
Submitted:
25 September 2025
Posted:
25 September 2025
You are already at the latest version
Abstract
Synapses, abundant in the brain, are structures needed for life. Our introduction, based on forms of such structures published few decades ago, helped introducing recent concepts of health and diseases. Developing axons govern their growth by cell-to cell communication, axon guidance, synapse orientations. The assembly of synapses requires the organization and function of pre-synaptic and post-synaptic neuron terminals, connected to each other by extracellular thin clefts. Upon synapse stimulation the clefts expand up to several folds while pre- and post-synaptic thickness remain unchanged. In additional responses neurons co-operate with astrocytes and EVs, the latter dependent on extracellular and also on intracellular spaces. Glial cells and/or EV secretions induce neurons by various effects including traveling changes. Pre-synaptic responses are defined canonical if based on neurotransmitter release; non-canonical when dependent without release and also when discharged not of neurotransmitters but of EVs. Healthy and diseases depend on other general properties, such as those defined molecularly. Among neurodegenerative diseases, attention is specified by various properties of Alzheimer’s and other diagnoses. Critical identifications can be due to astrocyte and microglia cells or multiple effects induced by EVs. At present the complexity of the therapies is developing innovative initiatives.
Keywords:
Alzheimer’s
; amyloid-β
; Arc
; astrocyte
; axon
; biomarker
; canonical
; cell-to-cell
; clathrin
; cleft
; clinical/therapeutic
; dendrite
; endocytic
; exocytosis
; glia
; microglia
; navigation
; neurotransmitter
; polyphenolic
; pre-synaptic/post-synaptic
; reclustered
; recycling
; special issue
; spine
; synaptic vesicle
; strokes
; SubType
; tau
; trails/impact
; triplets
1. Introduction
The present Special Issue (SI) deals with synapses, in particular about these structures, especially those existing in the central nervous system (CNS), with innovations developed during the last several years. Taking into account the developments that take place during such Introduction, I noted that at least half of the presented information is due to well-known events published in early years, i.e. before 2018. Large fractions of the additional information, published more recently, has been recognized directly related or connected to previous events. Because of the nature and of their coordinations, the properties presented in this Introduction appear to anticipate the mechanisms and processes that are presented in the following Sections of this review.
The first step presented here includes the axon, the first component of the synapses located in the somato-dendritic compartment of neurons. Such axons are distinct in various properties: composition, diameter and length [1]. The formation of axons includes progressive extension accompanied by navigation and orientation. The length of axons varies beaucoup among synapses. All together, however, the axons account for a high fraction of the whole neuronal volume [2]. In most neurons the axon is single, followed however by multiple branches. Its activity depends mostly on voltage-gated ion channels, necessary for the activity of their action potentials. Membrane trafficking of axons brings membrane and proteins to the plasma membrane, maintaining the polarized distribution of proteins into neuronal subdomains [3].
As widely accepted since many years, synapses are based on two coordinate neuron fractions, inducing the cell-to-cell signaling events necessary for the brain function. Upon their contact, neuron fractions undergo dramatic structural changes becoming one pre-synaptic, the other post-synaptic. This connection is the key of the synaptic structure and function [4]. The initial processes start from the activation of the pre-synaptic terminals including the exocytoses of their synaptic vesicles filled of neurotransmitter, distributed to selected sites of the plasma membrane. The discharged neurotransmitters are received by the corresponding receptors at the surfaces of post-synaptic sites. Vesicle exocytoses are followed by membrane retrieval, either rapid re-uptake or ensuing recycling of endocytic membranes [5].
The post-synaptic activation of specific receptors occurs at dendrites, the numerous, much shorter protrusions where the signaling activations or inhibitions are received. Dendrites, all covered of spines at their surface membrane, respond by ligand-gated ion channels producing depolarization or hyperpolarization of graded potentials. The essential property of dendrite signaling depends on the possible multiplicity of their plasma membrane rafts covered by specific receptors, appropriately distributed at close distance from the pre-synaptic discharge. In response to their receptor activation, the post-synaptic signaling grows up by multiplicity, the summing up and circulation of their responses [6,7,8].
For many decades the synapses, including two neurons of their cell-to-cell signaling events, were considered necessary for normal brain function. In addition neurons were considered positive also for many aspects of diseases in the central nervous system. At the beginning of 2000 the presence and the role of the extra brain cells, the glial cells, became present and active at synapses. In 2005 the astrocytes, the most important glial cells of the brain, were first recognized present at the extra-role of tripartite synapses [9]. At the same time they were shown to play roles in synapse formation [10]. In 2010 the astrocyte results were confirmed and extended by the control of synapse activities: establishment, functions and plasticity [11] (Table 1). Following times have extended the need of astrocytes in several key functions of synapses, including secretion. Moreover, astrocytes are relevant in many cases, not only in health but also in diseases [12]. In addition they are not the only glial cell type involved. In synapses, distinct and highly interesting effects are induced also by microglia [13].
