Submitted:
26 September 2025
Posted:
30 September 2025
Read the latest preprint version here
Abstract
There is an urgent need to evaluate the toxicity of xenobiotics and complex mixtures the prevention the degradation of water quality sustaining aquatic ecosystems. A simple biogenic chemical pathway based on malate formation from pyruvate (pyr) and glyoxalate (glyox) pathway is proposed as quick and cheap screening tool for toxicity assessment. The assay is based on pyr and glyox (aldol) pathway leading to biologically relevant precursors such as oxaloacetate and malate. Incubation of pyr and glyox at 40-70oC in the presence of reduced iron (FeII) led to formation of malate following the first 3 h of incubation. The addition of various xenobiotics/contaminants (silver, copper, zinc, cerium IV, samarium III, dibytlphthalate, 1,3-diphenylguanidine, carbon-walled nanotube, nanoFe2O3 and polystyrene nanoparticles) led to inhibitions in malate at various degrees. Based on the concentration inhibiting malate concentrations by 20% (IC20), the following potencies were observed: silver < copper ~1.3-diphenylguanidine ~ carbon walled nanotube < zinc ~ samarium < dibutylphthalate ~ samarium < Ce(IV) < nFeO3< polystyrene nanoplastics. The IC20 values were also significantly correlated with the reported trout acute lethality data making this test a potential alternative test for trout. The per-glyox was also tested on surface water extracts (C18) and identified the most contaminated sites from large cities and municipal wastewater effluents. The inhibition potencies of the selected test compounds revealed that not only pro-oxidants are associated to toxicity but chemicals hindering enolate formation, nucleophilic attack of carbonyls and dehydration involved in aldol condensation reactions were also associated to toxicity. The per-glyox pathway is based on a fundamental aspect of chemical reactions during the emergence of life and represents a unique tool to identify toxic compounds individually and in complex mixtures.

Keywords:
prebiotic chemistry
; toxicity
; alternative method
; fish toxicity
; pyruvate
; glyoxalate
; aldo condensation
Introduction
Aquatic ecosystems are subjected to multiple sources of pollution releasing thousands of human- made xenobiotics though water (wastewaters and rainfall runoffs), air (dust, volatile compounds) and solids (solid waste, electronic waste, etc). It is generally accepted by the scientific community that measuring the physico-chemical properties of water requires toxicity assessments, which integrates the whole mixture effects and unknown chemicals [1]. The on-going toxicity evaluation of these complex mixtures is nowadays an integral part of ecotoxicological risk assessment strategies. They sometimes measured toxicity at different levels of biological organization i.e., at the biochemical, cellular, individual and multiple species/community levels. In the context of environmental protection strategies and law enforcement, bioassays investigating the toxicological properties of these chemicals and mixtures represent an on-going effort for ensuring sustainability of our economy. Bioassays usually encompass the selection of sentinel species at specific levels of the food chain where fish toxicity information is mandatory since it is protected and recognized by law [2]. For example, a test battery encompassing bacteria, algae/plants, invertebrates (worm, crustacean and bivalve) and vertebrates (fish and amphibians) have the advantage to not only determine the toxic potential of xenobiotics but in identifying the most sensitive species [3] and elaborate a toxicity distribution often used for probabilistic risk assessments of chemicals of emerging interests [4]. Fish toxicity testing for regulatory purposes leads to the sacrifice of tens of thousands of fish per country. In this context, the use of new alternative methods to reduce fish sacrifice is welcome not only for ethical reasons but reducing the cost of fish handling and exposure experiments during screening of either specific chemicals or environmental samples usually from agriculture/municipal/industrial wastewaters and street leachates. Alternatives should be at least as sensitive and predict toxicity to the fish assay of interest especially for complex mixtures being released into water bodies. Different types of alternatives exist albeit not systematically validated: fish at the hatching/larval stage (thought not to perceive pain), alternate organism (usually an invertebrate) and, cell cultures/lines, biosensors such as enzymes, biochemical pathways and artificial intelligence modelling in identifying potentially toxic chemicals based on various chemical properties. For example, the Hydra vulgaris bioassay and the peroxidase biosensor hold promise as alternatives given their high sensitivity and predictability towards rainbow trout acute lethality test for law enforcement [5,6]. Metabolic pathways have yet to be considered as alternative tests although their high potential as cost-effective alternatives. Metabolic pathways essentially determine whether xenobiotics could dampen parts of crucial metabolic networks such as the tricarboxylic acid (TCA) cycle for energy metabolism and cellular respiration involving the 5 universal intermediates: acetate, pyruvate, oxaloacetate, succinate and α-ketoglutarate [7]. These metabolites are formed in all life forms as precursors of the building blocks and functions: amino acids, lipids and nucleic acids.
In the selection of metabolic pathways as potential candidates for alternatives, primitive metabolic pathways thought to originate in early Earth before the formation of organized cells could be ideal candidates. These pathways intervene during the first biochemical reactions of life during the Hadean era of Earth [8]. Hence the use of protometabolic networks could serve as potential toxicity analogs and perhaps as fish alternatives provided that the generated toxicity data is sensitive and predictive towards fish. The TCA cycle is fundamental for cellular respiration and energy metabolism (ATP) and has been an attractive target for protobiotic chemistry research given that virtually all anabolic pathways in the metabolic pathways chart originate from the TCA cycle (metabolic pathways chart) [9]. Genomic studies suggest that an ancestor of the TCA cycle was formed during the formation of early metabolic pathways in the making of intermediates such as sugars, amino acids and lipids. So-called the reverse TCA (rTCA), it was autocatalytic before the formation of enzymes provided the intermediates were maintained in the cycle leading to the addition of carbons from the reduction of CO2 (abundant during early earth). The capacity of iron to promote the rTCA variant from simple prebiotic percursors - pyruvate (pyr) and glyoxalate (glyox) was experimentally established [10]. In addition, Pyr and glyox could be produced from abiotic processes of atmospheric CO2 fixation by reduced iron [11]. Reduced iron (FeII) was/is one of the most abundant metals in Earth’s crust and large amounts of FeII prior to the rise of oxygen was readily available [12]. The reaction is surprisingly simple requiring only heating a mixture of pyr, glyox and FeII under inert conditions (N2 atmosphere) for a few hours. The studies revealed the formation of highly interconnected pathways capable of both anabolic and catabolic activities involving 5 main reaction mechanisms: decarboxylation, reduction/oxidation, dehydration/hydration, and aldol/retro-aldol reactions. Aldol condensation, in turn, involves the formation of enolate ions (pyr), nucleophilic attack to the carbonyl/aldehyde carbon and dehydration (Figure 1). It is conceivable that xenobiotics hinfering these prebiotic reactions have the potential to initiate toxicity.
