
Submitted:
28 September 2025
Posted:
02 October 2025
You are already at the latest version
Abstract
Background: Migraine is a prevalent neurological condition that is frequently observed in clinical practice and is often associated with insomnia. Insomnia can exacerbate and precipitate migraine attacks, with both conditions exerting a reciprocal influence on one another. cerebellum Crus (Crus) regions are significantly associated with the pathophysiology of migraine and insomnia.
The relationship between cerebellar crus functional alterations and migraine-associated insomnia remains unclear. This study utilizes resting-state functional magnetic resonance imaging (rs-fMRI) to examine functional alterations in the cerebellar crus of patients with migraine and concurrent insomnia.
Methods: Participants underwent resting-state functional magnetic resonance imaging. Subsequently, the disparity in amplitude of low-frequency fluctuations (ALFF) values among groups was analyzed, followed by functional connectivity (FC) investigations employing the cerebellum Crus as seed regions.
Results: Migraine patients frequently experience neuropsychological disorders and insomnia, which are interconnected. Both groups of migraine with insomnia (MwI) and migraine without insomnia (MwoI) exhibit elevated ALFF of the left Crus I and Crus II relative to the healthy controls (HCs) group, with the MwI group showing more pronounced alterations. Furthermore, there is diminished FC between the right Crus I and the left superior parietal lobule (SPL) and inferior parietal lobule (IPL) in both the MwI and MwoI cohorts. A negative correlation was discovered between the ALFF of the left Crus I and Crus II and the Pittsburgh Sleep Quality Index (PSQI) scores in the MwoI group. In the MwoI and MwI cohorts, anomalous FC of the right Crus I with the left SPL and IPL exhibited a positive connection with PSQI scores.
Conclusion: This study identified abnormalities in localized functional activity within the cerebellar crus and in FC with other brain regions among MwI. These findings further clarify the neural mechanisms linking migraines and insomnia, making them useful for identifying neuroimaging biomarkers and exploring therapeutic targets.
Keywords:
migraine
; insomnia
; cerebellum Crus
; ALFF
; functional connectivity
Introduction
Migraine, a persistent neurological condition, affects about one billion individuals globally and ranks as the second greatest cause of disability across all age demographics, with a greater prevalence among women. Migraines often co-occur with other conditions, such as insomnia, which can both trigger and exacerbate migraine attacks. Quality sleep can alleviate migraine episodes. Nonetheless, the neurobiological mechanisms connecting insomnia to migraines are largely unexplored [1].
The resting-state functional magnetic resonance imaging (rs-fMRI) approach has become extensively utilized in the study of migraine and insomnia. Previous studies indicate that the cerebellum contributes critically to pain processing and analgesic pathways in migraines [2,3], participates in the pain matrix network [4,5,6], and serves as a candidate imaging biomarker for chronicity risk [7]. Accumulating evidence indicates that the bilateral posterior cerebellar lobes play an essential part in the migraine pathophysiological mechanisms [9]. Notably, as important regions of the posterior cerebellum, the cerebellum Crus (Crus) have been established as primary inhibitory elements in nociceptive processing [10]. The cerebellum is also closely associated with insomnia. Particularly, the bilateral Crus I and II located in the posterior lobe participate in arousal modulation and sensory prediction through thalamic coupling [8,9,10]. Previous studies implicate the posterior cerebellum, particularly the crus, in the pathogenesis linking migraine and insomnia, suggesting it may represent a shared pathological pathway in individuals with both conditions. However, to date, no relevant neuroimaging evidence has been established.
This study employed amplitude of low-frequency fluctuations (ALFF) to detect local variations in brain functional activity, followed by seed-based functional connectivity (FC) analyses using bilateral cerebellar Crus I and II as seeds to examine differences in intrinsic FC networks among migraine subtypes. We aimed to characterize alterations in cerebellar functional properties in migraineurs with or without insomnia in order to examine their correlation with sleep disturbances.
Methods
Participants
This study recruited fifty-eight patients diagnosed with migraine without aura (MwoA) from the Fifth Affiliated Hospital of Anhui Medical University. The sample was made up of twenty-four migraine with insomnia (MwI), thirty-four migraine with no insomnia (MwoI), and thirty-one healthy controls (HCs) patients. Right-handed individuals of Chinese Han descent underwent neuropsychological assessments and rs-fMRI scans.
The diagnostic criteria for MwoA are derived from the International Classification of Headache Disorders, Third Edition (ICHD-3). Insomnia was identified using the International Classification of Sleep Disorders, third edition (ICSD-3). A PSQI score exceeding 7 indicates symptoms of insomnia. The MwoI group meets the diagnostic criteria for MwoA, while the MwI group requires concurrent diagnoses of MwoA and insomnia. Diagnoses of MwoA were established by two senior attending neurologists in accordance with the ICHD-3. Patients with discrepant assessments were excluded.
Participants underwent neuropsychological assessments (the PHQ-9, PSQI, and GAD-7 scales) before imaging. In addition to these, migraine patients reported the frequency of their headaches (days/month) over the previous three months, the severity of their headaches on the Visual Analog Scale (VAS), the duration of their headaches, and their level of disability using the Migraine Disability Assessment (MIDAS).
Exclusion criteria were (1) history of smoking, alcohol consumption, and coffee drinking. (2) History of neurological and psychiatric disorders other than migraine (3) Use of sedatives or anxiolytic/antidepressant medications within the past week, or prophylactic migraine medication within the past three months. (4) Contraindications to MRI. For HCs, participants were required to have good sleep quality and stable mood, with exclusion criteria identical to those for the MwI and MwoI groups.
