Submitted:
26 September 2025
Posted:
29 September 2025
You are already at the latest version
Abstract
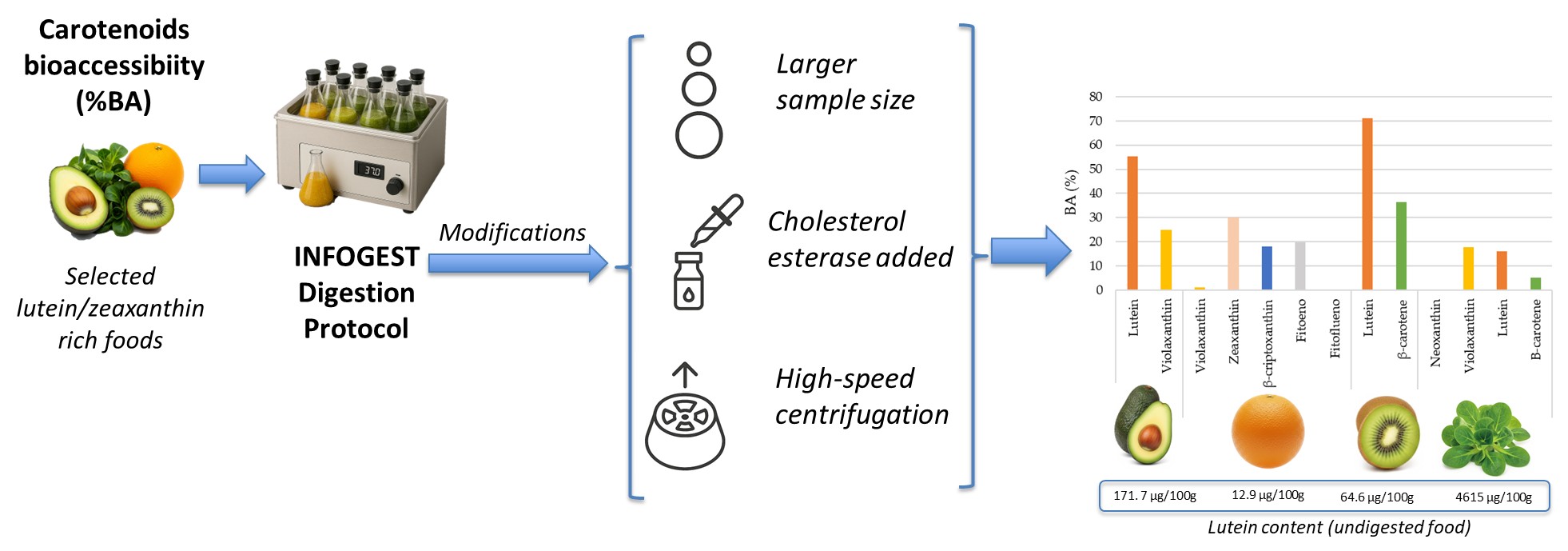
Background/Objectives: This study aimed to evaluate the bioaccessibility (BA) of lutein and other carotenoids in selected fruits (avocado, kiwi, orange) and a leafy green vegetable (lamb's lettuce) using a modified INFOGEST in vitro digestion protocol. The modifications were specifically designed to enhance the release, micellarization, and subsequent quantification of carotenoids. Methods: The standardized static in vitro digestion model proposed by the COST Action INFOGEST was adapted. Key modifications included using a larger sample size (20 g) and optimizing centrifugation conditions (20,000 G for 5 minutes) to efficiently transfer carotenoids to the aqueous-micellar phase. Quantification was done by HPLC. Results: Lamb's lettuce was rich in lutein (4615.03 µg/100g) and β-carotene (2156.42 µg/100g); avocado was a notable source for lutein (171.67 µg/100g); kiwi contained lutein (64.55 µg/100g); and orange was a source of β-cryptoxanthin (73.39 µg/100g) and zeaxanthin (12.85 µg/100g) in the Spanish population. Regarding stability and BA, lutein in avocado showed high recovery (95.84%) and a BA of 55.32%. Kiwi exhibited excellent lutein stability (73-80% recovery) and the highest BA for lutein (71.01%), with β-carotene at 36.39%. Orange had low violaxanthin BA (1.02%) but intermediate values for zeaxanthin (29.96%) and β-cryptoxanthin (18.03%). In contrast, lamb's lettuce showed low recoveries and significant losses, with very low β-carotene BA (5.08%). Conclusions: sample weight and centrifugation speed are crucial modifications for accurate carotenoid BA assessment. Carotenoid BA varies depending on the specific carotenoid and the food matrix. This highlights the importance of adapting in vitro digestion protocols to the characteristics of lipophilic compounds and food matrices.
Keywords:
Carotenoids
; lutein
; bioaccessibility
; In vitro digestion
; INFOGEST
1. Introduction
Carotenoids are natural, fat-soluble pigments synthesised by plants, algae, and certain bacteria. They are recognised for their various biological activities. These compounds have a wide variety of molecular structures, giving them antioxidant and pro-vitamin properties (some, such as β-carotene, are precursors of vitamin A), as well as important roles in vision and immune health [1]. Within this family, lutein and zeaxanthin are dietary xanthophylls with well-established roles in ocular health, particularly in protecting the macula from oxidative damage and age-related degeneration [2]. Dietary intake of lutein and zeaxanthin have been associated with improvements in blood levels, macular pigment optical density and visual function [3,4], yet their effective bioavailability remains a limiting factor for its health-promoting efficacy [1,3].
Human absorption of carotenoids, especially from raw foods, is generally inefficient due to physical barriers in plant tissues, competition among lipophilic compounds, and limited solubilization. Therefore, measuring bioaccessibility (BA) in vitro is critical for designing interventions that enhance carotenoid intake [1,5].
The INFOGEST protocol has been widely adopted as a standardized model to simulate human gastrointestinal digestion in vitro studies. However, several publications have indicated that this protocol may require specific adaptations when applied to lipophilic compounds such as carotenoids, due to their low solubility in aqueous environments and the limited hydrolysis efficiency of their esterified forms [5,6]. In this regard, several studies have successfully adapted the protocol to analyse carotenoids, with modifications typically involving additional steps to account for their lipophilic nature and the need for efficient micellarization and extraction. Specifically, adaptations often include enhanced micelle separation and carotenoid extraction, where after the digestion phases, methods are incorporated to efficiently separate the micellar fraction containing the solubilized carotenoids, commonly involving centrifugation at high speeds to isolate the aqueous phase with mixed micelles, followed by exhaustive extraction with organic solvents to recover the carotenoids for quantification [5,6]. Furthermore, optimization of enzyme concentrations or activities may be applied. For example, inclusion of specific gastric lipases may be considered as an effective adaptation to the standard INFOGEST protocol, since although it outlines the basic enzymatic setup, the addition of enzymes such as rabbit gastric extract can significantly enhance the lipolysis of plant-based food matrices [7]. These adaptations collectively aim to provide a more accurate estimation of carotenoid BA, overcoming the inherent limitations of the general INFOGEST protocol for these important lipophilic compounds.
Therefore, in the present study, methodological modifications in the original INFOGEST protocol [8] were introduced to improve the release and micellarization of carotenoids in good sources of lutein, and posterior quantification. These included using a larger sample size (20 g instead of the standard 5 g), the incorporation of cholesterol esterase in the intestinal phase to facilitate xanthophyll ester hydrolysis [9], the assay of different centrifugation methods to remove the undigested sample and to conduct experiments with different solvents (diethyl ether: hexane: dichloromethane and diethyl ether: methanol: methyl tert-butyl ether to carry out the extraction of carotenoids in the digested food.
The aim of this study was to evaluate the BA of lutein and other carotenoids in select fruits (avocado, kiwi, and orange) and leafy green vegetables (lamb's lettuce) using an in vitro digestion following the INFOGEST protocol [8] with modifications aimed at enhancing carotenoid release, micellization, and quantification.
2. Materials and Methods
2.1. Reagents
The following reagents were used to prepare the simulated digestion fluids: KCl (Merck, Darmstadt, Germany), MgCl2(H2O)6 and (NH4)2CO3 (Sigma- Aldrich, Missouri, USA), KH2PO4, NaHCO3, Na Cl, HCl and CaCl2(H2O)2 (Panreac, Barcelona, Spain).
Standard digestive enzymes namely α-amylase from porcine pancreas (EC 3.2.1.1), pepsin from porcine gastric mucosa (EC 3.4.23.1), cholesterol esterase from porcine pancreas (EC 3.1.1.13), bile from bovine and ovine (EC 232-369-0), and pancreatin from porcine pancreas (EC 232-468-9) were purchased from Sigma-Aldrich (Missouri, USA).
Acetone, tert-butyl methyl ether (tBME), methanol (MetOH), ethanol, hexane, dichloromethane (DCM), petroleum ether (PE) and diethyl ether (DE), were obtained from Análisis Vínicos (Tomelloso, Spain). Lutein (xanthophyll from marigold), zeaxanthin, α- and β-carotene, β-cryptoxanthin, violaxanthin, neoxanthin, phytoene, phytofluene, tocopherol acetate, potassium hydroxide (KOH), pyrogallic acid and trimethylamine were obtained from Sigma Aldrich (Madrid, Spain).
2.2. Samples
Avocados (Hass), kiwi (Oscar®, variety Hayward), oranges (Nave-Late), and lamb´s lettuces (Machuca®, in pre-packaged 150g trays) were purchased from Mercamadrid in Madrid, Spain, in March 2017. These foods were supplied weekly to volunteers participating in an intervention study [10]. From these, 3 pieces of each food (2 trays in the case of lamb’s lettuce) were saved for analysis (both fresh and after undergoing the digestion process).
