
Submitted:
21 September 2025
Posted:
22 September 2025
You are already at the latest version
Abstract
Monensin, a polyether ionophore widely used in veterinary medicine, has recently emerged as a multimodal anticancer candidate. This review provides a comprehensive overview of preclinical studies showing that monensin exerts selective cytotoxic and anti-progression effects across a wide range of malignancies. Its activity spans breast cancer to leukemia, reflecting both versatility and significant therapeutic potential. In breast cancer, monensin reduces proliferation, induces apoptosis, and enhances chemosensitivity, suggesting a role in combination therapies. In prostate cancer, it disrupts androgen receptor signaling, induces oxidative stress, and triggers mitochondria-dependent apoptosis. In pancreatic cancer, it suppresses EGFR signaling and promotes programmed cell death, while in ovarian and cervical cancers, it inhibits proliferation, migration, and invasion by modulating EGFR and MEK/ERK pathways and enhancing SUMOylation. In renal carcinoma, monensin induces cell-cycle arrest, autophagy, and apoptosis, whereas in bladder and squamous cell carcinoma, it interferes with EGFR-related signaling and lectin-mediated interactions. It selectively kills liver cancer cells through intracellular Na⁺ overload and mitochondrial damage, and in thyroid cancer, it disrupts cellular respiration and AMPK/mTOR signaling. In glioblastoma, it exhibits both anti-tumor and anti-angiogenic effects, while in hematologic malignancies such as leukemia and lymphoma, it induces apoptosis, cell-cycle arrest, and glycosylation alterations. Collectively, these findings highlight monensin as a highly promising, broad-spectrum anticancer candidate. Its multifaceted mechanisms of action and consistent efficacy across diverse tumor types provide a compelling rationale for continued preclinical evaluation and potential clinical translation. These insights position monensin as an innovative therapeutic avenue, offering new hope for the development of versatile and effective cancer treatments.
Keywords:
ionophore antibiotics
; apoptosis
; cell-cycle arrest
; EGFR/MEK/ERK signaling
; oxidative stress
; breast cancer
; prostate cancer
; glioblastoma
1. Introduction
Cancer remains one of the most significant health challenges in the world, with nearly ten million deaths per year, and is the second most common cause of death in every population [1]. Despite the tremendous advances in early diagnosis, targeted therapy, and immunotherapy, resistance to treatment and tumor heterogeneity continue to limit long-term outcomes [2]. Therefore, there is a significant need to identify new anticancer approaches that are effective and ready for translation into the clinical setting. In this regard, drug repurposing offers an innovative opportunity to develop new cancer therapies in a time- and cost-efficient manner, and to exploit agents that are already efficacious and use-approved in other indications as effective anticancer therapies.
Isolated from Streptomyces cinnamonensis, Monensin is a polyether ionophore antibiotic that has been used in veterinary medicine for many years [3]. Monensin is well recognized as a treatment for coccidial infections, a means to improve feed efficiency in ruminants, and a strategy to enhance growth performance in poultry [4]. Monensin has been administered to animals for decades, and it has a well-established pharmacological profile, which includes consideration of previously and well-studied ion-transporting properties that allow for the selective movement of cations across biological membranes [5]. Historically, Monensin has been primarily considered in discussions of animal health; however, evidence is beginning to emerge from preclinical cancer studies, suggesting that Monensin demonstrates impressive anticancer activity [6].
There are possibly several methods by which Monensin exhibits anticancer activity, as summarized below. The specific mechanism by which Monensin disrupts Na⁺ and H⁺ gradients leads to a disruption of cell ion homeostasis [7], typically accompanied by intracellular acidification, associated mitochondrial toxicity, and evident cell death [9]. Monensin can also exacerbate oxidative stress initiated by other agents, such as antibiotics, which produce reactive oxygen species (ROS) [8] and activate an associated set of stress-handling transcription factors, including ATF3 and CHOP, thereby increasing the apoptosis of malignancies [10,11,12]. Monensin does not solely demonstrate generalized cytotoxicity; instead, it acts within a complex network of oncogenic and chemotactic signaling pathways that include the PI3K/Akt/mTOR, Wnt/β-catenin, MAPK/ERK, STAT3, and NF-κB pathways, which are associated with cancer progression, therapy resistance, and metastasis [13,14,15].
In addition to its direct anti-proliferative effects, Monensin has direct activity against tumor invasion and metastasis, including the inhibition of matrix metalloproteinases (MMPs) and the epithelial–mesenchymal transition (EMT) [16,17,18,19]. Another very enticing aspect of Monensin is its ability to overcome multidrug resistance (MDR) by modulating drug efflux pumps and also inducing chemosensitization of tumor cells to conventional cytotoxic agents, including doxorubicin, cisplatin, paclitaxel, and gemcitabine [20]. Monensin has also shown considerable promise for synergistic combination in preclinical studies, suggesting That It can be developed as either an anticancer compound or applied as an adjuvant to existing chemotherapy agents [21].
Even with the encouraging data above, there are challenges associated with considering its use in human applications. Monensin is known to have cardiotoxicity and neurotoxicity at high concentrations and raises questions regarding its therapeutic window for use in humans [22]. However, developments in drug delivery systems, including nanoparticle encapsulation, liposomal formulations, and targeted formulations, could offer an avenue to circumvent these concerns while harnessing Monensin's strong anti-cancer potential. Thus, it is critical to consider monensin not only as a veterinary antibiotic but also as a translational research target in oncology.
In summary, Monensin represents a notable example of drug repurposing in oncology. It has multiple mechanisms of action, including disrupting ion transport systems and antagonizing signaling pathways, placing it as one of the most promising therapeutic agents against cancer. In this review, we present a comprehensive examination of the pharmacological domain, molecular mechanisms, and preclinical data supporting the anticancer application of Monensin, along with its limitations and future directions for clinical use.
2. Pharmacological Background of Monensin
Monensin is a polyether ionophore antibiotic isolated initially from Streptomyces cinnamonensis in the 1960s [23]. Due to its ability to selectively transport monovalent cations across biological membranes, it has been extensively used in veterinary medicine as a coccidiostat and growth promoter. The pharmacological uniqueness of monensin arises from its function as an ionophore: it forms lipid-soluble complexes with sodium (Na⁺) and other cations, exchanging them for protons (H⁺) across lipid bilayers. This activity disrupts transmembrane ion gradients, intracellular pH regulation, and ultimately cellular homeostasis [24].
Monensin’s pharmacological effects are not limited to ionic transport. Once localized in membranes, endosomes, and the Golgi apparatus, monensin alters vesicular trafficking and protein secretion. It interferes with N-glycosylation and glycoprotein processing, resulting in the accumulation of immature proteins in intracellular compartments. These effects are particularly detrimental to highly proliferative cells, which rely on efficient protein synthesis, trafficking, and secretion to sustain malignant growth and intercellular signaling [25,26].
At the mitochondrial level, monensin increases Na⁺ influx, which disrupts mitochondrial membrane potential, decreases oxidative phosphorylation, and leads to ATP depletion and oxidative stress. These bioenergetic alterations selectively compromise the survival of metabolically active cancer cells, while exerting minimal toxicity toward normal, slowly proliferating cells [27].