Imaging Data
The MRI scans were conducted with a Siemens Verio 3.0-Tesla machine with a 64-channel head coil from Siemens Corporation, Erlangen, Germany. T1-weighted volumes of high resolution were acquired via a gradient-echo sequence with a TR/TE value of 2200/2.46 ms, an 8-degree flip angle, a 230 × 230 mm field of view, a 256 × 256 acquisition matrix, and a 0.9 mm slice thickness without an interslice gap. Each volume comprised 208 contiguous slices and was acquired with a single excitation. Resting-state BOLD data were acquired over 8.17 minutes using echo-planning scans with a TR/TE value of 2000/30 ms, an 80-degree flip angle, a 96×96 acquisition matrix, and a 2 mm slice thickness without interslice gaps. A total of 84 interleaved slices were captured using simultaneous multislice imaging with a multiband factor of 3, resulting in 240 volumes per run. During the exam, participants were instructed to close their eyes, remain awake, and utilize foam earplugs to alleviate discomfort caused by noise.
First Data Processing
Using the rs-fMRI Data Analysis Toolkit Enhanced Version 1.28 (RESTplus 1.28) and the twelfth version of the Statistical Parametric Mapping software (SPM12), imaging data were processed. Data preprocessing includes discarding the initial 10 volumes, correcting motion, normalizing to MNI space using DARTEL, applying linear detrending to eliminate confounding covariates (global signals, white matter, cerebrospinal fluid, and six head motion parameters), filtering in the 0.01–0.08 Hz range, and spatial Gaussian smoothing (FWHM = 6 mm). Subjects exhibiting head movement exceeding 2 millimeters or 2 degrees in any direction were excluded.
The Computation of ALFF
The Fourier transform algorithm is applied to compute the power spectrum’s square root, named the Amplitude of Low Frequency Fluctuation (ALFF), for each voxel within the 0.01–0.08 Hz range. Each voxel's ALFF gets normalized through dividing it by the mean ALFF of the entire brain, producing a normalized ALFF map. For statistical comparison, each subject's ALFF map is converted to z-scores by centering on the whole-brain mean and scaling with the voxel-level whole-brain standard deviation.
FC Analysis from Seeds
After completing ALFF and ReHo analyses, we designated the bilateral Crus I and Crus II of the cerebellum as areas of interest (ROIs). These were independently identified and obtained from the Automated Anatomical Labeling Atlas 3 to produce corresponding binary masks. These masks were subsequently intersected with the whole-brain gray matter mask to confine the analysis to gray matter regions and exclude non-gray matter voxels. The resulting intersection mask ultimately served as the ROIs for subsequent FC analysis. The average time course of the ROI was correlated with that of the remaining brain voxels, followed by correlation analysis. The correlation coefficients were standardized with Fisher's Z transformation.
Statistical Analysis
Data were analyzed utilizing SPSS version 27.0, with a significance level established at p < 0.05. A one-way ANOVA assessed continuous demographic and neuropsychological variables across three groups. Chi-square tests evaluated categorical variables, controlling for age, gender, and education. Partial correlation analysis examined the relationships between headache characteristics and neuropsychological indicators within the migraine group. At the level of individual voxels, ANOVA was conducted in SPM12 to compare ALFF and FC values, with subsequent pairwise post-hoc tests employed to determine differences among each group pair. The analyses took into account gender, age, years of education, and scores related to anxiety and depression for adjustment. Statistical significance was assessed using a voxelwise threshold of p < 0.001, complemented by cluster-level family-wise error (FWE) correction at FWEp < 0.05. Moreover, analyses of partial correlation were carried out to investigate the links between brain dysfunction and clinical traits in migraineurs while adjusting for gender, age, and education (p<0.05). The extraction of signals from atypical brain regions was conducted using the RESTplus V1.28 toolbox. This tool calculates the mean signal within each FWE-corrected cluster and performs a Fisher Z-transformation.
Results
Characteristics of Demographics and Clinical Aspects
Eighty-nine participants meeting inclusion criteria were analyzed (Table 1). When comparing clinical and demographic factors among the three groups, no notable differences in gender, age, or educational achievement were observed; significant discrepancies were noted in PHQ-9, GAD-7, and PSQI scores (p<0.001), with post hoc analysis revealing that the MwI group's scores exceeded those of the other groups. Correlation between migraine characteristics and PHQ-9, GAD-7, and PSQI scores within pooled migraine (MwI and MwoI) patients: In a study involving 58 migraine patients, significant positive correlations were observed: MIDAS scores with headache frequency (r=0.74, p<0.001), attack duration with VAS scores (r=0.41, p=0.002), GAD-7 with PHQ-9 scores (r=0.87, p<0.001), and PSQI with both PHQ-9 (r=0.62, p<0.001) and GAD-7 (r=0.67, p<0.001), as shown in Figure 1.
Abbreviations match those listed in Table 1.
ALFF
A one-way ANOVA indicated a significant difference in ALFF values among groups in the left Crus I and II (F=7.17, p=0.001). Post hoc tests indicated significantly elevated ALFF levels in both the MwoI group (p=0.03) and the MwI group (Bonferroni-corrected p<0.001) relative to healthy controls (Table 2; Figure 2). The MwI group demonstrated a higher level of significance.