2.3. Extraction, Saponification and HPLC Analysis of Carotenoids
The fruits were manually peeled, the seeds were removed, and the edible portion was homogenized in a hand blender for sufficient time to obtain a puree. The lamb's lettuce was homogenized directly using the same method. The amounts necessary for carotenoid analysis or in vitro digestion assay were separated. To carotenoids analysis in non-digested foods, each sample was prepared in triplicate, with each replicate ranging from 10 to 20 grams based on its colour intensity. These homogenized samples were meticulously ground with celite in a mortar. The mixture was then subjected to repeated extractions with acetone until the resulting extract appeared colourless. The organic extract was subsequently transferred to a separatory funnel containing a 1:1 mixture of DE:PE. Acetone was then systematically removed by successive washes with water until the aqueous layer showed no colour. Any remaining traces of water were absorbed using anhydrous sodium sulphate. The purified sample was then evaporated to dryness using a rotary evaporator. For analysis, the dry residue was redissolved in an injection solvent consisting of tBME and MetOH (50:50) and passed through a 0.45 µm filter to remove particulates [11,12].
To hydrolyse carotenoid esters and clear the carotenoid extract of interfering substances, such as chlorophylls and unwanted lipids, the extraction of carotenoids (from raw foods) was followed by alkaline saponification of the extract according to the procedure of Granado et al. [13] with slight modifications [12]. For analysis, aliquots from each food extract (400 µL) were saponified. The process involved mixing the aliquot with 0.1 M pyrogallic acid in ethanol and 30% KOH in MetOH, followed by 7 minutes of ultrasonication. The extract was then washed with water, followed by extraction with a 1:1 DE:PE (50:50) mixture, using vortexing and centrifugation. Finally, the supernatant was dried under nitrogen, reconstituted in MetOH:tBME (50:50), and prepared for HPLC injection.
Carotenoid concentrations were determined by high performance liquid chromatography (HPLC) using a system consisting of a model 600 pump, a Rheodyne injector and a 2998 diode array detector (DAD) (Waters, Milford, MA, USA) and a C30 YMC column (5 µm, 250 × 4.6 mm i.d.) (Waters, Wilmington, MA, USA) with a guard column (Aquapore ODS type RP-18) at room temperature (25 °C). Mobile phase was formed by MetOH with 0.1% trimethylamine (solvent A) and tBME (solvent B) in a linear gradient. At baseline, 25, 55 and 60 min the ratios of the solvents were 95:5, 70:30, 35:65 and 95:5. The detection was performed at 450 nm for carotenoids, phytoene at 285 nm, phytofluene at 370 nm and 285 nm for tocopherol acetate (internal standard). Chromatograms were processed using Empower 2 software (Waters, Milford, MA, USA). Quantification was performed using standard curves.
2.4. Selection of Digestion Protocol and the Extraction Method for Carotenoids from Digested Foods
Two different protocols of in vitro digestion were tested using fresh oranges (Citrus sinensis, L.) (Figure 1) before the in vitro digestion of the foods selected for the study. Both were based on the standardized static in vitro digestion model proposed by the COST action INFOGEST [8] plus cholesterol esterase (3,077 U/mL). The digestion process included oral, gastric, and small intestinal phases, followed by the separation of the supernatant. For both protocols, the oral phase involved 3.5 mL oral fluid, 25 µL CaCl2, and 0.5 mL α-amylase (1,500 U/mL) at 37ºC for 2 minutes. The gastric phase involved 7.5 mL gastric fluid, 5 µL CaCl2, and 1.6 mL pepsin (25,000 U/mL), with pH adjusted to 3 with HCl at 37ºC. The small intestine phase included 10 mL intestinal fluid, 40 µL CaCl2, 5 mL pancreatin (800 U/mL) and 2.5 mL bile (160 mM), and 1 mL cholesterol esterase (3,077 U/mL), with pH adjusted to 7 with NaOH at 37ºC. Upon completion each phase, samples were placed in an ice bath. In accordance with the INFOGEST protocol, enzymes should ideally be incorporated based on their experimentally determined activity to ensure physiological relevance and reproducibility. However, in our study, enzymatic activities were not directly measured; instead, we relied on the activity values provided by the manufacturer, which were used as reference.
This approach has been adopted in previous applications of the INFOGEST method, particularly when technical constraints prevent direct measurement. Given that enzymes were sourced from reputable suppliers with standardized quality controls, this method is considered acceptable within the framework of the protocol and supports reproducibility under practical laboratory conditions.
- Protocol 1: 5 g homogenized sample was mixed with reagents specified by INFOGEST [8] plus cholesterol esterase (3,077 U/mL). The phases lasted 2 minutes (oral), 2 hours (gastric), and 2 hours (duodenal phase). After digestion, centrifugation was performed at 10,000 G for 10 minutes, which were the conditions we followed in previous studies [12].
- Protocol 2: Similar to Protocol 1, but a 20 g sample was used and, and, as a result, the enzyme concentration was also quadrupled (aimed to achieve higher absorbance values and reduce analytical determination errors). Furthermore, the gastric phase was shortened to 1 hour, and centrifugation was performed at 20,000 G for 5 minutes (to transfer carotenoids from the duodenal digesta to the aqueous-micellar phase). The centrifugation conditions were selected in based on different assays previously conducted on kiwis (Actinidia deliciosa) that were extracted, digested following protocol 1 and centrifuged at the last step following different conditions: 5,000 G, 20 min; 10,000 G, 5 min; 10,000 G, 10 min; 10,000 G, 20 min and 20,000 G, 5 min. Centrifugation assays conducted on kiwis (Actinidia deliciosa) showed the following BA percentages: 8.9% (5,000 G, 20 min), 10% (10,000 G, 5 min), 6.4% (10,000 G, 10 min), 5.1% (10,000 G, 20 min), and 16.6% (20,000 G, 5 min).
To extract carotenoids from the digested food fraction, a mixture of DE:MetOH:tBME (30:35:35) was used, as it yielded the best results after extracting a digested kiwi sample following protocol 2. Following this digestion, carotenoids were extracted using: a) DE:Hexane:DCM (30:35:35) (using a Hexane:DCM mixture (1:5 v/v) as this is what we use for carotenoid extraction in human serum [14] and b) DE:MetOH:tBME (30:35:35) (as MetOH and tBME are the mobile phases used in HPLC analysis). In both cases, DE was used as an intermediate polarity solvent. Briefly, the digesta fraction was mixed with the selected solvent mixture for extraction and stirred for 2 minutes on a stirring plate. Subsequently, after adding 10% NaCl, the mixture was centrifuged (10,000 G, 10 min), and the organic phase was collected. The process was repeated until the aqueous phase was colourless. After combining the extracted organic phases, the solvent was removed using a rotary evaporator, and the residue was reconstituted in MetOH:tBME (50:50). After extraction, the extract was saponified prior to chromatographic analysis, following the previously explained protocol. As already mentioned, solvent mixture b) (DE:MetOH:tBME (30:35:35)) offered the best results and was therefore used to extract carotenoids from the remaining digested food samples.
Using the β-cryptoxanthin results obtained from the digested orange extract analysed following each of the previously mentioned protocols, percentage of BA was calculated according to equation (1) as the ratio between the concentration of carotenoid in the micellar aqueous phase (supernatant) and its respective concentration in the fruit. The main findings regarding β-cryptoxanthin BA in orange showed a significant difference between the two protocols: 1.1% (with protocol 1) and 47.1% (with protocol 2). Protocol 2 yielded better BA values and was therefore used for the digestion of the remaining food samples.
(Nutrient content in the digested food/Nutrient content in the food) x 100
2.5. Digestion of Foods
We adapted a standardized static in vitro digestion model, originally proposed for food by the INFOGEST COST action in a consensus paper [8], to specifically evaluate carotenoid BA. Thus, the previously named "protocol 2" was followed to perform the digestion of the selected foods. Initially, eight 20 g samples of each food (which had been homogenized using a blender) were prepared to begin the digestion process. At each stage, two samples were removed to analyse the carotenoid content without centrifugation (digesta - a mixture of liquids, enzymes, and partially digested food) and to study the stability of carotenoids (and any possible alterations these compounds might undergo during in vitro digestion). Extraction of the carotenoids was performed using DE:MetOH:tBME (30:35:35), and analysis was done by HPLC. This yielded two samples from the oral phase, two from the gastric phase, two from the intestinal phase, and two that underwent complete digestion (mixed micelles). These last two samples were subjected to optimal centrifugation conditions determined through tests with kiwi (20,000 G for 5 minutes).
3. Results
3.1. Carotenoids Content in Fruits and Vegetables
The analysis of the undigested carotenoid content across various fruits and vegetables revealed distinct profiles for each (Figure 2, Table 1).
Avocado was found to be a notable source of both lutein, with 171.67 ug/100 g, and violaxanthin, at 128.80 ug/100 g. Kiwi presented a moderate carotenoid profile, with lutein (64.55 ug/100 g) being more abundant than β-carotene (35.50 ug/100 g). Orange, while containing a broader spectrum of carotenoids, showed its highest concentration in β-cryptoxanthin (73.39 ug/100 g) and violaxanthin (51.03 ug/100 g), with zeaxanthin and phytoene present in smaller quantities (12.85 ug/100 g and 7.33 ug/100g respectively). Finally, lamb's lettuce stood out as exceptionally rich in lutein (4615.03 ug/100 g) and β-carotene (2156.42 ug/100 g). It also contained substantial amounts of violaxanthin (1535.81 ug/100 g) and neoxanthin (940.31 ug/100 g), indicating a diverse and high carotenoid content.
3.2. Stability and BA of Carotenoids After the Digestion Process
The stability and BA of the most abundant carotenoids found in the foods are shown in Table 2 and Figure 3. The recovery (%) and loss (%) of each main carotenoid in the digesta fraction provides information about the stability of the carotenoids during in vitro gastrointestinal digestion.
