
Submitted:
27 September 2025
Posted:
29 September 2025
You are already at the latest version
Abstract
Chirality, the handedness of molecules, is a universal feature of biochemistry, yet Earth life exhibits a striking asymmetry: proteins are built from L-amino acids, while nucleic acids incorporate D-sugars. Explaining how this homochirality emerged is a central question in origin-of-life research, with proposed mechanisms ranging from stochastic “frozen accidents” to astrophysical biases, parity violation, and chemical amplification. A complementary approach has recently gained momentum. The synthesis of “mirror” biomolecules, in which every component is built from the opposite enantiomers. Advances in peptide chemistry, nucleic acid synthesis, and enzyme engineering have yielded functional mirror polymerases, ligases, ribosomal proteins, and aptamers, demonstrating that life’s molecular logic is, in principle, invariant under reflection. These achievements not only probe the plausibility of alternative biochemical worlds but also enable practical applications, including nuclease-resistant therapeutics, biostable data storage, and orthogonal biocontained systems. At the same time, the prospect of constructing a self-replicating mirror organism raises profound biosafety and ethical challenges. This review summarizes the current theories of homochirality, highlights milestones in synthetic mirror biology, and evaluates their implications for understanding life’s origins, for the search for extraterrestrial biospheres, and for the future of biotechnology. Together, these insights suggest that life’s chemistry is contingent rather than predetermined, and that mirror life represents both a powerful experimental probe and a transformative technological frontier.
Keywords:
chirality
; homochirality
; mirror life
; enantiomers
; prebiotic chemistry
; synthetic biology
; D-amino acids
; L-DNA
; xenobiology
; origin of life
; astrobiology
; mirror organisms
; Spiegelmers
; racemic crystallography
; orthogonal biocontainment
Introduction
Life’s molecular building blocks exhibit chirality (handedness); existing in left- or right-handed forms that are non-superimposable mirror images, like left vs. right gloves. Terrestrial biology is strikingly homochiral; DNA and RNA incorporate only right-handed sugars, while proteins consist exclusively of left-handed amino acids (Blackmond, 2019; Rohden et al., 2021). “Mirror life” refers to a hypothetical biology built from the opposite enantiomers, essentially a complete mirror-image version of life’s chemistry. The concept dates back to Louis Pasteur, who speculated in 1860 about a “mirror-image world of biology” after discovering molecular asymmetry (Sallembien et al., 2022). Although no mirror-based life has been found in nature, modern advances allow scientists to synthesize mirror versions of biomolecules and even contemplate entire mirror organisms in principle (Wang et al., 2016; Chen et al., 2022; Fan et al., 2021). The chirality of the amino acid alanine is illustrated in Figure 1 as an example.
Chirality in Biological Systems
A molecule is chiral if it has a non-superimposable mirror image. The two mirror forms (enantiomers) have identical chemical formulas but differ in 3D orientation, often labeled “left-handed” (L) or “right-handed” (D) based on how they rotate polarized light or relate to a reference compound. This property is critical in biology; like a left hand vs. right hand, a chiral molecule will interact differently with other chiral structures, a principle analogous to how a left glove fits only a left hand. Life’s chemistry relies on this stereospecific recognition; enzymes, for example, typically bind only substrates of the correct chirality (Blackmond, 2019).
Remarkably, all known life uses only one chirality for each class of fundamental biomolecules. Specifically, virtually all amino acids in proteins are the L-form (left-handed), with rare exceptions like achiral glycine. Organisms built their proteins exclusively from L-amino acids; the D-enantiomers are not used in ribosomal protein synthesis (Rohden et al., 2021; Harrison et al., 2023). The sugars in DNA and RNA (ribose/deoxyribose) are D-form (right-handed). Thus, the sugar-phosphate backbone of nucleic acids has D-chirality. Likewise, many metabolic sugars (glucose, etc.) in biology are D-form (Blackmond, 2019; Sallembien et al., 2022). Many other chiral co-factors, vitamins, and metabolites in cells are also homochiral (e.g., only one enantiomer is biologically active). Even cell membranes reflect chirality, phospholipid glycerol backbones in bacteria/eukaryotes are one enantiomer (sn-glycerol-3-phosphate) while archaea uniquely use the opposite enantiomer; sn-glycerol-1-phosphate (Rohden et al., 2021). Notably, that is one rare case where life evolved the use of both mirror forms in different lineages, though each organism’s membrane is homochiral.
Using uniform handedness is thought to be essential for biopolymers to fold and function. Mixed-chirality polymers (containing both L- and D-monomers) generally do not fold into stable, functional structures (Harrison et al., 2023). For example, a peptide with alternating D- and L-amino acids tends to form irregular structures instead of the well-ordered helices or sheets that pure L-peptides form. Enzymes are highly shape-specific, so a wrong-handed substrate often won’t fit the active site, akin to a left hand not fitting a right-hand glove (Milton et al., 1992; Weinstock et al., 2014). Homochirality enables precise molecular recognition (DNA strands aligning, enzymes binding substrates, etc.), which is likely a prerequisite for self-replication and life’s origin (Blackmond, 2019; Rohden et al., 2021). Indeed, life’s replication machinery (DNA polymerases, ribosomes) requires consistent chirality; a D-sugar backbone in DNA will not pair properly with an L-sugar strand, and an L-amino acid tRNA cannot be read by a ribosome evolved for L-amino acids (Wang et al., 2016; Xu et al., 2017). Comparison of molecular chirality in Earth life versus hypothetical mirror life is summarized in Table 1.
A direct consequence of homochirality is that mirror-image biomolecules are generally not recognized or utilized by Earth biology. For instance, L-glucose (the left-handed form of glucose) tastes sweet like normal D-glucose, but organisms cannot readily metabolize it as an energy source (Vater & Klussmann, 2015). Similarly, our enzymes that act on L-amino acid substrates won’t work on D-amino acids (Milton et al., 1992; Weinstock et al., 2014). This isolation cuts both ways; a “mirror” organism built from D-amino acids and L-sugars would find our nutrients, enzymes, and signals incompatible, and vice versa (Rohden et al., 2021; Harrison et al., 2023). We will later discuss how this fact underlies both potential uses of mirror systems (e.g., molecules that resist biodegradation) and dangers (e.g., mirror pathogens evading immune detection) (Adamala et al., 2024). Chirality in Earth life and hypothetical mirror life comparison is visualized in Figure 2.
Why Does Earth’s Life Use One Handedness?
The dominance of L-amino acids and D-sugars in life raises the question: Was this choice arbitrary or driven by deeper forces? In principle, chemistry in the lab without chiral influences produces racemic mixtures (equal left/right) (Blackmond, 2019). So how and why did biology “break symmetry” to pick one hand? Scientists have explored several hypotheses.
Chance “Frozen Accident”
Life’s chirality may have been a random choice that became locked in. In a racemic primordial soup, if by chance one self-replicating system happened to use L-amino acids and gained a slight foothold, it would propagate that handedness. Competing proto-life using D-amino acids might have been outcompeted or simply failed to arise. This scenario suggests that mirror life was equally likely; Earth could have been a D-amino acid world if a few initial conditions were different (Blackmond, 2019). Many researchers lean toward chance combined with amplification mechanisms to explain homochirality (Sallembien et al., 2022).
Cosmic or Physical Bias
Alternatively, some asymmetry in physics or the environment might have biased early chemistry. One oft-cited idea is circularly polarized light in space (Bailey et al., 1998). Certain astronomical sources emit circularly polarized UV light, which can destroy one enantiomer of chiral molecules slightly more than the other. Meteorites reaching Earth have indeed shown significant enantiomeric excesses; often an excess of the same handedness used by life (e.g., an excess of left-handed amino acids in meteorite samples) (Cronin & Pizzarello, 1997). For example, analyses of carbon-rich meteorites found several with a surplus of L-aspartic and L-glutamic acid (up to ~15% ee), while their D-counterparts were depleted (Cronin & Pizzarello, 1997). This suggests an extraterrestrial asymmetry seeding Earth; “the molecule in excess was the left-handed amino acid that is found exclusively in life on Earth”. Such an excess could have been caused by polarized starlight preferentially destroying the D-form in space (Bailey et al., 1998). If true, early Earth’s organic inventory might already have had a slight L-bias, which life then amplified.
