Submitted:
24 September 2025
Posted:
25 September 2025
You are already at the latest version
Abstract
Current health interventions, including dietary supplements, deliver bioactive compounds and ingredients to target various areas of human welfare, such as support for brain health, metabolic effects, energy function, and women's health. Gut wellbeing, including those that target the gut microbiome, is categorized as a separate domain. However, research has highlighted that the gut microbiome is a metabolically active organ, deeply interconnected in almost all areas of host health, not an isolated target. Thus, there is a need for solutions to intentionally design dual-targeting of both host cells and the gut microbiome, allowing for enhanced function and long-term effects. Based on this gap, we propose a new category within biotic solutions, termed co-biotics. Co-biotics is therefore “a substrate comprising of bioactive molecular compounds that, when delivered to permit both host metabolism and microbial accessibility, simultaneously modulate biological processes in both the host and its resident microbiota, to confer targeted health benefits". This paper will outline examples, as well as delivery requirements for co-biotic ingredients, which collectively allow for framing of these next-generation host-microbiome health solutions.
Keywords:
Co-biotics
; microbiome
; host-microbiome interactions
; dietary supplements
; bioactives
Introduction
Many health interventions, such as pharmaceuticals, supplements, functional foods, and nutritional strategies, aim to deliver bioactive compounds that exert discernible effects on human physiology. These interventions typically influence health domains (e.g., metabolic regulation, reproductive health, neurocognitive function), and the gastrointestinal tract is mostly regarded as a conduit for absorption, not a complementary axis by which to drive positive outcomes across each of these domains. On the contrary, the gut microbiome represents a metabolically active organ system and acts as an essential metabolic interface through which systemic physiological processes are coordinated to enhance health outcomes, including via their microbial products.
The co-evolutionary relationship between animals and their gut microbiota spans millions of years, resulting in obligate metabolic interdependencies. The human host has adapted to rely on microbes to support multiple biological functions across various host physiologies, with perturbations negatively impacting various areas of health and disease [1,2]. Some gut microbial species also show signatures of co-diversification and symbiotic specialization with the host and display limited survivability outside the human host [3]. Furthermore, the gut microbiome participates in multi-directional communication with extra-intestinal organs including via what has been termed the gut-organ axes [4], though it is not a linear connection and preferably should be designated gut-organ system.
Collectively, research has highlighted the gut microbiome’s role as an integrative regulator of host homeostasis, and a clear foundational target to enhance physiological function.
Microbiome
Foundation of the Gut-Organ Axes
The average human body contains roughly 30 trillion human cells and 38 trillion bacterial cells, with the vast majority of the latter residing in the colon, where pH, transit time, and substrate availability are favorable [5]. For decades, we have understood the importance of the gut microbiome in regulating our gut and digestive health, with direct impacts on local and systemic immune health [6]. However, evolving research has made it clear that the gut microbiome does not solely impact gut health - it is pivotal to many distinct areas of human health through direct and indirect mechanisms. Though we question the use of the term ‘axis’, for the purpose of this article, we will accept this microbial-host network being called the “gut-organ axis” [4] and from that, these axes are bi- or multi-directional pathways between the gut and non-gut organ systems, and communicate via neural, endocrine, immune, humoral, and metabolic pathways.
Amongst the gut-organ axes, the gut-brain axis is well studied, and an important regulator of several aspects of brain health, including cognition, mood, stress, anxiety, as well as developmental and neurodegenerative diseases [7,8]. Direct and indirect signaling are responsible for many gut-brain axis communications. For instance, short chain fatty acids (SCFA) produced by the gut microbiome have been shown preclinically to act on the vagus nerve to directly impact behavior [9,10]. In addition, a number of microbes are able to synthesize neurotransmitters (e.g., GABA, cortisol, dopamine) and modulate the regulation of both microbial- and/or host-derived neurotransmitters such as serotonin [11,12,13,14]. With the role of gut microbes being evident in brain health, probiotics targeting the gut-brain axis have been clinically-studied [15,16,17,18].
Another example of the gut-organ axis that is an emerging area of interest is the gut-skin axis. The association of dysbiotic gut microbiota with numerous inflammatory skin conditions has been well established; diseases such as atopic dermatitis, psoriasis, and acne are linked to dysbiosis, gut barrier disruptions, and its direct modification of the immune system and immune response [19,20]. Clinical studies using specific orally administered probiotic strains as well as prebiotics have shown improvement in various skin conditions via this gut-skin axis [21,22,23]. As research into the gut-skin axis continues to expand, several consumer products are following suit with the growth of “beauty from within” categories that are now ubiquitous on shelves.
The gut-organ axis is ever-expanding, and now includes the gut-lung axis, gut-heart axis, gut-liver axis, gut-hormone axis, gut-bladder axis, gut-hair follicle axis, and even the gut-eye axis [24,25,26,27,28,29,30]. While research in some of these areas is still preliminary, the large repertoire highlights that almost every aspect of human health is indeed tied to the health of the gut microbiome, and continued research will inevitably uncover more links and direct mechanisms of these interactions.
Modern Microbiome Disruption & Associated Human Health
The human microbiome, despite being a master of rapid adaptation, is vulnerable to stresses of modern living, with profound implications for systemic health. Multiple convergent factors, including dietary, pharmacological, and environmental, can destabilize gut microbial communities and diminish their capacity to support host physiology [31,32,33,34]. For example, intensive agricultural practices, soil nutrient depletion, and the widespread consumption of ultra-processed foods have collectively reduced the micronutrient density of the human diet [35]. These changes are believed to lower the likelihood of adequate micronutrient availability and increase preferential absorption in the small intestine, leaving limited quantities available to the colonic microbiota.
In parallel, exogenous agents such as pharmaceuticals (e.g., broad-spectrum antibiotics), food additives (e.g., emulsifiers) and agricultural pesticides, may directly perturb or deplete gut microbial populations [36,37,38,39]. Beyond xenobiotic exposures, lifestyle-associated factors including chronic psychological stress, insufficient or dysregulated sleep, age, and low levels of physical activity have each been shown to negatively modulate microbiome structure and function [40,41,42]. Taken together, various aspects of modern lifestyles can expose one to dysbiosis and gut microbial disruption.
As such, given the central role of the gut microbiome in regulating host physiology through multiple gut-organ axes, these disruptions represent a critical bottleneck to achieving optimal health outcomes. Interventions that target host physiology in isolation, without concomitant support for the microbiome, are therefore unlikely to realize their full therapeutic or preventive potential. Existing supplementation strategies that focus solely on the host often overlook the essential contributions of the microbiome in determining nutrient bioavailability, metabolic signaling, inflammatory status, and immune regulation. Without addressing the microbial ecosystem, most existing interventions provide only partial or transient benefits, as their efficacy is constrained by an impaired or unstable gut community. In therapeutics, the role of the microbiome in drug efficacy is an area of intense research, and the bidirectional influence of drugs and microbiome is being uncovered [33,43]. Due to strong links between the gut microbiome and therapeutic outcomes, precision medicine based on the patient’s gut microbiome, or co-administration of probiotics are areas currently being explored [44,45]. Overall, there is a need for health solutions that are intentionally designed to support both host and microbial systems in tandem, thereby enhancing resilience, optimizing function, and unlocking the full potential of biotic supplementations.