Pharmacologically, monensin also intersects with key oncogenic pathways. By altering endosomal and Golgi pH, it modulates receptor trafficking and degradation, thereby influencing pathways such as the EGFR, mTOR, STAT3, and Wnt/β-catenin pathways. These pleiotropic effects contribute to cell cycle arrest, induction of apoptosis, and inhibition of migration and invasion in diverse cancer models. Significantly, its synergistic interactions with established chemotherapeutics and targeted agents (e.g., EGFR inhibitors, oxaliplatin, and rapamycin) further demonstrate its therapeutic potential and justify its potential repurposing in oncology [28,29,30,31].
In summary, the pharmacological background of monensin highlights a compound that exerts multi-faceted cellular stress through ion imbalance, mitochondrial dysfunction, and disruption of protein trafficking. These distinct mechanisms create vulnerabilities in cancer cells that can be therapeutically exploited, positioning monensin as a unique candidate for anticancer drug development.
3. Molecular and Cellular Mechanisms of Anticancer Action
3.1. Induction of Cell Death Pathways
Monensin induces multiple forms of programmed cell death, primarily apoptosis. By disrupting ionic balance and mitochondrial integrity, it triggers the activation of Bax, the release of cytochrome c, and the caspase cascade, culminating in apoptotic cell death. In glioma and other solid tumors, monensin sensitized cells to TRAIL-mediated apoptosis by promoting endoplasmic reticulum (ER) stress, upregulating death receptor 5 (DR5), and downregulating c-FLIP, thereby overcoming resistance mechanisms. In some models, monensin also induces autophagic cell death through Golgi stress and impaired vesicular trafficking. Together, these mechanisms highlight its ability to selectively eliminate malignant cells via distinct cell death pathways [32,33].
Together, these data suggest that monensin activates both intrinsic and extrinsic apoptotic pathways.
3.2. Oxidative Stress and Mitochondrial Dysfunction
Monensin enhances reactive oxygen species (ROS) generation, either independently or synergistically with other agents, leading to oxidative stress and mitochondrial toxicity. ROS accumulation activates stress-responsive transcription factors such as ATF3 and CHOP, which promote apoptosis in tumor cells. At the mitochondrial level, sodium influx driven by monensin destabilizes the membrane potential, impairs oxidative phosphorylation, and reduces ATP production, rendering metabolically active cancer cells more susceptible to damage than quiescent normal cells. The ability of antioxidants to attenuate monensin-induced cytotoxicity in prostate cancer further underscores the central role of ROS in its anticancer mechanism [34,35].
3.3. Disruption of Ion Homeostasis
As a sodium ionophore, monensin perturbs intracellular Na⁺/H⁺ gradients, causing cytoplasmic acidification and subsequent bioenergetic collapse. This ionic imbalance also interferes with calcium homeostasis, further amplifying mitochondrial dysfunction and apoptosis. In hepatocellular carcinoma (HCC), tumor cells with inherently elevated sodium levels display selective sensitivity to monensin, which exacerbates Na⁺ overload and induces cell death without harming normal hepatocytes. Such ion-based vulnerabilities provide a tumor-selective therapeutic window for monensin [36,37,38].
3.4. Modulation of Oncogenic Signaling Pathways
Monensin disrupts multiple oncogenic pathways critical for tumor progression. It interferes with receptor trafficking in the Golgi and endosomal compartments, thereby inhibiting EGFR signaling and enhancing the efficacy of EGFR inhibitors in breast, ovarian, and pancreatic cancers. Monensin also downregulates the PI3K/Akt/mTOR axis and STAT3 signaling, resulting in the suppression of proliferation and survival. Furthermore, it inhibits canonical Wnt/β-catenin signaling by promoting LRP6 degradation, reducing β-catenin levels, and downregulating target genes such as cyclin D1 and SP5, effects validated in colorectal and cervical cancer models. These pleiotropic actions make monensin a potent modulator of signaling plasticity in cancer cells [39,40,41].
These findings confirm that monensin exerts pleiotropic effects on oncogenic signaling, highlighting its potential as a signaling modulator in cancer therapy.
3.5. Inhibition of Tumor Progression and Metastasis
Monensin exhibits anti-metastatic activity by targeting processes such as epithelial–mesenchymal transition (EMT) and extracellular matrix remodeling. It suppresses EMT-driven motility by altering Golgi pH, inducing exocytosis of GOLIM4, and thereby impairing cytoskeletal dynamics. Additionally, monensin reduces the activity of matrix metalloproteinases (MMPs), such as MMP7, and inhibits the secretion of pro-metastatic cytokines. Through modulation of ADAMTS-1 localization and riboflavin/endosomal trafficking, it further limits the migration and invasiveness of cancer cells. Collectively, these findings position monensin as a multi-targeted inhibitor of invasion and metastasis [42,43].
By targeting both intracellular signaling and extracellular matrix remodeling, monensin effectively interferes with metastatic processes.
3.6. Overcoming Drug Resistance
One of the most clinically relevant properties of monensin is its ability to overcome multidrug resistance (MDR). By modulating drug efflux pumps such as P-glycoprotein, monensin restores intracellular accumulation of chemotherapeutics like doxorubicin and vincristine. In colon cancer, it re-establishes nuclear localization of doxorubicin in ATP7A-expressing resistant cells, thereby reversing drug resistance. Moreover, monensin synergizes with conventional chemotherapeutics (such as cisplatin, paclitaxel, and gemcitabine) and targeted agents (including EGFR and mTOR inhibitors), thereby enhancing cytotoxicity and chemosensitivity in resistant cancers. This capacity to sensitize tumors to therapy underscores its translational value as an adjuvant in oncology [44]. Thus, monensin represents a promising adjuvant capable of reversing MDR and enhancing therapeutic efficacy.
Figure 1.
Types of human cancers investigated for the anticancer effects of monensin.