Parity Violation
On a fundamental physics level, the weak nuclear force violates mirror symmetry (P-symmetry). Calculations suggest this could make one enantiomer of certain amino acids ever so slightly more stable (on the order of 1 × 10⁻¹⁷in energy difference) (Quack, 2002). In theory, this parity-violating energy difference might favor L-amino acids and D-sugars (as observed in life). However, the effect is extremely small, and experiments have not confirmed it has any biochemical significance (Quack, 2002). Most scientists consider it unlikely to be the decisive factor in homochirality’s origin, given the vast gap between subatomic energy tweaks and the robust 100% homochirality of biology.
Terrestrial Chemical Processes
A variety of auto-amplification mechanisms have been demonstrated in the lab that can take a tiny initial enantiomeric excess and amplify it to near-pure homochirality. For instance, Frank’s autocatalysis model (1953) showed theoretically that a compound that catalyzes its own production while inhibiting its mirror image can drive a system to an enantiopure state. In 2006, Blackmond and colleagues experimentally achieved such amplification; starting from a small L-excess of an amino acid, they used crystallization cycles to amplify it to a large excess (Blackmond, 2019). They further showed that an L-excess amino acid could induce an excess in sugar chirality in an RNA precursor (Sallembien et al., 2022). Such chemical amplification could have occurred on early Earth in evaporation ponds, crystallizing mixtures, or surface catalysis on chiral minerals (Soai et al., 1995).
Given these possibilities, the prevailing view is that life’s handedness was likely a contingent outcome, a blend of some initial bias (possibly from meteorites or random fluctuation) followed by chemical and biological amplification (Blackmond, 2019; Sallembien et al., 2022). In other words, Earth’s biology might have turned out reversed if the dice had rolled differently. Importantly, nothing in known chemistry absolutely requires L-amino acids over D (or D-sugars over L), the choice could be seen as arbitrary from a purely chemical standpoint. Indeed, experimental models of prebiotic chemistry can produce either enantiomer; one experiment yielded a slight D-excess of certain amino acids under specific conditions, underscoring that homochirality could have gone either way (Soai et al., 1995).
Did Life Need Homochirality from the Start?
It’s debated whether a fully homochiral set of building blocks was necessary for the origin of life or a result of it. Some argue that molecular replication and the origin of genetic polymers (like RNA) demand a high degree of chirality purity; a “genetic takeover” can only happen once one handedness dominates (Blackmond, 2019). Others, like Blackmond, point out that life might tolerate some chirality heterogeneity in its earliest stages. Perhaps early self-replicating molecules worked with a majority chirality and gradually refined to exclusive use (Sallembien et al., 2022). Either way, once one form gained supremacy, life would become locked in because any inclusion of opposite-handed monomers in polymers tends to break function (Harrison et al., 2023). Thus, life likely selected one hand and stuck with it, leading to the universal homochirality we see today.
In summary, Earth’s use of L-amino acids and D-sugars appears to be a largely historical outcome, not an absolute requirement of biochemistry. A “mirror life” with reversed chirality is perfectly plausible chemically, a fact that motivates researchers to explore it further (Rohden et al., 2021).
Synthetic Mirror Biomolecules: D-Proteins, L-DNA, and More
Advances in synthetic chemistry and biotechnology now allow scientists to construct mirror versions of biomolecules and even envision assembling a mirror cell. This field, sometimes dubbed xenobiology or mirror biology, has achieved several milestones (Harrison et al., 2023; Rohden et al., 2021). Here are the key experiments and studies aimed at creating or investigating mirror life, from individual molecules to partial biological systems.
Researchers have chemically synthesized numerous mirror-image biomolecules by using enantiomerically flipped building blocks (D-amino acids in place of L, and L-nucleotides in place of D). These efforts demonstrate that, in isolation, many biological macromolecules have functional mirror twins.
In 2016, Ting Zhu and colleagues created a mirror-image DNA polymerase, a DNA-copying enzyme composed entirely of D-amino acids. They chemically synthesized a D-amino acid version of an enzyme and showed it could perform DNA replication and transcription on mirror-DNA templates (Wang et al., 2016). This was a landmark proof-of-concept of a “mirror central dogma” element. However, the initial mirror enzyme was small and low-efficiency. The group later synthesized a much larger, thermostable D-polymerase (Dpo4) ~352 amino acids in size (Xu et al., 2017; Pech et al., 2017; Peplow, 2016). By 2021, they reported a 90 kDa mirror version of Pfu DNA polymerase (from Pyrococcus furiosus), engineered in two fragments that self-assemble into a functional enzyme (Fan et al., 2021). This mirror Pfu polymerase can accurately amplify L-DNA (mirror DNA) in PCR-like reactions. Achieving such a large D-protein by total chemical synthesis was a major milestone, one of the largest D-proteins ever made, enabling “kilobase-sized mirror-gene” amplification.
With mirror polymerases in hand, researchers have begun constructing and using L-DNA (mirror DNA). L-DNA has the same double-helical structure as natural DNA but with left-handed deoxyribose sugars; it is sometimes called “enantiomeric DNA” or a Spiegel (mirror) polymer. A striking finding is that L-DNA is completely stable against natural nucleases and biochemistry; it’s invisible to normal enzymes. Fan et al. (2021) demonstrated this by encoding a piece of text (Louis Pasteur’s 1860 paragraph on molecular asymmetry) in an L-DNA sequence and showing it could be stored in the environment without degradation. An L-DNA “barcode” mixed into pond water remained intact and amplifiable a full year later, whereas a normal D-DNA control was destroyed within a day by microbes and nucleases. This illustrates how mirror genetic material can serve as robust “orthogonal” information storage, unreadable and uneditable by existing life. Researchers also developed mirror RNA transcription; e.g., a mirror version of T7 RNA polymerase (a ~100 kDa enzyme) was synthesized in 2022, capable of transcribing L-RNA from L-DNA templates (Chen et al., 2022; Fan et al., 2021; Rohden et al., 2021). This completes more steps of a mirror central dogma.
Beyond polymerases, other enzymes have been mirrored. In 2019, Weidmann et al. chemically synthesized a mirror-image DNA ligase entirely from D-amino acids. Remarkably, this D-ligase was enzymatically active; it could ligate L-DNA ends just as the normal ligase joins D-DNA (Weidmann et al., 2019). A commentary hailed this as “an important step for the creation of artificial life,” noting that preparing fully D-proteins is an “elegant approach to assemble completely orthogonal living systems”. Other examples include mirror versions of protein kinases, proteases, and metabolic enzymes constructed in research settings, often to test their folding and function. Each mirror enzyme typically functions only on substrates of the same chirality, highlighting strict stereospecificity (Meledin & Brik, 2019). For example, a natural L-enzyme that acts on an L-peptide will have a D-mirror counterpart that acts with equal proficiency on the corresponding D-peptide substrate (Milton et al., 1992; Weinstock et al., 2014).
An exciting application has been mirror-image aptamers (“Spiegelmers”). These are short DNA or RNA strands made of L-nucleotides that can bind targets like proteins. Because they are made of the unnatural mirror form, they resist degradation by nucleases. Companies like Noxxon Pharma have developed L-RNA aptamers that bind disease-related proteins, aiming for therapies that last longer in the body. One such L-aptamer drug (Spiegelmer NOX-A12) has progressed to clinical trials for cancer and other diseases (Vater & Klussmann, 2015). Until recently, producing mirror aptamers was laborious. In 2022, Chen et al. used a mirror-image PCR and selection system to evolve L-DNA aptamers directly from large random libraries. They performed iterative rounds of mirror PCR (using a mirror polymerase) to enrich L-DNA sequences that bind human thrombin. The selected L-DNA aptamers functioned as high-stability thrombin inhibitors, surviving even in human serum where normal DNA aptamers were quickly destroyed (Peplow, 2016). This demonstrated directed evolution in mirror space, greatly expanding the toolkit of mirror-life molecular technology (Chen et al., 2022).
