downregulated by pro-inflammatory triggers such as lipopolysaccharide (LPS) and upregulated by adiponectin in microglial cells. Moreover, adiponectin activates AMPK and suppresses NF-κB signalling, thereby reducing the synthesis of pro-inflammatory cytokines and chemokines [23].
To complement these findings, pharmacological inhibition through specific AdipoRs antagonists has been employed to dissect the contribution of adiponectin signalling to hippocampal function. This bidirectional approach—demonstrating both the beneficial effects of receptor activation and the detrimental consequences of its blockade—provides strong evidence for a causal role of adiponectin in mediating the neuroplastic and cognitive benefits of exercise. For example, administration of the AdipoRs antagonist ADP400 to mice undergoing physical training reduces synaptic protein expression and abolishes exercise-induced memory improvements, while anxiety-related behaviors remain unaffected [26]. Independent studies further support the essential role of adiponectin signalling: peripheral inhibition of AdipoRs via ADP400 leads to a broad spectrum of deleterious effects, including increased β-amyloid accumulation in the hippocampus in Alzheimer’s disease models, heightened anxiety-like behaviour, impaired cognitive performance, and disrupted hippocampal neurogenesis [153].
Taken together, these findings highlight that beyond its established role in metabolic regulation, adiponectin is a central mediator of neuroplastic processes. By translating external beneficial stimuli, such as physical activity, into enhanced hippocampal plasticity, resilience to stress, and improved cognitive and emotional function, adiponectin orchestrates multiple intersecting signalling pathways—including AMPK, BDNF, and Notch. These pathways do not act in isolation; rather, they dynamically interact to stimulate neurogenesis while safeguarding neuronal integrity against stress and inflammatory damage. This sophisticated profile positions adiponectin as a promising molecular bridge between lifestyle factors and brain health, with potential therapeutic relevance for neuropsychiatric disorders.
5. Conclusions and Future Directions
In this review, we have highlighted adiponectin as a pivotal hormone at the crossroads between metabolic health, brain plasticity, and emotional regulation—domains that are profoundly disrupted in depression. Both preclinical and clinical evidence indicates that, upon crossing the BBB, adiponectin can modulate synaptic plasticity, neurogenesis, and neuronal survival, processes central to the pathophysiology of depressive disorders. These neuroprotective effects are mediated through multiple pathways, most notably AMPK, PPAR-α, and BDNF signalling, alongside Notch activation and the upregulation of anti-apoptotic markers.
The rationale of this review was to examine the mechanisms by which exercise-induced adiponectin exerts its beneficial effects on the brain, particularly in counteracting the neural dysfunctions associated with depression. What emerges is a molecule with considerable translational promise, meriting deeper exploration.
First, its potential as a biomarker for both depression severity and treatment response has gained increasing empirical support. For example, Machado-Vieira et al. (2017) [107] reported that systemic adiponectin abnormalities predicted a favourable response to ketamine in patients with major depressive disorder, a finding echoed in studies of SSRI- and SNRI-treated individuals [154]. In the context of rising treatment-resistant depression and the modest efficacy of conventional antidepressants, interest is growing in alternative or complementary strategies that target the biological underpinnings of mood disorders. Because circulating adiponectin is responsive to lifestyle modifications [73], it could serve as a useful stratification tool to identify patients most likely to benefit from interventions such as structured exercise. Physical activity not only elevates adiponectin levels but also enhances mood and neuroplasticity [110], raising the prospect of adiponectin as a dynamic biomarker to monitor efficacy in exercise-based treatments. Similarly, pharmacological approaches that enhance adiponectin signalling, such as AdipoRon, a synthetic AdipoR agonist [151], offer the possibility of mimicking the neurogenic and mood-regulating benefits of exercise in patients unable to engage in regular physical activity. Dietary strategies, including adherence to the Mediterranean diet, have also been linked to higher adiponectin levels and lower depressive symptomatology [155,156], further supporting its role as a potential biomarker of response. Monitoring adiponectin in these contexts may be particularly valuable in depression subtypes with metabolic comorbidities, such as insulin resistance or obesity, where identifying a distinct “metabotype” could guide more personalized and integrative therapeutic approaches.
Second, while preclinical data strongly support the neuroprotective and pro-neurogenic actions of adiponectin, substantial translational research is still required to determine how best to harness these pathways clinically. Future mechanistic studies should focus on AdipoR function in brain regions central to mood regulation, such as the hippocampus and prefrontal cortex [101,102] and explore how adiponectin signalling intersects with other neurotrophic and inflammatory networks to identify new drug targets. Clinically, randomized trials are needed to test whether interventions that elevate adiponectin—through exercise, diet, or pharmacological AdipoR agonists [108] can improve depressive symptoms.
Given the higher baseline adiponectin levels typically observed in women [89] and its possible role in stress resilience, it will also be important to examine sex-specific responses and whether adiponectin modulation benefits particular subgroups, such as patients with metabolic syndrome or atypical depression.
Metformin, an insulin sensitizer widely used in type 2 diabetes, provides a useful pharmacological model in this regard. Beyond its metabolic effects, Metformin increases adiponectin levels [157] and exerts neuroprotective actions via AMPK activation, with evidence of reversing depressive symptoms and normalizing Hypothalamic–Pituitary–Adrenal Axis dysfunction [158]. However, its limited selectivity for AdipoRs underscores the need for more targeted strategies. Nonetheless, Metformin illustrates the therapeutic potential of leveraging adiponectin pathways in developing next-generation treatments for depression and its metabolic comorbidities.
Beyond adiponectin, another promising avenue of research lies in glucagon-like peptide-1 receptor agonists (GLP1RA), which are widely used in the treatment of type 2 diabetes and obesity. Emerging evidence suggests that GLP1RA exert central effects that extend well beyond glycaemic control, including modulation of neuroinflammation, enhancement of neurogenesis, and promotion of synaptic plasticity [159,160]. Preclinical studies demonstrate that GLP1R activation enhances hippocampal BDNF expression and reduces microglial activation, thereby attenuating stress-induced structural and functional impairments in brain regions critical for mood regulation [161]. Clinical observations also support a potential antidepressant role, with GLP1RA treatment associated with improvements in mood, cognitive function, and overall quality of life in patients with metabolic syndrome or diabetes, populations characterized by high rates of comorbid depression [162]. In this context, Semaglutide, a GLP-1RA, represent a mechanistically grounded pharmacological option. Widely used for diabetes and obesity, GLP-1RAs improve glucose and lipid metabolism, reduce systemic inflammation, and modulate central neuroenergetic regulation [163,164,165]. Emerging evidence, including a recent meta-analysis, suggests that GLP-1RAs may also improve depressive symptoms independently of weight loss [162]. Moreover, a recent study in patients with mild depressive symptoms demonstrated that Semaglutide is safe in individuals with MDD, led to clinically significant weight loss, and enhanced global cognition [166]. However, no randomized controlled trial to date has specifically evaluated its efficacy in mood disorder populations stratified by immune metabolic dysfunctions. Interestingly, GLP1RA may interact with adiponectin signalling, as both pathways converge on AMPK activation and inflammatory modulation, suggesting potential synergistic effects in promoting neuroplasticity and emotional resilience. Exploring this intersection could open new therapeutic opportunities, where GLP1R agonists not only address metabolic dysfunction but also improve depressive symptoms, representing another promising step toward integrated “metabocentric” strategies for depression treatment.
Looking forward, interdisciplinary collaboration will be crucial for translating adiponectin research into clinical practice. Integrating psychiatry, endocrinology, and metabolism could yield innovative interventions that address both the mental and physical dimensions of depression. Advances in biomarker science, particularly the use of adiponectin in combination with other metabolic and inflammatory indicators, may further refine depression subtyping and support more precise, personalized treatment strategies. Ultimately, by bridging the gap between metabolic dysfunction and mood disorders, adiponectin may pave the way toward a new generation of “metabocentric” therapies that expand our approach to treating depression and related conditions.
