Submitted:
29 September 2025
Posted:
29 September 2025
You are already at the latest version
Abstract
Objectives: Postprandial variations in plasma triacylglycerol (TAG) responses to veg-etable oils are well established, but their origins remain unclear. This study examined the variability of postprandial plasma long-chain omega-3 fatty acids (LC omega-3) in response to commonly used supplements and foods and considers the biological im-plications of this variability.
Methods: A literature review was conducted to identify postprandial studies that re-ported variability in plasma LC omega-3 responses to supplementation. Studies were included if variability could be expressed as the coefficient of variation (CV) for the area under the curve (AUC).
Results: Twenty-one studies encompassing 36 different treatments were identified. Supplements included LC omega-3 in the form of TAG, monoacylglycerols (MAG), free fatty acids (FFA), ethyl esters (EE), EE with emulsification agents, and whole foods. Variability was consistently observed across all forms; 65% of treatments showed a CV >50% for the AUC. Gastrointestinal (GI) symptoms were reported in some studies, suggesting possible malabsorption.
Conclusions: Substantial inter-individual variability existed in postprandial LC omega-3 responses, independent of the chemical form of supplementation. This variability likely reflects differences in absorption, enterocyte metabolism, and possibly malabsorption. Postprandial variability may therefore contribute significantly to observed differences in tissue LC omega-3 status.
Keywords:
omega-3 fatty acids
; long-chain omega-3 fatty acids
; postprandial metabolism
; inter-individual variability
; triacylglycerol
; Monacylglycerol
; free fatty acids
; ethyl esters
; lipid absorption
; supplementation
; malabsorption
; enterocyte metabolism
1. Introduction
After meals, the body produces triacyglycerol (TAG)-rich lipoproteins (chylomicrons) and their remnants. Elevated postprandial triglyceride levels have been linked to a higher risk of cardiovascular events, such as heart attacks and strokes. Studies indicate that chylomicron and remnant levels may be more predictive of such events than fasting TAG measurements [1,2,3].
Variability in postprandial TAG responses between individuals is well known and is likely to drive advances in the use of personalised nutrition. Recent studies have been reported postprandial variability for meals containing oils such as palm oil or high-oleic sunflower oil. Newman et al. [4] studied the postprandial TAG response to a mixed ingredients meal containing palm oil in 340 subjects and reported significant variability in responses over 6-h, including that approx. 2% of subjects showed minimal plasma TAG appearance by 6-h. In another study by Berry et al. [5], more than 1000 people consumed 8 meals differing in macronutrient composition with the fats/oils being mainly derived from high-oleic sunflower oil. Large inter-individual postprandial variations (over 6-h) were reported in TAG, glucose and insulin responses following consumption of identical meals (the coefficient of variation (CV) in the postprandial responses for TAG was 103%, and 68% for glucose, and 59% for insulin). Amongst the statistically derived determinants of variability for postprandial TAG, serum lipid markers had the greatest influence (>20%), serum glycemic markers (>10%), other serum markers (>8%), anthropometry (8%), microbiome (7%), and gender (6%). Meal macronutrients (3.6%) and genetic variants (0.8%) had minimal impacts on predictions for postprandial lipaemia.
There have been many LC omega-3 supplementation studies reported in the literature over the past 30+ years [6,7], but few report the data for individual subjects or refer to variability. Several medium to long-term studies reported the individual data or refer to the variability in the blood levels of LC omega-3 following supplementation [8,9,10,11,12,13]. We hypothesised that one contributor to the variability in plasma and erythrocyte (RBC) LC omega-3 levels following supplementation was variability which occurred during digestion and absorption of the LC omega-3. Therefore, we investigated the variability in the plasma LC omega-3 levels in the postprandial period. We found consistent and substantial variability in the postprandial response to LC omega-3 supplementation and hypothesised that postprandial variability could be a significant contributor to variability in plasma and RBC LC omega-3 levels.
2. Materials and Methods
Literature search strategy and selection criteria of articles
A systematic search was conducted in two databases—PubMed and Embase—up to June 2025. We used the following key words treated as title/abstract for the literature search: (“plasma docosahexaenoic acid” OR “plasma DHA” OR “plasma eicosapentaenoic acid” OR “plasma EPA” OR “plasma docosapentaenoic acid” OR “plasma DPA” OR “polyunsaturated fatty acid” OR “plasma LC omega-3 fatty acid” OR “plasma n-3 fatty acid” OR “fish oil” OR “krill” OR “algal oil” OR “wax esters” OR “ethyl esters” OR “FFA” OR “MAG” OR “n-3 FA enriched foods” OR “LC omega-3 enriched foods”) AND (“postprandial” OR “acute”). Our search was restricted to studies in humans and studies published in English. The references of retrieved relevant articles were reviewed to identify potential publications. AJS and DL independently conducted the literature search, identified potential studies, and extracted detailed information from each included article. Inclusion criteria were controlled trials with postprandial design; the exposure of interest was any type of dietary long chain LC omega-3 fatty acid, the endpoint of interest was plasma LC omega-3 concentration expressed as area under the curve. We excluded studies in animals, controlled trials without mean±SD/SEM, non-original research (reviews, editorials, or commentaries), abstracts, unpublished studies, and duplicated studies.
3. Results
We identified 30 original LC omega-3 postprandial studies which reported area under the curve data (AUC). In this report, we only considered 21 of these since they reported the AUC curve data as mean with either SD or SEM. This allowed the expression of the variability by converting the mean responses to a coefficient of variation (CV, SD/mean * 100). Studies reporting 25th and 75th percentile or other ranges were not included as it was not possible to derive a CV for the responses. The lipid type of LC omega-3 supplements varied considerably and included fish oils (FO) rich in TAG, lipid digestion products containing LC omega-3 (MAG and FFA), ethyl esters (EE) of EPA + DHA, krill oil typically rich in phospholipids (PL), algal oil (rich in polar lipids), EE of EPA + DHA mixed with novel emulsification agents and foods/meals rich in TAG containing LC omega-3.
Only two studies reported individual responses. Kohler et al. [12] compared the effects of a single dose of fish oil or krill oil (1700 mg EPA+DHA) on LC omega-3 levels in plasma PL and TAG fractions over a 72-h period in 15 adult subjects. There were large inter-individual variations in the incremental area under the curve (iAUC) to both the fish oil (rich in TAG) and the krill oil (rich in PL). The individual responses for fish oil and krill oil were not the same for the plasma PL and plasma TAG. Furthermore, in the plasma TAG fraction, for krill oil 2/15 subjects showed no response, while 1/15 (a different subject) showed no response to the fish oil; also 1/15 subjects for the fish oil group showed no response in the PL fraction. The reported results were converted into mean CV for the iAUC72h responses (SD/Mean *100). For the fish oil, the CVs for EPA+DHA in plasma PL and plasma TAG were 38% and 76%, respectively. For the krill oil, the CVs for EPA+DHA in plasma PL and plasma TAG were 37% and 72%, respectively. These results indicated that postprandial variability was evident for two different dietary lipid supplements, namely LC omega-3-TAG in the case of the fish oil and LC omega-3-PL (+FFA and TAG) in the case of krill oil. The other study reporting individual data was by Stonehouse et al. 2022 [14], in which the effects on plasma DHA levels of a single meal of soup or rice crackers with encapsulated algal DHA or capsules of DHA (440mg DHA in each treatment) were studied over a 24-h period in 27 male subjects. There was substantial variability between individuals for each treatment. The CVs for iAUC for plasma DHA were 68% for the soup meal, 76% for the rice crackers meal and 87% for the capsules of algal oil TAG.
Results of the variability of the 21 postprandial studies are summarised below and presented in detail in Table A1.
- TAG oil studies (fish oil, re-esterified fish oil, blended fish oil, sardine/anchovy oil, algal oil, n=6 studies). The variability, expressed as CV for the AUC or iAUC, for EPA, DHA or EPA+DHA in plasma TAG or PL ranged from 38% to 102%, with 7 of 10 data points exceeding 50%. These studies involved from 7 to 27 subjects over periods from 8h to 72h and used doses which ranged from 415mg up to 1700mg of EPA+DHA [12,14,15,16,17,29].