The purpose of this study was therefore to examine the pyr-glyox pathway to probe the effects of selected xenobiotics in the formation of the end product malate. The hypothesis states that xenobiotics capable of hindering malate formation from the primitive pyr-glyox pathway are related to toxicity. The influence of 10 well known contaminants of emerging interests (plasticizers, endocrine disruptors, plastic/carbon/iron-based nanoparticles, tire wear substances, metals and rare earth elements) toward the inhibition of the pyr-glyox pathway was examined and compared to fish mortality data. Moreover, malate production was examined in real-life river water and wastewater extracts to determine the most impacted/polluted samples. An attempt was made to understand the universal mechanisms associated to the initiation of toxicity beyond the oxidative stress paradigm during the initiation of toxicity [13].
Materials and Methods
Chemicals and Water Extraction
The following chemicals were purchased at 99% purity for copper (CuCl2), silver (AgNO3), samarium (SmCl3), cerium (CeCl4), zinc (ZnSO4), the plasticizer/endocrine disruptor dibutylphthalate (DBP) and tire related compound 1,3-diphenylguanidine (1,3-DPG) were purchased from Sigma Chemical Company (On, Canada). They were prepared in MilliQ water at 1mg/mL with the exception DBP and 1,3-DPG which were prepared in ethanol at the same concentration. Polystyrene nanoparticles of 20 nm diameter were purchased from ThermoFischer Scientific (USA). It was diluted in MilliQ water at 0.1%. Carbon nanotubes (CNT) of 5-10 nm internal diameter/1-3 µm fiber length and iron (II) nanoparticles (nFe(II)2O3, 30 nm diameter) were purchased at US Research Nanomaterials (Houston, TX, USA). They were diluted at 10 mg/mL in MilliQ water to prevent aggregation [14]. The stock solutions were prepared the day before the exposure experiments. All concentrations are nominal concentrations.
Surface waters, untreated wastewaters (influent) and treated wastewaters (effluent) were collected as composites of surface waters (3 x 1l samples per sites) and 24h composites (1 l) for surface waters and municipal wastewaters respectively. The surface water were collected in the Saint-Lawrence in the Montreal area: at the mouth of the Chateaughay river (CHA) on the south shore of the island of Montreal, one site 8 km downstream the City of Montréal (Downs), at the marina of the city of Lavaltrie (LA) located some 40 km downstream of the island of Montreal on the north shore of the Saint-Lawrence river. The untreated (influent) and treated wastewaters (effluent) were collected from typical city with a population of circa 100 000 residents using a primary clarifier (ferric chloride to remove phosphates) followed by an activated sludge and chlorine disinfection (sodium sulfite for dichlorination) treatments. A volume of 200 ml sample was filtered on 0.8 µm cellulose acetate membrane for the removal suspended solids and the filtrate passed through a C18 solid phase extraction cartridge (Bond Elut, 100 mg bed mass), washed with 5 mL of MilliQ water and eluted with 1 mL ethanol corresponding to 200 X concentrates. The ethanol extracts were kept at 4oC until analysis the same week.
Pyruvate-Glyoxylate Assay
The pyruvate-glyoxylate assay was prepared following a modification of a previously reported methodology [9,10]. The main modifications were the reduction of reactant concentrations to increase its sensitivity to exposure to various chemicals and complex mixtures in the environment and the optimal detection of the end products malate and oxaloacetate using an enzyme detection system. Briefly, 1 mM of sodium pyruvate, 2 mM glyoxalate and 4 mM FeSO4 were prepared in 1 mL of MilliQ water and increasing concentrations of the selected xenobiotics or water extracts. The mixture was degassed under N2 stream and incubated at 70oC for 3h. The control contained only MilliQ water or the equivalent amount of ethanol (C18 extract solvent). The reaction was cooled down at room temperature and the levels of malate were determined by malate dehydrogenase enzyme system. A 10-20 µl sample of the reaction mixture was mixed with 1 mM NAD+ in the assay buffer composed of 140 mM NaCl, 1 mM MgCl2 and 20 mM Tris base, pH 8.5 in 96-well dark microplates. The reaction was initiated by adding 10 µL of 10 units of malate dehydrogenase (10 units of MDH; Sigma Chemical Company, On, Canada). An operational blank was composed of NAD+ alone and malate standards (0.05-0.1 mM) in the presence of NAD+ and MDH was also included. The formation of reduced NADH was measured for 20 min at 1 min intervals by fluorescence at 350 nm excitation and 450 nm emission wavelengths using a microplate reader (Synergy IV, Biotek instruments, CA, USA). The rate of NADH formation was calculated for 10 min and used to calculate malate concentration in duplicate assay samples. The data was expressed as µmole/ml malate. The rainbow trout or other fish species lethality data were provided in the literature with the above compounds for comparison of malate inhibition with toxic effects on trout survival. With the exception of PsNPs and FeII- based nanoparticles (nFe3O4), the acute lethality tests were performed following the standard protocol of Environment and Climate Change Canada [15].
Data Analysis
The experiments were repeated N=3 times and the date were subjected to an analysis of variance after checking for data homogeneity using the Levene’s test. Critical differences between controls and exposure concentrations were determined by the Least Square Difference test. The calculation of the concentration of the test samples that inhibited malate formation by 20% (IC20) was calculated based on visual analysis of the concentration-response responses. The lethal concentration that caused trout mortality by 50% (LC50) was calculated by the Spearman-Karber methodology [16]. The data were expressed in mg/L for single substances and X fold concentration for complex mixtures (C18 ethanol extracts) where 1 X concentration correspond to the original undiluted water sample.
