
Submitted:
22 September 2025
Posted:
23 September 2025
You are already at the latest version
Abstract
We investigated whether recreational exercise (RE) influences systemic inflammation in Hashimoto’s thyroiditis (HT) across different disease severity groups. We analyzed 403 participants from the Croatian Biobank of Patients with HT (CROHT), including 173 controls and 230 HT patients (euthyroid, levothyroxine [LT4]-treated, and hypothyroid). Serum levels of 92 inflammatory proteins were measured using the Olink® Target 96 Inflammation panel, and exercise status was assessed via structured questionnaires. Linear regression revealed distinct protein associations depending on thyroid status. In controls, RE was associated with reduced MMP-10 and FGF-5, reflecting cardiovascular and muscle benefits. In euthyroid patients, RE was associated with decreased CXCL9 and TRAIL, implicating reduced type 1 inflammation and vascular risk. LT4-treated patients showed increases in IL-15RA and IL-24 with RE, suggesting improved muscle metabolism and anti-inflammatory effects. In hypothyroid patients, RE was associated with reduced CCL20 and increased HGF, while changes in TRANCE and TWEAK indicated mixed effects on bone and immune regulation. Notably, RE was associated with reduced CXCL9 and CCL20, two proteins previously linked to HT risk. Overall, RE is associated with distinct changes in inflammatory profiles across HT disease severity groups, with the most favourable responses observed in LT4-treated patients, suggesting synergy with hormone therapy.
Keywords:
autoimmune thyroiditis
; physical activity
; Olink inflammatory proteins
; CXCL9
; CCL20
; IL-24
; IL-15RA
1. Introduction
Hashimoto’s thyroiditis (HT) is one of the most common autoimmune disorders with marked prevalence in women. The pathogenesis of HT is characterised by lymphocytic infiltration of thyroid tissue and the production of thyroglobulin and thyroid-peroxidase antibodies (TgAb and TPOAb, respectively). Various cytokines, particularly tumour necrosis factor (TNF), interleukins (ILs), and CC chemokines, play important roles in the onset and modulation of the immune response in HT[1,2]. Chronic immune-mediated destruction of thyroid tissue leads to reduced thyroid hormone secretion that ultimately results in hypothyroidism. Although levothyroxine (LT4) replacement therapy remains the standard treatment for hypothyroidism, many patients continue to experience residual symptoms and immune dysregulation despite achieving biochemical euthyroidism[3,4]. In fact, HT is frequently associated with a broad spectrum of systemic effects, including symptom load, altered immune function, cardiovascular risk, and musculoskeletal impairment[5,6]. For this reason, many studies have focused on investigating ways to improve the overall health of patients with HT, including lifestyle modifications such as adopting regular physical activity.
Recent studies have highlighted the potential of “structured physical activity” in improving health outcomes in thyroid disease. For example, a meta-analysis of seven randomized controlled trials reported that long-term exercise interventions, when combined with standard therapy, significantly reduced serum thyroid-stimulating hormone (TSH) and increased free thyroxine (fT4) levels, suggesting that exercise may serve as an effective non-pharmacological addition to standard therapy in hypothyroidism management[7]. Moreover, regular daily physical activity was associated with improved immune parameters, such as lower levels of inflammatory markers (CRP and fibrinogen) and decreased counts of certain white blood cells in men and women from the large NANHES cohort[8]. The data on the potential anti-inflammatory effect of physical activity in HT is limited; however, the effect of regular exercise on modulating cytokine levels and other inflammatory markers was shown for various other autoimmune diseases[9,10]. In brief, physical activity was shown to increase T-regulatory cells, reduce immunoglobulin secretion, and shift the Th1/Th2 balance toward lower Th1 activity. It was also found to stimulate muscle-derived IL-6, which acts as a myokine to promote anti-inflammatory effects via IL-10 induction and IL-1β inhibition[11]. A 20-year systematic review of exercise effects on autoimmune diseases supports the development of exercise intervention programmes for autoimmune disease patients due to its beneficial anti-inflammatory effect[12].
Given the largely unknown molecular changes induced by physical activity in HT, particularly those involving inflammation and immune modulation, we conducted a study to address this gap. We investigated whether recreational exercise (RE) can modulate systemic inflammation in individuals with HT by using a comprehensive Olink® Target 96 Inflammation panel, a high-throughput proteomic platform enabling sensitive and multiplexed detection of immune proteins. Serum samples were obtained from the Croatian biobank of patients with Hashimoto’s thyroiditis (CRO-HT), a nationally recognized and deeply phenotyped biorepository focused on autoimmune thyroiditis in the Croatian population[13,14]. This unique resource provides an ideal foundation for integrative, translational analyses linking lifestyle factors to molecular disease signatures.
The aim of this study was to examine the effect of RE on inflammatory protein levels in individuals with HT, considering both the presence and progression of the disease. The ultimate goal was to determine whether RE can have beneficial effects in reducing inflammation by modulating immunopathological pathways associated with HT.