- PL-rich oil studies (krill oil, PL-enhanced fish oil, polar-rich algal oil, n=6 studies). The variability, expressed as CV for the AUC or iAUC, for EPA, DHA or EPA+DHA in plasma TAG or PL ranged from 28% to 76%, with 5 of 9 data points exceeding a CV of 50%. These studies involved from 10 to 24 subjects over periods from 10h to 72h and used doses which ranged from 206mg up to 1700mg of EPA+DHA [12,15,18,19].
- FFA studies (Epanova, OM3-CA, unspecified, n=3 studies). The variability, expressed as CV for the AUC or iAUC, for EPA, DHA or EPA+DHA in plasma ranged from 34% to 53%, with 3 of 5 data points equal to or exceeding a CV of 50%. These studies involved from 14 to 26 subjects over 24h and used doses which ranged from 3264mg up to 4000mg of EPA+DHA [20,21,22].
- Monoacylglycerol (MAG) studies (2-MAG, 1(3)-MAG, MAG unspecified, n=3 studies). The variability, expressed as CV for the AUC, for EPA, DHA or EPA+DHA in plasma ranged from 19% to 93%, with 2 of 4 data points equal to or exceeding a CV of 50%. These studies involved from 7 to 24 subjects over 24h and used doses which ranged from 1247mg up to 3000mg of EPA+DHA [17,21,23].
- Ethyl ester of LC omega-3 studies (Omacor, KD Pharma or unspecified sources, n=9 studies). The variability, expressed as CV for the AUC or iAUC, for EPA, DHA or EPA+DHA in plasma or plasma PL ranged from 32% to 154%, with 9 of 12 data points exceeding a CV of 50%. These studies involved from 10 to 40 subjects over 24h to 72h and used doses which ranged from 680mg up to 3360mg of EPA+DHA [15,20,21,23,24,25,26,27,28].
- Wax ester study (Calanus finmarchicus oil, n=1). The variability, expressed as CV for the iAUC, for EPA+DHA in plasma was 42%. This study involved 18 subjects over 72h and used a dose of 416mg of EPA+DHA. Wax esters are considered mostly undigestible due to the poor efficacy of the carboxylester lipase and the poor solubility of the wax esters. However, this study showed that the bioavailability of the wax esters was similar to that of the comparative EE group [27].
- Ethyl esters with enhanced emulsification properties (n=4 studies). The variability, expressed as CV for the AUC or iAUC, for EPA, DHA or EPA+DHA in plasma or whole blood ranged from 31% to 137%, with 4 of 6 data points exceeding a CV of 50%. These studies involved from 12 to 40 subjects over 24h to 72h and used doses which ranged from 374mg up to 1680mg of EPA+DHA. The data revealed that the enhanced emulsification properties significantly improved the bioavailability of the EE compared with standard EE, as judged by the AUC, but variability was still evident [24,25,26,29].
- Whole food studies (TAG) (herring, oils, foods with fish oil, novel foods, n=4 studies). The variability, expressed as CV for the iAUC for EPA, DHA or EPA+DHA in ranged from 21 to 138%, with 7 of 10 data points exceeding a CV of 50%. The studies involved from 17 to 27 subjects over 6h to 24h and used doses which ranged from 415mg up to 3200mg of EPA+DHA. These studies demonstrated that the food matrix did not alter the highly variable postprandial plasma LC omega-3 responses to whole food containing TAG-rich in LC omega-3 [14,30,31,32].
- In summary, the variability in the plasma or blood LC omega-3 levels of the 21 postprandial studies revealed that there was significant variability in the data independent of the type of lipid supplement. More than 64% of the treatments had a CV in excess of 50% for the AUC or iAUC for plasma EPA, plasma DHA, whole blood EPA+DHA or plasma EPA+DHA (Table 1).
4. Discussion
We showed that in all studies there was significant variability in the postprandial accretion of LC omega-3 into plasma (and whole blood) over periods ranging from 6h to 72h. Furthermore, the variability was independent of the lipid type (structure) of the LC omega-3 supplement, or for LC omega-3 present in whole foods. There have been few investigations into the causes of the variability of LC omega-3 responses at the postprandial level, however there are some indications from the literature which suggest that some of the variability could result from some of the LC omega-3 supplements escaping digestion in the duodenum and being excreted into the colon.
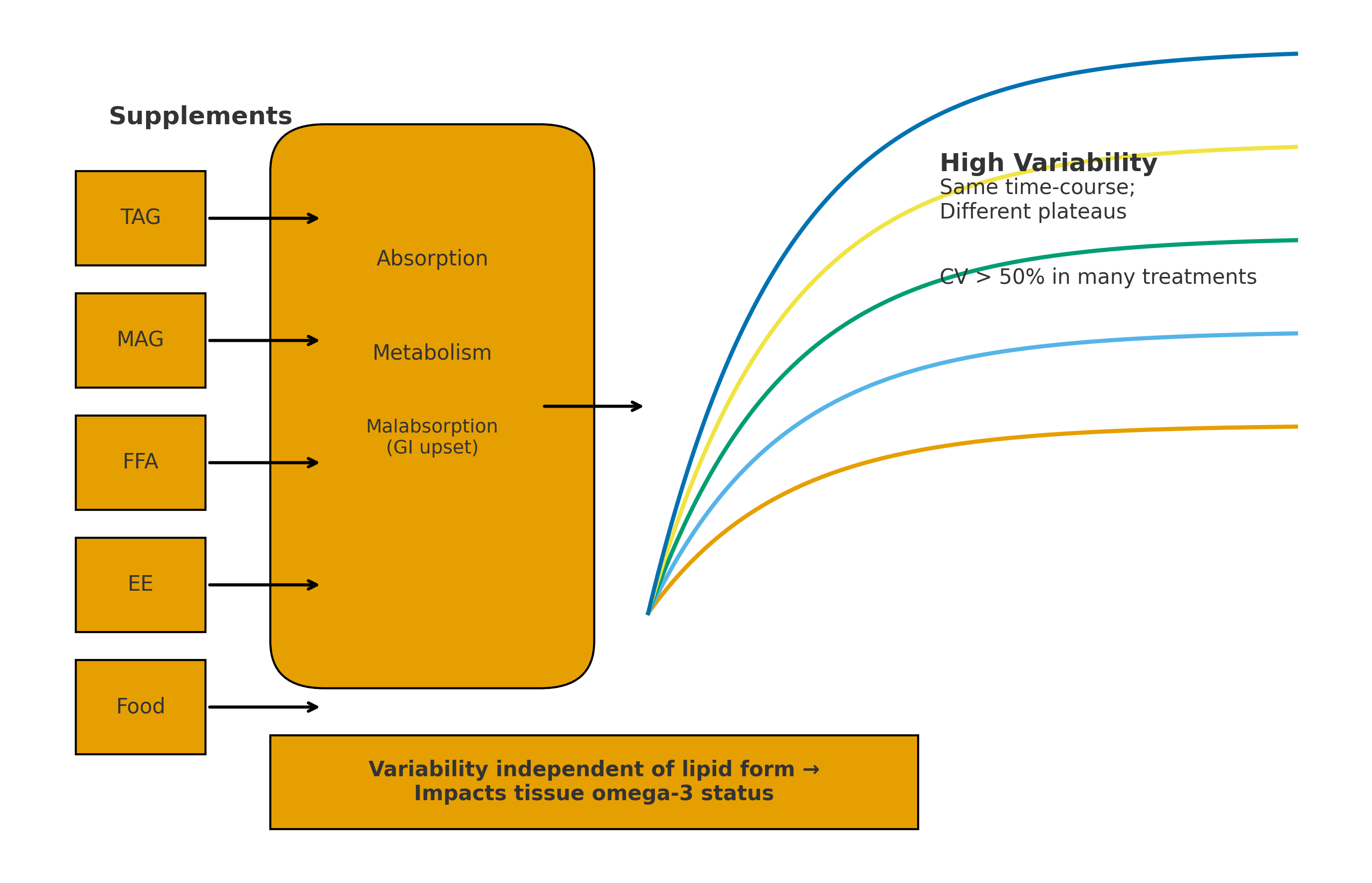
To examine which stages of digestion, absorption and re-assembly of fatty acids into TAG and PL in the enterocyte, the studies can be looked at from the stages of digestion and absorption, as outlined in Figure 1.