Results and Discussion
The pyr-glyox reaction involves aldol condensation reaction for the 2 carbon addition of pyr to glyox as depicted in Figure 1A. The newly formed metabolite hydroxyglutarate from glyox and pyr undergoes a series of hydration/dehydration, oxido/reductive and additional aldol condensation reactions forming more complex molecules such as citrate, succinate, malate and oxaloacetate (Figure 1B). The source of electrons is assured by Fe(II), which represents one of the most abundant and available metal in Earth crust prior to the Grand Oxidation event [12]. When glyox and pyr are added at a molar ration 2:1 in the presence of 4 equivalents of reduced Fe(II) in MilliQ water, malate (and oxaloacetate) are detected following 1 to 4 hr incubation at 70oC (Figure 2). Longer exposure times to 24h lead to increased levels of malate as well using the original protocol but was malate levels did not increased after 4 h when using lower concentrations of pyr, glyox and FeII. Indeed, the initial protocols used concentration in the 0.1 to 0.4 M range, representing concentration levels much higher than environmental contaminants. The concentration was downscaled to the mM range, which improved the sensitivity of the assay towards environmental pollutants permitting malate detection using malate dehydrogenase assay. In these conditions, longer exposure times (24 h) at 70oC did not lead to significant increase in malate, probably by saturation of the reaction or heat degradation. Hence, for toxicity experiments the following concentrations of the reagents were used: 1 mM Pyr, 2 mM glyoxalate and 4 mM Fe(II) at 70oC for 1-3h.
The pyr-glyox reaction was examined with increasing concentrations of the selected environmental contaminants (Figure 3A). The figure shows that increasing the concentrations of Ag, Zn and 1,3-DPG decreased production of malate at various intensities. Not only the pro-oxidants compounds Ag+ decreased malate production but zinc and the tire reducer 1,3-DPG as well. This suggests that not only oxidative chemicals can inhibit the pyr-glyox reaction but other chemicals as well. It is generally thought that the oxidative properties compounds are one of the main drivers of toxicity of xenobiotics [13]. The concentration that inhibits malate concentration by 20% (IC) was calculated and reported in Table 1. The IC20 spans from 1 to 150 mg/L with the following decreasing potency to inhibit the pyr-glyox reaction: Ag > copper ~1.3-diphenylguanidine ~ carbon walled nanotube > zinc ~ samarium > dibutylphthalate ~ samarium > Ce(IV) > nFeO3 > polystyrene nanoplastics. In respect to fish toxicity, the LC50 ranged from 0.1 to 100 mg/L in the following decreasing order of toxicity: silver > copper > zinc ~ samarium ~ dibutylphthalate > 1.3-diphenuylguanidine > carbon walled nanotubes > nFe2O3 > polystyrene nanoplastics. Based on these distributions, it appears that the per-glyox pathway is as sensitive to oxidants than fish based on copper and Ag responses albeit at concentrations 10 times higher than for rainbow trout. This could be attributed to the initial concentration of pyr (87 mg/L) and glyox (140 mg/L) used here but could be reduced at lower concentrations permitted by the malate detection system in place. The per-glyox reaction seems to respond more to electron donors (reductants) than rainbow trout such as 1,3-DPG and carbon (graphene) walled nanotubes in the presence of reduced iron Fe(II). The pyr-glyox pathway involves not only oxidative reactions but on aldol condensation as well. Aldol condensation involves 3 major steps: 1) the formation of the enolate ion from the mobility of hydrogen at the α carbon next to the carbonyl (pyr), 2) nucleophilic attack at aldehyde carbon (glyox) and followed by 3) dehydration of the carbonyl compound (i.e, hydroxyketogluatarate in Figure 1B) leading to unsaturated carbons. The mobility of hydrogen at the α carbon (of an Cδ-----Hδ+ ) next to carbonyl (pyruvate) could be inhibited by xenobiotics such as guanidines or acidic groups (carboxylic acid, thiols) in alkaline conditions [17]. Hence, this encompasses a large range of polar compounds such as pesticides, pharmaceuticals and industrial pollutants. For example, guanidines are used in the production of plastics (plastic tubing in households) and tire rubbers such as 1.3-DPG and 2-cyanoguanidine [18]. They are also considered chaotropic agents able to denature proteins and nucleic acids [19,20]. The former study also showed that excess oxidized FeIII, cadmium, lead, manganese and aluminum were also able to inhibit this reaction perhaps at the second step involving the nucleophilic reaction of the enolate ion. The guanidine containing drug metformin was shown to inhibit aconitase reaction involving the dehydration of isocitrate to citrate in the tricarboxylic acid cycle suggesting interactions at the 3rd step of aldol condensation as well [21]. Acrylamide and nucleophile maleimide were also shown to disrupt the cyclic production of thiol from amino acids thioester conjugates, which also involve nucleophilic reactions with thiols as with aldol reactions [22]. This suggests that electron donors/nucleophiles (RSH, maleimides, amines etc) could inhibit the pyr-glyox pathway but at different steps of the aldol condensation reactions. The aldol condensation of 2-keto-4-hydroxybutyrate from glyox and pyr catalyzed by aldolase in bovine liver and Escherichia coli also revealed inhibitions by various agents [23]. It was found that halides (anions) and carboxylic acids were inhibitors while esters were not as potent inhibitors of the condensation reaction of glyox and pyr. Mono-, di-, and tricarboxylic acids were proportionally inhibitory and hydroxypyruvate (blocks the mobility of H+ at the α carbon and the formation of enolate ion) was the most potent inhibitor. Increasing the concentration of glyox relative to pyr was also inhibitory perhaps through the enzyme Inactivation by this aldehyde. Hence, the pyr-glyox reaction involves reactions beyond the classic oxidative paradigm of toxic compounds. Indeed, compounds able to disrupt the formation of enols from the mobility of hydrogen of α carbon adjacent to carbonyl, block/competitive nucleophilic reaction to aldehydes and dehydration steps are involved in the pyr-glyox pathway where inhibitions in these reactions could initiate toxicity at the molecular level.