Author Contributions
Conceptualization, P.G. and A.C.; methodology, P.G., I.D., G.P., M.G.D.B., V.B. and N.C.; investigation, P.G., I.D., G.P., M.G.D.B., V.B. and N.C.; writing—original draft preparation, P.G., I.D., G.P., M.G.D.B., V.B. and N.C.; writing—review and editing, A.C.; supervision, A.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Italian Ministry of Health to N.C. and A.C. (Ricerca Corrente). The PhD Scholarship to P.G. and G.P. were supported by European Union NextGenerationEU.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| BDNF | Brain-Derived Neurotrophic Factor |
| CNS | Central Nervous System |
| LTP | Long-Term Potentiation |
| AHN | Adult Hippocampal Neurogenesis |
| NSC | Neural Stem Cells |
| SGZ | Subgranular Zone |
| DG | Dentate Gyrus |
| LTD | Long-Term Depression |
| L-LTP | Late Phase Long-Term Potentiation |
| TRKB | Tropomyosin Receptor Kinase B |
| PLC | Phospholipase C |
| PI3K | Phosphatidylinositol-3 Kinase |
| mTOR | Mammalian Target Of Rapamycin |
| AMPK | AMP-Activated Protein Kinase |
| SSRI | Selective Serotonin Reuptake Inhibitor |
| WAT | White Adipose Tissue |
| HMW | High Molecular Weight |
| ER | Endoplasmatic Reticulum |
| PPAR | Proliferator-Activated Receptor |
| TZD | Thiazolidinedione |
| TNF-α | Tumour Necrosis Factor Alpha |
| IL-6 | Interleukin-6 |
| NF-κB | Nuclear Factor kB |
| BBB | Blood Brain Barrier |
| CSF | Cerebrospinal Fluid |
| IFN-α | Interferon Alpha |
| HCV | Hepatitis C Virus |
| BAT | Brown Adipos Tissue |
| GLUT4 | Glucose Transporter Type 4 |
| LPS | Lipopolysaccharide |
| GLP1RA | Glucagone-Like Peptide 1 Receptor Agonist |
References
- Tartt, A.N.; Mariani, M.B.; Hen, R.; Mann, J.J.; Boldrini, M. Dysregulation of Adult Hippocampal Neuroplasticity in Major Depression: Pathogenesis and Therapeutic Implications. Mol Psychiatry 2022, 27, 2689–2699. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Nie, H.; Zhang, Y.; Chen, Y.; Peng, Z.; Cai, M.; Wei, H.; Qin, P.; Dong, H.; Xiong, L. Recombinant Human Thioredoxin-1 Promotes Neurogenesis and Facilitates Cognitive Recovery Following Cerebral Ischemia in Mice. Neuropharmacology 2014, 77, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Borsini, A.; Giacobbe, J.; Mandal, G.; Boldrini, M. Acute and Long-Term Effects of Adolescence Stress Exposure on Rodent Adult Hippocampal Neurogenesis, Cognition, and Behaviour. Mol Psychiatry 2023, 28, 4124–4137. [Google Scholar] [CrossRef] [PubMed]
- Du Preez, A.; Onorato, D.; Eiben, I.; Musaelyan, K.; Egeland, M.; Zunszain, P.A.; Fernandes, C.; Thuret, S.; Pariante, C.M. Chronic Stress Followed by Social Isolation Promotes Depressive-like Behaviour, Alters Microglial and Astrocyte Biology and Reduces Hippocampal Neurogenesis in Male Mice. Brain Behav Immun 2021, 91, 24–47. [Google Scholar] [CrossRef]
- Lucassen, P.J.; Meerlo, P.; Naylor, A.S.; van Dam, A.M.; Dayer, A.G.; Fuchs, E.; Oomen, C.A.; Czéh, B. Regulation of Adult Neurogenesis by Stress, Sleep Disruption, Exercise and Inflammation: Implications for Depression and Antidepressant Action. Eur Neuropsychopharmacol 2010, 20, 1–17. [Google Scholar] [CrossRef]
- Boldrini, M.; Underwood, M.D.; Hen, R.; Rosoklija, G.B.; Dwork, A.J.; John Mann, J.; Arango, V. Antidepressants Increase Neural Progenitor Cells in the Human Hippocampus. Neuropsychopharmacology 2009, 34, 2376–2389. [Google Scholar] [CrossRef]
- Boldrini, M.; Hen, R.; Underwood, M.D.; Rosoklija, G.B.; Dwork, A.J.; Mann, J.J.; Arango, V. Hippocampal Angiogenesis and Progenitor Cell Proliferation Are Increased with Antidepressant Use in Major Depression. Biol Psychiatry 2012, 72, 562–571. [Google Scholar] [CrossRef]
- Moreno-Agostino, D.; Wu, Y.-T.; Daskalopoulou, C.; Hasan, M.T.; Huisman, M.; Prina, M. Global Trends in the Prevalence and Incidence of Depression:A Systematic Review and Meta-Analysis. Journal of Affective Disorders 2021, 281, 235–243. [Google Scholar] [CrossRef]
- Church, T.S.; Thomas, D.M.; Tudor-Locke, C.; Katzmarzyk, P.T.; Earnest, C.P.; Rodarte, R.Q.; Martin, C.K.; Blair, S.N.; Bouchard, C. Trends over 5 Decades in U. S. Occupation-Related Physical Activity and Their Associations with Obesity. PLoS One 2011, 6, e19657. [Google Scholar] [CrossRef]
- Hamilton, M.T.; Hamilton, D.G.; Zderic, T.W. Sedentary Behavior as a Mediator of Type 2 Diabetes. Med Sport Sci 2014, 60, 11–26. [Google Scholar] [CrossRef]
- Lopresti, A.L.; Hood, S.D.; Drummond, P.D. A Review of Lifestyle Factors That Contribute to Important Pathways Associated with Major Depression: Diet, Sleep and Exercise. J Affect Disord 2013, 148, 12–27. [Google Scholar] [CrossRef]
- Yau, S.-Y.; Lee, T.H.-Y.; Li, A.; Xu, A.; So, K.-F. Adiponectin Mediates Running-Restored Hippocampal Neurogenesis in Streptozotocin-Induced Type 1 Diabetes in Mice. Front Neurosci 2018, 12, 679. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M.; Bishop, N.C.; Stensel, D.J.; Lindley, M.R.; Mastana, S.S.; Nimmo, M.A. The Anti-Inflammatory Effects of Exercise: Mechanisms and Implications for the Prevention and Treatment of Disease. Nat Rev Immunol 2011, 11, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Cao, L.; Zhou, D. The Impact of Physical Activities on Adolescents’ Rule Consciousness: The Chain Mediation Effect of Friendship Quality and Emotional Intelligence. Front Public Health 2025, 13, 1581016. [Google Scholar] [CrossRef]
- Pearce, M.; Garcia, L.; Abbas, A.; Strain, T.; Schuch, F.B.; Golubic, R.; Kelly, P.; Khan, S.; Utukuri, M.; Laird, Y.; et al. Association Between Physical Activity and Risk of Depression: A Systematic Review and Meta-Analysis. JAMA Psychiatry 2022, 79, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Dishman, R.K.; McDowell, C.P.; Herring, M.P. Customary Physical Activity and Odds of Depression: A Systematic Review and Meta-Analysis of 111 Prospective Cohort Studies. Br J Sports Med 2021, 55, 926–934. [Google Scholar] [CrossRef]
- Roshanaei-Moghaddam, B.; Katon, W.J.; Russo, J. The Longitudinal Effects of Depression on Physical Activity. General Hospital Psychiatry 2009, 31, 306–315. [Google Scholar] [CrossRef]
- Vallance, J.K.; Winkler, E.A.H.; Gardiner, P.A.; Healy, G.N.; Lynch, B.M.; Owen, N. Associations of Objectively-Assessed Physical Activity and Sedentary Time with Depression: NHANES (2005–2006). Preventive Medicine 2011, 53, 284–288. [Google Scholar] [CrossRef]
- Berlin, A.A.; Kop, W.J.; Deuster, P.A. Depressive Mood Symptoms and Fatigue After Exercise Withdrawal: The Potential Role of Decreased Fitness. Psychosomatic Medicine 2006, 68, 224–230. [Google Scholar] [CrossRef]
- Soini, E.; Rosenström, T.; Määttänen, I.; Jokela, M. Physical Activity and Specific Symptoms of Depression: A Pooled Analysis of Six Cohort Studies. Journal of Affective Disorders 2024, 348, 44–53. [Google Scholar] [CrossRef]
- Blume, G.R.; Royes, L.F.F. Peripheral to Brain and Hippocampus Crosstalk Induced by Exercise Mediates Cognitive and Structural Hippocampal Adaptations. Life Sci 2024, 352, 122799. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, X.; Lu, X.-Y. Adiponectin Exerts Neurotrophic Effects on Dendritic Arborization, Spinogenesis, and Neurogenesis of the Dentate Gyrus of Male Mice. Endocrinology 2016, 157, 2853–2869. [Google Scholar] [CrossRef]