A number of individual D-proteins have been synthesized to study their properties. Notably, a protein made of D-amino acids will fold into the exact mirror tertiary structure of the L-protein. X-ray crystallography of racemic mixtures (mixed D- and L-protein) can even yield better crystal packing, a technique called racemic crystallography used to solve structures that might be hard to crystallize otherwise (Yeates & Kent, 2012). Fewer than 30 distinct D-proteins had been reported by 2021. Examples include D-forms of enzymes like D-HIV protease (Milton et al., 1992), D-interleukin, and various D-peptide hormones (Weinstock et al., 2014). In all cases, the D-protein is a functional mirror of the L-protein, except that it is inert to normal biology (e.g., a D-enzyme won’t interact with L-substrates unless the substrate is achiral or via non-specific contacts) (Rohden et al., 2021).
These achievements confirm a critical point. Biochemistry is symmetric with respect to chirality. Under the same physical conditions, a protein made of D-amino acids is just as viable in structure and catalysis as its L-counterpart, only oriented in reverse (Harrison et al., 2023). There appear to be no insurmountable internal barriers to mirror life’s chemistry; the challenge is more about engineering and assembling a complete mirror system (Rohden et al., 2021). Chiral orthogonality in partial ribosome assembly, displayed in Figure 3.
Assembling Mirror-Life Systems
Constructing a fully autonomous “mirror organism” is a daunting task, but theoretical blueprints are emerging. It essentially requires rebuilding every major cellular component in opposite chirality: the genome (as L-DNA), the transcription/translation apparatus (L-RNA, D-ribosome, D-enzymes), and metabolic pathways (with mirror enzymes and co-factors) (Rohden et al., 2021; Harrison et al., 2023).
The ribosome, the molecular machine that translates mRNA into protein, is a top target for mirroring, but also one of the most complex (consisting of >50 proteins and 3 large RNA molecules). As an intermediate step, Zhu’s group synthesized mirror versions of several ribosomal proteins (D-amino acid versions of E. coli ribosomal proteins L5, L18, L25, each ~100–200 amino acids). They showed in 2019 that these mirror ribosomal proteins fold correctly and can assemble in vitro with a mirror (L-form) 5S rRNA to form partial mirror ribonucleoprotein complexes (Rohden et al., 2021). Crucially, they observed chiral specificity; the D-proteins did not bind normal 5S rRNA, only the L-5S, and vice versa. This proved that mixing chiral components in the ribosome doesn’t work; you cannot plug a mirror protein into a normal ribosome or vice versa, but a fully mirrored complex can assemble and is structurally analogous to the natural one. While a complete mirror ribosome (with all subunits and RNAs) has not been built yet, this was a step toward a mirror translation system. Simplified translation schemes are also being attempted; for example, using ribozymes called flexizymes to charge mirror tRNAs with D-amino acids, bypassing the need for dozens of mirror tRNA-synthetase enzymes (Peplow, 2016). In 2021, Chen et al. demonstrated charging L-form tRNAs with D-amino acids and even produced a small protein in a partially mirrored translation system (though not yet a fully functional mirror ribosome). These advances suggest it’s feasible to establish mirror protein synthesis, given enough engineered components (Chen et al., 2022; Peplow, 2016).
The simplest “organisms” to mirror might be viruses, since they have small genomes and few proteins (relying on a host to replicate). In theory, one could synthesize a mirror virus, e.g., an RNA virus with L-RNA genome and D-protein capsid. However, such a virus would not infect normal cells (the mirror RNA wouldn’t be recognized by L-ribosomes, and the D-capsid wouldn’t attach to L-receptors). It would require a mirror host cell to replicate. Thus, mirror viruses remain a thought experiment until mirror cells exist. One study in the 2000s did synthesize a polio virus from scratch (the natural form) to show de novo virus creation is possible; a mirror version would be the next leap. Some have proposed creating a mirror bacteriophage that could infect a mirror bacterium as part of a mirror ecosystem, but this remains speculative (Rohden et al., 2021).
Visionaries like genomics pioneer J. Craig Venter and synthetic biologist George Church have outlined how one might build a mirror microbe. One approach starts with a minimal cell (like Mycoplasma with ~500 genes, which Venter’s team has synthesized in normal form) and systematically replaces each component with its mirror version (Rohden et al., 2021). This would involve chemically synthesizing a mirror genome (L-DNA) and mirror ribosomes and enzymes, then encapsulating them in either a mirror membrane or an achiral membrane. Notably, membranes pose a special challenge; phospholipids are chiral molecules too. Church suggested using achiral lipid molecules for the first mirror cells to avoid needing mirror lipids. (Alternatively, one could use the archaeal lipid chirality as a “mirror” to bacterial, since life already tolerates that difference.) Another approach is building a proto-cell with a simplified metabolism that can copy, mirror DNA, and grow. Dr. Kate Adamala, for example, worked on assembling mirror versions of a protocell system (a liposome with a minimal gene set) as a testbed before ethical concerns shifted her focus (Adamala et al., 2024).
In parallel, computational biology has examined whether mirror proteins would interact with normal biomolecules. Predictably, they do not; a mirror enzyme’s binding pocket is the mirror image of the normal one, so it won’t bind the natural substrate. Computational docking studies confirm, for instance, that antibiotic drugs will not fit mirror bacterial targets (e.g., a mirror ribosome’s peptidyltransferase center has opposite chirality, so current antibiotics cannot bind it). This provides in silico evidence that a mirror microbe would be inherently resistant to our antibiotics and likely unrecognized by our immune proteins (Harrison et al., 2023; Adamala et al., 2024).
In summary, scientific progress has yielded individual mirror components of life: DNA, RNA, enzymes, even partial ribosomes, and shown that they can function properly in isolation. However, constructing a fully self-sustaining mirror organism has not yet been achieved. It remains a grand challenge requiring integration of all these pieces. Estimates vary, but some experts suspect it may take a decade or more of focused research to build even a simple mirror bacterium (Rohden et al., 2021). The efforts so far, though, have taught us a great deal about the feasibility and behavior of mirror-biochemistry, as discussed next.
Symmetry, Compatibility, and Constraints
Experiments confirm that for every biochemical mechanism in normal life, an equivalent mirror mechanism is possible. DNA replication, transcription, ligation, and even translation can, in principle, all operate with reversed chirality. Mirror polymerases accurately copy mirror DNA (Wang et al., 2016; Fan et al., 2021), mirror ligases join DNA ends (Weidmann et al., 2019), and mirror ribosomal proteins assemble with mirror RNA (Rohden et al., 2021; Meledin & Brik, 2019). There is no known biochemical reaction that inherently “fails” when done with opposite chirality, except that it must be done on consistently opposite substrates. This underscores a profound symmetry; the laws of chemistry (electromagnetism) are indifferent to chirality (Quack, 2002; Peplow, 2016). A mirror-image cell would obey the same biophysics as a normal cell, achieving the same complexity and function, just reflected. The only slight asymmetry comes from weak nuclear forces, but as noted, those effects are negligibly small for biochemistry (Quack, 2002).
While mirror life as a whole is viable, mixing chirality within one system is usually deleterious. The research uniformly shows that chiral specificity is strict; mirror enzymes do not interact productively with normal molecules and vice versa. For example, the mirror ribosomal proteins bound only mirror rRNA, not natural rRNA (Rohden et al., 2021). A mirror polymerase will not copy a normal DNA template (the geometry doesn’t match), and a normal polymerase won’t accept L-nucleotide substrates (Wang et al., 2016; Xu et al., 2017). If one tried to feed L-glucose to a normal bacterium, it would starve despite an energy-rich sugar being present, because its enzymes are specific for D-glucose (Vater & Klussmann, 2015; Fan et al., 2021; Peplow, 2016). These findings reinforce that any life form must internally choose one chirality consistently, otherwise its informational polymers and catalytic networks break down due to stereo-conflicts. This is likely why Earth life, once it “chose” L-amino acids and D-sugars, had to remain homochiral to thrive (Harrison et al., 2023).
One striking lesson is that a mirror biosphere would be biochemically isolated from our own. The experiments illustrate a strong form of orthogonality; normal organisms can’t eat or metabolize mirror-organic material efficiently, and mirror organisms couldn’t directly utilize normal nutrients (Harrison et al., 2023; Adamala et al., 2024). For instance, mirror cyanobacteria (if they existed), producing L-glucose and other mirror sugars, would create piles of energy-rich compounds that normal organisms cannot use. Conversely, a mirror microbe in our soil might sit surrounded by D-glucose and L-amino acids and yet starve, unless it has some racemase enzyme or a source of mirror nutrients. One exception noted is that some Earth bacteria have enzymes (racemases, epimerases) that can convert certain D-amino acids or L-sugars to the usable form. For example, a few strains can slowly metabolize L-glucose after converting it to D-glucose, and bacteria incorporate a few D-amino acids into cell walls via specialized enzymes (Rohden et al., 2021). These are rare capabilities, but they suggest that given enough time, one form of life could evolve to partially exploit the other form’s resources. Generally, however, mirror life and standard life would not compete for most biochemical resources; they occupy “orthogonal” niches.