Landscape of Biotic Innovation
Biotics are generally regarded as living or once-living components that support human health, through the lens of the microbiome [46,47]. There are several types of biotics currently defined and well-recognized by the scientific community including Probiotics, Prebiotics, Synbiotics, and Postbiotics. However, there is often conflation among biotic innovations.
- Probiotics are defined as “Live microorganisms that, when administered in adequate amounts, confer a health benefit on the host.” [48]. Supplements that use probiotic ingredients for a health benefit must be formulated at a clinically relevant dose, with well-documented strains, delivered to ensure the appropriate viability of the strains at the end of shelf life. The majority of probiotics are intended for the intestinal tract, supporting gut and digestive health, with a few documented to have systemic effects working along the gut-organ axes. Of note, fermented foods are not probiotics and they have a separate definition [49].
- Prebiotics are defined as “A substrate that is selectively utilized by host microorganisms conferring a health benefit.” [50]. While prebiotics are commonly believed to be only fiber, this is not the case as prebiotic substances can be non-fiber, such as polyphenols, and certain types of oligosaccharides that are selectively metabolized. As with probiotics, prebiotics should be formulated at clinically studied doses for its intended effects. Unfortunately, the terms prebiotic and probiotic are widely misused, leading to incorrect perceptions that foods like onions, cereals, or garlic are prebiotic (without defining the compound, its amount and the dosage needed to confer a health benefit), and that for example any Lactobacillus or Bifidobacterium strain is a probiotic (without performing appropriate clinical studies to prove the strain’s efficacy and competitiveness).
- Synbiotics are defined as “A mixture comprising of live microorganisms and substrate(s) selectively utilized by host microorganisms that confers a health benefit on the host.” [51]. Synbiotics may be complementary, where the prebiotic ingredients are not targeting the co-administered probiotic strains, or synergistic, where the prebiotics are selectively utilized by the co-administered probiotic. The benefit of synbiotics, especially synergistic synbiotics, is that it promotes the survival of the bacterial strain that feeds on the prebiotic substrate in the host, and the health benefits of both ingredients may be amplified.
- Lastly, postbiotics are defined as “A preparation of inanimate microorganisms and/or their components that confers a health benefit on the host.” [52]. Recently, the International Probiotic Association (IPA) led a global effort to standardize the commercial definition of postbiotics, and further classified postbiotics into four distinct subcategories: (1) complex non-viable microbial preparations (CX; inactive microbial cells/fractions in unpurified culture medium), (2) intact non-viable microbial cells (IC; inactive whole microbial cells, separated from culture medium), (3) fragmented microbial cells (FC; fragmented microbial cells, separated from culture medium), and (4) microbial metabolic products (MM; metabolic products of microbial cells within their unpurified or partially purified culture medium) [53]. The global acceptance of the definition and these four subcategories remains to be achieved. While purified microbial metabolites have been purposely excluded by both ISAPP and IPA in their published definitions of postbiotics, many believe that they are also important considerations in this category. Nevertheless, postbiotics provide manufacturing benefits over probiotics since they do not contain viable microbial cells. However, they must still be clinically validated and show health benefits on the intended host. Interestingly, postbiotics do not have to be derived from a clinically validated probiotic strain, so long as the postbiotic preparation has a proven health benefit.
The four biotic categories listed above are either seeding or feeding the microbiome, or in the case of postbiotics, are derived from the microbes to provide health benefits. In essence, these interventions are highly targeted and specific in function; either supplying the carbon-containing energetic substrates for microbial metabolism, introducing specific microbes into the host microbiome, or administering inactivated bacterial cells/fragments or metabolic end products to confer a clear benefit. In simplified terms, biotic supplements directly support the microbiome (and indirectly/consequently, the host) while non-biotic supplements directly support the human host. This current construct fails to provide a definition for a class of compounds that can be directly utilized by - and supports the function of - both microbes and human cells simultaneously. To resolve this gap in supplement health solutions, we propose a new category within biotics termed co-biotics.
As will be presented below, the term co-biotics recognizes that microbial and host systems are not independent entities but operate within a shared biochemical and physiological continuum, thereby expanding the scope of microbiome-directed interventions and offering a unifying framework for supporting health in living species.
Co-Biotics
A New Category of Biotics
The term co-biotic was used in a 2014 case study of a patient with gastrointestinal dysbiosis. The term was not defined, and the product comprised prebiotic inulin, an antioxidant blueberry pomace extract, and an oat preparation of purified beta-glucan, and was designed to restore gut dysbiosis [54]. Companies have also used co-biotic as a brand, some containing yeast, others having multivitamins or probiotics, plus dietary compounds such as sugar or starch along with artificial colour and preservatives.
In order to bring some order to the field, we propose that co-biotics be defined as “A substrate comprising of bioactive molecular compounds that, when delivered to permit both host metabolism and microbial accessibility, simultaneously modulate biological processes in both the host and its resident microbiota, to confer targeted health benefits” (Table 1, Figure 1).
A co-biotic does not include living microbes. Co-biotic ingredients can include vitamins, minerals, and other compounds that are known to act on both human and microbial processes. This is distinct from prebiotics, where the host health outcome is a direct result of the targeted microbial changes; the host effect in co-biotics does not necessarily require microbiome modulation or metabolism as a precursor step, and may be a distinct benefit from the microbial target. Co-biotic ingredients target health functions that are co-regulated by these processes, such as nutrient absorption and energy production. Careful selection of co-biotic ingredients in a product therefore allows for comprehensive support of whole-body health compared to traditional host-only interventions. This recognizes the importance of the microbiome and its interaction with ingested compounds.
Co-biotics at the product-level must be formulated with an ingredient that is a co-biotic, and importantly, be delivered in a way that ensures access by both host and microbiome cells. The product should support one or more specific health benefits in a targeted way.
Co-Biotic Examples
Co-biotic ingredients can be derived from various sources, but do not include microorganisms (i.e., probiotics). One example of a co-biotic ingredient is ginseng; such as Panax quinquefolius (American ginseng) or Panax ginseng (Asian ginseng) (Figure 2). American and Asian ginseng has been clinically proven to provide various host benefits in brain health, particularly cognitive functions related to working memory and attention, as well as certain aspects of mood [55,56,57,58]. Mechanistically, ginsenosides, the primary active component of ginseng, have been shown to directly affect neurotransmission in host cells, regulate neuronal ion channels, and provide neuroprotective effects through antioxidant and anti-inflammatory abilities [59,60,61]. Ginsenosides’ interaction with the gut microbiota is also well established. Gut microbes are capable of bioconverting primary ginsenosides into more bioavailable and bioactive secondary forms of ginsenosides for host benefit [62]. In addition, ginseng administration has been shown both preclinically and clinically to significantly upregulate beneficial microbes such as Bifidobacterium and Lactobacillus species and increase production of beneficial microbial metabolites such as SCFA [55,63].