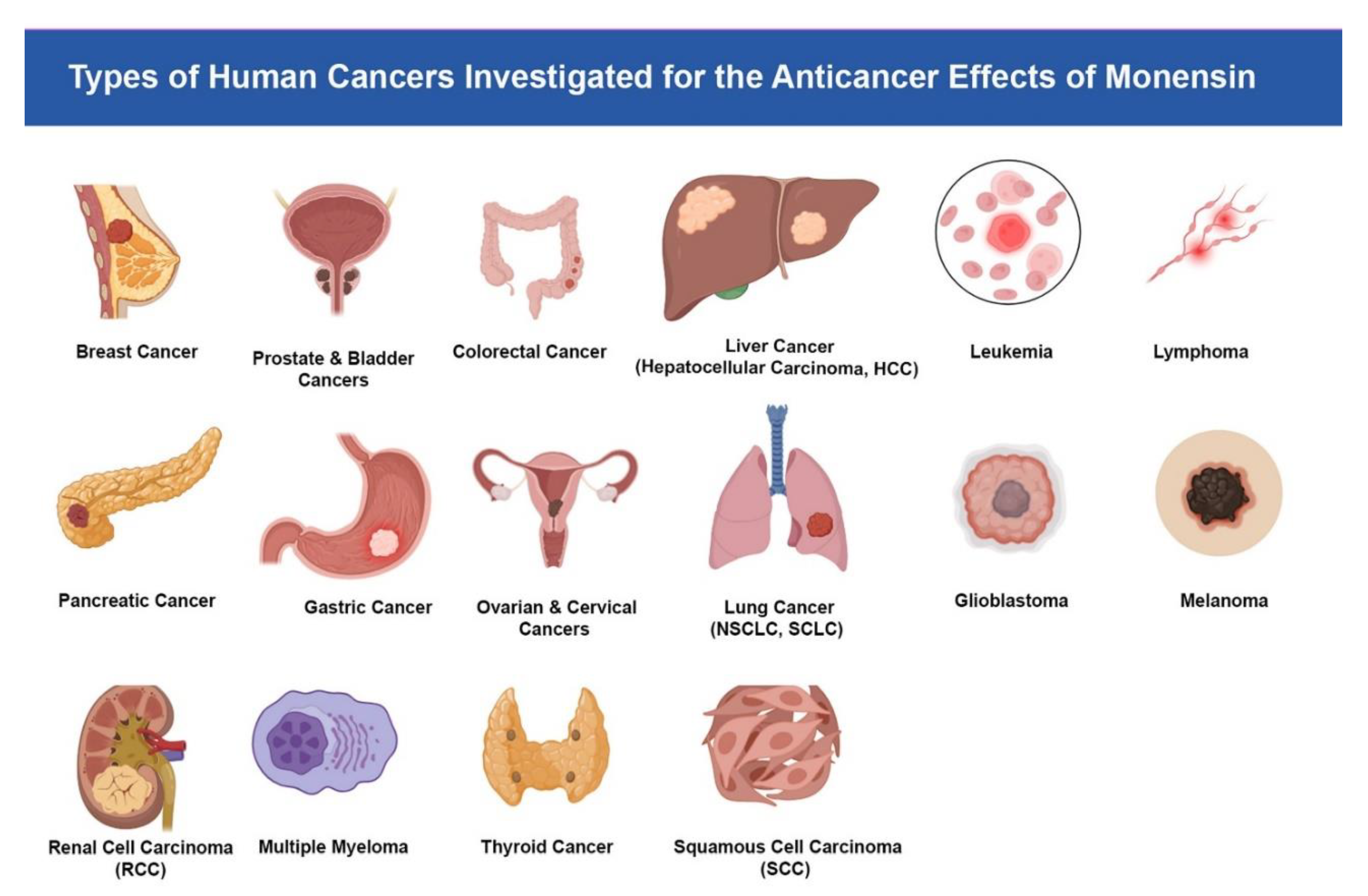
The illustration summarizes the spectrum of malignancies in which monensin has been studied in preclinical research, including breast, prostate and bladder, colorectal, liver (hepatocellular carcinoma), leukemia, lymphoma, pancreatic, gastric, ovarian and cervical, lung (NSCLC and SCLC), glioblastoma, melanoma, renal cell carcinoma, multiple myeloma, thyroid cancer, and squamous cell carcinoma. Reported effects across these tumor types include inhibition of proliferation, induction of apoptosis, disruption of signaling pathways, and chemosensitization.
4. Preclinical Evidence in Specific Cancer Types
Breast Cancer
Triple-Negative Breast Cancer (TNBC)
Breast cancer remains the most commonly diagnosed malignancy among women worldwide, with aggressive subtypes such as triple-negative breast cancer (TNBC) and therapy-resistant HER2-positive tumors continuing to present major therapeutic challenges. Recently, monensin has gained considerable attention for its anticancer potential across multiple breast cancer models. In an organoid-based system, Monensin and several synthetic analogs—particularly compound 12—exhibited more potent and more selective cytotoxicity compared to Monensin itself. These compounds induced DNA fragmentation, inhibited cell migration, reduced the CD44⁺/CD24⁻/low stem-like subpopulation, and downregulated SOX2 expression, thereby impairing the self-renewal of these cells. Proteomic analysis revealed the dysregulation of extracellular matrix–related pathways and a strong upregulation of the tumor suppressor TIMP2, supporting its potential as an anti-metastatic agent [45].
In triple-negative breast cancer (TNBC), a highly aggressive subtype with limited therapeutic options, monensin disrupted Na⁺ homeostasis. In vivo, administration of 8 mg/kg resulted in a marked increase in intracellular Na⁺ levels in tumor tissue, leading to tumor shrinkage and extensive necrosis, without affecting healthy organs. This cytotoxicity was Na⁺-dependent, confirming a tumor-specific mechanism of action [46].
Beyond its ionophore activity, monensin can also induce Golgi stress, triggering selective autophagy (Golgi phagy) and impairing cancer cell proliferation [47,48]. High expression of the Golgi-resident receptor TM9SF3, which is essential for Golgiphagy, is associated with a poor prognosis in breast cancer, further underscoring the therapeutic relevance of this pathway [48].
When combined with the EGFR inhibitor erlotinib, monensin synergistically suppressed the proliferation, migration, invasion, and stemness markers (SOX2 and CD133) in TNBC. This effect was mediated by simultaneous inhibition of EGFR/ERK and PI3K/AKT/mTOR signaling pathways [49]. Moreover, in an orthotopic breast cancer model, nanoparticle-based delivery of monensin (Hyb-NP system) significantly inhibited metastasis by forming a Golgi-associated drug reservoir, enhancing intracellular retention and blocking secretion of metastasis-related cytokines [50].
Several mechanistic insights further support monensin’s anticancer role. It interferes with epithelial-to-mesenchymal transition (EMT) and CSC survival by targeting Golgi function [51]. It regulates the compartmentalization of ADAMTS-1, thereby reducing mesenchymal cell migration [52]. It was also identified as one of the effective lysosome-targeting drugs against invasion in HER2-positive, drug-resistant cancers [53]. In addition, monensin suppresses proliferation and migration by downregulating UBA2 [54], disrupts sodium-dependent phosphate transport to reduce invasiveness [55], and alters riboflavin homeostasis by interfering with endosomal trafficking [56].
Collectively, these findings highlight monensin as a multi-targeted agent against breast cancer, with effects on CSCs, EMT, Golgi-associated stress responses, and key oncogenic signaling pathways.
4.2. Prostate Cancer
Prostate cancer is one of the most frequently diagnosed malignancies in men, and despite advances in androgen-deprivation therapy, resistance and progression to advanced stages remain significant challenges. Current treatments for advanced prostate cancer are limited, prompting the need for new therapeutic strategies.
Several studies have demonstrated that monensin exhibits potent anticancer activity in prostate cancer cells through multiple mechanisms. At very low doses, monensin reduces androgen receptor (AR) levels, induces apoptosis, and increases oxidative stress. Significantly, its cytotoxic effects are enhanced when combined with antiandrogen drugs but are attenuated by antioxidants such as vitamin C, underscoring the role of ROS in its mechanism of action [57].
In vitro analyses further revealed that monensin strongly inhibits prostate cancer cell growth by reducing viability in a dose-dependent manner, inducing G1-phase cell-cycle arrest, and triggering apoptosis. Mechanistically, monensin disrupts Ca2⁺ homeostasis and enhances ROS production, leading to mitochondrial-dependent apoptosis. The reversal of these effects by ROS inhibitors and calcium channel blockers confirmed that monensin acts via ROS- and Ca2⁺-mediated pathways [58].
Resistance to radiation therapy poses another significant clinical problem, affecting 20–30% of prostate cancer patients. Studies investigating monensin as a radiosensitizer have demonstrated that it enhances radiation-induced cytotoxicity in LNCaP (P53⁺/AR⁺) cells but not in PC3 (P53⁻/AR⁻) cells, with Bcl-xL overexpression likely contributing to the observed resistance in PC3 models [59,60].