- Nicolas, S.; Cazareth, J.; Zarif, H.; Guyon, A.; Heurteaux, C.; Chabry, J.; Petit-Paitel, A. Globular Adiponectin Limits Microglia Pro-Inflammatory Phenotype through an AdipoR1/NF-κB Signaling Pathway. Front Cell Neurosci 2017, 11, 352. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liang, Y.; Chen, K.; Yau, S.-Y.; Sun, X.; Cheng, K.K.-Y.; Xu, A.; So, K.-F.; Li, A. Potential Involvement of Adiponectin Signaling in Regulating Physical Exercise-Elicited Hippocampal Neurogenesis and Dendritic Morphology in Stressed Mice. Front Cell Neurosci 2020, 14, 189. [Google Scholar] [CrossRef]
- Kempermann, G.; Gage, F.H.; Aigner, L.; Song, H.; Curtis, M.A.; Thuret, S.; Kuhn, H.G.; Jessberger, S.; Frankland, P.W.; Cameron, H.A.; et al. Human Adult Neurogenesis: Evidence and Remaining Questions. Cell Stem Cell 2018, 23, 25–30. [Google Scholar] [CrossRef]
- Zheng, M.; Zhang, B.; Yau, S.S.Y.; So, K.-F.; Zhang, L.; Ou, H. Exercise Preconditioning Alleviates Ischemia-Induced Memory Deficits by Increasing Circulating Adiponectin. Neural Regen Res 2025, 20, 1445–1454. [Google Scholar] [CrossRef]
- Mukherjee, S.; Manahan-Vaughan, D. Role of Metabotropic Glutamate Receptors in Persistent Forms of Hippocampal Plasticity and Learning. Neuropharmacology 2013, 66, 65–81. [Google Scholar] [CrossRef]
- Minal, N.; Nilesh, W.; Akanksha, K. Epigenetic Regulation in Neuroplasticity: Key to Understanding and Treating Neurological Diseases. Zhongguo Ying Yong Sheng Li Xue Za Zhi 2025, 41, e20250011. [Google Scholar] [CrossRef]
- Cramer, S.C.; Sur, M.; Dobkin, B.H.; O’Brien, C.; Sanger, T.D.; Trojanowski, J.Q.; Rumsey, J.M.; Hicks, R.; Cameron, J.; Chen, D.; et al. Harnessing Neuroplasticity for Clinical Applications. Brain 2011, 134, 1591–1609. [Google Scholar] [CrossRef]
- Mudgal, S.K.; Nath, S.; Chaturvedi, J.; Sharma, S.K.; Joshi, J. Neuroplasticity in Depression: A Narrative Review with Evidence-Based Insights. Psychiatr Danub 2022, 34, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Ferrarelli, F. Is Neuroplasticity Key to Treatment Response in Depression? Maybe So. Am J Psychiatry 2022, 179, 451–453. [Google Scholar] [CrossRef]
- Yuan, M.; Yang, B.; Rothschild, G.; Mann, J.J.; Sanford, L.D.; Tang, X.; Huang, C.; Wang, C.; Zhang, W. Epigenetic Regulation in Major Depression and Other Stress-Related Disorders: Molecular Mechanisms, Clinical Relevance and Therapeutic Potential. Signal Transduct Target Ther 2023, 8, 309. [Google Scholar] [CrossRef]
- Chen, F.; Bertelsen, A.B.; Holm, I.E.; Nyengaard, J.R.; Rosenberg, R.; Dorph-Petersen, K.-A. Hippocampal Volume and Cell Number in Depression, Schizophrenia, and Suicide Subjects. Brain Res 2020, 1727, 146546. [Google Scholar] [CrossRef] [PubMed]
- Cotter, D.; Mackay, D.; Chana, G.; Beasley, C.; Landau, S.; Everall, I.P. Reduced Neuronal Size and Glial Cell Density in Area 9 of the Dorsolateral Prefrontal Cortex in Subjects with Major Depressive Disorder. Cereb Cortex 2002, 12, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Rajkowska, G.; O’Dwyer, G.; Teleki, Z.; Stockmeier, C.A.; Miguel-Hidalgo, J.J. GABAergic Neurons Immunoreactive for Calcium Binding Proteins Are Reduced in the Prefrontal Cortex in Major Depression. Neuropsychopharmacology 2007, 32, 471–482. [Google Scholar] [CrossRef]
- Tunc-Ozcan, E.; Peng, C.-Y.; Zhu, Y.; Dunlop, S.R.; Contractor, A.; Kessler, J.A. Activating Newborn Neurons Suppresses Depression and Anxiety-like Behaviors. Nat Commun 2019, 10, 3768. [Google Scholar] [CrossRef]
- Surget, A.; Belzung, C. Adult Hippocampal Neurogenesis Shapes Adaptation and Improves Stress Response: A Mechanistic and Integrative Perspective. Mol Psychiatry 2022, 27, 403–421. [Google Scholar] [CrossRef]
- Nicolas, S.; Veyssière, J.; Gandin, C.; Zsürger, N.; Pietri, M.; Heurteaux, C.; Glaichenhaus, N.; Petit-Paitel, A.; Chabry, J. Neurogenesis-Independent Antidepressant-like Effects of Enriched Environment Is Dependent on Adiponectin. Psychoneuroendocrinology 2015, 57, 72–83. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, F.; Zhai, M.; He, M.; Hu, Y.; Feng, L.; Li, Y.; Yang, J.; Wu, C. Hyperactive Neuronal Autophagy Depletes BDNF and Impairs Adult Hippocampal Neurogenesis in a Corticosterone-Induced Mouse Model of Depression. Theranostics 2023, 13, 1059–1075. [Google Scholar] [CrossRef]
- Abe, Y.; Erchinger, V.J.; Ousdal, O.T.; Oltedal, L.; Tanaka, K.F.; Takamiya, A. Neurobiological Mechanisms of Electroconvulsive Therapy for Depression: Insights into Hippocampal Volumetric Increases from Clinical and Preclinical Studies. J Neurochem 2024, 168, 1738–1750. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Feng, Y.; Hong, Z.; Yin, M.; Zheng, H.; Zhang, L.; Hu, X. High-Frequency Repetitive Transcranial Magnetic Stimulation Promotes Neural Stem Cell Proliferation after Ischemic Stroke. Neural Regen Res 2024, 19, 1772–1780. [Google Scholar] [CrossRef]
- Zhao, R.; Tian, X.; Xu, H.; Wang, Y.; Lin, J.; Wang, B. Aerobic Exercise Restores Hippocampal Neurogenesis and Cognitive Function by Decreasing Microglia Inflammasome Formation Through Irisin/NLRP3 Pathway. Aging Cell 2025, e70061. [Google Scholar] [CrossRef]
- Bliss, T.V.P.; Cooke, S.F. Long-Term Potentiation and Long-Term Depression: A Clinical Perspective. Clinics (Sao Paulo) 2011, 66 Suppl 1, 3–17. [CrossRef]
- Duman, R.S.; Aghajanian, G.K. Synaptic Dysfunction in Depression: Potential Therapeutic Targets. Science 2012, 338, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Marsden, W.N. Synaptic Plasticity in Depression: Molecular, Cellular and Functional Correlates. Prog Neuropsychopharmacol Biol Psychiatry 2013, 43, 168–184. [Google Scholar] [CrossRef]
- Bin Ibrahim, M.Z.; Benoy, A.; Sajikumar, S. Long-term Plasticity in the Hippocampus: Maintaining within and ‘Tagging’ between Synapses. The FEBS Journal 2022, 289, 2176–2201. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Liu, L.; Zhang, G.; Wang, X.; Gu, T.; Luo, Q.; Sha, S.; Du, Y.; Wu, C.; Chen, L. Activation of Transient Receptor Potential Vanilloid 4 Impairs Long-Term Depression in Nucleus Accumbens and Induces Depressive-like Behavior. Neuropharmacology 2025, 273, 110429. [Google Scholar] [CrossRef]
- Abraham, W.C.; Jones, O.D.; Glanzman, D.L. Is Plasticity of Synapses the Mechanism of Long-Term Memory Storage? npj Sci. Learn. 2019, 4, 9. [Google Scholar] [CrossRef]
- Aleksandrova, L.R.; Wang, Y.T.; Phillips, A.G. Evaluation of the Wistar-Kyoto Rat Model of Depression and the Role of Synaptic Plasticity in Depression and Antidepressant Response. Neuroscience & Biobehavioral Reviews 2019, 105, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Csabai, D.; Wiborg, O.; Czéh, B. Reduced Synapse and Axon Numbers in the Prefrontal Cortex of Rats Subjected to a Chronic Stress Model for Depression. Front Cell Neurosci 2018, 12, 24. [Google Scholar] [CrossRef]
- Mahati, K.; Bhagya, V.; Christofer, T.; Sneha, A.; Shankaranarayana Rao, B.S. Enriched Environment Ameliorates Depression-Induced Cognitive Deficits and Restores Abnormal Hippocampal Synaptic Plasticity. Neurobiology of Learning and Memory 2016, 134, 379–391. [Google Scholar] [CrossRef]
- Ely, B.A.; Nguyen, T.N.B.; Tobe, R.H.; Walker, A.M.; Gabbay, V. Multimodal Investigations of Reward Circuitry and Anhedonia in Adolescent Depression. Front Psychiatry 2021, 12, 678709. [Google Scholar] [CrossRef]
- Liu, W.; Ge, T.; Leng, Y.; Pan, Z.; Fan, J.; Yang, W.; Cui, R. The Role of Neural Plasticity in Depression: From Hippocampus to Prefrontal Cortex. Neural Plasticity 2017, 2017, 1–11. [Google Scholar] [CrossRef]
