
Submitted:
26 September 2025
Posted:
29 September 2025
You are already at the latest version
Abstract
Alcohol use disorder (AUD) is a significant medical problem and there is great need for developing effective treatment strategies. Brain-Derived Neurotrophic Factor (BDNF) has been shown to play a role in regulating numerous pharmacological and motivational effects of alcohol. We have shown that chronic alcohol-induced escalation of drinking is accompanied by a deficit in BDNF levels in medial prefrontal cortex (mPFC). This study examined whether exercise (wheel-running) attenuates excessive alcohol drinking via increased BDNF expression, thereby mitigating the deficit in mPFC. Adult male C57BL/6J mice were given scheduled (2-hr/day) access to a running wheel in the home-cage 1-hr following opportunity to drink alcohol for 2-hr/day. After six weeks, mice were further separated into groups that received chronic alcohol vapor or control (air) inhalation exposure. Results indicated that alcohol consumption did not alter wheel-running and exercise did not alter alcohol intake during the 6-week baseline. Exercise increased BDNF mRNA and protein expression in mPFC, reversed chronic alcohol-induced reduction in BDNF levels, and attenuated escalated alcohol drinking. Systemic administration of a TrkB receptor antagonist (ANA-12) reversed the beneficial effects of wheel-running in the model. Together, these data provide support for exercise as a potentially effective intervention strategy for treating AUD.
Keywords:
Alcohol consumption
; Alcohol dependence
; Exercise
; Wheel-running
; BDNF (Brain-Derived Neurotrophic Factor)
; TrkB receptors (Tropomyosin receptor kinase-B receptors)
; Mouse
; Alcohol use disorder
1. Introduction
Alcohol use disorder (AUD) constitutes a major public health concern in the United States and world-wide. Epidemiology studies have indicated a steady rise in alcohol misuse and prevalence of AUD, with the Covid-19 pandemic having added fuel to this upward trajectory [1,2,3]. Excessive alcohol use has serious medical consequences, and alcohol misuse results in tremendous economic, social, and personal costs to society [4,5,6]. Despite significant advances in our understanding of biological mechanisms and environmental factors that influence motivation to engage in harmful levels of alcohol drinking, there remains a critical need for developing new and more effective therapeutic interventions for AUD [7,8,9,10].
AUD is a chronic relapsing disorder, and relapse represents a major challenge to treatment efforts. Currently, there are four medications approved by the FDA for treatment of AUD: disulfiram, oral naltrexone, acamprosate, and a long-acting injectable form of naltrexone. While these medications are effective for some individuals, there remains a great need for identifying and developing new treatment strategies for AUD. Preclinical studies have played a key role in discovery of new targets with promising therapeutic potential. Using our mouse model of alcohol dependence involving repeated cycles of chronic intermittent ethanol (CIE) vapor exposure, we conducted an unbiased genomic analysis of brain tissue and showed significant down-regulation in expression of Bdnf mRNA in medial prefrontal cortex (mPFC) of CIE-exposed mice compared to nondependent controls [11].
Brain-Derived Neurotrophic Factor (BDNF), a member of the neurotrophin family of growth factors, plays an important role in the development and maintenance of the nervous system. BDNF is widely distributed and highly expressed in mammalian brain, it is secreted from neurons in an activity-dependent manner, and BDNF interacts with tropomyosin receptor kinase B (TrkB) receptors and low-affinity p75 neurotrophin (p75NT) receptors to regulate a multitude of cellular processes [12]. BDNF binding at TrkB receptors results in phosphorylation of the receptor and activation of MAPK/ERK1/2, PLCg, and PI3K intracellular signaling pathways. BDNF signaling in brain has been implicated in a number of neuropsychiatric disorders, including AUD [13].
Humans with AUD are reported to have lower circulating levels of BDNF [14], although mixed results have been obtained in studies examining serum BDNF levels in abstinent individuals with a history of AUD diagnosis. [15]. The TrkB (Ntrk2) gene locus on chromosome 9 was shown to be associated with alcohol dependence [16], and a functional polymorphism in the Bdnf gene (Val66Met) has been implicated in AUD, with evidence that the Val66 allelic variant confers higher risk for relapse [17] while the 66Met variant is associated with earlier onset and greater severity of alcohol dependence [18]. Interestingly, mice carrying the homolog of the human 66Met allele (68Met) showed excessive alcohol consumption compared to wildtype Val68 mice [19]. Additional evidence for a connection between BDNF and alcohol-related behaviors comes from studies showing increased drinking in several genetic mouse and rat models with known deficiencies in BDNF expression [13]. Moderate alcohol consumption in mice was reported to increase Bdnf mRNA in the hippocampus [20] while high levels of intake achieved in a binge drinking rat model were associated with suppressed BDNF expression in hippocampus, an effect reversed by administration of a TrkB agonist [21]. Moderate alcohol intake increased Bdnf mRNA levels in dorsolateral (but not dorsomedial) striatum [22]. Also, while viral-mediated decreased BDNF expression in dorsal striatum increased alcohol drinking [22], increasing BDNF expression in this brain region reduced alcohol consumption, effects shown to be mediated by BDNF-TrkB receptor interaction [23]. A similar negative relationship between BDNF activity and alcohol consumption was shown for the central and medial (but not basolateral) nuclei of the amygdala [24]. We and others have shown reduced BDNF expression in mPFC following chronic alcohol self-administration [25,26] and CIE vapor exposure [11,26,27]. We also showed that replenishing BDNF levels in the mPFC either by direct injection of the neurotrophin or viral-mediated overexpression of BDNF reduced or prevented, respectively, escalation of dependence-related alcohol drinking [28]. While elevating BDNF activity in brain appears to have therapeutic potential in treating AUD, from a clinical standpoint, systemically administering the neurotrophic factor is not practical since it does not readily penetrate the blood brain barrier [29]. Consequently, we turned our attention to an alternative approach that increases central BDNF activity – physical exercise.
Exercise has received increased attention as an effective, non-pharmacological treatment approach for various neuropsychiatric illnesses, including PTSD and alcohol/drug addiction. Physical activity (wheel-running, treadmill) has been shown to generally reduce self-administration of alcohol and other drugs such as cocaine, methamphetamine, heroin, and nicotine [30,31]. Free (24-hr) access to wheel-running reduced home-cage alcohol consumption in several studies [32,33,34,35,36,37], although others reported different outcomes [38,39]. Numerous factors likely explain differences in study outcomes, including species/genotype, duration of exercise, timing of exercise during different stages of addiction, and temporal relationship between access to exercise and alcohol/drug availability [30,40,41]. Sex is another factor that significantly influences the effects of exercise on alcohol/drug addiction in animals and humans [40,42]. Clinical studies have reported exercise to reduce ethanol drinking [43,44] and reduce craving and relapse vulnerability in detoxified alcoholics [44,45]. The role of exercise in the treatment of AUD is likely multifaceted, as it improves physical and mental health [46], reduces stress [47] and, owing to its rewarding effects, it provides an alternative (rewarding-substitute) to alcohol/drug use [30,33]. Indeed, the stress-alleviating effects of exercise has led to its consideration as an adjunct treatment for veterans seeking treatment for alcohol/drug addiction [48]. From a mechanistic perspective, exercise (aerobic and resistance) is known to elevate BDNF activity in brain [30,42,49,50].
The present series of studies were conducted to test the general hypothesis that physical activity (wheel-running) reduces escalated alcohol consumption in our mouse model of alcohol dependence. It was hypothesized that exercise-induced increased BDNF production in brain would mitigate chronic alcohol-induced deficits in BDNF expression, thereby, attenuating escalated alcohol drinking. To more directly test whether this effect is related to increased BDNF activity at TrkB receptors, we examined whether administering a TrkB receptor antagonist would reverse the beneficial effects of exercise in our model of excessive alcohol drinking.
2. Results
While some studies have shown exercise reduces home-cage alcohol preference and consumption in mice, in many cases access to wheel-running and alcohol was continuous (24-hr) and simultaneous [33,34,35,36]. Since these are essentially mutually exclusive behaviors (wheel-running vs. drinking alcohol (or water) from a sipper tube), we chose to examine an exercise regimen that involved limited access (2-hr/day) to running wheels in the home-cage. Further, to ensure that exercise did not merely alter alcohol intake due to elevated exercise-induced thirst, alcohol was presented for 2-hr/day in the home-cage in a free choice (2-bottle) situation (15% ethanol vs. water) 1-hr prior to the 2-hr access to running wheels.
2.1. Limited Daily Access to Wheel-Running Increases BDNF Protein Expression in Medial Prefrontal Cortex and Dentate Gyrus
To determine whether daily scheduled access to an activity wheel in the home-cage increased BDNF expression in brain, adult male C57BL/6J mice were given access to activity wheels (WH groups) or not (NWH group) in the home cage for 2-hr/day starting at 3-hr into the dark phase for 6 weeks. Wheel-running was tracked to calculate distance, and mice were sacrificed at 24-hr (WH-24-hr) or 1 week (WH-1-wk) following the last session (half the NWH group was sacrificed at each of these time points and then combined for analysis) (N= 12 mice/group). Bilateral medial prefrontal cortex (mPFC) and dentate gyrus (DG) was dissected on ice using 1-mm microdissection punches, with the mouse brain atlas serving as a guide [51]. Samples were stored for protein analysis by ELISA. mPFC was targeted because we had previously shown that chronic alcohol exposure reduced BDNF levels in this brain region [11,27]. The DG was included in our analysis since this brain region has been shown to be highly responsive to exercise effects on BDNF activity [52,53].