Initial findings revealed that the analysed fruits contained higher levels of carotenoids compared to the digested ones, suggesting that these compounds were partially lost during the in vitro digestion. In avocado, lutein exhibits high recovery after the oral phase (83.41%) and the duodenal phase (95.84%), although it drops in the gastric phase (31.14%). Violaxanthin in avocado shows low oral recovery (32.07%) and high loss (67.93%). In orange, violaxanthin experiences considerable loss, reaching 81.52% by the end of digestion. Zeaxanthin shows a "recovery" above 100% in the oral and gastric phases, which could indicate release or transformation. β-cryptoxanthin in orange maintains moderate recovery, while phytoene shows high loss. Kiwi demonstrates good stability for both lutein and β-carotene. Lutein has consistently high recovery (around 73-80%) and low losses across all phases. β-carotene in kiwi also shows high initial recovery (99.75%), though it decreases to 63.44% in the duodenal phase. In contrast, lamb's lettuce exhibits low recoveries and high losses for most of its carotenoids. Neoxanthin and violaxanthin have very low recoveries and losses exceeding 66% and 80% respectively. Lutein and β-carotene in lamb's lettuce also show very high losses, surpassing 83% by the end of digestion.
The BA results reveal significant variability among the carotenoids and food matrices studied (Figure 3). In avocado, lutein exhibits a notably high BA of 55.32%, indicating good availability for absorption. Violaxanthin in avocado also shows reasonable BA at 24.80%. In orange, the BA of violaxanthin is very low, only 1.02%, suggesting poor release or micellar incorporation. Zeaxanthin in orange shows a BA of 29.96%, while β-cryptoxanthin and phytoene present intermediate values of 18.03% and 19.78% respectively. Kiwi stands out for the high BA of lutein, reaching 71.01%, making it an excellent source of this bioavailable carotenoid. β-carotene in kiwi also demonstrates significant BA at 36.39%. In contrast, lamb's lettuce generally shows low BA for its carotenoids. Violaxanthin and lutein have bioaccessibilities of 17.66% and 16.12% respectively. β-carotene in lamb's lettuce exhibits particularly low BA, with only 5.08%, implying that only a small fraction of this carotenoid is potentially absorbable.
4. Discussion
The present study includes the in vitro digestion analysis of three fruits (avocado, kiwi, orange) and lamb's lettuce, which were used in a dietary intervention in healthy subjects to assess the effect on lutein status markers [10]. Specifically, these foods were used because they are rich in lutein or are good contributors to lutein intake in the Spanish population [15]. The carotenoid content in the analysed foods aligned with reported values in scientific literature, while also exhibiting expected variations due to factors such as specific cultivar, ripeness, growing conditions, and analytical methodologies [16].
The carotenoid profile observed in the studied fruits and vegetables highlight notable compositional differences and reinforces their relevance as sources of these bioactive compounds. Hass avocado is a notable source of carotenoids like lutein and violaxanthin. It's important to note that the concentration of these compounds can vary significantly due to factors such as variety, ripeness, growing conditions, and even the specific sampling area within the fruit [17,18]. Post-harvest handling can also lead to a decrease in certain carotenoids, with neoxanthin experiencing the most pronounced loss when stored at 20 °C [19]. The carotenoid levels in Hass avocados are also closely linked to their total fat content, which can vary considerably [20].
In citrus fruits such as Nave-Late oranges major identified carotenoids include β-cryptoxanthin, violaxanthin, zeaxanthin, and phytoene. Although lutein is commonly reported in oranges [21], it was not detected in the samples analysed in this study. Studies on oranges consistently identify β-cryptoxanthin as a major carotenoid, though significant variation exists depending on the maturity stage and variety [22]. For instance, while violaxanthin can be the predominant carotenoid in mature Navel and Valencia oranges, accumulating significantly during ripening phytoene and phytofluene may be present in trace amounts or not detected, indicating their minimal contribution to the carotenoid profile of these standard varieties [23].
Hayward kiwi also demonstrated relevant concentrations of lutein and β-carotene, supporting its role as a dietary source of these carotenoids. Previous studies across orchards in central Italy found lutein to be consistently present at higher levels than β-carotene, although variation among orchards was notable [24]. Environmental conditions, orchard microclimates, and handling practices after harvest were all considered influential factors affecting final composition.
Finally, lamb's lettuce samples show very high concentrations of lutein and β-carotene, being a vegetable widely recognized as an excellent source of these carotenoids. While values can vary when compared to other leafy greens like spinach, where lutein is also a predominant carotenoid along with violaxanthin and all-E-neoxanthin [25,26], lamb's lettuce can be considered as a carotenoid-rich leafy green.
4.1. Selection of Digestion Protocol and the Extraction Method for Carotenoids from Digested Foods
In vitro digestion models, such as the INFOGEST protocol [8,27] are valuable tools for estimating nutrient BA. However, their application to lipophilic compounds like carotenoids presents specific challenges. Despite standardization efforts, the literature reveals that the original INFOGEST protocol is not always optimal for precisely evaluating carotenoid BA, particularly due to the nature of carotenoid esters and the critical need for efficient separation of the micellar fraction [28,29]. For these reasons, and to optimize the assessment of carotenoid BA in the analysed fruits and vegetables, several modifications were implemented to the standardized INFOGEST protocol, and two different protocols were tested for the in vitro digestion of oranges. In both protocols, and although not appearing in the protocol proposed by INFOGEST [8,27], considering that carotenoid esters are not detected in vivo under usual dietary conditions [4] and are poorly hydrolysed under the conditions of the INFOGEST protocol adapted for carotenoids, cholesterol esterase was included during the gastric phase of digestion [28,30]. Furthermore, the original INFOGEST protocol recommends using 5 grams of solid food or 5 mL of liquid food as the initial sample for in vitro digestion. This amount is mixed with an equal volume of simulated salivary fluid (SSF), meaning a 1:1 weight/volume ratio between food and SSF during the oral phase. In protocol 2, a larger food sample (20 g) was used to achieve higher absorbance values and reduce analytical errors. This larger quantity has also been employed by other authors [28] to evaluate carotenoid BA in tropical fruits. On the other hand, the INFOGEST protocol also does not specify exact centrifugation parameters such as speed or duration, leaving this open to adjustments depending on the sample type and the study's objective. Centrifugation parameters, specifically speed and duration, exert a critical influence on the accurate assessment of carotenoid BA in static in vitro digestion models [30]. The physical forces applied during centrifugation can impact the stability of fragile mixed micelles and lipid droplets, potentially altering their integrity and, thus, the amount of carotenoids effectively transferred into the aqueous phase [29]. Insufficient centrifugal force or time can result in an incomplete sedimentation of solid particles and unmicellized carotenoid aggregates. This leads to a supernatant contaminated with non-bioaccessible forms, thereby artificially inflating the measured BA values. Conversely, overly aggressive centrifugation (excessively high speeds or prolonged durations) risks the unintended sedimentation of larger, yet physiologically relevant, mixed micelle structures or lipid droplets, which would consequently underestimate the true bioavailable fraction of carotenoids [31,32]. For this reason, the chosen centrifugation parameters directly dictate the purity and composition of the separated supernatant, which in turn profoundly affects the efficiency and precision of subsequent carotenoid extraction and quantification, making proper optimization crucial for reliable BA data [29,31]. In this context, Granado-Lorencio et al. [9] compared low-speed centrifugation with overnight static decantation at room temperature and observed that the latter provided more representative BA estimations in fruits and vegetables, particularly in matrices with high viscosity or fibre content. They concluded that centrifugation might underestimate carotenoid recovery due to the unintended sedimentation of relevant mixed micelles and lipid droplets. In the present study, protocol 1 employed the conditions previously used in our laboratory [12], while protocol 2 applied the conditions that yielded the best results (20,000 G and 5 min) after conducting multiple centrifugation trials with a digested kiwi sample under this protocol to compare the recovery of free and esterified carotenoids in the supernatants. These conditions are consistent with those used by other authors and are recommended when applying the INFOGEST protocol adapted to carotenoids [5].
It’s worth mentioning that, in this study, centrifugation was employed to obtain the supernatant containing carotenoids released from the food matrix, and no filtration step was used to separate micelles specifically. Several previous works have opted for direct extraction from the supernatant without micelle isolation, particularly when dealing with complex food systems or when the goal was to compare relative BA across experimental treatments [33,34,35]. Our decision to omit filtration aligns with established methodology in carotenoid research and was motivated by the importance of maintaining sample integrity for later analytical procedures. Although this strategy may slightly compromise the precision of absolute BA values, it remains a valid approach for comparative assessments of carotenoid liberation during digestion.
In conclusion, when applying the standardized static in vitro digestion model for food to assess carotenoid BA, a higher sample weight and increased centrifugation speed for separating the micellar fraction should be considered to obtain precise and improved results. The centrifugation step is particularly crucial for obtaining the fraction containing micellized carotenoids.
4.2. Stability of Carotenoids During Digestion
This study examines the recovery of specific carotenoids (neoxanthin, violaxanthin, lutein, zeaxanthin, β-cryptoxanthin, phytoene, and β-carotene) in avocado, orange, kiwi, and lamb's lettuce throughout the in vitro digestion phases (oral, oral + gastric and complete gastrointestinal phase), as well as their BA. The results, presented in Table 2, highlight the complex interaction between food matrix structure, carotenoid type, and digestive conditions—factors that significantly influence their fate.
