Submitted:
22 September 2025
Posted:
24 September 2025
You are already at the latest version
Abstract
Table grapes suffer significant losses from issues such as fungal infections, cracking, and berry shattering, which affect them both in the vineyard during ripening and throughout postharvest storage. Current control methods, such as sulfur dioxide (SO2) treatments, are increasingly constrained by potential fruit damage and regulatory limitations, prompting a search for sustainable alternatives. This review proposes sorbitol as an innovative preharvest solution to enhance table grape quality through improved calcium (Ca2+) transport. Ca is a vital macronutrient for cell wall integrity and fruit resistance; however its inherent low mobility in the phloem restricts its effective delivery to developing fruits, particularly after veraison. Sorbitol, a naturally occurring sugar-alcohol, acts as a "vector" by forming stable, soluble complexes with Ca, thereby facilitating its crucial translocation to fruit tissues. Preharvest foliar applications of these calcium-sorbitol complexes have demonstrated numerous benefits-, improving fruit firmness, reducing the incidence of cracking and shattering, mitigating fungal decay, and boosting antioxidant activity. These effects collectively enhance overall fruit quality and extend storability. Future investigations will aim to further clarify the molecular mechanisms involved and explore sorbitol’s potential to complex with other poorly mobile nutrients and plant elicitors, opening new avenues for sustainable crop management.
Keywords:
preharvest treatment
; fruit ripening
; nutritional quality
; polyols
; postharvest
1. Introduction
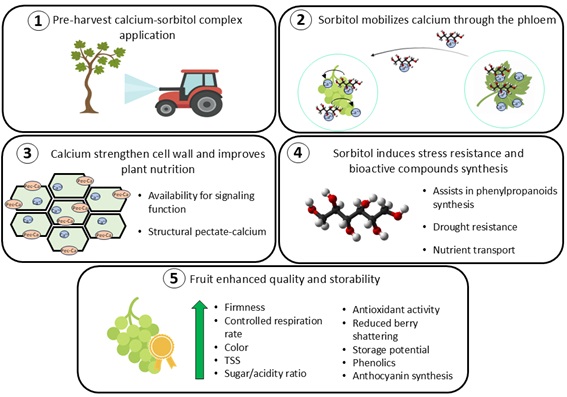
Grapes are the fruit of the Vitis genus which grows in clusters of berries. Table grapes are highly valued in the agri-food sector and appreciated by consumers for their complex flavour as well as its nutritional value given by many compounds such as sugars, organic acids, fibre, minerals, vitamins, flavones, flavanones, volatile compounds, pigments and, and anthocyanins in the case of red table grapes [1]. As a non-climacteric fruit, grapes are required to be harvested at an adequate maturity point. Otherwise, the accumulation of sugars, organic acids and pigments may not reach minimum requirements for commercialization, as well as the functional compounds related to human health such as polyphenols [2]. The synthesis of stilbenes, flavonoids, and phenolic acids proceeds through the phenylpropanoid pathway. Key branch-point enzymes involved in this pathway include phenylalanine ammonia-lyase (PAL), chalcone synthase (CHS), stilbene synthase (STS), anthocyanidin reductase (ANR), leucoanthocyanidin reductase (LAR), and UDP-glucose: flavonoid 3-O-glucosyltransferase (UFGT) [3]. These enzymes participate in the biosynthesis of phenolic compounds, including anthocyanins, which are directly associated with red pigmentation. However, table grapes face several challenges during ripening process and storage period that can result in significant economic losses and food waste. Given the growing demand for sustainable alternatives to current solutions such as hormone treatments (abscisic acid, gibberellins) or other chemicals like sulfur dioxide (SO₂), which pose environmental limitations and concerns, this review proposes and explores the potential of polyols, particularly sorbitol, as an innovative agricultural tool to enhance Ca transport in table grapes. It details how the complexation of Ca, an essential macronutrient with low mobility in the phloem and vital for cellular integrity and fruit resilience, with sorbitol can overcome this transport limitation, boosting its beneficial effects on fruit quality, shelf-life, and resistance to biotic and abiotic stresses, in alignment with sustainability goals.
2. Main Challenges in Table Grape Production and Postharvest
Major table grape issues include fungal infections, cracking, pedicel abscission, dehydration of berries and rachis, softening or colour loss [4]. These physiological disorders can disrupt plant cell homeostasis leading to an increase in enzymatic activity and the formation of reactive oxygen species (ROS), which affect the cell integrity and ultimately leading to a shortened shelf-life period for the grapes [5]. The current strategies used for controlling table grapes issues and ensuring fruit quality may be limited in the context of a global market that prioritizes food waste reduction and developing more sustainable strategies and practices for food production. The purpose of this review is to delve into the potential of sorbitol, Ca, and calcium-sorbitol complexes as sustainable solutions for enhancing the quality and shelf-life of table grapes.
2.1. Fungal Incidence and Decay
Postharvest decay of table grapes represents one of the primary causes of fruit loss reaching up to 40 % of world production [6]. The high moisture content of table grapes makes the fruit particularly susceptible to fungal infections, with grey rot caused by Botrytis cinerea being especially prevalent. Botrytis cinerea can proliferate under typical table grape storage conditions (0 °C to 4 °C) [7], as well as other fungal pathogens, including Alternaria alternata, Rhizopus oryzae, Colletotrichum acutatum, and Penicillium expansum [8,9,10,11]. Optimal temperature maintenance is mandatory to ensure quality of table grapes during transport, storage and commercialization, highlighting the need for fungi control in order to minimize fruit loss. As Ca is a key component in cell wall structure and a signalling agent, fruits with a Ca2+ deficiency are prone to fungal infestation [12]. Fungal infection control is commonly addressed with SO2 treatments [13] due to its antifungal properties. However, excessive SO2 exposure may cause fruit damage such as bleaching, discolouration, pitting and cracking [14].
2.2. Cracking
Fruit cracking devalues both commercial value and visual appeal of the grapes [15,16], while making the fruit vulnerable to biotic infections [17,18,19]. This disorder is defined as the ‘breacking, cracking, or splitting of the berry skin’ [20]. Cracking occurs during the third phase of grape development, when grapes begin to soften, sugars accumulate, and the rapid expansion of pulp cells exceeds the mechanical properties of the cuticle, leading to the cuticle’s incapability to sustain its integrity [20,21]. Several factors can lead fruit cracking: 1- Environmental factors such as rain and high humidity during developmental stages, especially during blossom or harvest are “serious limitations in profitable grape production”, high humidity and increasing temperature accelerate fruit cracking, with a linear increase in cracking as the temperature rises from 10 to 40 °C [20]; 2- The structure and composition of the cell wall from the pericarp directly influence mechanical properties and cracking vulnerability [15,22,23]; 3- Vegetal hormones, such as abscisic acid (ABA), auxin, giberellins (GA), ethylene and jasmonic acid are involved in fruit cracking [15,24]; 4- Nutrient deficiency , especially of Ca and Boron (B), can increase fruit cracking incidence as Ca is related to cell wall integrity [25], and its deficiency is tightly related to many disorders, and B has a role in cell wall development, cell division and sugar transport [20].
2.3. Berry Shattering
Berry drop (shattering) is a phenomenon where the berry detaches from the pedicel due to the degradation of pectins and celluloses along with the increased hydrolytic enzymes in the abscission zone [26]. Various factors can be attributed to shattering such as environmental conditions [25], genetic predisposition [27] or the use of growth regulators like ethephon and ABA [28,29]. Berry abscission happens in a specific location called the abscission zone (AZ) which is located between the pedicel and the berry. Cells from AZ differ physiologically from other cells and after specific signalling, generally enabled by plant hormones, hydrolytic enzymes are synthetized, degrading the cell wall and therefore causing abscission [30,31]. Shattering usually occurs during handling in harvest, transport, and storage. However, in extreme cases such as high temperatures (≥ 40 °C) during the fruit set, this phenomenon can occur during on-tree ripening, as was observed [32]. Berry shattering is a common issue during storage, transport, and commercialization of table grapes, causing important economic losses [31].
2.4. Colour Loss and Browning
Colour is highly determinant in consumer acceptance [33]. Browning during postharvest storage is a common issue in table grapes which that can be caused by enzymatic and non-enzymatic reactions [34]. Irregularities and lack of colour have been observed to be related with low expression of ripening-related genes involved in ABA biosynthesis [28,35] under high temperatures [36] and a lack of sunlight exposure [37] during ripening season.
2.5. Dehydration and Rachis Browning
Rachis development of colour is also indicative for table grapes storage suitability, as irregularities in size and browning of this part could indicate dehydration and a predisposition to advanced senescence [33]. For the consumer, a green rachis is a sign of freshness while a brown one indicates senescence or degradation. This perception is an “important cause of rejection on the consumer’s side and fruit loss” [38,39]. The rachis is the principal structure from which berries hang, representing approximately 4 % of the total mass of the grape cluster, but its respiration rate is 11 to 28 times higher than the berries [39,40]. Its high metabolic rate makes it particularly susceptible to dehydration and enzymatic browning during postharvest. Rachis browning is influenced by many factors that encompasses environment factors, grape physiology and agronomic handling, such as: 1- Water loss is “considered the main factor in rachis browning” [39,41]. Table grapes are highly sensitive to dehydration, especially the rachis, due to the absence of the epidermal layer and waxy cuticle present in berries [41]; 2- Low temperatures and high relative humidity are the most suitable conditions in order to reduce rachis browning and water loss [40,41]; 3- Packaging technologies such as sealed packages with low density polyethylene (LDPE) or microperforated film, are effective in reducing water loss and delaying browning [41]; 4- Biochemical and physiological processes, such as enzymatic browning, are associated to polyphenol oxidase (PPO) activity, which oxides phenolic compounds (the substrate) into quinones, causing the brown colour [40,41,42].
3. Current Strategies for Quality Control of Table Grapes
Contemporary strategies for maintaining table grape quality are multifaceted, encompassing both pre- and postharvest interventions. Preharvest protocols, such as girdling, a practice of removing a ring of bark from the trunk or canes to enhance berry size and hasten maturity, are widely employed. Furthermore, the preharvest application of plant growth regulators or elicitors can influence fruit development, colour, and resistance to disease. Postharvest, maintaining fruit integrity is crucial. Modified atmosphere packaging (MAP) is a primary technology used to slow respiration and senescence by altering the gas composition surrounding the fruit. The application of sulfur dioxide (SO2) remains a standard fungicidal treatment to control Botrytis cinerea (gray mold), though concerns over residue and potential bleaching have spurred research into alternatives. Promising new approaches include the application of natural compounds like essential oils, which exhibit antimicrobial properties. These can be used alone or in conjunction with other technologies, such as the application of 1-methylcyclopropene (1-MCP), a compound that inhibits ethylene action, or chitosan nanoemulsions, which form a protective, semi-permeable coating on the fruit surface. These combined strategies aim to extend shelf-life and preserve the sensory and nutritional qualities of table grapes.
3.1. Preharvest Strategies
3.1.1. Girdling
Girdling is an ancient and widely adopted horticultural practice in viticulture, particularly for table grapes, involving the precise removal of a narrow ring of bark, typically 3 to 6 mm in width, from the trunk or canes of the vine [43,44,45,46].This technique fundamentally alters the plant's physiological transport mechanisms by interrupting the phloem's downward flow of photosynthates, essential mineral nutrients, and plant hormones from canopy to the roots [45,46,47]. Consequently, these vital resources are redirected and accumulate in the plant parts situated above the girdle, significantly benefiting the developing fruit clusters and berries [45,46,47,48,49]. This localized accumulation leads to elevated concentrations of sugars, auxins (IAA), and abscisic acid (ABA) within the berries, which are crucial for fruit development and ripening [46]. As a preharvest strategy, girdling is primarily applied to achieve several commercial objectives, including promoting substantial increases in berry size and overall bunch weight, and accelerating the fruit's maturation process [21,44,47,48,50,51]. However, the effectiveness and outcomes of girdling are highly cultivar-dependent, sensitive to the precise timing of application, and influenced by environmental conditions, sometimes requiring considerable labour or posing potential physiological stress to the vine [44,49,51].
The impact of girdling on table grape quality extends across a diverse range of parameters, albeit with notable variability based on cultivar and application context. While girdling is often effective in deepening skin colour, especially in red varieties like 'Hanxiangmi' and 'Crimson Seedless', and can advance maturation [49,52,53,54], its influence on sugar content and acidity can be inconsistent. Some studies reported increased total soluble solids, while others observe a decrease in sugar content or an elevation in titratable acidity, for example in 'Italia' grapes [44,49,53,55,56]. Importantly, research indicates that girdling typically leads to a reduction in berry cuticle thickness, which may heighten the fruit's susceptibility to cracking, although certain Ca treatments appear to mitigate this adverse effect [21,57]. Furthermore, girdling can significantly modify fruit texture. Varieties such as 'Áron' and 'Melinda' have shown increased berry hardness and skin thickness postgirdling, presenting a pattern that deviates from the natural softening trend observed during normal ripening despite higher sugar levels [45]. In terms of aromatic profiles, girdling alone might reduce the overall concentration of volatile compounds, but when synergistically combined with other interventions like foliar potassium fertilization, it can markedly enhance the accumulation of key aromatic compounds, particularly terpenes, contributing to a more complex and desirable flavor in cultivars like 'Hanxiangmi' [49,56]. These complex and often cultivar-specific responses underscore the need for tailored application strategies to optimize table grape quality.
3.1.2. Hormone Applications
Preharvest application of elicitors represents a robust and eco-friendly strategy for enhancing the quality and yield of table grapes, addressing challenges such as fungal decay and suboptimal colouration [58,59,60,61]. Key elicitors include salicylic acid (SA) and its derivatives acetylsalicylic acid (ASA) and methyl salicylate (MeSA), as well as methyl jasmonate (MeJA) and oxalic acid (OA), all of which are naturally occurring plant compounds [62,63,64,65,66,67,68,69,70]. These compounds act as plant hormones, orchestrating various physiological processes, including defense responses and ripening [62,64,68,71,72]. Treatments with SA, ASA, and MeSA, particularly MeSA at 0.1 mM, have demonstrated significant efficacy in alleviating disease caused by Botrytis cinerea in cultivars like 'Crimson' and 'Magenta' [60]. This disease control is primarily mediated through the stimulation of the plant's antioxidant system, increasing the activity of enzymes such as ascorbate peroxidase (APX), catalase (CAT), and peroxidase (POD) [60,73]. Furthermore, these salicylate applications lead to higher total acidity, increased content of bioactive compounds (e.g., total phenolics and anthocyanins), and enhanced antioxidant activity in treated berries compared to controls [59,60]. Similarly, MeJA treatments, depending on concentration, can significantly impact grape ripening, with lower doses (e.g., 0.1 and 1 mM) accelerating maturation and increasing vine yield, while higher concentrations (e.g., 5 and 10 mM) may delay ripening and reduce yield [58,59]. This dual effect of MeJA underscores the importance of optimal concentration for desired agronomic and commercial outcomes, including improved berry size, firmness, and total soluble solids content [58].
The beneficial effects of elicitors extend beyond disease resistance, profoundly influencing various quality attributes of table grapes. Salicylates enhance the concentration of total phenolics and anthocyanins, which are crucial for pigmentation and possess significant antioxidant activity, contributing to the fruit's health benefits [49,59,60,65,66,67,69,70,74,75,76]. For example, preharvest salicylate treatments led to significantly higher concentrations of five identified anthocyanins (delphinidin-3-glucoside, cyanidin-3-glucoside, petunidin-3-glucoside, peonidin-3-glucoside, and malvidin-3-glucoside) in both 'Crimson' and 'Magenta' cultivars [59,60]. These treatments also boost the levels of important organic acids, such as ascorbic, succinic, and fumaric acids [67,77], with MeSA at 0.1 mM proving most effective in this regard [60]. The increased activity of antioxidant enzymes (APX, CAT, POD) induced by salicylates helps to scavenge reactive oxygen species (ROS), which accumulate under pathological and senescence conditions, thereby delaying ripening and senescence processes and maintaining berry quality, firmness, and colour during cold storage [59,60,73,77]. MeJA treatments at appropriate concentrations (e.g., 0.1 mM) not only accelerate berry ripening and improve crop yield but also increase total phenolics and individual anthocyanin concentrations, leading to enhanced homogeneous pigmentation and higher firmness and total soluble solids at harvest [58,78,79,80,81,82,83]. These effects translate into improved market value due to earlier harvests and fruit with superior organoleptic and nutritional properties [58].