Beyond apoptosis induction, monensin exerts broader effects on prostate cancer biology. It suppresses AR activity and downregulates oncogenes such as MYC and ERG, while also reducing cancer stem cell markers, promoting cell differentiation, and decreasing cell motility. These effects collectively impair tumor growth and metastatic potential [61,62].
Taken together, monensin demonstrates promising therapeutic potential in prostate cancer by combining direct cytotoxic effects, radiosensitization, and suppression of oncogenic signaling pathways. Its activity is particularly compelling when used in combination with agents that augment oxidative stress, highlighting its value as a candidate for novel therapeutic strategies in advanced prostate cancer.
4.3. Colorectal Cancer
Colorectal cancer (CRC) remains one of the most prevalent and lethal malignancies worldwide, and several studies have explored the potential of monensin as a repurposed anticancer agent in this context. Recent findings demonstrate that monensin modulates innate immune signaling pathways by decreasing the expression of TLR4, TLR7, and IRF3 mRNA in CRC cells in a dose- and time-dependent manner, suggesting that its anticancer activity may involve the suppression of TLR/IRF3-driven inflammatory signaling [63].
In addition to its immunomodulatory effects, monensin directly interferes with the proliferation and survival of cancer cells. It has been shown to significantly inhibit cell proliferation, migration, and cell cycle progression, while inducing apoptosis and G1 arrest in CRC cells. Mechanistically, monensin downregulated IGF1R expression and impacted multiple oncogenic transcriptional regulators, including Elk1, AP-1, and Myc/Max, further supporting its potential as a therapeutic candidate for CRC [64].
One of the most significant findings comes from studies linking monensin to Wnt/β-catenin signaling, a pathway frequently deregulated in colorectal carcinogenesis. Monensin was identified as a potent inhibitor of the Wnt pathway, as it blocked LRP6 phosphorylation, promoted LRP6 degradation, reduced intracellular β-catenin levels, and downregulated Wnt target genes, such as cyclin D1 and SP5. These molecular effects resulted in reduced proliferation of CRC cells, and in vivo, monensin suppressed tumor growth in multiple intestinal neoplasia (Min) mice, underscoring its translational potential [65].
Earlier mechanistic work further confirmed that monensin induces cell cycle arrest at the G1 or G2/M phases in colon cancer cells, mediated by reduced expression of CDK2, CDK4, CDK6, cyclin D1, and cyclin A, along with increased p27 expression and enhanced CDK binding. This was associated with Rb hypophosphorylation, reduced kinase activity, and induction of apoptosis through mitochondrial dysfunction, Bax activation, and caspase-3 cleavage [66].
Interestingly, monensin also influences drug resistance mechanisms. In colon cancer, expression of the copper-transporting ATPase ATP7A is associated with multidrug resistance (MDR) against agents such as cisplatin, paclitaxel, etoposide, SN-38, and doxorubicin. Remarkably, monensin was able to alter doxorubicin distribution, restoring its nuclear localization in ATP7A-expressing cells, and thereby counteracting resistance mechanisms [67]. However, other studies indicate that by mimicking Na⁺/H⁺ exchanger (NHE) activity, monensin can increase intracellular pH and decrease drug accumulation in resistant HT29 colon carcinoma cells, highlighting its complex role in chemoresistance [69].
Furthermore, monensin was reported to modulate the tumor microenvironment by inhibiting the secretion of metalloproteinases (MMPs) and urokinase plasminogen activator (uPA), both of which are critical mediators of invasion and metastasis. At the same time, monensin increased uPAR release, mainly from normal cells, suggesting context-dependent effects on protease regulation [68].
Another promising application is the use of monensin as a potentiator of immunotoxin therapies. Studies demonstrated that liposomal or antibody-targeted formulations of monensin significantly enhanced the cytotoxicity of transferrin receptor- and CEA-directed immunotoxins against CRC cells, both in vitro and in vivo [70,71,72,73]. Notably, monensin accelerated protein synthesis inhibition and increased immunotoxin potency by more than 100-fold in some models, without increasing off-target toxicity, making it a potent adjuvant to targeted toxin-based therapies.
Finally, broad pharmacological screenings confirmed the exceptional potency of monensin, with EC₅₀ values below 0.1 µM, ranking it among the most active agents against colon tumor cell proliferation compared to other drug classes [74].
Collectively, these findings suggest that monensin acts on multiple critical oncogenic processes in CRC, including:
- Immune signaling modulation (TLR/IRF3)
- Growth and survival pathways (IGF1R, Elk1, AP-1, Myc/Max)
- Canonical Wnt/β-catenin signaling
- Cell cycle regulation and apoptosis
- Drug resistance mechanisms (ATP7A, NHE)
- Invasion and metastasis (MMPs, uPA/uPAR)
- Potentiation of immunotoxins
Together, these data strongly support monensin as a multifunctional therapeutic candidate with the potential to be repurposed for the treatment of colorectal cancer.
4.4. Liver Cancer (Hepatocellular Carcinoma, HCC)
Hepatocellular carcinoma (HCC) is the most common primary liver malignancy and is associated with poor prognosis due to late diagnosis and limited effective therapies. HCC cells exhibit altered ion homeostasis, characterized by elevated intracellular sodium (Na⁺) levels compared to normal hepatocytes, which may represent a selective vulnerability.
Recent studies have shown that monensin selectively targets HCC cells by further increasing intracellular Na⁺. This leads to mitochondrial Na⁺ overload, energy depletion, impaired oxygen consumption, and enhanced intracellular water lifetime, ultimately inducing cell death and tumor regression. Importantly, monensin exhibits minimal effects on normal hepatocytes or healthy liver tissue, highlighting Na⁺ homeostasis as a druggable and selective target for HCC therapy [75,76].
These findings suggest that monensin holds promise as a therapeutic agent for HCC, exploiting the unique ionic vulnerabilities of tumor cells while sparing healthy liver tissue. Its selective mechanism provides a potential strategy to improve treatment efficacy and reduce systemic toxicity in liver cancer management.
4.5. Leukemia
Leukemia represents a heterogeneous group of hematologic malignancies characterized by uncontrolled proliferation of abnormal white blood cells, leading to impaired hematopoiesis and immune dysfunction. Among them, acute myeloid leukemia (AML) is particularly aggressive, with a poor prognosis and limited therapeutic options due to frequent relapse and drug resistance. Novel therapeutic agents with selective cytotoxicity against leukemic cells are therefore urgently needed.
Monensin has demonstrated vigorous anti-leukemic activity in both in vitro and in vivo models. In AML cells, monensin inhibits proliferation at low micromolar concentrations (IC₅₀ ~0.5 μM) by inducing G1/G2-M cell cycle arrest through the modulation of CDKs, cyclins, and p27, while simultaneously promoting apoptosis via the Bax, caspase, and mitochondrial pathways. In AML mouse models, monensin treatment significantly reduced tumor growth and increased the expression of apoptotic markers. Similar inhibitory effects were observed in myeloma and lymphoma cells, reinforcing its broad efficacy across hematologic malignancies [77,78].