In general, carotenoid recovery showed a decreasing trend as the digestion process progressed, which aligns with the inherent degradation of these compounds, known to be sensitive to factors such as pH, enzymatic activity, and oxidation during gastrointestinal transit. This pattern was clearly observed across all foods evaluated, although the extent of loss varied. The gastric phase—characterized by acidic pH and pepsin activity—is often a critical point for carotenoid stability, as previously described in the literature. The duodenal phase, with its elevated pH, presence of oxygen, and the action of bile salts and lipases, may further contribute to carotenoid degradation and isomerization [29]. However, as shown in the study by Granado-Lorencio et al. [30], carotenoid stability during the salivary, gastric, and duodenal phases was quite high, exceeding 75% in most of the foods they analysed (broccoli, spinach, sweet corn, carrot, tomato puree, red pepper, kiwi, orange, pineapple, and loquat), regardless of the type of food matrix (green or non-green vegetable or fruit), which supports the remarkable resistance of these compounds to digestive conditions.
In avocado, lutein showed a notable fluctuation in its recovery (and associated loss), with a remarkable increase in its recovery up to 95.84% in the complete digestion phase. This recovery could suggest a delayed release or a matrix isomerization/restructuring that would allow for its greater extraction and subsequent detection. Violaxanthin, on the other hand, showed a low initial oral recovery (32.07%) and was not detected in subsequent phases. It is important to consider that violaxanthin is an epoxide carotenoid, susceptible to acid hydrolysis and non-enzymatic transformations during digestion. These reactions may lead to the formation of derivatives such as auroxanthin or mutatoxanthin, which are not always detectable unless specific techniques like HPLC-MS/MS are employed [36].
The orange exhibited a varied behaviour among its carotenoids. Zeaxanthin demonstrated notable stability, with an apparent increase during the oral + gastric phase (125.29%), later stabilizing at 84.44% in the complete gastrointestinal phase. This increase could indicate efficient release from its matrix, or enhanced analytical extraction compared to the initial sample. In contrast to previous findings in citrus (mandarin) [28], β-cryptoxanthin experienced greater loss in the final digestion phase (49.65%) than in the gastric phase (26.57%). This behaviour is consistent with what has been described for its esterified forms (also in mandarin) [28]. Given the limited lipase activity during the gastric phase, it is plausible that the esters remained relatively intact during simulated digestion and were subsequently hydrolysed during the analytical saponification process. This would favour the release of the free form, leading to greater detection in extracts from the gastric phase. Data from Granado-Lorencio et al. [9] support this hypothesis, showing that β-cryptoxanthin esters require cholesterol esterase for effective hydrolysis, as human pancreatic lipase alone was insufficient to efficiently release the free form. However, it's also important to note that the type of esterase might have limited specificity in hydrolysing different carotenoid esters. While the hydrolysis of xanthophyll esters (like lutein and zeaxanthin) was over 90% in foods like pineapple, it did not reach 40% in most food matrices—particularly low in fruits such as loquat and orange—highlighting the influence of matrix type on the release of free forms. Phytoene showed significant degradation from the beginning of the digestive process, with limited recovery in the oral phase (29%) and losses around 70% across all phases. This early degradation may be attributed to the low chemical stability of phytoene under oxidative and enzymatic conditions, as well as its limited structural protection within the orange matrix. Unlike more conjugated carotenoids, phytoene has fewer conjugated double bonds, making it more susceptible to oxidation in the presence of oxygen and to digestive enzymes [37,38]. Additionally, its low affinity for lipid structures could hinder its incorporation into mixed micelles, reducing its protection against intestinal degradation [39]. Violaxanthin, as observed in avocado, suffered significant loss from the oral to intestinal phases. In line with this, Granado-Lorencio et al. [9] described a notable decrease in epoxidized carotenoids during the gastric phase, which was particularly critical for compounds such as violaxanthin and neoxanthin. This degradation was attributed to acid hydrolysis and epoxide group rearrangement, leading to new derivatives that may not be detected through conventional chromatographic methods.
In kiwi, lutein exhibited high stability throughout the in vitro digestive process, with recoveries of 80.33% in the oral phase, 78.33% in the oral + gastric phase, and 73.71% in the complete gastrointestinal phase. This remarkable stability may be attributed to kiwi’s cellular structure, which is rich in bioactive compounds and relatively soft, facilitating carotenoid release, as well as the presence of actinidin, a proteolytic enzyme that may aid in the disruption of the protein matrix [40]. Meanwhile, β-carotene also showed excellent initial recovery (99.75% in the oral phase), followed by a moderate loss in the complete gastrointestinal phase (36.56%), suggesting some susceptibility to degradation. This high initial recovery could be linked to kiwi's soft cellular structure and low insoluble fibre content, which facilitates mechanical disruption and carotenoid release during the oral phase. However, the moderate loss in the complete gastrointestinal phase (36.56%) suggests some susceptibility of β-carotene to degradation under more aggressive physiological conditions, such as the presence of bile salts and pancreatic enzymes. These results are consistent with those described by Granado-Lorencio et al. [30], except for the β-carotene loss in the duodenal phase. They observed over 80% stability for lutein and over 90% for β-carotene in kiwi during the salivary, gastric, and duodenal phases, confirming their resistance to simulated digestive conditions.
Lamb's lettuce exhibited the lowest overall stability for all detected carotenoids. Lutein showed very low recovery (27.03% oral, 12.07% oral + gastric and 13.58% complete gastrointestinal phase). Similarly, violaxanthin (20.05% complete gastrointestinal phase) and β-carotene (16.14% complete gastrointestinal phase) demonstrated significant reductions. Studies conducted on other leafy green vegetables, such as spinach, have also shown high loss values for these carotenoids when their fresh forms were subjected to digestion. In this regard, lutein, β-carotene, and violaxanthin exhibited, respectively, 24%, 60%, and 95% losses in the study by Courraud et al. [41], and losses of 76% (lutein) and 50% (β-carotene) in the work by Rich et al. [42]. Granado-Lorencio et al. [30] described that lettuce showed the lowest carotenoid stability during in vitro digestion, with losses exceeding 25% in all phases, which aligns with the low values observed in lamb's lettuce. As for neoxanthin, it was detected only in the oral phase of the in vitro digestion process, while it was not observed in the oral + gastric or complete gastrointestinal phases. This behaviour can be attributed to its high structural sensitivity to aggressive digestive conditions. In particular, the acidic environment of the gastric phase favours the opening of the epoxide ring and the transformation of neoxanthin into derivatives like neochromes, which are not detectable by conventional chromatographic analysis methods [43,44]. Additionally, the action of pancreatic enzymes and bile salts in the duodenal phase can promote further degradation or hinder its efficient incorporation into micelles, complicating its subsequent recovery [45]. In contrast, during the oral phase—characterized by a neutral pH and the absence of intensive digestive enzymes—neoxanthin remains stable, allowing for its detection.
In general terms, lutein (in avocado and kiwi) and β-carotene (in kiwi) are more stable in fruits than in lamb's lettuce, whereas violaxanthin exhibits very low stability in both types of food.
4.3. Carotenoids BA
The findings in the present work demonstrate a generally higher BA for lutein in fruits (avocado, orange, and kiwi) compared to leafy green vegetables (lamb's lettuce), which is consistent with existing literature [46]. While the carotenoid content in vegetables is typically higher than in fruits, especially for lutein, β-carotene, neoxanthin, and violaxanthin, numerous studies have reported better carotenoid transfer to micelles from fruits than from vegetables. Xanthophylls (lutein, zeaxanthin, β-cryptoxanthin) are often highly bioaccessible in fruits (ranging from 50% to 100%), possibly due to their localization in chromoplasts dissolved within oil droplets, which may facilitate their release [46]. This pattern was confirmed by Granado-Lorencio et al. [30], who observed that green vegetables such as spinach, lettuce, and broccoli exhibited lower transfer rates of lutein and β-carotene to the micellar phase compared to fruits like kiwi, orange, and loquat. These differences were attributed to the intracellular localization of carotenoids (e.g., in chloroplasts) and the presence of dietary fibres that limit their release. Accordingly, positive correlations were identified between the fat content of foods and the BA of carotenoids like lutein and β-carotene, while negative correlations with fibre content. Although these differences were not statistically significant in all cases, they suggest that lipid composition may favour the incorporation of carotenoids into mixed micelles.
In avocado, the BA values for lutein and violaxanthin are notably high (55.32% and 24.80%, respectively). As an oleaginous fruit, avocado contains lipid-soluble carotenoids embedded in a lipid-rich matrix composed primarily of monounsaturated fatty acids and phospholipids [47], which facilitates their dispersion and subsequent micellization during digestion—a critical step for intestinal absorption. However, it should be noted that the fruit ripening stage affects the bioavailability of fat-soluble carotenoids from avocado; therefore, these values may vary depending on the maturity stage of the analysed sample [48].
Although kiwi is not a lipid-rich fruit like avocado, its matrix structure may play a crucial role in carotenoid BA. The specific composition of the cell wall and the organization of chromoplasts where carotenoids are stored in kiwi could permit efficient release, as the progressive solubilization of pectic polysaccharides and loss of galactose during ripening loosens the cell wall matrix, while the differentiation of chromoplasts into carotenoid-rich crystalline or globular structures enhances BA [49]. Furthermore, lutein in fruits is often found in free or esterified forms with fatty acids, and the latter might be more bioavailable than unesterified forms, depending on the enzymatic capacity for hydrolysis in the intestine. In the study by Granado-Lorencio et al. [30], lutein in kiwi exhibited a 59% incorporation into the micellar phase, while β-carotene reached 35%, confirming their relatively high BA compared to other fruits evaluated. In the present study, lutein BA was considerably higher (71.01%) than the values reported for other non-fatty fruits, in which intestinal BA of lutein typically remains below 50% [30]. Regarding β-carotene, BA was lower than that observed for lutein (36.39%). This value aligns with previous studies such as Mapelli-Brahm et al. [38], who reported intestinal BA of β-carotene below 50% in non-fatty fruits like mango and papaya, and highlighted that food matrix and processing degree significantly influence carotenoid stability. Similarly, Reboul et al. [37] noted that although β-carotene is more stable than other oxygenated carotenoids, it can undergo oxidation and isomerization during intestinal digestion, thereby affecting its micellar incorporation. Within this context, kiwi stands out as an efficient source of β-carotene, demonstrating near-complete initial release and moderate intestinal degradation, making it a promising fruit to enhance intake of lipophilic carotenoids in low-fat diets.