Additional examples of co-biotic ingredients are quercetin, a flavonoid found in many fruits and vegetables, and resveratrol, a polyphenol that is abundant in the skin and seeds of grapes and select berries. Quercetin supplementation has been shown to directly contribute to a diverse range of host benefits, including for the cardiovascular, immune, and muscular systems. For instance, human studies have shown that quercetin supplementation can reduce blood pressure, inflammation, and increase muscle strength [64,65,66]. Mechanistically, quercetin has shown direct anti-inflammatory effects by inhibition of inflammatory cytokine production in host cells and exerts its antioxidant properties by directly regulating and upregulating host glutathione, heme-oxygenase-1, and superoxide dismutase-1 levels [67,68]. Quercetin has a direct impact on the flavonoid-degrading bacteria of the gut microbiome, such as Eubacterium ramulus and Clostridium orbiscindens. One study found that healthy adults consuming a flavonoid-free diet displayed decreased total fecal bacteria, including a 40-75% decrease in E. ramulus. Supplementation with quercetin increased total bacterial counts, including E. ramulus to levels higher than baseline [69]. Notably, E. ramulus is a beneficial microbe known to produce the important SCFA butyrate (which help colonocyte integrity), and work cooperatively with other health-promoting microbes in the gut [70]. C. orbiscindens has been shown to convert quercetin to metabolites such as 3,4-dihydroxyphenylacetic acid, and while its clinical significance has yet to be established, C. orbiscindens is prevalent in the population and likely significantly contributes to gut microbial metabolism of quercetin [71].
Resveratrol has been clinically shown to have a variety of health benefits in areas of metabolic, cardiovascular, and immune systems [72,73,74]. Specifically, resveratrol reduces insulin resistance and inflammatory biomarkers and increases endothelial functions. Mechanistically, resveratrol acts on host cells by activating SIRT1, a deacetylase that is a master regulator in various cellular processes. Resveratrol-dependent SIRT1 activation has been shown to increase mitochondrial function and turnover, protect against cellular oxidative stress, and inhibit inflammatory responses [75,76,77]. Resveratrol can be directly metabolized by certain gut microbes, and several metabolic outputs such as lunularin, 3,4-dihydroxy-trans-stilbene, and resveratroloside, have been identified and demonstrated to have beneficial impacts on gut health [78,79,80,81]. Furthermore, preclinical studies have shown that resveratrol supplementation can enrich beneficial microbes such as Lactobacillus and Bifidobacterium species, while potentially decreasing trimethylamine (TMA)-producing microbes (associated with cardiovascular disease) by remodeling the gut microbiome [82,83].
Vitamins, minerals, and co-factors such as some B vitamins, may act as co-biotic ingredients. The human health benefits of these essential B vitamins have been long established, and their impact on energy metabolism, skin, vision, cardiac function, and nervous system are well substantiated [84,85]. B vitamins are also essential and play a vital role in microbial physiology and ecology. Many bacteria are able to biosynthesize B vitamins, including thiamin (vitamin B1), riboflavin (vitamin B2), niacin (vitamin B3), pantothenic acid (vitamin B5), pyridoxine (vitamin B6), biotin (vitamin B7), folate (vitamin B9), and cobalamin (vitamin B12) [86]. These vitamins are not only involved in several microbial processes, but the presence of auxotrophs can create competition and affect the overall microbial diversity and ecology. How oral vitamin supplementation impacts the gut microbiome is only recently being better understood; research suggests both direct effects on microbial composition as well as indirect effects via alteration of the host physiology (e.g., to the intestinal lumen) can impact resident microbes [87]. Microbiome-targeted vitamins, including B1 and B2, can increase microbial diversity, enrich for beneficial taxa, and promote the production of SCFA [87,88,89,90,91].
Analysis of the gut microbiome in human diseases is shedding light into the deep connectivity of microbes and the host. For instance, a meta-analysis of Parkinson’s disease (PD) across 5 countries identified a decrease in SCFA-producing species, such as Faecalibacterium prausnitzii, as well as a decrease in fecal microbial riboflavin (vitamin B2) biosynthetic genes in PD patients [92]. In addition, the decrease in these genes correlated with decreases in fecal SCFA. While the SCFA-producing species are also riboflavin producers, the lack of microbiota-accessible riboflavin is believed to have a negative impact on the overall gut microbiome, reducing total SCFA production, which can subsequently lead to increased intestinal permeability. Increased intestinal permeability has been proposed as a signature of PD that may aggravate PD pathology via the gut-brain axis [93]. Interestingly, a preclinical study showed that supplementation of colon-targeted microencapsulated riboflavin increases SCFA-producers, as well as SCFA production [89].
Together, these examples illustrate how co-biotic ingredients extend beyond host-only interventions by simultaneously nourishing microbial ecosystems and supporting systemic health, underscoring their potential as a more comprehensive framework for health solutions.
Delivery Mechanisms
Ensuring Microbiome Delivery
An important component of a co-biotic is that it must not only be absorbed by the host but must also be accessible by the gut microbiome, which in highest abundance resides in the colon. Active ingredients delivered via food matrices, in contrast to traditional supplements, have very different absorption profiles and bioavailability. For instance, food matrices may provide synergistic or carrier-like effects for certain active components of food that are lacking in supplement forms [94]. Food matrices may also contain absorption inhibitors not present in supplement formats [95]. For certain fast-absorbing actives, their delivery in traditional supplement forms has indirectly ignored distal segments of the microbiome, by not allowing the colonic microbes access to these active compounds due to their faster absorption in the upper GI tract. While the distal small intestine does indeed house a large number of microbes, the vast majority of the gut microbiota are strict anaerobes that reside in the colon [96,97]. Co-biotics that target these taxa should ensure availability in the colon.
Additional parameters such as processing techniques (e.g., spray drying), excipients, and other active ingredients all impact bioavailability, which influences ingredient availability for both the host and microbiome. While some compounds, likely polyphenols, saponins, and tannins, are able to reach the colon without special delivery mechanisms due to their poor bioavailability, other compounds, like essential vitamins, will be readily and quickly absorbed in the small intestine, preventing utilization by colonic microbes [98,99,100]. In order to ensure efficient delivery to the colonic microbiome if the colon is an essential target, the co-biotic ingredient’s absorption profile should be considered. In cases where ingredients are absorbed or degraded prior to reaching the colon, appropriate targeted delivery systems may be required for the ingredient to function as a co-biotic.
Several colon-targeted oral delivery systems including modified capsules, microencapsulation with various polymers, and synthesis of ingredients into prodrugs, can be utilized for co-biotic formulations. A popular form of a modified capsular delivery system is the dual capsule system consisting of an inner and outer capsule, where the inner capsule ingredients exhibit a delayed release profile; modifications of the capsule composition can further alter the release profile to have a more specific targeted effect on the colonic environment (Figure 3). For single layer capsules, enteric coating can provide targeted delivery of ingredients into the colonic space. One of the more popular enteric coatings for drug delivery are polymethacrylate-based copolymers, which is also available for dietary supplement applications [101]. Vitamins delivered in these types of copolymers have successfully been shown to induce microbiome and microbiome-related changes correlating with a healthy ecosystem [90].
Probiotics are commonly microencapsulated to ensure delivery of viable cells to the colonic space, and several companies have developed microencapsulation technologies [102]. Microencapsulation generally utilizes polysaccharides such as sodium alginate, lipid polymers, or other natural polymers like shellac, and delays release of the ingredients to the colon, allowing for targeted microbiome effects [103,104,105,106]. Microencapsulation is an effective way to deliver co-biotic ingredients to the distal small intestine or the colon.