Oxalic acid (OA), another natural compound, offers a distinct mechanism for enhancing table grape quality, particularly colour in poorly pigmented cultivars like 'Magenta' [61]. Preharvest OA treatment (e.g., 5 mM) significantly increases endogenous abscisic acid (ABA) content and ABA glucose ester (ABA-GE) in berries, a critical hormone that regulates ripening [61]. This elevation in ABA is mediated by the upregulation of the VvNCED1 gene, which plays a key role in ABA biosynthesis and subsequent anthocyanin accumulation [61,84,85,86]. The accumulation of ABA directly contributes to improved colour and higher individual and total anthocyanin content at harvest[59,61,78,87,88,89,90,91,92]. OA treatments also lead to significantly lower glucose and fructose content but higher tartaric, ascorbic, and succinic acids at harvest, indicating a modulated ripening profile [61]. Moreover, OA boosts the activity of antioxidant enzymes (APX, CAT, POD) during berry development and maintains their activity during postharvest storage, thereby delaying the loss of firmness and colour and reducing susceptibility to postharvest decay caused by Botrytis cinerea [61,62,64,91,93]. Compared to other elicitors, OA at 5 mM has shown particularly strong effects, for instance example, a 2.4-fold increase in skin total anthocyanins compared to a 1.5-fold increase with MeSA, making it a highly effective tool for improving colour and overall quality in low-pigmented table grapes [61].
3.2. Postharvest Strategies
3.2.1. Modified Atmosphere Packaging Technology
Several postharvest strategies addressing table grape issues have been considered. Among these, modified atmosphere packaging (MAP) is a convenient, minimally invasive method to maintain quality of grapes during storage [94,95,96,97,98]. In MAP technology, the package headspace is usually replaced with specific concentration of atmospheric gases such as O2, CO2, and N, as well as other compounds such as ozone (O3), ethanol or nitric oxide (NO). Low concentrations of O2 are used but not low enough to produce anoxia, and moderate concentration of CO2, aiming to slow down berries decay, reducing the respiration rate and hindering the proliferation of microorganisms, especially moulds. CO2 and O3 prestorage treatment atmospheres are useful to control berry decay as well as to maintain quality traits of the fruit. Admane et al. (2018) [5] successfully maintained table grape quality with CO2 and O3 prestorage treatments following 45 days of storage at 0 °C with MAP (2 % CO2 - 5 % CO2). However, CO2 treatment caused organoleptic quality loss and lower anthocyanin content whereas 20 μL L−1 O3 treatment enhanced anthocyanin content as well as maintained good flavour and visual appeal [5]. Ethanol at 2.25 ppm was found to retain berry firmness during storage period. Nevertheless, increased cracking incidence, acetaldehyde and ethyl acetate accumulation, which may lead to unpleasant aromas and off-flavours [13] while NO at 300 μL L⁻¹ has been showed induction of the expression of VvSOD, VvCAT, VvPOD2, and VvGR for 60 days at 0 °C, thereby activating antioxidant enzyme activity and alleviating oxidative damage caused by ROS [99].
3.2.2. Sulfure Dioxide Applications: Benefits and Limitations
SO2 has been widely used since 1925, playing a major role in table grapes preservation strategies [5,100] . It is stated that, more than just a biocide, SO2 has a metabolic effect, promoting the defense mechanism of the plant [101]. The observed mechanism of action suggest an elevated expression of genes related to the production of H2O2, thereby augmenting respiratory burst oxidase homolog and superoxide dismutase (SOD) enzymatic activity [102] as well as the upregulation of genes related to glutathione synthesis and cyclic pathways [101], while the genes associated with ethylene signalling and cell wall degradation were downregulated [103]. Additionally, other authors reported physiological effects such as higher firmness of vegetal tissue due to SO2 and H2O2 mediating effects on cell wall pectin demethylation [104], a process associated with key enzymes such as pectin methyl esterase, polygalacturonase, pectate lyase and β-galactosidase [15]. Considering the broad and extensive use of SO₂ in maintaining postharvest quality of table grapes during cold storage, SO2 packaging technologies such as perforated SO₂-generating plastic liners and SO₂-generating pads have been designed and evaluated to enable a controlled and sustained release of the gas throughout the storage period [105]. Although the use of SO2 can be effective in controlling berry decay, there are major limitations related to this treatment. SO2 injury can express as bleaching, discolouration and fruit pitting [14]. Also, European Regulation (EC) Nº 1333/2008 on food additives limits the usage of SO2 sulphites in fresh table grapes to 10 mg kg-1.
3.2.3. Alternative Solutions
Other treatments aimed at preserving grapes include active packaging with essential oils, such as tea tree oil [106], oregano essential oils [107], thymol, eugenol or carvacrol [108]. However, detrimental effects of effective doses of these essential oils make them perceptible at the time of consumption with persistent aromas from the essential oils and off-odors occasionally detected in the fruit. Thus, strategies using essential oils in addition to 1-MCP application [109] or chitosan nanoemulsion [110] are gaining popularity in order to reduce EO doses in postharvest treatments.
3.3. The Need for Sustainable and Innovative Solutions
As global retailers and large-scale production groups continue to expand, private and public regulations are expected to become more demanding in regulatory restrictions at both national and international levels, along with growing consumer concern [111]. This evolving regulatory and market environment highlights the need for alternative solutions that can maintain table grape quality and extend shelf-life without compromising sensory or physicochemical attributes, while also meeting environmental, economic, and social sustainability goals. In this context, preharvest treatments with polyols represent a promising approach to enhancing the quality, durability, and productivity of table grapes.
4. The Crucial Role of Ca in Fruit Quality at Harvest and Postharvest
4.1. Ca as a Structural Component, Signaling Agent, and Its Mobility in the Plant
Ca is an essential macronutrient that plays a multifaceted role in plant physiology. It is an integral component the the cell wall, where it helps to stabilize pectin molecules as calcium-pectate, contributing to the cell rigidity and integrity. This structural role is fundamental for maintaining turgor pressure, which is essential for cell expansion and overall plant growth. Furthermore, Ca has a key role in cellular function and communication, as it is involved in the stabilization of membranes, regulating their permeability and fluidity [112]. Nevertheless, a defining characteristic of Ca is its low mobility through the phloem [113,114,115]. This results in limited translocation of Ca from mature leaves to developing tissues, such as young leaves, fruits, and sink organs, and the potential occurrence of physiological disorders, such as blossom end rot (BER).
Ca ions (Ca2+) are integral to the signaling networks of eukaryotic cells, including plants. They act as secondary messengers that connect external stimuli to internal cellular responses. The concentration of cytosolic Ca is regulated by a complex system of Ca channels, pumps, and sensors, which maintain homeostasis and facilitate the generation of Ca signals in response stimulus, such as hormones, biotic and abiotic stresses, and developmental factors [116]. In response to external signals, an early event in the signaling process consists of a translocation of Ca²⁺ from extracellular space or from internal stores, where Ca²⁺ is concentrated, into the cytosol, raising the cytosolic free Ca²⁺. The specific characteristics of the resulting Ca signal, such as its frequency, amplitude, and waveform are shaped by both the type and intensity of the stress faced [117].
Signaling is regulated by elemental components that control the magnitude and duration of cytosolic Ca fluctuations [117]. Ca²⁺-permeable ion channels allow the influx of Ca²⁺ ions into the cytosol in response to drought stress. The opening of these channels is often triggered by various environmental factors, leading to an increase in intracellular Ca levels. Ca²⁺/protons (H⁺) antiporters help maintain Ca homeostasis by exchanging Ca²⁺ for H⁺ across the plasma membrane. This exchange is vital for regulating cellular pH and Ca concentrations during drought stress. Ca²⁺-ATPases actively pump Ca²⁺ out of the cytosol, helping to restore Ca homeostasis after a transient increase in Ca levels, which is essential for resetting the Ca signaling pathways and preparing the plant for subsequent stress events. The initial key elements of the Ca signaling are Ca²⁺ channels and Ca²⁺ sensor proteins. Most of these sensor proteins are characterized by the presence of EF-hands, a conserved helix-loop-helix structural domains that coordinate Ca²⁺ ions through an acidic loop region [118]. In plants, the main types of EF-hand containing Ca2+ binding proteins include calmodulins, calmodulin-like proteins, calcineurin B-like proteins, and calcium-dependent protein kinases (CDPKs) [119]. CDPKs play a significant role in regulating secondary metabolism through two main pathways. They phosphorylate enzymes directly involved in metabolic processes, such as PAL, and they modify transcription factors that control the expression of genes linked to secondary metabolite biosynthesis. PAL initiates the biosynthesis of phenylpropanoids by catalyzing the deamination of L-phenylalanine into trans-cinnamic acid and ammonia [120]. This enzyme is key to the formation of various polyphenolic compounds in plants, including flavonoids, lignins, and other phenylpropanoids, which contribute to structural integrity and stress responses. Under drought conditions ABA is synthesized, triggering a signaling chain in guard cells that lead to stomatal closure, thereby minimizing water loss. A critical step in this response is the elevation of cytosolic Ca²⁺ levels, facilitated by its flux through both plasma membrane and intracellular Ca²⁺ channels. This rise in Ca²⁺ is essential for stomatal closure, which is induced by ABA, and involves hyperpolarization-activated and voltage-dependent channels, some of which are modulated by ROS [117].
4.2. Physiological Disorders Associated with Ca Deficiency
Calcium deficiency disorders usually have multifactorial causes that lead to lower Ca2+ concentrations in a specific plant tissue, resulting in diseases triggered by the shortage of mineral. Among Ca2+ deficiency symptoms, bitter pit is commonly observed in apples [121,122] as dark circular lesions beneath the skin surface of the fruit. The affected spots are brown, dry, and porous texture, tending to appear as necrotic tissue, and are commonly located at the calyx area. In tomatoes and pepper, deficiency may lead to BER, showing a sunken dark area at the vegetables end [122,123]. In pineapples [124,125,126] and loquat [127,128], internal browning may appear with Ca deficiency, when fruit is stored at chilling injury-promoting temperatures. Mangoes tend to manifest spongy tissues when Ca nutrition is insufficient [129]. Nevertheless, preharvest exogenous foliar treatments with Ca solutions have proven to be an effective solution to alleviate these symptoms.
4.3. Effect of Ca Application on Fruit Quality
Foliar applications of Ca solutions are a common agricultural practice (Table 1). The application of sprayed Ca salts during fruit development limits the breakdown of cell walls, preventing softening and delaying fruit ripeness, as high presence of Ca inhibits polygalacturonase activity, an enzyme responsible of pectin degradation [130]. In peaches, some authors [131] observed higher accumulation of polyphenol content as well as antioxidant activity, but a lack of modifications in fruit firmness, sugars, and acids content which gives peaches their main quality attributes, whereas others reported a 34.2-44.7 % firmness increase in Ca treated canned peaches cv. ‘Andross’ [132]. Ca applications also proved to improve storability of peach fruits. Ca chloride (CaCl2) solutions at 1 % improved weight loss, cell membrane ion leakage (IL), and brown rot incidence as increased phenolic content, antioxidant activity, and enzymatic activity after 30 days of storage at 8 ± 2 °C, 50 % RH [133]. In ‘Fengtangly’ variety, CaCl2 applications delayed softening by regulating cell wall metabolism during storage at 4 ± 1 °C and 90 ± 5 % RH for 70 days [134]. In cherry fruit, applications of 1 mM of CaCl2 during tree dormancy effectively improved Ca nutrition, cracking incidence, and sweet cherry fruit quality characteristics [135]. When treatments applied 30 days before harvest, Ca caseinate, CaCl2, Ca hydroxide (Ca(OH)2),and Ca nitrate (Ca(NO3)2) were effective to reduce cracking, with CaCl2 and Ca(OH)2 being the most effective compounds, reducing cracking by 62 % and 66 % respectively but only CaCl2 improved cherry firmness, and none of Ca treatments modified sugars or organic acids [136].
4.4. Effect of Ca Application on Table Grapes Quality
Preharvest Ca treatments strengthened the cell wall of grape berries, retaining turgor and delaying cellular lipid catabolism, thereby prolonging shelf-life [141]. The activities of PG and PE enzymes have also been reduced, delaying pectin degradation and alleviating weight loss, decay incidence, MDA content, and relative conductivity [143].
A recent study indicates that Ca ascorbate (CaAs), a Ca salt containing ascorbic acid, “significantly improved berry firmness and reduced browning and colour change indices at harvest and during storage” in ‘Thompson Seedless’ table grapes [145]. It also significantly decreased weight loss and reduced berry abscission and decay spread rates during storage [145].
Cracking is “closely related to the strength and elongation of the peel” [147,148]. Changes in the composition of cell wall components and the activities of metabolic enzymes regulated by cell wall–related genes may affect the occurrence of fruit cracking [23,148,149,150]. Enzymes such as PG, PME, beta-galactosidase (β-Gal), cellulase (Cx), and expansins (EXP) are involved in cracking. Elevated activities of PG, β-Gal and Cx in genotypes prone to cracking indicate a role in the breakdown of the cell wall [15,23,151]. Ca treatments can reduce PG and Cx activities, slowing down pectin and cellulose degradation and resulting in higher peel integrity [15]. The cuticle is a “protecting barrier against external or internal stresses” and pathogens [18,152,153,154]. Cuticle thinning, especially during the last stages of fruit development makes it vulnerable to microcracks [18,155]. Preharvest foliar spraying of Ca is a promising strategy to reduce fruit cracking, as Ca is essential for strengthening and stabilizing the structure of the cell wall and membrane [20]. CaCl₂ has been shown, particularly at the flowering stage, to reduce cracking incidence by approximately 4-to-10 fold compared with the control group [15]. This effect is attributed to increased Ca absorption during this period, leading to higher Ca content in the peel, improved mechanical properties, fruit hardening, and reduced internal pressure [15]. A research study in ‘El-Bayadi’ table grapes showed that 1 % CaCl2 preharvest spraying “significantly reduced rotting incidence during storage compared to the control” [156]. CaCl2 also reduced the extracellular PG activity of Botrytis cinerea by up to 90 % [12,157]. Furthermore CaCl2 “had a direct influence on the conidia and hyphae of Botrytis cinerea, causing conidial malformation and cytoplasmic disorganization” [12].
Ca transport to the berries decreases drastically post-veraison due to a reduction in transpiration rate and xylem functionality [12,141]. This suggests that Ca deficiency in some grape varieties is more common than previously thought, and that the post-veraison period offers the greatest opportunity to artificially modify berry Ca content.
Foliar treatments were also effective to preserve postharvest life and quality. In ‘Thompson Seedless’ variety, concentrations of 5 and 10 % enhanced phytochemicals, including the berry total acids, total phenolics, flavonoids and carotenoids contents, and total antioxidant activity, as well as ascorbate peroxidase (APX), superoxide dismutase (SOD), and catalase (CAT) enzyme activities. They also decreased total soluble solids content, malondialdehyde (MDA), ion leakage (IL), and pH. After 2 months of storage at 1 ± 1 °C, treated fruit showed lower decay, weight loss, colour change, IL, and browning [142]. This advantage of spraying Ca on leaves was also previously observed [140]. Fruit treated with Ca(NO3)2 at 2 % and stored at 1 °C were found to have a higher activity of antioxidant enzymes (SOD, CAT, APX, and guaiacol peroxidase (GPX)), and lower weight loss, chilling injury (CI), MDA, IL, and fungal incidence compared to untreated fruit. In addition, combined treatments of Ca(NO3)2 at 2 % and zinc sulfate at 1 % achieved the highest performance. Combined treatments of CaCl₂ at 0.016 % with magnesium oxide (MgO) at 0.056 % significantly reduced weight loss, berry shattering, and decay percentage of ‘Flame seedless’ variety after 12 days at 10 °C [142]. Moreover, Ca applications were observed to stimulate genes related to the biosynthesis of anthocyanins in table grapes [158], facilitating the accumulation of antioxidant compounds.