In the attempt to provide a toxicological meaning towards this prebiotic reaction network, the malate IC20 was compared with the rainbow trout acute lethality tests (Figure 3B). The data show that concentrations able to reduce malate production was significantly correlated to trout toxicity (LC50) data (r=0.84; p<0.001). This suggests that compounds able to inhibit aldol-condensation and/or redox properties are involved in the toxicity of these substances. The oxidative properties of xenobiotics represent one of the most fundamental interactions in toxicity initiation [13,24]. From the 10 selected compounds, 3 were nanoparticles (nFe2O3, PSNPs and CNT) and were shown to reduced malate production at relatively higher concentrations, in keeping with their relatively low toxicity (compared to Ag or Cu) in fish and hydra in previous studies [25,26]. However, plastic nanoparticles produce oxidative stress at the mg/L range in hydra [26,27] suggesting impacts at longer exposure times. In this respect, the pyr-glyox pathway could serve as generic test for toxicity screening of miscellaneous chemicals and mixtures. Keeping this in mind, we used the per-glyox reaction on C18 ethanolic extracts of various surface waters and municipal effluents (Figure 4). The samples consisted of various environmental liquids such as river water (CHA, LA, Downs), untreated (influents) and treated effluents. The data revealed that river water samples did not produce significant changes in the reaction at concentration reaching 20X of the water sample. The downstream site (DOWNS, 10 km downstream) of a 1.8 million population city inhibited the pyr-glyox pathway at 20 X concentration and to some extent at 4X but not at environmentally realistic concentration since no significant changes occurred at < 1 X (0.8) concentration. While the influent did not produce any significant changes in malate levels, the treated municipal effluent however, significantly inhibited malate formation at concentrations of 0.8 X and higher. This suggests that this effluent might pose a risk to aquatic life near the discharge point. In order to preserve anonymity of the city involved, fish toxicity of this effluent was previously reported on some occasions. This effluent is produced from an activated sludge effluent with residency times of 10-20 days where ferric chloride for phosphate removal and disinfection (UV) steps was applied. Activated sludges of a city of similar size revealed substantial amounts of plastic (polystryrene) polymers (10-20 µg/L) and polyaromatic hydrocarbons (30-40 µg/L) in effluents [28]. The levels of melamine and polystyrene nanoplastics in the effluents were higher than in the influent, suggesting that sustained effluent treatment breaks down the suspended organic matter leading to the release of dissolved components such as melamine-based products (paints and panels) and plastic nanoparticles [29]. This could explain why stronger inhibitions were observed in this secondary treated aeration sludge effluents than the other samples. Moreover, 1,3-DPG (0.04 µg/L) and cyanoguanidine (2.4 µg/L), constituents of tire and tire wear dusts, are finding their way in the dissolved fraction of municipal effluents [30].

In conclusion, a rapid and cost-effective chemical test is proposed as a screening tool for miscellaneous environmental liquids. The chemical is based on biochemical reactions between biogenic precursors pyr and glyox as primitive biochemical pathways thought to originate in early earth. The inhibition potencies of the selected test compounds revealed that not only pro-oxidants are associated to toxicity but chemicals hindering enolate formation, nucleophilic attack of carbonyls and dehydration. Furthermore, comparative analysis with fish toxicity (LC50) of per-glyox inhibitions revealed a significant correspondence making this reaction test a potential alternative methodology for fish toxicity of xenobiotics.
References
- Ten Brink, B.J.E.; Woudstra, J.H. Towards an effective and rational water management: the aquatic outlook project-integrating water management, monitoring and research. Eur. Water Pollut. Control 1991, 1, 20–27. [Google Scholar]
- Fishery Act. Published by the Minister of Justice 2025, R.S.; c. F-14, s. Fishery Act. Published by the Minister of Justice 2025, R.S.; c. F-14, s. 1, Canada, 111p.
- Costan, G.; Bermingham, N.; Blaise, C.; Ferard, J.F. Potential ecotoxic effects probe (PEEP): a novel index to assess and compare the toxic potential of industrial effluents. Environ. Toxicol. Wat. Qual. 1993, 18, 117–140. [Google Scholar] [CrossRef]
- Yang, H.; Sun, F.; Liao, H.; Guo, Y.; Wang, J.; Wu, F. Quantitative characteristics and multiple probabilistic risk assessment of small-sized microplastics in the middle and lower reaches of the Hanjiang River, China. Mar. Poll. Bull. 2025, 220, 118404. [Google Scholar] [CrossRef]
- Farley, G.; Bouchard, P.; Faille, C.; Trottier, S.; Gagné, F. Towards the standardization of Hydra vulgaris bioassay for toxicity assessments of liquid samples. Ecotoxicol. Environ. Saf. 2025, 290, 117560. [Google Scholar] [CrossRef]
- Gagné, F.; Roubeau Dumont, E.; Chantale, A. The effects of selected metals and rare earth elements on the peroxidase toxicity assay. Adv. Earth Environ. Sci, 2025, 6, 1–8. [Google Scholar]
- Braakman, R.; Smith, E. The compositional and evolutionary logic of metabolism. Phys. Biol 2013, 10, 011001. [Google Scholar] [CrossRef]
- Nogal, N.; Sanz-Sanchez, M.; Vela-Gallego, S.; Ruiz-Mirazo, K.; de la Escosura, A. The protometabolic nature of prebiotic chemistry. Chem. Soc. Rev. 2023, 52, 7359. [Google Scholar] [CrossRef]
- Tran, P.Q.; Adam, Z.R.; Fahrenbach, A.C. Prebiotic Reaction Networks in Water. Life 2020, 10, 352. [Google Scholar] [CrossRef] [PubMed]
- Muchowska, K.B.; Varma, S.J.; Moran, J. Synthesis and breakdown of universal metabolic precursors promoted by Iron. Nature 2019, 569, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Varma, S.J.; Muchowska, K.B.; Chatelain, P.; Moran, J. Native iron reduces CO2 to intermediates and end-Products of the acetyl-CoA Pathway. Nat. Ecol. Evol. 2018, 2, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Emsley, J. The Elements 2019, Oxford University Press, NY, USA, 256 pp.
- Villamena, F.; Forman, H.J. Molecular basis of oxidative stress: chemistry, toxicology, disease pathogenesis, diagnosis, and therapeutics, John Wiley & Sons, Inc. 25 ISBN:9781119790266. 20 January.
- Shrestha, S.; Wang, B.; Dutta, P. Nanoparticle processing: Understanding and controlling aggregation. Adv. Coll. Interf. Sci. 2020, 279, 102162. [Google Scholar] [CrossRef]
- Environment Canada. Biological test method: acute lethality test using rainbow trout environmental protection series 2007, report EPS 1/RM/9.