Prodrugs are inactive or partially inactive forms of an active ingredient, that are designed to undergo biotransformation to release the active ingredient at a specific site in the GI tract [107,108]. Prodrug systems can be designed for targeted colonic delivery. Tributyrin is a prodrug of butyrate, with the butyric acid esterified to a glycerol backbone. Butyrate chains are released from the glycerol backbone through intestinal lipases. In preclinical studies, tributyrin has been shown to have better colonic delivery of butyrate than traditional butyrate salts [109]. Ultimately, effectiveness of a co-biotic depends both on the selection of the active ingredient and on ensuring its delivery to the targeted segment of the microbiome, whether it be the small intestine of the colon.
Conclusions
As the scientific understanding of human health advances, so too must the frameworks through which interventions are conceptualized. Co-biotics represent an emerging paradigm that explicitly accounts for the bidirectional and interdependent relationship between the human host and its microbiome. This construct moves beyond single-domain targeting to acknowledge that sustained health outcomes require engagement of shared biochemical and/or physiological processes across host and microbial systems. Current and future research efforts will continue to characterize precise mechanisms of action by which compounds exert co-biotic effects, as well as to identify additional agents capable of modulating these dual pathways. Parallel innovation in delivery systems will also be essential to ensure that such interventions achieve appropriate localization, both at sites of host absorption and within the relevant niches of the gastrointestinal microbiome. Collectively, these developments hold the potential to vastly expand the scope of nutritional and therapeutic strategies, as well as their health promoting capacity, establishing co-biotics as a unifying framework for next-generation microbiome-host interventions.
Acknowledgments
The authors would like to thank Marco Pane, Mizue Naito, and Tye Jensen for their perspectives on this review.
Conflicts of Interest
The authors have no conflicts of interest other than JV and GR being scientific advisors for Seed Health.
References
- Davenport, ER; et al. The human microbiome in evolution. BMC Biol 2017, 15, 127. [Google Scholar] [CrossRef]
- Moeller, AH; et al. Cospeciation of gut microbiota with hominids. Science 2016, 353, 380–382. [Google Scholar] [CrossRef]
- Suzuki, TA; et al. Codiversification of gut microbiota with humans. Science 2022, 377, 1328–1332. [Google Scholar] [CrossRef]
- Ahlawat S, Asha null, Sharma KK. Gut-organ axis: a microbial outreach and networking. Lett Appl Microbiol 2021, 72, 636–668. [Google Scholar] [CrossRef]
- Sender R, Fuchs S, Milo R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol 2016, 14, e1002533. [Google Scholar] [CrossRef]
- Wang, J; et al. Gut-microbiota-derived metabolites maintain gut and systemic immune homeostasis. Cells 2023, 12, 793. [Google Scholar] [CrossRef] [PubMed]
- Loh, JS; et al. Microbiota-gut-brain axis and its therapeutic applications in neurodegenerative diseases. Signal Transduct Target Ther 2024, 9, 37. [Google Scholar] [CrossRef]
- Morais LH, Schreiber HL, Mazmanian SK. The gut microbiota-brain axis in behaviour and brain disorders. Nat Rev Microbiol 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Li, Z; et al. Butyrate reduces appetite and activates brown adipose tissue via the gut-brain neural circuit. Gut 2018, 67, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H; et al. Depletion of acetate-producing bacteria from the gut microbiota facilitates cognitive impairment through the gut-brain neural mechanism in diabetic mice. Microbiome 2021, 9, 145. [Google Scholar] [CrossRef]
- Bravo, JA; et al. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci U S A 2011, 108, 16050–16055. [Google Scholar] [CrossRef]
- Bs, S; et al. Evaluation of GABA production and probiotic activities of Enterococcus faecium BS5. Probiotics Antimicrob Proteins 2021, 13, 993–1004. [Google Scholar] [CrossRef]
- Li, J; et al. Effects of Bifidobacterium breve 207-1 on regulating lifestyle behaviors and mental wellness in healthy adults based on the microbiome-gut-brain axis: a randomized, double-blind, placebo-controlled trial. Eur J Nutr 2024, 63, 2567–2585. [Google Scholar] [CrossRef]
- Yano, JM; et al. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef]
- Chong, HX; et al. Lactobacillus plantarum DR7 alleviates stress and anxiety in adults: a randomised, double-blind, placebo-controlled study. Benef Microbes 2019, 10, 355–373. [Google Scholar] [CrossRef]
- Walden, KE; et al. A randomized controlled trial to examine the impact of a multi-strain probiotic on self-reported indicators of depression, anxiety, mood, and associated biomarkers. Front Nutr 2023, 10, 1219313. [Google Scholar] [CrossRef]
- Wallace CJK, Milev RV. The efficacy, safety, and tolerability of probiotics on depression: clinical results from an open-label pilot study. Front Psychiatry 2021, 12, 618279. [Google Scholar] [CrossRef] [PubMed]
- Wang, H; et al. Bifidobacterium longum 1714TM strain modulates brain activity of healthy volunteers during social stress. Am J Gastroenterol 2019, 114, 1152–1162. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Sanchez, M; et al. The gut-skin axis: a bi-directional, microbiota-driven relationship with therapeutic potential. Gut Microbes 2025, 17, 2473524. [Google Scholar] [CrossRef]
- Mahmud, MR; et al. Impact of gut microbiome on skin health: gut-skin axis observed through the lenses of therapeutics and skin diseases. Gut Microbes 2022, 14, 2096995. [Google Scholar] [CrossRef] [PubMed]
- Buhaș, MC; et al. Transforming psoriasis care: probiotics and prebiotics as novel therapeutic approaches. Int J Mol Sci 2023, 24, 11225. [Google Scholar] [CrossRef] [PubMed]
- Manzhalii E, Hornuss D, Stremmel W. Intestinal-borne dermatoses significantly improved by oral application of Escherichia coli Nissle 1917. World J Gastroenterol 2016, 22, 5415–5421. [Google Scholar] [CrossRef]