In this way, it can be confirmed that the use of foliar Ca on fruit is a useful tool to strengthen the plant’s resistance and activate defense mechanisms against both biotic and abiotic stresses. In general, the authors report improvements in firmness, weight loss, phenolic content, antioxidant activity and enzymatic activity, while fruit quality parameters related to sugars and organic acids remain unaffected by Ca application alone. Due to calcium’s low mobility, applying sorbitol, which promotes translocation of nutrients through the plant can be an important strategy to enhance calcium’s optimal nutrition benefits, as well as to boost the synergistic effects of sorbitol, in a suitable nutrient-polyol preharvest treatment.
5. Polyols as Physiological Tools and Nutrients Vector
Polyols, or sugars alcohol, are sugar-like carbohydrates that mainly have hydroxyl groups (-OH) groups attached. Polyols are widely used in food products, are Generally Recognised as Safe (GRAS) by the U.S. Food and Drug Administration and are included in the European Food Safety Agency food additive list (EC Nº 1333/2008). By their structural complexity, they can be found in monosaccharides forms (sorbitol, mannitol, and xylitol), disaccharides (maltitol and isomalt) and oligosaccharides, such as hydrogenated starch hydrolysates [159,160].
Furthermore, polyols have been repeatedly stated to be naturally present in higher plants [161]. They are a reduced form of ketose and aldose sugars, being sorbitol, mannitol, and galactitol the most abundant in plants. Besides the common hexitol structure among these three polyols, only mannitol and sorbitol are photosynthesis products, as is sucrose. Polyols are related to primary metabolism and possess several physiological functions, including nutrient transportation, energy source, and signalling modulation, increasing tolerance against drought, saline stress, and both biotic and abiotic stresses [162]. Salinity is one of the major environmental challenges to plants, as it is known to repress plant development and production in the first instance, followed by osmotic stress and water loss caused by the severe ion imbalance produced by Na+ and Cl- accumulation. In response, plants develop a succession of metabolic mechanisms in order to restore homeostasis, which encompasses the production of antioxidant enzymes, polyamines, NO, and osmoprotectants such as sorbitol [162].
5.1. Sorbitol: Properties and Role in Plant Physiology
Sorbitol is a 6-carbon sugar-alcohol derived from its sugar aldose, glucose. This polyol can naturally be found in fruits from Rosacea family, such as apples [163], pears [164], nectarines [165], apricots [166], plums [167], or peaches [168], as well as other plant species in lower contents [169]. Besides the known role of sorbitol as a carbon source in apples, pears peaches, apricots and cherries among other fruits, enzymes and genes of sorbitol metabolism has been found in other species initially not regarded as sorbitol-producers, like table grapes [170], as well as physiological effects when sorbitol is externally applied in blood oranges [171,172]. Sorbitol is photosynthesized mainly in matured leaves, in contrast to younger ones, which do not have enough photosynthetic capability to fully generate this metabolite, as well as sucrose [173]. In fruit that naturally accumulate sorbitol, its biosynthesis involves two key enzymes: sorbitol-6-phosphate dehydrogenase (S6PDH), which reduces glucose-6-phosphate (G6P) to sorbitol-6-phosphate (S6P) using NADPH, and sorbitol-6-phosphate phosphatase (S6PP), which converts S6P into free sorbitol. Its activity highly dependent on Mg+2 [161]. Then, sorbitol can be metabolized in either fructose or glucose. The main pathway involves sorbitol dehydrogenase which is NAD⁺-dependent (NAD-SDH), catalysing the reversible conversion of sorbitol to fructose. Sorbitol dehydrogenase, which is NADP+-dependant (NADP-SDH), also contributes to sorbitol transformation, producing glucose instead. Although this enzymatic reaction has a lower affinity for sorbitol, it is still reversible, whereas sorbitol oxidase (SOX), while still less active than NAD-SDH, irreversibly converts sorbitol to glucose. The activity of both SDH enzymes are induced by Ca2+ and Mg2+ availability [174].
In table grapes, sorbitol has a major role in cellular homeostasis. Under abiotic stress conditions such as high salinity and drought, polyols function as regulators thanks to their osmotic potential and capability to retain water, mitigating damage risk and stabilizing proteins and enzymes [175]. Furthermore, polyols also have a scavenging function against ROS, which are formed under certain moments of stress, preventing oxidative damage and subsequent plant deterioration and reduced productivity [176]. In grapevines, an increase in the VvPLT1 transporter was observed in response to salt and water stress, which has significantly higher affinity to sorbitol and mannitol resulting in polyol accumulation in grown berry tissue, therefore establishing a causal link between abiotic stress, polyol accumulation, and genetic response [177]. Sugar alcohol applications have also been proven to stimulate plant growth regulators [3], as well as PAL and UFGT activity, with the subsequently increase in anthocyanin, stilbenes, and total polyphenols content [3,172].
Regarding foliar fertilization, the presence of smaller particles, surfactants that reduce surface tension, and the use of optimal pH and concentration levels further enhance nutrient penetration and absorption, ultimately improving the overall efficiency of foliar fertilization [178]. In particular, sorbitol has proven to be an effective surfactant for foliar fertilization, as it modifies the morphology, the vapor pressure values and the distribution of fertilizer droplets through shape, gravity, vapor pressure and salt aggregation modulation [179].
The European Union´s (EU) strict regulation of fertilizers makes polyols a preharvest treatments interesting strategies to control issues aggravated by climate-change and environmental changes. In the EU, polyol regulations are encompassed in the legislation on fertilizers and additives, Regulation (EC) Nº 2003/2003, establishing the basis for the authorization of fertilizer products, including chelating and complexing agents, a description that fits polyols well. Although this regulation does not specifically mention polyols, their inclusion in the list of chelating agents may be an important step torward regulating these compounds. These directives promote the use of sustainable agricultural practices and the reduction of the environmental impact of chemical products. Nevertheless, the individual agricultural policy of each country could include modifications regarding the use of chelating agents that fit within the framework of these directives.
5.2. Complexation Mechanism and Transport of Sorbitol
Foliar applications of these nutrients complexed with sorbitol have been to improve resistance to various physiological stresses and disorders (Table 2). In this context, some authors refer to polyols as “vectors” [115,180] due to their role in forming carrier-mediated complexes with nutrients, thereby enhancing the transport of agrochemical products into specific targeted plant organs. In grapes, phloem transportation of nutrients may be an important way to improve fruit quality with preharvest treatments, as transport via xylem is non-functional after veraison berry growth [181]. Although the detailed mechanism by which sorbitol enhances nutrient transportation through phloem is still a subject of study [182], some researchers suggest that polyols facilitate mobility of other nutrients as they are able to behave as reducing agents, dissolving inorganic compounds and preventing particle aggregation through the process of dissolving minerals at high temperatures, thanks to theirs high dielectric constants [183,184]. Sorbitol’s multiple -OH groups could enhance the solubility of Ca salts by facilitating the formation of calcium-sorbitol complexes. These complexes are stabilized within a hydration sphere through hydrogen bonding in an aqueous medium (Figure 1). This process of cation stabilization is favoured by the redox potential of Ca2+ (-2.87 V) [185], small ionic radius (1.00 Å) [186], and the high enthalpy of complexation [187], which is promoted under acidic conditions, particularly at pH levels below 4. A temperature of 50 °C, pH 4, and a duration of 45 min, were shown to be the optimum conditions for stabilizing the chelating reaction of inorganic Ca with sorbitol, along with a specific mass ratio of sorbitol with Ca ion of 1.45-1.94 [188]. Computational modelling [187] and experimental data [189] both confirm that the presence of sorbitol increases Ca salt solubility, likely due to the formation of energetically favourable and highly hydrated complexes.
In order to overcome the low mobility of Ca in the phloem, the vectorization process is employed [115]. Vectorization involves associating compounds with limited translocation within the plant to a ‘vector’ that facilitates and controls their distribution. Commonly used vectors include amino acids, carboxylic acids, phenolic acids, sugars, and polyols (such as sorbitol, mannitol, and xylitol) [115]. It has been shown that calcium-sorbitol complex treatments on vegetables increased the total Ca concentration, especially the pectin-bound Ca, in basal, intermediate, and young leaves, as well as in fruits (up to 76% in pepper fruits) [115]and table grapes [25]. This is crucial, as pectin synthesis and the binding of Ca to these structures are closely linked to the total Ca concentration in plant tissues.
6. Applications and Effect of Calcium-Sorbitol Complexes in Table Grape Quality
6.1. Enhancing Ca Transport and Other Nutrients
Spraying foliar fertilizers and phytosanitary products to the aerial parts of plants is an effective tool for crop management and sustainability [178]. Applying fertilizers directly to the aerial parts of the plant can be beneficial in minimizing nutrient loss and preventing soil contamination. However, to achieve an adequate supply of macronutrients, foliar fertilization should be considered a supplementary strategy which can be particularly effective in specific situations, such as poor soil conditions, nutrients with limited phloem mobility, or to improve compatibility for nutrient distribution, thereby avoiding sole reliance on root uptake. For instance, in acidic soils, P solubility can be significantly restricted, whereas in alkaline soils, the availability of micronutrients such as Zn, F, and Mn is often diminished. Under arid conditions, foliar feeding becomes especially valuable, as the uptake of macronutrients like K is limited during drought-induced water stress [190]. Foliar application is also advantageous when nutrient uptake through roots is compromised by competitive interactions. For example, the use of urea to supply N when its absorption competes with polysaccharides at the root level [191]. Additionally, foliar fertilization can be used for the bio-fortification of the nutritional quality of crops in order to improve human health and nutrition by increasing the content of essential elements such as zinc [192] and selenium [193]. Furthermore, foliar nutrient enrichment can be used to alleviate physiological disorders in plants.
This expanded agronomic practice has been successfully used to apply methyl jasmonate [194], algae-based fertilizers [195], amino acids [196], and minerals [144,197], among other phytoproducts [198]. A different approach toward transportation has been proposed by Zhang et al. (2021) [199], who applied glycinebetaine in rice (Oryza sativa L.), chelating Al3+ in the plant roots, which is a detrimental element for plant function, and increasing Ca2+ concentrations up to 25 and 28 % in shoots and roots, respectively.
Minerals are important nutrients for plants though some of them are known to have low mobility through the phloem. Physicochemical factors restrict the efficient long-distance transport of Ca. Due to cation exchange within the xylem, Ca competes with other ions including H+ for binding sites on the walls of xylem vessels and pit membranes, making pH a determinant factor in modulating Ca mobility. Moreover, Ca is able to form low soluble or insoluble compounds, such as Ca oxalate, which restricts its translocation. Due to its limited mobility in the phloem, Ca primarily accumulates in transpiring aerial tissues (leaves). Thus, Ca transport to low transpiration sink organs, such as developing fruits, is often restricted, particularly after the fruit has evolved beyond the fruit set stage [114].
B is an essential micronutrient required for the growth of higher plants with an essential role in cell wall expansion. Most of B content is located in the cell wall and was considered phloem-immobile, meaning it could not be effectively redistributed within the plant. In 1996, Brown and Hu observed that despite being considered phloem-immobile, B was free and mobile in Pyrus, Malus, and Prunus species [200], which are major sorbitol producers, and later in celery (Apium Graveloens L.) [201], a mannitol producer specie. Proposed models for B polyol-dependent transport are attributed to the structural and chemical properties of B. Most of B within the plant can be found in the form of cis-diol esters complexed in cell wall pectins and polygalacturonans, being functionally immobile. In contrast, free B is primarily present as boric acid (H₃BO₃) in the cytoplasm and as the anion borate B(OH)4- (Figure 2), as it is the main reaction in aqueous solutions [202], where the pH of 7.5 favours this molecular form. The small molecular size and high membrane permeability of H3BO3 enable its diffusion from the phloem to near xylem vessels, returning to the leaves and preventing B from reaching sink organs [203]. H3BO3 naturally reacts with esters with polyhydroxy compounds like polyols (Figure 1), forming stable complexes [201] that are phloem mobile. This finding suggests that polyols are crucial for B mobility through the phloem and for reaching sink organs in plants.
Nevertheless, treatments of B have been reported as effective when applications are made from early stages of fruit setting, as treated almond trees were observed to achieve a higher concentration in flower buds, flowers, and bulbs. Will et al. (2012) [204] reported that foliar absorption B is significantly greater when applied to the abaxial (lower) leaf surface compared to the adaxial (upper) surface in both lychee and soybean, highlighting the importance of targeting the correct leaf side to enhance uptake, especially considering the thick and waxy characteristics of lychee leaves acknowledged by the authors. Also, the application of both sorbitol and CaCl₂ enhanced foliar B absorption, likely by reducing relative humidity. This humectant effect is particularly valuable for fertilizers like boric acid, promoting better nutrient uptake through prolonged leaf surface wetness [204]. In a similar way as B, Ca is expected to achieve higher mobility through the phloem when complexed with sorbitol (Figure 3).
6.2. Effects on Fruit Quality and Stress Resistance
In potatoes, chelated calcium-sorbitol fertilizer revealed better performance than solely inorganic Ca fertilizers, promoting yield as well as the absorption and utilization of N, P and K nutrients [188]. When Ca, zinc and sorbitol were sprayed individually, quality and yield traits in potato increased, including total soluble solids, dry matter, protein, and starch content. Furthermore, the three solutions combined resulted in the plants with the highest quality values [205]. This effect has also been observed when sorbitol is mixed with B and Ca applied to mangoes [129], achieving higher yield and quality traits. However, other authors observed a higher absorption rate of B but less translocated content of the mineral, probably due to a high B application, inducing toxicity in the soybean plants [206]. In peanuts, dose ranging from 1.5 to 1.8 g L-1 of chelated calcium-sorbitol treatments significantly increased yield production by up to 28.6 %, and fat content by 5.0 %, potassium and Ca content in peanut kernels by 98.6 and 55.3 %, respectively, and dry matter accumulation by 37.9 % in comparison to untreated peanut plants [207]. In wheat, sorbitol spray treatments with nanoparticles of Cu in addition to zeolite fertilizer contributed to a higher nutrient uptake and accumulation of the plants, leading to higher yield rates [208], highlighting the importance of symbiotic strategies with polyols in order to maximize crop performance.
The chelated calcium-sorbitol complex has been applied to white seedless table grapes of the ‘Doña María’ variety, which is included in a Protected Designation of Origin (PDO) and is required to achieve a minimum of 12.5 °Brix in total soluble solids [25]. Foliar applications during on-tree ripening improved the mobility of Ca intro the fruit as calcium-pectate, or ‘bound calcium’, resulting in increased fruit firmness and reduced ABA and MDA contents. Polyphenols synthesis was stimulated, leading to higher antioxidant activity. In chelated calcium-sorbitol treated grapes, malic acid degradation was preserved while greater amounts of glucose and fructose were shown [25]. Overall, these changes resulted in a superior maturity index while preserving good storability traits.
Calcium-sorbitol chelated treatments promoted anthocyanin biosynthesis in ‘Sanguinelli’ blood oranges through stimulation of the phenylpropanoid pathway, of which the PAL enzyme is key to triggerimg the synthesis of flavonoids [172]. After 17 days of the first treatment and during the on-tree ripening stage to harvest date, total phenolic and antioxidant activity was higher in treated fruits in every fraction: juice, flavedo, albedo, and pulp. The same behaviour was observed for anthocyanin content, minerals content, individual sugars, and individual acids. When blood oranges were stored at 8 °C, anthocyanin levels significantly increased. However, this increase was more prominent in treated fruits as well as hesperidin and narirutin. Overall, chelated calcium-sorbitol treatment preserved bioactive compounds in blood oranges, such as sugars and acids, resulting in an improved sensory quality after 30 days of storage [171].
Other sugar alcohols, such as inositol, have shown positive effects in wine grapes by enhancing yield, soluble solids, tannin, and anthocyanin contents when applied at doses between 2.4 and 3.6 L hm⁻². These results suggest that polyols may contribute to improved Ca absorption; however, higher doses have been associated with detrimental effects on leaf photosynthetic performance [211]. In honeydew melons, chelated Ca with mannitol resulted in elevated mesocarp Ca concentrations, elevated fruit firmness, and enhanced marketability without a modification in the sugar content of fruit [214].