A practical outcome of mirror experiments is recognizing that mirror biomolecules are often more stable in a normal environment. We saw how L-DNA resists nuclease degradation for orders of magnitude longer than regular DNA (Fan et al., 2021). Similarly, D-peptide drugs are not cleaved by our proteases (which target L-peptide bonds) (Milton et al., 1992; Weinstock et al., 2014). This stability is a double-edged sword; it makes mirror molecules attractive for drug design (longer half-life in the body) (Vater & Klussmann, 2015), and data storage (as DNA vaults that microbes can’t eat) (Fan et al., 2021), but it also means if mirror organisms or biochemicals were released, our ecosystem’s normal recycling pathways (decomposition, predation) wouldn’t eliminate them effectively. A mirror leaf falling from a tree might not be broken down by standard fungi or bacteria, potentially leading to the accumulation of litter that normal decomposers can’t digest (Adamala et al., 2024).
The step-by-step successes (polymerase, ligase, ribosomal subunits, etc.) show that mirror life is feasible; no known scientific law forbids it. The challenge is overwhelmingly one of engineering and labor. Chemical synthesis of large D-proteins is extremely time-consuming, often requiring solid-phase peptide synthesis in many segments and clever “ligation” chemistry to stitch segments together (Xu et al., 2017; Pech et al., 2017). For reference, the 90 kDa mirror Pfu polymerase had to be synthesized in two halves (each ~45 kDa), purified, and refolded together. The mirror T7 RNA polymerase (~100 kDa) was done in three fragments (Fan et al., 2021; Chen et al., 2022). To make a mirror E. coli (~1000 different proteins) by today’s methods is far beyond reach. However, each technological improvement (e.g., better chemical ligation methods, automated D-peptide synthesizers, or partial biological production of mirror blocks via engineered ribozymes) brings the goal closer (Harrison et al., 2023). Researchers ultimately imagine a bootstrapping scenario; once a minimal mirror ribosome or polymerase system is established, it could begin self-replicating mirror components, greatly accelerating progress. In essence, the first mirror cell will be hardest to build, but after that, it could propagate and perhaps even be engineered to produce other mirror biomaterials for us (Rohden et al., 2021).
In summary, the research so far teaches that mirror life is a realistic possibility in principle, with chemistry that mirrors our own. Yet it would be biochemically self-contained, unable to exchange genetic information or metabolites with standard life (Adamala et al., 2024). It validates that homochirality is a contingent feature, not a fixed requirement, since all critical biomolecules have functional mirror equivalents (Blackmond, 2019). And it highlights practical constraints; until we have mirror organisms to carry some load, creating large mirror systems will require enormous synthetic effort.
Discussion
Research into mirror life feeds directly into theories about how life began and why it favors one chirality. If we can create a mirror version of a living system, it would strengthen the idea that life’s emergence did not depend on any special “handed” property of physics or Earth, that life could have started with opposite chirality just as well (Blackmond, 2019; Rohden et al., 2021). This underscores that Earth’s homochirality was likely a result of symmetry-breaking processes, chance or slight biases, rather than an inevitable outcome (Sallembien et al., 2022). Moreover, attempts to build mirror life force us to revisit origin-of-life pathways; could prebiotic chemistry have produced a self-replicator in a racemic environment? The difficulty of cross-chirality interactions suggests that some early symmetry-breaking was probably necessary so that polymers could reliably replicate (a strand of RNA of mixed chirality might not fold or function). By experimenting with mirror nucleic acids and enzymes, scientists can test origin-of-life hypotheses in “chirality-neutral” ways. For example, one could attempt to start an RNA replication cycle with L-RNA and see if it is any harder or easier than with D-RNA; if there’s no difference, it implies life’s choice was truly arbitrary. So far, mirror biochemical reactions seem just as efficient, implying no intrinsic advantage to Earth’s L/D configuration (Wang et al., 2016; Xu et al., 2017; Weidmann et al., 2019; Rohden et al., 2021).
Another origin implication is the idea of a “second genesis” on Earth. If life could start in one-handedness, could it have started independently in the opposite handedness, and we just haven’t found it? Some scientists speculated about a possible “shadow biosphere” (Rohden et al., 2021), undiscovered microbial life using different chemistry co-existing with standard life. The fact that we haven’t clearly found any mirror-life pockets (e.g., no known organism uses L-sugars in DNA) suggests that if such a lineage ever began, it either died out or was outcompeted early. It is difficult for mirror life to hide in plain sight, since any mirror organism would leave telltale signs (e.g., producing only D-amino acids in its environment). Our analyses of environmental organic chirality haven’t revealed such anomalies beyond what meteorites contribute (Cronin & Pizzarello, 1997; Bailey et al., 1998). This makes it likely that Earth had a single origin of homochirality that set the pattern, rather than parallel origins with different handedness. Still, building a mirror cell in the lab, a “second genesis” under human guidance, might shed light on what minimal conditions are required for life’s spark independent of existing biology (Adamala et al., 2024).
Finally, mirror life experiments provide a compelling demonstration of life’s modularity. By swapping chirality, we learn which aspects of biochemistry are “hardwired” and which are flexible. The success of mirror polymerases and ribozymes indicates that the core logic of the central dogma doesn’t depend on specific molecular handedness. This reinforces the view that life’s origin is rooted in information and self-replication processes that are chemically agnostic to handedness (Wang et al., 2016; Fan et al., 2021; Chen et al., 2022). In summary, mirror life research suggests that if you rerun the tape of life’s origin, you could get the opposite chirality, and everything would function equivalently, a profound insight into the contingency of life’s chemistry.
One of the most exciting implications of mirror life is for extraterrestrial life. If life exists elsewhere in the universe, will it share our molecular handedness, or could it be a “mirror” of ours? Nothing about Earth’s environment uniquely determines that L-amino acids had to be chosen, so it’s quite plausible that alien biochemistries might use D-amino acids and L-sugars (or even a mix, though as discussed, pure homochirality is likely for any complex life) (Blackmond, 2019; Sallembien et al., 2022). This means our search for life on other planets should be careful not to assume Earth-like chirality. For instance, the Viking landers on Mars in the 1970s applied experiments that added Earth-chiral nutrients to soil to see if anything metabolized them. If Martian microbes (hypothetically) were of opposite chirality, those experiments might have given false negatives, since mirror bugs wouldn’t eat our L-amino acids or D-sugars. Thus, life-detection instruments in future missions are being designed to test for chirality (Rohden et al., 2021). One idea is to include a chiral separation or optical activity measurement; detection of a strong excess of either L- or D-enantiomers in Martian soil organics would be a hint of life, regardless of which hand is in excess (Blackmond, 2019). In fact, the signature of biology might simply be homochirality itself, no matter which hand, because non-biological chemistry produces racemic mixtures. As Blackmond put it, “the homochirality of biological molecules is a signature of life”. So finding either an L-bias or a D-bias in organic molecules beyond Earth would be a tantalizing sign of alien life (mirror or not) (Cronin & Pizzarello, 1997; Bailey et al., 1998).
If we ever encounter an actual alien organism, imagine a microorganism from Europa or an alien microbe fossil in a meteorite, determining its chirality will be one of the first tasks. A mirror life form would pose unique challenges; biological incompatibility would mean we likely couldn’t digest alien plants or animals for food if their chirality is opposite (a trope exploited in science fiction). The Expanse novel Cibola Burn illustrates colonists unable to eat native flora because of a chirality mismatch. Likewise, our pathogens probably couldn’t infect aliens of opposite chirality, and vice versa, which might be a relief regarding panspermia concerns (Harrison et al., 2023; Adamala et al., 2024). However, if alien life shares the same chirality as us, that raises deeper questions of common origin or deterministic processes (some have speculated that if life spread via meteorites, it could impose the same chirality across planets). Most scientists expect that life on another world could well be mirror-handed relative to us, given the arbitrary nature of the choice and the evidence from meteorites that some regions of space produce one excess (e.g., left-handed amino acids) while others might produce the opposite under different polarized radiation environments (Cronin & Pizzarello, 1997; Bailey et al., 1998).