2. Materials and Methods
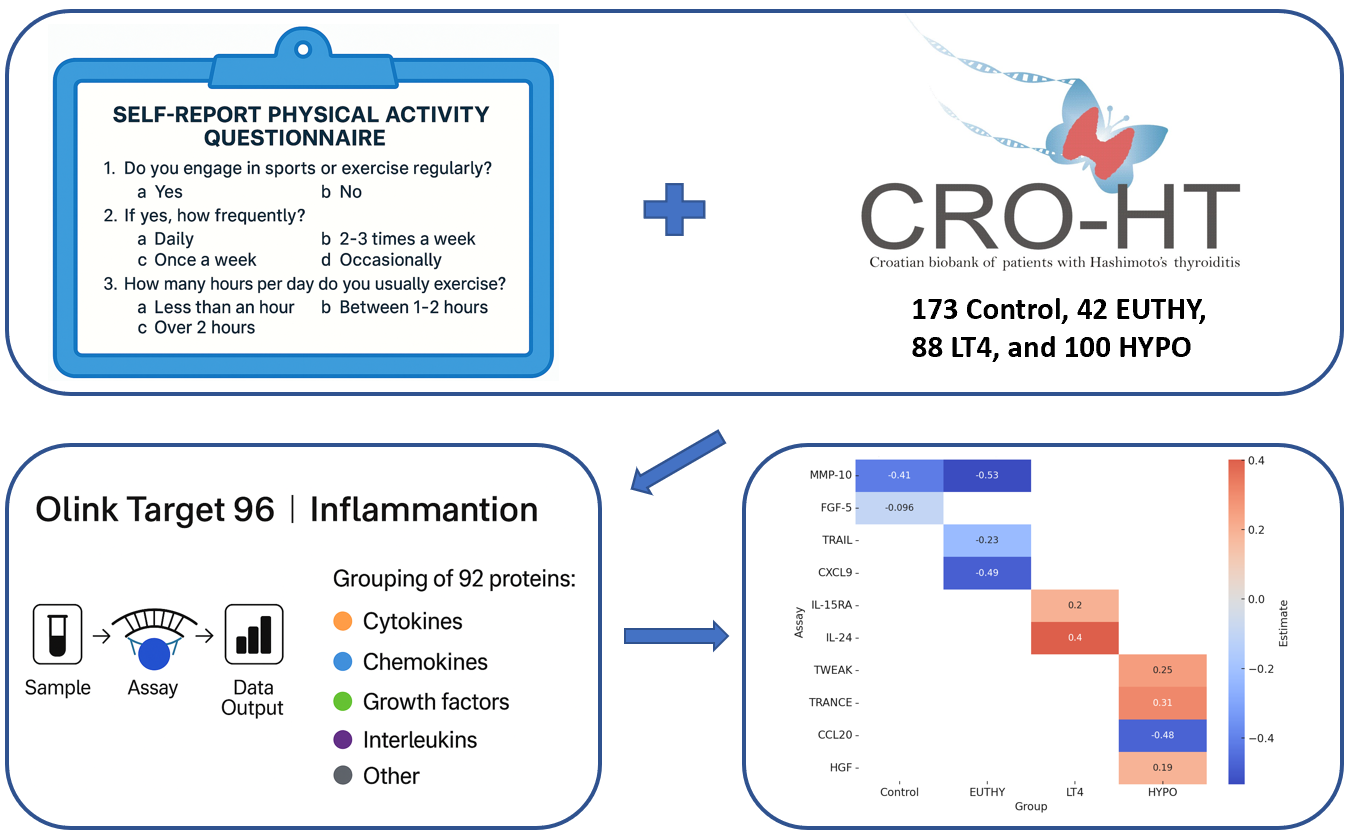
Subjects: This study included a total of 403 individuals: 230 individuals with HT (93% female) and 173 control participants (94.2% female) from the CRO-HT biobank[14]. The enrolment of study participants was performed by nuclear medicine specialists at the Outpatient Clinic for Thyroid Disorders, Clinical Department of Nuclear Medicine at University Hospital of Split. Diagnosis of HT followed established clinical guidelines[15], with participants undergoing clinical and biochemical evaluation. Controls were also selected based on clinical and laboratory examinations, and were required to be euthyroid with no signs of HT or other thyroid disorders. Additionally, structured questionnaires were administered to gather detailed information on personal and family history of autoimmune diseases, medical background, and various environmental and lifestyle factors, including diet, physical activity, smoking habits, and reproductive health. All participants were adults of white European descent living in iodine-sufficient southern Croatia[16]. Further details on the biobank, diagnostic criteria, and phenotype collection methods are available in our published studies[17,18].
We gathered data on RE using a self-report questionnaire consisting of three items: (1) Do you engage in sports or exercise regularly? (2) If yes, how frequently? (3) How many hours per day do you usually exercise? The scoring criteria for RE are presented in Table 1. For analytical purposes, we grouped participants into two groups: those who indicated no engagement in sports or reported only occasional sport activity were grouped in the “Without RE” group, whereas those who exercised (score 4, 8, or 16) were grouped in the “RE” group.
Blood samples were collected from all participants, and levels of TSH and thyroid hormones, including triiodothyronine (T3), thyroxine (T4), fT4, and fT3, as well as TgAb and TPOAb antibodies, were measured using LIAISON chemiluminescence immunoassays (DiaSorin Saluggia, Italy).
The Olink Target 96 Inflammation panel was used to measure 92 inflammatory proteins using the proximity extension assays (PEA) from the stored serum (at −80°C). The protein levels were expressed in Normalized Protein Expression (NPX) values, reflecting the relative abundance of each protein. Samples were successfully measured, with 95% passing quality control. Details on PEA technology can be found in a published study[19].
Statistical analyses. We divided HT patients into 3 groups based on their disease severity: 1) EUTHY group with 42 euthyroid HT patients (TSH, 0.3–3.6 mlU/L); 2) LT4 group with 88 patients receiving LT4 therapy, and 3) HYPO group with 100 hypothyroid patients (TSH>3.6 mlU/L). For analytical purposes, the groups were further divided into subgroups based on exercise status - those that perform exercise and those that do not (RE or Without RE). To assess differences in median RE values between the subgroups within each severity group, a Kruskal-Wallis test was performed, followed by pairwise Wilcoxon rank sum tests. Linear regression models were used to examine the association between NPX levels and exercise status (RE and Without RE), adjusting for variables including age, gender, and BMI, within each group (Control, EUTHY, LT4, HYPO). For each assay, the models were fitted separately, and p-values were calculated for each predictor, with p-values adjusted using the Benjamini-Hochberg method to control for multiple comparisons.
3. Results
Clinical characteristics of the Control group and the three groups of patients are shown in Table 2. The distribution of individuals who perform exercise and those who do not (RE or Without RE), across all groups, is shown in Figure 1. The proportion of individuals engaged in RE is highest in the Control group and decreases toward the HYPO group, while the opposite trend is observed for non-exercisers, with the lowest proportion in Controls and the highest in HYPO (Figure 1). In line, the Kruskal-Wallis test shows statistically significant difference in the median RE values among all four groups (χ² = 12.025, df = 3, p = 0.0073), while pairwise comparisons identified a significant difference between the Control and HYPO (p = 0.0043), while no significant differences are found between the other pairwise comparisons.
In Table 3, we present significant proteins that are differentially expressed between individuals who engage in RE and those who do not, across 4 groups. Among Controls, RE was linked to reduced MMP-10 and FGF-5. In EUTHY patients, RE was associated with decreased MMP-10, CXCL9, and TRAIL. In LT4 patients, RE was associated with increased IL-15RA and IL-24, while in HYPO patients, RE was associated with increased TWEAK, TRANCE, and HGF, and a decreased CCL20. Only proteins with statistically significant adjusted p-values were included in the main results, while the results for all 92 proteins across 4 groups are provided in Supplementary Table S1.
4. Discussion
Through the analysis of 92 inflammation-related proteins in individuals from the CRO-HT biobank, we identified distinct proteins associated with RE in both healthy subjects and HT patients, stratified according to disease severity. Many identified proteins overlap with molecular pathways involved with cardiovascular regulation, musculoskeletal health, and thyroid autoimmunity.