7. Conclusions and Future Perspectives
Adverse conditions for growing table grapes in certain regions caused by climate change and the poor suitability of arid soils highlight the need for more sustainable solutions. Ca has proven to be an effective tool in preventing physiological disorders in table grapes and other crops. Additionally, sorbitol has shown the ability to promote defense mechanisms against both biotic and abiotic stresses, while also improving fruit quality. Combining sorbitol with Ca may offer beneficial effects, not only due to their individual properties, but also thanks to sorbitol’s synergistic role in enhancing Ca transport, helping it reach sink organs more efficiently. Furthermore, sorbitol complexation with other low-mobility nutrients and elicitors could enhance their effects, opening new research opportunities involving polyols. Therefore, calcium-sorbitol treatments contribute to reducing postharvest food losses and generate economic benefits for producers and farmers by improving marketable yield and extending shelf-life. At the same time, it provides affordable, sustainable, and environmentally friendly treatment that is most likely compatible with global agricultural practices. In conclusion, calcium-sorbitol treatments represent an alternative tool, offering a nutrient-based fertilization option when fruit development or specific conditions require a complementary approach to ensure and enhance fruit quality and preservation.
Author Contributions
Conceptualization, D.M-R. and A.G.; writing—review and editing, A.G, A.S-G., M.E.G-P. M.H.D., J.M.V. and D.M-R.; funding acquisition, J.M.V. and D.M-R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by MICIU/AEI /10.13039/501100011033 and by FEDER, UE, grant number PID2022-137282OB-I00
Acknowledgments
The authors thank the Generalitat Valenciana, the Conselleria of Education, Universities and Employment, for the funding Alberto Guirao Carrascosa´s PhD-scholarships (CIACIF/2022/270), which enable him to undertake doctoral studies, and to the European Social Fund for co-financing these grants.
Conflicts of Interest
The authors declare no conflicts of interest, financial or otherwise, that could have influenced the research outcomes or conclusions presented in this work.
References
- Moradi, S.; Koushesh Saba, M.; Sadeghi, S.; Inglese, P.; Liguori, G. Changes in Biochemical and Bioactive Compounds in Two Red Grape Cultivars during Ripening and Cold Storage. Agronomy 2024, 14. [Google Scholar] [CrossRef]
- Akram, M.T.; Qadri, R.; Khan, M.A.; Atak, A.; Liaquat, M.; Hussain, T.; Khan, M.M.; Azam, M.; Hasan, M. Ul Comparative Assessment of Bioactive Compounds, Fruit Quality Attributes and Sugar Profiling in Early Maturing Table Grape (Vitis Vinifera L.) Cultivars from Pothohar, Pakistan. Applied Fruit Sci 2024, 66, 983–995. [Google Scholar] [CrossRef]
- Conde, A.; Badim, H.; Dinis, L.T.; Moutinho-Pereira, J.; Ferrier, M.; Unlubayir, M.; Lanoue, A.; Gerós, H. Stimulation of Secondary Metabolism in Grape Berry Exocarps by a Nature-Based Strategy of Foliar Application of Polyols. Oeno One 2024, 58. [Google Scholar] [CrossRef]
- Ban, Z.; Zhang, S.; Niu, C.; Liu, L.; Cao, K.; Li, L.; Wu, Z.; Wang, L.; Chen, C.; Zhu, Y. Potential Role of Exogenous Melatonin Involved in Postharvest Quality Maintenance of Vitis Labrusca × Vinifera ‘Kyoho. ’ J Sci Food Agric 2023, 103, 6243–6251. [Google Scholar] [CrossRef]
- Admane, N.; Genovese, F.; Altieri, G.; Tauriello, A.; Trani, A.; Gambacorta, G.; Verrastro, V.; Di Renzo, G.C. Effect of Ozone or Carbon Dioxide Pre-Treatment during Long-Term Storage of Organic Table Grapes with Modified Atmosphere Packaging. LWT 2018, 98, 170–178. [Google Scholar] [CrossRef]
- Solairaj, D.; Guillaume Legrand, N.N.; Yang, Q.; Zhang, H. Isolation of Pathogenic Fungi Causing Postharvest Decay in Table Grapes and in Vivo Biocontrol Activity of Selected Yeasts against Them. Physiol Mol Plant Pathol 2020, 110. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, J.; Wang, S.; Ke, Y.; Ren, T.; Wang, Y. Mechanism Analysis of Amphotericin B Controlling Postharvest Gray Mold in Table Grapes. Foods 2025, 14. [Google Scholar] [CrossRef]
- Pedrotti, C.; Marcon, Â.R.; Delamare, A.P.L.; Echeverrigaray, S.; da Silva Ribeiro, R.T.; Schwambach, J. Alternative Control of Grape Rots by Essential Oils of Two Eucalyptus Species. J Sci Food Agric 2019, 99, 6552–6561. [Google Scholar] [CrossRef] [PubMed]
- Quintieri, L.; Fancello, F.; Caputo, L.; Sorrentino, A.; Zara, S.; Lippolis, V.; Cervellieri, S.; Fanelli, F.; Corvino, A.; Pace, B.; et al. Effect of Gaseous Citral on Table Grapes Contaminated by Rhizopus Oryzae ITEM 18876. Foods 2022, 11. [Google Scholar] [CrossRef]
- Torres-Palazzolo, C.; Ferreyra, S.; Iribas, F.; Chimeno, V.; Rojo, M.C.; Casalongue, C.; Fontana, A.; Combina, M.; Ponsone, M.L. Biocontrol of Alternaria Alternata in Cold-Stored Table Grapes Using Psychrotrophic Yeasts and Bioactive Compounds of Natural Sources. Int J Food Microbiol 2024, 415. [Google Scholar] [CrossRef]
- Chen, L.; Zhao, C.; Yan, T.; Li, B.; Wang, S.; Gong, D.; Long, D. Antifungal Potentiality of Non-Volatile Compounds Produced from Hanseniaspora Uvarum against Postharvest Decay of Table Grape Fruit Caused by Botrytis Cinerea and Penicillium Expansum. Postharvest Biol Technol 2025, 222. [Google Scholar] [CrossRef]
- Mundy, D.C.; Elmer, P.; Wood, P.; Agnew, R. A Review of Cultural Practices for Botrytis Bunch Rot Management in New Zealand Vineyards. Plants 2022, 11. [Google Scholar] [CrossRef]
- Soldateli, F.J.; Both, V.; Thewes, F.R.; Wendt, L.M.; Ludwig, V.; Rossato, F.P.; Thewes, F.R.; Batista, C.B.; Brackmann, A.; Wagner, R. Overall Quality, Phenolic Compounds, and Volatile Profile of ‘BRS Isis’ Seedless Table Grapes after Long-Term Storage: Effect of Ethanol and High CO2. Sci Hortic 2023, 312. [Google Scholar] [CrossRef]
- Zoffoli, J.P.; Latorre, B.A.; Naranjo, P. Hairline, a Postharvest Cracking Disorder in Table Grapes Induced by Sulfur Dioxide. Postharvest Biol Technol 2008, 47, 90–97. [Google Scholar] [CrossRef]
- Shi, Y.; Li, B.J.; Su, G.; Zhang, M.; Grierson, D.; Chen, K.S. Transcriptional Regulation of Fleshy Fruit Texture. J Integr Plant Biol 2022, 64, 1649–1672. [Google Scholar] [CrossRef]
- Yu, J.; Li, J.; Hong, Z.; Zeng, Q.; Fu, Y.; Deng, R.; Xu, K.; Huang, Z.; Zhu, M. VvPIP1;1 Plays a Role in Grape Berry Cracking by Regulating Water Uptake. Hortic Plant J 2025, 11, 1506–1516. [Google Scholar] [CrossRef]
- Khadivi-Khub, A. Physiological and Genetic Factors Influencing Fruit Cracking. Acta Physiol Plant 2015, 37. [Google Scholar] [CrossRef]
- Santos, M.; Egea-Cortines, M.; Gonçalves, B.; Matos, M. Molecular Mechanisms Involved in Fruit Cracking: A Review. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef]
- Simon, G. Review on Rain Induced Fruit Cracking of Sweet Cherries (Prunus Avium L.), Its Causes and Possibilities of Prevention. Int J Hortic Sci 2006, 12, 3–17. [Google Scholar] [CrossRef]
- Dashrath, S.; Nrcg, R.; Parhe, S.D.; Bhagwat, S.; Rajya, M.; Bagaitdar Sangh Pune, D. Berry Cracking; Its Causes and Remedies in Grapes-A Review. Trend in Biosciences 2017, 10, 2. [Google Scholar]
- La Spada, P.; Continella, A.; Dominguez, E.; Heredia, A.; Gentile, A. Improving Cuticle Thickness and Quality Traits in Table Grape Cv. ‘Italia’ Using Pre-Harvest Treatments. Plants 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Brüggenwirth, M.; Knoche, M. Cell Wall Swelling, Fracture Mode, and the Mechanical Properties of Cherry Fruit Skins Are Closely Related. Planta 2017, 245, 765–777. [Google Scholar] [CrossRef]
- Yang, Z.; Wu, Z.; Zhang, C.; Hu, E.; Zhou, R.; Jiang, F. The Composition of Pericarp, Cell Aging, and Changes in Water Absorption in Two Tomato Genotypes: Mechanism, Factors, and Potential Role in Fruit Cracking. Acta Physiol Plant 2016, 38. [Google Scholar] [CrossRef]
- Wang, J.; Gao, X.; Ma, Z.; Chen, J.; Liu, Y. Analysis of the Molecular Basis of Fruit Cracking Susceptibility in Litchi Chinensis Cv. Baitangying by Transcriptome and Quantitative Proteome Profiling. J Plant Physiol 2019, 234–235, 106–116. [Google Scholar] [CrossRef]
- Guirao, A.; Valverde, J.M.; Díaz-Mula, H.M.; Valero, D.; Serrano, M.; Martínez-Romero, D. Role of Pre-Harvest Sorbitol–Calcium Treatments in Controlling Berry Drop in Bagged Table Grapes of the “Doña María” Variety. Horticulturae 2024, 10. [Google Scholar] [CrossRef]
- Deng, Y.; Wu, Y.; Li, Y.; Zhang, P.; Yang, M.; Shi, C.; Zheng, C.; Yu, S. A Mathematical Model for Predicting Grape Berry Drop during Storage. J Food Eng 2007, 78, 500–511. [Google Scholar] [CrossRef]
- Meneses, M.; García-Rojas, M.; Muñoz-Espinoza, C.; Carrasco-Valenzuela, T.; Defilippi, B.; González-Agüero, M.; Meneses, C.; Infante, R.; Hinrichsen, P. Transcriptomic Study of Pedicels from GA3-Treated Table Grape Genotypes with Different Susceptibility to Berry Drop Reveals Responses Elicited in Cell Wall Yield, Primary Growth and Phenylpropanoids Synthesis. BMC Plant Biol 2020, 20. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Calderón, Á.; Falagán, N.; Terry, L.A.; Alamar, M.C. Biomarkers of Postharvest Resilience: Unveiling the Role of Abscisic Acid in Table Grapes during Cold Storage. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef]
- Rizzuti, A.; Aguilera-Sáez, L.M.; Gallo, V.; Cafagna, I.; Mastrorilli, P.; Latronico, M.; Pacifico, A.; Matarrese, A.M.S.; Ferrara, G. On the Use of Ethephon as Abscising Agent in Cv. Crimson Seedless Table Grape Production: Combination of Fruit Detachment Force, Fruit Drop and Metabolomics. Food Chem 2015, 171, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. lin; Zhang, R. guang Effects of ABA Content on the Development of Abscission Zone and Berry Falling After Harvesting of Grapes. Agric Sci China 2009, 8, 59–67. [Google Scholar] [CrossRef]
- Zhu, M.; Zheng, L.; Zeng, Y.; Yu, J. Susceptibility of Two Grape Varieties to Berry Abscission. Sci Hortic 2022, 304. [Google Scholar] [CrossRef]
- Teker, T.; Soltekin, O. Berry Shattering Phenomena in Vineyards: The Influence of Maximum Temperatures during Flowering Period in an Extreme Year. Sci Hortic 2023, 321. [Google Scholar] [CrossRef]
- Khalil, U.; Rajwana, I.A.; Razzaq, K.; Farooq, U.; Saleem, B.A.; Brecht, J.K. Quality Attributes and Biochemical Changes in White and Colored Table Grapes as Influenced by Harvest Maturity and Ambient Postharvest Storage. South African Journal of Botany 2023, 154, 273–281. [Google Scholar] [CrossRef]
- Sabir, F.K.; Sabir, A.; Payli, T.; Unal, S. Exogenous Melatonin Treatments Maintain Berry Quality and Bioactive Compounds of Minimally Processed Table Grapes (‘Crimson Seedless’) During Cold Storage. Applied Fruit Science 2024, 66, 1609–1617. [Google Scholar] [CrossRef]
- Zenoni, S.; Savoi, S.; Busatto, N.; Tornielli, G.B.; Costa, F. Molecular Regulation of Apple and Grape Ripening: Exploring Common and Distinct Transcriptional Aspects of Representative Climacteric and Non-Climacteric Fruits. J Exp Bot 2023, 74, 6207–6223. [Google Scholar] [CrossRef] [PubMed]
- Ovadia, R.; Oren-Shamir, M.; Kaplunov, T.; Zutahy, Y.; Lichter, A.; Lurie, S. Effects of Plant Growth Regulators and High Temperature on Colour Development in “Crimson Seedless” Grapes. J Hortic Sci Biotech 2013, 88, 387–392. [Google Scholar] [CrossRef]
- Shinomiya, R.; Fujishima, H.; Muramoto, K.; Shiraishi, M. Impact of Temperature and Sunlight on the Skin Coloration of the ‘Kyoho’ Table Grape. Sci Hortic 2015, 193, 77–83. [Google Scholar] [CrossRef]
- Bahar, A.; Kaplunov, T.; Alchanatis, V.; Lichter, A. Evaluation of Methods for Determining Rachis Browning in Table Grapes. Postharvest Biol Technol 2017, 134, 106–113. [Google Scholar] [CrossRef]
- Lichter, A. Rachis Browning in Tablegrapes. Aust J Grape Wine Res 2016, 22, 161–168. [Google Scholar] [CrossRef]
- Hamie, N.; Zoffoli, J.P.; Tarricone, L.; Verrastro, V.; Pérez-Donoso, A.G.; Gambacorta, G. Rachis Browning and Water Loss Description during Postharvest Storage of ‘Krissy’ and ‘Thompson Seedless’ Table Grapes. Postharvest Biol Technol 2022, 184. [Google Scholar] [CrossRef]
- Lichter, A.; Kaplunov, T.; Zutahy, Y.; Daus, A.; Alchanatis, V.; Ostrovsky, V.; Lurie, S. Physical and Visual Properties of Grape Rachis as Affected by Water Vapor Pressure Deficit. Postharvest Biol Technol 2011, 59, 25–33. [Google Scholar] [CrossRef]
- Lo’ay, A.A.; Taha, N.A. Evaluation Rachis Browning Phenomena of ‘Superior Seedless’ Vines Grafted on Different Rootstocks during Shelf Life. Sci Hortic 2020, 261. [Google Scholar] [CrossRef]
- Tóth, A.M. Precision Canopy Management of the Grapevine: Early Defoliation and Girdling. Acta Carolus Robertus 2020, 107–118. [Google Scholar] [CrossRef]
- Timpanaro, N.; Ferlito, F.; Amenta, M.; Torrisi, B.; Allegra, M.; Rapisarda, P.; Romeo, F. V. Effect of Branch Girdling on Berry Traits, Chemical and Sensory Characteristics of ‘Italia’ and ‘Victoria’ Table Grapes. N Z J Crop Hortic Sci 2023, 51, 52–68. [Google Scholar] [CrossRef]
- Tóth, A.M.; Zsófi, Z.; Veres, S. Cane Girdling Influence on the Berry Texture Properties of Three Table Grape Varieties. Horticulturae 2022, 8. [Google Scholar] [CrossRef]
- Goren, R.H.M.; Goldsmichmidt, E.E. Girdling: Physiological Horticultural Aspects, Janick, John Wiley & Sons, Inc, J Horticultural Reviews 2004, 30 ISBN 9780471354208.