In astrobiology, researchers also consider whether both forms could arise on one planet. If a planet had two separate genesis events, one L-based and one D-based, would one inevitably extinguish the other? Our Earth experience and the incompatibility of coexisting chiral biochemistry suggest they would largely ignore each other except for competition over achiral resources like minerals, sunlight, etc. Possibly, one might gain an edge and dominate, for example, if one evolves photosynthesis first, it could fill the atmosphere with oxygen toxic to the other (Rohden et al., 2021). So, it could be rare to have a long-term stable dual biosphere; one would likely outcompete or outlive the other, resulting in a single-handed planet. That said, on a planet with separated niches (e.g., different oceans or an underground vs. surface biosphere), perhaps two chirality domains could persist without contact. These are speculative scenarios that mirror-life science allows us to ponder more concretely.
In summary, mirror life broadens our perspective in the search for life in the cosmos. It urges us not to be “handedness chauvinists”; alien life may be our molecular mirror. To detect it, we must look for the hallmark of chirality excess in organics and perhaps design experiments that cater to both L and D worlds. In a sense, mirror life research is training us to recognize life “as we don’t know it” by rehearsing with a version that is chemically alien but conceptually the same as us.
Beyond theoretical considerations, mirror biology has very practical implications here on Earth. Studying mirror molecules sharpens our fundamental understanding of biomolecular interactions. For example, by creating a D-enzyme and comparing its kinetics on a D-substrate vs. the L-enzyme on L-substrate, we test how much of the enzymatic specificity is pure geometry. So far, results show mirror enzymes are as efficient as normal ones for their mirror substrates, reinforcing that enzyme catalysis is primarily stereocomplementary shape-matching (Milton et al., 1992; Weinstock et al., 2014; Harrison et al., 2023). Racemic crystallography (growing crystals of mixed D/L protein) has helped solve structures that were previously elusive, improving drug target knowledge (Yeates & Kent, 2012). Mirror proteins also allow unique biophysical experiments, such as forming mirror-image complexes; one can mix an L-protein and its D-protein in equal parts. They cannot dimerize in the normal way, since each is the other's mirror shape, but they can co-crystallize in symmetric lattices, revealing interesting properties of protein folding and association. All this contributes to basic biochemistry and structural biology
The pharmaceutical world is intensely concerned with chirality; many drugs have an active enantiomer and an inert or even harmful mirror enantiomer (e.g., thalidomide’s tragic enantiomeric effects). Mirror life research provides tools to obtain pure enantiomers of complex molecules. A mirror microbe or enzyme could potentially produce a drug molecule in the opposite chirality to what a normal enzyme would make. Enantiopure drug synthesis could thus be aided by mirror biocatalysts. Moreover, mirror peptides and aptamers are promising therapeutics because they evade breakdown. Several D-peptide drugs (mirror-image peptides) are in development as protease-resistant inhibitors of diseases, for instance, D-peptides that block viral entry by binding viral proteins (HIV, COVID-19) have been researched, leveraging the fact that human proteases won’t degrade D-peptides. Mirror aptamers (Spiegelmers), as mentioned, are in trials for diseases like cancer and inflammation. These tend to have lower immunogenicity and longer circulation (Vater & Klussmann, 2015; Chen et al., 2022). One fascinating case is a mirror-image antibody or monobody; scientists have created D-protein scaffolds that bind targets just like an antibody would, but being D, they aren’t recognized by the immune system, making them stealthy. A recent study even generated a mirror-image binding protein (monobody) against a cytokine (MCP-1) using a combination of mirror display techniques and chemical synthesis. Such binders could serve as long-lasting drugs. Additionally, mirror life raises possibilities of novel biomaterials; for example, D-collagen or D-spider silk might have different properties or be impervious to typical enzymes, useful for medical implants that resist degradation (Harrison et al., 2023).
Mirror organisms, if ever created, would be a new kind of tool for synthetic biology. They could serve as biosecure factories; a mirror bacterium could produce valuable compounds without risk of being infected by wild-type viruses or contaminated by regular microbes, since those can’t interact biochemically (Rohden et al., 2021). As noted in one concept, a mirror microbe in an industrial fermenter would be immune to all normal phages that often plague fermentation processes. Moreover, if it escaped, one might expect it to die off due to a lack of food. This notion of orthogonal containment, organisms that can’t exchange genes or metabolites with natural ecosystems, has been discussed as a safety measure in biotechnology. However, recent risk analyses complicate this rosy picture; If a mirror organism is photosynthetic or can tap achiral resources (like mirror cyanobacteria using CO₂ and sunlight), it would survive and potentially proliferate unchecked, because no natural predator eats it, and our antibiotics wouldn’t kill it. This worst-case scenario has led scientists to call for caution (Adamala et al., 2024). In late 2024, a consortium of biologists (including Nobel laureates) published a report and a science editorial warning that a mirror microbe could pose “unprecedented and irreversible” risks if released. They pointed out that mirror bacteria might evade immune defenses and cause lethal infections in plants and animals, essentially acting as novel pathogens that our bodies can’t even recognize. And if they were established in the environment, we would have no effective antibiotics or predators to control them. Given these concerns, the group urged a moratorium on creating any fully functional mirror life until rigorous safeguards and global consensus are in place. This is a striking example of scientists anticipating risks before the technology fully materializes, a commendable instance of proactive ethics in synthetic biology (Adamala et al., 2024).
In response to these issues, some suggest that mirror life work should continue only at the molecular level, but not culminate in a self-replicating mirror organism until safety strategies (like built-in lethal genes or absolute containment) are foolproof. Others emphasize that mirror chemistry still holds immense promise in a controlled scope, for instance, using mirror molecular machinery in closed systems to carry out novel chemistry or store information (the concept of “chiral encryption” where data encoded in L-DNA can’t be read without a mirror polymerase).
Finally, mirror life research has philosophical and educational implications; it challenges our definition of life’s universals. It demonstrates that we can consider, and potentially create, life that is fundamentally biochemically isolated from us. This compels us to refine our criteria for life. It also inspires public imagination, appearing in science fiction as mirror-image humans or ecosystems, and prompting discussions about how we would relate to an alien or synthetic life with such a peculiar difference (Blackmond, 2019; Rohden et al., 2021).
Conclusion
The exploration of “mirror life” has progressed from Pasteur’s 19th-century speculation to an active interdisciplinary pursuit at the frontiers of chemistry and biology (Sallembien et al., 2022). Chirality, the handedness of molecules, proves to be a crucial but ultimately flexible facet of life. Earth’s organisms uniformly use left-handed amino acids and right-handed sugars, a fact that likely arose from stochastic symmetry-breaking and became frozen in due to life’s need for homochirality (Blackmond, 2019; Cronin & Pizzarello, 1997; Bailey et al., 1998). There appears to be nothing inevitable about that choice; research shows that a fully functional biochemistry can be built from the opposite set of building blocks, and that such a mirror biochemistry would operate by the same principles of molecular recognition and catalysis (Wang et al., 2016; Xu et al., 2017; Weidmann et al., 2019; Rohden et al., 2021).
Synthetic biology has taken impressive strides toward constructing a mirror life’s toolkit. Mirror-image polymerases, ligases, aptamers, and even partial ribosomes have been created, demonstrating the viability of mirror genetic processes (Fan et al., 2021; Chen et al., 2022; Rohden et al., 2021). These experiments underscore that the phenomena of life, information storage, self-replication, and catalysis are fundamentally invariant under reflection (Harrison et al., 2023). In practical terms, mirror biomolecules are already yielding real-world benefits in medicine; long-lived therapeutics, novel drug designs, and technology; biostable data storage in L-DNA (Vater & Klussmann, 2015; Adamala et al., 2024). At the same time, they highlight how segregated a mirror biosphere would be from our own, essentially invisible to each other’s biology (Harrison et al., 2023). This raises both exciting possibilities (e.g., orthogonal biocontained systems) and serious biosafety concerns; a mirror pathogen could evade all our defenses (Adamala et al., 2024).