To gain deeper insight into the potential immunomodulatory effects of physical activity, we compared our findings with those from our previous study conducted on the same cohort, which identified proteins associated with the risk of developing HT. Notably, two proteins—CXCL9 and CCL20—previously linked to increased risk for HT in our CRO-HT biobank (manuscript under review, shown in Supplementary Table S2) are also significantly associated with RE status in our current analysis. Interestingly, both exhibited opposite effects: they showed upregulation with the advancement of HT but also downregulation associated with physical activity. Also, one of the proteins with the greatest fold increase, IL-24, is known for its tolerogenic properties in autoimmunity[20]. Therefore, our results highlight the potential of physical activity to counteract autoimmune processes relevant to HT.
In the text below, we discuss all our findings in the light of the main functions of significant proteins, with a focus on their immunological and other physiological functions potentially influenced by physical activity.
Controls: we observed a modest reduction in circulating levels of two proteins in healthy individuals who are physically active compared to non-active controls: matrix metalloproteinase-10 (MMP-10) with a 1.04-fold decrease in mean NPX and fibroblast growth factor 5 (FGF-5) with a 1.36-fold decrease in mean NPX.
MMP-10 is a zinc-dependent endopeptidase involved in extracellular matrix (ECM) degradation and in physiological processes like development, reproduction, and tissue remodelling[21]. While MMP-10 has not been directly linked to physical activity, MMP-2 and MMP-9 were observed to decrease with aerobic exercise and weight loss, indicating reduced inflammation and ECM activity[22]. On the contrary, elevated MMP-10 is associated with cardiovascular disease (CVD), promoting inflammation, plaque formation, and heart failure progression[23,24,25]. To summarise, our data show that exercise is associated with MMP-10 reduction that may potentially contribute to cardiovascular protection, reduced inflammation, and reduced ECM. The same effect is also seen in our euthyroid HT patients undergoing RE.
FGF-5 is a glycosaminoglycan-binding protein that plays roles in embryonic development, cell proliferation, morphogenesis, tissue repair, and tumour progression. It is particularly relevant to skeletal muscle, where it promotes fibroblast proliferation and inhibits muscle development[26], modulates metabolism during myogenesis[27], and suppresses muscle fiber growth, as shown in FGF5 knockout models[28]. Its reduction with exercise may support muscle hypertrophy and improved skeletal-muscular balance by reducing fibroblast proliferation and shifting metabolism towards greater energy efficiency during physical activity. Our findings are the first to indicate potential modulation of FGF-5 with physical activity. On the other side, a more commonly studied protein in the context of exercise, FGF-21[29,30,31], did not show a significant change in our cohort (Supplementary Table S1).
EUTHY: we observed a modest reduction in circulating levels of three proteins in physically active euthyroid patients with HT: already mentioned MMP-10 with a 1.055-fold decrease in mean NPX, TNF-related apoptosis-inducing ligand (TRAIL) with a 1.028-fold decrease in mean NPX, and C-X-C motif chemokine ligand 9 (CXCL9) with a 1.07-fold reduction in mean NPX.
TRAIL is a cytokine that induces apoptosis in tumour cells, and plays a role in vascular biology, exhibiting anti-inflammatory and anti-atherosclerotic effects[32]. Clinically, lower TRAIL levels are linked to higher CVD risk and mortality[33,34]. While little is known about TRAIL’s response to physical activity, one study in obese individuals found no significant change following long-term exercise[35]. Our finding of decreased TRAIL with RE is unexpected, given exercise’s cardiovascular benefits, and may reflect unique immune or metabolic dynamics in euthyroid HT, a population already at increased cardiovascular risk[6,36]. Further research is needed to clarify the mechanisms underlying this inverse relationship.
CXCL9 is a pro-inflammatory cytokine induced by interferon-gamma and is central to type 1 (T1) inflammatory responses[37]. Elevated CXCL9 levels have been linked to reduced musculoskeletal and physical function, lower activity levels, and greater frailty in older adults[38,39,40], while exercise has been shown to lower CXCL9 and improve metabolic and immune parameters in obesity[41]. As already pointed out, we previously observed a marked increase in CXCL9 levels in patients who progress from an euthyroid to a hypothyroid state. These findings suggest that physical activity may help regulate CXCL9 signalling, potentially improving musculoskeletal function and reducing chronic inflammation, and delaying hypothyroidism in HT.
LT4 group: we observed a marked increase in circulating levels of two proteins in physically active HT patients that are treated with LT4, compared to the inactive ones: interleukin 15 receptor subunit alpha (IL-15RA) with a 1.54-fold increase in mean NPX and interleukin 24 (IL-24) with a bold 5.3-fold increase in mean NPX.
IL-15RA is a pro-inflammatory cytokine receptor that binds IL-15 with high affinity, promoting activation of CD8+ T cells and NK cells[42,43]. It also regulates IL-15 production, secretion, and stability[44]. Beyond its role in immunology, IL-15 is involved in muscle metabolism, contributing to fat loss, improved insulin sensitivity, and reduced hepatic steatosis[45,46]. Consistent with our results, a meta-analysis showed that acute, but not chronic, exercise increases IL-15 levels shortly after activity[47]. Our results suggest that increased IL-15RA may contribute to enhanced metabolic outcomes in physically active LT4-treated patients.
IL-24 is a member of the IL-10 cytokine family with both pro- and anti-inflammatory roles and roles in tumour suppression[48]. IL-24 is also implicated in autoimmunity, where it helps regulate pathogenic Th17 responses that may aid in resolving tissue inflammation[20]. To date, IL-24 has not been investigated in the context of physical activity, rendering this a novel finding that highlights its potential role as a mediator of RE–induced immunoregulation.
HYPO group: we observed an increase in circulating levels of three proteins in physically active hypothyroid HT patients: TNF-related weak inducer of apoptosis (TWEAK) with 1.028-fold increase in mean NPX, tumor necrosis factor ligand superfamily member 11 (TNFSF11 or TRANCE) with a 1.063-fold increase in mean NPX values, and hepatocyte growth factor (HGF) with a 1.016-fold increase in mean NPX values. We also observed a decrease in C-C motif chemokine ligand 20 (CCL20) with a 1.075-fold decrease in mean NPX values.
TWEAK is a cytokine from the TNF ligand superfamily that exists in soluble and membrane-bound forms and is involved in apoptosis, inflammation (e.g., IL-8 induction), and angiogenesis via endothelial cell proliferation and migration[49,50,51]. Regular physical activity has been shown to increase circulating soluble TWEAK (sTWEAK), potentially helping balance pro- and anti-inflammatory responses[52]. However, TWEAK was also found to impair skeletal muscle by reducing mitochondrial content and angiogenesis[53,54]. Thus, in skeletal muscle tissue, TWEAK suppresses angiogenesis and promotes muscle wasting and impaired regeneration. This contrasts with its pro-angiogenic role seen in other tissues or contexts. These findings, along with our data, highlight TWEAK’s complex, context-dependent roles in inflammation and metabolism, warranting further study in hypothyroid populations undergoing exercise.