- Lipases and carboxylesterases. LC omega-3 lipid supplements requiring enzymatic digestion (TAG, PL, ethyl esters and wax esters) all showed substantial variability (72% of treatments had CV>50%). This implies/indicates that within these studies there were individuals with either high, medium or low plasma accretion of LC omega-3. This suggests variability in the lipid digesting enzymes and/or the micelle formation which facilitate the enzymatic processes. To partially address the issue of micelle formation, it was shown that the ethyl ester preparations with enhanced micelle formation properties led to significantly higher plasma LC omega-3 accretion than ethyl esters, however despite this, the enhanced preparations still showed high CV values (31-115%). Thus, factors other than micelle formation are likely playing a role in the observed variability. In the case of those individuals with putatively low plasma LC omega-3 accretion, some of the undigested LC omega-3 oils will lead to malabsorption of the oils meaning the undigested lipids will pass into the large bowel and be subject to faecal excretion.
- Lipid digestion products (FFA and MAG). These lipid products also showed considerable variability in their accretion into plasma (67% of treatments had CV>50%). Again, there will be individuals with either high, medium or low plasma accretion of LC omega-3 and in the latter case the malabsorption will mean some of these unabsorbed products (FFA & MAG) will pass into the large bowel and be excreted.
We examined whether the variability was related to the dose of the LC omega-3 supplement, as shown in Figure 2a. It was clear that dose was not a significant factor in the variability (r2 = 0.002, p>0.00). Another possible source of variability could be the analytical method and/or laboratory conditions. Since all studies examined the postprandial availability of at least two lipid types, we plotted the pairs of treatments conducted in each laboratory as shown in Figure 2b. It was evident from this that there was a weak relationship between the pairs of studies, with the regression explaining on 28% of the variance, and with only 10/27 of the treatment pairs having a CV within 10% of each other. This suggests that neither the analytical method nor the laboratory conditions were likely to major sources of the postprandial variability reported here. We then compared the variability between supplements requiring enzymatic digestion (TAG + PL) with lipid digestion products (FFA + MAG). While these studies were not conducted in the same individuals, there was no significant difference between the pre- and post-lipid digestion lipid supplements (TAG+PL, CV = 58±18% (n=19 studies, mean±SD) versus FFA+DAG, CV= 49±20% (n=9 studies) p>0.05). This matter requires a detailed investigation comparing pre- and post-lipase supplements in the same subjects,
Since variability was found with all the different lipid formulations, it is logical that some of the variability was in the post-digestion and emulsification steps and the absorption of digested lipids into the enterocytes via fatty acid transporters, and/or assembly of the absorbed lipids in the endoplasmic reticulum into TAG and PL (including relevant enzymes such as acylCoA synthetase; 1-acyl-glycerol-3-phosphate acyltransferase; MAG acyltransferase; DAG acyltransferase; acetylCoA-cholesterol acyltransferase), and then export of these lipids in chylomicrons into the lymph [33,34]. Wang et al. (34) note that intestinal fatty acid absorption involves multiple genes, however it is not known which steps play a critical role in absorption of intestinal fatty acids; furthermore, they report that little is known about the variability in the multi-step processes of absorption of lipid digestion products or the variability in the enzymatic activities involved in the assembly of chylomicron lipids, either with specific reference to LC omega-3 or other common fatty acids.
Malabsorption of LC omega-3 oils is referred to as a side effect of LC omega-3 oil consumption in the literature [35,36] with effects reported including burping, fishy taste and gastrointestinal (GI) upsets such as looser stools, flatulence, malodorous stools and diarrhea. A review of five trials which used fish oils in the prevention of restenosis following coronary angioplasty reported that there was a strong relationship between the dose of fish oil and GI side effects (trend p<0.0001) [35]. There has been inconsistent reporting of adverse events in either postprandial or longer-term LC omega-3 studies, however GI-related adverse effects were reported in three of the postprandial studies referred to above (Table A1). An example of GI side effects of LC omega-3 ingestion was reported in a 24-h postprandial study by Cuenoud et al. [21] which used doses of approx. 3g of LC omega-3; in this study, there were 22 treatment emergent adverse effects in 11/24 subjects including nausea, diarrhea, and headache. Since these effects were not consistently reported in all subjects, malabsorption of LC omega-3 could contribute to inter-individual variability.
These GI-related events support the notion that some of the ingested fish oil escapes digestion and passes into the colon and is possibly excreted in the faeces. In support of this, a review by Costantini et al. [37] reported that dietary LC omega-3 supplementation can exert effects on the intestinal environment in humans including significantly altering the composition of the gut microbiome, likely indicating that some of the ingested LC omega-3 escaped digestion and absorption in the duodenum and exerted effects on the composition of the microbiota in the large intestine.
One consequence of variability in the postprandial absorption and subsequent uptake of LC omega-3 by the liver would a variability of the effect of LC omega-3 in lowering plasma TAG levels. Such variability has been frequently reported and was summarised in a recent review by Rundblad et al. (38) which identified a number of determinants of the variability, including genetic variants, epigenetics and gene expression profiles, gut microbiota and habitual intake of LC omega-3. This review did not consider variability in the postprandial response to LC omega-3 supplementation as a potentially influential factor.
Several postprandial and longer-term studies have identified individuals who showed little or no increase in plasma or RBC LC omega-3 levels [8,9,10,15]. These subjects could be categorised as non-responders, a topic which has not been pursued in the literature in the context of LC omega-3 supplementation. A recent study designed to precisely characterise the individual glucose responses to high carbohydrate or high fat meals used an n=1 trial design (39). Perhaps this approach could be adopted in characterising individuals’ responses to LC omega-3 supplements? Following the identification of such individuals, this would presumably lead to further studies investigating the proteomic and genomic characteristics of these individuals.
Future Directions
We hypothesise that the variability in the digestion and absorption of LC omega-3 fatty acids, resulting in a variable postprandial response, would lead to variability in plasma and red blood cell LC omega-3 levels following supplementation.
The implications of variability in red blood cell LC omega-3 levels, if supported by the above hypothesis, are that the utility of RBC omega 3 as a measure of status might be called into question if it is found that there are low and high responders to LC omega-3 intakes. Furthermore, this could have significant implications for LC omega-3 trials: for example, should low responders be excluded from trials? Finally, there are significant implications for consumers taking LC omega-3 supplements, including whether low responders gain any biological benefits from consuming LC omega-3 supplements? These matters will all need further study.
5. Conclusions
Postprandial variability in response to TAG ingestion is a common phenomenon, not restricted to oils rich in LC omega-3. This paper highlighted the postprandial variability in response to LC omega-3 oils, with the suggestion that part of the variability could stem from [a] maldigestion of these oils leading to excretion into the large bowel, and possibly in faeces and [b] variability in the absorption of the FFA and/or 2-MAG, either into the enterocytes, and/or the processing of the absorbed lipids into chylomicrons and/or the export of these into the lymph. The paper proposes that postprandial variability in plasma LC omega-3 responses might contribute to variations in LC omega-3 levels in both plasma and RBC following LC omega-3 supplementation
Author Contributions
AJS conceived the idea of this work and both AJS and DL wrote the manuscript.