Mechanistically, monensin acts as a novel MYB inhibitor, suppressing the proliferation of AML cells while sparing normal hematopoietic cells, suggesting potential as a targeted therapy [79]. Beyond direct cytotoxicity, monensin blocks the morphogenesis of Friend murine leukemia virus, thereby preventing viral-driven leukemogenesis [80]. In HL-60 leukemia cells, it disrupts myeloperoxidase maturation and trafficking within the ER–Golgi pathway, highlighting an additional mechanism interfering with leukemic differentiation [81]. Evidence from uveal melanoma models further suggests that monensin-mediated suppression of RhoA signaling may also contribute to its anti-leukemic effects [82].
Clinically, monensin enhanced the intracellular delivery and cytotoxicity of the anti-CD5 immunotoxin T101-RTA in B-cell chronic lymphocytic leukemia (B-CLL), overcoming drug resistance and supporting its potential role as an adjuvant in targeted immunotherapies [83].
4.6. Lymphoma
Lymphomas are a heterogeneous group of hematologic malignancies arising from B- or T-lymphocytes and represent a significant clinical challenge due to their diverse biology and resistance mechanisms. Monensin, a Na⁺ ionophore, has been extensively studied for its anticancer effects in lymphoma, demonstrating potent activity through multiple mechanisms.
At the cellular level, monensin strongly inhibits the proliferation of human lymphoma cell lines with an IC₅₀ of ~0.5 μM, inducing cell cycle arrest at G1 and/or G2-M phases. This is mediated by the modulation of key regulatory proteins, including CDK4, cyclin A, cdc2, p21, and p27, resulting in reduced CDK- and cdc2-associated kinase activity. In addition, monensin triggers apoptosis by inducing loss of mitochondrial membrane potential, indicating that it suppresses lymphoma growth through a dual mechanism of cell cycle arrest and apoptosis [84,88].
In B-cell lymphomas, monensin has been shown to increase CD20 surface expression by downregulating MYC, a repressor of the MS4A1 gene that encodes CD20. This effect enhances the efficacy of anti-CD20 monoclonal antibodies and CAR-T cell therapies, offering a promising strategy to overcome therapeutic resistance in non-Hodgkin lymphoma [85].
In T-cell lymphoma, monensin alters cell surface glycosylation, decreasing terminal modifications such as sialylation and fucosylation while increasing high-mannose glycans. Although surface antigen expression (e.g., Thy-1) remains unchanged, these glycosylation alterations impair tumor cell migration and homing, as evidenced by reduced spleen localization after intravenous injection [86,87].
Additional studies highlight other mechanisms of action. Monensin functions as an ER stress inducer in B lymphoma cells (CH12), where loss of TFG (a regulator of ER homeostasis and autophagy) sensitizes cells to apoptosis, revealing its potential to exploit ER stress vulnerabilities [89]. It also interferes with receptor trafficking, such as blocking endosome–lysosome degradation of ovine prolactin in Nb2 lymphoma cells, thereby altering hormone signaling [90]. Moreover, monensin blocks the endocytic uptake of synthetic alkyl-lysophospholipids, such as Et-18-OCH₃, thereby preventing the inhibition of phosphatidylcholine biosynthesis and reducing apoptosis, which underscores its role in modulating membrane-mediated apoptotic pathways [91].
Collectively, these findings indicate that monensin exerts multi-faceted antitumor effects in lymphoma, including cell cycle regulation, apoptosis induction, modulation of glycosylation and trafficking, and enhancement of immunotherapy efficacy. Such pleiotropic actions highlight its promise as a candidate for therapeutic strategies in lymphoma treatment.
4.7. Pancreatic Cancer
Pancreatic cancer is one of the most lethal malignancies, characterized by late diagnosis, aggressive progression, and poor responsiveness to chemotherapy. Resistance to conventional chemotherapeutic agents, such as gemcitabine, remains a significant barrier to effective treatment. Hence, novel therapeutic strategies targeting key oncogenic pathways are urgently needed.
Monensin has been reported to exert potent anticancer effects in chemo-resistant pancreatic cancer cells by interfering with epidermal growth factor receptor (EGFR) signaling. Wang et al. (2018) demonstrated that monensin significantly inhibited cell proliferation and colony formation in resistant pancreatic cancer cell lines. Mechanistically, monensin reduced the phosphorylation of EGFR and its downstream effectors, thereby disrupting critical pathways involved in survival and proliferation [92].
In addition to its in vitro effects, monensin also suppressed tumor growth in vivo, further supporting its therapeutic potential. By specifically targeting the EGFR pathway—a central driver of pancreatic cancer progression and chemoresistance—monensin offers a promising avenue for overcoming drug resistance in this highly aggressive malignancy.
Collectively, these findings suggest that monensin may serve as an effective adjuvant agent in pancreatic cancer therapy, particularly for patients with disease that is resistant to conventional treatments. Future studies should focus on exploring its synergistic potential with standard chemotherapeutics and elucidating its role in the broader signaling network that drives pancreatic tumorigenesis.
4.8. Gastric Cancer
Gastric cancer (GC) remains one of the most lethal malignancies worldwide, mainly due to late diagnosis, therapeutic resistance, and tumor heterogeneity. In recent years, particular attention has been directed toward the role of cancer stem-like cells (CSCs) in driving tumor initiation, metastasis, and resistance to standard therapies. Several studies have highlighted the potential of monensin to target these aggressive subpopulations in gastric tumors selectively.
Recent work demonstrated that HMGA1, a chromatin-binding protein frequently upregulated in GC, enhances cancer stem-like features such as self-renewal, migration, and drug resistance. Importantly, HMGA1-overexpressing gastric cancer cells exhibited increased sensitivity to monensin, indicating that this ionophore may exploit vulnerabilities associated with CSC phenotypes [93]. Similarly, the use of a SOX2 reporter system identified gastric CSCs that were highly responsive to monensin treatment. In these models, monensin significantly impaired the viability and clonogenicity of CSCs, suggesting that it may serve as a valuable adjunct for targeting stemness-driven disease progression [94].
Beyond its role in CSC modulation, monensin’s unique ability to interfere with vesicular trafficking and membrane dynamics may also enhance targeted drug delivery in gastric cancer. A classic study on the RS-1541 compound, delivered via the low-density lipoprotein (LDL) pathway, revealed efficient uptake and cytotoxicity in St-4 human gastric cancer cells. This finding underscores the potential of exploiting vesicular transport mechanisms, a process in which monensin is known to play a modulatory role [95].
Taken together, these studies position monensin as a promising candidate in the treatment of gastric cancer, with dual potential: (i) targeting CSC populations that drive therapy resistance and recurrence, and (ii) modulating intracellular trafficking pathways to improve drug delivery efficiency. Future research should focus on integrating monensin into combination therapies with CSC-targeted agents and LDL-based nanocarriers, aiming to overcome the major therapeutic hurdles in gastric cancer.
4.9. Ovarian Cancer
Ovarian cancer is the most lethal gynecologic malignancy, often diagnosed at advanced stages and associated with poor prognosis due to high rates of recurrence and the development of chemotherapy resistance. Despite improvements in surgery and systemic therapy, most patients relapse within 12–24 months, underscoring the urgent need for new therapeutic approaches.
Recent studies have highlighted monensin as a promising agent for the treatment of ovarian cancer. In vitro experiments on multiple ovarian cancer cell lines have demonstrated that monensin suppresses cell proliferation, migration, invasion, and DNA replication in a dose- and time-dependent manner. At concentrations as low as 0.2 μM, monensin reduced proliferation, while higher doses (5 μM) completely blocked growth within 24 hours. Monensin also impaired cell motility and wound healing capacity, and induced G2/M phase arrest. Mechanistically, it enhanced MEK1 SUMOylation, leading to inhibition of the MEK-ERK signaling pathway, which was further validated in xenograft models where monensin significantly suppressed tumor growth [96].
