Submitted:
01 October 2025
Posted:
01 October 2025
You are already at the latest version
Abstract
Hantaviridae (order Bunyavirales) is a family of negative-sense, single-stranded RNA viruses. To date, several genetically distinct hantaviruses have been found in the same species of shrews and moles. In this report, we describe Biya River virus (BIRV), a novel hantavirus detected in the Eurasian water shrew (Neomys fodiens), the principal host of Boginia virus (BOGV). Genetic analysis of the complete L-, M-, and partial S-genomic segments showed that BIRV shared a common evolutionary origin with shrew-borne Altai (ALTV) and Lena (LENV) viruses, belonging to the Mobatvirus genus, and was distantly related to BOGV and other shrew- and mole-borne orthohantaviruses. Ancient cross-species transmission of hantaviruses, with subsequent diversification within the Soricinae subfamily in Eurasia might have shaped the evolutionary history of BIRV, ALTV and LENV.
Keywords:
Hantaviridae
; hantavirus
; mobatvirus
; shrew
; evolution
; Russia
1. Introduction
Hantaviridae (order Bunyavirales), a family of negative-sense single-stranded RNA viruses, has a genome comprising small (S), medium (M), and large (L) segments, which encode a nucleocapsid (N) and occasionally nonstructural (NSs) protein, envelope glycoproteins (Gn and Gc), and an RNA-dependent RNA polymerase (RdRP), respectively. More than 47 distinct species of hantaviruses identified in rodents, moles, shrews, and bats are classified into one of four genera (Loanvirus, Mobatvirus, Orthohantavirus and Thottimvirus) [1].
The human pathogens are rodent-associated hantaviruses (genus Orthohantavirus) [2]. Initial hypothesis of the co-evolution of rodent-borne hantaviruses and their hosts have been changed after discovery of hantaviruses in shrews (family Soricidae), moles (family Talpidae) and bats (order Chiroptera) [3,4]. The evolutionary history of hantaviruses in moles, shrews, and bats is much more complex and includes cases of interspecies transitions and genome reassortment [5,6,7,8,9]. Serological evidence of human infection has been obtained for several hantaviruses from insectivores. Thus, specific antibodies have been identified in residents of Africa (Gabon and Côte d’Ivoire) to the Bowe virus (BOWV) from Thereses`s shrew (Crocidura theresae) and the Uluguru virus (ULUV) from Geata mouse shrew (Myosorex geata) [10]. In Asia (Thailand, Sri Lanka), antibodies to hantaviruses from insectivores, Altai virus (ALTV) and Thottapalayam virus (TPMV) have been detected in humans [11].
Studies of soricid- and talpid-borne hantaviruses have identified several pairs of significantly different hantaviruses that share a natural host. The pair of such viruses was discovered in the common shrews (Sorex araneus), natural carrier of the Seewis virus (SWSV), which is widespread in the habitat of the species, from Western Europe to the Baikal region [12,13,14]. A new virus, named Altai (ALTV), was found co-circulating with SWSV in the same host species in Western Siberia and in European countries [6,15]. The other pair of hantaviruses was identified in European moles (Talpa europaea), originally known as the natural hosts of Nova virus (NVAV) [16]. A second hantavirus, named Bruges virus (BRGV), found in the same host and in the same habitats in European countries [17]. And the Iberian mole (Talpa occidentalis) has been shown to harbor NVAV, BRGV and Asturias virus (ASTV) in northwestern Spain [18]. The pair, Lena virus (LENV) and Artybash variant (ARTV) of SWSV, was identified among Laxman’s shrews (S. caecutiens) in the Far Eastern and Siberian Russia [19]. These findings highlight the complexity of hantavirus-reservoir relationships. ALTV and LENV viruses are members of the genus Mobatvirus, which includes most of hantaviruses associated with bats; their pairs, SWSV and ARTV, belong to the genus Orthohantavirus, which includes all other hantaviruses carried by multiple species of shrews. Based on this data, a hypothesis that they originate as a result of interspecies transfer has been proposed [6,19].
In Russia, studies of hantaviruses carried by insectivores have been conducted mainly in Siberia and the Far East. Seven species of hantaviruses were identified, five of which were discovered for the first time and were traditionally named after the place where they were discovered. This list includes viruses Kenkeme (KKMV), ARTV, ALTV, LENV, Academ (ACDV) [6,15,19,20,21]. Hantaviruses SWSV and Yakeshi (YKSV), circulating in Russia, were previously identified in neighboring countries [13,22]. In the European part of the country one of the suspected new hantaviruses has been discovered. Artesian virus (ATSV) was identified in the lesser shrew (C. suaveolens) and was most closely related to viruses found in other Crocidura species from Africa [23].
Boginia virus (BOGV) has been identified among Eurasian water shrews (N. fodiens) in Poland and Finland [5,24]. N. fodiens inhabits the forest zone from Western Europe to the Pacific coast. The presence of hantaviral antigens in the Eurasian water shrew was reported in the European part of Russia [25]. In this study we screened N. fodiens captured in Western Siberia for hantaviruses. We present the genetic and phylogenetic analyses of of highly distinct hantaviruses harbored by the same host, suggesting both co-evolution and species-shift in their evolution history.
2. Materials and Methods
2.1. Trapping and Sample Collection
During August 2019-2020 and September 2022, shrews were trapped in the Altai Republic, and during June to August 2021–2025 in the Kemerovo Oblast of Western Siberia, Russia. The study was conducted in accordance with the Declaration of Helsinki and met the guideline requirements of the order of the Russian High and Middle Education Ministry (No. 742 issued on 13 November, 1984) and by the Federal Law of the Russian Federation (No. 498-FZ issued on 19 December, 2018). Field procedures and protocols were approved by the Institutional Animal Care and Use Committee of the Institute of Systematics and Ecology of Animals (Protocols 2020-02, 2021-01). The study did not involve endangered or protected species.
Collection sites (Figure 1) in the Altai Republic were located near Teletskoye Lake (51.79424 N/87.30447 E), and around the settlement Azhendarovo (54.76237 N/87.03094 E and 54.74537 N/87.02093 E) in the Kemerovo Oblast. Lung samples were collected aseptically and stored in RNAprotect® (Qiagen, Hilden, Germany) before analysis.
2.2. RNA Extraction and RT-PCR Analysis
Total RNA was extracted from lung tissues, using the RNeasy MiniKit (Qiagen, Hilden, Germany), then reverse transcribed, using the Expand reverse transcriptase (Roche, Basel, Switzerland) with universal oligonucleotide primer (OSM55, 5`–TAGTAGTAGACTCC–3`), designed from the conserved 3` end of the S-, M-, and L-segments of hantaviruses. For initial screening by nested RT-PCR, previously described genus-specific oligonucleotide primers targeting the partial L-segment sequence were used [26]. Oligonucleotide primer sequences for PCR, specific for BIRV were designed from consensus regions of ALTV, LENV and other hantaviruses. Partial S-segment was amplified using nested PCR with primers SAF20: 5`–TCNGGNGCHCHNGCAAANAHCCA–3` and S974R: 5`–TCNGGNGCHCHNGCAAANAHCCA–3`; then SAF1: 5`–GGAGCAYAAAGGRAATAGGA –3` and S974R. Short-size amplicons were separated by electrophoresis on 1.2% agarose gels and purified using the QIAQuick Gel Extraction Kit (Qiagen, Hilden, Germany). DNA was sequenced directly using an ABI Prism 310 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA).