Funding
This research received no external funding.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
Nil.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| LC omega-3 | Long chain omega-3 |
| TAG | Triacylglycerol |
| PL | Phospholipids |
| FFA | Free fatty acids |
| MAG | Monoacylglycerol |
| EE | Ethyl esters |
| CV | Coefficient of Variation |
| AUC | Area Under the Curve |
| iAUC | Incremental Area Under the Curve |
| GI | Gastrointestinal |
| RBC | Red Blood Cell |
| DHA | Docosahexaenoic Acid |
| EPA | Eicosapentaenoic Acid |
| DPA | Docoapentaenoic Acid |
Appendix A

Table A1.
Study details of the LC omega-3 postprandial studies grouped by the type of lipid supplement.
Table A1.
Study details of the LC omega-3 postprandial studies grouped by the type of lipid supplement.
| Supplement type |
Dose & subject details |
Study details |
Postprandial CV for the AUC/iAUC |
Comments | Reference |
| TAG oils | |||||
| TAG (fish oil) | 1700 mg EPA+DHA, n=15 (8F, 7M) healthy adults from northern Finland. | 72-h study, with breakfast (29g fat), standardised lunch, dinner and supper. |
CV for iAUC Plasma PL EPA+DHA 59.2±22.2 (SD), CV= 38% Plasma TAG EPA+DHA 35.0±26.5 (SD) CV= 76% (iAUC % x h). |
Nine adverse events: 1 after fish oil, 5 after krill meal and 3 after the fish oil (note double entries for fish oil which might be an error). Most events were classified as mild, and included loose stools. | Kohler et al. 2015 (12) |
| Re-esterified fish oil, (rTAG) | 1680mg EPA+DHA, n= 12 (M) healthy adult from Germany. | 72-h study, capsules with breakfast (30.1g fat). Standardised meals consumed throughout study. |
CV for AUC Plasma PL EPA+DHA, 59.8±36.8 (SD), CV= 62%. (AUC - % x h). |
Adverse events not mentioned | Schuchardt et al. 2011 (15) |
|
TAG (blended fish oil) |
452 mg EPA+DHA, n=20 (10F, 10M) healthy young adults from the UK. | 8-h study with breakfast (49.5g fat). |
CV for iAUC F: Plasma TAG EPA+DHA 63.1±1.8 (SEM) (SD 48.2), CV= 76% M: Plasma TAG EPA+DHA 84.4±7.4 (SEM) (SD 31.7), CV= 38% F: Plasma PC EPA+DHA 159.9±25.1 (SEM) (SD 112.2), CV= 70% M: Plasma PC EPA+DHA 167.7±30.3 (SEM) (SD 135.4) CV= 81% (iAUC umol/L x h). |
No reported adverse effects were reported during trial | West et al. 2019 (16) |
| TAG (fish oil) | 4.5g fish oil (19.8% EPA, 7.9% DHA, 1247mg EPA+DHA), n=7 healthy subjects from the UK. | 24-h study, no breakfast. |
CV for AUC Plasma EPA 25.3±11.2 (SD), CV= 44% (no units provided for AUC) |
Did not report any adverse effects | Wakil et al. 2010 (17) |
| PL-rich oils | |||||
| PL-rich (Krill oil) | 1700 mg EPA+DHA as krill oil, n=15 (8F, 7M) healthy adults from northern Finland. | 72-h study, with breakfast (29g fat), standardised lunch, dinner and supper |
CV for iAUC Plasma PL EPA+DHA 89.1±33.4 (SD), CV= 38% Plasma TAG EPA+DHA 24.5±17.6 (SD), CV= 76% (iAUC % x h). |
Adverse events mild. One subject reported increased defecation. | Kohler et al. 2015 (12) |
|
PL-rich (Krill oil) |
1680mg EPA+DHA, n= 12 M healthy. from Germany. | A 72-h study, capsules with breakfast (30.1g fat). Standardised meals consumed throughout study. |
CV for AUC Plasma PL EPA+DHA 80.0±34.7 (SD), CV= 43% (AUC % x h). |
KO had high FFA levels containing EPA and DHA (22%, 19% of total content, respectively). Adverse events not mentioned | Schuchardt et al. 2011 (15) |
| PL-rich (Krill oil) | 206mg EPA+DHA, n=24 (14F, 10M), healthy subjects from USA (48% non-Hispanic/Other, 52% Hispanic/Latino). | A 24-h study, no breakfast just water. Low fat lunch/dinner |
CV for iAUC Plasma EPA+DHA 448±248 (SD) CV = 55% (iAUC nmol/ml x hr) |
One adverse event - upper resp tract inf. – judged to be unrelated to treatment. | Guarneiri et al. 2023 (18) |
| PL-enhanced Fish oil | 337mg EPA+DHA, n=24 (14F, 10M), healthy subjects from USA (48% non-Hispanic/Other, 52% Hispanic/Latino).. | A 24-h study, no breakfast just water. Low fat lunch/dinner |
CV for iAUC Plasma EPA+DHA 440±286 (SD) – CV = 65% (iAUC nmol/ml x hr) |
One adverse event - upper resp tract inf. – judged to be unrelated to treatment. | Guarneiri et al. 2023 (18) |
| PL-rich (Krill oil) | 1.02g EPA, 0.54g DHA, n=10 (10M), healthy subjects from Germany. | A 10-hr study, 55g breakfast |
CV for iAUC Plasma EPA 137±39 (SD), CV= 28% Plasma DHA 70±39 (SD), CV= 56% (iAUC ug/ml/hr) |
Adverse events not mentioned | Kagan et al. 2013 (19) |
| Polar rich algal oil (incl. glycolipids & PC) | 1.5g EPA, n=10 (M), healthy subjects from Germany. | A 10-hr study, 55g breakfast |
CV for iAUC Plasma EPA 277±135 (SD), CV= 49% Plasma DHA 65±45 (SD) CV= 69% (iAUC ug/ml/hr) |
Adverse events not mentioned | Kagan et al. 2013 (19) |
| FFA | |||||
| FFA (Epanova) | 4 x 1g cap of each type. FFA contained 0.55g EPA, 0.22g DHA/g. Dose = 2200mg EPA, 880 mg DHA, n=26 healthy subjects from USA (3 non-Hispanic/Other, 23 Hispanic/Latino). |
24-h study, low fat breakfast (<10%en fat). The study was conducted at the end of 2-week trial taking 4g FFA/day. |
CV for iAUC Plasma EPA + DHA geom mean 19.1, CV= 34% (iAUC nmol x hr/ml) |
Adverse events not mentioned. | Offman et al. 2013 (20) |
|
FFA (OM3-FFA) |
1748mg EPA + 1516mg DHA as FFA, n=24, (15F/9M) normal healthy adults (but finished with n=21-23/group) from Switzerland. | A 24-h study. No breakfast but fat-free lunch & dinner. |
CV for AUC Plasma EPA 1496±734 (SD), CV= 49% Plasma DHA 1356±676 (SD), CV= 50% (AUC nmol-h/ml). |
There were 22 adverse events (mild) reported by 11 subjects including nausea, diarrhea, headache. | Cuenoud et al. 2020 (21) |
| FFA (OM3-CA) probably EPANOVA | 4g EPA+DHA as FFA, n=14 (7F/7M), healthy Chinese subjects. | A 24-h study; supplements consumed with low fat (<10%) breakfast. |
CV for iAUC Plasma EPA 975.6±517.3 (SD), CV= 53% Plasma DHA 273.6±144 (SD), CV= 53% (iAUC h x ug/mL) |
In a continuation of the postprandial study, subjects were provided supplement for 14 days, and 6/14 subjects reported at least one mild adverse event (diarrhea). | Jing et al. 2020 (22) |
| MAG | |||||
|
MAG (Maxsimil 3020) |
3000 mg EPA+DHA (1800 mg EPA, 1200mg DHA), n=20 (10F, 10M), healthy subjects from Canada | A 24h study, with breakfast 20% fat |
CV for AUC Plasma EPA+DHA 89.6±17.0 (SD), CV= 19%. (AUC mg/dl x h) |