For origin-of-life research, the mirror life concept reinforces that life’s emergence was not predestined to one chirality; we likely live in one of two possible symmetry states, chosen by chance or slight bias and then amplified (Soai et al., 1995; Blackmond, 2019). For astrobiology, it broadens our expectation of what alien life might be like; we must be prepared to find “life, even down to the handedness of its molecules (Rohden et al., 2021; Blackmond, 2019). And for synthetic biology, it represents both the ultimate challenge and a cautionary frontier; the ability to engineer a parallel form of life orthogonal to all existing life (Adamala et al., 2024). Achieving a mirror organism, if done responsibly, would be a profound technical and scientific milestone, essentially proving that life is a phenomenon that transcends specific chemistry, as long as information and self-replication can be instantiated in that chemistry (Harrison et al., 2023).
Acknowledgments
I am extremely grateful to Prof. Nuran Çiçek for her mentorship, guidance, and steadfast support, as well as for the confidence and trust she placed in me throughout this process.
Competing Interests
Authors declare no competing interests.
Funding
No external funding was received.
Data and Code Availability
No new datasets or code were generated.
Author Contributions
Conceptualization: A. Şems
References
- Adamala, K. P. , Agashe, D., Belkaid, Y., Bittencourt, D. M. C., Cai, Y., Chang, M. W., Chen, I. A., Church, G. M., Cooper, V. S., Davis, M. M., Devaraj, N. K., Endy, D., Esvelt, K. M., Glass, J. I., Hand, T. W., Inglesby, T. V., Isaacs, F. J., James, W. G., Jones, J. D. G., Kay, M. S., … Zuber, M. T. Confronting risks of mirror life. Science 2024, 386, 1351–1353. [Google Scholar] [CrossRef]
- Bailey, J. , Chrysostomou, A., Hough, J. H., Gledhill, T. M., McCall, A., Clark, S., Menard, F., & Tamura, M. Circular polarization in star-formation regions: implications for biomolecular homochirality. Science 1998, 281, 672–674. [Google Scholar]
- Blackmond D., G. The Origin of Biological Homochirality. Cold Spring Harbor perspectives in biology 2019, 11, a032540. [Google Scholar] [CrossRef]
- Chen, J. , Chen, M., & Zhu, T. F. Directed evolution and selection of biostable L-DNA aptamers with a mirror-image DNA polymerase. Nature biotechnology 2022, 40, 1601–1609. [Google Scholar] [CrossRef]
- Cronin, J. R. , & Pizzarello, S. Enantiomeric excesses in meteoritic amino acids. Science 1997, 275, 951–955. [Google Scholar] [CrossRef]
- Fan, C. , Deng, Q., & Zhu, T. F. Bioorthogonal information storage in L-DNA with a high-fidelity mirror-image Pfu DNA polymerase. Nature biotechnology 2021, 39, 1548–1555. [Google Scholar] [CrossRef]
- Harrison, K. , Mackay, A. S., Kambanis, L., Maxwell, J. W. C., & Payne, R. J. Synthesis and applications of mirror-image proteins. Nature reviews. Chemistry 2023, 7, 383–404. [Google Scholar] [CrossRef]
- Jiang, W. , Zhang, B., Fan, C., Wang, M., Wang, J., Deng, Q., Liu, X., Chen, J., Zheng, J., Liu, L., & Zhu, T. F. Mirror-image polymerase chain reaction. Cell discovery 2017, 3, 17037. [Google Scholar] [CrossRef]
- Meledin, R. , & Brik, A. Mirroring Life's Building Blocks. Cell chemical biology 2019, 26, 616–619. [Google Scholar] [CrossRef]
- Milton, R. C. , Milton, S. C., & Kent, S. B. Total chemical synthesis of a D-enzyme: the enantiomers of HIV-1 protease show reciprocal chiral substrate specificity [corrected]. Science 1992, 256, 1445–1448. [Google Scholar] [CrossRef] [PubMed]
- Pech, A. , Achenbach, J., Jahnz, M., Schülzchen, S., Jarosch, F., Bordusa, F., & Klussmann, S. A thermostable d-polymerase for mirror-image PCR. Nucleic acids research 2017, 45, 3997–4005. [Google Scholar] [CrossRef]
- Peplow, M. Mirror-image enzyme copies looking-glass DNA. Nature 2016, 533, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Quack, M. How important is parity violation for molecular and biomolecular chirality? Angewandte Chemie (International ed. in English) 2002, 41, 4618–4630. [Google Scholar] [CrossRef]
- Rohden, F. , Hoheisel, J. D., & Wieden, H. J. Through the looking glass: milestones on the road towards mirroring life. Trends in biochemical sciences 2021, 46, 931–943. [Google Scholar] [CrossRef]
- Sallembien, Q. , Bouteiller, L., Crassous, J., & Raynal, M. Possible chemical and physical scenarios towards biological homochirality. Chemical Society Reviews 2022, 51, 3436–3476. [Google Scholar] [CrossRef]
- Soai, K. , Kawasaki, T., & Matsumoto, A. Asymmetric autocatalysis of pyrimidyl alkanol and its application to the study on the origin of homochirality. Accounts of chemical research 2014, 47, 3643–3654. [Google Scholar] [CrossRef]
- Vater, A. , & Klussmann, S. Turning mirror-image oligonucleotides into drugs: the evolution of Spiegelmer(®) therapeutics. Drug discovery today 2015, 20, 147–155. [Google Scholar] [CrossRef]
- Wang, Z. , Xu, W., Liu, L., & Zhu, T. F. A synthetic molecular system capable of mirror-image genetic replication and transcription. Nature Chemistry 2016, 8, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, M. T. , Jacobsen, M. T., & Kay, M. S. Synthesis and folding of a mirror-image enzyme reveals ambidextrous chaperone activity. Proceedings of the National Academy of Sciences of the United States of America 2014, 111, 11679–11684. [Google Scholar] [CrossRef] [PubMed]
- Xu, W. , Jiang, W., Wang, J., Yu, L., Chen, J., Liu, X., Liu, L., & Zhu, T. F. Total chemical synthesis of a thermostable enzyme capable of polymerase chain reaction. Cell discovery 2017, 3, 17008. [Google Scholar] [CrossRef]
- Yeates, T. O. , & Kent, S. B. Racemic protein crystallography. Annual review of biophysics 2012, 41, 41–61. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chirality of the amino acid alanine. Top: structural formulas of L-alanine (S-alanine, left) and its mirror image D-alanine (R-alanine, right), showing the non-superimposable arrangement of substituents around the central α-carbon atom. Bottom; ball-and-stick models of the two enantiomers, highlighting their mirror symmetry. Although L- and D-alanine share identical chemical composition, they cannot be superimposed, and their distinct handedness underlies stereospecific interactions in biology. (Blackmond, 2019; Cronin & Pizzarello, 1997). Created in BioRender. Sems, A. (2025) https://BioRender.com/kwzrgw9.
Figure 1.
Chirality of the amino acid alanine. Top: structural formulas of L-alanine (S-alanine, left) and its mirror image D-alanine (R-alanine, right), showing the non-superimposable arrangement of substituents around the central α-carbon atom. Bottom; ball-and-stick models of the two enantiomers, highlighting their mirror symmetry. Although L- and D-alanine share identical chemical composition, they cannot be superimposed, and their distinct handedness underlies stereospecific interactions in biology. (Blackmond, 2019; Cronin & Pizzarello, 1997). Created in BioRender. Sems, A. (2025) https://BioRender.com/kwzrgw9.