From the knowledge presented up to now we can conclude that synapses are produced and governed by neurons cooperating with the two types of glial cells. Although correct, this conclusion is not complete. In the list of actions involved in the generation and functions of synapses we need to include also the extracellular vesicles (EVs), structures generated and released by all types of cells [14]. At synapses the cells producing EVs are neurons, astrocytes and microglia. Of the various types of EV in many organs, one is very abundant in synapses, the exosome of type 2, generated and then accumulated within cytoplasmic multi vesicular bodies (MVBs) [14]. These exosomes are involved in their generation and in various relevant functions, highly important in synapses. The properties of EVs will be discussed in the following Sections of this review
In the course of this Introduction the properties of synapses, including the two, pre- and post-components, have been presented in sequence, anticipating their coordinate structure and function. In general terms, however, the two main components of synapses have been found to require distinct properties dependent on their innovative developments. Based on these considerations I have decided to organize my presentation of synapses in two sections. The present review has been focused primarily on the pre-synaptic form, illustrated and discussed in terms of health, diseases and therapies. In this presentation the post-synaptic forms are not yet complete. In the near future they will be organized in in a second form, completing the presentation of our synapses.
2. Axons and Synapses
In the presentation of these synapses, the first investigated areas include neuronal axons, the structures that maintain the pre-synaptic order, necessary for their transmission [15]. Within all synaptic neurons, axons are peculiar structures characterized by many specific proteins [15,16]. Here I will concentrate the presentation of general concepts of axons with ensuing special interests about synapses.
Canonical versus non-canonical properties refer to two distinct neuronal processes released by exocytosis: the pre-synaptic discharge by exocytosis of synaptic vesicles loaded with neurotransmitters (left); and exocytosis of MVBs filled with EVs (right).
A and B refer to different conditions of synaptic terminals connected by cleft. A: during weak stimulation clefts are thin. B: during strong stimulation the pre-synaptic and MVB exocytoses are stimulated. In addition the clefts space enlarge, with thicker space and longer trees connecting pre- and post-synapses. During such stimulation the thickness of pre- and post-synapses does not change substantially.
Several general properties of neurons, and also their variously observed details, depend not only on axons but also on their multiple, associated EVs, in particular those of neuronal origin. The most important axonal functions include synapse formations, cell-to cell communications, axon guidances, and synapse orientations [15,16]. Synaptic axons, because of their role in both electrical and molecular signaling, are vital for neuronal operations. Their functions, however, are not always independent. Their interactions with EVs, critical for their physiology, are very important also for many pathological processes [17,18,19].
The studies of synaptic terminals are often carried out not only by many EVs, but also by two neuronal terminals: pre-synaptic of the various post-synaptic structures, assembled together by the third intermediate connector, the thin cleft (Figure 1). Interestingly, analyses of synapses by high-resolution microscopy have revealed the relevance of the three components that carry out functions distinct from each other. In response to cell stimulation, cleft connections induce the enlargement of the extracellular space between pre-synaptic and post-synaptic structures (Figure 1) [20]. In such excitatory conditions the thickness of synaptic clefts undergo several times of increase. These results are exciting because the enlargement of clefts appears as a direct regulator of synapses. In contrast the thickness of the pre- and post-synaptic structures remain largely unchanged, controlled by space occupied by factors that are not enlarged [21]. The confirmation of exciting results [21,22,23] demonstrates that the changes of clefts, established in the course of stimulation, are due to the activation of protein trees connecting the pre-synaptic to post-synaptic sites. The essential neurotransmitter release and the post-synaptic sites, dependent on rafts of cleft surface oriented in front of the pre-synaptic sites, govern the functional receptors at least potentially activated [21,22,23,24].
Another important aspect of synaptic organization and function are the tripartite structures. In addition to the pre- and post-neuronal sites, which are included in all types of synapses, the tripartite forms include a third site, due to astrocyte connections associated to post-neuronal sites [9,24]. Summing up, therefore, bipartite synapses depend on the two, pre- and post-neuronal sites. For the tripartite synapses, the bipartites include also the third portions composed by astrocytes, present as additions to synapses.
In addition to synapses based on bipartite and tripartite sites, with the latter including astrocytes, others exist which in contrast include also cytoskeleton-associated proteins, the activity-regulated cytoskeletal–associated protein Arc [25]. In stimulated conditions the Arc concentration is higher and more active in the post-synaptic than in the pre-synaptic sites, contributing to both and also to the third component cleft, i.e. the three integrated sites [25]. Same Arc protein participates to the distribution of a misfolded form of the tau proteins, active both in the cytoplasm and at the surface of neurons. In established Arc-dependent releases the EVs play significant roles in neurodegeneration by elimination of intracellular tau together with the increase of extracellular tau, the latter accumulated at neuronal surfaces [26].
3. Astrocytes and Other Glial Cells
The results reported in [17,20] are focused on synaptic fusions due to exocytoses of neuronal nature, including the pre-synaptic neurotransmitter release and the post-synaptic activated receptors. Because of the two trans-synaptic components, these processes are traditionally called bipartite. Two additional aspects of these synapses have been emphasized during the last decades. The first deals with astrocytes. These cells are always present, however only few of them are protective, able to restore synaptic functions in response to the establishment of their damages. Moreover, some of these effects have been found to depend on molecules secreted or released by astrocytes [27,28] (Table 1). In other conditions, the interaction of astrocytes with synapses has been shown to take place only upon secretion of proteins released by microglia activation [29].