- Togawa, N; et al. Improvement of skin condition and intestinal microbiota via Heyndrickxia coagulans SANK70258 intake: A placebo-controlled, randomized, double-blind, parallel-group comparative study. Nutr Burbank Los Angel Cty Calif 2024, 126, 112533. [Google Scholar] [CrossRef]
- Campagnoli, LIM; et al. Targeting the gut-eye axis: an emerging strategy to face ocular diseases. Int J Mol Sci 2023, 24, 13338. [Google Scholar] [CrossRef]
- Dang AT, Marsland BJ. Microbes, metabolites, and the gut-lung axis. Mucosal Immunol 2019, 12, 843–850. [Google Scholar] [CrossRef]
- Hsu CL, Schnabl B. The gut-liver axis and gut microbiota in health and liver disease. Nat Rev Microbiol 2023, 21, 719–733. [Google Scholar] [CrossRef]
- Zhang, Y; et al. The gut-heart axis: unveiling the roles of gut microbiota in cardiovascular diseases. Front Cardiovasc Med 2025, 12, 1572948. [Google Scholar] [CrossRef]
- Salazar, AM; et al. Gut-bladder axis enters the stage: Implication for recurrent urinary tract infections. Cell Host Microbe 2022, 30, 1066–1069. [Google Scholar] [CrossRef]
- Tian, Y; et al. Fecal microbiota transplantation promotes hair growth through gut microbiome and metabolic regulation. Life Sci 2025, 379, 123887. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y. Exploring clues pointing toward the existence of a brain-gut microbiota-hair follicle axis. Curr Res Transl Med 2024, 72, 103408. [Google Scholar] [CrossRef] [PubMed]
- Ahn J, Hayes RB. Environmental influences on the human microbiome and implications for noncommunicable disease. Annu Rev Public Health 2021, 42, 277–292. [Google Scholar] [CrossRef]
- Brown, K; et al. Diet-induced dysbiosis of the intestinal microbiota and the effects on immunity and disease. Nutrients 2012, 4, 1095–1119. [Google Scholar] [CrossRef]
- Vich Vila, A; et al. Impact of commonly used drugs on the composition and metabolic function of the gut microbiota. Nat Commun 2020, 11, 362. [Google Scholar] [CrossRef]
- Onali, T; et al. Berry supplementation in healthy volunteers modulates gut microbiota, increases fecal polyphenol metabolites and reduces viability of colon cancer cells exposed to fecal water- a randomized controlled trial. J Nutr Biochem 2025, 141, 109906. [Google Scholar] [CrossRef]
- Bhardwaj, RL; et al. An alarming decline in the nutritional quality of foods: the biggest challenge for future generations’ health. Foods 2024, 13, 877. [Google Scholar] [CrossRef]
- Abdelsalam, NA; et al. Toxicomicrobiomics: the human microbiome vs. pharmaceutical, dietary, and environmental xenobiotics. Front Pharmacol 2020, 11, 390. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B; et al. Randomized controlled-feeding study of dietary emulsifier carboxymethylcellulose reveals detrimental impacts on the gut microbiota and metabolome. Gastroenterology 2022, 162, 743–756. [Google Scholar] [CrossRef] [PubMed]
- Lindell AE, Zimmermann-Kogadeeva M, Patil KR. Multimodal interactions of drugs, natural compounds and pollutants with the gut microbiota. Nat Rev Microbiol 2022, 20, 431–443. [Google Scholar] [CrossRef]
- Whelan, K; et al. Ultra-processed foods and food additives in gut health and disease. Nat Rev Gastroenterol Hepatol 2024, 21, 406–427. [Google Scholar] [CrossRef]
- Madison A, Kiecolt-Glaser JK. Stress, depression, diet, and the gut microbiota: human-bacteria interactions at the core of psychoneuroimmunology and nutrition. Curr Opin Behav Sci 2019, 28, 105–110. [Google Scholar] [CrossRef]
- Neroni, B; et al. Relationship between sleep disorders and gut dysbiosis: what affects what? Sleep Med 2021, 87, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wegierska, AE; et al. The connection between physical exercise and gut microbiota: implications for competitive sports athletes. Sports Med Auckl NZ 2022, 52, 2355–2369. [Google Scholar] [CrossRef]
- Weersma RK, Zhernakova A, Fu J. Interaction between drugs and the gut microbiome. Gut 2020, 69, 1510–1519. [Google Scholar] [CrossRef] [PubMed]
- Javdan, B; et al. Personalized mapping of drug metabolism by the human gut microbiome. Cell 2020, 181, 1661–1679.e22. [Google Scholar] [CrossRef]
- Purdel, C; et al. Exploring the potential impact of probiotic use on drug metabolism and efficacy. Biomed Pharmacother Biomedecine Pharmacother 2023, 161, 114468. [Google Scholar] [CrossRef]
- Nataraj, BH; et al. Evolutionary concepts in the functional biotics arena: a mini-review. Food Sci Biotechnol 2021, 30, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Revankar NA, Negi PS. Biotics: An emerging food supplement for health improvement in the era of immune modulation. J Parenteral Ent Nutr 2024, 39, 311–329. [Google Scholar] [CrossRef]
- Hill, C; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Marco, ML; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on fermented foods. Nat Rev Gastroenterol Hepatol 2021, 18, 196–208. [Google Scholar] [CrossRef]
- Gibson, GR; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Swanson, KS; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat Rev Gastroenterol Hepatol 2020, 17, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat Rev Gastroenterol Hepatol 2021, 18, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Guglielmetti, S; et al. Commercial and regulatory frameworks for postbiotics: an industry-oriented scientific perspective for non-viable microbial ingredients conferring beneficial physiological effects. Trends Food Sci Technol 2025, 163, 105130. [Google Scholar] [CrossRef]
- Greenway F, Wang S, Heiman M. A novel cobiotic containing a prebiotic and an antioxidant augments the glucose control and gastrointestinal tolerability of metformin: a case report. Benef Microbes 2014, 5, 29–32. [Google Scholar] [CrossRef]
- Bell, L; et al. A randomized, placebo-controlled trial investigating the acute and chronic benefits of American Ginseng (Cereboost®) on mood and cognition in healthy young adults, including in vitro investigation of gut microbiota changes as a possible mechanism of action. Eur J Nutr 2022, 61, 413–428. [Google Scholar] [CrossRef]