Did not mention of adverse events. Does not mention positional distribution of MAG. | Chevalier and Plourde 2021 (23) |
|
MAG Enzymatically produced oil followed by distillation, with EPA main FA. |
4.5g MAG oil (19.8% EPA, 7.9% DHA; 1247mg EPA+DHA), n=7 healthy subjects from the UK. | A 24hr study, no breakfast |
CV for AUC Plasma EPA 13.4±12.5 (SD), CV= 93%. (no units provided) |
Did not report any adverse effects | Wakil et al. 2010 (17) |
|
1(3)-MAG |
1655mg EPA + 1275mg DHA, n=24, (15F/9M) normal healthy adults (but finished with n=21-23/group ) from Switzerland. | A 24-h study. No breakfast but fat-free lunch & dinner. |
CV for AUC Plasma EPA 1486±626 (SD), CV= 42% Plasma DHA 1206±600 (SD), CV= 50% (AUC nmol-h/ml). |
There were 22 adverse events (mild) reported by 11 subjects including nausea, diarrhea, headache. | Cuenoud et al. 2020 (21) |
| Ethyl esters (EE) | |||||
| Ethyl esters (Lovanza) | 1860mg EPA, 1500mg DHA. n=26/arm (10F, 16M), healthy subjects from USA (4 non-Hispanic/Other, 22 Hispanic/Latino). | A 24-h study, low fat breakfast (<10%en fat) postprandial study treatment conducted at the end of 2-weeks trial. |
CV for iAUC Plasma EPA + DHA 3.32, CV= 76%. (iAUC nmol x hr/ml) |
Adverse events not mentioned | Offman et al. 2013 (20) |
|
Ethyl esters (fish oil origin) |
680mg EPA+DHA, n=30 (20F,10M) from Australia. | A 24hr study, with breakfast (2g fat) |
CV for iAUC Plasma EPA+DHA 150.2±40.5 (SEM) (221 SD), CV = 147%. (units not supplied) |
Adverse events not mentioned | Bremmell et al. 2020 (24) |
|
Ethyl esters (GmbH) |
1680mg EPA+DHA, n= 12 healthy M . from Germany. | A 72-h study, capsules with breakfast (30.1g fat). Standardised meals consumed throughout study. |
CV for AUC Plasma PL EPA+DHA: 47.5±38.4 (SD), CV= 81% (AUC % x h). |
Adverse events not mentioned. | Schuchardt et al. 2011 (15) |
|
Ethyl esters (Omacor) |
Ethyl esters 3080mg (1700mg EPA, 1380mg DHA), n=24, F15/M9 normal healthy adults (but finished with n=21-23/group) from Switzerland. | A 24-h postprandial study. No breakfast but fat-free lunch & dinner. |
CV for AUC Plasma EPA: 163±252 (SD), CV= 154% Plasma DHA: 562±695 (SD), CV= 124% (AUC nmol-h/ml). |
There were 22 adverse events (mild) reported by 11 subjects including nausea, diarrhea, headache. | Cuenoud et al. 2020 (21 |
|
Ethyl esters (Omacor) |
1000mg ethyl esters omega-3, n=12 (M) healthy adults from Republic of Korea. | A 72-h study, breakfast, 15-20% fat 20 min before single dose of capsules with 200mL water. |
CV for AUC Plasma EPA 914±464 (SD), CV= 41% Plasma DHA 404±216, CV= 53% (AUC ug/ml x h). |
No adverse events were reported | Kang et al., 2023 (25) |
|
Ethyl esters |
3360mg EPA+DHA, n=40 (20F/20M) healthy adults from Switzerland and the UK. | A 48-h study, no breakfast, capsules consumed with water, but low-fat lunch (<15g) and standard meals thereafter. |
CV for iAUC Plasma EPA+DHA 46.96±31.05 (SD), CV= 66%. (iAUC ug x h/mL/g). |
No adverse events were reported. | Qin et al., 2017 (26) |
|
Ethyl esters (Lovaza) |
840mg EPA+DHA as EE, n= 18(9F, 9M) from USA. | A 72-h study, breakfast with 23g fat |
CV for iAUC Plasma EPA+DHA 764±93 (SEM) (395 SD), CV= 52%. (iAUC ug x h/mL) |
No significant changes in GI symptoms over study period | Cook et al. 2016 (27) |
|
Ethyl esters (no brand mentioned) |
3g EPA+DHA 1800 mg EPA, 1200mg DHA, n=20 (10F, 10M), healthy subjects from Canada. | A 24h study, with breakfast 20% fat |
CV for AUC Plasma EPA+DHA 38.4±12.4 (SD), CV = 32%. (AUC mg/dl x h) |
Did not mention of adverse events. | Chevalier and Plourde 2021 (23) |
| EPA and DHA ethyl esters (KD Pharma) | 2024mg EPA, 1921 mg DHA (oil form), n=10 (M) healthy subjects from Germany. | A 72-h cross-over study, with breakfast (36.6g fat) |
CV for iAUC EPA: Plasma EPA 2461±279 (SEM), 882 SD, CV= 36% DHA: Plasma DHA 1021±170 (SEM), 538 SD, CV= 53% (iAUC ug/ml) |
No adverse effects were reported. | Smieta et al. 2025 (28) |
| Wax ester | |||||
| Wax ester (Calanus finmarchicus oil) |
4g oil containing 260mg EPA and 156 mg DHA, n= 18(9F, 9M) from USA. | A 72-h study, breakfast with 23g fat |
CV for iAUC Plasma EPA+DHA 931±92 (SEM) (391 SD), CV= 42%. (iAUC ug x h/mL) |
No significant changes in GI symptoms over study period | Cook et al. 2016 (27) |
| EE with enhanced emulsification properties | |||||
| Self-emulsifying drug delivery system, Aquacelle ethyl esters | 680mg EPA+DHA Aquacelle EE n=30 (18F, 12M), healthy subjects from Australia. | A 24-hr study, with breakfast (2g fat) |
CV for AUC Plasma EPA+DHA 460.3±69.8 (SEM) (381 SD), CV = 83% (units not supplied) |
Adverse events not mentioned | Bremmell et al. 2020 (24) |
| Ethyl esters of omega-3 with enhanced solubility | 580mg novel liquid microcrystalline nanoparticle ethyl esters of omega-3, n=12 (M) healthy adults from Republic of Korea. | A 72-h study, breakfast, 15-20% fat 20 min before single dose of capsules with 200mL water. |
CV for AUC Plasma EPA 929±285 (SD), CV= 31% Plasma DHA 541±287, CV= 53% (AUC ug/ml x h). |
No adverse events were reported. | Kang et al., 2023 (25 |
|
Ethyl esters with self microemulsifying delivery system (SMEDS) containing ethyl esters |
1680mg EPA+DHA, n=40 (20F/20M) healthy adults from Switzerland and the UK. | A 48-h study, no breakfast, capsules consumed with water, but low-fat lunch (<15g) and standard meals thereafter. |
CV for iAUC Plasma EPA+DHA 358±141 (SD), CV= 39% (iAUC ug x h/mL/g). |
No adverse events were reported |
Qin et al., 2017 (26 |
| TAG either enteric coated or as microencapsulated emulsified form (LipoMicel, LMF) | Single meal, cross-over study. The standard and enteric-coated capsules contained 600mg omega-3 (400mg EPA + 200mg DHA); the microencapsulated form contained 374mg omega-3 (200mg EPA; 133 mg DHA; 41 mg DPA)., n=12 healthy adults (gender not specified) from Canada. | A 24-h study with breakfast (bagel, cream cheese, jam); lunch and dinner provided (no details) | CV for iAUC Blood total LC omega-3 Standard capsule 1498.9±443 (SEM), 1534 SD, CV=102% Enteric-coated capsule 2057.2±813.7 (SEM), 2819 SD, CV= 137% Microencapsulated capsule 16150±5454 (SEM), 18892 SD, CV= 117%. (iAUC ng/mL x h) |
No side effects were reported during the study. | Ibi et al. 2024 (29) |