- Reynolds, A.G.; De Savigny, C. Influence of Girdling and Gibberellic Acid on Yield Components, Fruit Composition, and Vestigial Seed Formation of ‘Sovereign Coronation’ Table Grapes. Hort Science 2004, 39, 3–541. [Google Scholar] [CrossRef]
- Abu-Zahra, T. Berry Size of Thompson Seedless as Influenced by the Application of Gibberellic Acid and Cane Girdling. Pak J Bot 2010, 42, 3–1755. [Google Scholar]
- Chen, T.; Xu, T.; Shen, L.; Zhang, T.; Wang, L.; Chen, Z.; Wu, Y.; Yang, J. Effects of Girdling and Foliar Fertilization with K on Physicochemical Parameters, Phenolic and Volatile Composition in ‘Hanxiangmi’ Table Grape. Horticulturae 2022, 8. [Google Scholar] [CrossRef]
- Ferrara, G.; Mazzeo, A.; Netti, G.; Pacucci, C.; Matarrese, A.M.S.; Cafagna, I.; Mastrorilli, P.; Vezzoso, M.; Gallo, V. Girdling, Gibberellic Acid, and Forchlorfenuron: Effects on Yield, Quality, and Metabolic Profile of Table Grape Cv. ‘Italia’. Am J Enol Vitic 2014, 65, 381–387. [Google Scholar] [CrossRef]
- Soltekin, O.; Candemir, A.; Altindisli, A. Effects of Cane Girdling on Yield, Fruit Quality and Maturation of (Vitis Vinifera L.) Cv. ‘Flame Seedless’. BIO Web Conf 2016, 7, 01032. [Google Scholar] [CrossRef]
- Yamane, T.; Shibayama, K. Effects of Trunk Girdling and Crop Load Levels on Fruit Quality and Root Elongation in ‘Aki Queen’ Grapevines. J Japan Soc Hort Sci 2006, 75, 6–439. [Google Scholar] [CrossRef]
- İşçi, B.; Kacar, E.; Altındişli, A. The Effects of Some Exogenous Applications on Quality in ‘Crimson Seedless’ Grape. Erwerbs-Obstbau 2020, 62, 87–100. [Google Scholar] [CrossRef]
- Zhu, M.; Yu, J.; Xu, Y.; Yang, G. Effect of Girdling on Anthocyanin Content and Quality of Spine Grape Berries. J Plant Growth Regul 2022, 41, 65–73. [Google Scholar] [CrossRef]
- Crupi, P.; Antonacci, D.; Savino, M.; Genghi, R.; Perniola, R.; Coletta, A. Girdling and Gibberellic Acid Effects on Yield and Quality of a Seedless Red Table Grape for Saving Irrigation Water Supply. Eur J Agron 2016, 80, 21–31. [Google Scholar] [CrossRef]
- Xi, X.; Zha, Q.; He, Y.; Tian, Y.; Jiang, A. Influence of Cluster Thinning and Girdling on Aroma Composition in ‘Jumeigui’ Table Grape. Sci Rep 2020, 10. [Google Scholar] [CrossRef]
- Trentham, W.R.; Sams, C.E.; Conway, W.S. Histological Effects of Calcium Chloride in Stored Apples. J Amer Soc Hort Sci 2008, 133, 4–487. [Google Scholar] [CrossRef]
- García-Pastor, M.E.; Serrano, M.; Guillén, F.; Castillo, S.; Martínez-Romero, D.; Valero, D.; Zapata, P.J. Methyl Jasmonate Effects on Table Grape Ripening, Vine Yield, Berry Quality and Bioactive Compounds Depend on Applied Concentration. Sci Hortic 2019, 247, 380–389. [Google Scholar] [CrossRef]
- García-Pastor, M.E.; Zapata, P.J.; Castillo, S.; Martínez-Romero, D.; Valero, D.; Serrano, M.; Guillén, F. Preharvest Salicylate Treatments Enhance Antioxidant Compounds, Color and Crop Yield in Low Pigmented-Table Grape Cultivars and Preserve Quality Traits during Storage. Antioxidants 2020, 9, 1–17. [Google Scholar] [CrossRef]
- García-Pastor, M.E.; Giménez, M.J.; Zapata, P.J.; Guillén, F.; Valverde, J.M.; Serrano, M.; Valero, D. Preharvest Application of Methyl Salicylate, Acetyl Salicylic Acid and Salicylic Acid Alleviated Disease Caused by Botrytis Cinerea through Stimulation of Antioxidant System in Table Grapes. Int J Food Microbiol 2020, 334. [Google Scholar] [CrossRef]
- García-Pastor, M.E.; Giménez, M.J.; Serna-Escolano, V.; Guillén, F.; Valero, D.; Serrano, M.; García-Martínez, S.; Terry, L.A.; Alamar, M.C.; Zapata, P.J. Oxalic Acid Preharvest Treatment Improves Colour and Quality of Seedless Table Grape ‘Magenta’ Upregulating on-Vine Abscisic Acid Metabolism, Relative VvNCED1 Gene Expression, and the Antioxidant System in Berries. Front Plant Sci 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Wan, Y.; Qin, G.; Xu, Y. Induction of Defense Responses against Alternaria Rot by Different Elicitors in Harvested Pear Fruit. Appl Microbiol Biotechnol 2006, 70, 729–734. [Google Scholar] [CrossRef]
- Kawano, T.; Furuichi, T. Salicylic Acid: a Plant Hormone, first edition, Hayat, S; A. A. Springer Dordrecht, 2007, 10, ISBN–10. [Google Scholar]
- Zheng, X.; Jing, G.; Liu, Y.; Jiang, T.; Jiang, Y.; Li, J. Expression of Expansin Gene, MiExpA1, and Activity of Galactosidase and Polygalacturonase in Mango Fruit as Affected by Oxalic Acid during Storage at Room Temperature. Food Chem 2012, 132, 849–854. [Google Scholar] [CrossRef]
- Giménez, M.J.; Valverde, J.M.; Valero, D.; Guillén, F.; Martínez-Romero, D.; Serrano, M.; Castillo, S. Quality and Antioxidant Properties on Sweet Cherries as Affected by Preharvest Salicylic and Acetylsalicylic Acids Treatments. Food Chem 2014, 160, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Giménez, M.J.; Valverde, J.M.; Valero, D.; Díaz-Mula, H.M.; Zapata, P.J.; Serrano, M.; Moral, J.; Castillo, S. Methyl Salicylate Treatments of Sweet Cherry Trees Improve Fruit Quality at Harvest and during Storage. Sci Hortic 2015, 197, 665–673. [Google Scholar] [CrossRef]
- Giménez, M.J.; Serrano, M.; Valverde, J.M.; Martínez-Romero, D.; Castillo, S.; Valero, D.; Guillén, F. Preharvest Salicylic Acid and Acetylsalicylic Acid Treatments Preserve Quality and Enhance Antioxidant Systems during Postharvest Storage of Sweet Cherry Cultivars. J Sci Food Agric 2017, 97, 1220–1228. [Google Scholar] [CrossRef]
- Dar, T.A.; Uddin, M.; Khan, M.M.A.; Hakeem, K.R.; Jaleel, H. Jasmonates Counter Plant Stress: A Review. Environ Exp Bot 2015, 115, 49–57. [Google Scholar] [CrossRef]
- Martínez-Esplá, A.; Serrano, M.; Valero, D.; Martínez-Romero, D.; Castillo, S.; Zapata, P.J. Enhancement of Antioxidant Systems and Storability of Two Plum Cultivars by Preharvest Treatments with Salicylates. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef]
- Martínez-Esplá, A.; Zapata, P.J.; Valero, D.; Martínez-Romero, D.; Díaz-Mula, H.M.; Serrano, M. Preharvest Treatments with Salicylates Enhance Nutrient and Antioxidant Compounds in Plum at Harvest and after Storage. J Sci Food Agric 2018, 98, 2742–2750. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhang, D.; Zhang, H.; Jiang, G.; Su, X.; Qu, H.; Jiang, Y.; Duan, X. Physiological and Biochemical Response of Harvested Plum Fruit to Oxalic Acid during Ripening or Shelf-Life. Food Research International 2011, 44, 1299–1305. [Google Scholar] [CrossRef]
- Kumar, D. Salicylic Acid Signaling in Disease Resistance. Plant Science 2014, 228, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Min, D.; Li, F.; Ji, N.; Meng, D.; Li, L. Synergistic Effects of l -Arginine and Methyl Salicylate on Alleviating Postharvest Disease Caused by Botrysis Cinerea in Tomato Fruit. J Agric Food Chem 2017, 65, 4890–4896. [Google Scholar] [CrossRef]
- Chen, J.Y.; Wen, P.F.; Kong, W.F.; Pan, Q.H.; Zhan, J.C.; Li, J.M.; Wan, S.B.; Huang, W.D. Effect of Salicylic Acid on Phenylpropanoids and Phenylalanine Ammonia-Lyase in Harvested Grape Berries. Postharvest Biol Technol 2006, 40, 64–72. [Google Scholar] [CrossRef]
- Oraei, M.; Panahirad, S.; Zaare-Nahandi, F.; Gohari, G. Pre-Véraison Treatment of Salicylic Acid to Enhance Anthocyanin Content of Grape (Vitis Vinifera L.) Berries. J Sci Food Agric 2019, 99, 5946–5952. [Google Scholar] [CrossRef]
- Blanch, G.P.; Gómez-Jiménez, M.C.; del Castillo, M.L.R. Exogenous Salicylic Acid Improves Phenolic Content and Antioxidant Activity in Table Grapes. Plant Foods for Human Nutrition 2020, 75, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Habibi, F.; Ramezanian, A.; Rahemi, M.; Eshghi, S.; Guillén, F.; Serrano, M.; Valero, D. Postharvest Treatments with γ-Aminobutyric Acid, Methyl Jasmonate, or Methyl Salicylate Enhance Chilling Tolerance of Blood Orange Fruit at Prolonged Cold Storage. J Sci Food Agric 2019, 99, 6408–6417. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Esplá, A.; Zapata, P.J.; Valero, D.; García-Viguera, C.; Castillo, S.; Serrano, M. Preharvest Application of Oxalic Acid Increased Fruit Size, Bioactive Compounds, and Antioxidant Capacity in Sweet Cherry Cultivars (Prunus Avium L.). J Agric Food Chem 2014, 62, 3432–3437. [Google Scholar] [CrossRef]
- Flores, G.; Ruiz del Castillo, M.L. Variations in Ellagic Acid, Quercetin and Myricetin in Berry Cultivars after Preharvest Methyl Jasmonate Treatments. Journal of Food Composition and Analysis 2015, 39, 55–61. [Google Scholar] [CrossRef]
- Flores, G.; Ruiz del Castillo, M.L. Accumulation of Anthocyanins and Flavonols in Black Currants (Ribes Nigrum L.) by Pre-Harvest Methyl Jasmonate Treatments. J Sci Food Agric 2016, 96, 4026–4031. [Google Scholar] [CrossRef]
- Portu, J.; Santamaría, P.; López-Alfaro, I.; López, R.; Garde-Cerdán, T. Methyl Jasmonate Foliar Application to Tempranillo Vineyard Improved Grape and Wine Phenolic Content. J Agric Food Chem 2015, 63, 2328–2337. [Google Scholar] [CrossRef]
- Portu, J.; López, R.; Baroja, E.; Santamaría, P.; Garde-Cerdán, T. Improvement of Grape and Wine Phenolic Content by Foliar Application to Grapevine of Three Different Elicitors: Methyl Jasmonate, Chitosan, and Yeast Extract. Food Chem 2016, 201, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Gil-Muñoz, R.; Fernández-Fernández, J.I.; Crespo-Villegas, O.; Garde-Cerdán, T. Elicitors Used as a Tool to Increase Stilbenes in Grapes and Wines. Food Research International 2017, 98, 34–39. [Google Scholar] [CrossRef]
- Sun, C.; Zhu, C.; Tang, Y.; Ren, D.; Cai, Y.; Zhou, G.; Wang, Y.; Xu, L.; Zhu, P. Inhibition of Botrytis Cinerea and Control of Gray Mold on Table Grapes by Calcium Propionate. Food Quality and Safety 2021, 5. [Google Scholar] [CrossRef]
- Young, P.R.; Lashbrooke, J.G.; Alexandersson, E.; Jacobson, D.; Moser, C.; Velasco, R.; Vivier, M.A. The Genes and Enzymes of the Carotenoid Metabolic Pathway in Vitis Vinifera L. BMC Genomics 2012, 13. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Gambetta, G.A.; Wada, H.; Krasnow, M.N.; Cramer, G.R.; Peterlunger, E.; Shackel, K.A.; Matthews, M.A. Characterization of Major Ripening Events during Softening in Grape: Turgor, Sugar Accumulation, Abscisic Acid Metabolism, Colour Development, and Their Relationship with Growth. J Exp Bot 2016, 67, 709–722. [Google Scholar] [CrossRef]
- Valero, D.; Díaz-Mula, H.M.; Zapata, P.J.; Castillo, S.; Guillén, F.; Martínez-Romero, D.; Serrano, M. Postharvest Treatments with Salicylic Acid, Acetylsalicylic Acid or Oxalic Acid Delayed Ripening and Enhanced Bioactive Compounds and Antioxidant Capacity in Sweet Cherry. J Agric Food Chem 2011, 59, 5483–5489. [Google Scholar] [CrossRef]
- Deng, J.; Bi, Y.; Zhang, Z.; Xie, D.; Ge, Y.; Li, W.; Wang, J.; Wang, Y. Postharvest Oxalic Acid Treatment Induces Resistance against Pink Rot by Priming in Muskmelon (Cucumis Melo L.) Fruit. Postharvest Biol Technol 2015, 106, 53–61. [Google Scholar] [CrossRef]
- Razzaq, K.; Khan, A.S.; Malik, A.U.; Shahid, M.; Ullah, S. Effect of Oxalic Acid Application on Samar Bahisht Chaunsa Mango during Ripening and Postharvest. LWT 2015, 63, 152–160. [Google Scholar] [CrossRef]
- Razavi, F.; Hajilou, J. Enhancement of Postharvest Nutritional Quality and Antioxidant Capacity of Peach Fruits by Preharvest Oxalic Acid Treatment. Sci Hortic 2016, 200, 95–101. [Google Scholar] [CrossRef]
- Zhu, Y.; Yu, J.; Brecht, J.K.; Jiang, T.; Zheng, X. Pre-Harvest Application of Oxalic Acid Increases Quality and Resistance to Penicillium Expansum in Kiwifruit during Postharvest Storage. Food Chem 2016, 190, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Kok, D.; Bal, E. Changes on Bioactive Compounds and Electrochemical Characteristics of Cv. ‘Horoz Karası’ Table Grape (V. Vinifera L.) Induced by Various Doses of Preharvest Applications of Benzoic Acid, Citric Acid and Oxalic Acid at Berry Setting and Verasion Periods. Erwerbs-Obstbau 2019, 61, 17–24. [Google Scholar] [CrossRef]
- Zheng, X.; Tian, S.; Gidley, M.J.; Yue, H.; Li, B. Effects of Exogenous Oxalic Acid on Ripening and Decay Incidence in Mango Fruit during Storage at Room Temperature. Postharvest Biol Technol 2007, 45, 281–284. [Google Scholar] [CrossRef]
- Abdusattarov, B.; Safarov, A.; Toshmatov, B. Determination of the Influence of Packaging Measures of a Variety of Husayni Grapes. In Proceedings of the E3S Web of Conferences; EDP Sciences, April 14 2023, 381. [Google Scholar] [CrossRef]
- Kulapichitr, F.; Asensio, C.; Arpaia, M.L.; Walse, S.; Obenland, D. Effect of Controlled Atmosphere Storage on Key Volatiles and Sensory Perception of Muscat Grapes. ACS Food Science and Technology 2024, 4, 766–772. [Google Scholar] [CrossRef]
- Liguori, G.; Sortino, G.; Gullo, G.; Inglese, P. Effects of Modified Atmosphere Packaging and Chitosan Treatment on Quality and Sensorial Parameters of Minimally Processed Cv. ‘Italia’ Table Grapes. Agronomy 2021, 11. [Google Scholar] [CrossRef]
- Martínez-Romero, D.; Guillén, F.; Castillo, S.; Valero, D.; Serrano, M. Modified Atmosphere Packaging Maintains Quality of Table Grapes. J Food Sci 2003, 68, 1838–1843. [Google Scholar] [CrossRef]