- Finney, D.J. Statistical Method in Biological Assay; Hafner Publishing Company: New York, NY, USA, 1964; 668p. [Google Scholar]
- Bhajiwala, H.M.; Patil, H.R.; Gupta, V. Studies of amino acids for inhibition of aldol condensation and dissolution of polymeric product of aldehyde in alkaline media. Appl. Petrochem. Res. 2013, 3, 17–23. [Google Scholar] [CrossRef]
- dos Santos, M.M.; Snyder, S.A. Occurrence of polymer additives 1,3-Diphenylguanidine (DPG), N-(1,3-Dimethylbutyl)-N′-phenyl-1,4-benzenediamine (6PPD), and chlorinated byproducts in drinking water: contribution from plumbing polymer materials. Environ. Sci. Technol. Lett. 2023, 10, 10–885. [Google Scholar] [CrossRef]
- Maxwell, K.L.; Bona, D.; Liu, C.; Arrowsmith, C.H.; Edwards, A.M. Refolding out of guanidine hydrochloride is an effective approach for high-throughput structural studies of small proteins. Protein Sci. 2003, 12, 2073–80. [Google Scholar] [CrossRef] [PubMed]
- Blanco, F.; Kelly, B.; Sánchez-Sanz, G.; Trujillo, C.; Alkorta, I.; Elguero, J.; Rozas, I. Non-covalent interactions: complexes of guanidinium with DNA and RNA nucleobases. J. Phys. Chem. B 2013, 117, 11608–16. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, A.; Peana, M.; Arshad, M.; Butnariu, M.; Menzel, A.; Bjørklund, G. Krebs cycle: activators, inhibitors and their roles in the modulation of carcinogenesis Arch. Toxicol. 2021, 95, 1161–1178. [Google Scholar] [CrossRef]
- Cafferty, B.J.; Wong, A.S.Y.; Semenov, S.N.; Belding, L.; Gmür, S.; Huck, W.T.S.; Whitesides, G.M. Robustness, entrainment, and ybridization in dissipative molecular networks, and the origin of life. J. Am. Chem. Soc. 2019, 141, 8289–8295. [Google Scholar] [CrossRef]
- Grady, S.R.; Wang, J.K.; Dekker, E.E. Steady-state kinetics and inhibition studies of the aldol condensation reaction catalyzed by bovine liver and Escherichia coli 2-keto-4-hydroxyglutarate aldolase. Biochemistry 1981, 20, 2497–2502. [Google Scholar] [CrossRef]
- Deavall, D.G.; Martin, E.A.; Horner, J.M.; Roberts, R. Drug-Induced Oxidative Stress and Toxicity. J Toxicol. 2012, 645460. [Google Scholar] [CrossRef]
- Ali, D.; Falodah, F.A.; Almutairi, B.; Alkahtani, S.; Alarifi, S. 2020. Assessment of DNA damage and oxidative stress in juvenile Channa punctatus (Bloch) after exposure to multi-walled carbon nanotubes. Environ. Toxicol. 2020, 35, 359–367. [Google Scholar] [CrossRef]
- Auclair, J.; Quinn, B.; Peyrot, C.; Wilkinson, K.J.; Gagné, F. Detection, biophysical effects, and toxicity of polystyrene nanoparticles to the cnidarian Hydra attenuata. Environ. Sci. Poll. Res. Int. 2020, 27, 11772–11781. [Google Scholar] [CrossRef]
- Auclair, J.; Roubeau Dumont, E.; Gagné, F. Lethal and sublethal toxicity of nanosilver and carbon nanotube composites to Hydra vulgaris—A toxicogenomic approach. Nanomaterials 2024, 14, 1955. [Google Scholar] [CrossRef] [PubMed]
- André, C.; Smyth, S.A.; Gagné, F. The peroxidase toxicity assay for the rapid evaluation of municipal effluent quality. Water Emerg. Contam. Nanoplast. 2025, 101, 2. [Google Scholar]
- Gagné, F.; André, C.; Smyth, S.-A. Screening of municipal effluents with the peroxidase toxicity assay. Disc. Wat. 2024, 4, 101. [Google Scholar]
- Ichihara, M.; Asakawa, D.; Yamamoto, A.; Sudo, M. Quantitation of guanidine derivatives as representative persistent and mobile organic compounds in water: method development. Anal. Bioanal. Chem. 2023, 415, 1953–1965. [Google Scholar] [CrossRef]
- Brooke, D.N.; Nielsen, I.R.; Dobson, S.; Howe, P.D. Environmental hazard assessment: Di-(2-ethylhexyl) phthalate. TSD 2, 1991. UK Department of the Environment, Garston, United Kingdom.
- Brix, K.V.; Tellis, M.S.; Crémazyc, A.; Wood, C.M. Characterization of the effects of binary metal mixtures on short-term uptake of Ag, Cu, and Ni by rainbow trout (Oncorhynchus mykiss) Aquatic Toxicol. 2016, 180, 236–246.
- Delahaut, V.; Chen, P.J.; Tan, S.W.; Wu, W.L.; Rašković, B.; Salvado, M.S.; Bervoets, L.; Blust, R.; De Boeck, G. Toxicity and bioaccumulation of Cadmium, Copper and Zinc in a direct comparison at equitoxic concentrations in common carp (Cyprinus carpio) juveniles. PLoS One 2020, 15, e0220485. [Google Scholar] [CrossRef]
- Dubé, M.; Auclair, J.; Hanana, H.; Turcotte, P.; Gagnon, C.; Gagné, F. Gene expression changes and toxicity of selected rare earth elements in rainbow trout juveniles. Comp. Biochem. Physiol. 2019, 223C, 88–95. [Google Scholar] [CrossRef]
- Chen, P.J.; Tan, S.W.; Wu, W.L. Stabilization or oxidation of nanoscale zerovalent iron at environmentally relevant exposure changes bioavailability and toxicity in medaka fish. Environ. Sci. Technol. 2012, 46, 8431–8239. [Google Scholar] [CrossRef]
Figure 1.
The pyruvate glyoxalate pathway. The pyr-glyox pathway starts with aldol reaction depicted in A. The pathway between pyruvate and glyoxalate in the presence of reduced Fe2+ (B). The cycle involves aldol/retroaldol condensation (orange lines, e.g., pyr + glyox → hydroxyglutarate and isocitrate→succinate + glyoxalate for aldol and retroaldol reactions respectively), hydration/retrohydration (blue line, e.g., hydroxyglutarate→oxopentenedioate), oxidative decarboxylation (green arrows oxalohydroxyglutarate→isocitrate + CO2), reduction/oxidation (violet arrows, e.g., succinate→fumarate). From [9].
Figure 1.