On the other hand, orange exhibits more variable behaviour. Violaxanthin shows a low BA value (1.01%), while zeaxanthin (29.96%), β-cryptoxanthin (18.03%), and phytoene (19.76%) display somewhat higher values. The generally low BA observed may be related to the potential degradation of more labile carotenoids (such as epoxides), or to interactions with matrix components that sequester them as digestion progresses. The presence of pectins and other non-starch polysaccharides in orange may form a network that, while facilitating initial release, could hinder complete micellization of certain carotenoids during the final phases [50]. In this regard, Granado-Lorencio et al. [9] observed that the hydrolysis of xanthophyll esters like β-cryptoxanthin in orange was incomplete (<40%), which limits the availability of the free form for micellar incorporation. Indeed, consistent with our results, these authors described micellar BA below 40% for this carotenoid. Phytoene stands out for its relatively high BA, frequently reported as having comparable or even higher bioaccessibilities than xanthophylls. The value obtained in the present study (19.78%) aligns with those reported for mandarin cv. Clementine (C. clementina) (20–30%) [49] and mandarin cvs. ‘Ponkan’, ‘Rio’, and ‘Murcott’ (22–32%) [28]. In contrast, studies conducted on more processed matrices such as tomato or apricot have described significantly higher phytoene BA (up to 97%), suggesting that the intact cellular structure of whole orange may limit both its release and stability [38]. These results reinforce the need to consider the food matrix and digestive environment when evaluating the BA of linear carotenoids like phytoene. The study by Granado-Lorencio et al. [30] also showed that phytoene BA in fruits like orange was below 20%, further emphasizing the influence of the food matrix on micellization efficiency.
For lamb’s lettuce the BA of both lutein and β-carotene showed low values (16.12% and 5.08%, respectively). This aligns with previously reported data, which demonstrate that, compared to fruits, dark green leafy vegetables tend to exhibit significantly lower carotenoid BA (typically ranging between 5–10%). This phenomenon may be attributed to the strong association of carotenoids with chloroplast protein structures, which are encapsulated by dense cell walls rich in fibre, acting as a physical barrier that limits carotenoid release into the intestinal lumen during digestion. Furthermore, the low lipid content in these vegetables may also reduce micellization efficiency [30,46,52]. In addition, the presence of dietary fibre, pectin, and mucilaginous compounds can increase chyme viscosity, thereby decreasing the diffusion of liberated carotenoids and impeding micelle formation, which further limits their BA. For this reason, processing techniques such as steaming or oil addition can significantly improve this BA [53,54].
In summary, the BA findings found in the present study demonstrate that lutein is more bioaccessible than β-carotene in both lamb's lettuce (16.12% vs. 5.08%) and kiwi (71.01% vs. 36.39%). This aligns with previous studies reporting higher BA for xanthophylls (lutein, zeaxanthin, violaxanthin, neoxanthin) compared to carotenes (β-carotene) [46,55]. Furthermore, the high BA of lutein in avocado and kiwi is consistent with the hypothesis that lipids (in avocado) and matrix structure (in kiwi) facilitate its release. Other investigations into lutein bioavailability from various plant sources confirm that dietary sources differ in their capacity to elevate plasma lutein levels, a variability attributed to differences in BA based on the specific food matrix [30,56].
5. Conclusions
This study has demonstrated the critical importance of adapting standardized in vitro digestion protocols, such as that from the INFOGEST, for the accurate evaluation of the BA of lipophilic compounds like carotenoids. Our findings confirm that specific modifications, including increasing the sample weight and optimizing centrifugation conditions to 20,000 G and 5 min., are essential to enhance the release and micellarization of carotenoids, more accurately reflecting their digestive fate.
The results obtained underscore the inherent variability in carotenoid BA, which depends not only on the type of carotenoid but also, crucially, on the food matrix in which they are embedded [30]. Thus, several key elements dictate the efficiency with which these bioactive compounds are released from the food matrix and become available for micelle formation and subsequent intestinal absorption. These include the lipid composition of the food, the integrity and composition of the cell wall (including fiber, pectin, and mucilage), the intracellular localization of carotenoids (chromoplasts/chloroplasts), and whether xanthophylls are in free or esterified form. For instance, while violaxanthin was present in notable amounts in both avocado and orange, its observed BA from orange was notably low. Likewise, significantly higher lutein BA was observed in kiwi and avocado compared to lamb's lettuce.
In summary, this work contributes to advancing in vitro methodology in carotenoid research, providing evidence for key adaptations to obtain more robust and physiologically relevant BA data. These insights are fundamental for future nutritional research, the development of optimized dietary strategies for carotenoid intake, and the formulation of functional foods with improved BA.
Author Contributions
Conceptualization, B.O.A.; methodology, B.O.A, E.R.R, R.S.E., M.S.P; formal analysis, E.R.R.; investigation, E.R.R, R.S.E., M.S.P.; resources, B.O:A.; data curation, E.R.R.; writing—original draft preparation, E.R.R.; writing—review and editing, B.O.A.; supervision, B.O:A.; project administration, B.O.A.; funding acquisition, B.O:A. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by a grant from the Instituto de Salud Carlos III (Ministerio de Ciencia e Innovación, Spain) (grant PI16/01991) and co-funded by the ERDF/European Social Fund.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
During the preparation of this manuscript/study, the author(s) used Gemini and Copilot for the purpose of creating images for the graphical abstract. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| O | Oral phase |
| G | Gastric phase |
| D | Duodenal phase |
| BA | Bioaccessibility |
References
- Rodríguez-Concepción, M.; Ávalos, F.; Bonet, M.L.; Boronat, A.; Gómez-Gómez, L.; Hornero-Méndez, D.; Limón, C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef]
- Chew, E.Y.; Clemons, T.E.; Agrón, E.; Domalpally, A.; Keenan, T.D.L.; Vitale, S.; Weber, C.; Smith, D.C.; Christen, W.; AREDS2 Research Group. Long-term Outcomes of Adding Lutein/Zeaxanthin and ω-3 Fatty Acids to the AREDS Supplements on Age-Related Macular Degeneration Progression: AREDS2 Report 28. JAMA Ophthalmol. 2022, 140, 692–698. [Google Scholar] [CrossRef]