- Dormal, V; et al. Effect of hydroponically grown red Panax Ginseng on perceived stress level, emotional processing, and cognitive functions in moderately stressed adults: a randomized, double-blind, placebo-controlled study. Nutrients 2025, 17, 955. [Google Scholar] [CrossRef]
- Reay JL, Scholey AB, Kennedy DO. Panax ginseng (G115) improves aspects of working memory performance and subjective ratings of calmness in healthy young adults. Hum Psychopharmacol 2010, 25, 462–471. [Google Scholar] [CrossRef]
- Scholey, A; et al. Effects of American ginseng (Panax quinquefolius) on neurocognitive function: an acute, randomised, double-blind, placebo-controlled, crossover study. Psychopharmacol 2010, 212, 345–356. [Google Scholar] [CrossRef]
- Gong, L; et al. Neuroprotective mechanisms of ginsenoside Rb1 in central nervous system diseases. Front Pharmacol 2022, 13, 914352. [Google Scholar] [CrossRef]
- Radad K, Moldzio R, Rausch W-D. Ginsenosides and their CNS targets. CNS Neurosci Ther 2011, 17, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Xue, J-F; et al. Ginsenoside Rb1 promotes neurotransmitter release by modulating phosphorylation of synapsins through a cAMP-dependent protein kinase pathway. Brain Res 2006, 1106, 91–98. [Google Scholar] [CrossRef]
- Zhao, L; et al. The interaction between ginseng and gut microbiota. Front Nutr 2023, 10, 1301468. [Google Scholar] [CrossRef]
- Kim YK, Yum K-S. Effects of red ginseng extract on gut microbial distribution. J Ginseng Res 2022, 46, 91–103. [Google Scholar] [CrossRef]
- Edwards, RL; et al. Quercetin reduces blood pressure in hypertensive subjects. J Nutr 2007, 137, 2405–2411. [Google Scholar] [CrossRef]
- Mury, P; et al. Quercetin reduces vascular senescence and inflammation in symptomatic male but not female coronary artery disease patients. Aging Cell 2025, 24, e70108. [Google Scholar] [CrossRef]
- Nishikawa, T; et al. Quercetin ingestion alters motor unit behavior and enhances improvement in muscle strength following resistance training in older adults: a randomized, double-blind, controlled trial. Eur J Nutr 2025, 64, 117. [Google Scholar] [CrossRef]
- Sun, H-T; et al. Quercetin suppresses inflammatory cytokine production in rheumatoid arthritis fibroblast-like synoviocytes. Exp Ther Med 2021, 22, 1260. [Google Scholar] [CrossRef]
- Xu, D; et al. Antioxidant activities of quercetin and its complexes for medicinal application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef]
- Simmering, R; et al. The growth of the flavonoid-degrading intestinal bacterium, Eubacterium ramulus, is stimulated by dietary flavonoids in vivo. FEMS Microbiol Ecol 2002, 40, 243–248. [Google Scholar] [CrossRef]
- Rodriguez-Castaño, GP; et al. Bacteroides thetaiotaomicron starch utilization promotes quercetin degradation and butyrate production by Eubacterium ramulus. Front Microbiol 2019, 10, 1145. [Google Scholar] [CrossRef]
- Schoefer, L; et al. Anaerobic degradation of flavonoids by Clostridium orbiscindens. Appl Environ Microbiol 2003, 69, 5849–5854. [Google Scholar] [CrossRef]
- Delpino FM, Figueiredo LM. Resveratrol supplementation and type 2 diabetes: a systematic review and meta-analysis. Crit Rev Food Sci Nutr 2022, 62, 4465–4480. [Google Scholar] [CrossRef]
- Koushki M, Dashatan NA, Meshkani R. Effect of resveratrol supplementation on inflammatory markers: a systematic review and meta-analysis of randomized controlled trials. Clin Ther 2018, 40, 1180–1192.e5. [Google Scholar] [CrossRef]
- Mohammadipoor, N; et al. Resveratrol supplementation efficiently improves endothelial health: A systematic review and meta-analysis of randomized controlled trials. Phytother Res 2022, 36, 3529–3539. [Google Scholar] [CrossRef]
- Yang, X; et al. The role and mechanism of SIRT1 in resveratrol-regulated osteoblast autophagy in osteoporosis rats. Sci Rep 2019, 9, 18424. [Google Scholar] [CrossRef]
- Yun, J-M; et al. Resveratrol up-regulates SIRT1 and inhibits cellular oxidative stress in the diabetic milieu: mechanistic insights. J Nutr Biochem 2012, 23, 699–705. [Google Scholar] [CrossRef]
- Zhu, X; et al. Activation of Sirt1 by resveratrol inhibits TNF-α induced inflammation in fibroblasts. PloS One 2011, 6, e27081. [Google Scholar] [CrossRef]
- Dong, Z; et al. Discovery of an ene-reductase initiating resveratrol catabolism in gut microbiota and its application in disease treatment. Cell Rep 2025, 44, 115517. [Google Scholar] [CrossRef]
- Li, M; et al. Resveratrol and its derivates improve inflammatory bowel disease by targeting gut microbiota and inflammatory signaling pathways. Food Sci Hum Wellness 2022, 11, 22–31. [Google Scholar] [CrossRef]
- Prakash, V; et al. Resveratrol as a promising nutraceutical: implications in gut microbiota modulation, inflammatory disorders, and colorectal cancer. Int J Mol Sci 2024, 25, 3370. [Google Scholar] [CrossRef]
- Yao, M; et al. Gut microbiota composition in relation to the metabolism of oral administrated resveratrol. Nutrients 2022, 14, 1013. [Google Scholar] [CrossRef]
- Chen, M; et al. Resveratrol attenuates trimethylamine-N-oxide (TMAO)-induced atherosclerosis by regulating TMAO synthesis and bile acid metabolism via remodeling of the gut microbiota. mBio 2016, 7, e02210–e02215. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M; et al. Effect of a low dose of dietary resveratrol on colon microbiota, inflammation and tissue damage in a DSS-induced colitis rat model. J Agric Food Chem 2009, 57, 2211–2220. [Google Scholar] [CrossRef]
- EFSA panel on dietetic products, nutrition and allergies (NDA). Scientific opinion on substantiation of health claims related to thiamine and energy-yielding metabolism (ID 21, 24, 28), cardiac function (ID 20), function of the nervous system (ID 22, 27), maintenance of bone (ID 25), maintenance of teeth (ID 25), maintenance of hair (ID 25), maintenance of nails (ID 25), maintenance of skin (ID 25) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J 2009, 7, 1222. [Google Scholar] [CrossRef]