TRANCE regulates immune responses and bone resorption by promoting osteoclast differentiation via RANK, counterbalanced by osteoprotegerin (OPG), which inhibits this interaction[55,56]. Disruption of the TRANCE–OPG axis—via elevated TRANCE or reduced OPG—contributes to bone loss, as seen in postmenopausal osteoporosis[57,58]. Although we observe a modest TRANCE increase with RE, we also observe a non-significant decrease in OPG and OPG/TRANCE ratio, suggesting a non-pathological TRANCE increase. With respect to exercise, a decrease in TRANCE is common[59], although it may vary by sex and age[60]. Thus, the modest TRANCE increase observed in our physically active hypothyroid patients may not be pathologic, but rather restorative, especially if balanced by unchanged or mildly decreased OPG.
CCL20 is a chemokine that plays a central role in immune regulation by recruiting immature dendritic cells, memory T cells, and B cells via CCR6[61]. As already mentioned, the CCL20 increase is associated with HT (manuscript under review). A key aspect of HT immunopathology involves Th17 cells[62], and emerging evidence suggests that CCL20 signalling can promote the conversion of regulatory T cells (Tregs) into pathogenic Th17 cells[63]. Thus, the observed reduction in CCL20 with RE may reflect exercise-induced mitigation of Th17-driven inflammation, particularly evident in hypothyroid HT patients. This aligns with prior studies reporting decreased CCL20 levels after physical activity, supporting its role in lowering systemic inflammation[64,65].
HGF is a pleiotropic growth factor that promotes epithelial proliferation, angiogenesis, and tissue repair, while also exerting anti-inflammatory and anti-apoptotic effects, particularly in the heart and brain[66]. It supports bone regeneration through enhanced osteoblast activity and neovascularization, though it may also influence bone resorption under specific conditions[67,68]. Exercise is known to transiently elevate HGF levels[69,70], which aligns with our study, and suggests that this response may aid cardiovascular, muscular, vascular, and bone health in hypothyroid individuals.
Taken together, this study offers compelling evidence that RE modulates inflammatory proteins involved in HT, with potential benefits extending to autoimmune regulation, cardiovascular health, bone metabolism, and muscle maintenance. Below, we summarise our findings and discuss them by integrating the roles of individual proteins with the effects of physical activity, highlighting their interconnected impact on multiple physiological processes.
Immunomodulation and autoimmunity: RE was associated with the downregulation of proinflammatory chemokines CXCL9 and CCL20, both linked to HT risk in our previous study. The upregulation of IL-24, a cytokine with anti-inflammatory and immunoregulatory roles in Th17 cells, supports a broader anti-autoimmune shift potentially induced by RE.
Cardiometabolic regulation: Exercise-induced changes in MMP-10 and HGF point to improvements in cardiovascular and metabolic profiles. The downregulation of MMP-10, a protease involved in vascular inflammation, and upregulation of HGF, a tissue-reparative and angiogenic factor, suggest a cardioprotective shift. Although TRAIL showed a paradoxical decrease, its interpretation may require further investigation within the unique immunometabolic context of euthyroid HT.
Skeletal muscle and metabolic adaptation: The observed decrease in FGF-5, a known inhibitor of muscle growth and regulator of fibroblast activity, alongside an increase in IL-15RA, which promotes muscle metabolism and insulin sensitivity, suggests that RE supports muscle hypertrophy and metabolic efficiency—particularly in LT4-treated patients. Reduced CXCL9 also associates with musculoskeletal benefits.
Osteoimmunological Crosstalk: Proteins such as TRANCE, CCL20, and HGF—each with dual roles in inflammation and bone remodelling—responded to RE in HT patients, highlighting the intersection between immune and skeletal systems. These findings reinforce the concept of osteoimmunology, particularly relevant in hypothyroid patients, where both bone metabolism and immune balance are dysregulated.
Finally, we also observe differences in inflammatory profiles associated with RE depending on the disease severity group. For example, in healthy controls, RE led to favourable but expected changes, such as reductions in MMP-10 and FGF-5, reflecting improved cardiovascular and muscular profiles. However, since these participants are euthyroid and disease-free, the clinical significance of these changes is relatively limited compared to the HT subgroups. The LT4 group demonstrates the clearest, most favourable, and least ambiguous molecular responses to RE—spanning metabolic, muscular, and immunoregulatory domains. These responses suggest that exercise acts synergistically with hormone replacement, potentially restoring a more balanced physiological state than either treatment alone. In contrast, euthyroid HT patients exhibited a potentially adverse drop in TRAIL, which may reflect a vulnerable state—HT pathology without hormonal compensation (as in LT4). Untreated hypothyroid patients showed mixed responses—some beneficial (increase in HGF and decrease in CCL20), others ambiguous (increase in TWEAK and in TRANCE), raising concerns for bone health and inflammation. These findings highlight group-specific effects of exercise, with LT4-treated individuals emerging as the most responsive and physiologically balanced subgroup.
Advantages and limitations: The use of the same well-defined cohort across both the current RE-focused analysis and a previous study on HT risk enables a unique within-subject comparison, revealing biologically meaningful and opposing trends in key proteins such as CXCL9 and CCL20. However, certain limitations should be acknowledged. The observational nature of the study precludes causal inference, and the absence of longitudinal or functional clinical outcomes limits interpretation regarding long-term or symptomatic benefits. Finally, our findings require validation in independent cohorts.
5. Conclusions
In our study, we identified several proteins known to be associated with cardiovascular/ metabolic (MMP-10, HGF, TRAIL, IL-15RA) and skeletal systems (TRANCE, CCL20, HGF), indicating that the physiological phenotype of RE is well captured at the molecular level. The opposing regulation of HT-risk proteins (CXCL9 and CCL20) by RE is particularly noteworthy and supports the hypothesis that physical activity could mitigate autoimmune progression. Finally, this study shows the combined beneficial effects of LT4 treatment and RE in patients with HT.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Table S1: The results of the linear regression model between NPX levels of 92 inflammatory proteins and RE status across 4 groups. Supplementary Table S2: The results of ANOVA analysis between NPX values and HT-disease status.
Author Contributions
Conceptualization, M.V. and V.B.P.; Methodology, M.V., V.B.P. and V.Ž.; Software, V.Ž.; Validation, M.V., V.B.P. and V.Ž.; Formal Analysis, V.Ž.; Investigation, M.V., V.B.P., D.K., A.B.Ž., S.S., V.T.L., M.C. and A.P.; Resources, V.B.P.; Data Curation, V.B.P.; Writing – Original Draft Preparation, M.V., V.Ž. and V.B.P.; Writing – Review & Editing, A.B.Ž., S.S., D.K., V.T.L., M.C. and A.P.; Visualization, M.V., V.Ž. and V.B.P.; Supervision, V.B.P. and A.P.; Project Administration, V.B.P.; Funding Acquisition, V.B.P.