Wheel-running stabilized over the 6-week period (2.5-3 km/2-hr session) and did not differ between groups that were sacrificed at 24-hr or 1-wk following the last exercise session (Figure 1A). ANOVA indicated no significant main effect of Group [F(1,22)= 0.36, p= 0.55] or Group x Week interaction [F(5,110)= 0.52, p= 0.67]. Compared to BDNF levels in the NWH group, BDNF protein expression in mPFC was significantly elevated 24-hr following the 6-week exercise regimen, with a trend of elevated levels at the 1-wk time point (Figure 1B). In the DG, BDNF protein expression was increased at 24-hr, and this persisted at least 1-week following wheel-running (Figure 1C). Analysis of mPFC indicated a main effect of Group [F(2,22)= 4.91, p= 0.02] and post-hoc analyses showing BDNF levels higher in WH-24-hr compared to the NWH group (p< 0.05). Similarly, for DG there was a main effect of Group [F(2,26)= 6.36, p= 0.01] and the elevated levels of BDNF protein was evident at 24-hr and persisted for at least 1-wk following exercise (ps< 0.05).
2.2. Effects of Alcohol Drinking on Wheel-Running and Bdnf mRNA Levels in mPFC
Next we determined whether limited access to alcohol drinking influenced wheel-running and its ability to elevate Bdnf mRNA expression in the mPFC. Adult male C57BL/6J mice were divided into 4 groups based on scheduled home-cage wheel and/or alcohol access (N= 12/group). One group was given access to alcohol (15% v/v) for 1-hr starting at 3-hr into the dark cycle with no opportunity to exercise (NWH-EtOH group); a second group was treated the same but was given access to activity wheels for 2-hr/day for 6 weeks, starting 1-hr after the alcohol bottle was removed (WH-EtOH group); another group was given access to the same regimen of wheel-running without alcohol access (WH-H2O group); and the final group served as controls, receiving neither access to running wheels nor alcohol (NWH-H2O group).
Wheel-running (2-hr/day/5-days/week for 6 weeks) was not significantly altered by alcohol consumption that occurred 1-hr before access to the activity wheels (Figure 2). That is, wheel-running remained stable and did not differ between WH-EtOH and WH-H2O groups over the 6-week study (Group: [F(1,19)= 0.49, p= 0.52]; Group x Week: [F(5,95)= 0.59, p= 0.71]). Additionally, scheduled access to wheel-running did not significantly influence alcohol intake (Figure 3). That is, alcohol consumption was similar for WH-EtOH and NWH-EtOH groups over the 6-week study (Group: [F(1,22)= 0.66, p= 0.42]; Group x Week: [F(5,110)= 2.25, p= 0.06]).
At 24-hr following the final session, all mice were sacrificed, and mPFC and DG samples were dissected and frozen until qRT-PCR analysis was performed targeting Bdnf exon-I, exon-IV, and exon-IX. Bdnf exon-I and exon-IV transcripts were selected because they have been shown to be especially responsive to exercise and chronic alcohol, respectively [26,27,54], and Bdnf exon-IX encodes the pre-proBDNF protein [55,56]. Wheel-running (2-hr/day) with or without home-cage access to alcohol (2-hr/day) over 6 weeks significantly increased Bdnf mRNA expression (exons I, IV, and IX) in mPFC (Figure 4A) and DG (Figure 4B). That is, compared to controls (NWH-H2O group), both WH-H2O and WH-EtOH groups evidenced elevated Bdnf mRNA levels in mPFC and DG. ANOVA indicated a main effect Wheel access for Bdnf exon-I (mPFC: [F(1,32)= 16.09, p< 0.001] DG: [F(1,30)= 18.71, p< 0.001]); exon-IV (mPFC: [F(1,32)= 11.68, p< 0.01] DG: [F(1,30)= 6.19, p< 0.025]); and exon-IX (mPFC: [F(1,32)= 10.72, p< 0.01] DG: [F(1,34)= 18.01, p< 0.001]). Moderate level of alcohol consumption alone (NWH-EtOH group) did not significantly alter BDNF transcriptional activity in either brain region. Thus, limited daily access (2-hr/day) to activity wheels over 6 weeks increased Bdnf mRNA expression in mPFC and DG, and limited home-cage access to alcohol (2-hr/day) did not modify this effect.
2.3. Wheel-Running Reverses Chronic Alcohol-Induced Deficits in mPFC Bdnf mRNA Expression and Attenuates Escalated Alcohol Drinking
In a separate cohort of mice, we examined the effects of exercise in our CIE Drinking model. Adult male C57BL/6J mice were given free access to alcohol (15% v/v) in the home-cage 2-hr/day 5-days/week during a 6-week baseline period, with half the mice presented with a running wheel for 2-hr in the home-cage 1-hr after alcohol was removed. The remaining mice were left undisturbed. WH and NWH groups were then divided into two groups, receiving either weekly cycles of chronic intermittent ethanol (CIE) vapor exposure (NWH-CIE and WH-CIE groups) or control (air) exposure (NWH-CLT and WH-CTL groups) alternating with weekly test drinking days (Figure 5). Wheel-running access and alcohol drinking was suspended during inhalation exposure cycles.
Wheel-running remained relatively stable during the 6-week baseline period (Supplemental Figure S1). During test cycles, a significant Group x Week interaction [F(3,102)= 4.11, p< 0.01] indicated that wheel-running was lower in the WH-CIE group compared to WH-CTL mice during the last two test cycles (ps< 0.05). Also, wheel-running did not appear to influence alcohol metabolism, as suggested by the similar blood alcohol levels produced during each of the CIE exposure cycles (Supplemental Table S1).
As previously reported, CIE exposure resulted in escalation of alcohol consumption compared to CTL groups, but providing scheduled opportunity to exercise attenuated this increase in drinking (Figure 6A). Analysis of alcohol intake (g/kg) data revealed a significant Group x Test interaction [F(12,304)= 9.14, p< 0.001]. Further analysis indicated that alcohol intake was similar for all groups at the end of the baseline period. Alcohol intake was significantly higher for the NWH-CIE group compared to their baseline level of intake and consumption in the NWH-CTL group during each Test cycle (NWH-CIE > NWH-CTL; ps< 0.05). Exercise significantly reduced this effect during Tests 2, 3, and 4 (NWH-CIE > WH-CIE; ps< 0.05). Alcohol intake remained relatively stable across Baseline and Test cycles for the NWH-CTL group, and wheel-running alone (WH-CTL) did not alter alcohol drinking throughout the study.
A similar profile of results was obtained when data were expressed as a change from each animal’s baseline level of intake (Figure 6B). Analysis of these data indicated a significant Group x Test cycle interaction [F(9,228)= 2.53, p< 0.01]. The NWH-CIE mice evidenced an escalation of drinking, with alcohol intake increasing from 66% during Test 1 to more than doubling (105%) during Test 4 (ps< 0.05). This escalated drinking was significantly attenuated in CIE-exposed mice that had scheduled daily access to a running wheel in their home-cage during baseline and each test cycle (NWH-CIE > WH-CIE, ps< 0.05). Alcohol intake remained relatively unchanged throughout the study in NWH-CTL and WH-CTL groups.
A subset of mice from each group received a 5th cycle of CIE (or air) exposure and were then sacrificed to determine Bdnf mRNA expression in mPFC (Figure 7). Analysis of each Bdnf transcript revealed significant group differences (exon I: [F(3,31)= 21.50,p< 0.001; exon IV: [F(3,31)= 33.48, p< 0.001], exon IX: [F(3,31)= 27.64, p< 0.001]).Overall, CIE exposure alone (NWH-CIE group) produced a deficit in Bdnf mRNA levels relative to the NWH-CTL condition (exon I: p=0.03[F(1,31)= 25.86, p< 0.001]; exon IV: p= 0.004[F(1,1,31)= 52.95, p< 0.001]; exon IX: p=0.001[F(1,31)= 50.81, p< 0.001]). Exercise alone (WH-CTL group) significantly elevated Bdnf mRNA expression in mPFC and this served to completely block the deficit in Bdnf mRNA levels produced by CIE exposure (NWH-CIE > WH-CIE) (exon I: p= 0.001[F(1,31)= 37.72, p< 0.001]; exon IV: p= 0.001[F(1,31)= 45.53, p< 0.001]; exon IX: p= 0.002[F(1,31)= 27.96, p< 0.001]). That is, reduced Bdnf mRNA expression in the mPFC produced by CIE exposure was normalized when mice were given the opportunity to exercise. (exonI; p=0.017; exon IV: p= 0.003; exon IX: p= 0.004This suggests that BDNF activity plays an important role in mediating escalation of drinking associated with chronic alcohol exposure.
2.4. TrkB Receptor Antagonism Blocks the Ability of Exercise to Attenuate CIE-Induced Escalated Drinking
To more directly investigate whether increased BDNF activity at TrkB receptors contributes to the ability of exercise to reduce chronic alcohol-induced escalated drinking, we examined the effects of a TrkB receptor antagonist, ANA-12, in the CIE Drinking model. Adult male mice were separated into NWH-CTL, NWH-CIE, WH-CTL, and WH-CIE groups and treated as described in the previous study. After the 2nd CIE/Air exposure cycle, the groups were further separated into drug treatment conditions, receiving injections of Vehicle, 0.5, or 1.0 mg/kg ANA-12 immediately following the 2-hr daily wheel-running sessions.