The effects of astrocytes on synapses are not always covered by the preliminary action of microglia. Various effects of the latter cells have been found to induce relevant effects on CNS, including developments and plasticity [30,31]. Another mechanism similarly operative in synapses depends on cytokines, another form of regulator controlling general responses [32]. So far several glia and other cells have been reported regulating the activity of pre-synaptic terminals. In other conditions, however, such effects are induced not by the various types of cells already mentioned, but some of their EVs addressed primarily to synapses. The functional EV signaling released by glial cells requires relevant targets of cells achieved in the nervous system [33,34,35]. In such conditions the EVs, released by their cells of origin, can be implicated in short- and long-distance traveling, operative from local brain sites, addressed to distinct large areas of the body [33,34,35]. Summing up, in these conditions the distinct morphology of neurons is maintained for extended periods of adult life. The mechanisms that maintain the morphology of mature neurons depend, at least in part, on elevated release of glial EVs [35,36].
The brain areas affected by data from both neurons and glia, spread in the whole CNS, are quite variable. In some cases the formation of synapses induced by astrocytes are relevant for image recognition [37]. In other cases they are followed by ischemia [38]. The EVs generated by both neurons and astrocytes are spread in the whole CNS. The recent year integrated data in terms of signaling are still interpreted as canonical and non-canonical properties of neurotransmitter release (Figure 1). These properties are relevant when discussing events occurring at synapses [39,40,41,42].
Almost all the brain important processes include neurons and glial cells working together with various types of their EVs. Among these processes are the fusions of EVs either with specific neurons, such as those of synapses, or of neuronal diseases of various types [34,37,40,41,42]. Neurodegenerative diseases, occurring after EV treatments, are presented in the next Section.
4. From Mechanisms to Diseases and Therapies
This Section is focused on wide and complex functions of the various synapses active in the brain. Neurotransmission is sustained by discharge and refilling of synaptic vesicles (SVs) by neurotransmitters, taking place at critical surface sites of pre-synapses. Until recently, the SVs were believed to take place either by fusions, followed by rapid pore closures, or by clathrin-mediated endocytosis occurring at the plasma membrane. By these and additional synaptic processes discussed recently, the discharged SVs are refilled and re-clustered in the course of physiological events [40,41,42].
Recent studies have revealed details about the dynamics of nerve terminals. Such processes, specific of pre-synaptic responses, take place by unique mechanisms now defined as liquid-liquid phase separation (LLPS) accumulated in response to multiple electrical forces [43]. Under these conditions specific proteins involve in the neurodegenerative discharge of synaptic vesicles (SVs) are specifically confined to small membrane areas of nerve terminals where LLPSs are induced by the accumulation of aliphatic alcohols. The ensuing local accumulation governs the accurate distribution by clustering and recycling active zones that support the evoked, Ca2+-dependent fusions of SVs followed by the immediate discharge of their neurotransmitters. These results indicate that LLPS, once accumulated in the pre-synaptic terminals, induce relevant processes of such distribution. These processes include the expansion of cleft, the extracellular space between pre- and post-synaptic terminals, governed by enlarged trees [21,24] (see Figure 1), and also the rapid SV discharge. In contrast the process largely spares any spontaneous neurotransmission [44,45].
Upon their discharge the SVs, in the course of their physiological activity, undergo refilling and re-clustering [46,47]. Additional developments have been established about the relevance and function of soma and integration of synapses, located at significant distances from their sites of origin. Finally, the SV locations and their functions are continuously integrated from neurons and activated glial cells by ongoing EVs produced and released to the extracellular space. Years ago, the pre-synaptic functions of SVs had been defined by two properties: canonical, due to the co-localization of their SV exocytoses; and non-canonical, defined by the lack of neurodegeneration discharge [48]. Additional non-canonical responses have been recently interpreted based on EVs, first developed in the course of their pre-synaptic functions, then released to the extracellular space (Figure 1) [48].
SubSection A: Diseases. The pathology of the brain includes critical participation of synapses of great importance. Here I will start with a number of examples concerning important diseases. In order to mention a list of brain diseases, I include a number of treatments administered to different patients or properties of such diseases: psychiatry patients [49]; many diseases appearing in response to CNS [50]; responses inducing alterations of neuronal morphology [35]; biomarkers developed for CNS diseases [51]. These diseases affect considerable numbers, but not all treatments administered to patients.
The relevant step of this presentation deals with glial cells and their cooperation with synapses, already considered during the previous Section 3. A second summary of disease processes is presented at [52], a review from the Handbook of Clinical Neurology that introduces the synaptic neurons interactions with astrocytes, microglia and EVs generated by both these cell types [52]. Properties about the last two glial cells, astrocyte and microglia, and the EVs [53,54] extend in terms of diseases the health information previously anticipated in the Introduction [13,14], and then discussed in several cases.
The glial cells mentioned in [52], which are highly important in this Section, include especially the astrocyte [55] (Table 1). Astrocytes are excellent coordinators employing important chemical properties. From 2012 distinct signals secreted by astrocytes associated to neurons were found to act processes of synaptogenesis, maturation, and synaptic pruning [56]. Interestingly glypical 4, one astrocyte- secreted signal, was found to act on synapses of the hippocampus; its similar factor, glypican 6, on those of the cerebellum [56]. Employed glypicans have been found more effective when associated to heparan sulfate, inducing effects on synaptic development, neural plasticity and disorders [57]. Moreover, astrocytes were found to induce results with potential effects on synapses. For example neurocan, a protein secreted by astrocytes, has been found to induce inhibitory synapses and strong regulation of their formation [58]; the new form of glypcan 5 regulates synapse maturation and stabilization [59]; astrocyte regulation governs synapse formation, maturation and elimination [60] (Table 1).