Complementary evidence shows that monensin also inhibits ovarian cancer cell proliferation by inducing G1 arrest and apoptosis, while suppressing multiple cancer-related signaling pathways, including Elk1/SRF, AP1, NF-κB, STAT, and Wnt/β-catenin. Notably, it reduced EGFR expression and synergized with both EGFR inhibitors and the chemotherapeutic drug oxaliplatin, leading to enhanced suppression of cell proliferation and tumor growth in xenograft studies [97].
In addition, lysosome-targeting properties of monensin have attracted interest as part of broader strategies to inhibit tumor invasion, including in treatment-resistant HER2-positive cancers, further underscoring its potential as a repositioned anti-cancer agent [98].
Together, these findings establish monensin as a potent inhibitor of ovarian cancer progression, acting through multi-pathway suppression, induction of SUMOylation, and synergism with existing therapies, making it a strong candidate for therapeutic development in resistant ovarian malignancies.
4.10. Cervical Cancer
Cervical cancer remains one of the most prevalent gynecological malignancies worldwide, often associated with persistent human papillomavirus (HPV) infection and dysregulation of multiple oncogenic signaling pathways. Among these, the Wnt/β-catenin pathway plays a pivotal role in promoting proliferation, survival, epithelial-mesenchymal transition (EMT), and therapy resistance in cervical cancer cells.
Recent evidence has shown that monensin effectively inhibits the Wnt/β-catenin signaling cascade in cervical cancer models. Fu et al. (2024) reported that monensin treatment led to a marked reduction in β-catenin activity and its downstream transcriptional targets, resulting in suppressed cell growth and induction of apoptosis [99]. By targeting this pathway, monensin disrupts key molecular mechanisms that sustain cervical cancer progression, particularly those associated with cell cycle regulation and tumor invasiveness.
The inhibition of Wnt/β-catenin by monensin is particularly significant, as this pathway is frequently activated in HPV-driven carcinogenesis and contributes to chemoresistance. Thus, monensin not only exhibits direct cytotoxic effects but may also enhance the sensitivity of cervical cancer cells to conventional chemotherapeutics by modulating signaling plasticity.
Collectively, these findings suggest that monensin holds therapeutic promise in cervical cancer by selectively targeting the Wnt/β-catenin signaling pathway, a central driver of tumor survival and progression. Future studies should focus on validating its efficacy in vivo and exploring its potential use in combination therapy strategies for the management of cervical cancer.
4.11. Lung Cancer (NSCLC, SCLC)
Lung cancer remains the leading cause of cancer-related mortality worldwide, and despite significant advances in targeted therapies and immunotherapy, resistance and tumor recurrence continue to pose substantial challenges. Recent studies have highlighted monensin as a promising adjuvant agent that can interfere with multiple oncogenic processes in both non-small cell lung cancer (NSCLC) and small cell lung cancer (SCLC).
One of the most extensively studied mechanisms is the ability of monensin to function as an inhibitor of autophagy. By disrupting autophagosome–lysosome fusion, monensin blocks the autophagic process at its final stage. While its standalone cytotoxicity is limited, combination therapy with standard anticancer agents has demonstrated notable synergistic effects. For instance, co-treatment with rapamycin or erlotinib significantly enhanced cell cycle arrest and apoptosis in lung cancer cells, as evidenced by increased expression of pro-apoptotic proteins (Bax, cleaved caspase-3, and PARP) and downregulation of anti-apoptotic markers (Bcl-2 and Bcl-xL). DNA content analysis further revealed an accumulation of cells in the G1 phase, indicating that autophagy suppression may potentiate the efficacy of mTOR and EGFR inhibitors [100].
Another critical mechanism involves the suppression of epithelial–mesenchymal transition (EMT), a process strongly linked to metastasis and drug resistance in NSCLC. Monensin partially inhibited TGF-β–induced EMT, as reflected by reduced vimentin and ZEB1 expression and the preservation of E-cadherin. Functionally, this translated into a restoration of osimertinib sensitivity in EGFR-mutant NSCLC cells that had undergone EMT, suggesting that monensin may help overcome therapy resistance mediated by phenotypic plasticity [101]. Complementary findings revealed that monensin also disrupts EMT-driven migration by altering Golgi pH and inducing exocytosis of GOLIM4. This scaffold protein regulates the trafficking of talin 1 (TLN1), a protein involved in adhesion. This impairment of motility highlights a unique mechanism by which monensin affects lung cancer cell invasiveness [102].
Beyond EMT, monensin has been shown to alter receptor trafficking. In NSCLC cells harboring EGFR mutations, monensin caused perinuclear accumulation of EGFR by blocking its exit from the endocytic recycling compartment. This aberrant trafficking reduced the availability of mutant EGFR at the plasma membrane, potentially diminishing oncogenic signaling and interaction with Src kinases, while sparing wild-type EGFR-expressing cells [103].
Resistance to chemotherapy in lung cancer is often mediated by multidrug resistance (MDR) transport systems such as P-glycoprotein. In drug-resistant NSCLC cell lines, monensin disrupted ATP-dependent vesicular sequestration of chemotherapeutic agents, thereby enhancing nuclear accumulation of doxorubicin and vincristine. This suggests that monensin could act as a chemosensitizer in multidrug-resistant phenotypes [104].
Monensin has also been implicated in the regulation of tumor dormancy and reactivation. In a study of dormant tumor cells (DTCs) in the lung, monensin blocked the TGF-β1/SNAI2-mediated switch that reactivated dormant cells into a proliferative state, highlighting its potential role in preventing metastatic recurrence [105].
Finally, in the context of small cell lung cancer (SCLC), monensin was found to inhibit the secretion of laminin, a key extracellular matrix protein frequently dysregulated in SCLC. This effect reflects its broader impact on protein trafficking and secretion pathways, which could contribute to alterations in tumor-microenvironment interactions [106].
Taken together, these findings demonstrate that monensin exerts multifaceted effects in lung cancer, ranging from inhibiting autophagy and inducing apoptosis to suppressing EMT, disrupting receptor trafficking, reversing drug resistance, inhibiting dormant cell reactivation, and modulating extracellular matrix components. While preclinical evidence is compelling, clinical translation remains unexplored, positioning monensin as a promising but underutilized candidate for lung cancer therapy [100,101,102,103,104,105,106,107].
4.12. Glioblastoma
Glioblastoma Multiform (GBM)
Glioblastoma (GBM) is the most aggressive and common primary brain tumor in adults, characterized by rapid proliferation, high vascularization, and marked resistance to conventional therapies. Despite surgery, radiotherapy, and chemotherapy, the prognosis remains dismal, with a median survival of around 15 months. This highlights the urgent need for novel therapeutic strategies targeting both tumor cells and the supportive tumor microenvironment.