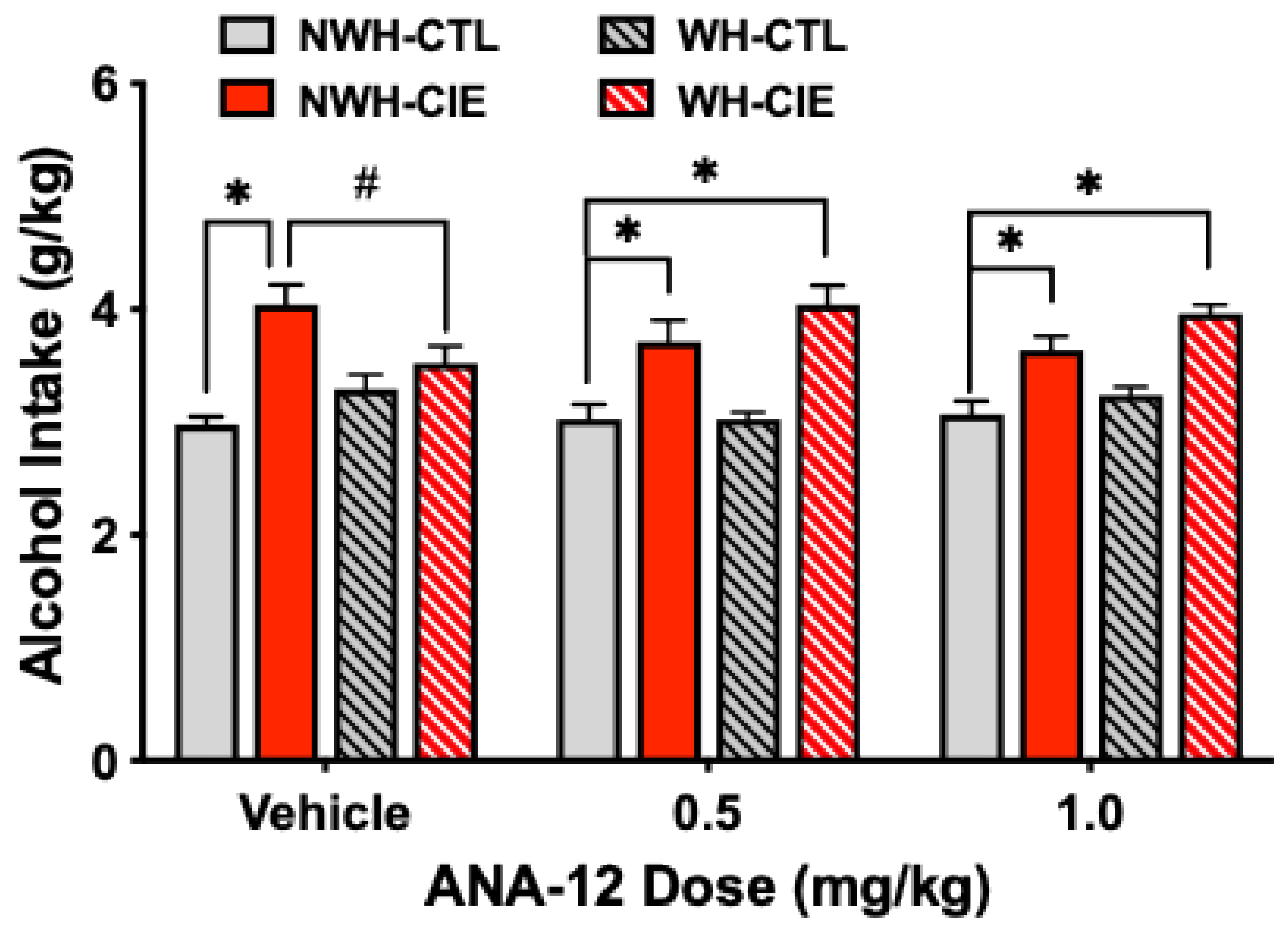
Analysis of alcohol consumption during Test 2 indicated a significant main effect of Group [F(3,102)=34.33, p<0.0001] and a significant Group x ANA-12 Dose interaction [F(6,102)=3.09, p<0.0081] (Figure 8). Post-hoc comparisons revealed higher alcohol intake in CIE-exposed mice compared to CTL mice with no access to running wheels (NWH-CIE > NWH-CTL), and ANA-12 treatment did not alter this effect (ps<0.05). Replicating our previous finding, exercise (wheel-running) significantly reduced alcohol intake in CIE-exposed mice that received Vehicle (NWH-CIE > WH-CIE; p< 0.05). However, the dampening effects of exercise on CIE-induced elevated drinking was eliminated in mice that were treated with the TrkB antagonist ANA-12. That is, alcohol intake in WH-CIE mice treated with either 0.5 mg/kg or 1.0 mg/kg ANA-12 did not differ from the corresponding NWH-CIE groups and these mice consumed significantly more alcohol in comparison to the NWH-CTL group (ps< 0.05). These results support the notion that exercise attenuates chronic alcohol-induced escalation of drinking by increasing BDNF activity at TrkB receptors.
3. Discussion
Results from this study demonstrate that providing mice with the opportunity to exercise 2-hr/day in their home-cage over several weeks is sufficient to elevate BDNF mRNA and protein expression in the hippocampus (dentate gyrus) and medial PFC. Chronic alcohol (CIE) exposure produced the expected escalation of alcohol drinking and, as we previously showed, this was accompanied by a significant reduction in Bdnf mRNA levels in mPFC. However, daily scheduled exercise attenuated escalated drinking in CIE-exposed mice without significantly altering alcohol intake in nondependent (CTL) mice. Further the daily scheduled exercise regimen mitigated chronic alcohol-induced reduction in Bdnf mRNA levels in mPFC. Finally, to more directly implicate a role for BDNF-TrkB receptor activity in this effect, the TrkB receptor antagonist ANA-12 was shown to block the ability of exercise to reduce CIE-induced escalated alcohol drinking.
It has been previously reported that continuous (unlimited) home-cage access to running wheels significantly increased BDNF mRNA and protein expression in the DG, an exercise responsive region [52,53]. Complimenting these findings, results from this study show that limited scheduled daily access (2-hr/day) to wheel-running over 6 weeks significantly increased BDNF mRNA and protein expression in the DG as well as the mPFC.
In the present series of studies, activity wheels were provided in the home-cage 1-hr following a 2-hr period of free-choice alcohol drinking. The temporal order and separation of these behaviors was implemented to avoid any immediate influence exercise may have on alcohol drinking due to the need for more calories or a general increase in thirst. Wheel-running (2-hr/day) did not alter alcohol consumption during a 6-week baseline period. Further, access to alcohol prior to wheel running did not affect running behavior (which remained relatively stable over 6 weeks), and alcohol intake did not influence the ability of exercise to elevate BDNF transcriptional activity in target brain regions.
It is interesting that this limited access exercise regimen selectively reduced excessive levels of alcohol drinking produced by CIE exposure while not altering more moderate stable levels of alcohol intake in non-dependent (CTL) mice. As previously noted, unlimited access to voluntary wheel-running has generally been shown to reduce alcohol consumption [32,33,34,35,36,37]. These studies have typically involved concurrent unlimited access to wheel running and alcohol. Other studies have examined effects of exercise on alcohol consumption under limited access conditions or when wheel-running was restricted to periods of abstinence, with mixed results [35,37,39]. Differences in study outcomes are likely related to various biological variables (e.g., species, strain, sex) as well as a host of experimental procedural factors (e.g., duration and temporal relationship of both exercise and alcohol access) [30,40,41,42]. All of these studies have examined the effects of physical activity on alcohol consumption using drinking models in nondependent animals. Here we demonstrate that scheduled limited access to wheel-running attenuates escalated alcohol drinking in dependent (CIE-exposed) mice. To our knowledge, there is only one other study that has examined the effects of wheel-running in dependent animals. Somkuwar et al. [57] reported that wheel-running access following CIE exposure in rats reduced alcohol relapse-like behavior using an operant reinstatement model. As such, these results complement findings of the present study and provide support for the potential therapeutic value of exercise in reducing excessive alcohol drinking and relapse vulnerability.
Studies have demonstrated a negative relationship between alcohol drinking behavior and BDNF expression in hippocampus [21], striatum [22,23], and amygdala [24]. In this study we show that increased alcohol consumption following chronic alcohol (CIE) exposure was associated with decreased Bdnf mRNA expression in mPFC. This aligns with our previous findings and that of others showing reduced BDNF expression in mPFC following chronic alcohol exposure [11,25,26,27]. Results from this study demonstrate that daily limited access to wheel-running not only elevated BDNF expression in mPFC but also reversed the chronic alcohol-induced reduction in Bdnf mRNA levels and, importantly, attenuated escalated voluntary alcohol drinking. These results support the notion that reversing the deficit in BDNF expression following chronic alcohol exposure can reduce alcohol consumption, and exercise may be an effective means to produce this effect.