- Rygvold, T.W.; Hatlestad-Hall, C.; Elvsåshagen, T.; Moberget, T.; Andersson, S. Long Term Potentiation-like Neural Plasticity and Performance-Based Memory Function. Neurobiology of Learning and Memory 2022, 196, 107696. [Google Scholar] [CrossRef]
- Baltaci, S.B.; Mogulkoc, R.; Baltaci, A.K. Molecular Mechanisms of Early and Late LTP. Neurochem Res 2019, 44, 281–296. [Google Scholar] [CrossRef]
- Kandel, E.R.; Dudai, Y.; Mayford, M.R. The Molecular and Systems Biology of Memory. Cell 2014, 157, 163–186. [Google Scholar] [CrossRef] [PubMed]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell Mol Neurobiol 2018, 38, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Panja, D.; Bramham, C.R. BDNF Mechanisms in Late LTP Formation: A Synthesis and Breakdown. Neuropharmacology 2014, 76, 664–676. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, A.; Cattane, N.; Begni, V.; Pariante, C.M.; Riva, M.A. The Human BDNF Gene: Peripheral Gene Expression and Protein Levels as Biomarkers for Psychiatric Disorders. Transl Psychiatry 2016, 6, e958. [Google Scholar] [CrossRef]
- Cavaleri, D.; Moretti, F.; Bartoccetti, A.; Mauro, S.; Crocamo, C.; Carrà, G.; Bartoli, F. The Role of BDNF in Major Depressive Disorder, Related Clinical Features, and Antidepressant Treatment: Insight from Meta-Analyses. Neuroscience & Biobehavioral Reviews 2023, 149, 105159. [Google Scholar] [CrossRef]
- Correia, A.S.; Cardoso, A.; Vale, N. BDNF Unveiled: Exploring Its Role in Major Depression Disorder Serotonergic Imbalance and Associated Stress Conditions. Pharmaceutics 2023, 15, 2081. [Google Scholar] [CrossRef]
- Kishi, T.; Yoshimura, R.; Ikuta, T.; Iwata, N. Brain-Derived Neurotrophic Factor and Major Depressive Disorder: Evidence from Meta-Analyses. Front. Psychiatry 2018, 8, 308. [Google Scholar] [CrossRef]
- Molendijk, M.L.; Spinhoven, P.; Polak, M.; Bus, B.A.A.; Penninx, B.W.J.H.; Elzinga, B.M. Serum BDNF Concentrations as Peripheral Manifestations of Depression: Evidence from a Systematic Review and Meta-Analyses on 179 Associations (N=9484). Mol Psychiatry 2014, 19, 791–800. [Google Scholar] [CrossRef]
- Odaira, T.; Nakagawasai, O.; Takahashi, K.; Nemoto, W.; Sakuma, W.; Lin, J.-R.; Tan-No, K. Mechanisms Underpinning AMP-Activated Protein Kinase-Related Effects on Behavior and Hippocampal Neurogenesis in an Animal Model of Depression. Neuropharmacology 2019, 150, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Cao, J.; Liu, X.; Meng, F.; Li, M.; Chen, B.; Zhang, J. AMPK Plays a Dual Role in Regulation of CREB/BDNF Pathway in Mouse Primary Hippocampal Cells. J Mol Neurosci 2015, 56, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, F.; Rossetti, A.C.; Racagni, G.; Gass, P.; Riva, M.A.; Molteni, R. Brain-Derived Neurotrophic Factor: A Bridge between Inflammation and Neuroplasticity. Front Cell Neurosci 2014, 8, 430. [Google Scholar] [CrossRef]
- Cattaneo, A.; Macchi, F.; Plazzotta, G.; Veronica, B.; Bocchio-Chiavetto, L.; Riva, M.A.; Pariante, C.M. Inflammation and Neuronal Plasticity: A Link between Childhood Trauma and Depression Pathogenesis. Front. Cell. Neurosci. 2015, 9. [Google Scholar] [CrossRef]
- Pedraz-Petrozzi, B.; Insan, S.; Spangemacher, M.; Reinwald, J.; Lamadé, E.K.; Gilles, M.; Deuschle, M.; Sartorius, A. Association between rTMS-Induced Changes in Inflammatory Markers and Improvement in Psychiatric Diseases: A Systematic Review. Ann Gen Psychiatry 2024, 23, 31. [Google Scholar] [CrossRef]
- Qian, X.; Zhong, Z.; Zhang, Y.; Qiu, L.; Tan, H. Fluoxetine Mitigates Depressive-like Behaviors in Mice via Anti-Inflammation and Enhancing Neuroplasticity. Brain Research 2024, 1825, 148723. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Luo, Y.; Liang, X.; Tang, J.; Wang, J.; Xiao, Q.; Qi, Y.; Li, Y.; Zhu, P.; Yang, H.; et al. Beneficial Effects of Running Exercise on Hippocampal Microglia and Neuroinflammation in Chronic Unpredictable Stress-Induced Depression Model Rats. Transl Psychiatry 2021, 11, 461. [Google Scholar] [CrossRef]
- Holmes, S.E.; Scheinost, D.; Finnema, S.J.; Naganawa, M.; Davis, M.T.; DellaGioia, N.; Nabulsi, N.; Matuskey, D.; Angarita, G.A.; Pietrzak, R.H.; et al. Lower Synaptic Density Is Associated with Depression Severity and Network Alterations. Nat Commun 2019, 10, 1529. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Martín-Rodríguez, A.; Martínez-Guardado, I.; Navarro-Jiménez, E.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. The Role of Adipokines in Health and Disease. Biomedicines 2023, 11, 1290. [Google Scholar] [CrossRef] [PubMed]
- Guenther, M.; James, R.; Marks, J.; Zhao, S.; Szabo, A.; Kidambi, S. Adiposity Distribution Influences Circulating Adiponectin Levels. Translational Research 2014, 164, 270–277. [Google Scholar] [CrossRef]
- Arias, C.; Álvarez-Indo, J.; Cifuentes, M.; Morselli, E.; Kerr, B.; Burgos, P.V. Enhancing Adipose Tissue Functionality in Obesity: Senotherapeutics, Autophagy and Cellular Senescence as a Target. Biol Res 2024, 57, 51. [Google Scholar] [CrossRef] [PubMed]
- Waki, H.; Yamauchi, T.; Kamon, J.; Ito, Y.; Uchida, S.; Kita, S.; Hara, K.; Hada, Y.; Vasseur, F.; Froguel, P.; et al. Impaired Multimerization of Human Adiponectin Mutants Associated with Diabetes. Journal of Biological Chemistry 2003, 278, 40352–40363. [Google Scholar] [CrossRef]
- Liu, M.; Liu, F. Regulation of Adiponectin Multimerization, Signaling and Function. Best Practice & Research Clinical Endocrinology & Metabolism 2014, 28, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metabolic Syndrome and Related Disorders 2015, 13, 423–444. [Google Scholar] [CrossRef]
- Tripathi, Y.B.; Pandey, V. Obesity and Endoplasmic Reticulum (ER) Stresses. Front. Immun. 2012, 3. [Google Scholar] [CrossRef]
- Park, H.S.; Lim, J.H.; Kim, M.Y.; Kim, Y.; Hong, Y.A.; Choi, S.R.; Chung, S.; Kim, H.W.; Choi, B.S.; Kim, Y.S.; et al. Resveratrol Increases AdipoR1 and AdipoR2 Expression in Type 2 Diabetic Nephropathy. J Transl Med 2016, 14, 176. [Google Scholar] [CrossRef]
- Li, J.; Zou, B.; Yeo, Y.H.; Feng, Y.; Xie, X.; Lee, D.H.; Fujii, H.; Wu, Y.; Kam, L.Y.; Ji, F.; et al. Prevalence, Incidence, and Outcome of Non-Alcoholic Fatty Liver Disease in Asia, 1999–2019: A Systematic Review and Meta-Analysis. The Lancet Gastroenterology & Hepatology 2019, 4, 389–398. [Google Scholar] [CrossRef]
- Wang, C.W.; Feng, S.; Covinsky, K.E.; Hayssen, H.; Zhou, L.-Q.; Yeh, B.M.; Lai, J.C. A Comparison of Muscle Function, Mass, and Quality in Liver Transplant Candidates: Results From the Functional Assessment in Liver Transplantation Study. Transplantation 2016, 100, 1692–1698. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Y.; Shao, H.; Liu, S.; Niu, Y.; Fu, L. Globular Adiponectin Ameliorates Insulin Resistance in Skeletal Muscle by Enhancing the LKB1-Mediated AMPK Activation via SESN2. Sports Medicine and Health Science 2023, 5, 34–41. [Google Scholar] [CrossRef]
- Blandin, A.; Amosse, J.; Froger, J.; Hilairet, G.; Durcin, M.; Fizanne, L.; Ghesquière, V.; Prieur, X.; Chaigneau, J.; Vergori, L.; et al. Extracellular Vesicles Are Carriers of Adiponectin with Insulin-Sensitizing and Anti-Inflammatory Properties. Cell Reports 2023, 42, 112866. [Google Scholar] [CrossRef]