- Matera, A.; Altieri, G.; Genovese, F.; Scarano, L.; Genovese, G.; Pinto, P.; Rashvand, M.; Elshafie, H.S.; Ippolito, A.; Mincuzzi, A.; et al. Impact of the Pre-Harvest Biocontrol Agent and Post-Harvest Massive Modified Atmosphere Packaging Application on Organic Table Grape (Cv. ‘Allison’) Quality during Storage. Appl Sci (Switzerland) 2024, 14. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, J.; Chen, Y.; Wei, J.; Wu, B. Nitric Oxide Treatment Maintains Postharvest Quality of Table Grapes by Mitigation of Oxidative Damage. Postharvest Biol Technol 2019, 152, 9–18. [Google Scholar] [CrossRef]
- Pretel, M.T.; Martínez-Madrid, M.C.; Martínez, J.R.; Carreño, J.C.; Romojaro, F. Prolonged Storage of ‘Aledo’ Table Grapes in a Slightly CO2 Enriched Atmosphere in Combination with Generators of SO2. LWT 2006, 39, 1109–1116. [Google Scholar] [CrossRef]
- Xing, S.; Wang, M.; Zhang, Z.; Yuan, Y.Y.; Song, Z.; Wu, B.; Wei, J. Sulfur Dioxide Enhances Postharvest Grape Resistance to Botrytis Cinerea by Promoting Glutathione Level. Sci Hortic 2024, 334. [Google Scholar] [CrossRef]
- Xing, S.; Tian, Q.; Zheng, Y.; Yuan, Y.Y.; Zhang, Z.; Zhang, Y.; Zhang, H.; Wei, J.; Wu, B. Sulfur Dioxide Enhances the Resistance of Postharvest Grape Berries to Gray Mold through Hydrogen Peroxide Signaling. Postharvest Biol Technol 2025, 221. [Google Scholar] [CrossRef]
- Yan, D.; Yi, H. Transcriptome Analysis Provides Insights into Preservation Mechanism of Postharvest Muscat Hamburg Grapes Treated with SO2. Sci Hortic 2024, 331. [Google Scholar] [CrossRef]
- Li, Z.; Tan, N.; Huang, J.; Wang, J.; Xiao, Y.; Xu, J.; Wang, Q.; Wu, B.; Luo, Z.; Xu, Y. H2O2-Mediated Cell Wall Remodeling and Pectin Demethylesterification Are Involved in Maintaining Postharvest Texture of Table Grape by Sulfur Dioxide. Food Chem 2025, 464. [Google Scholar] [CrossRef]
- de Aguiar, A.C.; Higuchi, M.T.; Yamashita, F.; Roberto, S.R. SO2-Generating Pads and Packaging Materials for Postharvest Conservation of Table Grapes: A Review. Horticulturae 2023, 9. [Google Scholar] [CrossRef]
- Reuveni, M.; Arroyo, C.J.; Ovadia, S. An Effective Hybrid Fungicide Containing Tea Tree Oil and Difenoconazole for Grape Powdery Mildew Management. Agric (Switzerland) 2023, 13. [Google Scholar] [CrossRef]
- El-Abbasy, U.K.; Abdel-Hameed, M.A.; Hatterman-Valenti, H.M.; El-Shereif, A.R.; Abd El-Khalek, A.F. Effectiveness of Oregano and Thyme Essential Oils as Alternatives for Sulfur Dioxide in Controlling Decay and Gray Mold and Maintaining Quality of ‘Flame Seedless’ Table Grape (Vitis Vinifera L.) during Cold Storage. Agronomy 2023, 13. [Google Scholar] [CrossRef]
- Guillén, F.; Zapata, P.J.; Martínez-Romero, D.; Castillo, S.; Serrano, M.; Valero, D. Improvement of the Overall Quality of Table Grapes Stored under Modified Atmosphere Packaging in Combination with Natural Antimicrobial Compounds. J Food Sci 2007, 72. [Google Scholar] [CrossRef]
- Loh, M.Y.; Adzahan, N.M.; Azman, E.M.; Koh, S.P.; Yusof, N.L. Enhancing Cold Tolerance and Quality Characteristics of Carica Papaya Linn through the Application of 1-Methylcyclopropene, Geranium and Lavender Oil. Int J Food Sci Technol 2024, 59, 3245–3257. [Google Scholar] [CrossRef]
- Dhanasekaran, S.; Liang, L.; Gurusamy, S.; Yang, Q.; Zhang, H. Chitosan Stabilized Lemon Essential Oil Nanoemulsion Controls Black Mold Rot and Maintains Quality of Table Grapes. Int J Biol Macromol 2024, 277. [Google Scholar] [CrossRef]
- Walsh, K.B. Postharvest Handling, third edition, Florkowski, W. J.; Banks, N.H; Shewfelt, R.L.; Prussia, S.E. Elsevier Inc., 2014, 8, ISBN–978. [Google Scholar]
- Nedjimi, B. Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M.; Fujita, M.; Oku, H.; Nahar, K.; Hawrylak-Nowak, B. Springer, Singapore, 2018, 9, pp. 205-220, ISBN 9789811090448.
- Fernández, V.; Brown, P.H. From Plant Surface to Plant Metabolism: The Uncertain Fate of Foliar-Applied Nutrients. Front Plant Sci 2013, 4. [Google Scholar] [CrossRef]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit Calcium: Transport and Physiology. Front Plant Sci 2016, 7. [Google Scholar] [CrossRef]
- Navarro-León, E.; López-Moreno, F.J.; Fernández, M.A.; Maldonado, J.J.; Yánez, J.; Blasco, B.; Ruiz, J.M. A New Calcium Vectoring Technology: Concentration and Distribution of Ca and Agronomic Efficiency in Pepper Plants. Agronomy 2022, 12. [Google Scholar] [CrossRef]
- Kim, Y.; Castroverde, C.D.M.; Kim, J.H. Natural Allelic Diversity of the Calcium Signaling Regulators in Plants. Mol Cells 2024, 47. [Google Scholar] [CrossRef]
- Shao, H.B.; Song, W.Y.; Chu, L.Y. Advances of Calcium Signals Involved in Plant Anti-Drought. C R Biol 2008, 331, 587–596. [Google Scholar] [CrossRef]
- Halling, D.B.; Liebeskind, B.J.; Hall, A.W.; Aldrich, R.W. Conserved Properties of Individual Ca2+-Binding Sites in Calmodulin. Proc Natl Acad Sci U S A 2016, 113, E1216–E1225. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, K. V.; Dubrovina, A.S. The Role of Calcium-Dependent Protein Kinase (CDPK) Genes in Plant Stress Resistance and Secondary Metabolism Regulation. Plant Growth Regul 2025, 105, 535–552. [Google Scholar] [CrossRef]
- Camm, E.L.; Towers, G.H.N. Phenylalanine Ammonia Lyase. Phytochemistry 1973, 12, 961–973. [Google Scholar] [CrossRef]
- Griffith, C.; Einhorn, T.C. The Effect of Plant Growth Regulators on Xylem Differentiation, Water and Nutrient Transport, and Bitter Pit Susceptibility of Apple. Sci Hortic 2023, 310. [Google Scholar] [CrossRef]
- Reitz, N.F.; Mitcham, E.J. Lignification of Tomato (Solanum Lycopersicum) Pericarp Tissue during Blossom-End Rot Development. Sci Hortic 2021, 276. [Google Scholar] [CrossRef]
- Reitz, N.F.; Shackel, K.A.; Mitcham, E.J. Differential Effects of Excess Calcium Applied to Whole Plants vs. Excised Fruit Tissue on Blossom-End Rot in Tomato. Sci Hortic 2021, 290. [Google Scholar] [CrossRef]
- Loekito, S.; Afandi; Afandi, A. ; Nishimura, N.; Koyama, H.; Senge, M. The Effects of Calcium Fertilizer Sprays during Fruit Development Stage on Pineapple Fruit Quality under Humid Tropical Climate. International Journal of Agronomy 2022, 2022. [Google Scholar] [CrossRef]
- Pusittigul, I.; Siriphanich, J.; Juntee, C. Role of Calcium on Internal Browning of Pineapples. Acta Hortic 2014, 1024. [Google Scholar] [CrossRef]
- Youryon, P.; Supapvanich, S.; Kongtrakool, P.; Wongs-Aree, C. Calcium Chloride and Calcium Gluconate Peduncle Infiltrations Alleviate the Internal Browning of Queen Pineapple in Refrigerated Storage. Hortic Environ Biotechnol 2018, 59, 205–213. [Google Scholar] [CrossRef]
- Hou, Y.; Liu, Y.; Zhao, L.; Zhao, Y.; Wu, Z.; Zheng, Y.; Jin, P. EjCML19 and EjWRKY7 Synergistically Function in Calcium Chloride-Alleviated Chilling Injury of Loquat Fruit. Postharvest Biol Technol 2023, 203. [Google Scholar] [CrossRef]
- Li, Z.; Wang, L.; Xie, B.; Hu, S.; Zheng, Y.; Jin, P. Effects of Exogenous Calcium and Calcium Chelant on Cold Tolerance of Postharvest Loquat Fruit. Sci Hortic 2020, 269. [Google Scholar] [CrossRef]
- Talang, H.D.; Dutta, P.; Mukhim, C.; Patil, S. Effect of Calcium, Boron and Sorbitol on Fruit-Set, Yield and Quality in Mango Cv. Himsagar. J Hort Sci, 2017, 11, 2, 166-169.
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of Macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, Third Edition, Elsevier Inc., 2011, pp. 135–189 ISBN 9780123849052.
- Maletsika, P.; Liava, V.; Sarrou, E.; Titeli, V.S.; Nasiopoulou, E.; Martens, S.; Karagiannis, E.; Grigoriadou, K.; Molassiotis, A.; Nanos, G.D. Foliar Calcium Effects on Quality and Primary and Secondary Metabolites of White-Fleshed ‘Lemonato’ Peaches. Horticulturae 2023, 9. [Google Scholar] [CrossRef]
- Manganaris, G.A.; Vasilakakis, M.; Diamantidis, G.; Mignani, I. Effect of Calcium Additives on Physicochemical Aspects of Cell Wall Pectin and Sensory Attributes of Canned Peach (Prunus Persica (L) Batsch Cv Andross). J Sci Food Agric 2005, 85, 1773–1778. [Google Scholar] [CrossRef]
- Shah, S.T.; Sajid, M.; Basit, A.; Haleema, B.; Khan, A.; Mohamed, H.I. Revealing Effects of Calcium-Based Formulations Focusing on Membrane Instability, Fruit Quality, Phytochemical Properties, and Disease Incidence in Peach (Prunus Persica) During Storage. Applied Fruit Science 2025, 67. [Google Scholar] [CrossRef]
- Wu, Y.; Dong, X.; Zhu, Y.; Chen, H. Influence of Pre-Harvest Sprays of Calcium Chloride on Fruit Storage Quality and Softening of ‘Fengtangli’ (Prunus Salicina L.) Plum. CYTA - Journal of Food 2025, 23. [Google Scholar] [CrossRef]
- Michailidis, M.; Polychroniadou, C.; Kosmidou, M.A.; Petraki-Katsoulaki, D.; Karagiannis, E.; Molassiotis, A.; Tanou, G. An Early Calcium Loading during Cherry Tree Dormancy Improves Fruit Quality Features at Harvest. Horticulturae 2021, 7. [Google Scholar] [CrossRef]
- Erogul, D. Effect of Preharvest Calcium Treatments on Sweet Cherry Fruit Quality. Not Bot Horti Agrobo 2014, 42, 1–150. [Google Scholar] [CrossRef]
- Guerra, M.; Sanz, M.Á.; Rodríguez-González, Á.; Casquero, P.A. Effect of Sustainable Preharvest and Postharvest Techniques on Quality and Storability of High-Acidity ‘Reinette Du Canada’ Apple. Horticulturae 2022, 8. [Google Scholar] [CrossRef]
- Reitz, N.F.; Mitcham, E.J. Impact of Postharvest Dips with Abscisic Acid, Prohexadione, Calcium, or Water on Bitter Pit Incidence and Apple Physiology. Postharvest Biol Technol 2025, 219. [Google Scholar] [CrossRef]
- Li, S.; Tao, S.; Zhang, J.; Wu, L.; Huan, C.; Zheng, X. Effect of Calcium Treatment on the Storability and Disease Resistance in Preharvest Bagging Mango Fruit during Room Temperature Storage. J Food Process Preserv 2020, 44. [Google Scholar] [CrossRef]
- Asgarian, Z.S.; Karimi, R.; Palou, L. Pre-Harvest Foliar Spraying of Calcium and Zinc Preserves Berries Quality and Mitigates Chilling Injury of Grape during Cold Storage. Sci Hortic 2024, 338. [Google Scholar] [CrossRef]
- Ciccarese, A.; Stellacci, A.M.; Gentilesco, G.; Rubino, P. Effectiveness of Pre- and Post-Veraison Calcium Applications to Control Decay and Maintain Table Grape Fruit Quality during Storage. Postharvest Biol Technol 2013, 75, 135–141. [Google Scholar] [CrossRef]
- Abd El-Wahab, S.M.; Abd El-Zaher, M.H.; Hussein, M.M. Prolonging Shelf Life of Flame Seedless Table Grapes with Pre-Harvest Calcium and Magnesium Treatment. In Proceedings of the BIO Web of Conferences; EDP Sciences, April 23 2025; Vol. 173. [Google Scholar] [CrossRef]
- Zhu, M.; Li, J.; Liu, Y.; Wang, Q.; Fan, Z.; Zeng, J.; Yu, J. Preharvest Nano-Calcium Reduces the Table Grape Berry Abscission by Regulating Ethylene Production During Storage. J Plant Growth Regul 2024, 43, 1400–1409. [Google Scholar] [CrossRef]
- Wang, G.; Wang, J.; Han, X.; Chen, R.; Xue, X. Effects of Spraying Calcium Fertilizer on Photosynthesis, Mineral Content, Sugar–Acid Metabolism and Fruit Quality of Fuji Apples. Agronomy 2022, 12. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Du, H.; Li, W. Impact of Preharvest Foliar Application of Calcium Ascorbate on Quality, Health Promoting Phytochemicals and Postharvest Life of Thompson Seedless Table Grape. J Plant Growth Regul 2025, 44, 879–890. [Google Scholar] [CrossRef]
- Park, Y.S.; Lee, J.C.; Kim, J.H.; Jeong, H.N.; Heo, J.Y. Effect of Calcium Nitrate Treatment on Reduction of Berry Shattering in ‘Cheongsoo’ Grape Cultivars. Not Sci Biol 2022, 14. [Google Scholar] [CrossRef]
- Lang, A.; During, H. Grape Berry Splitting and Some Mechanical Properties of the Skin. Vitis 1990, 29, 61–70. [Google Scholar]
- Zhang, C.; Wu, J. Yun; Cui, L. wen; Fang, J. Gui Mining of Candidate Genes for Grape Berry Cracking Using a Genome-Wide Association Study. J Integr Agric 2022, 21, 2291–2304. [Google Scholar] [CrossRef]
- Huang, X.M.; Wang, H.C.; Lu, X.J.; Yuan, W.Q.; Lu, J.M.; Li, J.G.; Huang, H.B. Cell Wall Modifications in the Pericarp of Litchi (Litchi Chinensis Sonn.) Cultivars That Differ in Their Resistance to Cracking. J Hortic Sci Biotech 2006, 81, 231–237. [Google Scholar] [CrossRef]
- Giné-Bordonaba, J.; Echeverria, G.; Ubach, D.; Aguiló-Aguayo, I.; López, M.L.; Larrigaudière, C. Biochemical and Physiological Changes during Fruit Development and Ripening of Two Sweet Cherry Varieties with Different Levels of Cracking Tolerance. Plant Physiol Biochem 2017, 111, 216–225. [Google Scholar] [CrossRef]
- Yu, J.; Zhu, M.; Bai, M.; Xu, Y.; Fan, S.; Yang, G. Effect of Calcium on Relieving Berry Cracking in Grape (Vitis Vinifera L.) “Xiangfei. ” PeerJ 2020, 8. [Google Scholar] [CrossRef]
- Lara, I.; Heredia, A.; Domínguez, E. Shelf Life Potential and the Fruit Cuticle: The Unexpected Player. Front Plant Sci 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Petit, J.; Bres, C.; Mauxion, J.P.; Bakan, B.; Rothan, C. Breeding for Cuticle-Associated Traits in Crop Species: Traits, Targets, and Strategies. J Exp Bot 2017, 68, 5369–5387. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Nguyen, N.; Hykkerud, A.L.; Häggman, H.; Martinussen, I.; Jaakola, L.; Karppinen, K. Developmental and Environmental Regulation of Cuticular Wax Biosynthesis in Fleshy Fruits. Front Plant Sci 2019, 10. [Google Scholar] [CrossRef]
- Winkler, A.; Brüggenwirth, M.; Ngo, N.S.; Knoche, M. Fruit Apoplast Tension Draws Xylem Water into Mature Sweet Cherries. Sci Hortic 2016, 209, 270–278. [Google Scholar] [CrossRef]