| Whole Foods | |||||
| Whole food (herring) rich in omega-3 TAG | Single meal, cross-over study. The meals contained 3.2g omega-3 (baked herring) and 2.7g omega-3, (pickled herring). n=17 (M) overweight subjects from Sweden. | A 7-h study, meal of baked or pickled herring (29g fat, EPA+DHA 22% in baked herring and 18% in pickled herring). |
Baked herring -CV for iAUC Plasma EPA 108.3± 14.5 (SEM) (59.8 SD), CV= 56% Plasma DHA 120.0± 20.0 (SEM) (82.5 SD), CV= 68% Pickled herring – CV for iAUC Plasma EPA 93.9±10.8 (SEM) (44.5 SD), CV= 48% Plasma DHA 101.1±13.3 (SEM) (54.8 SD), CV= 54% (iAUC mg/L x h). |
Did not report side effects. | Svelander et al., 2015 (30) |
| Butter or sunflower oil supplemented with fish oil TAG | Single meal, cross-over study. The meals contain 1.9g omega-3 plus either 38g butter or 32g sunflower oil. n=26 (18F, 8M) from Australia. | A 6-h study, meal of mashed potato plus butter or sunflower oil plus omega-3 (1.8g EPA+DHA). |
Butter + omega-3 - CV for iAUC Plasma total omega-3 161.55±34.2 (SD), CV= 21% Sunflower oil + omega-3 - CV for iAUC Plasma total omega-3 164.89±42.02 (SD), CV= 26% (no units provided) |
Did not report adverse events. | Dias et al. 2015 (31) |
| Standard meal or standard meal enriched with fish oil | Single meal, cross-over study. The standard meal contained 0.4g EPA+DHA, and enriched meal contained 2.3g EPA+DHA (1.2g EPA, 1.1g DHA), n=20 (10F, 10M) from UK. | A 6-h study, meals were a liquid emulsion together with toast and jam/marmalade (55-56g fat). |
Standard meal CV for iAUC Plasma TG EPA 0.5±0.1 (SEM), 0.45 SD, CV= 89% Plasma TG DHA -0.5±0.2 (SEM), 0.9 SD, CV= 180% Enriched meal CV for iAUC Plasma TG EPA 3.2±0.4 (SEM), 1.79 SD, CV= 56% Plasma TG DHA 1.3±0.4 (SEM), 1.79 SD, CV= 138% (units not mentioned) |
Did not report adverse events. |
Burdge et al. 2007 (32) |
| Soup or rice crackers with encapsulated algal DHA (HighDHA) or capsules of DHA (HighDHA) | Single breakfast, cross-over study. Each treatment contained 400mg DHA and 14-15mg EPA, n=27 (M) from Australia and Singapore. | A 24-h cross-over study, all treatments included standard breakfast (7.3g fat) and low-fat snacks, lunch and dinner. |
CV for iAUC Soup: Plasma DHA 8069±5500(SD), CV= 68% Rice Crackers : Plasma DHA 7367±5599 (SD), CV= 76% DHA capsule: Plasma DHA 9864±8603 (SD), CV= 87%. (iAUC ug/ml x hr) |
No adverse events were reported during the study | Stonehouse et al. 2022 (14) |
References
- Enkhmaa B, Ozturk Z, Anuurad E, Berglund L. Postprandial lipoproteins and cardiovascular disease risk in diabetes mellitus. Curr Diab Rep. 2010 Feb;10(1):61-9. doi: 10.1007/s11892-009-0088-4. PMID: 20425069; PMCID: PMC2821507.
- Bansal S, Buring JE, Rifai N, Mora S, Sacks FM, Ridker PM. Fasting compared with nonfasting triglycerides and risk of cardiovascular events in women. JAMA. 2007 Jul 18;298(3):309-16. doi: 10.1001/jama.298.3.309. PMID: 17635891.
- Kolovou GD, Mikhailidis DP, Kovar J, Lairon D, Nordestgaard BG, Ooi TC, Perez-Martinez P, Bilianou H, Anagnostopoulou K, Panotopoulos G. Assessment and clinical relevance of non-fasting and postprandial triglycerides: an expert panel statement. Curr Vasc Pharmacol. 2011 May;9(3):258-70. doi: 10.2174/157016111795495549. PMID: 21314632.
- Newman JW, Krishnan S, Borkowski K, Adams SH, Stephensen CB, Keim NL. Assessing Insulin Sensitivity and Postprandial Triglyceridemic Response Phenotypes With a Mixed Macronutrient Tolerance Test. Front Nutr. 2022 May 11;9:877696. doi: 10.3389/fnut.2022.877696. PMID: 35634390; PMCID: PMC9131925.
- Berry SE, Valdes AM, Drew DA, Asnicar F, Mazidi M, Wolf J, Capdevila J, Hadjigeorgiou G, Davies R, Al Khatib H, Bonnett C, Ganesh S, Bakker E, Hart D, Mangino M, Merino J, Linenberg I, Wyatt P, Ordovas JM, Gardner CD, Delahanty LM, Chan AT, Segata N, Franks PW, Spector TD. Human postprandial responses to food and potential for precision nutrition. Nat Med. 2020 Jun;26(6):964-973. doi: 10.1038/s41591-020-0934-0. Epub 2020 Jun 11. Erratum in: Nat Med. 2020 Nov;26(11):1802. doi: 10.1038/s41591-020-1130-y. PMID: 32528151; PMCID: PMC8265154.
- de Groot RHM, Emmett R, Meyer BJ. Non-dietary factors associated with n-3 long-chain PUFA levels in humans - a systematic literature review. Br J Nutr. 2019 Apr;121(7):793-808. doi: 10.1017/S0007114519000138. Epub 2019 Jan 28. PMID: 30688181; PMCID: PMC6521789.
- Alijani S, Hahn A, Harris WS, Schuchardt JP. Bioavailability of EPA and DHA in humans - A comprehensive review. Prog Lipid Res. 2024 Dec 28:101318. doi: 10.1016/j.plipres.2024.101318. Epub ahead of print. PMID: 39736417.
- Sparkes C, Sinclair AJ, Gibson RA, Else PL, Meyer BJ. High Variability in Erythrocyte, Plasma and Whole Blood EPA and DHA Levels in Response to Supplementation. Nutrients. 2020 Apr 8;12(4):1017. doi: 10.3390/nu12041017. PMID: 32276315; PMCID: PMC7231102.
- Von Schacky C. Omega-3 fatty acids vs. cardiac disease--the contribution of the omega-3 index. Cell Mol Biol (Noisy-le-grand). 2010 Feb 25;56(1):93-101. PMID: 20196973.
- Köhler A, Bittner D, Löw A, von Schacky C. Effects of a convenience drink fortified with n-3 fatty acids on the n-3 index. Br J Nutr. 2010 Sep;104(5):729-36. doi: 10.1017/S0007114510001054. Epub 2010 Apr 27. PMID: 20420756.
- Ly R, MacIntyre BC, Philips SM, McGlory C, Mutch DM, Britz-McKibbin P. Lipidomic studies reveal two specific circulating phosphatidylcholines as surrogate biomarkers of the omega-3 index. J Lipid Res. 2023 Nov;64(11):100445. doi: 10.1016/j.jlr.2023.100445. Epub 2023 Sep 18. PMID: 37730162; PMCID: PMC10622695.
- Köhler A, Sarkkinen E, Tapola N, Niskanen T, Bruheim I. Bioavailability of fatty acids from krill oil, krill meal and fish oil in healthy subjects--a randomized, single-dose, cross-over trial. Lipids Health Dis. 2015 Mar 15;14:19. doi: 10.1186/s12944-015-0015-4. PMID: 25884846; PMCID: PMC4374210.
- Flock MR, Skulas-Ray AC, Harris WS, Etherton TD, Fleming JA, Kris-Etherton PM. Determinants of erythrocyte omega-3 fatty acid content in response to fish oil supplementation: a dose-response randomized controlled trial. J Am Heart Assoc. 2013 Nov 19;2(6):e000513. doi: 10.1161/JAHA.113.000513. PMID: 24252845; PMCID: PMC3886744.