- Litvinova, L.; Atochin, D.; Vasilenko, M.; Fattakhov, N.; Zatolokin, P.; Vaysbeyn, I.; Kirienkova, E. Role of Adiponectin and Proinflammatory Gene Expression in Adipose Tissue Chronic Inflammation in Women with Metabolic Syndrome. Diabetol Metab Syndr 2014, 6, 137. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Walsh, K. Adiponectin as an Anti-Inflammatory Factor. Clinica Chimica Acta 2007, 380, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Ramírez, P.; Malmhäll, C.; Tliba, O.; Rådinger, M.; Bossios, A. Adiponectin/AdipoR1 Axis Promotes IL-10 Release by Human Regulatory T Cells. Front. Immunol. 2021, 12, 677550. [Google Scholar] [CrossRef]
- Basak, S.; Murmu, A.; Matore, B.W.; Roy, P.P.; Singh, J. Thiazolidinedione an Auspicious Scaffold as PPAR-γ Agonist: Its Possible Mechanism to Manoeuvre against Insulin Resistant Diabetes Mellitus. European Journal of Medicinal Chemistry Reports 2024, 11, 100160. [Google Scholar] [CrossRef]
- Hui, X.; Lam, K.S.; Vanhoutte, P.M.; Xu, A. Adiponectin and Cardiovascular Health: An Update. British J Pharmacology 2012, 165, 574–590. [Google Scholar] [CrossRef]
- Honda, M.; Tsuboi, A.; Minato, S.; Kitaoka, K.; Takeuchi, M.; Yano, M.; Kurata, M.; Wu, B.; Kazumi, T.; Fukuo, K. Association of Age and Anemia With Adiponectin Serum Levels in Normal-Weight Japanese Women. J Clin Med Res 2019, 11, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Takahashi, N.; Hileman, S.M.; Patel, H.R.; Berg, A.H.; Pajvani, U.B.; Scherer, P.E.; Ahima, R.S. Adiponectin Acts in the Brain to Decrease Body Weight. Nat Med 2004, 10, 524–529. [Google Scholar] [CrossRef]
- Yau, S.Y.; Li, A.; Hoo, R.L.C.; Ching, Y.P.; Christie, B.R.; Lee, T.M.C.; Xu, A.; So, K.-F. Physical Exercise-Induced Hippocampal Neurogenesis and Antidepressant Effects Are Mediated by the Adipocyte Hormone Adiponectin. Proc Natl Acad Sci U S A 2014, 111, 15810–15815. [Google Scholar] [CrossRef]
- Neumeier, M.; Weigert, J.; Buettner, R.; Wanninger, J.; Schäffler, A.; Müller, A.M.; Killian, S.; Sauerbruch, S.; Schlachetzki, F.; Steinbrecher, A.; et al. Detection of Adiponectin in Cerebrospinal Fluid in Humans. Am J Physiol Endocrinol Metab 2007, 293, E965–969. [Google Scholar] [CrossRef] [PubMed]
- Kusminski, C.M.; McTernan, P.G.; Schraw, T.; Kos, K.; O’Hare, J.P.; Ahima, R.; Kumar, S.; Scherer, P.E. Adiponectin Complexes in Human Cerebrospinal Fluid: Distinct Complex Distribution from Serum. Diabetologia 2007, 50, 634–642. [Google Scholar] [CrossRef]
- Clain, J.; Couret, D.; Planesse, C.; Krejbich-Trotot, P.; Meilhac, O.; Lefebvre d’Hellencourt, C.; Viranaicken, W.; Diotel, N. Distribution of Adiponectin Receptors in the Brain of Adult Mouse: Effect of a Single Dose of the Adiponectin Receptor Agonist, AdipoRON, on Ischemic Stroke. Brain Sci 2022, 12, 680. [Google Scholar] [CrossRef] [PubMed]
- Bloemer, J.; Pinky, P.D.; Smith, W.D.; Bhattacharya, D.; Chauhan, A.; Govindarajulu, M.; Hong, H.; Dhanasekaran, M.; Judd, R.; Amin, R.H.; et al. Adiponectin Knockout Mice Display Cognitive and Synaptic Deficits. Front Endocrinol (Lausanne) 2019, 10, 819. [Google Scholar] [CrossRef] [PubMed]
- Gorska-Ciebiada, M.; Ciebiada, M. Adiponectin and Inflammatory Marker Levels in the Elderly Patients with Diabetes, Mild Cognitive Impairment and Depressive Symptoms. IJMS 2024, 25, 10804. [Google Scholar] [CrossRef] [PubMed]
- Lehto, S.M.; Huotari, A.; Niskanen, L.; Tolmunen, T.; Koivumaa-Honkanen, H.; Honkalampi, K.; Ruotsalainen, H.; Herzig, K.-H.; Viinamäki, H.; Hintikka, J. Serum Adiponectin and Resistin Levels in Major Depressive Disorder. Acta Psychiatr Scand 2010, 121, 209–215. [Google Scholar] [CrossRef]
- Leo, R.; Di Lorenzo, G.; Tesauro, M.; Cola, C.; Fortuna, E.; Zanasi, M.; Troisi, A.; Siracusano, A.; Lauro, R.; Romeo, F. Decreased Plasma Adiponectin Concentration in Major Depression. Neurosci Lett 2006, 407, 211–213. [Google Scholar] [CrossRef]
- Zeugmann, S.; Quante, A.; Heuser, I.; Schwarzer, R.; Anghelescu, I. Inflammatory Biomarkers in 70 Depressed Inpatients with and without the Metabolic Syndrome. J Clin Psychiatry 2010, 71, 1007–1016. [Google Scholar] [CrossRef]
- Fábregas, B.C.; Vieira, É.L.M.; Moura, A.S.; Carmo, R.A.; Ávila, R.E.; Abreu, M.N.S.; Prossin, A.R.; Teixeira, A.L. A Follow-Up Study of 50 Chronic Hepatitis C Patients: Adiponectin as a Resilience Biomarker for Major Depression. Neuroimmunomodulation 2016, 23, 88–97. [Google Scholar] [CrossRef]
- Sun, F.; Lei, Y.; You, J.; Li, C.; Sun, L.; Garza, J.; Zhang, D.; Guo, M.; Scherer, P.E.; Lodge, D.; et al. Adiponectin Modulates Ventral Tegmental Area Dopamine Neuron Activity and Anxiety-Related Behavior through AdipoR1. Mol Psychiatry 2019, 24, 126–144. [Google Scholar] [CrossRef]
- D’Aprile, I.; Petrillo, G.; Zonca, V.; Mazzelli, M.; De Cillis, F.; Di Benedetto, M.G.; Riva, M.A.; Cattaneo, A. Sex-Specific Metabolic and Inflammatory Alterations in Adult Animals Vulnerable to Prenatal Stress Exposure. Progress in Neuro-Psychopharmacology and Biological Psychiatry 2025, 138, 111344. [Google Scholar] [CrossRef]
- Liu, L.; Tang, J.; Liang, X.; Li, Y.; Zhu, P.; Zhou, M.; Qin, L.; Deng, Y.; Li, J.; Wang, Y.; et al. Running Exercise Alleviates Hippocampal Neuroinflammation and Shifts the Balance of Microglial M1/M2 Polarization through Adiponectin/AdipoR1 Pathway Activation in Mice Exposed to Chronic Unpredictable Stress. Mol Psychiatry 2024, 29, 2031–2042. [Google Scholar] [CrossRef]
- Pousti, F.; Ahmadi, R.; Mirahmadi, F.; Hosseinmardi, N.; Rohampour, K. Adiponectin Modulates Synaptic Plasticity in Hippocampal Dentate Gyrus. Neuroscience Letters 2018, 662, 227–232. [Google Scholar] [CrossRef]
- Rizzo, M.R.; Fasano, R.; Paolisso, G. Adiponectin and Cognitive Decline. IJMS 2020, 21, 2010. [Google Scholar] [CrossRef] [PubMed]
- Furman, J.L.; Soyombo, A.; Czysz, A.H.; Jha, M.K.; Carmody, T.J.; Mason, B.L.; Scherer, P.E.; Trivedi, M.H. Adiponectin Moderates Antidepressant Treatment Outcome in the Combining Medications to Enhance Depression Outcomes Randomized Clinical Trial. Pers Med Psychiatry 2018, 9–10, 1–7. [CrossRef]
- Machado-Vieira, R.; Gold, P.W.; Luckenbaugh, D.A.; Ballard, E.D.; Richards, E.M.; Henter, I.D.; De Sousa, R.T.; Niciu, M.J.; Yuan, P.; Zarate, C.A. The Role of Adipokines in the Rapid Antidepressant Effects of Ketamine. Mol Psychiatry 2017, 22, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Permoda-Pachuta, A.; Malewska-Kasprzak, M.; Skibińska, M.; Rzepski, K.; Dmitrzak-Węglarz, M. Changes in Adipokine, Resitin, and BDNF Concentrations in Treatment-Resistant Depression after Electroconvulsive Therapy. Brain Sci 2023, 13, 1358. [Google Scholar] [CrossRef] [PubMed]
- Chow, L.S.; Gerszten, R.E.; Taylor, J.M.; Pedersen, B.K.; Van Praag, H.; Trappe, S.; Febbraio, M.A.; Galis, Z.S.; Gao, Y.; Haus, J.M.; et al. Exerkines in Health, Resilience and Disease. Nat Rev Endocrinol 2022, 18, 273–289. [Google Scholar] [CrossRef]
- Sirico, F.; Bianco, A.; D’Alicandro, G.; Castaldo, C.; Montagnani, S.; Spera, R.; Di Meglio, F.; Nurzynska, D. Effects of Physical Exercise on Adiponectin, Leptin, and Inflammatory Markers in Childhood Obesity: Systematic Review and Meta-Analysis. Child Obes 2018, 14, 207–217. [Google Scholar] [CrossRef]
- Bruun, J.M.; Helge, J.W.; Richelsen, B.; Stallknecht, B. Diet and Exercise Reduce Low-Grade Inflammation and Macrophage Infiltration in Adipose Tissue but Not in Skeletal Muscle in Severely Obese Subjects. Am J Physiol Endocrinol Metab 2006, 290, E961–967. [Google Scholar] [CrossRef]