Primers used for PCR amplification of the complete M-segment were M1F: 5`–TAGTAGTAGACTCCGCAARAA–3`; L-segment was amplified as two overlapping amplicons, L5-1: 5`–TTCTGCAGTAGTAGTAGACTCCG–3` and LR1: 5`-AACADTCWGTYCCRTCATC–3`; LF2: 5`–TGCWGATGCHACIAARTGGTC–3`and LBR3: 5`–CTGCAGTAGTAGTAKRCTCCGG–3`. Long-size amplicons were purified using the Mini Elute PCR Purification Kit (Qiagen, Hilden, Germany) and were used for libraries preparation with a NEBNext® Ultra™ II FS DNA Library Prep with Sample Purification Beads (NEB, Hitchin, Great Britain). Deep sequencing of libraries covering complete M- and L-segments of genome was performed on an Illumina MiSeq using the MiSeq reagent kit v3 (Illumina, San Diego, CA, USA). The full-length genomes were assembled de novo using MIRA assembly (version 4.9.6) and by the alignment of reads to known references with bwa-0.7.15 [27].
2.3. Genetic and Phylogenetic Analysis
Pairwise alignment and comparison of full-length coding regions of the M-, L-, and partial S-, and L-segment nucleotide and amino acid sequences of hantaviruses from N. fodiens, captured in Russia, with representative shrew-, mole-, rodent-, and bat-borne hantaviruses were performed, using the MUSCLE in MEGA version 11 [28]. Phylogenetic trees were generated using the Maximum Likelihood approach with the MEGA, under the best-fit general time-reversible model of nucleotide evolution with gamma-distributed rate heterogeneity and invariable sites (GTR + I + Г).
3. Results
3.1. Genetic Analysis
During 2019–2025, 64 Eurasian water shrews (Neomys fodiens), were captured at two localities in Western Siberia (Figure 1). N. fodiens were absent among shrews captured in locality Teletskoye in 2020 and 2022. RNAprotect®-preserved lung specimens were initially analyzed for hantavirus RNA by nested RT-PCR using oligonucleotide primers directed at the RdRp gene. Hantaviral RNA was detected in lung tissues from four of 64 Eurasian water shrews (Table 1).
Genetic analysis of the partial L-segment sequences showed that two significantly different hantaviruses were detected in N. fodiens (3 and 1 positive/64 tested). One of these viruses, detected in three Eurasian water shrews captured at the locality Azhendarovo in Kemerovo Oblast, was BOGV virus, previously found in the same species in Poland and Finland [5,24]. The 346-nucleotide fragments of the L-segment of BOGV showed 82.7–85.0% nucleotide (98.3% amino acid) similarity with BOGV strain 2074 from Poland. The genetic divergence between Siberian BOGV strains was 3.4–8.1% for nucleotide, while amino acid sequences were identical.
A separate genetically distinct hantavirus was identified in one of two Neomys fodiens, captured near the source of the Biya River, locality Teletskoye Lake, Altai Republic. This new hantavirus was designated Biya River (BIRV) according to the capture site. The difference of the partial L-segment nucleotide and amino acid sequences from other known hantaviruses exceeded 21.8% and 11.0%, divergence between BIRV and the new BOGV strains from Siberia was 28.9–33.3% and 33.1%, respectively. The BIRV-positive Eurasian water shrew sample was subjected to full-genome sequencing. The full-length M- and L-segments of the genome were obtained; for the S segment, only a 485-nucleotide fragment of the N protein gene was obtained.
The partial S-segment of BIRV (prototype strain Biya-Nf215/Russia/2019) displayed considerable divergence (>45.6% nucleotide and >23.9% amino acid) to other hantaviruses and was most closely related to strain HV/SC/C7-49.2/2022, recently discovered from the De Winton’s shrew (Chodsigoa hypsibia) in China [29].
The complete 3712-nucleotide M segment of BIRV contained a single ORF (positions 101–3523) encoding the 1140-amino acid glycoprotein precursor of the Gn and Gc glycoproteins, separated by a WAATA pentapeptide at position 649–653. The same motif was found in hantavirus HV/SC/C7-49.2/2022. Analysis of the complete M-coding sequence revealed more than 26.9% nucleotide and 22.6% amino acid sequence differences between BIRV and the most closely related HV/SC/C7-49.2/2022 and considerable divergence from other representative hantaviruses both at the nucleotide (>32.4%) and amino acid (>35.4%) levels. The M-segment sequence of BIRV showed 59.6% amino acid divergence from the partial 795-nucleotide M-segment sequence of BOGV (strain 2074), available in GenBank.
The 6537-nucleotide L segment of BIRV encoded an RdRp protein of 2146 amino acids in length. Pairwise alignment and comparison of the BIRV L segment with representative hantaviruses belonging to the four genera of the Mammantavirinae subfamily showed considerable divergence, ranging between 23.7–30.7% and 12.6–36% at the nucleotide and amino acid level, respectively.
The level of difference of the amino acid sequences of the partial N protein and GP, encoded by the S and M segments from other hantavirus species was more than 23.9%, and 22.6%, respectively, which meets the criteria for a new species and confirms the novelty of BIRV.
3.2. Phylogenetic Analysis
Phylogenetic trees, based on the coding regions of the full-length M-, L-, and partial S- and L-segments, were constructed using maximum-likelihood methods. In the tree, based on the partial L-segment, the viral sequences recovered from Eurasian water shrews were placed into two genera (Figure 2A). BOGV strains from Siberia and Europe occupied a separate branch within the genus Orthohantavirus, and were grouped according to the geographical principle. The BIRV sequence was most closely related to strain HV/SC/C7-49.2/2022 from China and grouped with ALTV and LENV, previously identified in Sorex shrews sampled in northern Eurasia. The strain Biya-Nf215/Russia/2019 was most closely related to members of the Mobatvirus genus in the S, M, and L phylogenetic trees (Figure 2).
4. Discussion
Here, we describe a new hantavirus, named BIRV, in the Eurasian water shrew, previously recognized as the reservoir host of BOGV, captured in locality Teletskoye Lake, Altai Republic. A new pair of highly divergent hantaviruses that have a common host was identified in Western Siberia. These hantaviruses were found at two different localities, 350 km apart. BOGV sequences were detected in N. fodiens, captured at a second locality, Azhendarovo in Kemerovo Oblast. Both capture sites represent the foothill areas of Altai and Kuznetsk Alatau, respectively. Previously published data demonstrate significant level of divergence in the genomes of hantaviruses detected in these foothill regions. Thus, a high level of partial L-segment divergence (up to 14.9% nucleotide) in each of the sites was established for the ACDV virus circulating among Siberian moles [30]. Similar, high divergence (up to 7.7% nucleotide) was shown for SWSV L-sequences from Teletskoye Lake [13]. The exact trapping sites of BOGV in Azhendarovo were within 2 km, but partial L-segment sequences also demonstrated high divergences (up to 8.1%). Our data of L-segment phylogeny suggest that there are two sub-lineages of BOGV in N. fodiens from Azhendarovo. This finding supports our previous suggestion that foothill areas were colonized by several different routes of shrews from separate refugia [13].