FC Analysis from Seeds
FC analysis, focusing on the right Crus I as the ROI, identified differences in the left SPL and IPL. Post-hoc tests revealed that, compared to HCs, the MwI and MwoI groups separately exhibited significantly decreased FC strength (Bonferroni-corrected p<0.001) (Table 2, Figure 3). The FC analyses using the left Crus I, left Crus II, or right Crus I as seed points, respectively, revealed no statistically significant differences.
Correlation Analyses
In the MwoI group, a negative correlation was identified between the ALFF value of Left Crus I and II and the PSQI (r = -0.42, p = 0.02), and in the MwI group, it was identified with PHQ-9 (r = -0.46, p = 0.03) and GAD-7 (r = -0.55, p = 0.008). In the combined migraine group, the FC strength between the right Crus I and the left SPL and left IPL was positively related to PHQ-9 scores (r = 0.29, p = 0.03) and GAD-7 scores (r = 0.27, p = 0.05), and in the MwoI group, it was observed with PHQ-9 (r = 0.41, p = 0.02) and PSQI (r = 0.38, p = 0.03); similarly, in the MwI group, the FC strength correlated positively with MIDAS (r = 0.56, p = 0.007), PHQ-9 (r = 0.47, p = 0.03), GAD-7 (r = 0.50, p = 0.02), and PSQI (r = 0.47, p = 0.03). as shown in Figure 4.
Discussion
This study investigated the FC network characteristics in individuals with MwI and MwoI. Both groups exhibited elevated ALFF values in the cerebellar network (left Crus I and II) compared to HCs, with a more significant increase noted in the MwI group. Additionally, changes in the right Crus I FC network were primarily noted within specific functional networks: the dorsal attention network (DAN), such as the left SPL, and the default mode network (DMN), such as the left IPL. Compared to the HCs group, both the MwI and MwoI groups had diminished FC strength with the left SPL and IPL. Elevated ALFF values in the left Crus I and II are associated with milder neuropsychological and sleep disturbances in migraine patients. Conversely, alterations in the right Crus I FC network reveal that increased FC values in the L-SPL and L-IPL correlate with more severe neuropsychological and sleep disturbances, as well as heightened migraine-related disability.
Among migraine patients, PSQI is positively correlated with both GAD-7 and PHQ-9, the attack duration is positively correlated with VAS, and MIDAS is positively correlated with headache frequency. Migraine patients frequently experience comorbid neuropsychological disorders and insomnia, which can trigger the exacerbation of their symptoms [11]. Therefore, emotional management and sleep improvement are particularly important for individuals with migraines.
Traditionally linked to motor control, the cerebellum is now recognized as integral to migraine-related pain transmission and analgesic pathways. Migraine sufferers show heightened local FC density in the cerebellum, inversely related to pain intensity.
Temporal dynamic irregularities in the cerebellum (mostly in bilateral Crus) among migraine sufferers are uncovered by Dynamic fALFF. These irregularities are linked to the clinical severity, indicating that the cerebellum serves as a crucial modulation hub in the migraine network [5].
The connection strength between the hypothalamus and the cerebellum (primarily in bilateral Crus) can serve as a candidate imaging biomarker for migraine severity and chronicity risk [7]. Fu's study has demonstrated abnormal cerebral perfusion in migraine patients, primarily involving the bilateral Crus, which correlates with clinical indicators (migraine attack frequency, disease duration, and disability severity) [12]. The activity in the Crus is heightened under pain stimulation and exhibits cognitive and emotional features, which are related to pain encoding [13]. The cerebellum predominantly exerts an inhibitory effect on nociception. The activation of the Crus is more pronounced in migraine patients compared to control subjects, serving to sustain its inhibitory function [14]. The aforementioned studies all support the notion that the cerebellum, particularly the Crus region, possesses a pain regulation mechanism in migraines.
The cerebellum is involved in sleep regulation by modulating monoaminergic neurotransmission and is closely connected to the sleep-wake cycle. Among individuals with chronic insomnia, the FC between the ascending arousal network and cerebellar networks, as well as the internal connections within the cerebellar networks, is enhanced [15,16]. Cerebellar lobules, mostly located in the posterior lobe (especially in Crus, VI, VIIb, and VIII) or the vermis, participate in arousal regulation and sensory prediction through coupling with the thalamus [8]. Cerebellar gray matter volume, especially in the posterior lobe, changes gradually as insomnia severity increases. Increased gray matter in the right Crus II may represent a compensatory mechanism [9]. The bilateral Crus exhibits significant alterations in FC with the frontoparietal network following sleep deprivation [10]. The bilateral cerebellar Crus serves as a crucial network node for motor interventions aimed at improving sleep [17].
Our study found heightened local brain activity in the left Crus among migraine patients, with ALFF values negatively correlating with anxiety, depression, and sleep disorder scale scores. This data supports the modulatory role of the cerebellum, specifically Crus, in migraineurs experiencing insomnia. However, this regulatory mechanism is impaired. Abnormal functional activity in the left Crus is a key pathological mechanism shared by migraine patients with insomnia, highlighting the cerebellar crus as a crucial focus in brain functional studies of migraine.