- Fonseca, T.R.; Mendes, T.T.; Ramos, G.P.; Cabido, C.E.T.; Morandi, R.F.; Ferraz, F.O.; Miranda, A.S.; Mendonça, V.A.; Teixeira, A.L.; Silami-Garcia, E.; et al. Aerobic Training Modulates the Increase in Plasma Concentrations of Cytokines in Response to a Session of Exercise. J Environ Public Health 2021, 2021, 1304139. [Google Scholar] [CrossRef]
- Jürimäe, J.; Purge, P.; Jürimäe, T. Adiponectin and Stress Hormone Responses to Maximal Sculling after Volume-Extended Training Season in Elite Rowers. Metabolism 2006, 55, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, R.R.; Aboudehen, K.S.; Carruth, A.K.; Durand, R.T.J.; Acevedo, E.O.; Hebert, E.P.; Johnson, L.G.; Castracane, V.D. Adiponectin Responses to Continuous and Progressively Intense Intermittent Exercise. Med Sci Sports Exerc 2003, 35, 1320–1325. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, M.A.; White, L.J.; McCoy, S.; Kim, H.-W.; Petty, T.; Wilsey, J. Plasma Adiponectin Response to Acute Exercise in Healthy Subjects. Eur J Appl Physiol 2004, 91, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Jamurtas, A.Z.; Theocharis, V.; Koukoulis, G.; Stakias, N.; Fatouros, I.G.; Kouretas, D.; Koutedakis, Y. The Effects of Acute Exercise on Serum Adiponectin and Resistin Levels and Their Relation to Insulin Sensitivity in Overweight Males. Eur J Appl Physiol 2006, 97, 122–126. [Google Scholar] [CrossRef]
- Yang, W.; Liu, L.; Wei, Y.; Fang, C.; Zhou, F.; Chen, J.; Han, Q.; Huang, M.; Tan, X.; Liu, Q.; et al. Exercise Ameliorates the FGF21-Adiponectin Axis Impairment in Diet-Induced Obese Mice. Endocr Connect 2019, 8, 596–604. [Google Scholar] [CrossRef]
- Cano-Montoya, J.; Bentes, A.; Pavez, Y.; Rubilar, P.; Lavoz, C.; Ehrenfeld, P.; Sandoval, V.; Martínez-Huenchullán, S. Metabolic Response After a Single Maximal Exercise Session in Physically Inactive Young Adults (EASY Study): Relevancy of Adiponectin Isoforms. Biomolecules 2025, 15, 314. [Google Scholar] [CrossRef]
- Narciso, P.H.; von Ah Morano, A.E.; Agostinete, R.R.; Werneck, A.O.; Giannopoulos, A.J.; Bell, M.; Antunes, B.M.; Lira, F.S.; Fernandes, R.A.; Klentrou, P. Cytokine and Adipokine Response Following High-Intensity Interval Running and Cycling in Female Adolescents. Eur J Appl Physiol 2025. [CrossRef]
- Vidal, P.; Stanford, K.I. Exercise-Induced Adaptations to Adipose Tissue Thermogenesis. Front Endocrinol (Lausanne) 2020, 11, 270. [Google Scholar] [CrossRef]
- Baldelli, S.; Aiello, G.; Mansilla Di Martino, E.; Campaci, D.; Muthanna, F.M.S.; Lombardo, M. The Role of Adipose Tissue and Nutrition in the Regulation of Adiponectin. Nutrients 2024, 16, 2436. [Google Scholar] [CrossRef]
- Félix-Soriano, E.; Sáinz, N.; Gil-Iturbe, E.; Castilla-Madrigal, R.; Celay, J.; Fernández-Galilea, M.; Pejenaute, Á.; Lostao, M.P.; Martínez-Climent, J.A.; Moreno-Aliaga, M.J. Differential Remodeling of Subcutaneous White and Interscapular Brown Adipose Tissue by Long-Term Exercise Training in Aged Obese Female Mice. J Physiol Biochem 2023, 79, 451–465. [Google Scholar] [CrossRef]
- Ryan, B.J.; Schleh, M.W.; Ahn, C.; Ludzki, A.C.; Gillen, J.B.; Varshney, P.; Van Pelt, D.W.; Pitchford, L.M.; Chenevert, T.L.; Gioscia-Ryan, R.A.; et al. Moderate-Intensity Exercise and High-Intensity Interval Training Affect Insulin Sensitivity Similarly in Obese Adults. J Clin Endocrinol Metab 2020, 105, e2941–2959. [Google Scholar] [CrossRef] [PubMed]
- Blümer, R.M.E.; van Roomen, C.P.; Meijer, A.J.; Houben-Weerts, J.H.P.M.; Sauerwein, H.P.; Dubbelhuis, P.F. Regulation of Adiponectin Secretion by Insulin and Amino Acids in 3T3-L1 Adipocytes. Metabolism 2008, 57, 1655–1662. [Google Scholar] [CrossRef]
- Hajri, T.; Tao, H.; Wattacheril, J.; Marks-Shulman, P.; Abumrad, N.N. Regulation of Adiponectin Production by Insulin: Interactions with Tumor Necrosis Factor-α and Interleukin-6. Am J Physiol Endocrinol Metab 2011, 300, E350–360. [Google Scholar] [CrossRef]
- Westerbacka, J.; Cornér, A.; Kannisto, K.; Kolak, M.; Makkonen, J.; Korsheninnikova, E.; Nyman, T.; Hamsten, A.; Fisher, R.M.; Yki-Järvinen, H. Acute in Vivo Effects of Insulin on Gene Expression in Adipose Tissue in Insulin-Resistant and Insulin-Sensitive Subjects. Diabetologia 2006, 49, 132–140. [Google Scholar] [CrossRef]
- Motta, V.F.; Aguila, M.B.; Mandarim-DE-Lacerda, C.A. High-Intensity Interval Training (Swimming) Significantly Improves the Adverse Metabolism and Comorbidities in Diet-Induced Obese Mice. J Sports Med Phys Fitness 2016, 56, 655–663. [Google Scholar]
- Pala, R.; Genc, E.; Tuzcu, M.; Orhan, C.; Sahin, N.; Er, B.; Cinar, V.; Sahin, K. L-Carnitine Supplementation Increases Expression of PPAR-Î3 and Glucose Transporters in Skeletal Muscle of Chronically and Acutely Exercised Rats. Cell Mol Biol (Noisy-le-grand) 2018, 64, 1–6. [Google Scholar] [CrossRef]
- Shirvani, H.; Mirnejad, R.; Soleimani, M.; Arabzadeh, E. Swimming Exercise Improves Gene Expression of PPAR-γ and Downregulates the Overexpression of TLR4, MyD88, IL-6, and TNF-α after High-Fat Diet in Rat Skeletal Muscle Cells. Gene 2021, 775, 145441. [Google Scholar] [CrossRef] [PubMed]
- Korbecki, J.; Bobiński, R.; Dutka, M. Self-Regulation of the Inflammatory Response by Peroxisome Proliferator-Activated Receptors. Inflamm Res 2019, 68, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Gao, X.; Ge, Q.; Tai, W.; Hao, X.; Shao, Q.; Fang, Z.; Chen, M.; Song, Y.; Gao, W.; et al. Tumor Necrosis Factor-α Reduces Adiponectin Production by Decreasing Transcriptional Activity of Peroxisome Proliferator-Activated Receptor-γ in Calf Adipocytes. J Dairy Sci 2023, 106, 5182–5195. [Google Scholar] [CrossRef]
- Monsalve, F.A.; Pyarasani, R.D.; Delgado-Lopez, F.; Moore-Carrasco, R. Peroxisome Proliferator-Activated Receptor Targets for the Treatment of Metabolic Diseases. Mediators Inflamm 2013, 2013, 549627. [Google Scholar] [CrossRef]
- Pajvani, U.B.; Hawkins, M.; Combs, T.P.; Rajala, M.W.; Doebber, T.; Berger, J.P.; Wagner, J.A.; Wu, M.; Knopps, A.; Xiang, A.H.; et al. Complex Distribution, Not Absolute Amount of Adiponectin, Correlates with Thiazolidinedione-Mediated Improvement in Insulin Sensitivity. J Biol Chem 2004, 279, 12152–12162. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Sun, J.; Huang, J.; Yu, X.; Yu, A.; He, Y.; Li, Q.; Yang, Z. Peroxisome Proliferator-Activated Receptor γ Enhances Adiponectin Secretion via up-Regulating DsbA-L Expression. Molecular and Cellular Endocrinology 2015, 411, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lam, K.S.L.; Yau, M.; Xu, A. Post-Translational Modifications of Adiponectin: Mechanisms and Functional Implications. Biochemical Journal 2008, 409, 623–633. [Google Scholar] [CrossRef]
- Mohammadi, A.; Gholamhoseinian, A.; Fallah, H. Zataria Multiflora Increases Insulin Sensitivity and PPARγ Gene Expression in High Fructose Fed Insulin Resistant Rats. Iran J Basic Med Sci 2014, 17, 263–270. [Google Scholar]
- Hargreaves, M.; Spriet, L.L. Skeletal Muscle Energy Metabolism during Exercise. Nat Metab 2020, 2, 817–828. [Google Scholar] [CrossRef]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of Cellular Energy Sensing and Restoration of Metabolic Balance. Mol Cell 2017, 66, 789–800. [Google Scholar] [CrossRef]
- Fang, H.; Judd, R.L. Adiponectin Regulation and Function. Compr Physiol 2018, 8, 1031–1063. [Google Scholar] [CrossRef]