The present-day distribution of mammals in northern Eurasia is largely the result of recolonization and dispersal of species in the period after the last glaciation [31]. In response to climate change, some mammal species have shifted their ranges in search of suitable conditions, persisting in large or small refugia [32,33]. The foothill regions of Altai and Kuznetsk Alatau are zones of secondary contacts of separately evolving lineages of moles and shrews and associated hantaviruses from different refugia [13,30].
The Eurasian water shrew, like many species of shrews, is widespread, but the animals prefer near-water biotopes. This is the reason for the noticeable difference in the number of animals caught in the two localities. In the Teletskoye Lake area, a relatively high number of water shrews was observed along the shoreline, while in the meadow clearings among the forest, where the trapping site is located, the number of N. fodiens was comparatively low. In the community of eight species of shrews, S. araneus, S. tundrensis, S. isodon, S. caecutiens, S. minutus, S. minutissimus, N. fodiens, C. sibirica, inhabiting this area, the predominant species is S. araneus (70.4%), while the proportion of the Eurasian water shrews is only 3.5% [34]. In 2019, 48 shrews including two N. fodiens (4.2%) were captured at this site, and those were absent among shrews captured in 2020 and 2022.
In Azhendarovo, the capture sites were located at the junction of the forest-steppe and taiga zones of the foothills of the Kuznetsk Alatau. The animals were caught in a floodplain meadow, an overgrown clearing in place of black taiga, and in an ecotone area between the floodplain meadow and the overgrown clearing. In addition to the above-mentioned species from Lake Teletskoye, the flat-skulled shrew (S. roboratus) is added to the shrew community of Azhendarovo. The dominant species is also the common shrew (40.1%). The proportion of the Eurasian water shrews varies from 3.6% to 6.7% of the community [35]. The location of the sites and the longer period of capture determined the higher number of N. fodiens captured in this locality during 2021–2025.
The Eurasian water shrew, the host of BIRV and BOGV, is widespread in the forest zone of Eurasia from the British Isles to the Pacific Ocean [36]. The geographic range includes most of Europe, Siberian and far-eastern Russia, Sakhalin Island, north Korea, northwestern Mongolia and China. BIRV was found in one locality of Western Siberia, while BOGV was detected in distant localities of its host geographic range, in Western Siberia and European countries, Poland and Finland [5,24]. Based on broad geographic range of other hantavirus pairs, ALTV and SWSV, LENV and ARTV, throughout the distribution of their hosts, we can suggest that BIRV also might be widespread in the forest zone of Eurasia and co-circulate with BOGV among N. fodiens. Co-circulation of hantaviruses in the same host species also raises the distinct possibility of co-infection and reassortment or recombination as a mechanism for rapid evolutionary change.
Analysis of nearly the whole genome of BIRV demonstrated that it represents a new hantavirus species, which belong to the Mobatvirus genus. Comparison of BIRV with available partial Siberian and European BOGV L- and M-segment sequences showed significant difference between the two viruses. However, we found inconsistency during comparative sequence analysis of BIRV and BOGV (strain 2074) from Poland based on available 783 nucleotides fragment of L-segment. Two parts of this fragment demonstrated different level of divergence with BIRV. Sequence, corresponding to 356 nucleotides fragment, positions (2980–3331) exhibited high nucleotide and amino acid sequences divergence (31.4% and 32.5%), while adjacent fragment 427 (2541–2968) nucleotides in length was much closer to BIRV (17.1% and 2.1%). Whether this means there might be evidence of recombination between BIRV and BOGV during co-infection, warrants further investigation. Undoubtedly, analysis based on the partial L- and M-segment sequences is inadequate, and full-genome of BOGV strains are needed to acquire a better understanding of the phylogenetic relationships and possible recombination events in the evolutionary history of BOGV.
Based on the complete M-, L- and partial S-segment phylogeny (Figure 2), BIRV and strain HV/SC/C7-49.2 from China shared a common ancestry with ALTV and LENV, forming together a sub-clade within the Mobatvirus genus. The natural host of HV/SC/C7-49.2, the De Winton’s shrew (Chodsigoa hypsibius), is endemic and widely distributed in central and southwestern China. The habitats of the natural hosts of ALTV (S. araneus), BIRV (Neomys fodiens) and LENV (S. caecutiens) overlap with each other and with Chodsigoa hypsibius in the Hengduan Mountains region, which ensures the possibility of contacts between species and interspecies transmission of associated viruses.
5. Conclusions
BIRV and BOGV represent genetically distinct hantaviruses that share the same host species. The finding of BIRV in N. fodiens supports the hypothesis that BIRV, ALTV and LENV arose from ancient host-switching events from another reservoir host with subsequent diversification within the Soricinae subfamily in Eurasia.
Author Contributions
Conceived and designed the experiments: L.N.Y.; collected the samples: S.A.A., E.M.L., T.A.D., A.V.K. and E.D.V.; performed the experiments: L.N.Y., N.A.S. and T.V.T.; analyzed the data: L.N.Y., S.A.A., E.M.L., and R.Y.; wrote and edited the paper: L.N.Y., S.A.A., and R.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the state assignment of FBRI SRC VB VECTOR, Rospotrebnadzor, Russia; the Federal Fundamental Scientific Research Program for 2021–2025, Russia (Project No. FWGS-2021-0002).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and met the guideline requirements of the order of the Russian High and Middle Education Ministry (No. 742 issued on 13 November, 1984) and by the Federal Law of the Russian Federation (No. 498-FZ issued on 19 December, 2018). Field procedures and protocols were approved by the Institutional Animal Care and Use Committee of the Institute of Systematics and Ecology of Animals (Protocol 2020-02, 2021-01). The study did not involve endangered or protected species.