Most of the synaptic changes dependent from astrocytes [49,50,51,52,53] induce effects analogous to those already reported in the previous Sections. Interesting here are the astrocyte-secreted protein effects including maturation, stabilization, plasticity, disorders, elimination [52,53]. The effects induced by distinct secreted forms of glypicans appear in various areas of the brain [49].
Neuroegenerative diseases have been found including Alzheimer’s (AD), amyotrophic lateral sclerosis, Parkinson’s and Huntington’s. These diseases are different, including however some common properties. Here is the state of flavinoids, a broad family of polyphenolic chemicals present in a wide variety of plants [61]. Because of their neuroprotective function, flavonoids have attracted large interest. Here I intend to emphasize their effects inducing reduction of neurodegenerative symptoms accompanied by prevention of synaptic loss and enhanced cognitive functions. For the future of these diseases, a lot of researches are expected [61,62]. Only a small fraction of patients, affected by genetic defects, exhibits a fraction of their disease at relatively young age. In most other patients, AD appears at, or after 60 years. Among the neurodegenerative diseases is AD, focusing on advanced results recently reported: the identification of genes with potential role of synaptic vesicles [62]; amyloid-β labeling of synapses in live human brain slice cultures [63]; correlation of glial reactivity with synapse dysfunctions across aging [64]. About AD therapy: advancing of amyloid-β implications [65]; computational strategies and strong innovations in treatment of AD diseases [66].
SubSection B: Therapy. Interest about therapy is considerable in the whole Section 4, directly emphasized in a number of previous publications [46-48 and others]. The present SubSection is dedicated to specific developments of AD and other neurodegenerative diseases. Many developments of these diseases have been reported recently. The advanced forms, first appropriately modified by reinforcing their addresses, are loaded by diseases and then released to their specific targets. Good answers and limitations by brain EVs have been reported to appropriate results [39,60,61,62,63,64]. Intense studies in the CNS have been dedicated to clinical and therapeutic potentials. The latter include ongoing developments and advanced methodologies employed in AD studies [62,63,64,65,66].
Here I intend to mention examples of recent development in the field of therapy. The first presented studies result in aging and therapeutics investigations in patients [67] followed by the development of EV drug delivery systems used to enhance neurological therapy [68]. Three following articles deal with other EVs involved in emerging development of neurodegenerative diseases, presented here in clinical terms [69,70,71], followed by a clinical implication about neurodegenerative diseases [72]. ]. In addition to the AD data presented here, information about therapy is given by references to other neurodegenerative diseases. The last three publications of this SubSection [73,74,75] deal with clinical implications about therapeutic agents different from AD: Huntington’s [73], ALS [74], multiple sclerosis [75].
5. Conclusions
The present study, concentrated preferentially on the pre-synaptic formation and function, depends on general aspects and also on various details concerning neurons, glial cells and EVs, discovered or simply identified during recent studies [31-35 and Figure 1]. The role of astrocytes, presented in the 1, 3 and 4 Sections, is illustrated in the Table 1 of this review.
Main components of axons, characterized by general specificities, play key roles in the development of neurons, the initial component of synapses. Properties of these axons depend on two mechanisms: 1. Heterogeneity of pre-synaptic structures; 2. Role of synaptic clefts governed by their trees [36,37,38,39,40,41].
The role of EVs, released by neurons and glial cells, is due to various functions: 1. Cell-to-cell and neuron-to-glia communications; axon guidance; synapse orientations; other processes; 2. Tripartite synapses generated by astrocyte addition to bipartite synapse forms; 3. Various ensuing processes including ischemic stroke and other CNS diseases [43,44,45,46,47,48,49,50,51,52].
Highly important synapses are needed for active mechanism operative on distinct forms and areas of diseases. Attention is dedicated to various forms of neurodegeneration treated by polyphenolic agents of innovative treatment [53,54,55,56,57,58,59,60,61,62,63,64,65,66]. The final innovative therapies are considered of potential interest for relevant, innovative improvements [67,68,69,70,71,72,73,74,75].
Authorships
The author has analyzed the improvements and innovative developments in the field of the presentation.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
Abbreviations
AD: Alzheimer’s; Arc, activity-related cytoskeletal protein; CNS, central nervous system; EV, extracellular vesicle; MVB, multi-vesicular body; SI, Special Issue; SV, synaptic vesicle.