- Tishinsky, J.M.; Dyck, D.J.; Robinson, L.E. Lifestyle Factors Increasing Adiponectin Synthesis and Secretion. Vitam Horm 2012, 90, 1–30. [Google Scholar] [CrossRef]
- Lee, T.H.; Christie, B.R.; van Praag, H.; Lin, K.; Siu, P.M.-F.; Xu, A.; So, K.-F.; Yau, S.-Y. AdipoRon Treatment Induces a Dose-Dependent Response in Adult Hippocampal Neurogenesis. Int J Mol Sci 2021, 22, 2068. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Sun, L.; Wang, J.; Sun, F.; Wang, W.; Wang, D.; Fan, X.; Liu, D.; Xu, Z.; Qiu, C.; et al. Role of Adiponectin-Notch Pathway in Cognitive Dysfunction Associated with Depression and in the Therapeutic Effect of Physical Exercise. Aging Cell 2021, 20, e13387. [Google Scholar] [CrossRef]
- Liu, B.; Liu, J.; Wang, J.-G.; Liu, C.-L.; Yan, H.-J. AdipoRon Improves Cognitive Dysfunction of Alzheimer’s Disease and Rescues Impaired Neural Stem Cell Proliferation through AdipoR1/AMPK Pathway. Exp Neurol 2020, 327, 113249. [Google Scholar] [CrossRef]
- Whitaker, R.H.; Cook, J.G. Stress Relief Techniques: P38 MAPK Determines the Balance of Cell Cycle and Apoptosis Pathways. Biomolecules 2021, 11, 1444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Guo, M.; Zhang, W.; Lu, X.-Y. Adiponectin Stimulates Proliferation of Adult Hippocampal Neural Stem/Progenitor Cells through Activation of P38 Mitogen-Activated Protein Kinase (p38MAPK)/Glycogen Synthase Kinase 3β (GSK-3β)/β-Catenin Signaling Cascade. J Biol Chem 2011, 286, 44913–44920. [Google Scholar] [CrossRef]
- Song, J.; Kang, S.M.; Kim, E.; Kim, C.-H.; Song, H.-T.; Lee, J.E. Adiponectin Receptor-Mediated Signaling Ameliorates Cerebral Cell Damage and Regulates the Neurogenesis of Neural Stem Cells at High Glucose Concentrations: An in Vivo and in Vitro Study. Cell Death Dis 2015, 6, e1844. [Google Scholar] [CrossRef]
- Lampada, A.; Taylor, V. Notch Signaling as a Master Regulator of Adult Neurogenesis. Front. Neurosci. 2023, 17, 1179011. [Google Scholar] [CrossRef]
- Shah, S.A.; Yoon, G.H.; Chung, S.S.; Abid, M.N.; Kim, T.H.; Lee, H.Y.; Kim, M.O. Novel Osmotin Inhibits SREBP2 via the AdipoR1/AMPK/SIRT1 Pathway to Improve Alzheimer’s Disease Neuropathological Deficits. Mol Psychiatry 2017, 22, 407–416. [Google Scholar] [CrossRef]
- Wang, N.; Zhu, S.; Chen, S.; Zou, J.; Zeng, P.; Tan, S. Neurological Mechanism-Based Analysis of the Role and Characteristics of Physical Activity in the Improvement of Depressive Symptoms. Rev Neurosci 2025. [CrossRef]
- Yu, L.; Wang, J.; Xia, Y.; Hou, W.; Yao, X.; Guo, Y.; Wang, J.; Wei, H.; Wang, S. Adiponectin Promotes Neurogenesis After Transient Cerebral Ischemia Through STAT3 Mediated BDNF Upregulation in Astrocytes. Neurochem Res 2023, 48, 641–657. [Google Scholar] [CrossRef]
- Nicolas, S.; Debayle, D.; Béchade, C.; Maroteaux, L.; Gay, A.-S.; Bayer, P.; Heurteaux, C.; Guyon, A.; Chabry, J. Adiporon, an Adiponectin Receptor Agonist Acts as an Antidepressant and Metabolic Regulator in a Mouse Model of Depression. Transl Psychiatry 2018, 8, 159. [Google Scholar] [CrossRef] [PubMed]
- Chabry, J.; Nicolas, S.; Cazareth, J.; Murris, E.; Guyon, A.; Glaichenhaus, N.; Heurteaux, C.; Petit-Paitel, A. Enriched Environment Decreases Microglia and Brain Macrophages Inflammatory Phenotypes through Adiponectin-Dependent Mechanisms: Relevance to Depressive-like Behavior. Brain Behav Immun 2015, 50, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.-H.; Ou, H.-N.; Yu, J.-S.; Yau, S.-Y.; Tsang, H.W.-H. Pharmacological Blocking of Adiponectin Receptors Induces Alzheimer’s Disease-like Neuropathology and Impairs Hippocampal Function. Biomedicines 2025, 13, 1056. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, F.; Branchi, I.; Poletti, S.; Lorenzi, C.; Bigai, G.; Colombo, C.; Zanardi, R. Adiponectin Predicts Poor Response to Antidepressant Drugs in Major Depressive Disorder. Human Psychopharmacology 2021, 36, e2793. [Google Scholar] [CrossRef] [PubMed]
- Houminer-Klepar, N.; Dopelt, K. Associations Between Mediterranean Diet, Processed Food Consumption, and Symptoms of Anxiety and Depression: Cross-Sectional Study Among Israeli Adults. Foods 2025, 14, 1485. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Cuenca, J.A.; Díaz-Jiménez, D.E.; Pineda-Juárez, J.A.; Mendoza-Mota, A.G.; Valencia-Aldana, O.D.; Núñez-Angeles, S.; Vera-Gómez, E.; Hernández-Patricio, A.; Loeza-Magaña, P.; Lara-Vargas, J.A.; et al. Effect of Mediterranean Diet in Combination with Isokinetic Exercise Therapy on Body Composition and Cytokine Profile in Patients with Metabolic Syndrome. Nutrients 2025, 17, 256. [Google Scholar] [CrossRef]
- Shams, H.A.; Aziz, H.M.; Al-Kuraishy, H.M. The Potential Effects of Metformin and/or Sitagliptin on Leptin/Adiponectin Ratio in Diabetic Obese Patients: A New Therapeutic Effect. J Pak Med Assoc 2024, 74, S241–S245. [Google Scholar] [CrossRef]
- Khedr, S.A.; Elmelgy, A.A.; El-Kharashi, O.A.; Abd-Alkhalek, H.A.; Louka, M.L.; Sallam, H.A.; Aboul-Fotouh, S. Metformin Potentiates Cognitive and Antidepressant Effects of Fluoxetine in Rats Exposed to Chronic Restraint Stress and High Fat Diet: Potential Involvement of Hippocampal c-Jun Repression. Naunyn Schmiedebergs Arch Pharmacol 2018, 391, 407–422. [Google Scholar] [CrossRef]
- Femminella, G.D.; Frangou, E.; Love, S.B.; Busza, G.; Holmes, C.; Ritchie, C.; Lawrence, R.; McFarlane, B.; Tadros, G.; Ridha, B.H.; et al. Evaluating the Effects of the Novel GLP-1 Analogue Liraglutide in Alzheimer’s Disease: Study Protocol for a Randomised Controlled Trial (ELAD Study). Trials 2019, 20, 191. [Google Scholar] [CrossRef]
- Salinas, C.B.G.; Lu, T.T.-H.; Gabery, S.; Marstal, K.; Alanentalo, T.; Mercer, A.J.; Cornea, A.; Conradsen, K.; Hecksher-Sørensen, J.; Dahl, A.B.; et al. Integrated Brain Atlas for Unbiased Mapping of Nervous System Effects Following Liraglutide Treatment. Sci Rep 2018, 8, 10310. [Google Scholar] [CrossRef]
- Reich, N.; Hölscher, C. The Neuroprotective Effects of Glucagon-like Peptide 1 in Alzheimer’s and Parkinson’s Disease: An in-Depth Review. Front. Neurosci. 2022, 16, 970925. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, P.; Wang, W.; Guo, L.; Pan, Q. The Antidepressant Effects of GLP-1 Receptor Agonists: A Systematic Review and Meta-Analysis. Am J Geriatr Psychiatry 2024, 32, 117–127. [Google Scholar] [CrossRef]
- Kanoski, S.E.; Hayes, M.R.; Skibicka, K.P. GLP-1 and Weight Loss: Unraveling the Diverse Neural Circuitry. Am J Physiol Regul Integr Comp Physiol 2016, 310, R885–895. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J. Mechanisms of Action and Therapeutic Application of Glucagon-like Peptide-1. Cell Metabolism 2018, 27, 740–756. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, S.L.; Rørth, R.; Jhund, P.S.; Docherty, K.F.; Sattar, N.; Preiss, D.; Køber, L.; Petrie, M.C.; McMurray, J.J.V. Cardiovascular, Mortality, and Kidney Outcomes with GLP-1 Receptor Agonists in Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis of Cardiovascular Outcome Trials. Lancet Diabetes Endocrinol 2019, 7, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Badulescu, S.; Gill, H.; Shah, H.; Brudner, R.; Phan, L.; Di Vincenzo, J.D.; Tabassum, A.; Armanyous, M.; Llach López, C.-D.; Rosenblat, J.D.; et al. Semaglutide for the Treatment of Cognitive Dysfunction in Major Depressive Disorder: A Double-Blind, Placebo-Controlled, Randomised, Phase 2 Clinical Trial 2025.