Funding
The formation of the CROHT biobank and measurements of Olink proteins were funded by the Croatian Science Foundation under projects UIP-11-2013-4950 and IP-2022-10-4071.
Institutional Review Board Statement
Study was approved by the Ethics Committee of the University of Split, School of Medicine (Classification no. 003-08/14-03/0001 and Registry no. 2181-198-03-04-14-0028; Classification no. 003-08/19-03/0003 and Reg. no. 2181-198-03-04-19-0019) and the Ethics Committee of University Hospital Split (Classification no. 530-02/13-01/11; Registry no. 2181-147-01/06/J.B.-14-2; Classification no. 500-03/18-01/80 and Reg. no. 2181-147-01/06/M.S.-18-2). The study’s compliance with the Code of Ethics and the Helsinki Declaration was confirmed by both Ethics Committees.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data supporting the conclusions of this article will be made available by the corresponding author upon request.
Acknowledgments
The authors have reviewed and edited the output and take full responsibility for the content of this publication. AI tools (such as ChatGPT and Perplexity) were utilized to enhance the English language quality and refine the writing style of the authors’ text.
Conflicts of Interest
The authors declare no conflict of interest.
References
- K. Wrońska, M. Hałasa, and M. Szczuko, “The Role of the Immune System in the Course of Hashimoto’s Thyroiditis: The Current State of Knowledge.,” Int J Mol Sci, vol. 25, no. 13, Jun. 2024. [CrossRef]
- Q.-Y. Zhang et al., “Lymphocyte infiltration and thyrocyte destruction are driven by stromal and immune cell components in Hashimoto’s thyroiditis.,” Nat Commun, vol. 13, no. 1, p. 775, Feb. 2022. [CrossRef]
- K. L. Groenewegen, C. F. Mooij, and A. S. P. van Trotsenburg, “Persisting symptoms in patients with Hashimoto’s disease despite normal thyroid hormone levels: Does thyroid autoimmunity play a role? A systematic review.,” J Transl Autoimmun, vol. 4, p. 100101, 2021. [CrossRef]
- S. Razvi, S. Mrabeti, and M. Luster, “Managing symptoms in hypothyroid patients on adequate levothyroxine: a narrative review.,” Endocr Connect, vol. 9, no. 11, pp. R241–R250, Nov. 2020. [CrossRef]
- R. M. McLean and D. N. Podell, “Bone and joint manifestations of hypothyroidism.,” Semin Arthritis Rheum, vol. 24, no. 4, pp. 282–90, Feb. 1995. [CrossRef]
- W.-H. Chen, Y.-K. Chen, C.-L. Lin, J.-H. Yeh, and C.-H. Kao, “Hashimoto’s thyroiditis, risk of coronary heart disease, and L-thyroxine treatment: a nationwide cohort study.,” J Clin Endocrinol Metab, vol. 100, no. 1, pp. 109–14, Jan. 2015. [CrossRef]
- H. Sundus, S. A. Khan, S. Zaidi, C. Chhabra, I. Ahmad, and H. Khan, “Effect of long-term exercise-based interventions on thyroid function in hypothyroidism: A systematic review and meta-analysis of randomized controlled trials.,” Complement Ther Med, vol. 92, p. 103196, Sep. 2025. [CrossRef]
- C. L. Klasson, S. Sadhir, and H. Pontzer, “Daily physical activity is negatively associated with thyroid hormone levels, inflammation, and immune system markers among men and women in the NHANES dataset.,” PLoS One, vol. 17, no. 7, p. e0270221, 2022. [CrossRef]
- G. S. Metsios, R. H. Moe, and G. D. Kitas, “Exercise and inflammation.,” Best Pract Res Clin Rheumatol, vol. 34, no. 2, p. 101504, Apr. 2020. [CrossRef]
- C. Weyh, K. Krüger, and B. Strasser, “Physical Activity and Diet Shape the Immune System during Aging.,” Nutrients, vol. 12, no. 3, Feb. 2020. [CrossRef]
- K. Sharif, A. Watad, N. L. Bragazzi, M. Lichtbroun, H. Amital, and Y. Shoenfeld, “Physical activity and autoimmune diseases: Get moving and manage the disease.,” Autoimmun Rev, vol. 17, no. 1, pp. 53–72, Jan. 2018. [CrossRef]
- B. Luo et al., “The anti-inflammatory effects of exercise on autoimmune diseases: A 20-year systematic review.,” J Sport Health Sci, vol. 13, no. 3, pp. 353–367, May 2024. [CrossRef]
- M. Cvek et al., “Vitamin D and Hashimoto’s Thyroiditis: Observations from CROHT Biobank,” Nutrients, vol. 13, no. 8, p. 2793, Aug. 2021. [CrossRef]
- L. Brčić et al., “Genome-wide association analysis suggests novel loci for Hashimoto’s thyroiditis,” J Endocrinol Invest, vol. 42, no. 5, pp. 567–576, May 2019. [CrossRef]
- S. H. S. Pearce et al., “2013 ETA Guideline: Management of Subclinical Hypothyroidism,” Eur Thyroid J, vol. 2, no. 4, pp. 215–228, 2013. [CrossRef]
- Z. Kusić et al., “Croatia has reached iodine sufficiency,” J Endocrinol Invest, vol. 26, no. 8, pp. 738–742, Aug. 2003. [CrossRef]
- M. Cvek et al., “Presence or severity of Hashimoto’s thyroiditis does not influence basal calcitonin levels: observations from CROHT biobank,” J Endocrinol Invest, vol. 45, no. 3, pp. 597–605, Mar. 2022. [CrossRef]
- M. Cvek et al., “Vitamin D and Hashimoto’s Thyroiditis: Observations from CROHT Biobank.,” Nutrients, vol. 13, no. 8, Aug. 2021. [CrossRef]
- L. Wik et al., “Proximity Extension Assay in Combination with Next-Generation Sequencing for High-throughput Proteome-wide Analysis,” Molecular & Cellular Proteomics, vol. 20, p. 100168, 2021. [CrossRef]
- Y. Zhong, X. Zhang, and W. Chong, “Interleukin-24 Immunobiology and Its Roles in Inflammatory Diseases.,” Int J Mol Sci, vol. 23, no. 2, Jan. 2022. [CrossRef]