- Stonehouse W, Klingner B, Tso R, Teo PS, Terefe NS, Forde CG. Bioequivalence of long-chain omega-3 polyunsaturated fatty acids from foods enriched with a novel vegetable-based omega-3 delivery system compared to gel capsules: a randomized controlled cross-over acute trial. Eur J Nutr. 2022 Jun;61(4):2129-2141. doi: 10.1007/s00394-021-02795-7. Epub 2022 Jan 18. PMID: 35041046; PMCID: PMC9106597.
- Schuchardt JP, Schneider I, Meyer H, Neubronner J, von Schacky C, Hahn A. Incorporation of EPA and DHA into plasma phospholipids in response to different omega-3 fatty acid formulations--a comparative bioavailability study of fish oil vs. krill oil. Lipids Health Dis. 2011 Aug 22;10:145. doi: 10.1186/1476-511X-10-145. PMID: 21854650; PMCID: PMC3168413.
- West AL, Miles EA, Lillycrop KA, Han L, Sayanova O, Napier JA, Calder PC, Burdge GC. Postprandial incorporation of EPA and DHA from transgenic Camelina sativa oil into blood lipids is equivalent to that from fish oil in healthy humans. Br J Nutr. 2019 Jun;121(11):1235-1246. doi: 10.1017/S0007114519000825. Epub 2019 Apr 12. PMID: 30975228; PMCID: PMC6658215.
- Wakil A, Mir M, Mellor DD, Mellor SF, Atkin SL. The bioavailability of eicosapentaenoic acid from reconstituted triglyceride fish oil is higher than that obtained from the triglyceride and monoglyceride forms. Asia Pac J Clin Nutr. 2010;19(4):499-505. PMID: 21147710.
- Guarneiri LL, Wilcox ML, Maki KC. Comparison of the effects of a phospholipid-enhanced fish oil versus krill oil product on plasma levels of eicosapentaenoic and docosahexaenoic acids after acute administration: A randomized, double-blind, crossover study. Nutrition. 2023 Oct;114:112090. doi: 10.1016/j.nut.2023.112090. Epub 2023 May 29. PMID: 37413768.
- Kagan ML, West AL, Zante C, Calder PC. Acute appearance of fatty acids in human plasma--a comparative study between polar-lipid rich oil from the microalgae Nannochloropsis oculata and krill oil in healthy young males. Lipids Health Dis. 2013 Jul 15;12:102. doi: 10.1186/1476-511X-12-102. PMID: 23855409; PMCID: PMC3718725.
- Offman E, Marenco T, Ferber S, Johnson J, Kling D, Curcio D, Davidson M. Steady-state bioavailability of prescription omega-3 on a low-fat diet is significantly improved with a free fatty acid formulation compared with an ethyl ester formulation: the ECLIPSE II study. Vasc Health Risk Manag. 2013;9:563-73. doi: 10.2147/VHRM.S50464. Epub 2013 Oct 1. PMID: 24124374; PMCID: PMC3794864.
- Cuenoud B, Rochat I, Gosoniu ML, Dupuis L, Berk E, Jaudszus A, Mainz JG, Hafen G, Beaumont M, Cruz-Hernandez C. Monoacylglycerol Form of Omega-3s Improves Its Bioavailability in Humans Compared to Other Forms. Nutrients. 2020 Apr 7;12(4):1014. doi: 10.3390/nu12041014. PMID: 32272659; PMCID: PMC7230359.
- Jing S, Zhang Z, Chen X, Miao R, Nilsson C, Lin Y. Pharmacokinetics of Single and Multiple Doses of Omega-3 Carboxylic Acids in Healthy Chinese Subjects: A Phase I, Open-Label Study. Clin Pharmacol Drug Dev. 2020 Nov;9(8):985-994. doi: 10.1002/cpdd.800. Epub 2020 Jun 21. PMID: 32567203.
- Chevalier L, Plourde M. Comparison of pharmacokinetics of omega-3 fatty acid supplements in monoacylglycerol or ethyl ester in humans: a randomized controlled trial. Eur J Clin Nutr. 2021 Apr;75(4):680-688. doi: 10.1038/s41430-020-00767-4. Epub 2020 Oct 3. PMID: 33011737; PMCID: PMC8035073.
- Bremmell KE, Briskey D, Meola TR, Mallard A, Prestidge CA, Rao A. A self-emulsifying Omega-3 ethyl ester formulation (AquaCelle) significantly improves eicosapentaenoic and docosahexaenoic acid bioavailability in healthy adults. Eur J Nutr. 2020 Sep;59(6):2729-2737. doi: 10.1007/s00394-019-02118-x. Epub 2019 Oct 21. PMID: 31637467.
- Kang KM, Jeon SW, De A, Hong TS, Park YJ. A Randomized, Open-Label, Single-Dose, Crossover Study of the Comparative Bioavailability of EPA and DHA in a Novel Liquid Crystalline Nanoparticle-Based Formulation of ω-3 Acid Ethyl Ester Versus Omacor® Soft Capsule among Healthy Adults. Int J Mol Sci. 2023 Dec 6;24(24):17201. doi: 10.3390/ijms242417201. PMID: 38139029; PMCID: PMC10743492.
- Qin Y, Nyheim H, Haram EM, Moritz JM, Hustvedt SO. A novel self-micro-emulsifying delivery system (SMEDS) formulation significantly improves the fasting absorption of EPA and DHA from a single dose of an omega-3 ethyl ester concentrate. Lipids Health Dis. 2017 Oct 16;16(1):204. doi: 10.1186/s12944-017-0589-0. PMID: 29037249; PMCID: PMC5644165.
- Cook CM, Larsen TS, Derrig LD, Kelly KM, Tande KS. Wax Ester Rich Oil From The Marine Crustacean, Calanus finmarchicus, is a Bioavailable Source of EPA and DHA for Human Consumption. Lipids. 2016 Oct;51(10):1137-1144. doi: 10.1007/s11745-016-4189-y. Epub 2016 Sep 7. PMID: 27604086.
- Schmieta HM, Greupner T, Schneider I, Wrobel S, Christa V, Kutzner L, Hahn A, Harris WS, Schebb NH, Schuchardt JP. Plasma levels of EPA and DHA after ingestion of a single dose of EPA and DHA ethyl esters. Lipids. 2025 Jan;60(1):15-23. doi: 10.1002/lipd.12417. Epub 2024 Sep 19. PMID: 39299684; PMCID: PMC11717491.
- Ibi A, Chang C, Kuo YC, Zhang Y, Du M, Roh YS, Gahler R, Hardy M, Solnier J. Evaluation of the Metabolite Profile of Fish Oil Omega-3 Fatty Acids (n-3 FAs) in Micellar and Enteric-Coated Forms-A Randomized, Cross-Over Human Study. Metabolites. 2024 May 7;14(5):265. doi: 10.3390/metabo14050265. PMID: 38786742; PMCID: PMC11123365.
- Svelander C, Gabrielsson BG, Almgren A, Gottfries J, Olsson J, Undeland I, Sandberg AS. Postprandial lipid and insulin responses among healthy, overweight men to mixed meals served with baked herring, pickled herring or baked, minced beef. Eur J Nutr. 2015 Sep;54(6):945-58. doi: 10.1007/s00394-014-0771-3. Epub 2014 Nov 22. PMID: 25416681.
- Dias CB, Phang M, Wood LG, Garg ML. Postprandial lipid responses do not differ following consumption of butter or vegetable oil when consumed with omega-3 polyunsaturated fatty acids. Lipids. 2015 Apr;50(4):339-47. doi: 10.1007/s11745-015-4003-2. Epub 2015 Mar 10. PMID: 25753895.