Data Availability Statement
GenBank accession numbers for newfound viruses are available in Table 1. Other presented data are available on request from corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Maes, P.; Adkins, S.; Alkhovsky, S.V.; Avšič-Županc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, E.; Blair, C.D.; Briese, T.; et al. Taxonomy of the order Bunyavirales: Second update 2018. Arch. Virol. 2019, 164, 927–941. [Google Scholar] [CrossRef]
- Jonsson, C.B.; Figueiredo, L.T.; Vapalahti, O. A global perspective on hantavirus ecology, epidemiology, and disease. Clin. Microbiol. Rev. 2010, 23, 412–444. [Google Scholar] [CrossRef]
- Yanagihara, R.; Gu, S.H.; Arai, S.; Kang, H.J.; Song, J.-W. Hantaviruses: rediscovery and new beginnings. Virus Res. 2014, 187, 6–14. [Google Scholar] [CrossRef]
- Zhang, Y.Z. Discovery of hantaviruses in bats and insectivores and the evolution of the genus Hantavirus. Virus Res. 2014, 187, 15–21. [Google Scholar] [CrossRef]
- Ling, J.; Sironen, T.; Voutilainen, L.; Hepojoki, S.; Niemimaa, J.; Isoviita, V.M.; Vaheri, A.; Henttonen, H.; Vapalahti, O. Hantaviruses in Finnish soricomorphs: Evidence for two distinct hantaviruses carried by Sorex araneus suggesting ancient host-switch. Infect. Genet. Evol. 2014, 27, 51–61. [Google Scholar] [CrossRef]
- Kang, H.J.; Gu, S.H.; Yashina, L.N.; Cook, J.A.; Yanagihara, R. Highly divergent genetic variants of soricid-borne Altai virus (Hantaviridae) in Eurasia suggest ancient host-switching events. Viruses 2019, 11, 857. [Google Scholar] [CrossRef]
- Kang, H.J.; Bennett, S.N.; Dizney, L.; Sumibcay, L.; Arai, S.; Ruedas, L.A.; Song, J.W.; Yanagihara, R. Host switch during evolution of a genetically distinct hantavirus in the American shrew mole (Neurotrichus gibbsii). Virology 2009, 388, 8–14. [Google Scholar] [CrossRef]
- Klempa, B. Reassortment events in the evolution of hantaviruses. Virus Genes 2018, 54, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Liphardt, S.W.; Kang, H.J.; Dizney, L.J.; Ruedas, L.A.; Cook, J.A.; Yanagihara, R. Complex history of codiversification and host switching of a newfound soricid-borne orthohantavirus in North America. Viruses 2019, 11, 637. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, P.; Tia, M.; Alabi, A.; Anon, J.C.; Auste, B.; Essbauer, S.; Gnionsahe, A.; Kigninlman, H.; Klempa, B.; Kraef, C.; et al. Human infections by non-rodent-associated hantaviruses in Africa. J. Infect. Dis. 2016, 214, 1507–1511. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Shimizu, K.; Nishigami, K.; Tsuda, Y.; Sarathukumara, Y.; Muthusinghe, D.S.; Gamage, C.D.; Granathne, L.; Lokupathirage, S.M.W.; Nanayakkara, N.; et al. Serological methods for detection of infection with shrew-borne hantaviruses: Thottapalayam, Seewis, Altai, and Asama viruses. Arch. Virol. 2021, 166, 275–280. [Google Scholar] [CrossRef]
- Song, J.-W.; Gu, S.H.; Bennett, S.N.; Arai, S.; Puorger, M.; Hilbe, M.; Yanagihara, R. Seewis virus, a genetically distinct hantavirus in the Eurasian common shrew (Sorex araneus). Virol. J. 2007, 4, 114. [Google Scholar] [CrossRef] [PubMed]
- Yashina, L.; Abramov, S.; Gutorov, V.; Dupal, T.; Krivopalov, A.; Panov, V.; Danchinova, G.; Vinogradov, V.; Luchnikova, E.; Hay, J.; Kang, H.J.; Yanagihara, R. Seewis virus: Phylogeography of a shrew-borne hantavirus in Siberia, Russia. Vector-Borne Zoonotic Dis. 2010, 10, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, M.; Radosa, L.; Rosenfeld, U.M.; Schmidt, S.; Triebenbacher, C.; Löhr, P.W.; Fuchs, D.; Heroldová, M.; Jánová, E.; Stanko, M.; Mošanský, L.; Fričová, J.; Pejčoch, M.; Suchomel, J.; Purchart, L.; Groschup, M.H.; Krüger, D.H.; Klempa, B.; Ulrich, R.G. Broad geographical distribution and high genetic diversity of shrew-borne Seewis hantavirus in Central Europe. Virus Genes 2012, 45, 48–55. [Google Scholar] [CrossRef]
- Yashina, L.N.; Abramov, S.A.; Zhigalin, A.V.; Smetannikova, N.A.; Dupal, T.A.; Krivopalov, A.V.; Kikuchi, F.; Senoo, K.; Arai, S.; Mizutani, T.; et al. Geographic distribution and phylogeny of soricine shrew-borne Seewis virus and Altai virus in Russia. Viruses 2021, 13, 1286. [Google Scholar] [CrossRef]
- Kang, H.J.; Bennett, S.N.; Sumibcay, L.; Arai, S.; Hope, A.G.; Mocz, G.; Song, J.-W.; Cook, J.A.; Yanagihara, R. Evolutionary insights from a genetically divergent hantavirus harbored by the European common mole (Talpa europaea). PLoS ONE 2009, 4, e6149. [Google Scholar] [CrossRef]
- Laenen, L.; Vergote, V.; Kafetzopoulou, L.E.; Wawina, T.B.; Vassou, D.; Cook, J.A.; Hugot. J.P.; Deboutte, W.; Kang, H.J.; Witkowski, P.T.; Köppen-Rung, P.; et al. A novel hantavirus of the European mole, Bruges virus, is involved in frequent Nova virus coinfections. Genome Biol. Evol. 2018, 10, 45–55. [Google Scholar] [CrossRef]
- Gu, S.H.; Miñarro, M.; Feliu, C.; Hugot, J.-P.; Forrester, N.L.; Weaver, S.C.; Yanagihara, R. Multiple lineages of hantaviruses harbored by the Iberian mole (Talpa occidentalis) in Spain. Viruses 2023, 15, 1313. [Google Scholar] [CrossRef]