Our previous work showed that replenishing BDNF levels by direct infusion of the neurotrophin into the mPFC reduced CIE-related escalation of drinking, Further, viral-mediated overexpression of BDNF in the mPFC was effective in preventing the emergence of escalated drinking in CIE-exposed mice [28]. In this study, physical activity (wheel-running) produced a similar effect in reducing CIE-induced elevated alcohol consumption. To further interrogate the hypothesized link between exercised-induced elevated BDNF expression and reduced alcohol drinking, we examined whether this effect of exercise could be reversed by administrating the TrkB receptor antagonist ANA-12. Results indicated that both doses of ANA-12 tested were equally effective in reversing the ability of exercise to reduce CIE-induced excessive drinking. ANA-12 treatment did not alter drinking in non-dependent mice. Similarly, elevating BDNF levels in mPFC did not alter alcohol intake in non-dependent mice [28]. Together, these results suggest that BDNF-TrkB signaling in the mPFC may be most relevant in modulating alcohol consumption under conditions that engender excessive levels of intake (i.e., models of chronic alcohol exposure that produce deficits in BDNF expression). However, since ANA-12 was systemically administered, it cannot be assumed that this outcome was due to blocking BDNF-TrkB receptor activity in the mPFC. Future studies will need to target pharmacological antagonism or genetic deletion of the receptor in specific brain sites to address this issue.
A limitation of this work is that only male subjects were evaluated in these experiments. Sex-related differences in the ability of physical activity to modulate alcohol/drug self-administration have been reported [40,42]. Sex has also been shown to influence the ability of exercise to induce central BDNF transcriptional activity [36,58]. Thus, it will be important to determine whether the effects of exercise on BDNF expression and CIE-related escalated alcohol drinking observed in males in this series of studies are similar or differ in females.
Finally, the present set of studies did not control for the presence of activity wheels in the home-cage. The presence of activity wheels may be viewed as providing environmental enrichment, a manipulation that is known to influence alcohol consumption [59] as well as increasing BDNF levels in brain [60]. Some studies have used a locked wheel to address this issue. These studies have produced mixed results, with some reports showing that presenting a locked wheel to animals did not alter alcohol intake [35,38] while other studies showed an increase in alcohol intake [32,61]. Future studies will need to investigate whether blocking access to physical activity (locked wheel) alters BDNF levels and excessive alcohol consumption associated with dependence.
In summary, daily limited access to physical activity in the form of wheel-running was effective in significantly reducing escalation of alcohol drinking in chronic alcohol (CIE) exposed mice. This exercise regimen was shown to elevate BDNF levels in brain as well as reverse chronic alcohol-induced deficits in Bdnf mRNA expression in mPFC. Additionally, systemic administration of the TrkB receptor antagonist ANA-12 blocked the ability of exercise to attenuate CIE-related excessive alcohol drinking, thereby implicating increased BDF-TrkB receptor activity as a significant contributor of this effect. Together, these results support the emerging evidence that exercise may be an effective adjunct to treatment for individuals with alcohol use disorder.
4. Materials and Methods
4.1. Subjects
Adult male C57BL/6J mice (Jackson Laboratories, Bar Harbor, ME) were individually housed under a 12-hr light/dark cycle (lights on at 0800-hr). Mice were provided with food (Diet #2918; Harlan Teklad, Madison, WI) and water ad libitum in a temperature and humidity-controlled AAALAC-accredited facility at the Medical University of South Carolina. All procedures were approved by the Institutional Animal Care and Use Committee and conducted in accordance with the NIH Guide for the Care and Use of Laboratory Animals.
4.2. Exercise: Scheduled Wheel-Running
Mice were given home-cage access to an activity wheel (Bioserve cat #S3174 and S3175) 2-hr/day beginning 6 hours in the dark cycle. The amount of exercise (distance ran) was tracked for each mouse and calculated using bike computers (Enkeeo Model BKV-1527) attached to the running wheels. The No-Wheel groups were left undisturbed in their home-cages.
4.3. Home Cage Limited Access Alcohol Drinking
Mice were given daily (Mon-Fri) free access to alcohol (15% v/v ethanol) in their home-cage, with water as the alternative fluid for 2-hr, starting 3-hr into the dark cycle. Alcohol was prepared fresh daily, and bottles were weighed before and after the 2-hour access period.
4.4. Chronic Intermittent Ethanol (CIE) Exposure
Mice were exposed to chronic intermittent alcohol (ethanol) vapor or air in inhalation chambers, as previously described [28,62,63,64,65]. Prior to each day of alcohol vapor exposure, mice were administered 1.6 g/kg alcohol with 1 mmol/kg pyrazole in saline, to stabilize blood ethanol concentration (BEC). Air exposed mice received only pyrazole in saline. Alcohol and pyrazole were administered intraperitoneally (IP) in a volume of 0.02 mL/g body weight. Mice were placed in alcohol vapor or air inhalation chambers (60 × 36x60 cm Plexiglas boxes) 16-hr/day for 4 days during each weekly exposure cycle.
Using an air stone, alcohol (95% ethanol) was vaporized, mixed with fresh air, and delivered to the alcohol chamber. Air alone was delivered to the air chambers. The concentration of alcohol in the chambers was monitored daily, and air and alcohol flow rates were adjusted to maintain BECs in the 200-250 mg/dl range throughout the exposure cycles. BECs were determined as previously described [28,62,65]. Briefly, blood was collected from the retro-orbital sinus using heparinized capillary tubes and assayed using an Analox Instrument analyzer (Lunenburg, MA, USA).
4.5. Brain Tissue Collection
Mice were sacrificed by decapitation and brains rapidly removed. The medial prefrontal cortex (mPFC) and dentate gyrus region of the hippocampus (HPC) were dissected on ice using 1-mm microdissection punches, with the mouse brain atlas serving as a guide [51]. Tissue samples were placed into RNA Later (Thermo Fisher; cat no. AM7021). Total RNA was extracted from the brain tissue samples using ReliaPrep RNA extraction kit (Promega; cat no. Z6012) according to the manufacturer’s instructions. Total RNA was quantified on a Denovix DS-11FX spectrophotometer.
4.6. qRT-PCRAssay
Quantitative real-time polymerase chain reaction (qRT-PCR) assays were performed using similar procedures to those previously published by our group [11]. For Bdnf exon I, IV, and IX mRNA measurements, cDNA was created using QuantiTect Reverse Transcription Kit (Qiagen; cat no 205314) according to manufacturer’s instructions. Custom designed TaqMan qRT-PCR primers were used to specifically target and amplify Bdnf exon (I, IV, IX) mRNA, were designed using Integrated DNA Technologies (IDT) primer designer online software and manufactured by Life Technologies (BDNF I, Forward: GATGCCAGTTGCTTTGTCTTC; Reverse: CCACCACTTGGTGTGACTTAT; Probe: TCGCCAAGGTGGATGAGAGTTGAA; BDNF IV, Forward: GCCTAGATCAAATGGAGCTTCT; Reverse: GCCGATATGTACTCCTGTTCTG; Probe: ACCTCCGCCATGCAATTTCCACTA; BDNF IX, Forward: GTGACAGTATTAGCGAGTGGG; Reverse: GGGATTACACTTGGTCTCGTAG; Probe: CAGTTGGCCTTTGGATACCGGGA). Cyclophilin (Ppia) (Life Technologies) was used as the reference gene, for data normalization. For the reactions, 10 µl of sample cDNA and TaqMan Universal Master Mix II, with UNG (Applied Biosystems; cat no 4440042), were loaded in triplicate into a 384-well optical PCR plate and analyzed on a BioRad CFX384 Real Time PCR system. Cycling parameters: 50° C for 2-min, 95° C for 10-min, followed by 40 amplification cycles with melting at 95° C for 15-sec, and annealing/extending at 60° C for 60 sec. Fluorescence readings are obtained after each cycle. The 2^-DDCT method [66] was used to calculated fold change in expression of the target gene relative to the reference gene (Ppia) using the CTL group as the reference condition.
4.7. ELISA Assay
Brain-derived neurotrophic factor (BDNF) protein was assayed using BDNF Duo ELISA (R & D Systems) as per manufacturer’s instructions. Briefly, brain tissue was lysed in 1% BSA in PBS pH 7.4 via sonication. Standards (0-1500 pg/mL) and samples were added in duplicate to a 96-well plate, coated with BDNF-capture antibody, and blocked with 1% bovine serum albumin (BSA). Plates were incubated at room temperature for 2-hr and washed with Wash buffer (0.1% Tween in PBS). Detection antibody was added to a 96-well microplate and incubated at room temperature for 2-hr and then washed with Wash buffer. Streptavidin-HRP was added to each well and incubated for 20 minutes. 50-µL Stop solution (ThermoFisher) was added to the well, and the plate was washed with Wash buffer.
4.8. ANA-12 Preparation
The TrkB antagonist, ANA-12 (Selleckchem Cat no S7745), was resuspended in 1% (DMSO) in phosphate-buffered saline (PBS) (Sigma Aldrich) at 0.1 mg/mL for 1 mg/kg and 0.05 mg/mL for 0.5 mg/kg doses. ANA-12 (and vehicle) was administered intraperitoneally (IP) in a volume of 0.01 mL/g body weight.