The SPL is a core component of the dorsal attention network, participating in spatial attention, sensory-motor integration, switching between internal and external attention, and the pointing and maintenance of working memory. It serves top-down control functions, and in migraine patients, abnormal brain activity is observed in the SPL during the processing of negative emotions [18]. The IPL's posterior region, overlapping with the default mode network, acts as an aspect in attention reorientation, context updating, and semantic and emotional processing. Within the DMN, the IPL integrates intense headache signals into the insula's pain processing and interoceptive network during migraines, aiming to mitigate pain perception via cognitive strategies. The SPL and IPL participate in pain perception and pain processing, and the weakening of their functional connections may reflect abnormalities in pain perception and attention in migraine patients [19]. Insufficient top-down inhibition of the SPL/IPL leads to difficulty in falling asleep at night and easy awakening [20]. Insomnia causes structural alterations in the SPL/IPL, impairing functions such as sensorimotor coordination, spatial cognition, attention, working memory, and emotional regulation in the parietal lobe [21]. The cerebellum can regulate the function of the DMN by way of its associations with the prefrontal and parietal cortex, indirectly affecting the overall function and integrity of the DMN. If the cerebellum's function or structure is impaired, it may not be able to regulate or coordinate the activity of the DMN effectively, thereby exacerbating the abnormal pain processing and stress response of the DMN in migraines [3,19].
Our study identifies diminished FC with the left parietal lobe in individuals suffering migraines compared to the HCs, and the FC strength correlates positively with sleep disorders, anxiety, depression, and disability in migraine sufferers. These findings indicate that the FC network between the cerebellar Crus and the DMN and DAN is imbalanced and reorganized in individuals with comorbid migraine and insomnia, and that the top-down regulation mechanism of migraines is compromised. The altered FC with the left parietal lobe may serve as a biomarker for individuals suffering from both migraine and insomnia.
Even though the present research offers neuroimaging proof of regional cerebellar functional irregularities and modified cerebellar FC in those with co-occurring migraine and insomnia, it has a few limitations. The cross-sectional design of this investigation complicates the establishment of a causal link among migraine, changes in FC networks, and local cerebellar dysfunction. In the future, additional longitudinal investigations will be required to verify this. Secondly, the limited sample size led to the absence of some relevant capabilities. Research in the future ought to concentrate on increasing the number of samples. Thirdly, the evaluation of the sleep scale is subjective, and this may lead to grouping bias. In future research, objective methods for measuring insomnia, like polysomnography, ought to be utilized. Finally, the group suffering from migraines was not differentiated further based on subtype or the period of onset. Subsequent analyses ought to take these elements into account.
Conclusions
Our research has identified abnormalities in the local functional activity within the cerebellar crus region and aberrant FC networks in migraine patients with comorbid insomnia. This discovery offers novel neuroimaging biomarkers for elucidating the mechanisms underlying migraine with comorbid insomnia. Abnormal localized activity in the cerebellar Crus, along with its aberrant FC to the left SPL and IPL, is associated with anxiety, depression, sleep disturbances, and disability in migraine patients with comorbid insomnia. These findings could be valuable for identifying therapeutic targets for migraine-associated neuropsychiatric disorders and insomnia in future research.
Funding