- Yashina, L.N.; Kartashov, M.Y.; Wang, W.; Li, K.; Zdanovskaya, N.I.; Ivanov, L.I.; Zhang, Y.Z. Co-circulation of distinct shrewborne hantaviruses in the far east of Russia. Virus Res. 2019, 272, 197717. [Google Scholar] [CrossRef]
- Kang, H.J.; Arai, S.; Hope, A.G.; Cook, J.A.; Yanagihara, R. Novel hantavirus in the flat-skulled shrew (Sorex roboratus). Vector Borne Zoonotic Dis. 2010, 10, 593–597. [Google Scholar] [CrossRef]
- Arai, S.; Kang, H.J.; Gu, S.H.; Ohdachi, S.D.; Cook, J.A.; Yashina, L.N.; Tanaka-Taya, K.; Abramov, S.A.; Morikawa, S.; Okabe, N.; et al. Genetic diversity of Artybash virus in the Laxmann’s shrew (Sorex caecutiens). Vector Borne Zoonotic Dis. 2016, 16, 468–475. [Google Scholar] [CrossRef]
- Yashina, L.N.; Ivanov, L.I.; Kompanets, G.G.; Zdanovskaya, N.I.; Kartashov, M.Yu. Shrew-borne hantaviruses (Hantaviridae: Orthohantavirus) in the Far East of Russia. Problems of Virology (Voprosy Virusologii). 2023, 68, 79–85. [Google Scholar] [CrossRef]
- Yashina, L.N.; Zinich, L.S.; Smetannikova, N.A.; Kartashov, M.; Kovalenko, I.S.; Yusupova, Z.S.; Konashenko, E.V.; Tikhonov, S.N. Hantaviruses (Hantaviridae) in Republic of Crimea. Mol. Genet. Microbiol. Virol. (In Russ.). 2024, 42, 37–42. [Google Scholar] [CrossRef]
- Gu, S.H.; Markowski, J.; Kang, H.J.; Hejduk, J.; Sikorska, B.; Liberski, P.P.; Yanagihara, R. Boginia virus, a newfound hantavirus harbored by the Eurasian water shrew (Neomys fodiens) in Poland. Virol. J. 2013, 10, 160. [Google Scholar] [CrossRef]
- Tkachenko, E.A.; Ivanov, A.P.; Donets, M.A.; Miasnikov, Y.A.; Ryltseva, E.V.; Gaponova, L.K.; Bashkirtsev,V. N.; Okulova, N.M.; Drozdov, S.G.; Slonova, R.A.; et al. Potential reservoir and vectors of haemorrhagic fever with renal syndrome (HFRS) in the U.S.S.R. Ann. Soc. Belg. Med. Trop. 1983, 63, 267–269. [Google Scholar] [PubMed]
- Klempa, B.; Fichet-Calvet, E.; Lecompte, E.; Auste, B.; Aniskin, V.; Meisel, H.; Barrier, P.; Koivogue, L.; Meulen, J.; Krüger, D.H. Novel hantavirus sequences in shrew, Guinea. Emerg. Infect. Dis. 2007, 13, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv. 2013, 16, 1303. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Cui, X.; Fan, K.; Liang, X.; Gong, W.; Chen, W.; He, B.; Chen, X.; Wang, H.; Wang, X.; Zhang, P.; et al. Virus diversity, wildlife-domestic animal circulation and potential zoonotic viruses of small mammals, pangolins and zoo animals. Nat. Commun. 2023, 14, 2488. [Google Scholar] [CrossRef]
- Yashina, L.N.; Panov, V.V.; Abramov, S.A.; Smetannikova, N.A.; Luchnikova, E.M.; Dupal, T.A.; Krivopalov, A.V.; Arai, S.; Yanagihara, R. Academ virus, a novel hantavirus in the Siberian mole (Talpa altaica) from Russia. Viruses 2022, 14, 309. [Google Scholar] [CrossRef]
- Pavelková Řičánková, V.; Robovský, J.; Riegert, J. Ecological structure of recent and last glacial mammalian faunas in northern Eurasia: The case of Altai-Sayan Refugium. PLoS ONE 2014, 9, e85056. [Google Scholar] [CrossRef] [PubMed]
- Provan, J.; Bennett, K.D. Phylogeographic insights into cryptic glacial refugia. Trends Ecol. Evol. 2006, 23, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Marková, S.; Horníková, M.; Lanier, H.C.; Henttonen, H.; Searle, J.B.; Weider, L.J.; Kotlík, P. High genomic diversity in the bank vole at the northern apex of a range expansion: The role of multiple colonizations and end-glacial refugia. Mol. Ecol. 2020, 29, 1730–1744. [Google Scholar] [CrossRef] [PubMed]
- Shchipanov, N.A. , Litvinov, Y.N.; Sheftel, B.I. Rapid method for estimating local biodiversity of a community of small mammals. Contemp. Probl. Ecol. 2008, 1, 596–602. [Google Scholar] [CrossRef]
- Ilyashenko, V.B.; Luchnikova, E.M.; Skalon, N.S.; Grebentschikov, I.S.; Kovalevsky, A.V. Long-term dynamics of small-mammal communities in anthropogenically disturbed territories in the south-east of West Siberia. IOP Conf. Ser.: Earth Environ. Sci. 2019, 224, 012055. [Google Scholar] [CrossRef]
- Bannikova, A.A.; Lebedev, V.S. Order Eulipotyphla. In The Mammals of Russia: A taxonomic and geographic reference; Pavlinov, I.Ya., Lissovsky, A.A., Eds.; KMK Sci. Press: Moscow, Russia, 2012; pp. 25–73. [Google Scholar]
Figure 1.
Map, showing the locations of the collection sites in Western Siberia, where hantavirus--infected Neomys fodiens were captured. (1) Azhendarovo, (2) Teletskoye. Detected hantaviruses were Biya River virus (BIRV) (red) and Boginia virus (BOGV) (blue). The inset shows the location of trapping area in Eurasia (yellow box).
Figure 1.
Map, showing the locations of the collection sites in Western Siberia, where hantavirus--infected Neomys fodiens were captured. (1) Azhendarovo, (2) Teletskoye. Detected hantaviruses were Biya River virus (BIRV) (red) and Boginia virus (BOGV) (blue). The inset shows the location of trapping area in Eurasia (yellow box).