- EFSA Panel on dietetic products, nutrition and allergies (NDA). Scientific opinion on the substantiation of health claims related to riboflavin (vitamin B2) and contribution to normal energy-yielding metabolism (ID 29, 35, 36, 42), contribution to normal metabolism of iron (ID 30, 37), maintenance of normal skin and mucous membranes (ID 31, 33), contribution to normal psychological functions (ID 32), maintenance of normal bone (ID 33), maintenance of normal teeth (ID 33), maintenance of normal hair (ID 33), maintenance of normal nails (ID 33), maintenance of normal vision (ID 39), maintenance of normal red blood cells (ID 40), reduction of tiredness and fatigue (ID 41), protection of DNA, proteins and lipids from oxidative damage (ID 207), and maintenance of the normal function of the nervous system (ID 213) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J 2010, 8, 1814. [Google Scholar] [CrossRef]
- Hossain KS, Amarasena S, Mayengbam S. B Vitamins and their roles in gut health. Microorganisms 2022, 10, 1168. [Google Scholar] [CrossRef]
- Pham, VT; et al. Vitamins, the gut microbiome and gastrointestinal health in humans. Nutr Res N Y N 2021, 95, 35–53. [Google Scholar] [CrossRef]
- Park, J; et al. Dietary Vitamin B1 intake influences gut microbial community and the consequent production of short-chain fatty acids. Nutrients 2022, 14, 2078. [Google Scholar] [CrossRef]
- Steinert, RE; et al. In vitro validation of colon delivery of vitamin B2 through a food grade multi-unit particle system. Benef Microbes 2024, 16, 253–269. [Google Scholar] [CrossRef]
- Pham, VT; et al. Effects of colon-targeted vitamins on the composition and metabolic activity of the human gut microbiome- a pilot study. Gut Microbes 2021, 13, 1–20. [Google Scholar] [CrossRef]
- Mckirdy, S; et al. Micronutrient supplementation influences the composition and diet-originating function of the gut microbiome in healthy adults. Clin Nutr 2025, 51, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Nishiwaki, H; et al. Meta-analysis of shotgun sequencing of gut microbiota in Parkinson’s disease. NPJ Parkinsons Dis 2024, 10, 106. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, CB; et al. Increased intestinal permeability correlates with sigmoid mucosa alpha-synuclein staining and endotoxin exposure markers in early Parkinson’s disease. PloS One 2011, 6, e28032. [Google Scholar] [CrossRef]
- Pressman P, Clemens RA, Hayes AW. Bioavailability of micronutrients obtained from supplements and food: A survey and case study of the polyphenols. Toxicol Res Appl 2017, 1, 2397847317696366. [Google Scholar] [CrossRef]
- Noonan SC, Savage GP. Oxalate content of foods and its effect on humans. Asia Pac J Clin Nutr 1999, 8, 64–74. [Google Scholar] [CrossRef]
- Donaldson GP, Lee SM, Mazmanian SK. Gut biogeography of the bacterial microbiota. Nat Rev Microbiol 2016, 14, 20–32. [Google Scholar] [CrossRef]
- Yersin S, Vonaesch P. Small intestinal microbiota: from taxonomic composition to metabolism. Trends Microbiol 2024, 32, 970–983. [Google Scholar] [CrossRef]
- Goncalves, A; et al. Fat-soluble vitamin intestinal absorption: absorption sites in the intestine and interactions for absorption. Food Chem 2015, 172, 155–160. [Google Scholar] [CrossRef]
- Rose, RC. Intestinal absorption of water-soluble vitamins. Proc Soc Exp Biol Med 1996, 212, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Solnier, J; et al. A pharmacokinetic study of different quercetin formulations in healthy participants: a diet-controlled, crossover, single- and multiple-dose pilot study. Evidence Based Complement Altern Med 2023, 2023, 9727539. [Google Scholar] [CrossRef] [PubMed]
- Thakral S, Thakral NK, Majumdar DK. Eudragit: a technology evaluation. Expert Opin Drug Deliv 2013, 10, 131–149. [Google Scholar] [CrossRef]
- D’Amico, V; et al. Microencapsulation of probiotics for enhanced stability and health benefits in dairy functional foods: a focus on pasta filata cheese. Pharmaceutics 2025, 17, 185. [Google Scholar] [CrossRef]
- Del Piano, M; et al. Evaluation of the intestinal colonization by microencapsulated probiotic bacteria in comparison with the same uncoated strains. J Clin Gastroenterol 2010, 44 (Suppl 1), S42–46. [Google Scholar] [CrossRef]
- Fangmann, D; et al. Targeted microbiome intervention by microencapsulated delayed-release niacin beneficially affects insulin sensitivity in humans. Diabetes Care 2018, 41, 398–405. [Google Scholar] [CrossRef]
- Jia, M; et al. The construction of a double-layer colon-targeted delivery system based on zein-shellac complex and gelatin-isomaltooligosaccharide Maillard product: In vitro and in vivo evaluation. Food Res Int 2025, 200, 115477. [Google Scholar] [CrossRef] [PubMed]
- Prakoeswa, CRS; et al. Lactobacillus plantarum IS-10506 supplementation reduced SCORAD in children with atopic dermatitis. Benef Microbes 2017, 8, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Rautio, J; et al. Prodrugs: design and clinical applications. Nat Rev Drug Discov 2008, 7, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Sinha VR, Kumria R. Colonic drug delivery: prodrug approach. Pharm Res 2001, 18, 557–564. [Google Scholar] [CrossRef]
- Ye, Z; et al. Oral tributyrin treatment affects short-chain fatty acid transport, mucosal health, and microbiome in a mouse model of inflammatory diarrhea. J Nutr Biochem 2025, 138, 109847. [Google Scholar] [CrossRef]
Figure 1.
Categories of microbiome-directed interventions, spanning probiotics, prebiotics, synbiotics, postbiotics, and co-biotics.
Figure 1.
Categories of microbiome-directed interventions, spanning probiotics, prebiotics, synbiotics, postbiotics, and co-biotics.
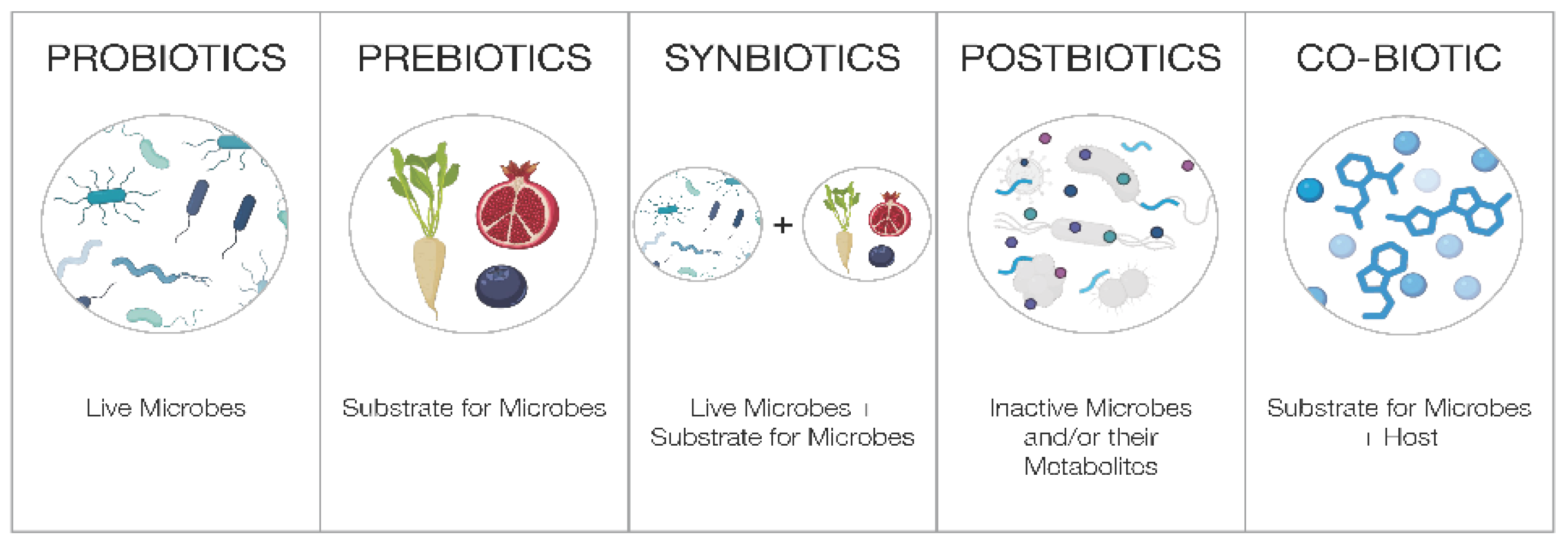
Figure 2.
Bi-directional interactions between ginseng, the gut microbiome, and the host. Ginsenosides can act directly on the host to positively impact cognition and mood. Ginsenosides are also metabolized by gut microbes into secondary ginsenosides and SCFAs which further influence host health.
Figure 2.
Bi-directional interactions between ginseng, the gut microbiome, and the host. Ginsenosides can act directly on the host to positively impact cognition and mood. Ginsenosides are also metabolized by gut microbes into secondary ginsenosides and SCFAs which further influence host health.