The pyruvate glyoxalate pathway. The pyr-glyox pathway starts with aldol reaction depicted in A. The pathway between pyruvate and glyoxalate in the presence of reduced Fe2+ (B). The cycle involves aldol/retroaldol condensation (orange lines, e.g., pyr + glyox → hydroxyglutarate and isocitrate→succinate + glyoxalate for aldol and retroaldol reactions respectively), hydration/retrohydration (blue line, e.g., hydroxyglutarate→oxopentenedioate), oxidative decarboxylation (green arrows oxalohydroxyglutarate→isocitrate + CO2), reduction/oxidation (violet arrows, e.g., succinate→fumarate). From [9].
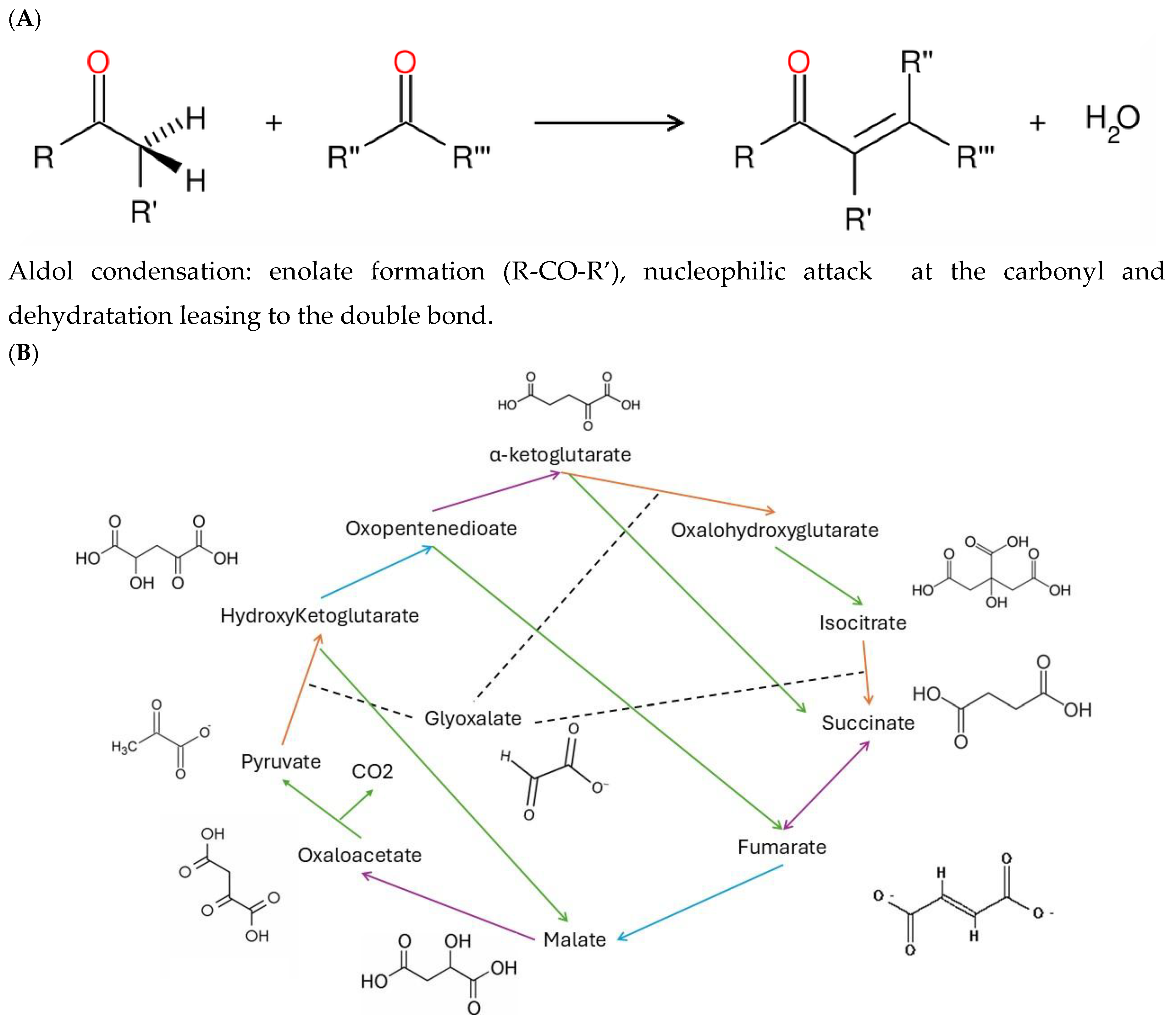
Figure 2.
Formation of malate following incubation of pyruvate and glyoxalate. Pyr, glyox and reduced iron (Fe2+) were allowed to react at 70oC for 1 and 3 h. Malate levels were determined by the NAD+ malate dehydrogenase system. A gradual increase in malate is observed relative to initial time 0h at 1 and 3h incubation time. The last column represents a malate standard (50 µM).
Figure 2.
Formation of malate following incubation of pyruvate and glyoxalate. Pyr, glyox and reduced iron (Fe2+) were allowed to react at 70oC for 1 and 3 h. Malate levels were determined by the NAD+ malate dehydrogenase system. A gradual increase in malate is observed relative to initial time 0h at 1 and 3h incubation time. The last column represents a malate standard (50 µM).