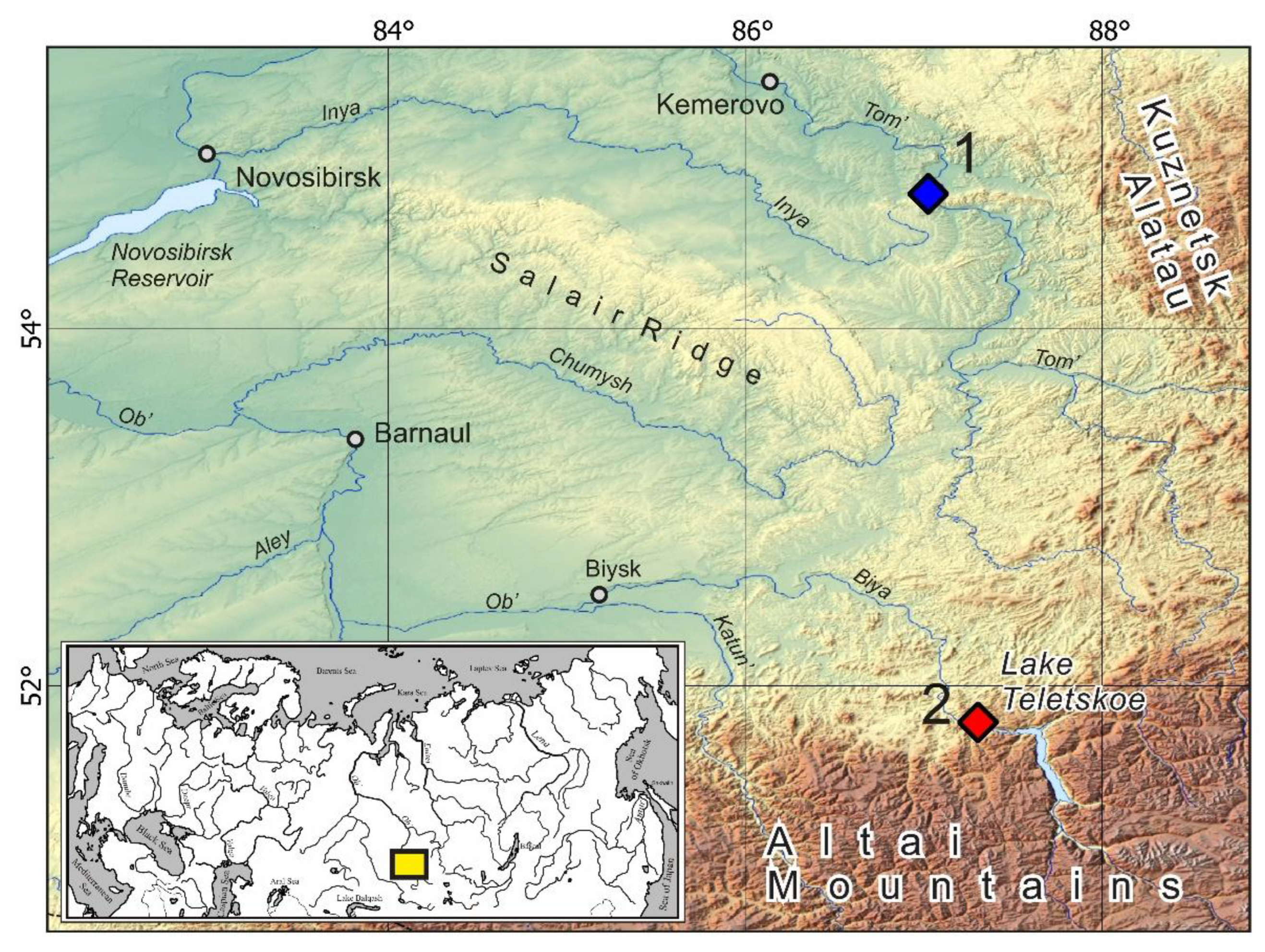
Figure 2.
Phylogenetic trees, generated by maximum-likelihood methods, were based on the alignment of the 353-nucleotide (L) L-segment sequences, and 485-nucleotide (S) S-segment (A); the complete coding regions of the M- and L-segment sequences (B) of newfound BIRV and other representative hantaviruses. Bootstrap values (> 70%) are shown at relevant nodes. The scale bar indicates the nucleotide substitutions per site. Colors (red, and blue bold lettering) highlight newfound hantaviruses. Phylogenetic positions of BIRV are shown in relation to shrew-borne hantaviruses: Altai mobatvirus (ALTV ALT302, S: MK340902; M: MK340903; L: MT648514, ALTV R SO5, S: ON720815; M: ON720819; L: ON720823, ALTV Uurainen, L: KJ136623 and ALTV Parabel-Sa44, S: MT560057; L: MN815789) from Sorex araneus, Lena mobatvirus (LENV Khekhtsir-Sc67, S: MH499470; M: MH499471; L: MH499472, LENV MSB146482, S: KM361043, and LENV Parnaya-Sc1217, S: MW505551; L: MW505552) from Sorex caecutiens, Seewis orthohantavirus (SWSV mp70, S: EF636024; L: EF636026 and SWSV Telet-Sa300, S: MN815797) from Sorex araneus, Seewis orthohantavirus (ARTV/Galkino-St2714, M: MG913806; L: MH499473) from Sorex tundrensis, Asikkala orthohantavirus (ASIV CZ/Beskydy/412, S: NC_043070; M: NC_043069; L: NC_043068) from Sorex minutus, Kenkeme orthohantavirus (KKMV MSB148794, S: GQ306148 and KKMV Fuyuan Sr326, S: NC_034559; M: NC_034565; L: NC_034560) from Sorex roboratus, Boginia virus (BOGV 2074, L: JX990965 and BOGV Laihia, L: KJ136642) from Neomys fodiens, Hantaviridae sp. (HV/SC/C7-49.2, S: OQ451885; M: OQ451886; L: OQ451887) from Chodsigoa hypsibia, Asama orthohantavirus (ASAV N10, S: EU929072; M: EU929075; L: EU929078) from Urotrichus talpoides, Nova mobatvirus (NVAV Te34, S: NC_034464, M: NC_034470, L: NC_034465) and Bruges orthohantavirus (BRGV BE/VieuxGenappeTE2013, S: MK340905; M: OL871119; L: NC_034402) from Talpa europaea, Academ orthohantavirus (ACDV Academ-Ta450/Russia, S: MK340905; M: OL871119; L: PX270319) from Talpa altaica, Thottapalayam thottimvirus (TPMV VRC66412, S: NC_010704, M: NC_010708, L: NC_010707) from Suncus murinus, Imjin thottimvirus (MJNV Cixi, S: NC_034558; M: NC_034557; L: NC_034564) from Crocidura lasiura, Jeju orthohantavirus (JJUV 10-11, S: NC_034398; M: NC_034404; L: NC_034399) from Crocidura shantungensis, Cao Bằng orthohantavirus (CBNV TC-3, S: NC_034484; M: EF543526; L: NC_034485) from Anourosorex squamipes, Bowé orthohantavirus (BOWV VN1512, S: NC_034405; M: KC631783; L: NC_034407) from Crocidura douceti. Also shown are representative rodent-borne hantaviruses, including Sin Nombre orthohantavirus (SNV NMH10, S: NC_005216; M: NC_005215; L: NC_005217), Prospect Hill orthohantavirus (PHV PH-1, S: M34011; M: X55129; L: EF646763), Tula orthohantavirus (TULV M5302v, S: NC_005227; M: NC_005228; L: NC_005226), Puumala orthohantavirus (PUUV Sotkamo, S: NC_005224; M: NC_005223; L: NC_005225), Dobrava/Belgrade orthohantavirus (DOBV/BGDV Ano-Poroia, S: NC_005233; M: NC_005234; L: NC_005235), Hantaan orthohantavirus (HTNV 76-118, S: NC_005218; M: NC_005219; L: NC_005222), and Seoul orthohantavirus (SEOV 80-39, S: NC_005236; M: NC_005237; L: NC_005238). Bat-borne hantaviruses include Brno loanvirus (BRNV 7/2012/CZE, S: KX845678; M: KX845679; L: KX845680) from Nyctalus noctula, Láibīn mobatvirus (LAIV BT20, S: KM102247; M: KM102248; L: KM102249) from Taphozous melanopogon, Xuân Sơn mobatvirus (XSV PR15, S: KC688335; M: NC_078261; L: NC_078262) from Hipposideros pomona, Quezon mobatvirus (QZNV MT1720/1657, S: NC_034400; M: NC_034393; L: NC_034401) from Rousettus amplexicaudatus, Lóngquán loanvirus (LQUV Rs-32, S: NC_043126; M: NC_043127 and L: NC_043125) from Rhinolophus sinicus, Robina mobatvirus (ROBV P17-14855, S: NC_055633; M: NC_055634; L: NC_055632) from Pteropus alecto, respectively.