This study was funded by the Anhui Provincial Department of Education, Key Natural Science Research Project of Higher Education Institutions (2023AH053157).
References
- Nahman-Averbuch H, Schneider VJ, Lee GR, et al. New insight into the neural mechanisms of migraine in adolescents: Relationships with sleep. Headache: The Journal of Head and Face Pain 2022;62(6):668-80. [CrossRef]
- Wei W, Xueyan Z, Xiaoyan B, et al. Gamma-aminobutyric acid and glutamate/glutamine levels in the dentate nucleus and periaqueductal gray with episodic and chronic migraine: a proton magnetic resonance spectroscopy study. J Headache Pain 2022;23(1). [CrossRef]
- Dóra D, Gyöngyi K, Gyula G, et al. Microstructural differences in migraine: A diffusion-tensor imaging study. Cephalalgia 2023;43(12). [CrossRef]
- Mi Ji L, Bo-Yong P, Soohyun C, et al. Increased connectivity of pain matrix in chronic migraine: a resting-state functional MRI study. J Headache Pain 2019;20(1). [CrossRef]
- Wei C, Hui W, Tianze S, et al. Dynamic changes in fractional amplitude of low-frequency fluctuations in patients with chronic insomnia. [CrossRef]
- Vasilis S V, Nikos K, Yiolanda C, et al. Neural correlates of pain acceptance and the role of the cerebellum: Functional connectivity and anatomical differences in individuals with headaches versus matched controls. Eur J Pain 2024;29(3). [CrossRef]
- Roberta M, Maria A R, Paola V, et al. Clinical correlates of hypothalamic functional changes in migraine patients. Cephalalgia 2021;42(0). [CrossRef]
- Yupeng G, Yun C, Yan S, et al. Thalamic network under wakefulness after sleep onset and its coupling with daytime fatigue in insomnia disorder: An EEG-fMRI study. J Affect Disord 2023;334(0). [CrossRef]
- Shumei L, Bin A W, Cheng L, et al. Progressive gray matter hypertrophy with severity stages of insomnia disorder and its relevance for mood symptoms. Eur Radiol 2021;31(8). [CrossRef]
- Rong-Jun N, Yi-Yan W, Wen-Jun P, et al. Differential effects of sleep deprivation on behavior and microglia in a brain-region-specific manner in young and aged male mice. Brain Behav Immun 2024;117(0). [CrossRef]
- Christian L, Hallie T, Cristina T, et al. Headache, depression and anxiety: associations in the Eurolight project. J Headache Pain 2016;17(0). [CrossRef]
- Tong F, Lindong L, Xiaobin H, et al. Cerebral blood flow alterations in migraine patients with and without aura: An arterial spin labeling study. J Headache Pain 2022;23(1). [CrossRef]
- Eric A M, Igor E, Gautam P, et al. Aversion-related circuitry in the cerebellum: responses to noxious heat and unpleasant images. J Neurosci 2011;31(10). [CrossRef]
- Jan M, Arne M. Functional and structural alterations in the migraine cerebellum. J Cereb Blood Flow Metab 2017;39(4). [CrossRef]
- Liang G, Kewu H, Fang C, et al. The role of ascending arousal network in patients with chronic insomnia disorder. Hum Brain Mapp 2022;44(2). [CrossRef]
- Shiqi L, Xi Y, Yuping Y, et al. Enhanced functional connectome of cerebellum in chronic insomnia patients. Brain Behav 2023;13(7). [CrossRef]
- Rongrong C, Shilei W, Qinzi H, et al. Exercise intervention in middle-aged and elderly individuals with insomnia improves sleep and restores connectivity in the motor network. Transl Psychiatry 2024;14(1). [CrossRef]
- Keva K, Joshua C, Christina H, et al. Differential fMRI neural synchrony associated with migraine during naturalistic stimuli with negative emotional valence. J Headache Pain 2025;26(1). [CrossRef]
- Faisal Mohammad A, Anders H, Stefano M, et al. Change in brain network connectivity during PACAP38-induced migraine attacks: A resting-state functional MRI study. Neurology 2015;86(2). [CrossRef]
- Yongli L, Enfeng W, Hongju Z, et al. Functional connectivity changes between parietal and prefrontal cortices in primary insomnia patients: evidence from resting-state fMRI. Eur J Med Res 2014;19(1). [CrossRef]
- Xiang G, Tao W, Shenglong X, et al. Sleep disorders causally affect the brain cortical structure: A Mendelian randomization study. Sleep Med 2023;110(0). [CrossRef]
Figure 1.
The correlation matrix of clinical features in the combined migraine groups.
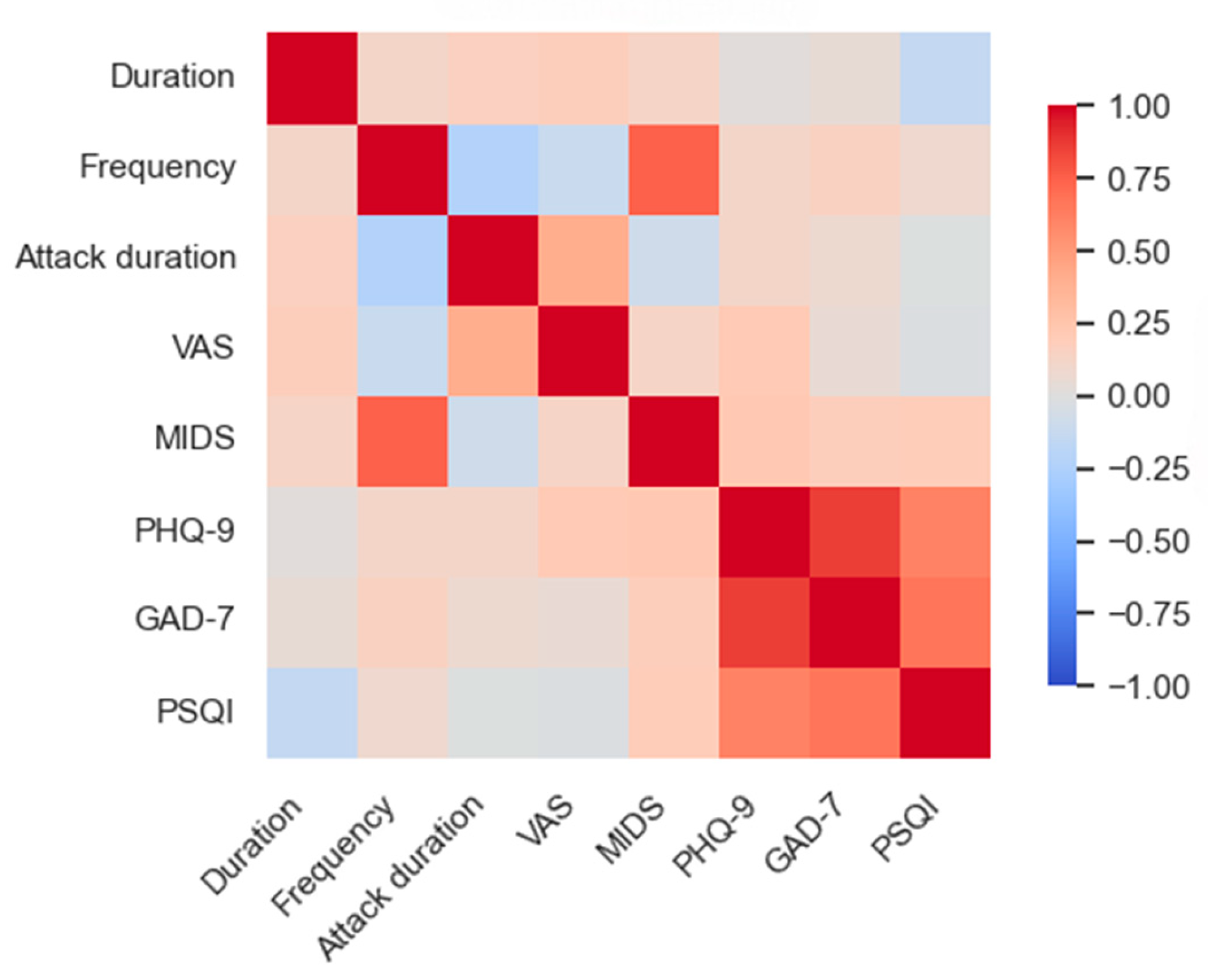
Figure 2.