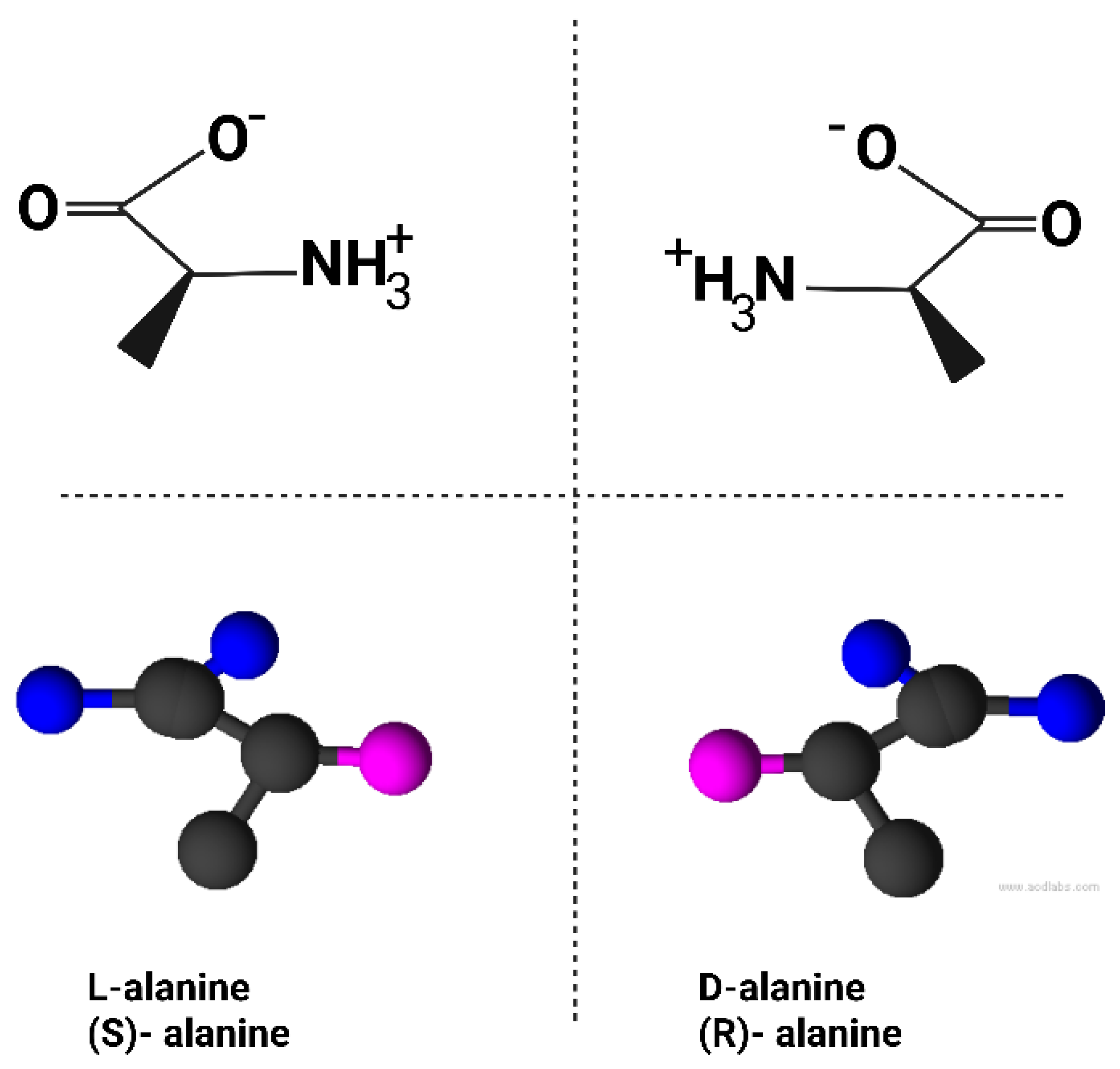
Figure 2.
Comparison of chirality in Earth life and hypothetical mirror life. Terrestrial proteins use L-amino acids, whereas mirror proteins would use D-amino acids (Milton et al., 1992; Weinstock et al., 2014). DNA and RNA on Earth are built on D-sugars, forming right-handed helices, while mirror nucleic acids would use L-sugars, forming left-handed helices (Wang et al., 2016; Fan et al., 2021). Membranes in bacteria/eukaryotes use sn-glycerol-3-phosphate backbones, while archaea employ sn-glycerol-1-phosphate; mirror organisms would invert these (Rohden et al., 2021). Metabolism on Earth depends on D-glucose, whereas mirror life would rely on L-glucose, which Earth enzymes cannot process (Vater & Klussmann, 2015). Created in BioRender. Sems, A. (2025) https://BioRender.com/j9q5v6w.
Figure 2.
Comparison of chirality in Earth life and hypothetical mirror life. Terrestrial proteins use L-amino acids, whereas mirror proteins would use D-amino acids (Milton et al., 1992; Weinstock et al., 2014). DNA and RNA on Earth are built on D-sugars, forming right-handed helices, while mirror nucleic acids would use L-sugars, forming left-handed helices (Wang et al., 2016; Fan et al., 2021). Membranes in bacteria/eukaryotes use sn-glycerol-3-phosphate backbones, while archaea employ sn-glycerol-1-phosphate; mirror organisms would invert these (Rohden et al., 2021). Metabolism on Earth depends on D-glucose, whereas mirror life would rely on L-glucose, which Earth enzymes cannot process (Vater & Klussmann, 2015). Created in BioRender. Sems, A. (2025) https://BioRender.com/j9q5v6w.
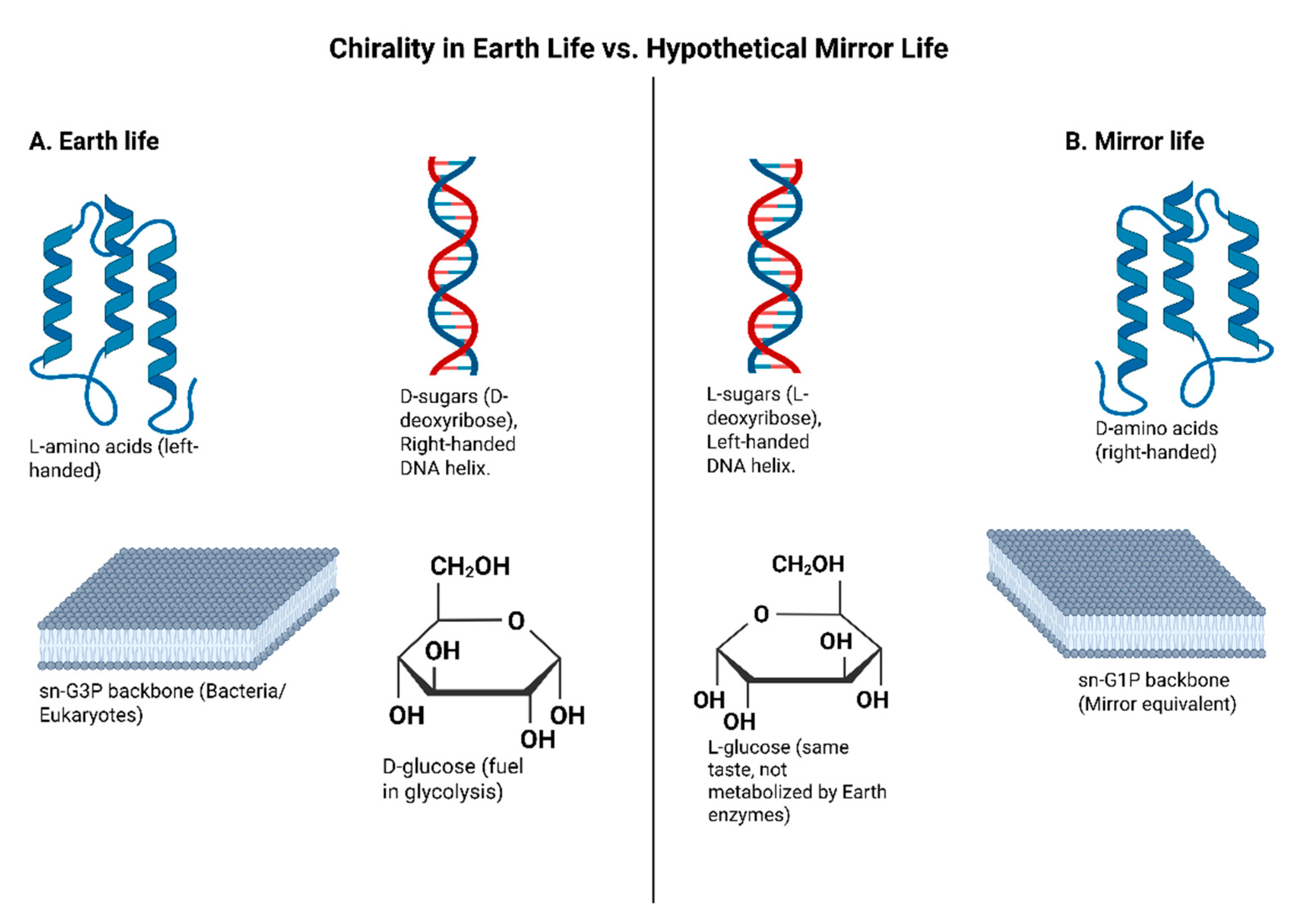
Figure 3.