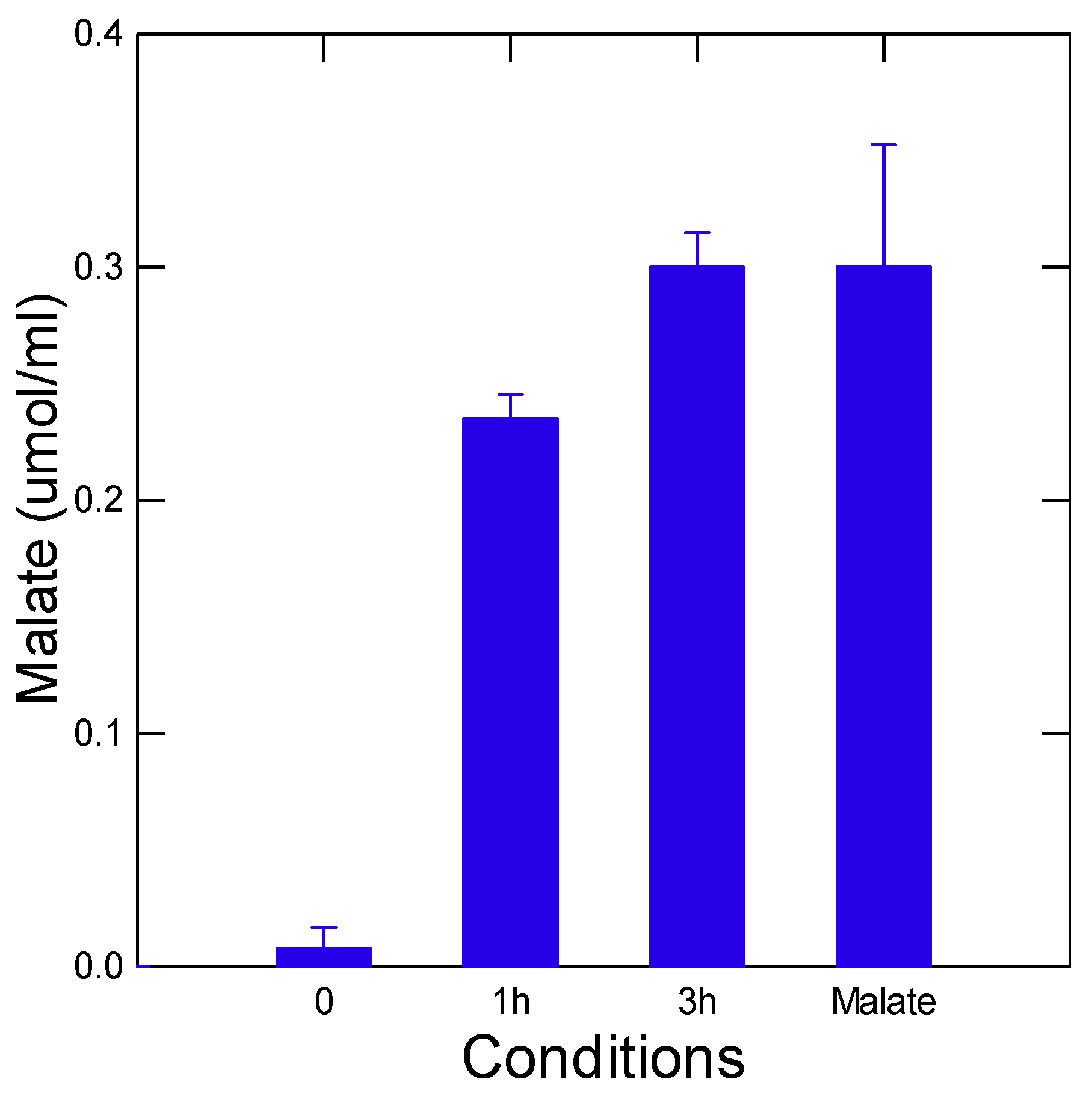
Figure 3.
Representation responses of malate formation inhibitions by selected toxic compounds towards rainbow trout. Representative inhibition profiles of malate production (A) and regression analysis of malate inhibition (log IC20 mg/l) and 96h trout mortality (log LC50 mg/l) (B) from reported literature data. The regression slope between the IC20 of the pyr-glyox reaction and trout 96h LC50 was obtained from N=10 compounds reported in Table 1: DBP, 1,3-DPG, Cu, Ag, Zn, Sm, Ce(IV), PSNPs, CWNT and nFe(II)3O4.
Figure 3.
Representation responses of malate formation inhibitions by selected toxic compounds towards rainbow trout. Representative inhibition profiles of malate production (A) and regression analysis of malate inhibition (log IC20 mg/l) and 96h trout mortality (log LC50 mg/l) (B) from reported literature data. The regression slope between the IC20 of the pyr-glyox reaction and trout 96h LC50 was obtained from N=10 compounds reported in Table 1: DBP, 1,3-DPG, Cu, Ag, Zn, Sm, Ce(IV), PSNPs, CWNT and nFe(II)3O4.
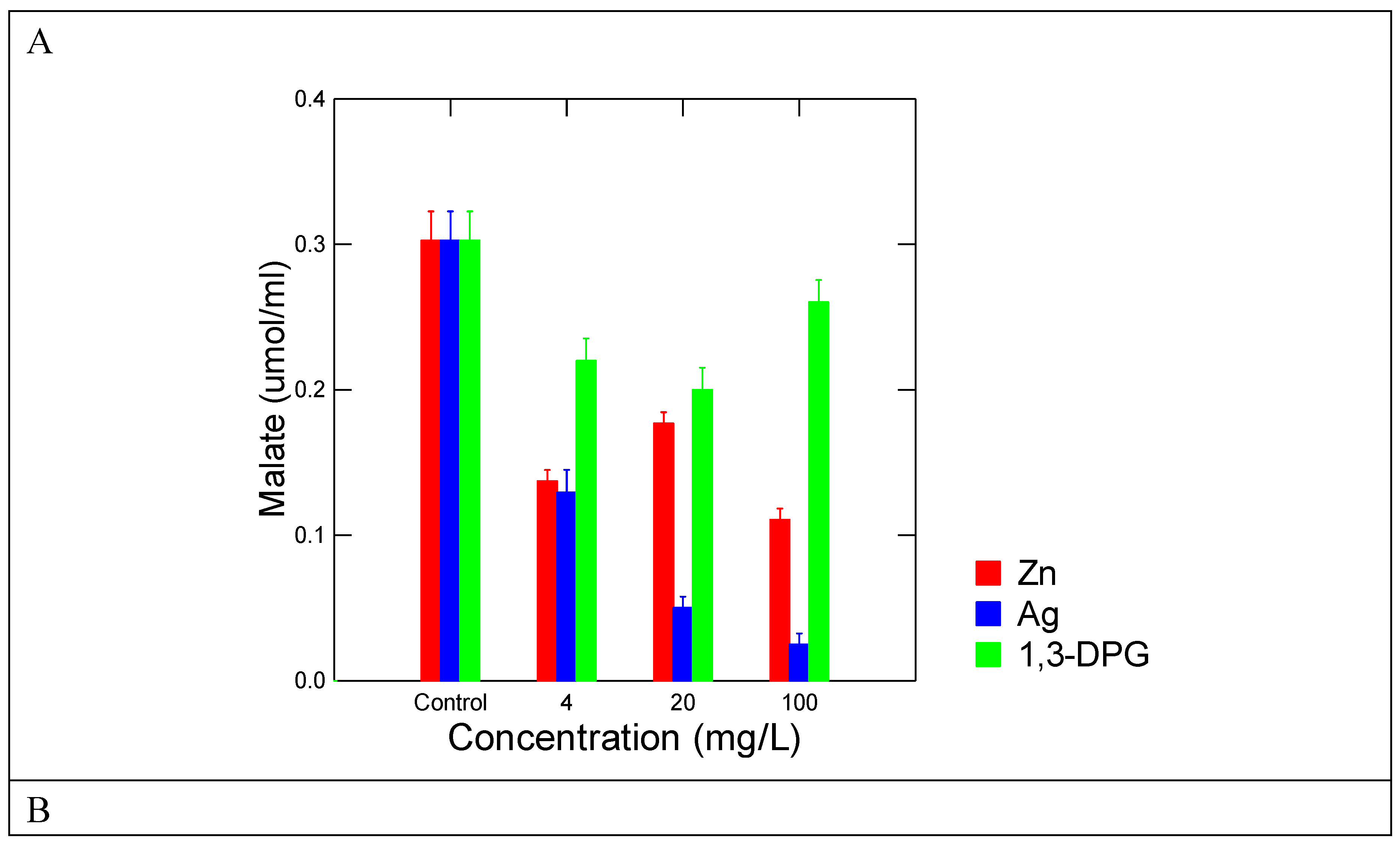
Figure 4.