References
- Leterrier C. The axonal initial segment: an updated viewpoint. J. Neurosci. 2018; 38: 2135-2145. [CrossRef]
- Rich S.K., Terman J.R. Axon formation, extension, and navigation: only neuroscience phenomenon. Curr. Opin. Neurobiol. 2018; 53: 174-182. [CrossRef]
- Wojnacki J., Galli T. Membrane duing development. Dev. Neurobiol, 2016; 76: 1185-11200. [CrossRef]
- Batool S., Raza H., Zaidi J., Riaz S., Hasan S., et al. Synapse formation from cellular and molecular mechanisms in neurodevelopmental and neurodegenerative disorders. J. Neurophysiol. 2019; 121: 1381-1397. [CrossRef]
- Scheiffele P. Cell-cell signaling during synapse formation in the CNS. Annu. Rev. Neurosci. 2003; 26: 485-508. [CrossRef]
- Meldolesi J. Post-synapses in the brain: role of dendrites and spine structures. Biomedicine 2022; 10: 1859. [CrossRef]
- Lopez-Hernandez T., Takenaka K.I., Mori Y., Kongpracha P., Sagamori S., et al. Clathrin-independent endocytic retrieval of synaptic vesicle protein mediated by the clathrin adaptor AP-2 at mammalian central synapses. Elife 2022; 11: e71198. [CrossRef]
- Saheki Y., De Camilli P. Synaptic vesicle endocytosis. Cold Spring Harb. Perspect. Biol. 2012; 4: a005645. [CrossRef]
- Peres G., Navarrete M., Araque A. Tripartite synapses: astrocytes process and control synaptic information. Trends Neurosci, 2009; 32: 421-431. [CrossRef]
- Allen N.J., Barres B.A. Signaling between glia and neurons: focus on synaptic plasticity. Curr. Opin. Neurobiol. 2005; 15: 542-548. [CrossRef]
- Eroglu C., Barres B.A. Regulation of synaptic connectivity by glia. Nature 2010; 468: 223-231. [CrossRef]
- Chung W.S., Baldwin K.T., Allen N.J. Astrocyte regulation of synapse formation, maturation, and elimination. Cold Spring Harb. Perspect. Bol. 2024; 16: a041352. [CrossRef]
- Pereira-Iglesias M., Maldonado-Teixido J., Melero H., Piriz J., Galea E., et al. Microglia as hunters or gatherers of brain synapses. Nat. Neurosci. 2025; 28: 15-23. [CrossRef]
- Mason A.J., Deppmann C., Winkler B. Emerging roles of neuronal extracellular vesicles at the synapse. Neuroscientist 2024; 30: 199-213. [CrossRef]
- Liu Y., Shual K., Sun Y., Zhu L., Wu X.M. Advances in the study of axon-associated vesicles. Front. Mol. Neurosci. 2022; 15: 1045778. [CrossRef]
- Chen S., Bao Q., Xu W., Zhai X. Extracellular particles: emerging insights into central nervous system diseases. J. Nanobiotechnology 2025; 23: 263. [CrossRef]
- Steele-Nicholson L.J., Andrews M.R. Axon-targeting motifs: mechanisms and applications of enhancing axonal localization of transmembrane proteins. Cells 2022; 11: 937. [CrossRef]
- Liu M., Teng T. Exosomes: new target for understanding axon guidance in the developing central nervous system. Front. Cell Dev. Biol. 2025; 12: 1510862. [CrossRef]
- Filannino F.M., Panaro M.A., Benameur T., Pizzolorusso I., Porro C. Extracellular vesicles in the central nervous system: a novel mechanism of neuronal cell communication. Int. J. Mol. Sci. 2024; 25: 1629. [CrossRef]
- Chen J., Tian C., Xiong X., Yang Y., Zhang J. Extracellular vesicles: new horizons in neurodegeneration. EBioMedicine 2025; 113: 105605. [CrossRef]
- Cole A.A., Reese T.S. Transsynaptic assemblies link domains of presynaptic and postsynaptic intracellular structures across the synaptic cleft. J. Neurosci. 2023; 43: 5883-5892. [CrossRef]
- Tao-Cheng J.H., Moreira S.L., Winters C.A., Reese T.S., Dosemeci A. Modifications of the synaptic cleft under excitatory conditions. Front. Synaptic Neurosci. 2023; 15: 1239098. [CrossRef]
- Muttathukunnel P., Frei P., Perry S., Dickman D., Muller M. Rapid homeostasis modulation of trans-synaptic nanocolumn rings. Proc. Natl. Acad. Sci. USA 2022; 119: e211904119. [CrossRef]
- Godavarthi S.K., Hiramoto M., Ignatyev Y., Levin J.B., Li H.Q., et al. Postsynaptic receptors regulate presynaptic transmitter stability through transsynaptic bridges. Proc. Natl. Acad. Sci. USA 2024; 121: e2318041121. [CrossRef]
- Ringsevjen H., Egbenya D.L., Bieler M., Davanger S., Hussein S. Activity-regulated cytoskeletal–associated protein (Arc) in presynaptic terminals and extracellular vesicles in hippocampal synapses. Front. Mol. Neurosci. 2023; 16: 1225533. [CrossRef]
- Tyagi M., Chadha R., de Hoog E., Sullivan K.R., Walker A.C., et al. Arc mediates intercellular tau transmission via extracellular vesicles. bioRxiv 2024; 22: 2024. [CrossRef]
- Walker C.D., Risher W.C., Risher M.L. Regulation of synaptic development by astrocyte signaling factors and their emerging roles in substance of abuse. Cells 2021; 9: 28. [CrossRef]