4.9. Data Analysis
Data were analyzed using ANOVA followed by Newman-Keuls post-hoc comparisons. Alpha was set at 0.05 to assess significant effects. Analyses were done using Statistica (Tibco Software Inc.; Palo Alto, CA).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization: HCB, MGS, AG, MFL; Experimental Work and Data Collection: MGS, AG, MFL; Statistical Analyses: HCB, MGS, AG, MFL; Writing: HCB, MGS, AG, MFL; Writing - review and editing: HCB and MFL. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the U.S. Department of Veterans Affairs Office of Research and Development grants (BLR&D I01BX000813, IK6BX006299) to HCB, and the U.S. National Institutes of Health, National Institute on Alcohol Abuse and Alcoholism grants (P50 AA010761, U01 AA014095) to HCB and grant (U24 AA029968) to MFL. MGS and AG were supported by an Institutional Training grant (T32 AA007474).
Institutional Review Board Statement
All procedures were approved by the Medical University of South Carolina Institutional Animal Care and Use Committee and conducted in accordance with the NIH Guide for the Care and Use of Laboratory Animals.
Data Availability Statement
Data used to support the findings of this study are available from the corresponding author upon request.
Acknowledgements
The authors would like to acknowledge Ms. Laura Ralston for her technical assistance. Some of the research findings in this report are based on a dissertation submitted to the College of Graduate Studies, Medical University of South Carolina by MGS in partial fulfillment of the requirements for his Ph.D. degree.
Conflicts of Interest
The authors disclose no conflicts of interest. The funding sources were not involved in the study design, data collection, analysis, interpretation, or preparation of the manuscript.
References
- Kerr, W.C. , et al., Longitudinal assessment of drinking changes during the pandemic: The 2021 COVID-19 follow-up study to the 2019 to 2020 National Alcohol Survey. Alcohol Clin Exp Res, 2022. 46(6): p. 1050-1061. [CrossRef]
- Koob, G.F., P. Powell, and A. White, Addiction as a Coping Response: Hyperkatifeia, Deaths of Despair, and COVID-19. Am J Psychiatry, 2020. 177(11): p. 1031-1037.
- White, A.M. , et al., Alcohol-Related Deaths During the COVID-19 Pandemic. JAMA, 2022. 327(17): p. 1704-1706.
- Kranzler, H.R. and M. Soyka, Diagnosis and Pharmacotherapy of Alcohol Use Disorder: A Review. JAMA, 2018. 320(8): p. 815-824.
- Rehm, J. , et al., Global burden of disease and injury and economic cost attributable to alcohol use and alcohol-use disorders. Lancet, 2009. 373(9682): p. 2223-33.
- Sacks, J.J. , et al., 2010 National and State Costs of Excessive Alcohol Consumption. Am J Prev Med, 2015. 49(5): p. e73-e79.
- Koob, G.F. , Alcohol Use Disorder Treatment: Problems and Solutions. Annu Rev Pharmacol Toxicol, 2024. 64: p. 255-275.
- Litten, R.Z. , et al., Five Priority Areas for Improving Medications Development for Alcohol Use Disorder and Promoting Their Routine Use in Clinical Practice. Alcohol Clin Exp Res, 2020. 44(1): p. 23-35.
- Ray, L.A., R. Green, and E. Grodin, Neurobiological Models of Alcohol Use Disorder in Humans. Am J Psychiatry, 2021. 178(6): p. 483-484.
- Witkiewitz, K., R. Z. Litten, and L. Leggio, Advances in the science and treatment of alcohol use disorder. Sci Adv, 2019. 5(9): p. eaax4043.
- Melendez, R.I. , et al., Brain region-specific gene expression changes after chronic intermittent ethanol exposure and early withdrawal in C57BL/6J mice. Addict Biol, 2012. 17(2): p. 351-64. [CrossRef]
- Reichardt, L.F. , Neurotrophin-regulated signalling pathways. Philos Trans R Soc Lond B Biol Sci, 2006. 361(1473): p. 1545-64.
- Ron, D. and A. Berger, Targeting the intracellular signaling "STOP" and "GO" pathways for the treatment of alcohol use disorders. Psychopharmacology (Berl), 2018. 235(6): p. 1727-1743.
- Nubukpo, P. , et al., Determinants of Blood Brain-Derived Neurotrophic Factor Blood Levels in Patients with Alcohol Use Disorder. Alcohol Clin Exp Res, 2017. 41(7): p. 1280-1287.
- Heberlein, A. , et al., BDNF and GDNF serum levels in alcohol-dependent patients during withdrawal. Prog Neuropsychopharmacol Biol Psychiatry, 2010. 34(6): p. 1060-4.
- Xu, K. , et al., Nucleotide sequence variation within the human tyrosine kinase B neurotrophin receptor gene: association with antisocial alcohol dependence. Pharmacogenomics J, 2007. 7(6): p. 368-79.
- Wojnar, M. , et al., Association between Val66Met brain-derived neurotrophic factor (BDNF) gene polymorphism and post-treatment relapse in alcohol dependence. Alcohol Clin Exp Res, 2009. 33(4): p. 693-702.
- Matsushita, S. , et al., Association study of brain-derived neurotrophic factor gene polymorphism and alcoholism. Alcohol Clin Exp Res, 2004. 28(11): p. 1609-12.
- Warnault, V. , et al., The BDNF Valine 68 to Methionine Polymorphism Increases Compulsive Alcohol Drinking in Mice That Is Reversed by Tropomyosin Receptor Kinase B Activation. Biol Psychiatry, 2016. 79(6): p. 463-73.
- Stragier, E. , et al., Ethanol-induced epigenetic regulations at the Bdnf gene in C57BL/6J mice. Mol Psychiatry, 2015. 20(3): p. 405-12.
- Briones, T.L. and J. Woods, Chronic binge-like alcohol consumption in adolescence causes depression-like symptoms possibly mediated by the effects of BDNF on neurogenesis. Neuroscience, 2013. 254: p. 324-34.
- Jeanblanc, J. , et al., Endogenous BDNF in the dorsolateral striatum gates alcohol drinking. J Neurosci, 2009. 29(43): p. 13494-502.
- Jeanblanc, J. , et al., BDNF-mediated regulation of ethanol consumption requires the activation of the MAP kinase pathway and protein synthesis. Eur J Neurosci, 2013. 37(4): p. 607-12.
- Pandey, S.C. , et al., Central and medial amygdaloid brain-derived neurotrophic factor signaling plays a critical role in alcohol-drinking and anxiety-like behaviors. J Neurosci, 2006. 26(32): p. 8320-31.
- Darcq, E. , et al., The Neurotrophic Factor Receptor p75 in the Rat Dorsolateral Striatum Drives Excessive Alcohol Drinking. J Neurosci, 2016. 36(39): p. 10116-27.
- Tapocik, J.D. , et al., microRNA-206 in rat medial prefrontal cortex regulates BDNF expression and alcohol drinking. J Neurosci, 2014. 34(13): p. 4581-8.
- Solomon, M.G. , et al., Brain Regional and Temporal Changes in BDNF mRNA and microRNA-206 Expression in Mice Exposed to Repeated Cycles of Chronic Intermittent Ethanol and Forced Swim Stress. Neuroscience, 2019. 406: p. 617-625.
- Haun, H.L. , et al., Increasing Brain-Derived Neurotrophic Factor (BDNF) in medial prefrontal cortex selectively reduces excessive drinking in ethanol dependent mice. Neuropharmacology, 2018. 140: p. 35-42. [CrossRef]
- Palasz, E. , et al., BDNF as a Promising Therapeutic Agent in Parkinson's Disease. Int J Mol Sci, 2020. 21(3).
- Lynch, W.J. , et al., Exercise as a novel treatment for drug addiction: a neurobiological and stage-dependent hypothesis. Neurosci Biobehav Rev, 2013. 37(8): p. 1622-44.
- Smith, M.A. and W.J. Lynch, Exercise as a potential treatment for drug abuse: evidence from preclinical studies. Front Psychiatry, 2011. 2: p. 82.
- Centanni, S.W. , et al., The impact of intermittent exercise on mouse ethanol drinking and abstinence-associated affective behavior and physiology. Alcohol Clin Exp Res, 2022. 46(1): p. 114-128.
- Darlington, T.M. , et al., Mesolimbic transcriptional response to hedonic substitution of voluntary exercise and voluntary ethanol consumption. Behav Brain Res, 2014. 259: p. 313-20.
- Darlington, T.M. , et al., Voluntary wheel running reduces voluntary consumption of ethanol in mice: identification of candidate genes through striatal gene expression profiling. Genes Brain Behav, 2016. 15(5): p. 474-90.
- Ehringer, M.A., N. R. Hoft, and M. Zunhammer, Reduced alcohol consumption in mice with access to a running wheel. Alcohol, 2009. 43(6): p. 443-52.
- Gallego, X. , et al., Voluntary exercise decreases ethanol preference and consumption in C57BL/6 adolescent mice: sex differences and hippocampal BDNF expression. Physiol Behav, 2015. 138: p. 28-36.
- Grigsby, K. , et al., Voluntary wheel-running reduces harmful drinking in a genetic risk model for drinking to intoxication. Alcohol, 2025. 128: p. 35-42.
- Ozburn, A.R., R. A. Harris, and Y.A. Blednov, Wheel running, voluntary ethanol consumption, and hedonic substitution. Alcohol, 2008. 42(5): p. 417-24.
- Werme, M. , et al., Running increases ethanol preference. Behav Brain Res, 2002. 133(2): p. 301-8.