- Böhm, V.; Lietz, G.; Olmedilla-Alonso, B.; Phelan, D.; Reboul, E.; Bánati, D.; Borel, P.; Corte-Real, J.; de Lera, A.R.; Desmarchelier, C.; et al. From carotenoid intake to carotenoid blood and tissue concentrations—Implications for dietary intake recommendations. Nutr. Rev. 2021, 79, 544–573. [Google Scholar] [CrossRef]
- Olmedilla-Alonso, B.; Granado-Lorencio, F.; Castro-Feito, J.; Herrero-Barbudo, C.; Blanco-Navarro, I.; Estévez-Santiago, R. Bioavailability of lutein from marigold flowers (free vs ester forms): A randomised cross-over study to assess serum response and visual contrast threshold in adults. Nutrients 2024, 16, 1415. [Google Scholar] [CrossRef]
- Xavier, A.A.O.; Mercadante, A.Z. A guide for the evaluation of in vitro bioaccessibility of carotenoids. Methods Enzymol. 2022, 674, 297–327. [Google Scholar] [CrossRef] [PubMed]
- Corte-Real, J.; Iddir, M.; Soukoulis, C.; Richling, E.; Hoffmann, L.; Bohn, T. Effect of divalent minerals on the bioaccessibility of pure carotenoids and on physical properties of gastro-intestinal fluids. Food Chem. 2016, 197, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Iddir, M.; Porras Yaruro, J.F.; Larondelle, Y.; Bohn, T. Gastric lipase can significantly increase lipolysis and carotenoid bioaccessibility from plant food matrices in the harmonized INFOGEST static in vitro digestion model. Food Funct. 2021, 12, 9043–9053. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Granado-Lorencio, F.; Olmedilla-Alonso, B.; Herrero-Barbudo, C.; Blanco-Navarro, I.; Pérez-Sacristán, B.; Blázquez-García, S. In vitro bioaccessibility of carotenoids and tocopherols from fruits and vegetables. Food Chem. 2007, 102, 641–648. [Google Scholar] [CrossRef]
- Olmedilla-Alonso, B.; Rodríguez-Rodríguez, E.; Beltrán-de-Miguel, B.; Sánchez-Prieto, M.; Estévez-Santiago, R. Changes in Lutein Status Markers (Serum and Faecal Concentrations, Macular Pigment) in Response to a Lutein-Rich Fruit or Vegetable (Three Pieces/Day) Dietary Intervention in Normolipemic Subjects. Nutrients 2021, 13, 3614. [Google Scholar] [CrossRef] [PubMed]
- de Rosso, V.V.; Mercadante, A.Z. Identification and quantification of carotenoids, by HPLC-PDA-MS/MS, from Amazonian fruits. J. Agric. Food Chem. 2007, 55, 5062–5072. [Google Scholar] [CrossRef]
- Estévez-Santiago, R.; Olmedilla-Alonso, B.; Fernández-Jalao, I. Bioaccessibility of provitamin A carotenoids from fruits: application of a standardised static in vitro digestion method. Food Funct. 2016, 7, 1354–1366. [Google Scholar] [CrossRef]
- Granado, F.; Olmedilla, B.; Gil-Martinez, E.; Blanco, I. A Fast, Reliable and Low-cost Saponification Protocol for Analysis of Carotenoids in Vegetables. J. Food Comp. Anal. 2001, 14, 479–489. [Google Scholar] [CrossRef]
- Olmedilla-Alonso, B.; Granado-Lorencio, F.; Blanco-Navarro, I. Carotenoids, retinol and tocopherols in blood: Comparability between serum and plasma (Li-heparin) values. Clin. Biochem. 2005, 38, 444–449. [Google Scholar] [CrossRef]
- Estévez-Santiago, R.; Beltrán-de-Miguel, B.; Olmedilla-Alonso, B. Assessment of dietary lutein, zeaxanthin and lycopene intakes and sources in the Spanish Survey of Dietary Intake (2009-2010). Int. J. Food Sci. Nutr. 2016, 67, 305–313. [Google Scholar] [CrossRef]
- Dias, M.G.; Borge, G.I.A.; Kljak, K.; Mandić, A.I.; Mapelli-Brahm, P.; Olmedilla-Alonso, B.; Pintea, A.M.; Ravasco, F.; Šaponjac, V.T.; Sereikaitė, J.; et al. European database of carotenoid levels in foods. Factors affecting carotenoid content. Foods 2021, 10, 912. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.Y.; Zhang, Y.; Wang, Y.; Wang, D.; Lee, R.P.; Gao, K.; Byrns, R.; Heber, D. California Hass avocado: profiling of carotenoids, tocopherol, fatty acid, and fat content during maturation and from different growing areas. J. Agric. Food Chem. 2009, 57, 10408–10413. [Google Scholar] [CrossRef] [PubMed]
- Ford, N.A.; Spagnuolo, P.; Kraft, J.; Bauer, E. Nutritional Composition of Hass Avocado Pulp. Foods 2023, 12, 2516. [Google Scholar] [CrossRef] [PubMed]
- Ashton, O.B.; Wong, M.; McGhie, T.K.; Vather, R.; Wang, Y.; Requejo-Jackman, C.; Ramankutty, P.; Woolf, A.B. Pigments in avocado tissue and oil. J. Agric. Food Chem. 2006, 54, 10151–10158. [Google Scholar] [CrossRef]
- Lu, Q.Y.; Zhang, Y.; Wang, Y.; Wang, D.; Lee, R.P.; Gao, K.; Byrns, R.; Heber, D. California Hass avocado: Profiling of carotenoids, tocopherol, fatty acid, and fat content during maturation and from different growing areas. J. Agric. Food Chem. 2009, 57, 10408–10413. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.G.; Olmedilla-Alonso, B.; Hornero-Méndez, D.; Mercadante, A.Z.; Osorio, C.; Vargas-Murga, L.; Meléndez-Martínez, A.J. Comprehensive database of carotenoid contents in ibero-american foods. A valuable tool in the context of functional foods and the establishment of recommended intakes of bioactives. J. Agric. Food Chem. 2018, 66, 5055–5107. [Google Scholar] [CrossRef]
- Rodrigo, M.J.; Cilla, A.; Barberá, R.; Zacarías, L. Carotenoid bioaccessibility in pulp and fresh juice from carotenoid-rich sweet oranges and mandarins. Food Funct. 2015, 6, 1950–1959. [Google Scholar] [CrossRef]
- Zacarías-García, J.; Cronje, P.J.; Diretto, G.; Zacarías, L.; Rodrigo, M.J. A comprehensive analysis of carotenoids metabolism in two red-fleshed mutants of Navel and Valencia sweet oranges (Citrus sinensis). Front. Plant Sci. 2022, 13, 1034204. [Google Scholar] [CrossRef]
- D’Evoli, L.; Moscatello, S.; Lucarini, M.; Aguzzi, A.; Gabrielli, P.; Proietti, S.; Battistelli, A.; Famiani, F.; Böhm, V.; Lombardi-Boccia, G. Nutritional traits and antioxidant capacity of kiwifruit (Actinidia deliciosa Planch., cv. Hayward) grown in Italy. J. Food Comp. Anal. 2015, 37, 25–29. [Google Scholar] [CrossRef]
- Li, H. , Deng, Z., Lu, J., Liu, H., & Chen, F. Effects of High-Temperature Frying of Spinach Leaves in Sunflower Oil on Carotenoids, Chlorophylls, and Tocopherol Composition. Front. Chem. 2017, 5, 19. [Google Scholar] [CrossRef]
- Bunea, A.; Andjelkovic, M.; Socaciu, C.; Bobis, O.; Neacsu, M.; Verhé, R.; Van Camp. Total and individual carotenoids and phenolic acids content in fresh, refrigerated and processed spinach (Spinacia oleracea L.). Food Chem. 2008, 108, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Petry, F.C.; Mercadante, A.Z. Impact of in vitro digestion phases on the stability and bioaccessibility of carotenoids and their esters in mandarin pulps. Food Funct. 2017, 8, 3951–3963. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.B.; Mariutti, L.R.; Mercadante, A.Z. An in vitro digestion method adapted for carotenoids and carotenoid esters: moving forward towards standardization. Food Funct. 2016, 7, 4992–5001. [Google Scholar] [CrossRef]
- Granado-Lorencio, F.; Olmedilla-Alonso, B.; Herrero-Barbudo, C.; Pérez-Sacristán, B.; Blanco-Navarro, I.; Blázquez-García, S. Comparative in vitro bioaccessibility of carotenoids from relevant contributors to carotenoid intake. J. Agric. Food Chem. 2007, 55, 10887–10892. [Google Scholar] [CrossRef]