- Adel, A.-Q.; Awad, M.A. Effect of Pre-Harvest Calcium Chloride and Ethanol Spray on Quality of “El-Bayadi” Table Grapes during Storage. Vitis 2013, 52, 2–61. [Google Scholar]
- Chardonnet, C.O.; Sams, C.E.; Trigiano, R.N.; Conway, W.S. Variability of Three Isolates of Botrytis Cinerea Affects the Inhibitory Effects of Calcium on This Fungus. APS, 2000, 90, 7–769. [Google Scholar]
- Zhu, M.; Yu, J.; Tang, W.; Fan, S.; Bai, M.; Chen, M.; Yang, G. Role of Calcium in Regulating Anthocyanin Accumulation in ‘Manicure Finger’ Grape Berries. Sci Hortic 2019, 256. [Google Scholar] [CrossRef]
- Grembecka, M. Sugar Alcohols - Their Role in the Modern World of Sweeteners: A Review. European Food Research and Technology 2015, 241. [Google Scholar] [CrossRef]
- Shankar, P.; Ahuja, S.; Sriram, K. Non-Nutritive Sweeteners: Review and Update. Nutrition 2013, 29, 1293–1299. [Google Scholar] [CrossRef]
- Pleyerová, I.; Hamet, J.; Konrádová, H.; Lipavská, H. Versatile Roles of Sorbitol in Higher Plants: Luxury Resource, Effective Defender or Something Else? Planta 2022, 256. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.; Huang, B. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. Int J Genomics 2014, 2014. [Google Scholar] [CrossRef]
- Tian, G.; Liu, C.; Xu, X.; Xing, Y.; Liu, J.; Lyu, M.; Feng, Z.; Zhang, X.; Qin, H.; Jiang, H.; et al. Effects of Magnesium on Nitrate Uptake and Sorbitol Synthesis and Translocation in Apple Seedlings. Plant Physiol Biochem 2023, 196, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shen, Y.; Liu, D.H.; Liu, J.; Zhang, J.; Wei, J.; Wang, C.L. A Sorbitol Transporter Gene Plays Specific Role in the Occurrence of Watercore by Modulating the Level of Intercellular Sorbitol in Pear. Plant Science 2022, 317. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Wu, J.; Li, Y.; Deng, W.; Cao, K.; Li, Z.; Wang, L. Effect of Postharvest Cold Storage and Subsequent Shelf-Life on Fruit Quality and Endogenous Phytohormones in Nectarine Fruit. Postharvest Biol Technol 2024, 218. [Google Scholar] [CrossRef]
- Gómez-Martínez, H. Variation of Fruit Quality Traits in Apricot as Sources for Nutraceutical Breeding. Ph.D. Thesis, IVIA, Valencia, October 2021. [Google Scholar]
- Niu, X.X.; Deng, L.Z.; Wang, H.; Wang, Q.H.; Xu, M.Q.; Li, S. Bin; Okaiyeto, S.A.; Xiao, H.W. Transformation of Cell Wall Pectin Profile during Postharvest Ripening Process Alters Drying Behavior and Regulates the Sugar Content of Dried Plums. Food Chem 2024, 458. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Su, M.; Du, J.; Zhang, X.; Li, X.; Zhang, M.; Hu, Y.; Huan, C.; Ye, Z. Crucial Roles of Sorbitol Metabolism and Energy Status in the Chilling Tolerance of Yellow Peach. Plant Physiol Biochem 2023, 204. [Google Scholar] [CrossRef]
- Lee, J. Sorbitol, Rubus Fruit, and Misconception. Food Chem 2015, 166, 616–622. [Google Scholar] [CrossRef]
- Conde, A.; Soares, F.; Breia, R.; Gerós, H. Postharvest Dehydration Induces Variable Changes in the Primary Metabolism of Grape Berries. Food Res Int 2018, 105, 261–270. [Google Scholar] [CrossRef]
- Guirao, A.; Valverde, J.M.; Solana-Guilabert, A.; Díaz-Mula, H.M.; Valero, D.; Martínez-Romero, D. Impact of Pre-Harvest Sorbitol and Sorbitol-Ca Complex Treatments on the Quality of Fresh Blood Oranges and their Juice after Storage at 8°C. Postharvest Biol Technol 2025, 227. [Google Scholar] [CrossRef]
- Guirao, A.; Martínez-Romero, D.; Solana-Guilabert, A.; Agulló, V.; Díaz-Mula, H.M.; Valverde, J.M. Influence of Preharvest Sorbitol and Calcium-Sorbitol Applications on the Ripening Process and Anthocyanin Biosynthesis in Blood Orange (Citrus Sinensis Cv. Sanguinelli). Food Chem 2025, 481. [Google Scholar] [CrossRef] [PubMed]
- Freitas, J.R.L.; Vendramini, P.H.; Melo, J.O.F.; Eberlin, M.N.; Augusti, R. An Appraisal on the Source-to-Sink Relationship in Plants: An Application of Desorption Electrospray Ionization Mass Spectrometry Imaging. J Braz Chem Soc 2018, 29, 17–23. [Google Scholar] [CrossRef]
- Doehlert, D.C. Ketose Reductase Activity in Developing Maize Endosperm. Plant Physiol 1987, 84, 830–834. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of Osmoprotectants in Improving Salinity and Drought Tolerance in Plants: A Review. Rev Environ Sci Biotechnol 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol Biochem 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Conde, A.; Regalado, A.; Rodrigues, D.; Costa, J.M.; Blumwald, E.; Chaves, M.M.; Gerós, H. Polyols in Grape Berry: Transport and Metabolic Adjustments as a Physiological Strategy for Water-Deficit Stress Tolerance in Grapevine. J Exp Bot 2015, 66, 889–906. [Google Scholar] [CrossRef]
- Fernández, V.; Brown, P.H. Foliar Fertilization: Scientific Principles and Field Pratices, First edition, International Fertilizer Industry Association, Paris, France, 2013, ISBN 979-10-92366-00-6.
- Carreón, Y.J.P.; Pereyra Zarate, A.A.; Pérez Sánchez, A.E.; Díaz-Hernández, O.; González-Gutiérrez, J. Surface Coating with Foliar Fertilizers. Coatings 2024, 14. [Google Scholar] [CrossRef]
- Wu, H.; Xu, H.; Marivingt-Mounir, C.; Bonnemain, J.L.; Chollet, J.F. Vectorizing Agrochemicals: Enhancing Bioavailability via Carrier-Mediated Transport. Pest Manag Sci 2019, 75, 1507–1516. [Google Scholar] [CrossRef]
- Bondada, B.R.; Matthews, M.A.; Shackel, K.A. Functional Xylem in the Post-Veraison Grape Berry. J Exp Bot 2005, 56, 2949–2957. [Google Scholar] [CrossRef]
- Dinant, S.; Lemoine, R. The Phloem Pathway: New Issues and Old Debates. C R Biol 2010, 333, 307–319. [Google Scholar] [CrossRef]
- Laurent, S.; Forge, D.; Port, M.; Roch, A.; Robic, C.; Vander Elst, L.; Muller, R.N. Magnetic Iron Oxide Nanoparticles: Synthesis, Stabilization, Vectorization, Physicochemical Characterizations and Biological Applications. Chem Rev 2008, 108, 2064–2110. [Google Scholar] [CrossRef]
- Miguel, M.G.; Lourenço, J.P.; Faleiro, M.L. Superparamagnetic Iron Oxide Nanoparticles and Essential Oils: A New Tool for Biological Applications. Int J Mol Sci 2020, 21, 1–24. [Google Scholar] [CrossRef]
- Deng, X.; Li, L.; Zhang, G.; Zhao, X.; Hao, J.; Han, C.; Li, B. Anode Chemistry in Calcium Ion Batteries: A Review. Energy Storage Mater 2022. [CrossRef]
- Junio, J.B.; Chirawatkul, P.; Conato, M.T.; Mercado, C.C. Substitution of Ca2+ in Calcite by Sn2+ and Sr2+ Cations through Ion Exchange Characterized by X-Ray Absorption and Photoelectron Spectroscopies. MRS Adv 2021, 6, 342–349. [Google Scholar] [CrossRef]
- Zabozlaev, A.A.; Oganesyan, É.T.; Pogorelov, V.I. Drug Synthesis Methods and Manufacturing Technology. Solubilization of Poorly Soluble Calcium Salts of Organic Acids with Sorbitol. Pharm Chem J 2007, 41, 8. [Google Scholar] [CrossRef]
- Li, P.; Geng, C.; Li, L.; Li, Y.; Li, T.; Wei, Q.; Yan, D. Calcium-Sorbitol Chelating Technology and Application in Potatoes. Am J Biochem Biotechnol 2020, 16, 96–102. [Google Scholar] [CrossRef]
- Mäikinen, K.K.; Söderling, E. Solubility of Calcium Salts, Enamel, and Hydroxyapatite in Aqueous Solutions of Simple Carbohydrates. Calcif Tissue Int 1984, 36, 67–71. [Google Scholar] [CrossRef]
- Restrepo-Diaz, H.; Benlloch, M.; Fernández-Escobar, R. Plant Water Stress and K+ Starvation Reduce Absorption of Foliar Applied K+ by Olive Leaves. Sci Hortic 2008, 116, 409–413. [Google Scholar] [CrossRef]
- Gholami, A.; Akhlaghi, S.; Shahsavani, S.; Farrokhi, N. Effects of Urea Foliar Application on Grain Yield and Quality of Winter Wheat. Commun Soil Sci Plant Anal 2011, 42, 719–727. [Google Scholar] [CrossRef]
- Asif, M.; Asad, M.S.; Safdar, M.E.; Khan, I.; Akhtar, N.; Hassan, M.A.; Moosa, M.; Baoyi, Z.; Almoallim, H.S.; Ansari, M.J. Agronomic Bio-Fortification of Zinc Improves the Yield and Quality of Fodder Oat. J Ecol Eng 2024, 25, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, U.; Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Ashraf, J.; Shah, A.N.; Nawaz, M. Mineral Biofortification in Crop Plants for Ensuring Food Security; Hasanuzzaman, M. , Muhammad, S.T., Mohsin, T., Adnan, N.S. Springer, 2023, 6, pp.139-157, ISBN 978-981-99-4090-5. 6.
- Bonasia, A.; Lazzizera, C.; Elia, A.; Conversa, G. Pre-Harvest Strategy for Improving Harvest and Post-Harvest Performance of Kale and Chicory Baby Leaves. Plants 2025, 14. [Google Scholar] [CrossRef] [PubMed]
- Mangena, P. A Review on the Role of Algae-Nanoparticles as a Sustainable Agricultural Strategy to Improve Resilience against Abiotic Stresses in Leguminous Crops. Plant Nano Biol 2025, 11. [Google Scholar] [CrossRef]
- Xie, P.; Yang, Y.; Li, Y.; Wang, Y.; Bai, B.; Prusky, D.; Li, Y.; Bi, Y. Preharvest Phenylalanine Spraying Alleviates Chilling Injury in Harvested Muskmelons by Maintaining Reactive Oxygen Species Homeostasis. Food Chem 2025, 466. [Google Scholar] [CrossRef]
- Li, S.; Guo, S.; Xu, J.; Cai, Z.X.; Ma, R.; Yu, M.; Shen, Z. Preharvest Calcium Chloride Treatments Enhance Post-Harvest Peach Fruit Resistance to Monilinia Fructicola-Induced Brown Rot Disease. Postharvest Biol Technol 2025, 222. [Google Scholar] [CrossRef]
- Ssemugenze, B.; Ocwa, A.; Kuunya, R.; Gumisiriya, C.; Bojtor, C.; Nagy, J.; Széles, A.; Illés, Á. Enhancing Maize Production Through Timely Nutrient Supply: The Role of Foliar Fertiliser Application. Agronomy 2025, 15. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, W.; Li, D.; Zhou, F.; Chen, X.; Li, C.; Yu, S.; Brestic, M.; Liu, Y.; Yang, X. Glycinebetaine: A Versatile Protectant to Improve Rice Performance against Aluminium Stress by Regulating Aluminium Uptake and Translocation. Plant Cell Rep 2021, 40, 2397–2407. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.H.; Hu, H. Phloem Mobility of Boron Is Species Dependent : Evidence for Phloem Mobility in Sorbitol-Rich Species. Ann Bot 1996, 77, 497–505. [Google Scholar] [CrossRef]
- Hu, H.; Penn, S.C.; Lebrilla, C.B.; Brown, P.H. lsolation and Characterization of Soluble Boron Complexes in Higher Plants' The Mechanism of Phloem Mobility of Boron. Plant physiol 1997, 113, 649–655. [Google Scholar] [CrossRef]
- Raposo, J.C.; Zuloaga, O.; Olazabal, M.A.; Madariaga, J.M. Development of a Modified Bromley Methodology for the Estimation of Ionic Media Effects on Solution Equilibria: Part 5. The Chemical Model of Boric Acid in Aqueous Solution at 25 °C and Comparison with Arsenious Acid. Fluid Phase Equilib 2003, 207, 81–95. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of Nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants, Third Edition, Elsevier Inc., 2011; pp. 191–248 ISBN 9780123849052.
- Will, S.; Eichert, T.; Fernández, V.; Müller, T.; Römheld, V. Boron Foliar Fertilization of Soybean and Lychee: Effects of Side of Application and Formulation Adjuvants. J Plant Nutr Soil Sci 2012, 175, 180–188. [Google Scholar] [CrossRef]
- Al-Jburi, S.M.Y.; Alshammari, M.F. The Effect of Spraying Zinc, Calcium, and Sorbitol on Qualitative Yield Traits of Potato Solanum Tuberosum L. Bionatura 2023, 8. [Google Scholar] [CrossRef]
- Will, S.; Eichert, T.; Fernández, V.; Möhring, J.; Müller, T.; Römheld, V. Absorption and Mobility of Foliar-Applied Boron in Soybean as Affected by Plant Boron Status and Application as a Polyol Complex. Plant Soil 2011, 344, 283–293. [Google Scholar] [CrossRef]
- Liu, C.; Li, Y.; Huo, W.; Li, T.; Wei, Q.; Huang, M.; Geng, C.; Yan, D. Effect of Sorbitol Calcium Chelate on Yield and Calcium Nutrient Absorption of Peanut. Am J Biochem Biotechnol 2021, 17, 160–173. [Google Scholar] [CrossRef]
- Abd El-Azeiz, E.H.; El-Sonbaty, A.E.; El-Sherpiny, M.A. Enhancing the Quantitative and Qualitative Traits of Wheat Grown with Low Mineral Nitrogen Level through Zeolite, Sorbitol and Copper. Egypt J Soil Sci 2024, 64, 1053–1067. [Google Scholar] [CrossRef]
- Maibam, M.; Subba, S.; Chhetri, S.; Majhi, D.; Lepcha, L. Effect of Micronutrient on Quality, Yield and Shelf Life of Mango (Mangifera Indica L.) Cv. Himsagar. Proggressive Horticulture 2024, 56. [Google Scholar] [CrossRef]
- Nasif, S.; Soliman, G.; Farag, M.; Ibrahim, M.; Abd, M.; Nasser, E.-H. Application of Exogenous Polyols, Amino Acids and Girdling to Improve (Vitis Vinifera L.) Crimson Seedless Cv. Berries Coloration, and Postharvest Quality. Am-Eurasian J Agric Environ Sci 2023, 23, 70–80. [Google Scholar] [CrossRef]
- Ma, T.; Hui, Y.; Zhang, L.; Su, B.; Wang, R. Foliar Application of Chelated Sugar Alcohol Calcium Fertilizer for Regulating the Growth and Quality of Wine Grapes. Int J Agric Biol Eng 2022, 15, 153–158. [Google Scholar] [CrossRef]
- Alvarez, R.C.F.; Prado, R.M.; Souza Júnior, J.P.; Oliveira, R.L.L.; Felisberto, G.; Deus, A.C.F.; Cruz, F.J.R. Effects of Foliar Spraying with New Zinc Sources on Rice Seed Enrichment, Nutrition, and Productivity. Acta Agric Scand B Soil Plant Sci 2019. [Google Scholar] [CrossRef]
- Alabdaly, M.M.M.; Abdullah, S.K. Influence of Potassium, Sugar Alcohol, and Boron on Growth and Chemical Composition of Melons under Protected Cultivation. Anbar J Agric Sci 2024, 22, 160–172. [Google Scholar] [CrossRef]
- Lester, G.E.; Grusak, M.A. Field Application of Chelated Calcium: Postharvest Effects on Cantaloupe and Honeydew Fruit Quality. Hort Technol 2004, 14, 1. [Google Scholar] [CrossRef]
Figure 1.