- Burdge GC, Sala-Vila A, West AL, Robson HJ, Le Fevre LW, Powell J, Calder PC. The effect of altering the 20:5n-3 and 22:6n-3 content of a meal on the postprandial incorporation of n-3 polyunsaturated fatty acids into plasma triacylglycerol and non-esterified fatty acids in humans. Prostaglandins Leukot Essent Fatty Acids. 2007 Jul;77(1):59-65. doi: 10.1016/j.plefa.2007.06.003. Epub 2007 Aug 10. PMID: 17693069.
- Shi Y, Burn P. Lipid metabolic enzymes: emerging drug targets for the treatment of obesity. Nat Rev Drug Discov. 2004 Aug;3(8):695-710. doi: 10.1038/nrd1469. PMID: 15286736.
- Wang TY, Liu M, Portincasa P, Wang DQ. New insights into the molecular mechanism of intestinal fatty acid absorption. Eur J Clin Invest. 2013 Nov;43(11):1203-23. doi: 10.1111/eci.12161. Epub 2013 Sep 18. PMID: 24102389; PMCID: PMC3996833.
- O’Connor GT, Malenka DJ, Olmstead EM, Johnson PS, Hennekens CH. A meta-analysis of randomized trials of fish oil in prevention of restenosis following coronary angioplasty. Am J Prev Med. 1992 May-Jun;8(3):186-92. PMID: 1385968.
- Belluzzi A, Brignola C, Campieri M, Camporesi EP, Gionchetti P, Rizzello F, Belloli C, De Simone G, Boschi S, Miglioli M, et al. Effects of new fish oil derivative on fatty acid phospholipid-membrane pattern in a group of Crohn’s disease patients. Dig Dis Sci. 1994 Dec;39(12):2589-94. doi: 10.1007/BF02087694. PMID: 7995183.
- Costantini L, Molinari R, Farinon B, Merendino N. Impact of Omega-3 Fatty Acids on the Gut Microbiota. Int J Mol Sci. 2017 Dec 7;18(12):2645. doi: 10.3390/ijms18122645. PMID: 29215589; PMCID: PMC5751248.
- Rundblad A, Sandoval V, Holven KB, Ordovás JM, Ulven SM. Omega-3 fatty acids and individual variability in plasma triglyceride response: A mini-review. Redox Biol. 2023 Jul;63:102730. doi: 10.1016/j.redox.2023.102730. Epub 2023 May 3. PMID: 37150150; PMCID: PMC10184047.
- Ma Y, Fu Y, Tian Y, Gou W, Miao Z, Yang M, Ordovás JM, Zheng JS. Individual Postprandial Glycemic Responses to Diet in n-of-1 Trials: Westlake N-of-1 Trials for Macronutrient Intake (WE-MACNUTR). J Nutr. 2021 Oct 1;151(10):3158-3167. doi: 10.1093/jn/nxab227. PMID: 34255080; PMCID: PMC8485912.
Figure 1.
Caption.

Figure 2.
(a) Relationship between the dose of supplement (mg) and variability of AUC (CV%); (b) relationship between CV’s (%) for pairs of treatments within the same laboratory.
Figure 2.
(a) Relationship between the dose of supplement (mg) and variability of AUC (CV%); (b) relationship between CV’s (%) for pairs of treatments within the same laboratory.
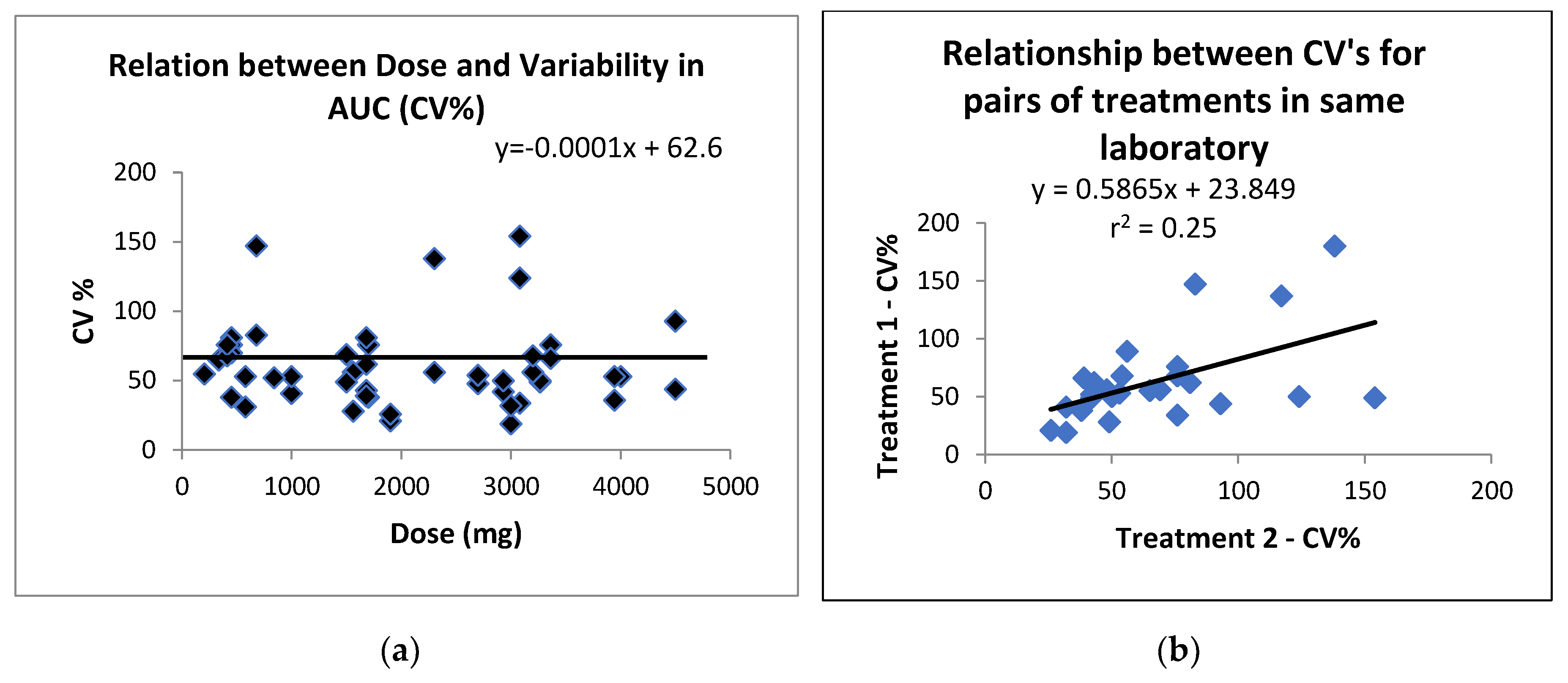
Table 1.
Summary of the postprandial variability in the area under the curve (AUC) or incremental AUC data for different chemical forms of the LC omega-3 supplements.
Table 1.
Summary of the postprandial variability in the area under the curve (AUC) or incremental AUC data for different chemical forms of the LC omega-3 supplements.
|
Omega-3 lipid supplement |
Number of treatments1 |
Variability in AUC (iAUC) expressed as coefficient of variation (CV)2 |
| TAG | 6 | 38-102%, with 7/10 data points3 >50% |
| PL or polar lipid | 6 | 28-76%, with 5/9 data points >50% |
| FFA | 3 | 34-53%, with 3/5 data points >50% |
| MAG | 3 | 19-93%, with 2/4 data points >50% |
| Ethyl esters (EE) | 9 | 32-154%, with 9/12 data points >50% |
| Wax esters | 1 | 0/1 data points >50% |
| EE with enhanced emulsification |
4 | 31-137%, with 4/6 data points >50% |
| Whole foods | 4 | 21-138%, with 7/10 data points > 50% |
| Total | 36 | 19-154%, with 37/57 data points >50% |
1Treatments = individual treatments within a study. 2Coefficient of variation (CV) = SD/mean*100. 3Data points = number of AUC outcomes (eg. TAG, PL, etc).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