- Petry, F.C.; Mercadante, A.Z. In vitro bioaccessibility of carotenoids from fruits and vegetables: A review. Food Res. Int. 2016, 88, 303–311. [Google Scholar]
- Reboul, E.; Borel, P. Digestion and Absorption of Carotenoids: The Physiologically Relevant Micellar Fraction. Curr. Nutr. Food Sci. 2011, 7, 273–280. [Google Scholar]
- Xu, Y.; Hu, T.; Hu, H.; Xiong, S.; Shi, K.; Zhang, N.; Mu, Q.; Xu, G.; Zhang, P.; Pan, S. Comparative evaluation on the bioaccessibility of citrus fruit carotenoids in vitro based on different intake patterns. Foods 2022, 11, 1457. [Google Scholar] [CrossRef] [PubMed]
- Failla, M.L.; Huo, T.; Thakkar, S.K. In vitro screening of relative bioaccessibility of carotenoids from foods. Asia Pac. J. Clin. Nutr. 2008, 17 (Suppl. 1), 200–203. [Google Scholar]
- Granado, F.; Olmedilla, B.; Herrero, C.; Pérez-Sacristán, B.; Blanco, I.; Blázquez, S. Bioavailability of carotenoids and tocopherols from broccoli: in vivo and in vitro assessment. Exp. Biol. Med. (Maywood) 2006, 231, 1733–1738. [Google Scholar] [CrossRef] [PubMed]
- Shimode, S.; Miyata, K.; Araki, M.; Shindo, K. Antioxidant activities of the antheraxanthin-related carotenoids, antheraxanthin, 9-cis-antheraxanthin, and mutatoxanthins. Oleo Sci. 2018, 67, 977–981. [Google Scholar] [CrossRef]
- Reboul, E.; Margier, M.; Desmarchelier, C.; Halimi, C.; Nowicki, M.; Borel, P.; Mapelli-Brahm, P. Comparison of the bioavailability and intestinal absorption sites of phytoene, phytofluene, lycopene and β-carotene. Food Chem. 2019, 297, 125232. [Google Scholar] [CrossRef]
- Mapelli-Brahm, P.; Corte-Real, J.; Meléndez-Martínez, A.J.; Bohn, T. Bioaccessibility of phytoene and phytofluene is superior to other carotenoids from selected fruit and vegetable juices. Food Chem. 2017, 229, 304–311. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, E.; Estévez-Santiago, R.; Sánchez-Prieto, M.; Olmedilla-Alonso, B. Status and dietary intake of phytoene and phytofluene in Spanish adults and the effect of a four-week dietary intervention. Nutrients 2022, 14, 2922. [Google Scholar] [CrossRef]
- Richardson, D.P.; Ansell, J.; Drummond, L.N. The nutritional and health attributes of kiwifruit: a review. Eur. J. Nutr. 2018, 57, 2659–2676. [Google Scholar] [CrossRef]
- Courraud, J.; Berger, J.; Cristol, J.P.; Avallone, S. Stability and bioaccessibility of different forms of carotenoids and vitamin A during in vitro digestion. Food Chem. 2013, 136, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Rich, G.T.; Bailey, A.L.; Faulks, R.M.; Parker, M.L.; Wickham, M.S.J.; Fillery-Travis, A. Solubilization of carotenoids from carrot juice and spinach in lipid phases: I. Modeling the gastric lumen. Lipids 2003, 38, 933–945. [Google Scholar] [CrossRef]
- Asai, A.; Terasaki, M.; Nagao, A. An epoxide-furanoid rearrangement of spinach neoxanthin occurs in the gastrointestinal tract of mice and in vitro: formation and cytostatic activity of neochrome stereoisomers. J. Nutr. 2004, 134, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Asai, A.; Yonekura, L.; Nagao, A. Low bioavailability of dietary epoxyxanthophylls in humans. Br. J. Nutr. 2008, 100, 273–277. [Google Scholar] [CrossRef]
- Rodrigues, D.B.; Chitchumroonchokchai, C.; Mariutti, L.R.B.; Mercadante, A.Z.; Failla, M.L. Comparison of two static in vitro digestion methods for screening the bioaccessibility of carotenoids in fruits, vegetables, and animal products. J. Agric. Food Chem. 2017, 65, 11220–11228. [Google Scholar] [CrossRef]
- O’Connell, O.; Ryan, L.; O’Brien, N. Xanthophyll carotenoids are more bioaccessible from fruits than dark green vegetables. Nutr. Res. 2007, 27, 258–264. [Google Scholar] [CrossRef]
- Bhuyan, D.J.; Alsherbiny, M.A.; Perera, S.; Low, M.; Basu, A.; Devi, O.A.; Barooah, M.S.; Li, C.G.; Papoutsis, K. The Odyssey of Bioactive Compounds in Avocado (Persea americana) and Their Health Benefits. Antioxidants 2019, 8, 426. [Google Scholar] [CrossRef]
- Cervantes-Paz, B.; Yahia, E.M.; Ornelas-Paz, J.J.; Victoria-Campos, C.I.; Perez-Martinez, J.D.; Reyes-Hernandez, J. Bioaccessibility of fat-soluble bioactive compounds (FSBC) from avocado fruit as affected by ripening and FSBC composition in the food matrix. Food Res. Int. 2021, 139, 109960. [Google Scholar] [CrossRef]
- Bhargava, N.; Ampomah-Dwamena, C.; Voogd, C.; Allan, A.C. Comparative transcriptomic and plastid development analysis sheds light on the differential carotenoid accumulation in kiwifruit flesh. Front. Plant Sci. 2023, 14, 1213086. [Google Scholar] [CrossRef]
- Cervantes-Paz, B.; Ornelas-Paz, J.J.; Ruiz-Cruz, S.; Rios-Velasco, C.; Ibarra-Junquera, V.; Yahia, E.M.; Gardea-Béjar, A.A. Effects of pectin on lipid digestion and possible implications for carotenoid bioavailability during pre-absorptive stages: A review. Food Res. Int. 2017, 99, 917–927. [Google Scholar] [CrossRef]
- Rodrigo, M.J.; Cilla, A.; Barberá, R.; Zacarías, L. Food carotenoid bioaccessibility in pulp and fresh juice from carotenoid-rich sweet oranges and mandarins. Food Funct. 2015, 6, 1950–1959. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Rodriguez-Amaya, D.B. Carotenoid Composition of Hydroponic Leafy Vegetables. J. Agric. Food Chem. 2003, 51, 2603–2607. [Google Scholar] [CrossRef]
- Gärtner, C.; Stahl, W.; Sies, H. Lycopene is more bioavailable from tomato paste than from fresh tomatoes. Am. J. Clin. Nutr. 1997, 66, 116–122. [Google Scholar] [CrossRef]
- Chacón-Ordóñez, T.; Carle, R.; Schweiggert, R. Bioaccessibility of carotenoids from plant and animal foods. J. Sci. Food Agric. 2019, 99, 3220–3239. [Google Scholar] [CrossRef] [PubMed]
- Olmedilla-Alonso, B.; Granado-Lorencio, F.; De Ancos, B.; Sánchez-Moreno, C.; Martín-Belloso, O.; Blanco, I.; Herrero-Barbudo, C.; Elez, P.; Plaza, L.; Cano, M.P. Greater bioavailability of xanthophylls compared to carotenes from orange juice (high-pressure processed, pulsed electric field treated, low-temperature pasteurised, and freshly squeezed) in a crossover study in healthy individuals. Food Chem. 2022; 371. [Google Scholar] [CrossRef]
- Riso, P.; Brusamolino, A.; Ciappellano, S.; Porrini, M. Comparison of lutein bioavailability from vegetables and supplement. Int. J. Vitam. Nutr. Res. 2003, 73, 201–205. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Scheme of two different in vitro digestion protocols tested to assess BA of β-cryptoxanthin from orange (Citrus sinensis, L.).
Figure 1.
Scheme of two different in vitro digestion protocols tested to assess BA of β-cryptoxanthin from orange (Citrus sinensis, L.).
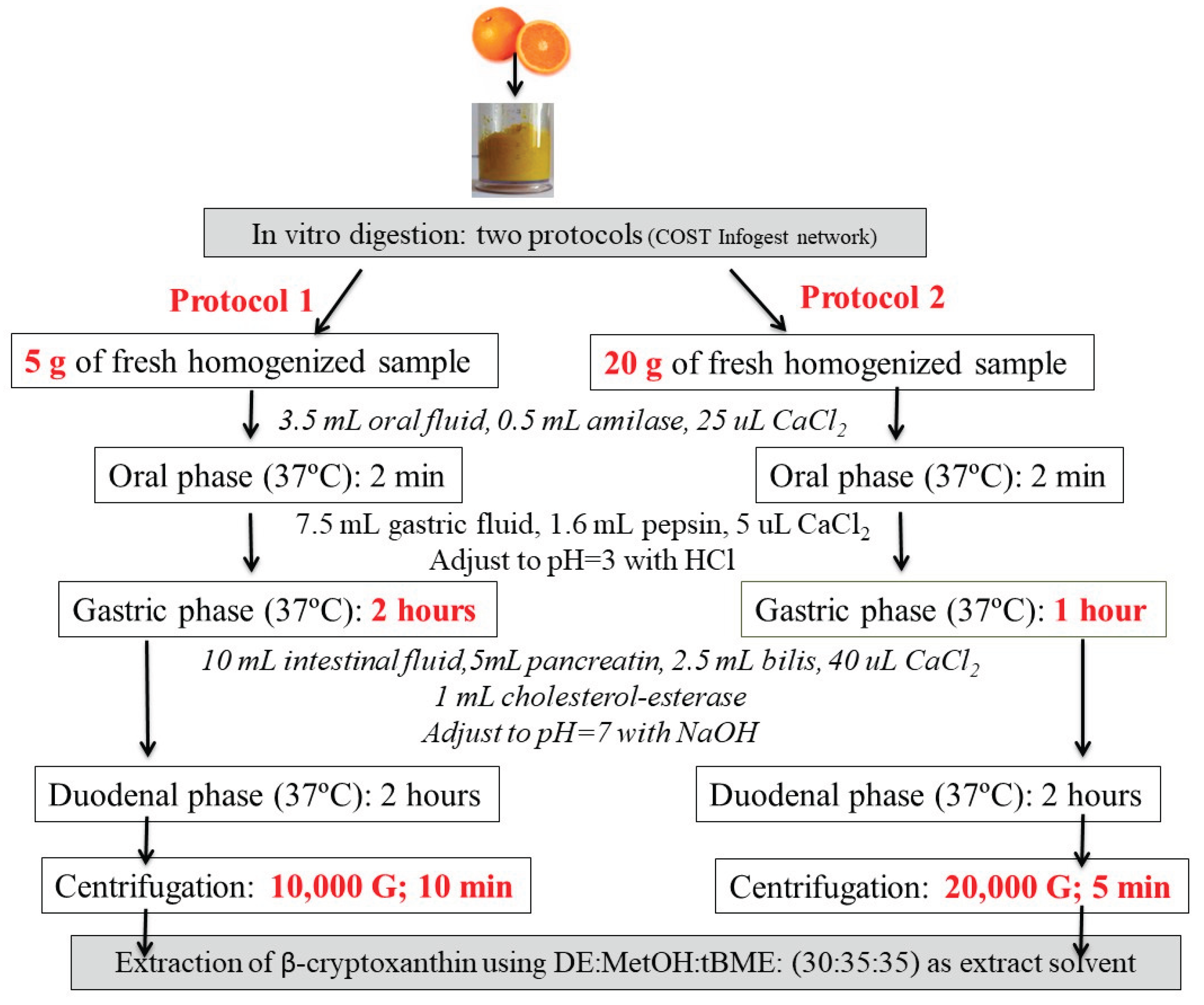
Figure 2.
Chromatograms recorded at 450 nm of: (a) undigested avocado; (b) kiwi; (c) saponified orange; (d) lamb’s lettuce.
Figure 2.
Chromatograms recorded at 450 nm of: (a) undigested avocado; (b) kiwi; (c) saponified orange; (d) lamb’s lettuce.
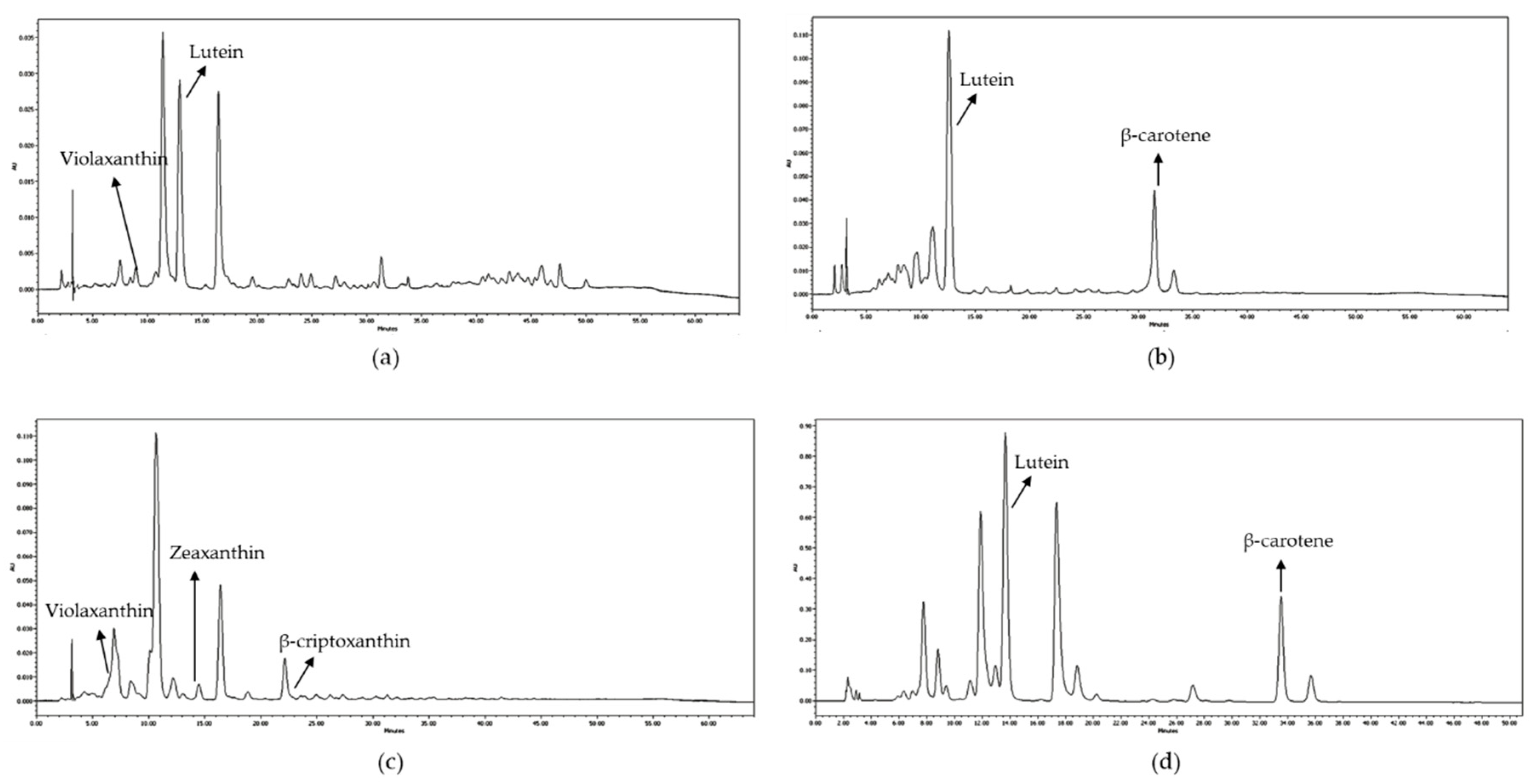
Figure 3.