ALFF among the three groups. The brain map (a) reveals differences in ALFF (red) within the left Crus I and II, and the violin plots (b) display their ALFF values across the three groups, with * * indicating p < 0.001 (Bonferroni correction). Abbreviations match those listed in Table 1 and 2.
Figure 2.
ALFF among the three groups. The brain map (a) reveals differences in ALFF (red) within the left Crus I and II, and the violin plots (b) display their ALFF values across the three groups, with * * indicating p < 0.001 (Bonferroni correction). Abbreviations match those listed in Table 1 and 2.
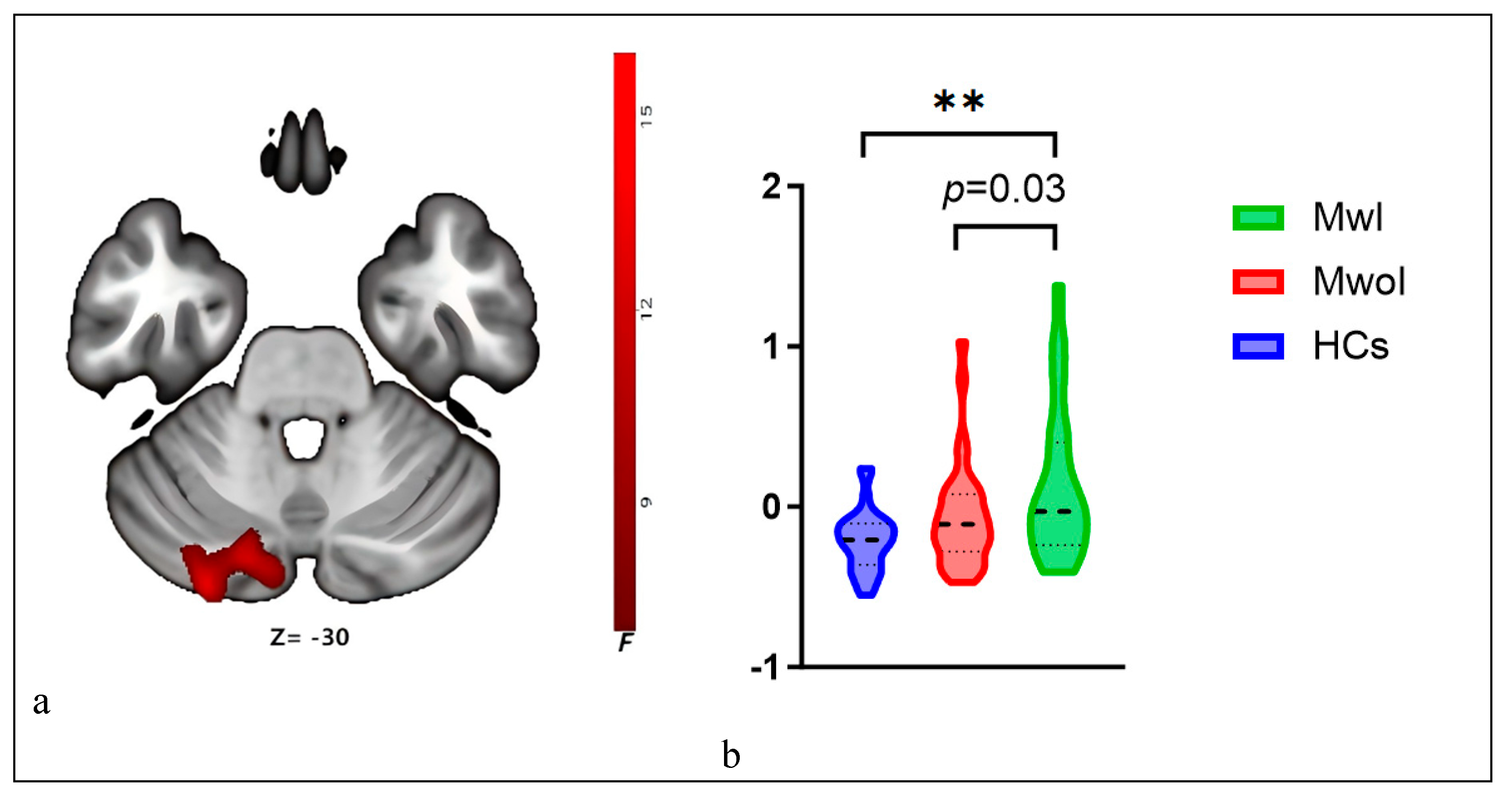
Figure 3.
The right CrusI FC network exhibits differences across three groups. a. The brain map reveals differences in the right Crus I FC (red) with the left SPL and IPL. b. The violin plots display its FC values across the three groups, with * * indicating p < 0.001 (Bonferroni correction). Abbreviations match those listed in Table 1 and Table 2.
Figure 3.
The right CrusI FC network exhibits differences across three groups. a. The brain map reveals differences in the right Crus I FC (red) with the left SPL and IPL. b. The violin plots display its FC values across the three groups, with * * indicating p < 0.001 (Bonferroni correction). Abbreviations match those listed in Table 1 and Table 2.