- J. Wozniak, J. Floege, T. Ostendorf, and A. Ludwig, “Key metalloproteinase-mediated pathways in the kidney.,” Nat Rev Nephrol, vol. 17, no. 8, pp. 513–527, Aug. 2021. [CrossRef]
- J. Jaoude and Y. Koh, “Matrix metalloproteinases in exercise and obesity.,” Vasc Health Risk Manag, vol. 12, pp. 287–95, 2016. [CrossRef]
- A. Purroy et al., “Matrix metalloproteinase-10 deficiency delays atherosclerosis progression and plaque calcification.,” Atherosclerosis, vol. 278, pp. 124–134, Nov. 2018. [CrossRef]
- L. Matilla et al., “A Role for MMP-10 (Matrix Metalloproteinase-10) in Calcific Aortic Valve Stenosis.,” Arterioscler Thromb Vasc Biol, vol. 40, no. 5, pp. 1370–1382, May 2020. [CrossRef]
- Y. Wei et al., “Type-specific dysregulation of matrix metalloproteinases and their tissue inhibitors in end-stage heart failure patients: relationship between MMP-10 and LV remodelling.,” J Cell Mol Med, vol. 15, no. 4, pp. 773–82, Apr. 2011. [CrossRef]
- K. L. Clase, P. J. Mitchell, P. J. Ward, C. M. Dorman, S. E. Johnson, and K. Hannon, “FGF5 stimulates expansion of connective tissue fibroblasts and inhibits skeletal muscle development in the limb.,” Dev Dyn, vol. 219, no. 3, pp. 368–80, Nov. 2000. [CrossRef]
- M. Chen et al., “Metabolic differences in MSTN and FGF5 dual-gene edited sheep muscle cells during myogenesis.,” BMC Genomics, vol. 25, no. 1, p. 637, Jun. 2024. [CrossRef]
- M.-M. Chen et al., “A MSTNDel73C mutation with FGF5 knockout sheep by CRISPR/Cas9 promotes skeletal muscle myofiber hyperplasia.,” Elife, vol. 12, Oct. 2024. [CrossRef]
- C. Liu et al., “The effects of exercise on FGF21 in adults: a systematic review and meta-analysis.,” PeerJ, vol. 12, p. e17615, 2024. [CrossRef]
- M. Peterson, K. A. Richardson, and L. Funderburk, “Effect of exercise on Fibroblast Growth Factor 21 levels in healthy males and females.,” PLoS One, vol. 20, no. 5, p. e0321738, 2025. [CrossRef]
- D. Cuevas-Ramos et al., “Exercise increases serum fibroblast growth factor 21 (FGF21) levels.,” PLoS One, vol. 7, no. 5, p. e38022, 2012. [CrossRef]
- F. Corallini, E. Rimondi, and P. Secchiero, “TRAIL and osteoprotegerin: a role in endothelial physiopathology?,” Front Biosci, vol. 13, pp. 135–47, Jan. 2008. [CrossRef]
- K. Kakareko, A. Rydzewska-Rosołowska, E. Zbroch, and T. Hryszko, “TRAIL and Cardiovascular Disease-A Risk Factor or Risk Marker: A Systematic Review.,” J Clin Med, vol. 10, no. 6, Mar. 2021. [CrossRef]
- G. Biolo, P. Secchiero, S. De Giorgi, V. Tisato, and G. Zauli, “The energy balance positively regulates the levels of circulating TNF-related apoptosis inducing ligand in humans.,” Clin Nutr, vol. 31, no. 6, pp. 1018–21, Dec. 2012. [CrossRef]
- C. Davenport, H. Kenny, D. T. Ashley, E. P. O’Sullivan, D. Smith, and D. J. O’Gorman, “The effect of exercise on osteoprotegerin and TNF-related apoptosis-inducing ligand in obese patients.,” Eur J Clin Invest, vol. 42, no. 11, pp. 1173–9, Nov. 2012. [CrossRef]
- P. İşgüven, Y. Gündüz, and M. Kılıç, “Effects of Thyroid Autoimmunity on Early Atherosclerosis in Euthyroid Girls with Hashimoto’s Thyroiditis.,” J Clin Res Pediatr Endocrinol, vol. 8, no. 2, pp. 150–6, Jun. 2016. [CrossRef]
- F. Liao, R. L. Rabin, J. R. Yannelli, L. G. Koniaris, P. Vanguri, and J. M. Farber, “Human Mig chemokine: biochemical and functional characterization.,” J Exp Med, vol. 182, no. 5, pp. 1301–14, Nov. 1995. [CrossRef]
- J. Li et al., “Association between Circulating Levels of CXCL9 and CXCL10 and Physical Frailty in Older Adults.,” Gerontology, vol. 70, no. 3, pp. 279–289, 2024. [CrossRef]
- D. H. Seo et al., “Chemokine CXCL9, a marker of inflammaging, is associated with changes of muscle strength and mortality in older men.,” Osteoporos Int, vol. 35, no. 10, pp. 1789–1796, Oct. 2024. [CrossRef]
- Y. C. Hu et al., “Physical activity affects DNA methylation-derived inflammation markers in a community-based Parkinson’s disease study.,” Brain Behav Immun Health, vol. 46, p. 101014, Jul. 2025. [CrossRef]
- W. He, H. Wang, G. Yang, L. Zhu, and X. Liu, “The Role of Chemokines in Obesity and Exercise-Induced Weight Loss.,” Biomolecules, vol. 14, no. 9, Sep. 2024. [CrossRef]
- D. M. Anderson et al., “Functional characterization of the human interleukin-15 receptor alpha chain and close linkage of IL15RA and IL2RA genes.,” J Biol Chem, vol. 270, no. 50, pp. 29862–9, Dec. 1995. [CrossRef]
- H. Lee, S.-H. Park, and E.-C. Shin, “IL-15 in T-Cell Responses and Immunopathogenesis.,” Immune Netw, vol. 24, no. 1, p. e11, Feb. 2024. [CrossRef]
- E. H. Duitman, Z. Orinska, E. Bulanova, R. Paus, and S. Bulfone-Paus, “How a cytokine is chaperoned through the secretory pathway by complexing with its own receptor: lessons from interleukin-15 (IL-15)/IL-15 receptor alpha.,” Mol Cell Biol, vol. 28, no. 15, pp. 4851–61, Aug. 2008. [CrossRef]
- S. Busquets, M. Figueras, V. Almendro, F. J. López-Soriano, and J. M. Argilés, “Interleukin-15 increases glucose uptake in skeletal muscle. An antidiabetogenic effect of the cytokine.,” Biochim Biophys Acta, vol. 1760, no. 11, pp. 1613–7, Nov. 2006. [CrossRef]