- Yamagata K. Astrocyte-induced synapse formation and ischemic stroke. J. Neurosci. Res. 2021; 99: 140-1431. [CrossRef]
- Faust T.E., LeeY.H., O’Connor C.D., Boyle M.A., Gunner G., et al. Microglia-associate crosstalk regulates synapse remodeling via Wnt signaling. Cell 2025; 188: 5212-5230. [CrossRef]
- Schafer D.P., Stevens B., Bennett M.L., Bennett F.C. Role of microglia in central nervous system development and plasticity. Cold Spring Harb. Perspect. Biol. 2024; 30: a041810. [CrossRef]
- Duran Laforet V., Schafer D.P. Microglia: activity-dependent regulators of neural circuits. Ann.N.Y. Acad. Sci. 2024; 1533: 38-50. [CrossRef]
- Zipp F., Bittner S., Schaefer D.P. Cytokines as emerging regulators of central nervous system synapses. Immunity 2023; 56: 914-925. [CrossRef]
- Blanchette C.R., Scalera L., Harris K.P., Zhao Z., Dresselhaus E.C., et al. Local regulation of extracellular vesicle traffic by the synaptic endocytic machinery. J. Cell Biol. 2022; 221: e202112094. [CrossRef]
- Dresselhaus E.C., Harris K.P., Blancette C.R., Koles K., Del Signore S.J., et al. ECRT disruption provides evidence against trans-synaptic signaling via extracellular vesicles. [CrossRef]
- Solana-Balaguer J., Campoy-Campos G., Matin-Flores N., Perez-Sisques L., Sitja-Roqueta L., et al. Neuron-derived extracellular vesicles contain synaptic proteins, promote spine formation, activate TrkB-mediated signaling and preserve neuronal complexity. J. Extracell. Vesicles 2023; 12: e12355. [CrossRef]
- Park S., Noblett N., Pitts L., Colavita A., Wehman A.M., et al. Dopey-dependent regulation of extracellular vesicles maintains neuron morphology. Curr. Biol. 2024; 34: 4920-4933.e11. [CrossRef]
- Ikezu T., Yang Y., Verderio C., Kramer-Albers E.M. Extracellular vesicle-mediated neuron-glia communications in the central nervous system. J. Neurosci. 2024; 44: e1170242024. [CrossRef]
- Lorenzo J., Rico-Gallego J.A., Binczak S., Jacquir S. Spiking neuron-astrocyte networks for image recognition. Neural Comput.2025; 37: 635-665. [CrossRef]
- Xia X., Wang Y., Quin Y., Zhao S., Zheng J.C. Exosome: a novel neurotransmission modulator or non-canonical neurotransmitter? Ageing Res. Rev. 2022; 74: 101558. [CrossRef]
- Nieves Torres D., Lee S.H. Inter-neuronal signaling mediated by small extracellular vesicles: wireless communication? Front. Mol. Neurosci. 2023; 16: 1187300. [CrossRef]
- Krishnan S., Klingauf J. The readily retrievable pool of synaptic vesicles. Biol. Chem. 2023, 404: 385-397. [CrossRef]
- Meldolesi J. Specific extracellular vesicles generated and operating at synapses, contribute to neuronal effects and signaling. Int. J. Mol. Sci. 2024, 25: 5103. [CrossRef]
- McCaig C.D. Synaptic physiology depends on electrical forces on liquid-liquid phase separation. Rev. Physiol. Biochem. Pharmacol. 2025; 187: 339-359. [CrossRef]
- Guzikowski N.J., Kavalali E.T. Functional specificity of liquid-liqid phase separatin at the synapse. Nat. Commun. 2024; 15: 10103. [CrossRef]
- Choi J., Rafiq N.M., Park D. Liquid-liquid phase separation in pre-synaptic nerve terminals. Trends Biochem. Sci. 2024; 49: 888-9009: 888-900. [CrossRef]
- Chanaday N.L., Cousin M.A., Milosevic I., Watanabe S., Morgan J.R. The synaptic vesicle cycle revisited: new insights into the modes and mechanisms. J. Neurosci. 2019; 39: 8209-8216. [CrossRef]
- Andres-Alonso M., Kreutz M.R., Karpova A. Autophagy and the endolysosomal system in presynaptic function. Cell Mol. Life Sci. 2021; 78: 2621-2639. [CrossRef]
- Brenna S., Krisp C., Altmeppen H.C., Magnus T., Puig B. Brain-derived extracellular vesicles in health and diseases: a methodological perspective. Int. J. Mol. Sci. 2021; 33: 1365. [CrossRef]
- Xu X., Iqbal Z., Xu L., When C., Duan L., et al. Brain-derived extracellular vesicles: potential diagnostic biomarkers for central nervous system diseases. Psychiatry Clin. Neurosci. 2024; 78: 83-96. [CrossRef]
- Chang C., Weiping L., Jibing C. Exosomal miRNA therapy for central nervous system injury diseases. Cell Mol. Neurobiol. 2024; 45: 3. [CrossRef]
- Zanirati G., Dos Santos P.G., Alcara A.M., Bruzzo F., Machado Ghilardi I., et al. Extracellular vesicles: the new generation of biomarkers and treatment for central nervous system diseases. Int. J. Mol. Sci. 2024; 25:7371. [CrossRef]
- Dzyubenko E., Hermann D.M. Neuroglia and extracellular matrix molecules. Handb. Clin. Neurol. 2025; 209: 197-211. [CrossRef]