Stabilisation of the calcium-sorbitol complex structure in an aqueous medium.
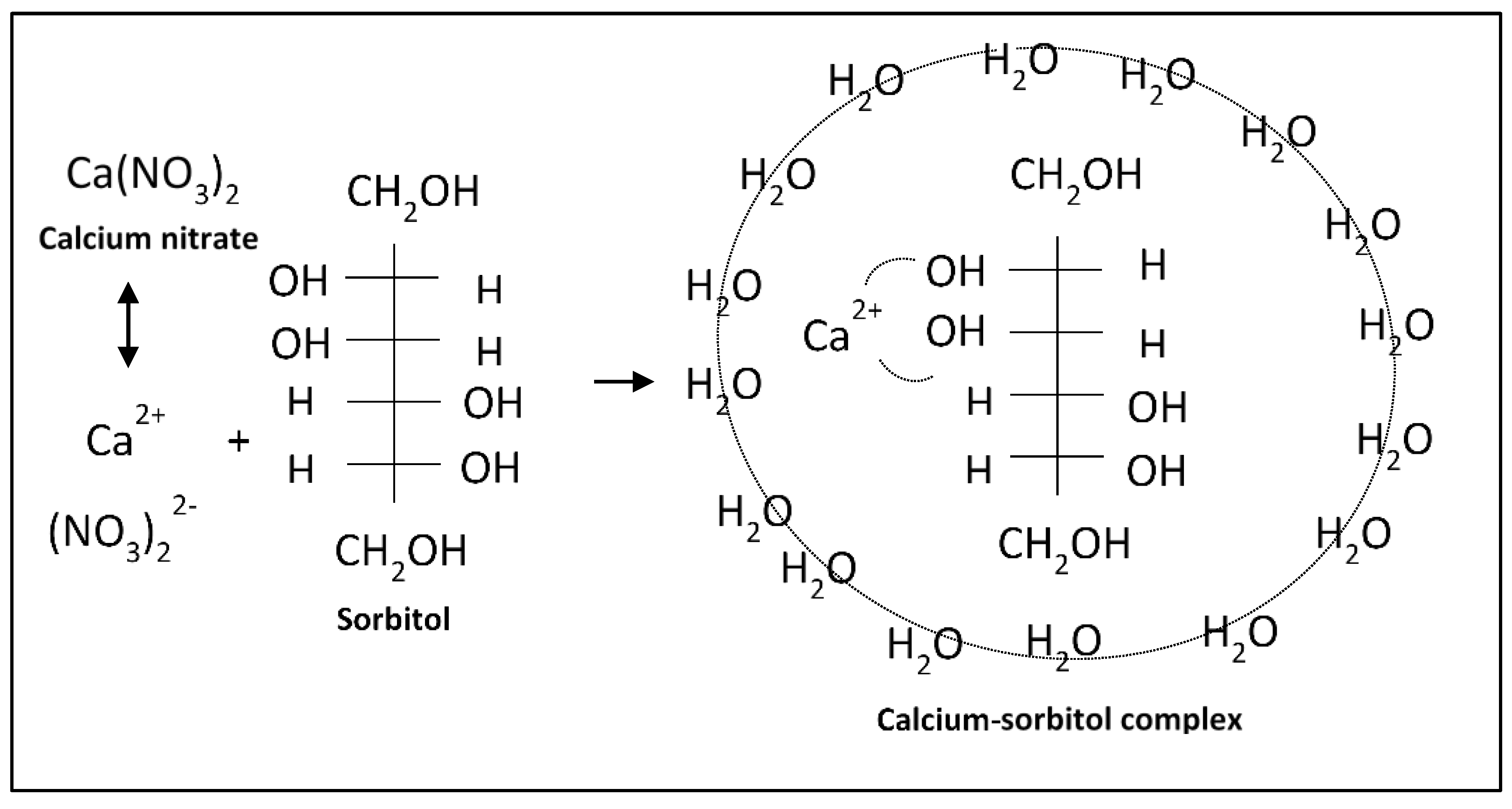
Figure 2.
Formation of the borate-sorbitol complex.
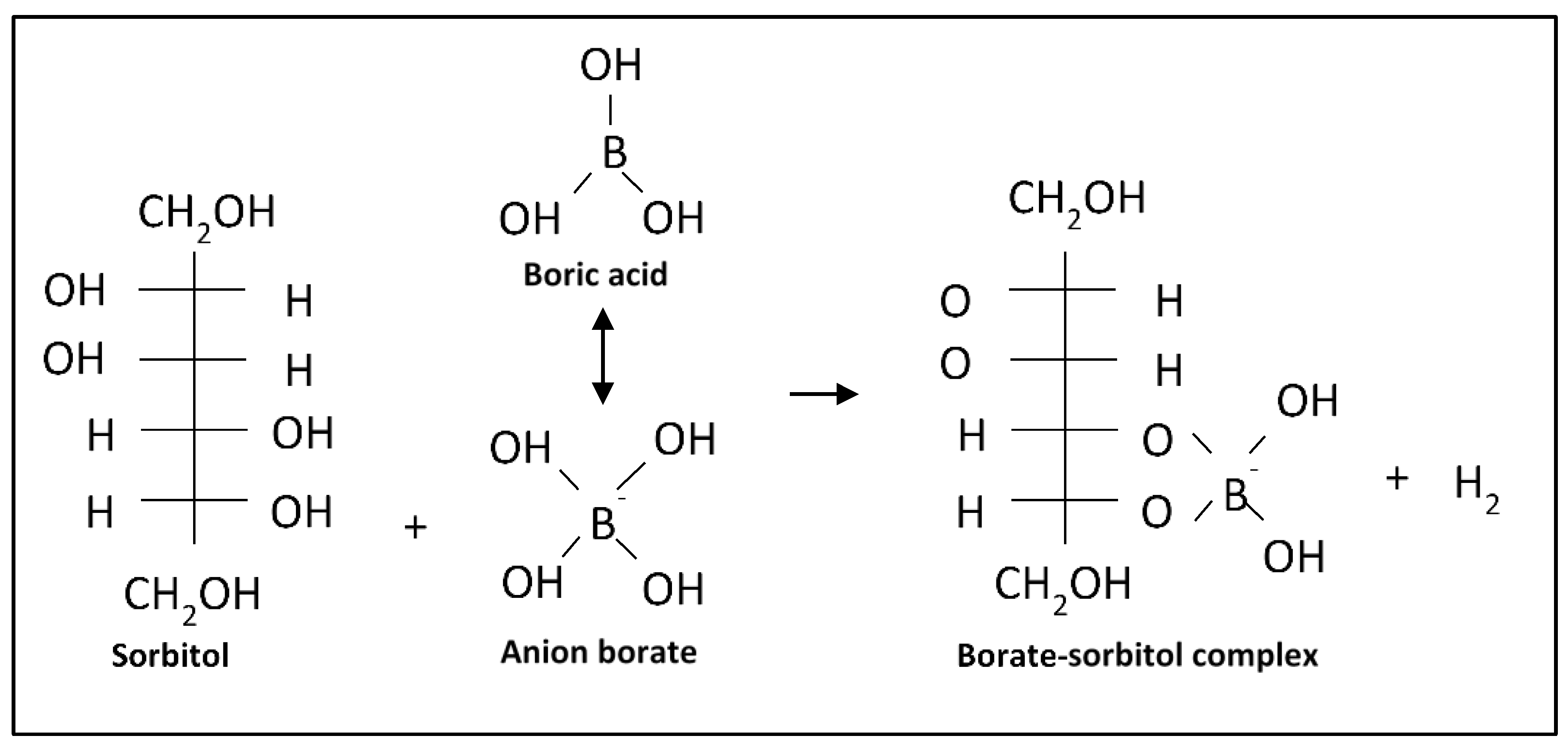
Figure 3.
Proposed transport mechanism of the calcium-sorbitol complex through the phloem.
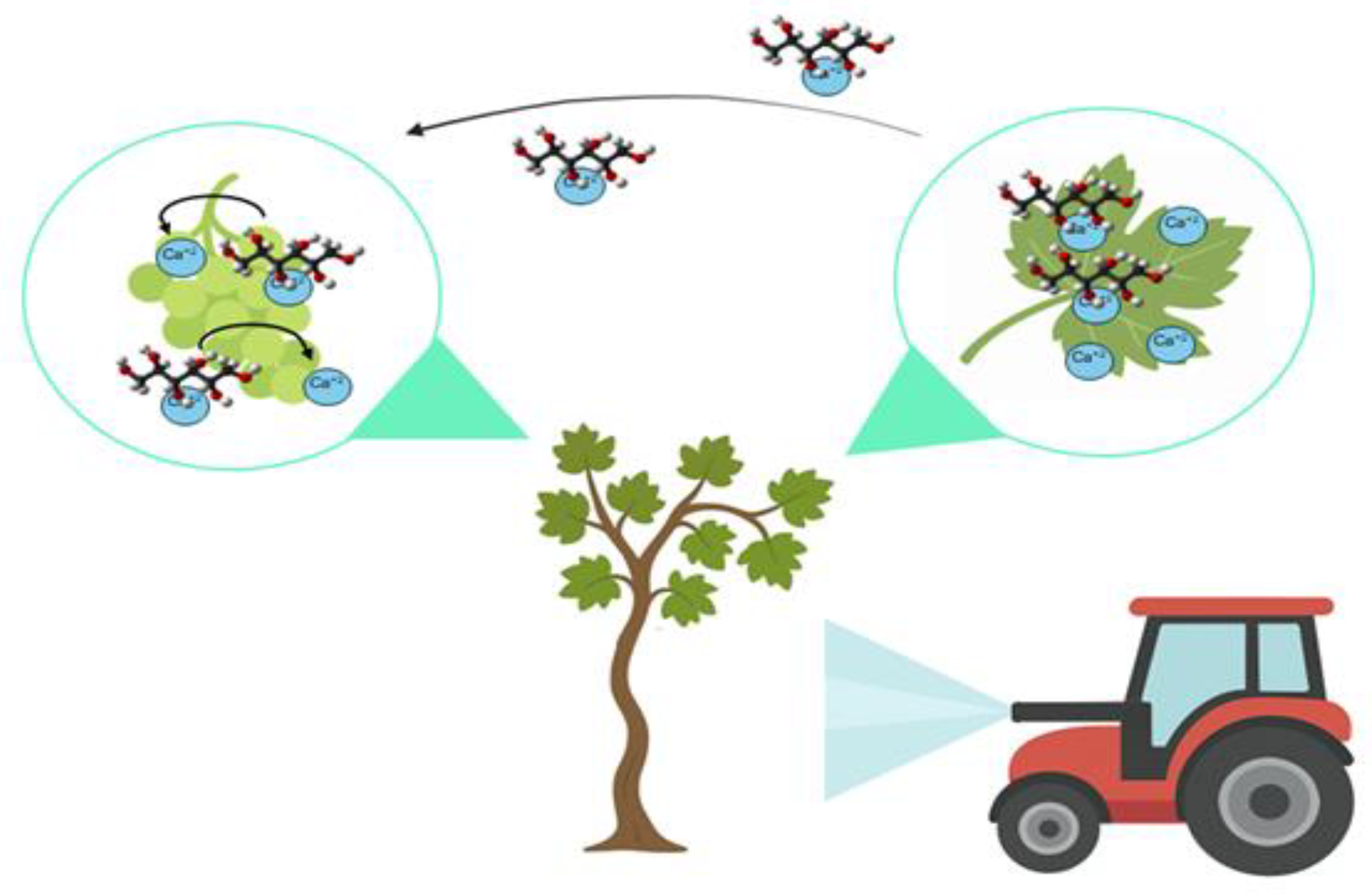

Table 1.
Effects of Ca nutrition on plant products.
| Fruit | Ca Products | Effects | References |
| Apple | Prohexadione-Ca CaCl2 CaCO3 |
-Alleviates bitter pit incidence | [121,137,138] |
| Tomato | CaCl2 | -Reduced blossom-end rot incidence and severity | [122,123] |
| Pineapple | CaCl2 Ca gluconate Ca oxide (CaO) Ca(NO3)2 Ca-boron |
-Reduced IL -Reduced MDA content -Reduced internal browning incidence |
[124,126] |
| Loquat | CaCl2 | -Reduced IL -Reduced MDA content -Reduced internal browning incidence -Promoted accumulation of proline, GABA and polyamines |
[127,128] |
| Mango | CaCl2 | -Enhanced antioxidant enzymatic activity | [139] |
| Peaches | Calcium-silicate CaCl2 Ca(NO3)2 Calcium sulphate (CaSO4) |
-Improved firmness -Reduced weight loss -Promoted bioactive compound content -Delayed softening -Limited rise of PG, PME, Cx, and β-Gal activities |
[131,132,133,134] |
| Sweet cherry | CaCl2 Ca(OH)2 Ca(NO3)2 Calcium caseinate |
-Improved firmness -Enhanced bioactive compounds content and antioxidant activity -Reduced cracking incidence |
[135,136] |
| Table grapes | Ca(NO3)2 CaCl₂ CaAs |
-Reduced weight loss -Reduced berry shattering -Reduced malic acid degradation -Reduced Botrytis cinerea incidence -Higher activity of antioxidant enzymes -Enhanced bioactive compounds content -Reduced MDA -Reduced IL -Inhibed ABA synthesis - Increased fruit firmness and Ca pectate -Inhibed ethylene production by supressing VvACO1 expression -Reduced cracking incidence |
[25,30,140,141,142,143,144,145,146] |
MDA- Malondialdehyde; ABA- Abscisic Acid; PG- Polygalacturonase; PME- Pectin Methylesterase; Cx- Cellulase; β-Gal- β-Galactosidase; GABA- Gamma-Aminobutyric Acid; VvACO1- 1-aminocyclopropane-1-carboxylate oxidase 1.
Table 2.
Physiological effects of applying mineral-sorbitol complexes to different plant.
| Plant | Minerals Applied | Effect | References |
| Mango | B Ca |
+ Total soluble solids +Carotenoids +Ascorbic acid +C: N ratio +Shelf-life +Yield -Acidity - Total soluble solids /acidity ratio |
[129,209] |
| Table grapes | Ca | + Total soluble solids + Total soluble solids /acidity ratio +Total Phenols +Colour and anthocyanidins -Berry shattering +Improved firmness -Respiration rate |
[3,25,177,210,211] |
| Wheat | Cu | +Enhanced nutrient uptake | [208] |
| Peanut | Ca | +Higher crop yield +Fat content +Mineral content +Dry matter |
[207] |
| Blood oranges | Ca | +Red colour of peel and pulp +Anthocyanin content +Naruritin and hesperidin content +Individual sugars accumulation +Enhanced organoleptic properties -K: Ca ratio |
[171,172] |
| Potatoes | Ca Zn |
+Tubers per plant +Tuber weight +Total yield +N, P, and K content |
[188,205] |
| Rice | Zn | +Yield +Protein synthesis +Seed production +Dry matter |
[212] |
| Melon | B | +Plant growth +Dry matter |
[213] |
| Lychee | B | +B absorption | [204] |
| Soybeans | B | +B absorption | [204,206] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