- H. Sun, Y. Ma, M. Gao, and D. Liu, “IL-15/sIL-15Rα gene transfer induces weight loss and improves glucose homeostasis in obese mice.,” Gene Ther, vol. 23, no. 4, pp. 349–56, Apr. 2016. [CrossRef]
- M. Khalafi et al., “Interleukin-15 responses to acute and chronic exercise in adults: a systematic review and meta-analysis.,” Front Immunol, vol. 14, p. 1288537, 2023. [CrossRef]
- L. Emdad et al., “Recent insights into apoptosis and toxic autophagy: The roles of MDA-7/IL-24, a multidimensional anti-cancer therapeutic.,” Semin Cancer Biol, vol. 66, pp. 140–154, Nov. 2020. [CrossRef]
- S. R. Wiley and J. A. Winkles, “TWEAK, a member of the TNF superfamily, is a multifunctional cytokine that binds the TweakR/Fn14 receptor.,” Cytokine Growth Factor Rev, vol. 14, no. 3–4, pp. 241–9, 2003. [CrossRef]
- C. N. Lynch, Y. C. Wang, J. K. Lund, Y. W. Chen, J. A. Leal, and S. R. Wiley, “TWEAK induces angiogenesis and proliferation of endothelial cells.,” J Biol Chem, vol. 274, no. 13, pp. 8455–9, Mar. 1999. [CrossRef]
- P. J. Donohue et al., “TWEAK is an endothelial cell growth and chemotactic factor that also potentiates FGF-2 and VEGF-A mitogenic activity.,” Arterioscler Thromb Vasc Biol, vol. 23, no. 4, pp. 594–600, Apr. 2003. [CrossRef]
- R. Schönbauer et al., “Regular Training Increases sTWEAK and Its Decoy Receptor sCD163-Does Training Trigger the sTWEAK/sCD163-Axis to Induce an Anti-Inflammatory Effect?,” J Clin Med, vol. 9, no. 6, Jun. 2020. [CrossRef]
- S. Sato et al., “TWEAK promotes exercise intolerance by decreasing skeletal muscle oxidative phosphorylation capacity.,” Skelet Muscle, vol. 3, no. 1, p. 18, Jul. 2013. [CrossRef]
- S. Sato, Y. Ogura, and A. Kumar, “TWEAK/Fn14 Signaling Axis Mediates Skeletal Muscle Atrophy and Metabolic Dysfunction.,” Front Immunol, vol. 5, p. 18, 2014. [CrossRef]
- T. Ono, M. Hayashi, F. Sasaki, and T. Nakashima, “RANKL biology: bone metabolism, the immune system, and beyond.,” Inflamm Regen, vol. 40, p. 2, 2020. [CrossRef]
- P. J. Kostenuik, “Osteoprotegerin and RANKL regulate bone resorption, density, geometry and strength.,” Curr Opin Pharmacol, vol. 5, no. 6, pp. 618–25, Dec. 2005. [CrossRef]
- S. Jabbar, J. Drury, J. N. Fordham, H. K. Datta, R. M. Francis, and S. P. Tuck, “Osteoprotegerin, RANKL and bone turnover in postmenopausal osteoporosis.,” J Clin Pathol, vol. 64, no. 4, pp. 354–7, Apr. 2011. [CrossRef]
- G. Chi, L. Qiu, J. Ma, W. Wu, and Y. Zhang, “The association of osteoprotegerin and RANKL with osteoporosis: a systematic review with meta-analysis.,” J Orthop Surg Res, vol. 18, no. 1, p. 839, Nov. 2023. [CrossRef]
- M. Tobeiha, M. H. Moghadasian, N. Amin, and S. Jafarnejad, “RANKL/RANK/OPG Pathway: A Mechanism Involved in Exercise-Induced Bone Remodeling.,” Biomed Res Int, vol. 2020, p. 6910312, 2020. [CrossRef]
- E. A. Marques et al., “Effects of resistance and aerobic exercise on physical function, bone mineral density, OPG and RANKL in older women.,” Exp Gerontol, vol. 46, no. 7, pp. 524–32, Jul. 2011. [CrossRef]
- A. Y. S. Lee, R. Eri, A. B. Lyons, M. C. Grimm, and H. Korner, “CC Chemokine Ligand 20 and Its Cognate Receptor CCR6 in Mucosal T Cell Immunology and Inflammatory Bowel Disease: Odd Couple or Axis of Evil?,” Front Immunol, vol. 4, p. 194, 2013. [CrossRef]
- Z.-R. Shi et al., “The Chemokine, CCL20, and Its Receptor, CCR6, in the Pathogenesis and Treatment of Psoriasis and Psoriatic Arthritis.,” J Psoriasis Psoriatic Arthritis, vol. 8, no. 3, pp. 107–117, Jul. 2023. [CrossRef]
- N. Kulkarni et al., “CCR6 signaling inhibits suppressor function of induced-Treg during gut inflammation.,” J Autoimmun, vol. 88, pp. 121–130, Mar. 2018. [CrossRef]
- A. K. Ziegler, S. M. Jensen, P. Schjerling, A. L. Mackey, J. L. Andersen, and M. Kjaer, “The effect of resistance exercise upon age-related systemic and local skeletal muscle inflammation.,” Exp Gerontol, vol. 121, pp. 19–32, Jul. 2019. [CrossRef]
- ”Effects of Resistance Exercise Intensity on Cytokine and Chemokine Gene Expression in Atopic Dermatitis Mouse Model,” J Mens Health, vol. 14, no. 2, p. 14, 2018. [CrossRef]
- K. Chen, K. K. Aenlle, K. M. Curtis, B. A. Roos, and G. A. Howard, “Hepatocyte growth factor (HGF) and 1,25-dihydroxyvitamin D together stimulate human bone marrow-derived stem cells toward the osteogenic phenotype by HGF-induced up-regulation of VDR.,” Bone, vol. 51, no. 1, pp. 69–77, Jul. 2012. [CrossRef]
- R. Zhen et al., “Hepatocyte growth factor improves bone regeneration via the bone morphogenetic protein-2-mediated NF-κB signaling pathway.,” Mol Med Rep, vol. 17, no. 4, pp. 6045–6053, Apr. 2018. [CrossRef]
- C. Huang et al., “Hepatocyte growth factor overexpression promotes osteoclastogenesis and exacerbates bone loss in CIA mice.,” J Orthop Translat, vol. 27, pp. 9–16, Mar. 2021. [CrossRef]
- S. Yasuda et al., “Exercise-induced hepatocyte growth factor production in patients after acute myocardial infarction: its relationship to exercise capacity and brain natriuretic peptide levels.,” Circ J, vol. 68, no. 4, pp. 304–7, Apr. 2004. [CrossRef]
- P. Wahl et al., “Effects of high intensity training and high volume training on endothelial microparticles and angiogenic growth factors.,” PLoS One, vol. 9, no. 4, p. e96024, 2014. [CrossRef]
Figure 1.