Influence of surface and wastewaters to the pyruvate-glyoxalate pathway. Surface water and municipal influent/effluent extracts were examined with the pyr-glyox assay. The identification of the tested compounds and exposure concentration (expressed as X fold concentration) are identified on the abscissa axe.
Figure 4.
Influence of surface and wastewaters to the pyruvate-glyoxalate pathway. Surface water and municipal influent/effluent extracts were examined with the pyr-glyox assay. The identification of the tested compounds and exposure concentration (expressed as X fold concentration) are identified on the abscissa axe.
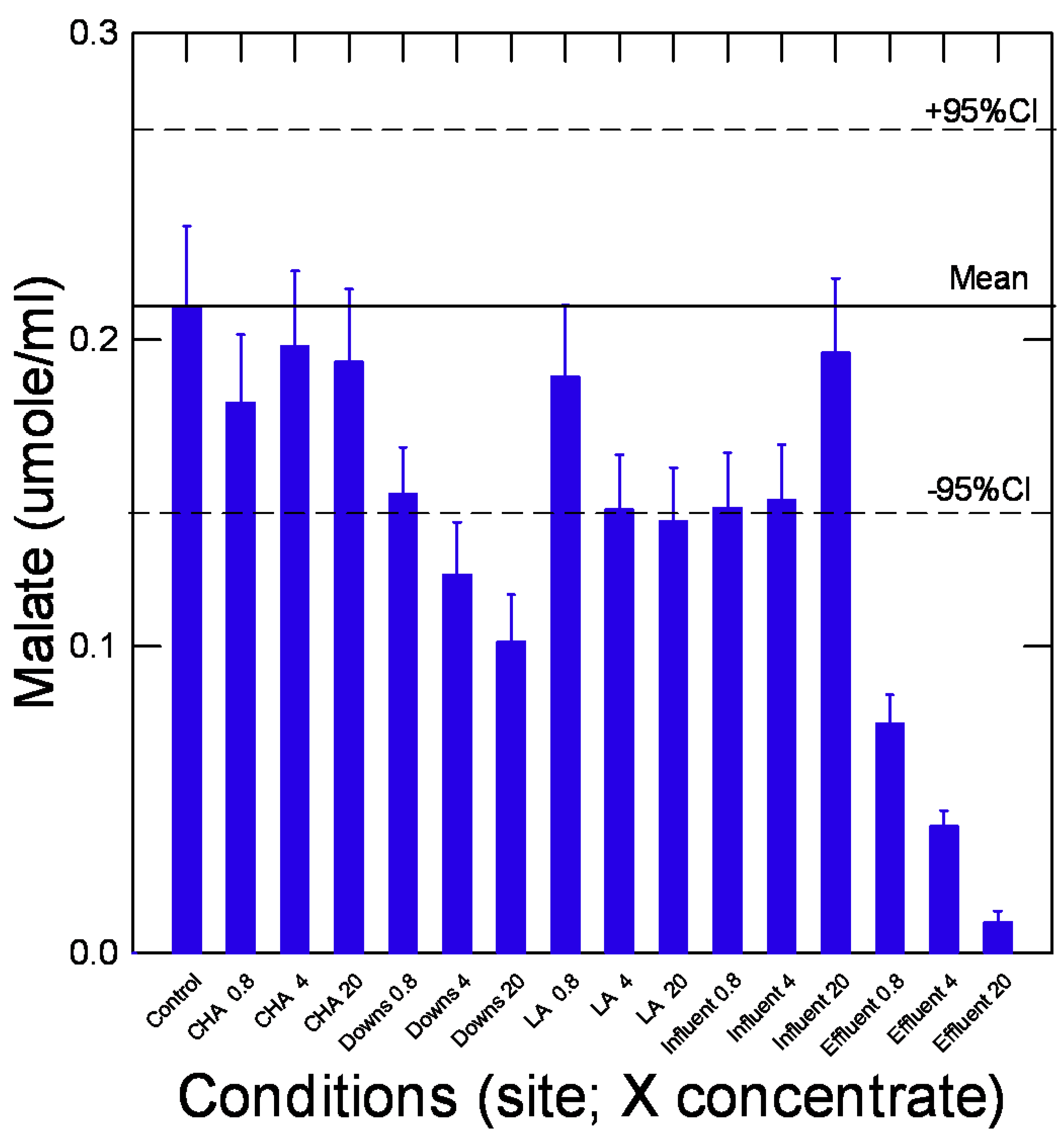

Table 1.
Selected compounds for the pyr-glyox reactions.
| Compounds |
Pyr-Glyox (IC20 mg/l) |
Trout toxicity (LC50 mg/l) |
References |
| Dibutylphthalate | 35 | 1.6 | [31] |
| 1,3-Diphenylguanidine | 12 | 11 | This lab |
| Copper(II) | 12 | 0.1 | [32] |
| Silver(I) | 1 | 0.02 | [32] |
| Zinc(II) | 41 | 1.6 | [32,33] |
| Samarium(III) | 24 | 2 | [34] |
| Cerium (IV) | 77 | 95 | [34] |
| nFe2O3 | 146 | 100 (Oryzias latipe embryo) |
[35] |
| Polystyrene nanoplastic (20 nm; uncoated) |
150 | >100 | This lab |
| Carbon walled nanotubes | 17 | 22 (Channa punctatus juvenile) |
[25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