Monensin and its analogs have emerged as promising agents against GBM. In organoid models, monensin induced apoptosis, DNA damage, autophagy, and PARP degradation, while reducing migration and suppressing critical oncogenic pathways, including STAT3, Akt, and GSK3β. Treatment of co-cultured cerebral organoids further demonstrated a significant reduction in tumor size, underscoring the therapeutic potential of monensin analogs [108]. Additionally, monensin demonstrated anti-angiogenic activity, effectively inhibiting GBM endothelial cell proliferation, migration, and capillary-like network formation at nanomolar concentrations. Mechanistically, these effects were mediated through suppression of VEGFR- and EGFR-driven signaling, with in vivo studies confirming reduced tumor vascularization and growth at tolerable doses [109].
Further insights reveal that monensin impacts endosomal pH regulation, mimicking NHE9 activity to alkalinize endosomes, thereby modulating receptor tyrosine kinase stability and stemness pathways such as STAT3/Oct4/Nanog [110]. It also interferes with N-glycan processing in glioblastoma cells, reducing glycoprotein surface expression, adhesion, and proliferation [111]. Monensin enhances the efficacy of anti-transferrin receptor immunotoxins by disrupting endosomal trafficking and facilitating the release of cytosolic toxins. Encapsulation in liposomes further enhanced its bioavailability and stability, resulting in potent in vitro and in vivo activity [112,113,114,115].
Other studies demonstrate that monensin can inhibit TGF-β maturation, suppressing downstream Smad2 signaling and potentially reducing tumor invasiveness [116]. It also blocks the secretion of tumor-promoting proteins such as α2-macroglobulin-like factors. It disrupts receptor recycling processes, including neuromedin B receptor (NMB-R) trafficking, thereby prolonging receptor desensitization and reducing glioblastoma cell signaling and proliferation [117,118].
Taken together, these findings suggest that monensin exhibits multi-targeted anti-GBM effects, including the induction of apoptosis, inhibition of angiogenesis, suppression of stemness, and interference with protein trafficking and receptor signaling. Such pleiotropic mechanisms highlight its potential as a candidate for therapeutic repositioning in glioblastoma treatment, particularly when combined with immunotoxins or other targeted therapies.
4.13. Melanoma
Melanoma is the most aggressive and lethal cutaneous malignancy, characterized by poor response to chemotherapy and a high degree of drug resistance, which calls for new therapeutic strategies urgently. Drug repurposing has emerged as a promising approach, and in this context, monensin has shown significant anti-melanoma activity.
Experimental studies have demonstrated that monensin is selectively toxic to melanoma cells while sparing normal human cells. It effectively inhibits proliferation and colony/sphere formation, promotes apoptosis, and induces terminal differentiation of melanoma stem cells, thereby reducing their stemness properties. Monensin treatment increases melanogenesis and arrests cells in the G0/G1 phase, decreasing S/G2/M populations in a dose-dependent manner, ultimately leading to growth inhibition [119].
In uveal melanoma, which is the most common primary intraocular malignancy, monensin was found to suppress cell viability, inhibit migration, and induce apoptosis. Importantly, it synergized with chemotherapy by inhibiting RhoA-mediated signaling, highlighting its potential as a radiosensitizer and a combination therapy agent [120].
At the molecular level, monensin interferes with Golgi apparatus-dependent glycosylation processes in melanoma cells. It disrupts the biosynthesis and secretion of tumor-associated fibronectin, alters the glycosylation of tissue plasminogen activator (t-PA), and selectively blocks the N-glycan processing of tyrosinase-related proteins (TRP-1 and TRP-2), which are critical for melanin synthesis and immune recognition [121,123,124]. Furthermore, monensin enhances tyrosinase enzymatic activity and stimulates L-tyrosine transport, both of which are closely linked to melanin biosynthesis [126,127].
Monensin also exerts its anti-melanoma effect by inhibiting the canonical Wnt/β-catenin signaling pathway, a pathway widely implicated in melanoma progression, stemness, and therapy resistance. Inhibition of this pathway reduces the expression of downstream targets such as cyclin D1 and SP5, thereby decreasing proliferation and tumorigenic potential [122].
In metastatic settings, monensin has been shown to potentiate the efficacy of immunotoxin therapies. Specifically, when combined with ricin A-chain immunotoxin, monensin significantly reduced pulmonary metastases both in vitro and in vivo, indicating a possible role as an adjuvant in immunotherapy [125].
Collectively, these findings demonstrate that monensin exerts multifaceted anti-melanoma activities, including the induction of apoptosis, promotion of differentiation, inhibition of stemness, disruption of glycoprotein processing, and modulation of key oncogenic pathways, such as the Wnt/β-catenin and RhoA pathways. These pleiotropic effects highlight its promise as a potential therapeutic or adjuvant agent in both cutaneous and uveal melanoma.
4.14. Renal Cell Carcinoma (RCC)
Renal cell carcinoma (RCC) is the most common type of kidney cancer in adults and encompasses multiple subtypes, including clear cell RCC (ccRCC) and TFE3-translocated RCC. RCC is often resistant to conventional chemotherapy, highlighting the need for novel targeted therapies.
Monensin has demonstrated significant anti-RCC activity across different subtypes. In TFE3-translocated RCC (UOK146), monensin induces cell death, disrupts mitochondrial function, activates autophagy, reduces cell migration, and downregulates the PRCC–TFE3 fusion transcript, suggesting its potential as a therapeutic candidate for this aggressive subtype [128].
In general, RCC cell lines, such as ACHN and Caki-2, are inhibited from proliferating by monensin, with an IC50 of approximately 2.5 μM, which induces cell cycle arrest at the G1 or G2-M phases. This is associated with downregulation of CDK2, CDK6, cdc2, cyclin A, and cyclin B1, along with upregulation of the cell cycle inhibitors p21 and p27, resulting in reduced Rb phosphorylation. Monensin also triggers mitochondria-mediated apoptosis, characterized by modulation of Bcl-2 family proteins, caspase activation, and loss of mitochondrial membrane potential [129].
Furthermore, bioinformatics analyses of ccRCC datasets identified CTLA4 as a prognostic biomarker and potential immunotherapeutic target. Connectivity Map (CMap) analyses suggested that monensin, along with quercetin and fenbufen, may be potential small-molecule drugs for ccRCC, although direct experimental validation is still needed [86=130].
These findings collectively indicate that monensin exerts anti-proliferative, pro-apoptotic, and autophagy-inducing effects in RCC, supporting its promise as a targeted therapeutic strategy for multiple RCC subtypes.
4.15. Multiple Myeloma
Multiple myeloma (MM) is a malignant plasma cell disorder characterized by the clonal proliferation of myeloma cells in the bone marrow, resulting in skeletal destruction, immune suppression, and systemic complications. Despite advances in proteasome inhibitors, immunomodulatory agents, and monoclonal antibodies, relapse and drug resistance remain significant challenges, highlighting the need for novel therapeutic strategies. Recent evidence suggests that monensin, a polyether ionophore antibiotic, may exert potent anti-myeloma effects through distinct mechanisms.
Early preclinical studies demonstrated that monensin induces growth inhibition and apoptosis in myeloma cell lines. In NCI-H929 myeloma cells, monensin triggered cell cycle arrest at the G1 phase, along with a significant increase in apoptotic cell populations. Mechanistically, these effects were accompanied by mitochondrial dysfunction, caspase activation, and PARP cleavage, underscoring the pro-apoptotic activity of monensin in plasma cell malignancies [131].