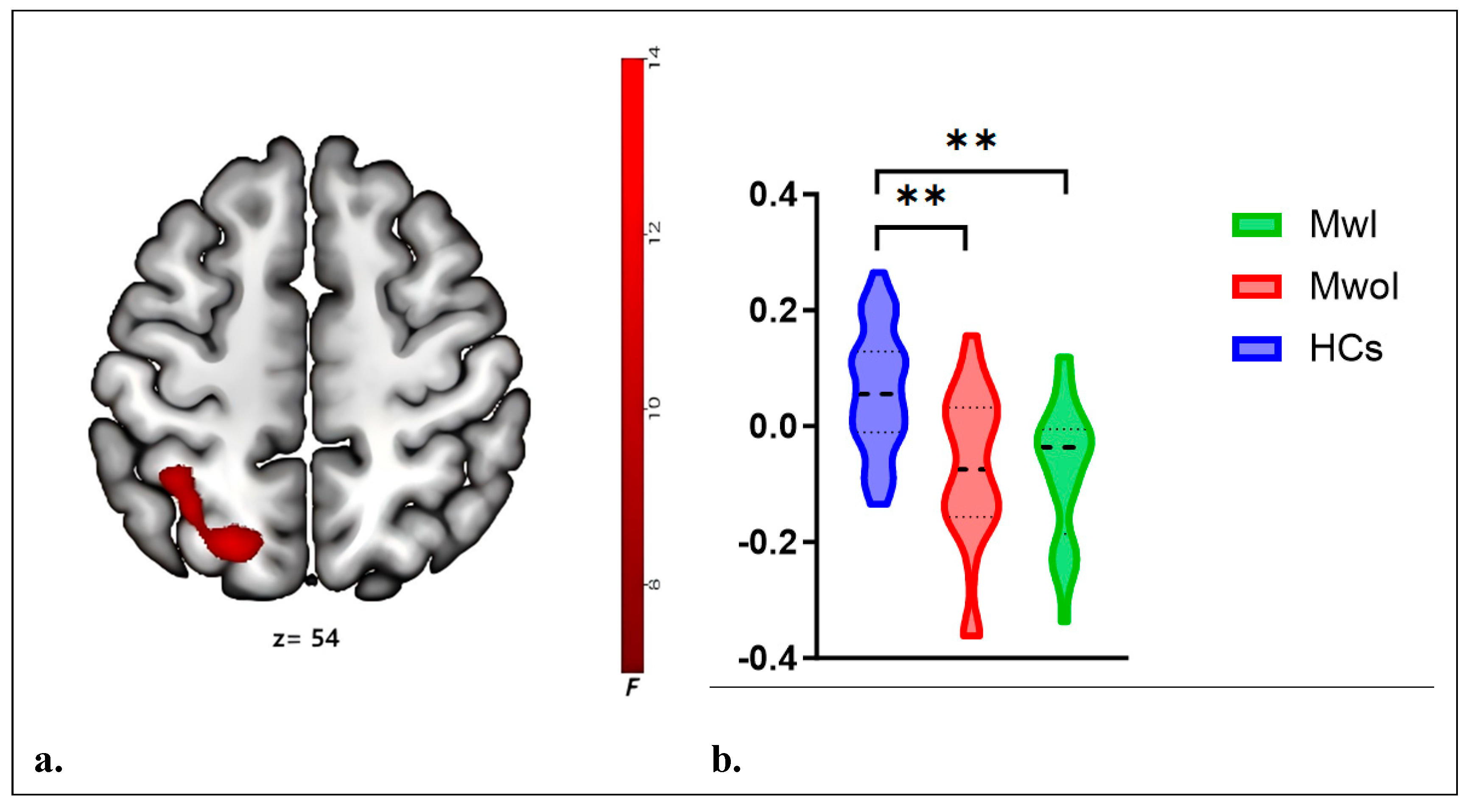
Figure 4.
Relationship between the ALFF values of left Crus I and left Crus II, as well as the FC strength in right Crus I, and clinical characteristics in migraine patients. The ALFF value had a negative relationship with the PSQI in the MwoI group (a) and with the PHQ-9 and GAD-7 in the MwI group (b). The FC strength with the left SPL and IPL was positively related with the PHQ-9 and GAD-7 in the pooled migraine group (c), with the PHQ-9 and the PSQI in the MwoI group (d), and with the MIDAS, PHQ-9, PSQI, and GAD-7 in the MwI group (e). Abbreviations match those listed in Table 1 and Table 2.
Figure 4.
Relationship between the ALFF values of left Crus I and left Crus II, as well as the FC strength in right Crus I, and clinical characteristics in migraine patients. The ALFF value had a negative relationship with the PSQI in the MwoI group (a) and with the PHQ-9 and GAD-7 in the MwI group (b). The FC strength with the left SPL and IPL was positively related with the PHQ-9 and GAD-7 in the pooled migraine group (c), with the PHQ-9 and the PSQI in the MwoI group (d), and with the MIDAS, PHQ-9, PSQI, and GAD-7 in the MwI group (e). Abbreviations match those listed in Table 1 and Table 2.