Distribution of individuals engaged in recreational exercise (light blue) and those not engaged (dark blue) across the four study groups.
Figure 1.
Distribution of individuals engaged in recreational exercise (light blue) and those not engaged (dark blue) across the four study groups.
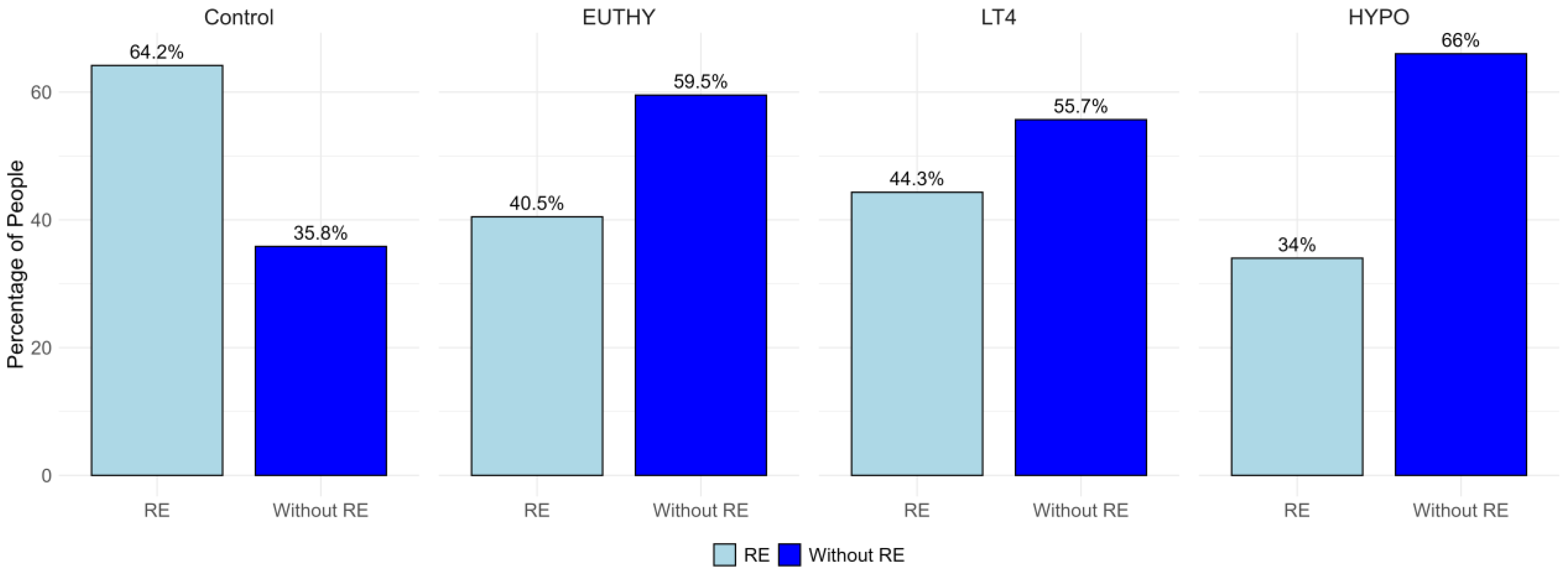

Table 1.
Calculation of the score for recreational exercise (RE).
| Less than an hour | Between 1-2 hours | Over 2 hours | |
|---|---|---|---|
| Daily | 16 | 16 | 16 |
| 2-3 times a week | 8 | 16 | 16 |
| Once a week | 4 | 8 | 8 |
| Occasionally | 0 | 0 | 0 |
Table 2.
Clinical characteristics of controls and HT patients across three disease severity groups.
| Phenotype | Control | EUTHY | LT4 | HYPO |
|---|---|---|---|---|
| N = 173 | N = 42 | N = 88 | N = 100 | |
| Mean (SD) | Mean (SD) | Mean (SD) | Mean (SD) | |
| Age, years | 39,24 (11,88) | 34,39 (12,67) | 40,48 (13,87) | 39,05 (13,22) |
| BMI, kg/m² | 23,58 (3,92) | 23,80 (3,80) | 24,21 (4,25) | 24,19 (4,22) |
| T3, nmol/L | 1,55 (0,22) | 1,62 (0,42) | 1,64 (0,32) | 1,51 (0,39) |
| T4, nmol/L | 102,92 (20,30) | 98,52 (22,92) | 114,47 (26,78) | 94,18 (27,05) |
| fT4, pmol/L | 12,89 (1,62) | 12,64 (1,53) | 13,09 (2,40) | 10,50 (2,50) |
| TSH, mIU/L | 1,61 (0,67) | 2,11 (0,96) | 4,89 (8,78) | 15,89 (24,93) |
| TgAt, IU/ml | 17,36 (17,46) | 730,52 (1177,64) | 575,48 (1126,23) | 618,80 (1048,81) |
| TpOAb, IU/ml | 5,73 (5,88) | 522,37 (483,10) | 546,01 (575,37) | 620,51 (692,42) |
| RE, hours/month | 5,80 (5,68) | 4,57 (6,34) | 5,45 (6,79) | 4,00 (6,15) |
*RE – recreational exercise.
Table 3.
Significant Associations Between Inflammatory Proteins and Recreational Exercise Across Study Groups.
Table 3.
Significant Associations Between Inflammatory Proteins and Recreational Exercise Across Study Groups.
| Control (RE = 111, Without RE = 62) | |||
| Assay | Estimate* | p-value | adjusted p-value |
| MMP-10 | -0,4133 | 0,0026 | 0,0065 |
| FGF-5 | -0,0957 | 0,0058 | 0,0292 |
| EUTHY (RE = 17, Without RE = 25) | |||
| Assay | Estimate | p-value | adjusted p-value |
| MMP-10 | -0,5335 | 0,0070 | 0,0175 |
| TRAIL | -0,2298 | 0,0150 | 0,0375 |
| CXCL9 | -0,4858 | 0,0165 | 0,0412 |
| LT4 (RE = 39, Without RE = 49) | |||
| Assay | Estimate | p-value | adjusted p-value |
| IL-15RA | 0,1958 | 0,0042 | 0,0126 |
| IL-24 | 0,4020 | 0,0098 | 0,0163 |
| HYPO (RE = 34, Without RE = 66) | |||
| Assay | Estimate | p-value | adjusted p-value |
| TWEAK | 0,2526 | 0,0019 | 0,0047 |
| TRANCE | 0,3096 | 0,0272 | 0,0340 |
| CCL20 | -0,4781 | 0,0218 | 0,0364 |
| HGF | 0,1936 | 0,0275 | 0,0459 |
*The estimate for exercise status represents the difference in NPX levels between individuals who engage in RE and those who do not.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