Chiral orthogonality in partial ribosome assembly. Earth ribosomal proteins (L-amino acids) bind only Earth rRNA (D-sugar backbone) to form functional complexes (top left). Mirror ribosomal proteins (D-amino acids) bind only mirror rRNA (L-sugar backbone) to form mirror complexes (top right). Cross-combinations fail to assemble; mirror proteins cannot bind Earth rRNA, and Earth proteins cannot bind mirror rRNA (bottom panels). These results demonstrate that ribosomal components are strictly chiral-specific, and mixed systems do not function (Rohden et al., 2021; Chen et al., 2022). Created in BioRender. Sems, A. (2025) https://BioRender.com/viqofah.
Figure 3.
Chiral orthogonality in partial ribosome assembly. Earth ribosomal proteins (L-amino acids) bind only Earth rRNA (D-sugar backbone) to form functional complexes (top left). Mirror ribosomal proteins (D-amino acids) bind only mirror rRNA (L-sugar backbone) to form mirror complexes (top right). Cross-combinations fail to assemble; mirror proteins cannot bind Earth rRNA, and Earth proteins cannot bind mirror rRNA (bottom panels). These results demonstrate that ribosomal components are strictly chiral-specific, and mixed systems do not function (Rohden et al., 2021; Chen et al., 2022). Created in BioRender. Sems, A. (2025) https://BioRender.com/viqofah.
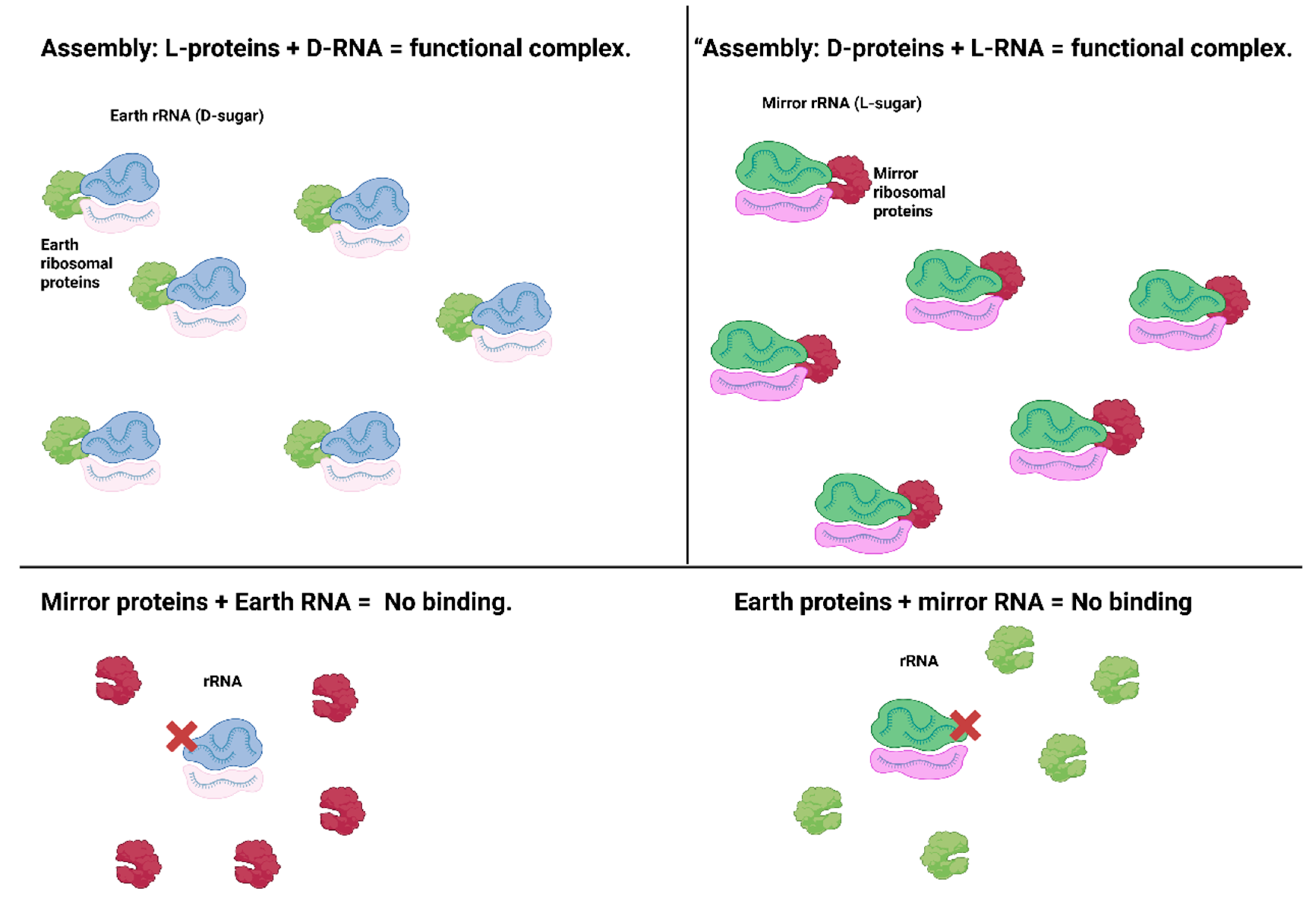

Table 1.
Comparison of molecular chirality in Earth life versus hypothetical mirror life. Proteins in terrestrial life are built exclusively from L-amino acids, whereas mirror life would employ D-amino acids (Milton et al., 1992; Weinstock et al., 2014). Nucleic acids on Earth use D-ribose or D-deoxyribose in RNA and DNA, forming right-handed helices; mirror life would instead use L-sugars, producing a left-handed double helix with identical base-pairing rules (Wang et al., 2016; Fan et al., 2021). Membrane chirality also differs; bacteria and eukaryotes use sn-glycerol-3-phosphate, while archaea employ the opposite sn-glycerol-1-phosphate; mirror life would invert these choices or potentially use achiral lipids (Rohden et al., 2021). In metabolism, Earth organisms process D-glucose as the primary sugar fuel; mirror organisms would rely on L-glucose, which tastes sweet to us but is not metabolized by our enzymes (Vater & Klussmann, 2015).
Table 1.
Comparison of molecular chirality in Earth life versus hypothetical mirror life. Proteins in terrestrial life are built exclusively from L-amino acids, whereas mirror life would employ D-amino acids (Milton et al., 1992; Weinstock et al., 2014). Nucleic acids on Earth use D-ribose or D-deoxyribose in RNA and DNA, forming right-handed helices; mirror life would instead use L-sugars, producing a left-handed double helix with identical base-pairing rules (Wang et al., 2016; Fan et al., 2021). Membrane chirality also differs; bacteria and eukaryotes use sn-glycerol-3-phosphate, while archaea employ the opposite sn-glycerol-1-phosphate; mirror life would invert these choices or potentially use achiral lipids (Rohden et al., 2021). In metabolism, Earth organisms process D-glucose as the primary sugar fuel; mirror organisms would rely on L-glucose, which tastes sweet to us but is not metabolized by our enzymes (Vater & Klussmann, 2015).
| Biochemical Component | Earth Chirality | Mirror Life Chirality (hypothetical) |
|---|---|---|
| Proteins (amino acids) | L-amino acids only (left-handed). | D-amino acids only (right-handed). |
| Nucleic acids (sugars) Example: DNA double helix |
D-sugars in RNA/DNA (right-handed). Built on D-deoxyribose; standard Watson-Crick base pairing. |
L-sugars in RNA/DNA (left-handed). It would be built on L-deoxyribose; it forms a mirror double helix. |
| Membrane phospholipids | G3P backbone in bacteria/eukaryotes (one enantiomer); G1P in archaea (mirror form). | Mirror life might use the opposite enantiomer of its Earth counterpart’s lipids. |
| Metabolic sugars (e.g., glucose) | D-glucose (dextrose) is used in glycolysis. | L-glucose same taste, but is not metabolized by Earth’s enzymes. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