Bioaccessibility (BA) (%) of the major carotenoids after in vitro digestion in the selected pulps fruits and lamb´s lettuce.
Figure 3.
Bioaccessibility (BA) (%) of the major carotenoids after in vitro digestion in the selected pulps fruits and lamb´s lettuce.
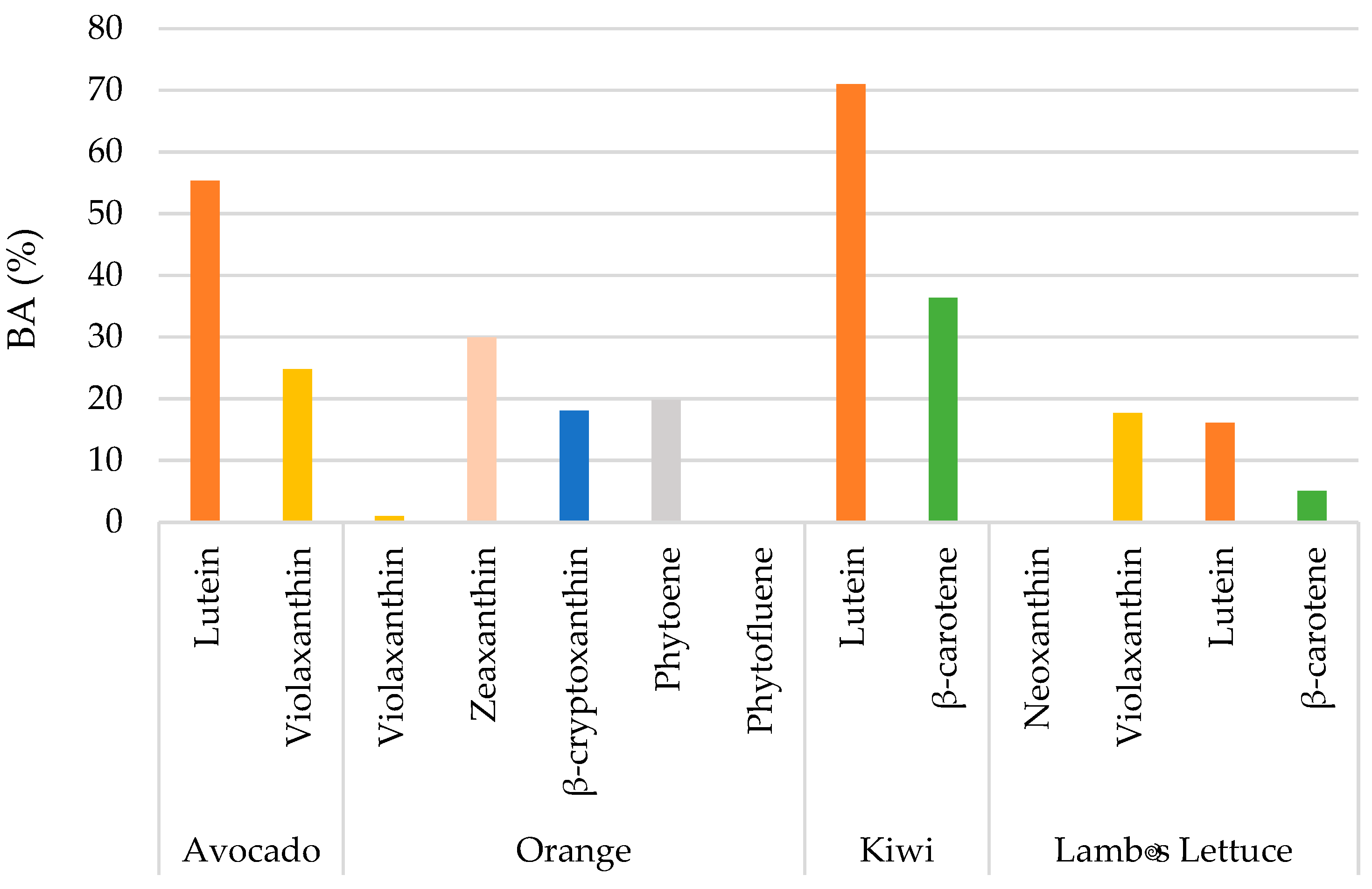

Table 1.
Carotenoids content (µg per 100 g) in the selected pulps fruits and lamb´s lettuce and digested materials after each phase of in vitro digestion.
Table 1.
Carotenoids content (µg per 100 g) in the selected pulps fruits and lamb´s lettuce and digested materials after each phase of in vitro digestion.
| Undigested food | O | O+G | O+G+D | Mixed micelles | |
|---|---|---|---|---|---|
| Avocado | |||||
| Lutein | 171.67 | 143.19 | 53.46 | 164.52 | 94.96 |
| Violaxanthin | 128.8 | 41.31 | n.d. | n.d. | 31.94 |
| Orange | |||||
| Violaxanthin | 51.03 | 44.82 | 19.33 | 9.43 | 0.52 |
| Zeaxanthin | 12.85 | 15.98 | 16.1 | 10.85 | 3.85 |
| β-cryptoxanthin | 73.39 | 54.32 | 53.89 | 36.95 | 13.23 |
| Phytoene | 7.33 | 1.94 | 1.85 | 2.16 | 1.45 |
| Phytofluene | Tr | n.d. | n.d. | n.d. | n.d. |
| Kiwi | |||||
| Lutein | 64.55 | 51.85 | 50.56 | 47.58 | 45.84 |
| β-carotene | 35.5 | 35.41 | 30.42 | 22.52 | 12.92 |
| Lamb´s lettuce | |||||
| Neoxanthin | 940.31 | 315.87 | n.d. | n.d. | n.d. |
| Violaxanthin | 1535.81 | 302.05 | n.d. | 307.93 | 271.19 |
| Lutein | 4615.03 | 1247.42 | 557.02 | 626.74 | 743.82 |
| β-carotene | 2156.42 | 697.5 | 682.08 | 347.97 | 109.54 |
O: oral phase; G: gastric phase; D: duodenal phase; Tr: traces; n.d.: Not detectable.
Table 2.
Recovery (%) and loss (%) of the major carotenoids after each phase of in vitro digestion in the selected pulps fruits and lamb´s lettuce, calculated in comparison to the undigested food matrix.
Table 2.
Recovery (%) and loss (%) of the major carotenoids after each phase of in vitro digestion in the selected pulps fruits and lamb´s lettuce, calculated in comparison to the undigested food matrix.
| Recovery (%) | Loss (%) | |||||
|---|---|---|---|---|---|---|
| O | O+G | O+G+D | O | O+G | O+G+D | |
| Avocado | ||||||
| Lutein | 83.41 | 31.14 | 95.84 | 53.46 | 164.52 | 94.96 |
| Violaxanthin | 32.07 | n.d. | n.d. | n.d. | n.d. | 31.94 |
| Orange | ||||||
| Violaxanthin | 87.83 | 37.88 | 18.48 | 12.17 | 62.12 | 81.52 |
| Zeaxanthin | 124.36 | 125.29 | 84.44 | -24.36 | -25.29 | 15.56 |
| β-cryptoxanthin | 74.02 | 73.43 | 50.35 | 25.98 | 26.57 | 49.65 |
| Phytoene | 26.47 | 25.24 | 29.47 | 73.53 | 74.76 | 70.53 |
| Phytofluene | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Kiwi | ||||||
| Lutein | 80.33 | 78.33 | 73.71 | 19.67 | 21.67 | 26.29 |
| β-carotene | 99.75 | 85.69 | 63.44 | 0.25 | 14.31 | 36.56 |
| Lamb´s lettuce | ||||||
| Neoxanthin | 33.59 | n.d. | n.d. | 66.41 | n.d. | n.d. |
| Violaxanthin | 19.67 | n.d. | 20.05 | 80.33 | n.d. | 79.95 |
| Lutein | 27.03 | 12.07 | 13.58 | 72.97 | 87.93 | 86.42 |
| β-carotene | 32.35 | 31.63 | 16.14 | 67.65 | 68.37 | 83.86 |
O: oral phase; G: gastric phase; D: duodenal phase; n.d.: Not detectable.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