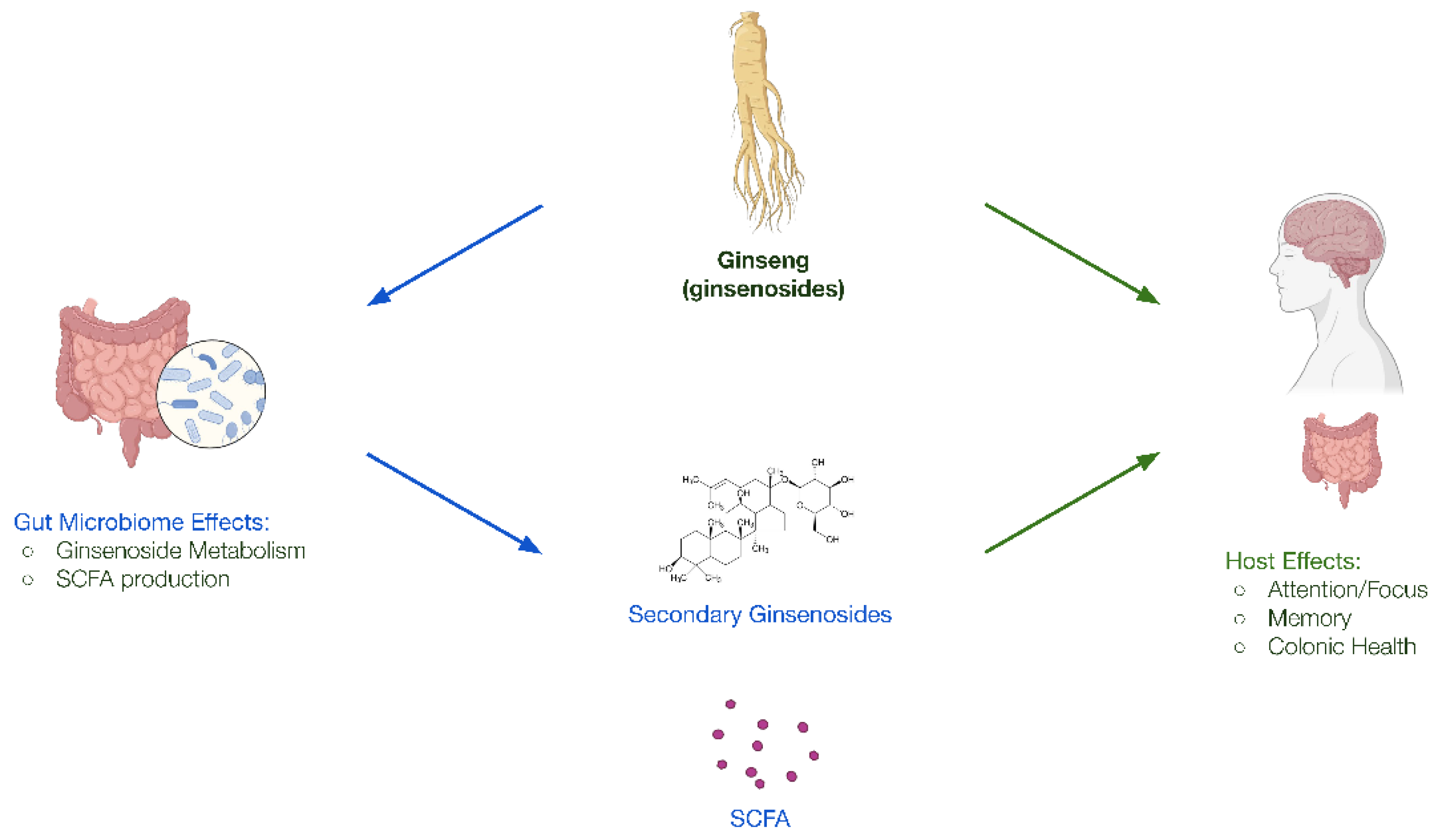
Figure 3.
Example of dual-capsule delivery technology for targeted delivery to the host and the colonic microbiome.
Figure 3.
Example of dual-capsule delivery technology for targeted delivery to the host and the colonic microbiome.
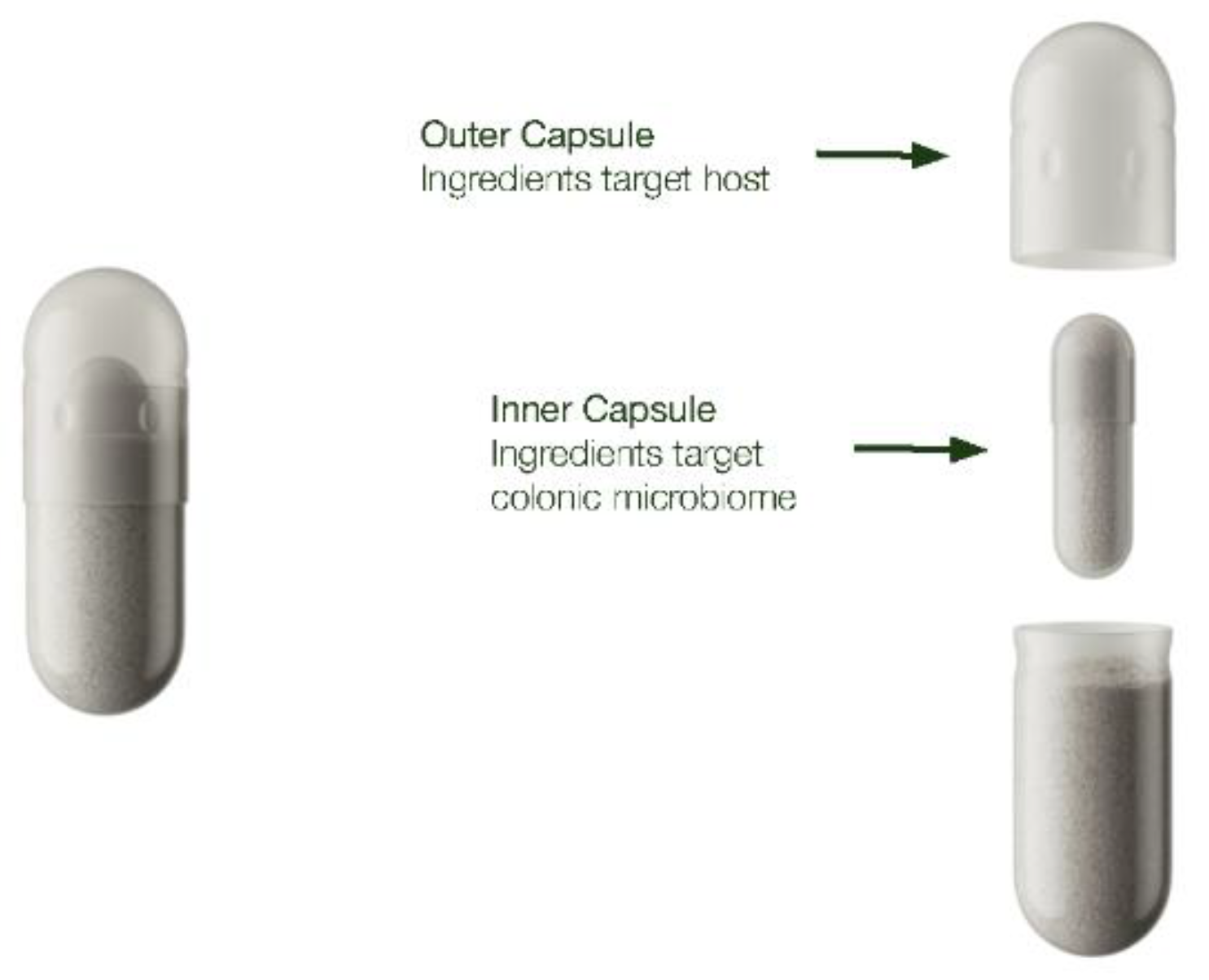

Table 1.
The Biotic Categories.
| Biotic Category | Definition | Examples |
|---|---|---|
| Probiotic | Live microorganisms that, when administered in adequate amounts, confer a health benefit on the host [45]. |
|
| Prebiotic | A substrate that is selectively utilized by host microorganisms conferring a health benefit [47]. |
|
| Synbiotic | A mixture comprising of live microorganisms and substrate(s) selectively utilized by host microorganisms that confers a health benefit on the host [48]. |
|
| Postbiotic | A preparation of inanimate microorganisms and/or their components that confers a health benefit on the host [49]. |
|
| Co-biotic | A substrate comprising of bioactive molecular compounds that, when delivered to permit both host absorption and microbial accessibility, simultaneously modulate biological processes in both the host and its resident microbiota, to confer a targeted health benefit |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