- Booher, W.C. J. Reyes Martinez, and M.A. Ehringer, Behavioral and neuronal interactions between exercise and alcohol: Sex and genetic differences. Genes Brain Behav, 2020. 19(3): p. e12632. [CrossRef]
- Leasure, J.L. , et al., Exercise and Alcohol Consumption: What We Know, What We Need to Know, and Why it is Important. Front Psychiatry, 2015. 6: p. 156.
- Lynch, W.J. , et al., Exercise as a Sex-Specific Treatment for Substance Use Disorder. Curr Addict Rep, 2017. 4(4): p. 467-481.
- Manthou, E. , et al., Role of exercise in the treatment of alcohol use disorders. Biomed Rep, 2016. 4(5): p. 535-545.
- Terry-McElrath, Y.M. and P.M. O'Malley, Substance use and exercise participation among young adults: parallel trajectories in a national cohort-sequential study. Addiction, 2011. 106(10): p. 1855-65; discussion 1866-7.
- Brown, R.A. , et al., A preliminary, randomized trial of aerobic exercise for alcohol dependence. J Subst Abuse Treat, 2014. 47(1): p. 1-9.
- Penedo, F.J. and J.R. Dahn, Exercise and well-being: a review of mental and physical health benefits associated with physical activity. Curr Opin Psychiatry, 2005. 18(2): p. 189-93.
- Greenwood, B.N. , et al., Exercise-induced stress resistance is independent of exercise controllability and the medial prefrontal cortex. Eur J Neurosci, 2013. 37(3): p. 469-78.
- Linke, S.E. , et al., The Go-VAR (Veterans Active Recovery): An Adjunctive, Exercise-Based Intervention for Veterans Recovering from Substance Use Disorders. J Psychoactive Drugs, 2019. 51(1): p. 68-77.
- Cotman, C.W., N. C. Berchtold, and L.A. Christie, Exercise builds brain health: key roles of growth factor cascades and inflammation. Trends Neurosci, 2007. 30(9): p. 464-72.
- Phillips, C. , Physical Activity Modulates Common Neuroplasticity Substrates in Major Depressive and Bipolar Disorder. Neural Plast, 2017. 2017: p. 7014146.
- Franklin, K.B.J. and G. Paxinos, The Mouse Brain in Stereotaxic Coordinates, Third Edition. 3rd ed. 2008, San Diego: Academic Press.
- Berchtold, N.C. , et al., Exercise primes a molecular memory for brain-derived neurotrophic factor protein induction in the rat hippocampus. Neuroscience, 2005. 133(3): p. 853-61.
- Liu, P.Z. and R. Nusslock, Exercise-Mediated Neurogenesis in the Hippocampus via BDNF. Front Neurosci, 2018. 12: p. 52.
- Sleiman, S.F. , et al., Exercise promotes the expression of brain derived neurotrophic factor (BDNF) through the action of the ketone body beta-hydroxybutyrate. Elife, 2016. 5. [CrossRef]
- Aid, T. , et al., Mouse and rat BDNF gene structure and expression revisited. J Neurosci Res, 2007. 85(3): p. 525-35.
- You, H. and B. Lu, Diverse Functions of Multiple Bdnf Transcripts Driven by Distinct Bdnf Promoters. Biomolecules, 2023. 13(4).
- Somkuwar, S.S. , et al., Wheel running reduces ethanol seeking by increasing neuronal activation and reducing oligodendroglial/neuroinflammatory factors in the medial prefrontal cortex. Brain Behav Immun, 2016. 58: p. 357-368.
- Islas-Preciado, D. , et al., Sex and BDNF Val66Met polymorphism matter for exercise-induced increase in neurogenesis and cognition in middle-aged mice. Horm Behav, 2023. 148: p. 105297.
- Camarini, R. , et al., Environmental enrichment and complementary clinical interventions as therapeutic approaches for alcohol use disorder in animal models and humans. Int Rev Neurobiol, 2024. 178: p. 323-354.
- Costa, G.A. , et al., Environmental Enrichment Increased Bdnf Transcripts in the Prefrontal Cortex: Implications for an Epigenetically Controlled Mechanism. Neuroscience, 2023. 526: p. 277-289.
- Piza-Palma, C. , et al., Oral self-administration of EtOH: sex-dependent modulation by running wheel access in C57BL/6J mice. Alcohol Clin Exp Res, 2014. 38(9): p. 2387-95.
- Becker, H.C. and M.F. Lopez, Increased ethanol drinking after repeated chronic ethanol exposure and withdrawal experience in C57BL/6 mice. Alcoholism: Clinical & Experimental Research, 2004. 28(12): p. 1829-1838.
- Becker, H.C. and M.F. Lopez, Animal Models of Excessive Alcohol Consumption in Rodents. Curr Top Behav Neurosci, 2024.
- Griffin, W.C. , 3rd, M.F. Lopez, and H.C. Becker, Intensity and duration of chronic ethanol exposure is critical for subsequent escalation of voluntary ethanol drinking in mice. Alcoholism: Clinical and Experimental Research, 2009. 33(11): p. 1893-1900.
- Griffin, W.C. , 3rd, et al., Repeated cycles of chronic intermittent ethanol exposure in mice increases voluntary ethanol drinking and ethanol concentrations in the nucleus accumbens. Psychopharmacology, 2009. 201(4): p. 569-580.
- Livak, K.J. and T.D. Schmittgen, Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods, 2001. 25(4): p. 402-8. [CrossRef]
Figure 1.
Exercise (2-hr/day) over 6 weeks increased BDNF protein expression in brain. Wheel-running remained relately stable over the last 4 weeks of exercise (A); daily scheduled limited access to running-wheels (2-hr/day) in the home-cage increased BDNF protein expression in mPFC 24-hr after the 6-week exercise regimen, with an increased trend at 1-wk (B) while elevetaed BDNF protein levels in DG was evident at 24-hr and peristed for at least 1 week following the exercise (C). Values are mean ± s.e.m. N= 8-10/group.
Figure 1.
Exercise (2-hr/day) over 6 weeks increased BDNF protein expression in brain. Wheel-running remained relately stable over the last 4 weeks of exercise (A); daily scheduled limited access to running-wheels (2-hr/day) in the home-cage increased BDNF protein expression in mPFC 24-hr after the 6-week exercise regimen, with an increased trend at 1-wk (B) while elevetaed BDNF protein levels in DG was evident at 24-hr and peristed for at least 1 week following the exercise (C). Values are mean ± s.e.m. N= 8-10/group.
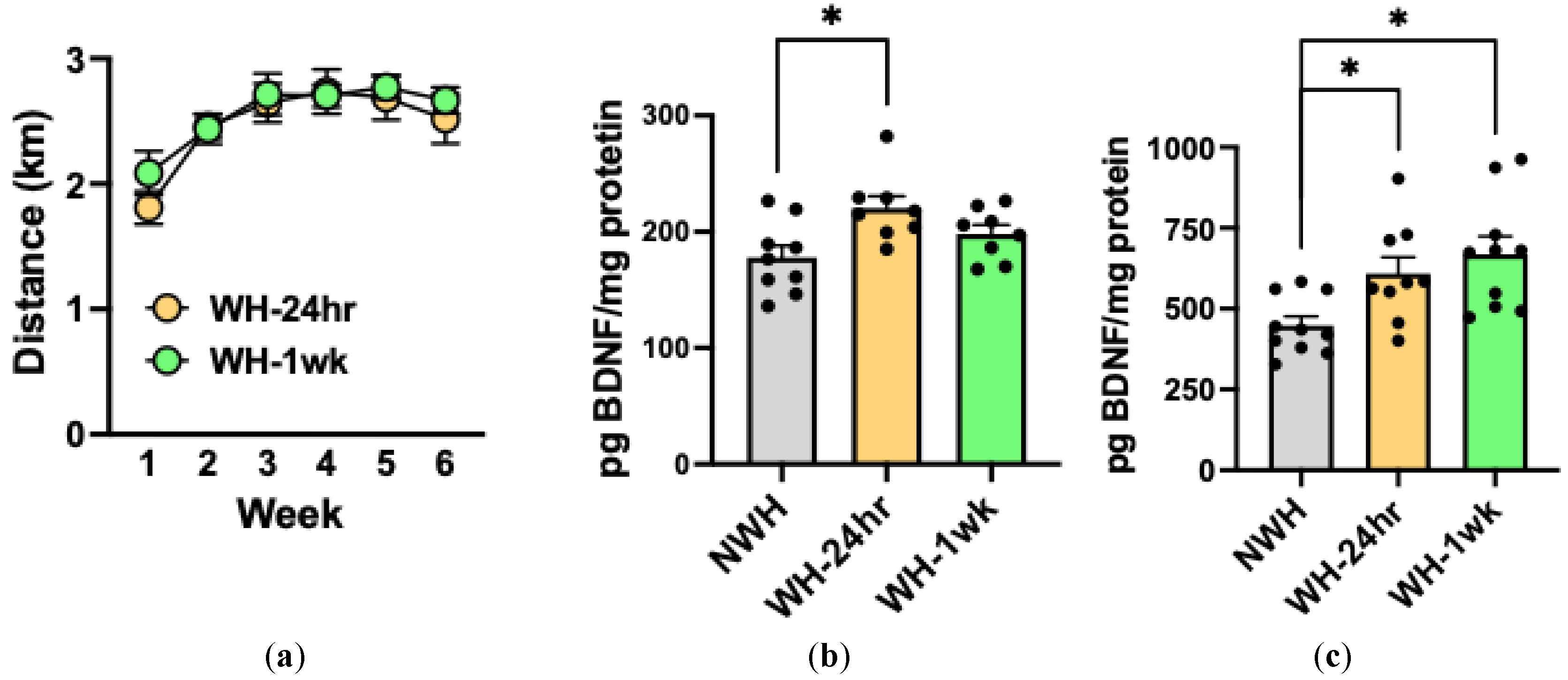
Figure 2.