- Shen H., Chen J., Liu M., Zhao M., Hu D., et al. Research progress of extracellular vesicles derived from mesenchymal stem cells in the treatment of neurodegenerative diseases. Front. Immunol. 2025; 16: 1496304. [CrossRef]
- Guo M., Hao Y., Feng Y., Li M., Mao Y., et al. Microglial exosomes in neurodegenerative disease. Front. Mol. Neurosci. 2021; 14: 630808. [CrossRef]
- Patel M.R., Weaver A.M. Astrocyte-derive small extracellular vesicles promote synapse formation via tubulin-2-mediated TGF-β signaling. Cell Rep. 2021; 34: 108829. [CrossRef]
- Allen N.J., Bennett M.L., Foo L.C., Wang G.X., Chakraborty C., et al. Astrocyte glypicans 4 and 6 promote formation of excitatory synapses via GluA1, AMPA receptor. Nature 2012; 486:410-4014. [CrossRef]
- Kamimura K., Maeda N. Glypian and heparin sulfate in synaptic development, neural plasticity, and neurological disorders. Front Neural Circuits 2021; 15: 59556. [CrossRef]
- Irala D., Wang S., Skers K., Nagendren L., Ulloa-Severino F.P., et al. Astrocyte-secreted neurocam controls inhibitory synapse formation and function. Neuron 2024; 112: 1657-1675. [CrossRef]
- Bosworth A., Contreras M., Sancho L., Salas H., Paumler A., et al. Astrocyte glypcan 5 regulates synapse maturation and stabilization. Cell Rep. 2025; 4: 115374. [CrossRef]
- Chung W.S., Baldwin K.T., Allen N.J. Astrocyte regulation of synapse formation, maturation, and elimination. Cold Spring Harb. Perspect. Biol. 2024; 16: a41352. [CrossRef]
- Kopalli S.R., Behi T., Kyada A., Rekha M.M., Kundias M., et al. Synaptic plasticity and neuroprotection: the molecular impact of flavonoids on neurodegenerative disease progression. Neuroscience 2025; 569: 161-183. [CrossRef]
- Zeng J, Zhang R., Hu H., Zhang C., Lu L. Integrative single cell RNA sequencing and mandelian randomization analysis reveal the potential role of synaptic vesicle cycling-related genes in Alzheimer’s disease. J. Prev. Alzheimer’s Dis. 2025; 12: 100097. [CrossRef]
- McGeachan R.I., Meftan S., Taylor L.W., Caterson J.H., Negro D., et al. Divergent actions of physiological and pathological amyloid-β on synapses in live human brain slice cultures. Nat. Commun. 2025; 16: 3753. [CrossRef]
- Rhoden F., Ferreira P.C.L., Bellaver B., Ferrari-Souza J.P., Aguzzoli C.S., et al. Glial reactivity correlates with synaptic dysfunction across aging and Alzheimer’s disease. Nat. Comm. 2025; 16: 5653. [CrossRef]
- Pokrzyk J. Kulczynska-Przybik A., Guzik-Makaruk E. Clinical presence of amyloid-β implication in the detection and treatment of Alzheimer's disease. Int. J. Mol. Sci. 2025; 26: 1935. [CrossRef]
- Paul J.K., Malik A., Azmal M., Gulzar T., Muhammad Talal R., et al. Advancing Alzheimer's therapy: computational strategies and treatment innovations. IBRO Neurosci. Rep. 2025; 18: 270-282. [CrossRef]
- Szczupak D., LjungQvist Brinson L., Kolarcik C.L. Brain connectivity, neural networks, and resilience in aging and neurodegeneration. Am. J. Pathol. 2025; 23: S0002--9440 (25)00027-6. [CrossRef]
- Vahab S.A., KV V., Kumar V.S. Exosome-based drug delivery system for enhanced neurological therapeutics. Drug Deliv. Transl. Res. 2025; 15: 1121-1138. [CrossRef]
- Wang L., Zhang X., Yang Z., Wang B., Gong H., et al. Extracellular vesicles: biological mechanisms and emerging therapeutic opportunities in neurodegenerative diseases. Transl. Neurodegener. 2024; 13: 60. [CrossRef]
- Malaguarnera M., Cabrera-Pastor A. Emerging role of extracellular vesicles as biomarkers in neurodegenerative diseases and their clinical and therapeutic potential in central nervous system pathologies. Int. J. Mol. Sci. 2024; 25: 10068. [CrossRef]
- Zhu Y., Wang F., Xia Y., Wang L., Lin H., et al. Research progress on astrocyte- derived extracellular vesicles in the pathogenesis and treatment of neurodegenerative diseases. Rev. Neurosci. 2024, 35: 855-875. [CrossRef]
- Vinaiphat A., Sze S.K., Clinical Implications of extracellular vesicles in neurodegenerative diseases. Expert Rev. Mol. Diagn. 2019; 19: 813-824. [CrossRef]
- Sharma S., Kaur I., Dubey N., Goswami N., Tanwar S.S. Berberine can be a potential therapeutic agent in treatment of Huntington’s disease: a proposed mechanistic insight. Mol. Neurobiol. 2025; May 16. [CrossRef]
- Burg T., Van Den Bosch L. Glycerophospholipids in ALS: insights into diseases mechanisms and clinical implication. Mol. Neurodegener. 2025; 20: 85. [CrossRef]
- De Angelis F., Nistri R., Wright S. Measuring disease progression in multiple sclerosis clinical drug trails and impact on future patient care. CNS Drugs 2025; 39: 55-80. [CrossRef]
Figure 1.