Figure 2.
Phylogenetic trees, generated by maximum-likelihood methods, were based on the alignment of the 353-nucleotide (L) L-segment sequences, and 485-nucleotide (S) S-segment (A); the complete coding regions of the M- and L-segment sequences (B) of newfound BIRV and other representative hantaviruses. Bootstrap values (> 70%) are shown at relevant nodes. The scale bar indicates the nucleotide substitutions per site. Colors (red, and blue bold lettering) highlight newfound hantaviruses. Phylogenetic positions of BIRV are shown in relation to shrew-borne hantaviruses: Altai mobatvirus (ALTV ALT302, S: MK340902; M: MK340903; L: MT648514, ALTV R SO5, S: ON720815; M: ON720819; L: ON720823, ALTV Uurainen, L: KJ136623 and ALTV Parabel-Sa44, S: MT560057; L: MN815789) from Sorex araneus, Lena mobatvirus (LENV Khekhtsir-Sc67, S: MH499470; M: MH499471; L: MH499472, LENV MSB146482, S: KM361043, and LENV Parnaya-Sc1217, S: MW505551; L: MW505552) from Sorex caecutiens, Seewis orthohantavirus (SWSV mp70, S: EF636024; L: EF636026 and SWSV Telet-Sa300, S: MN815797) from Sorex araneus, Seewis orthohantavirus (ARTV/Galkino-St2714, M: MG913806; L: MH499473) from Sorex tundrensis, Asikkala orthohantavirus (ASIV CZ/Beskydy/412, S: NC_043070; M: NC_043069; L: NC_043068) from Sorex minutus, Kenkeme orthohantavirus (KKMV MSB148794, S: GQ306148 and KKMV Fuyuan Sr326, S: NC_034559; M: NC_034565; L: NC_034560) from Sorex roboratus, Boginia virus (BOGV 2074, L: JX990965 and BOGV Laihia, L: KJ136642) from Neomys fodiens, Hantaviridae sp. (HV/SC/C7-49.2, S: OQ451885; M: OQ451886; L: OQ451887) from Chodsigoa hypsibia, Asama orthohantavirus (ASAV N10, S: EU929072; M: EU929075; L: EU929078) from Urotrichus talpoides, Nova mobatvirus (NVAV Te34, S: NC_034464, M: NC_034470, L: NC_034465) and Bruges orthohantavirus (BRGV BE/VieuxGenappeTE2013, S: MK340905; M: OL871119; L: NC_034402) from Talpa europaea, Academ orthohantavirus (ACDV Academ-Ta450/Russia, S: MK340905; M: OL871119; L: PX270319) from Talpa altaica, Thottapalayam thottimvirus (TPMV VRC66412, S: NC_010704, M: NC_010708, L: NC_010707) from Suncus murinus, Imjin thottimvirus (MJNV Cixi, S: NC_034558; M: NC_034557; L: NC_034564) from Crocidura lasiura, Jeju orthohantavirus (JJUV 10-11, S: NC_034398; M: NC_034404; L: NC_034399) from Crocidura shantungensis, Cao Bằng orthohantavirus (CBNV TC-3, S: NC_034484; M: EF543526; L: NC_034485) from Anourosorex squamipes, Bowé orthohantavirus (BOWV VN1512, S: NC_034405; M: KC631783; L: NC_034407) from Crocidura douceti. Also shown are representative rodent-borne hantaviruses, including Sin Nombre orthohantavirus (SNV NMH10, S: NC_005216; M: NC_005215; L: NC_005217), Prospect Hill orthohantavirus (PHV PH-1, S: M34011; M: X55129; L: EF646763), Tula orthohantavirus (TULV M5302v, S: NC_005227; M: NC_005228; L: NC_005226), Puumala orthohantavirus (PUUV Sotkamo, S: NC_005224; M: NC_005223; L: NC_005225), Dobrava/Belgrade orthohantavirus (DOBV/BGDV Ano-Poroia, S: NC_005233; M: NC_005234; L: NC_005235), Hantaan orthohantavirus (HTNV 76-118, S: NC_005218; M: NC_005219; L: NC_005222), and Seoul orthohantavirus (SEOV 80-39, S: NC_005236; M: NC_005237; L: NC_005238). Bat-borne hantaviruses include Brno loanvirus (BRNV 7/2012/CZE, S: KX845678; M: KX845679; L: KX845680) from Nyctalus noctula, Láibīn mobatvirus (LAIV BT20, S: KM102247; M: KM102248; L: KM102249) from Taphozous melanopogon, Xuân Sơn mobatvirus (XSV PR15, S: KC688335; M: NC_078261; L: NC_078262) from Hipposideros pomona, Quezon mobatvirus (QZNV MT1720/1657, S: NC_034400; M: NC_034393; L: NC_034401) from Rousettus amplexicaudatus, Lóngquán loanvirus (LQUV Rs-32, S: NC_043126; M: NC_043127 and L: NC_043125) from Rhinolophus sinicus, Robina mobatvirus (ROBV P17-14855, S: NC_055633; M: NC_055634; L: NC_055632) from Pteropus alecto, respectively.