Wheel-running is not altered by alcohol consumption (2-hr/day) in the home-cage. Values are mean ± s.e.m. N= 12/group.
Figure 2.
Wheel-running is not altered by alcohol consumption (2-hr/day) in the home-cage. Values are mean ± s.e.m. N= 12/group.
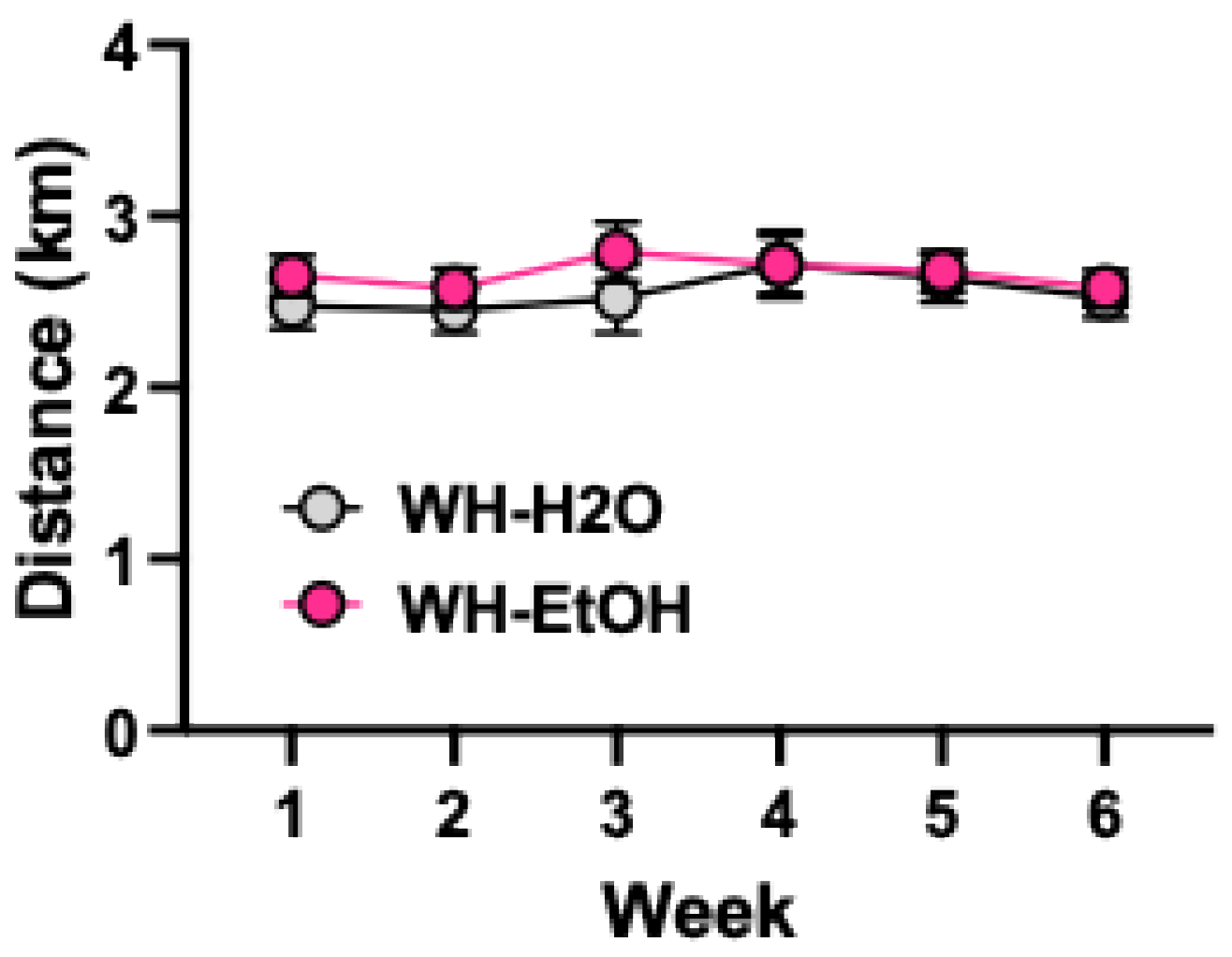
Figure 3.
Access to wheel-running (2-hr/day) does not alter alcohol consumption (2-hr/day) in the home-cage. Values are mean ± s.e.m. N= 12/group.
Figure 3.
Access to wheel-running (2-hr/day) does not alter alcohol consumption (2-hr/day) in the home-cage. Values are mean ± s.e.m. N= 12/group.

Figure 4.
Wheel-running (2-hr/day) for 6 weeks increases Bdnf mRNA expression (Exons I, IV, IX) in mPFC (A) and Dentate Gyrus (B). Values are means ± s.e.m. expressed as change from NWH-H2O condition. N= 6-11/group. * p< 0.05; ** p< 0.01; *** p< 0.001.
Figure 4.
Wheel-running (2-hr/day) for 6 weeks increases Bdnf mRNA expression (Exons I, IV, IX) in mPFC (A) and Dentate Gyrus (B). Values are means ± s.e.m. expressed as change from NWH-H2O condition. N= 6-11/group. * p< 0.05; ** p< 0.01; *** p< 0.001.
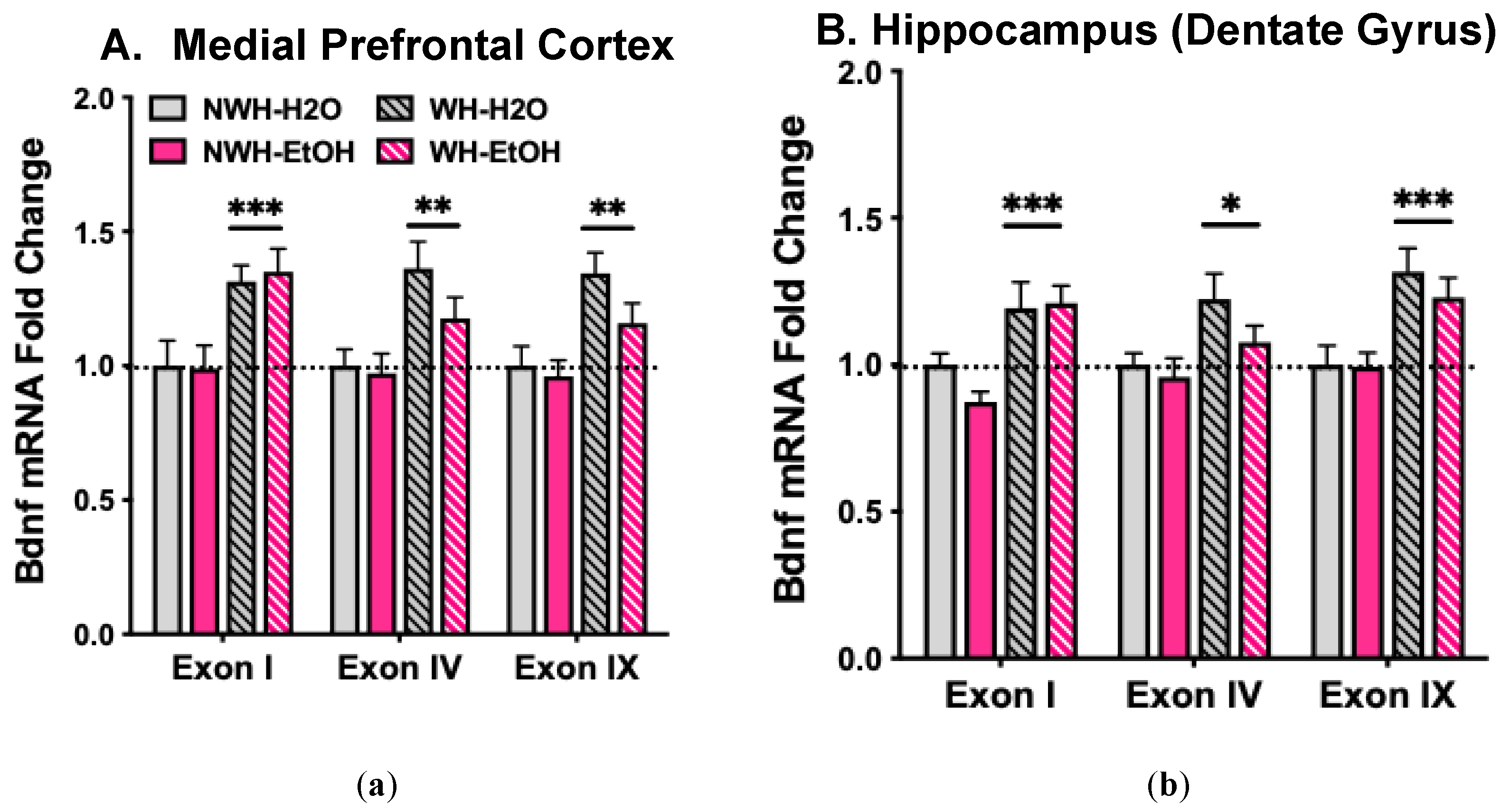
Figure 5.