Figure 1.
Adiponectin production induced by physical exercise in WAT.
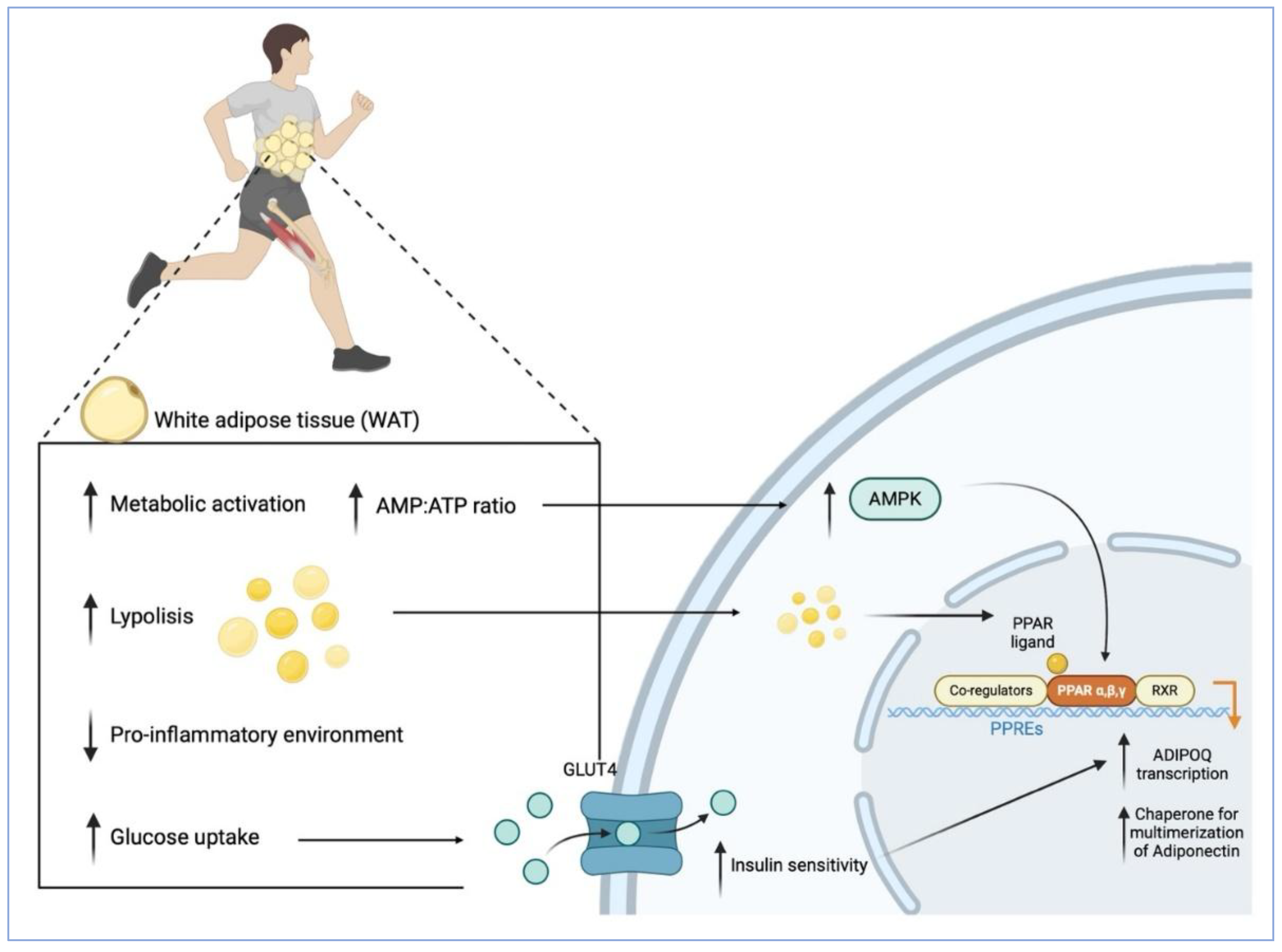
Figure 2.
Adiponectin Signalling Pathways Involved in Neurogenesis and Neuroplasticity.
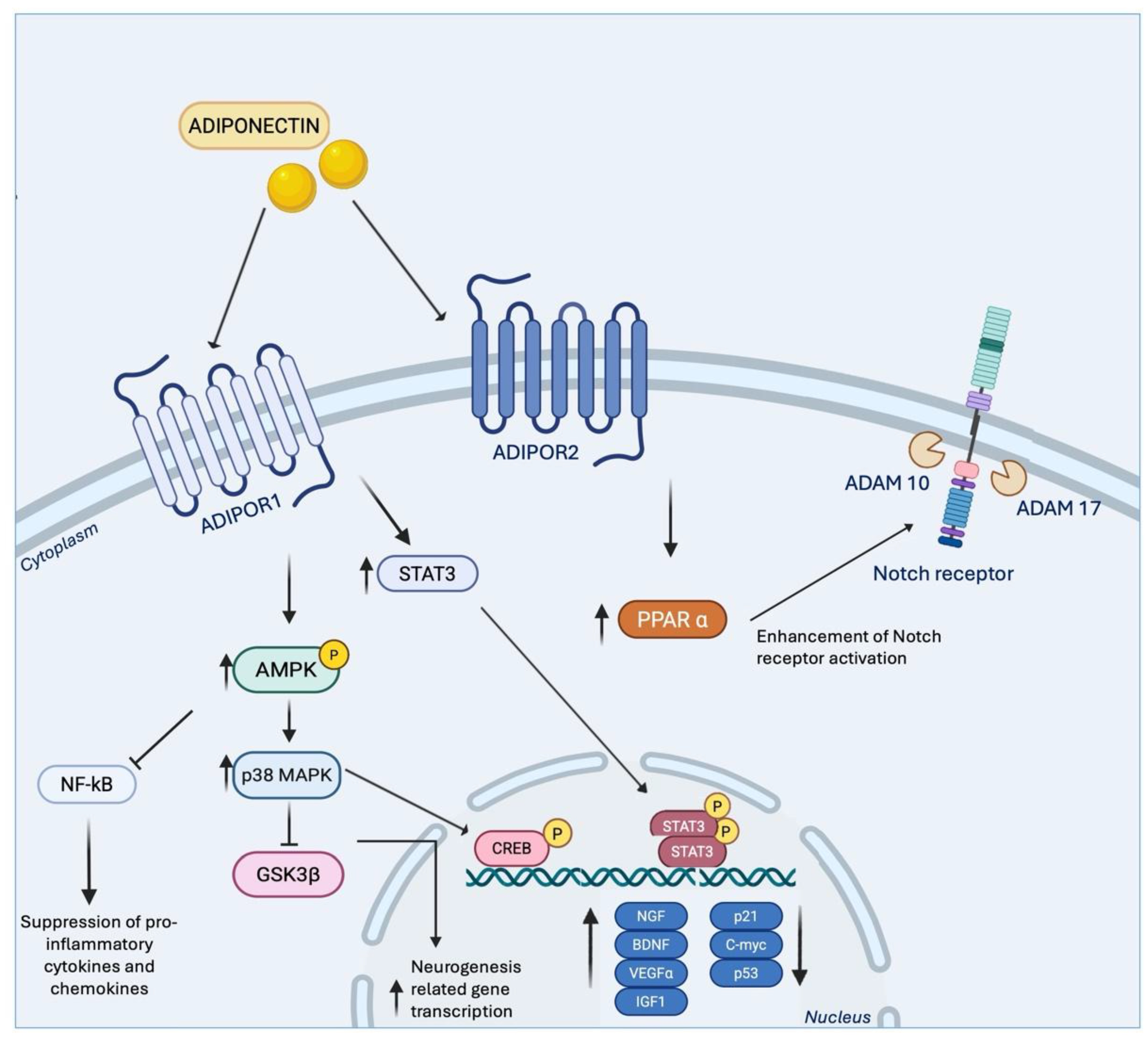
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