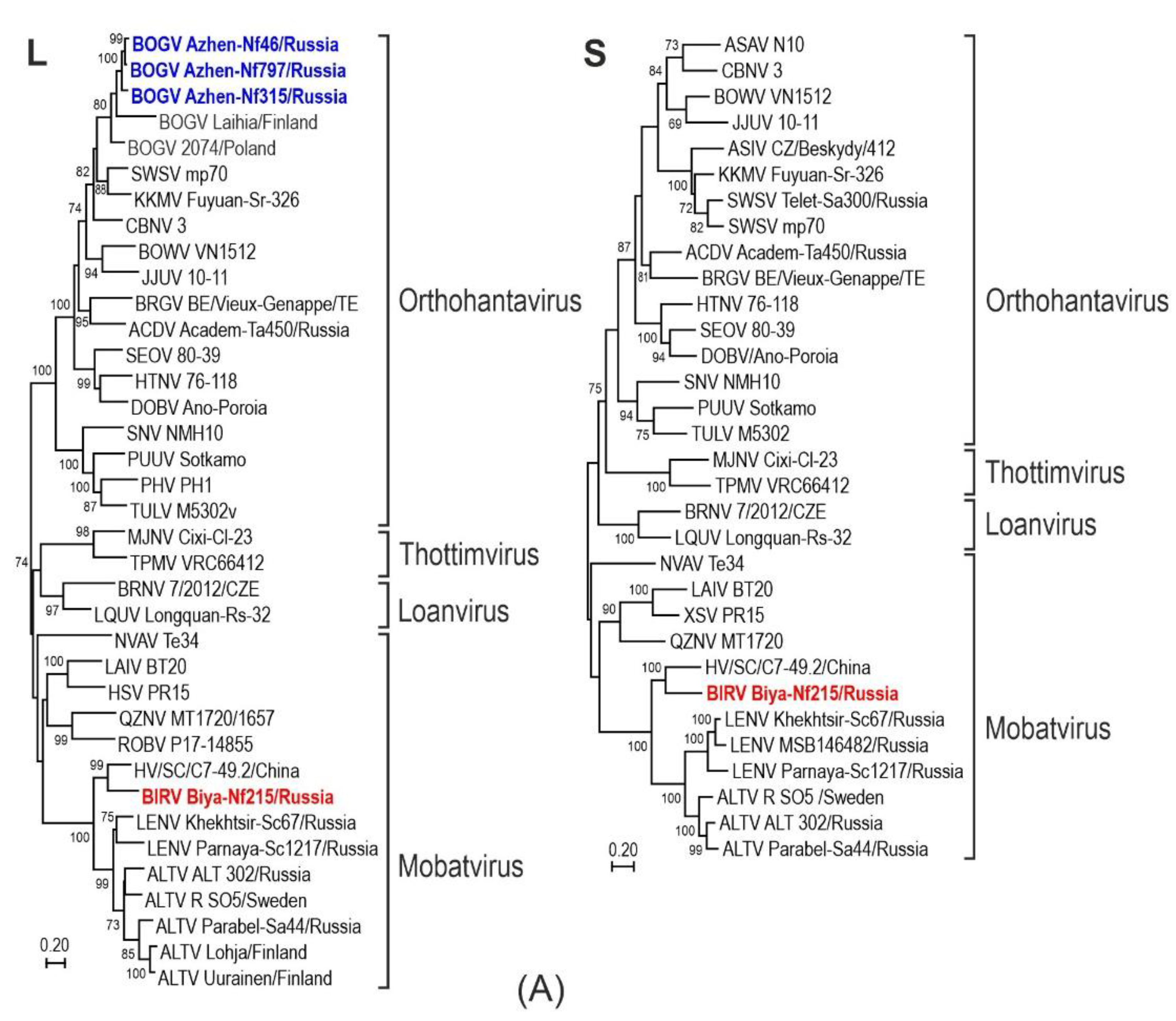
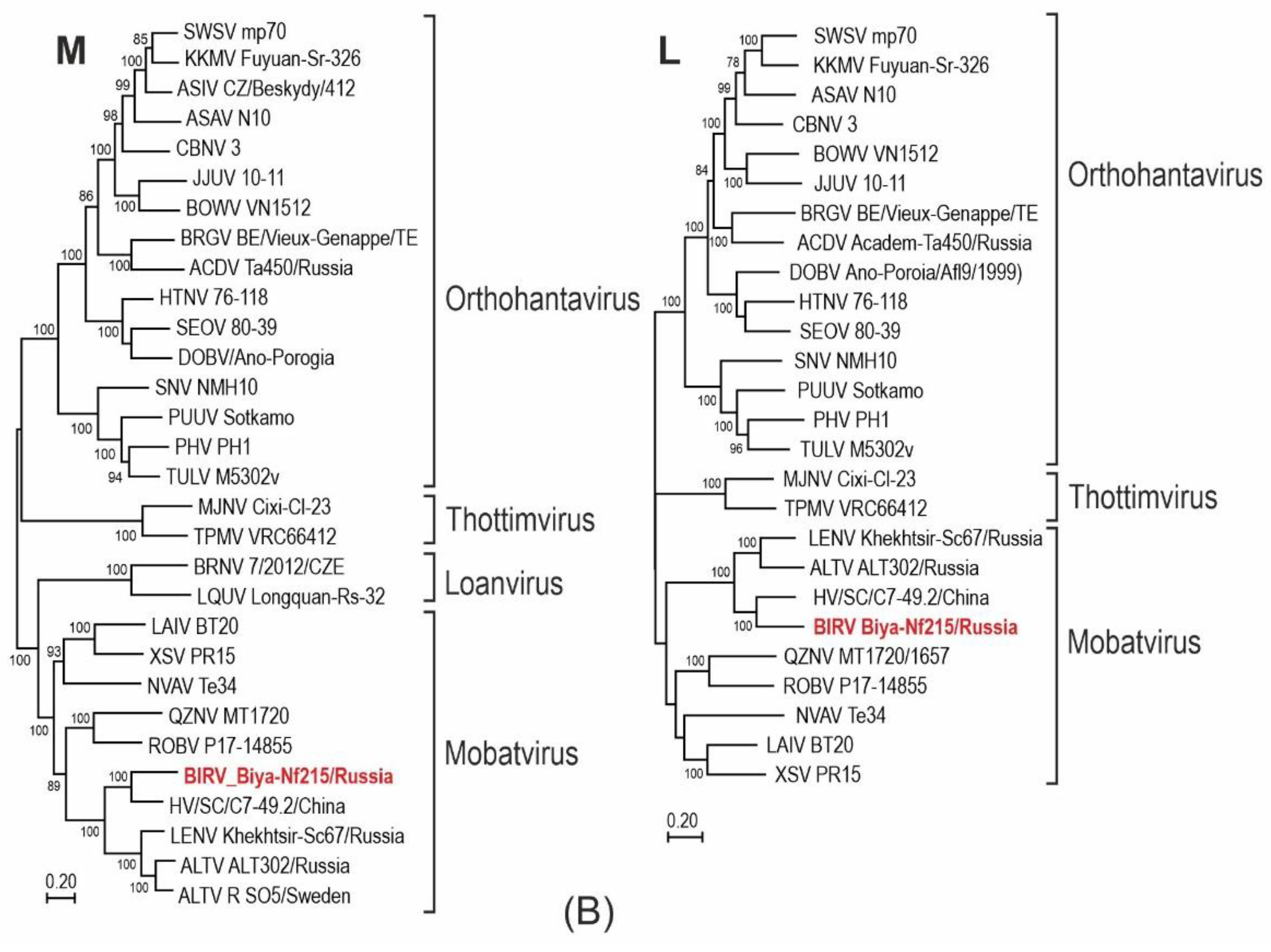

Table 1.
Prevalence of hantavirus RNA and hantavirus sequences in Neomys fodiens from Western Siberia, Russia.
Table 1.
Prevalence of hantavirus RNA and hantavirus sequences in Neomys fodiens from Western Siberia, Russia.
| Positive/tested | GenBank no. | |||||
| Capture site | Year | shrews | Virus strain | S | M | L |
| Altai Republic, Teletskoye | 2019 | 1/2 | BIRV_Biya-Nf215 | PQ355537 | PQ355538 | PQ355539 |
| Kemerovo Oblast, Azhendarovo | 2021 | 1/13 | BOGV_Azhen-Nf315 | - | - | PQ355534 |
| 2022 | 2/17 | BOGV_Azhen-Nf46 | - | - | PQ355535 | |
| BOGV_Azhen-Nf797 | - | - | PQ355536 | |||
| 2023 | 0/1 | - | - | - | - | |
| 2024 | 0/6 | - | - | - | - | |
| 2025 | 0/25 | - | - | - | - | |
“-“ sequences unavailable.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