Study design to examine effects of wheel-running on alcohol consumption in the CIE Drinking model and Bdnf mRNA expression in mPFC.
Figure 5.
Study design to examine effects of wheel-running on alcohol consumption in the CIE Drinking model and Bdnf mRNA expression in mPFC.

Figure 6.
Exercise (wheel-running) attenuates CIE-induced escalation of alcohol drinking. Weekly alcohol intake (g/kg) during last week of baseline and Test cycles 1-4 (A) and percent change in alcohol intake from baseline levels of intake (B). Values are mean ± s.e.m. (N= 20/group). * significantly differs from respective baseline and NWH-CTL group (ps< 0.05); # significantly differs from NWH-CIE group (ps< 0.05).
Figure 6.
Exercise (wheel-running) attenuates CIE-induced escalation of alcohol drinking. Weekly alcohol intake (g/kg) during last week of baseline and Test cycles 1-4 (A) and percent change in alcohol intake from baseline levels of intake (B). Values are mean ± s.e.m. (N= 20/group). * significantly differs from respective baseline and NWH-CTL group (ps< 0.05); # significantly differs from NWH-CIE group (ps< 0.05).
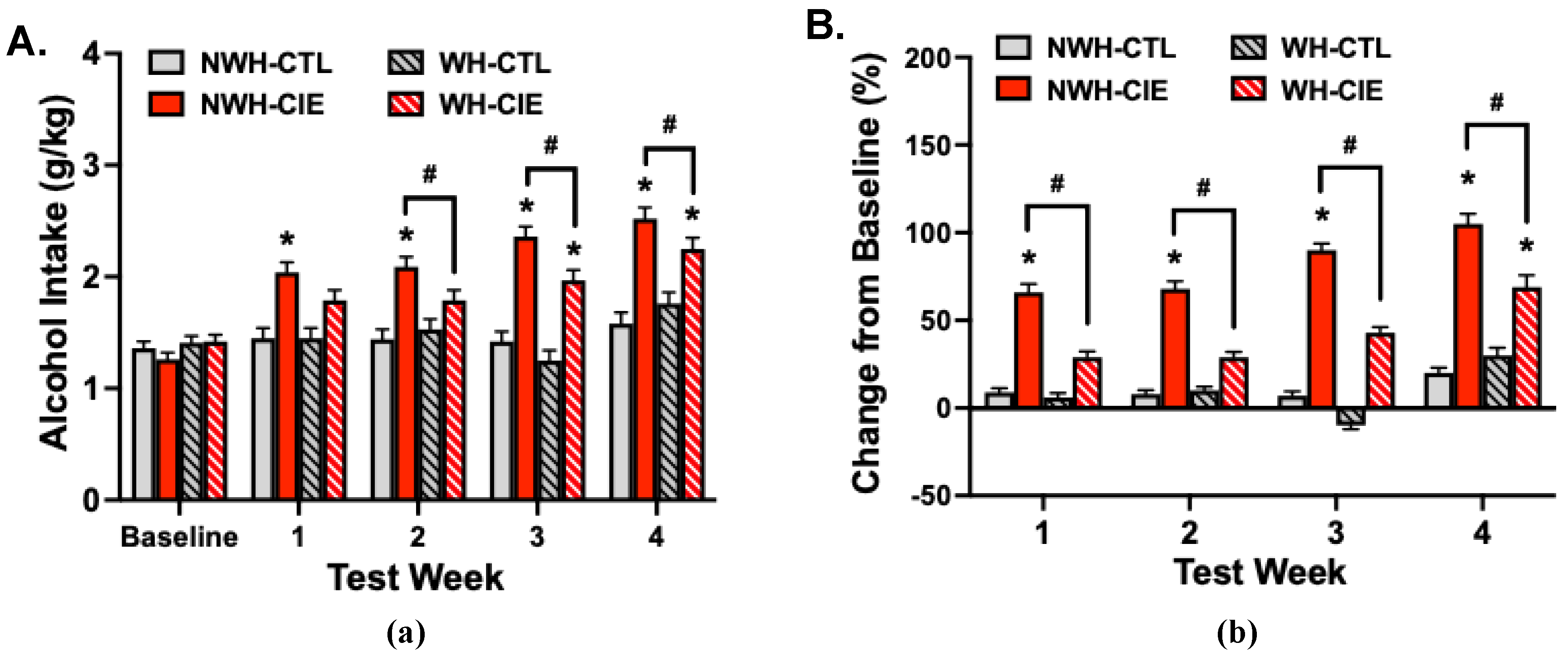
Figure 7.
Exercise mitigates chronic alcohol (CIE) induced deficits in Bdnf mRNA expression in mPFC. Values are mean ± s.e.m. N= 7-11/group. * significantly differs from NWH-CTL group (ps< 0.05); ^ significantly differs from NWH-CTL and WH-CIE groups (ps< 0.05).
Figure 7.
Exercise mitigates chronic alcohol (CIE) induced deficits in Bdnf mRNA expression in mPFC. Values are mean ± s.e.m. N= 7-11/group. * significantly differs from NWH-CTL group (ps< 0.05); ^ significantly differs from NWH-CTL and WH-CIE groups (ps< 0.05).
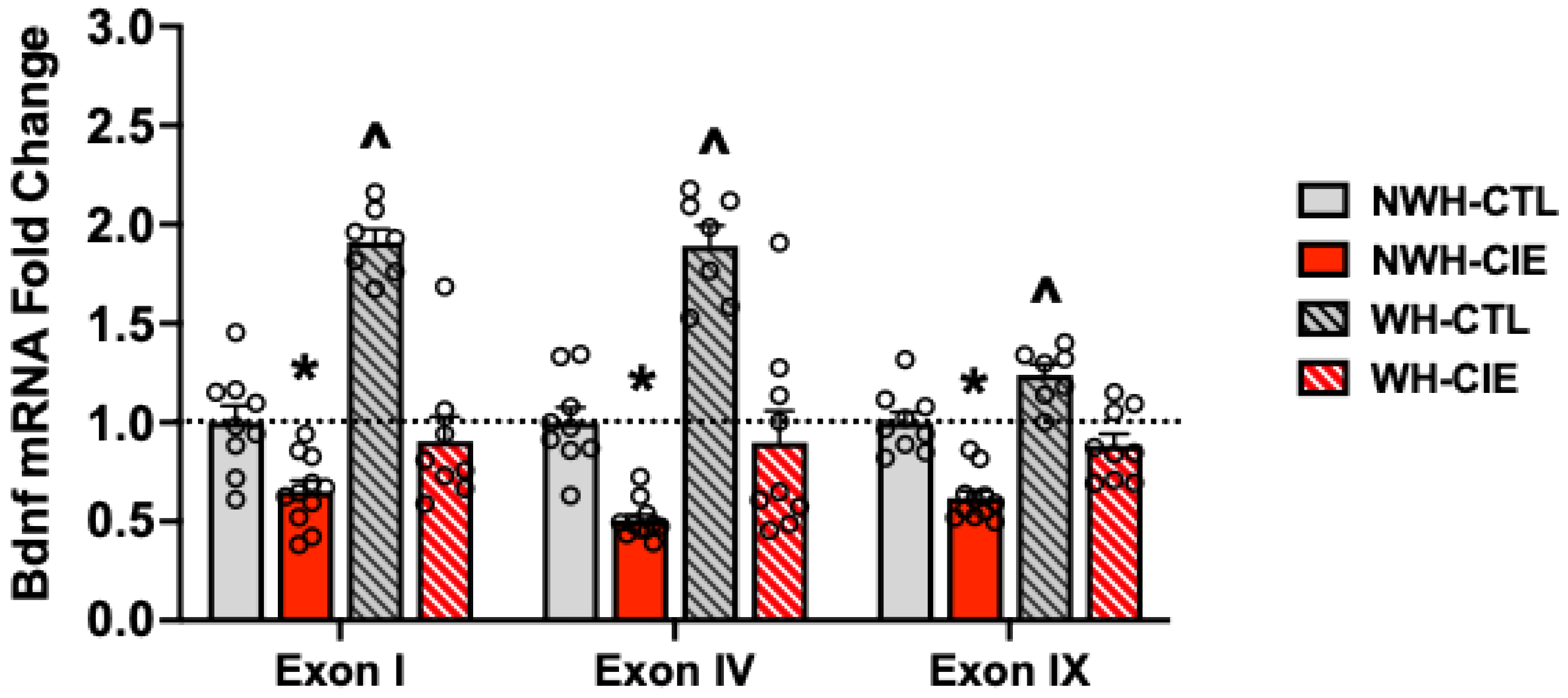
Figure 8.
The TrkB receptor antagonist ANA-12 blocks the ability of exercise to attenuate CIE-induced escalated alcohol drinking. Values are mean ± s.e.m. N= 9-11/group. * significantly differs from NWH-CTL group (ps< 0.05); # significantly differs from WH-CIE group (ps< 0.05).
Figure 8.
The TrkB receptor antagonist ANA-12 blocks the ability of exercise to attenuate CIE-induced escalated alcohol drinking. Values are mean ± s.e.m. N= 9-11/group. * significantly differs from NWH-CTL group (ps< 0.05); # significantly differs from WH-CIE group (ps< 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