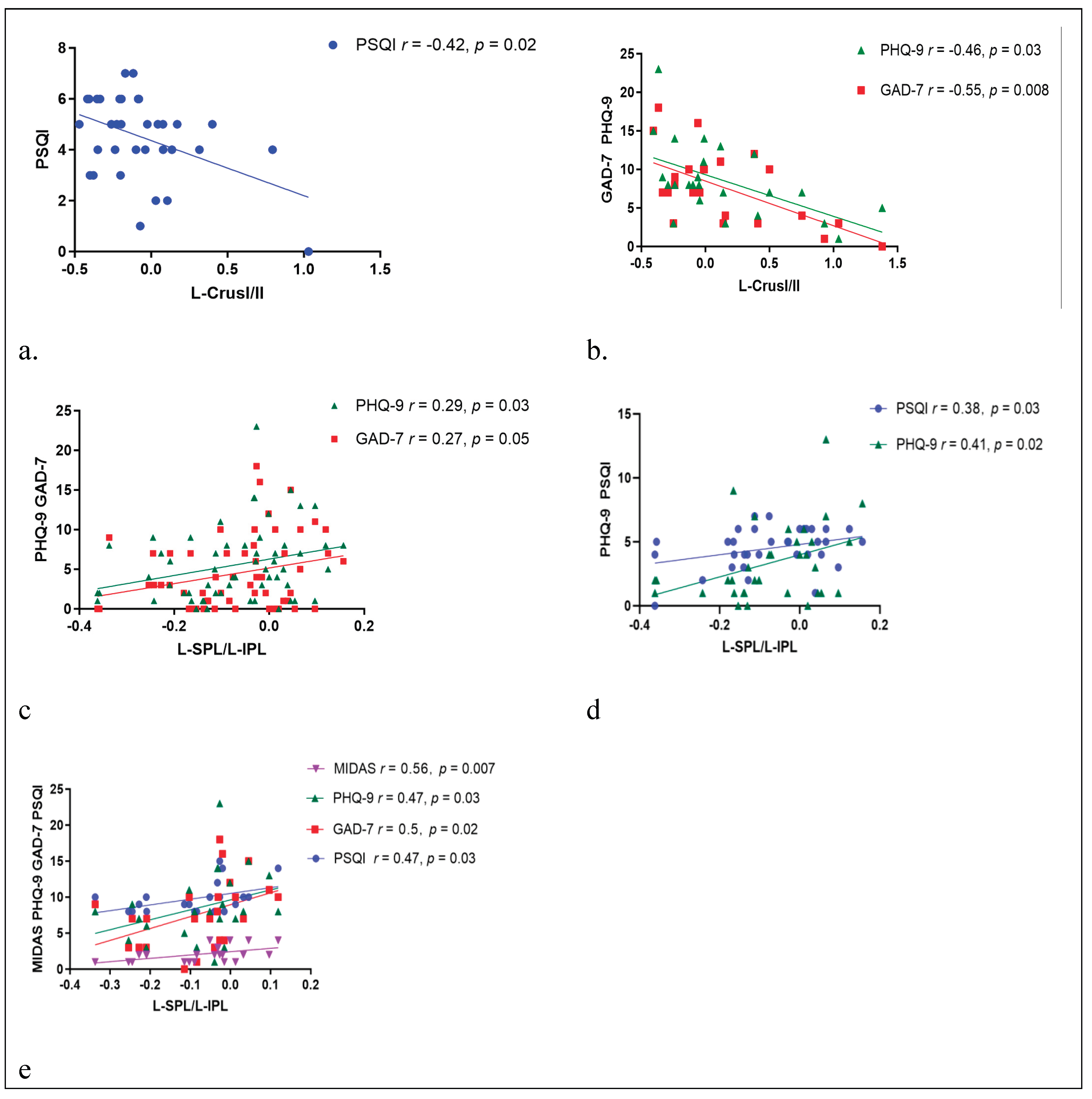

Table 1.
Clinical characteristic and demographics of all participants.
| HCs | MwI | MwoI | F/T/X2 | p-value | |
| Education(years) | 13.16±2.45 | 12.47±2.47 | 11.96±2.55 | 1.64 | 0.2 |
| Gender(M/F) | 28/3 | 31/3 | 24/0 | 2.39 | 0.3 |
| Age | 32.55±6.56 | 31.5±6.57 | 33.04±5.85 | 0.452 | 0.64 |
| Duration(years) | / | 6.53±6.11 | 5.83±3.8 | 0.493 | 0.21 |
| Frequency(Days/Month) | / | 4.06±3.92 | 4.5±4.68 | -0.39 | 0.43 |
| Attack duration(Hours/Time) | / | 15.82±19.69 | 15.67±15.66 | 0.03 | 0.55 |
| VAS | / | 5.88±1.25 | 6±1.77 | -0.3 | 0.22 |
| MIDAS | / | 1.91±0.97 | 2.08±1.06 | -0.64 | 0.81 |
| PHQ-9 | 1.48±0.89 | 3.35±2.97 | 8.58±4.87 | 35.62 | <0.001 |
| GAD-7 | 1.29±0.64 | 2.12±2.63 | 7.71±4.68 | 36.99 | <0.001 |
| PSQI | 4±1.29 | 4.5±1.62 | 9.92±2.1 | 101.87 | <0.001 |
Abbreviations: MwI: migraine with insomnia, MwoI: migraine with no insomnia, HCs: healthy controls. VAS: Visual Analogue Scale; PSQI: Pittsburgh Sleep Quality Index; PHQ-9: Patient Health Questionnaire-9; GAD-7: Generalized Anxiety Disorder-7.
Table 2.
group differences in ALFF and FC network .
| Brain regions within cluster | MNIcoordinate | |||||
| Cluster size | X | Y | Z | Peak F Score | ||
| ALFF |
Left CrusI/II |
86 | -27 | -87 | -30 | 14.27 |
| FC |
Left SPL/IPL |
148 | -33 | -57 | 54 | 11.77 |
Abbreviations: ALFF: amplitude of low-frequency fluctuations; FC: functional connectivity; Crus: cerebellar Crus. SPL: Superior Parietal Lobule; IPL: Inferior Parietal Lobule.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