Combination of pre- and post-synaptic terminals connected by clefts. This Figure includes changes introduced in the Figure 1 of [38].
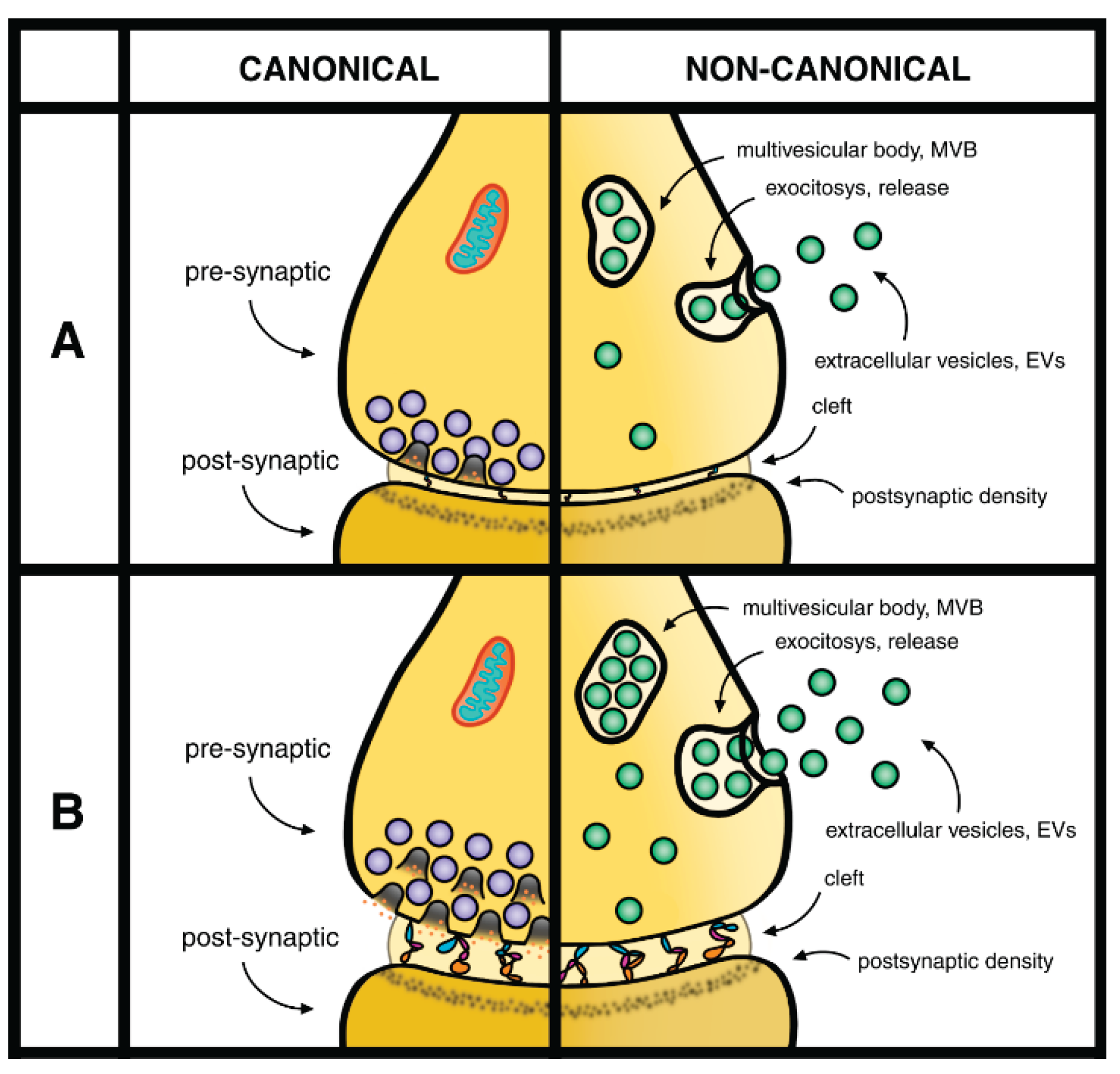

Table 1.
Role of astrocytes at synapses. Astrocytes are present in most sites of synapses appearing in this review. In three such sites the presentations of astrocytes are highly important.
Table 1.
Role of astrocytes at synapses. Astrocytes are present in most sites of synapses appearing in this review. In three such sites the presentations of astrocytes are highly important.
| Section 1. Introduction | In 2005, the role of astrocytes was two fold: accumulation of astrocytes in the post-synaptic site of tripartite synapses [9]; signaling between glia and neurons: focus on a synaptic process with their plasticity [10]. In 2010 the role of astrocytes is powerful on synaptic formation: function, plasticity and elimination [11]. Recent evidence has confirmed the astrocyte effects including astrocyte secretion [12]. |
| Section 3. Astrocytes and other Glial Cells | The important role of astrocytes revealed in these studies has included protection and restoration of synapses, induced by vesicle release and/or by astrocytes protein secretion [27,28]. |
| Section 4. From Mechanisms to Diseases and Therapies | Most of the synaptic changes dependent from astrocytes [49,50,51,52,53] induce effects analogous to those already reported in the previous Sections. Interesting here are the astrocyte-secreted protein effects including maturation, stabilization, plasticity, disorders, elimination [52,53]. The effects induced by distinct secreted forms of glypicans appear in various areas of the brain [49]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