More recently, novel insights have been gained into how myeloma cells internalize extracellular signals and interact with their microenvironment. A 2024 study identified a retrotransposon-derived DNA “zip code” that drives the internalization of myeloma cells via clathrin- and Rab5a-mediated endocytosis. Interestingly, monensin treatment was shown to disrupt this process, suggesting that it may interfere with tumor cell trafficking and microenvironmental interactions crucial for myeloma progression [132]. By altering endocytic trafficking, monensin may reduce the ability of myeloma cells to internalize survival-promoting signals, thereby sensitizing them to apoptosis and limiting their growth potential.
Taken together, these findings provide compelling evidence that monensin exerts dual anti-myeloma activity: (i) direct cytotoxicity through cell cycle arrest and apoptosis induction, and (ii) disruption of endocytosis-dependent survival pathways that are unique to malignant plasma cells. Although these findings are still limited to preclinical settings, they point toward the potential of monensin as an adjuvant therapeutic candidate in multiple myeloma, either alone or in combination with existing anti-myeloma agents.
4.16. Thyroid Cancer
Thyroid cancer is the most common endocrine malignancy, encompassing several histological subtypes, including papillary, follicular, medullary, and the highly aggressive anaplastic thyroid carcinoma (ATC). While most differentiated thyroid cancers respond well to surgery and radioactive iodine therapy, ATC remains refractory to conventional treatments, with a median survival of only a few months. This highlights the urgent need for novel therapeutic strategies targeting ATC and other resistant thyroid tumors.
Preclinical studies have shown that monensin exerts potent anti-thyroid cancer activity across multiple thyroid cancer cell lines, with the strongest sensitivity observed in ATC cells. Mechanistically, monensin disrupts mitochondrial respiration, leading to oxidative stress and mitochondrial damage. This is accompanied by AMPK activation and subsequent mTOR inhibition, ultimately suppressing cell proliferation and triggering apoptosis in ATC cells. These findings identify mitochondrial metabolism as a critical target for monensin in thyroid cancer, supporting its repurposing as a potential anti-ATC therapy [133].
In follicular thyroid carcinoma (FTC) models, monensin has also been shown to perturb Golgi complex integrity and alter intracellular protein localization. Specifically, in FTC133A cells, treatment with monensin caused expansion of the Golgi apparatus and a redistribution of PDE4A splice variant RD1 immunofluorescence from a polarized nuclear focus to a more diffuse perinuclear and cytoplasmic pattern. Similar effects were observed for the Tex1 antigen, consistent with disruption of the Golgi apparatus. These findings indicate that, in addition to mitochondrial dysfunction, monensin can modulate cellular organization and protein trafficking in thyroid cancer cells [134].
Collectively, these studies highlight the selective anti-cancer effects of monensin in thyroid malignancies, particularly in aggressive and treatment-resistant subtypes such as ATC, positioning it as a promising candidate for therapeutic repurposing.
4.17. Bladder Cancer
Bladder cancer is among the most prevalent malignancies of the urinary tract, with high recurrence rates and a strong need for novel targeted therapies. Conventional treatment strategies, including surgery and intravesical chemotherapy, often face limitations due to incomplete tumor eradication and drug resistance. Hence, the exploration of agents that modulate intracellular trafficking and drug uptake has gained increasing attention.
A study using human 5637 bladder cancer cells investigated the role of plant lectins as mediators of targeted drug delivery and uptake. It has been demonstrated that lectins can efficiently bind to and internalize into bladder cancer cells via receptor-mediated pathways, highlighting the therapeutic potential of compounds that exploit endocytosis and vesicular trafficking for selective targeting [135].
In this context, monensin emerges as a particularly relevant candidate, given its ability to alter Golgi function, vesicular trafficking, and endosomal acidification. By disrupting intracellular transport mechanisms, monensin may enhance or interfere with the uptake of drug–carrier complexes such as lectins, potentially increasing the cytotoxicity of co-administered anticancer agents. Additionally, since bladder cancer cells exhibit high rates of receptor-mediated internalization, monensin’s modulatory effect on clathrin-dependent pathways could be leveraged to sensitize tumor cells to targeted delivery approaches.
Although direct studies on monensin in bladder cancer remain limited, the mechanistic overlap between its known activity and the endocytic pathways described in 5637 bladder cancer cells strongly supports its exploration in this setting. Future studies should focus on combining monensin with lectin-based carriers or intravesical therapies to maximize drug accumulation in tumor tissue while reducing systemic toxicity.
4.18. Squamous Cell Carcinoma (SCC)
Squamous cell carcinoma (SCC) represents one of the most common epithelial malignancies, arising in tissues such as the skin, head and neck, and esophagus. SCC progression is tightly linked to aberrations in growth factor receptor signaling and membrane trafficking, processes in which monensin has shown significant modulatory potential.
One of the key molecular hallmarks of SCC is dysregulation of the epidermal growth factor receptor (EGFR). Early studies have demonstrated that EGFR undergoes complex glycosylation events that are critical for its proper membrane localization and ligand-binding activity. Alterations in glycosylation patterns, as observed in SCC cells, can influence receptor stability, signaling output, and response to therapy [136]. Additionally, investigations into the biosynthesis of EGFR in human SCC cell lines revealed that while some hyperproducing cells secrete truncated forms of the receptor, this phenomenon is not universal, highlighting the heterogeneous biology of EGFR regulation in SCC [137].
Beyond EGFR, SCC cells may also harbor genetic amplifications that affect receptor-related pathways. For example, UMSCC38 cells with amplification at chromosome 11q13 were found to synthesize a mutant, nonfunctional folate receptor, underscoring how receptor abnormalities can compromise cellular transport and folate metabolism in this type of cancer [138].
These findings are particularly relevant in the context of monensin. As a polyether ionophore that disrupts Golgi function, vesicular trafficking, and glycoprotein processing, monensin can interfere with the proper maturation and membrane localization of receptors, such as EGFR and the folate receptor. By perturbing these pathways, monensin may attenuate oncogenic signaling, sensitize SCC cells to targeted therapies, and overcome adaptive resistance mechanisms.
Taken together, while direct experimental evidence of monensin’s efficacy in SCC remains limited, the overlap between its cellular targets and the molecular vulnerabilities of SCC strongly suggests a therapeutic potential worth further exploration.
5. Advantages and Challenges
Advantages
Monensin offers several advantages as a potential candidate for repurposing as an anticancer drug. Its long-standing veterinary use provides a well-documented safety and pharmacological profile, which shortens the path to clinical evaluation compared with new drug discovery. Monensin acts through multiple complementary mechanisms, including disruption of ion balance, induction of oxidative stress, modulation of oncogenic signaling, inhibition of epithelial–mesenchymal transition (EMT), and reversal of multidrug resistance. These pleiotropic actions allow it to target a broad range of cancers. Importantly, malignant cells appear more sensitive than normal cells to monensin-induced ionic and metabolic stress, suggesting a degree of selectivity.
Challenges
At the same time, several obstacles limit its clinical application. Monensin can produce cardiotoxic, neurotoxic, and muscular side effects at higher doses, raising concerns about its therapeutic window. Its poor solubility and limited bioavailability reduce systemic usability and demand innovative formulations. While its multi-targeted nature enhances efficacy, it also increases the risk of off-target effects in healthy tissues. Moreover, current evidence is confined to laboratory and animal studies, with no clinical validation in humans. Regulatory and ethical issues linked to repurposing a veterinary antibiotic also remain unresolved.
6. Clinical Implications and Future Perspectives
Preclinical findings highlight monensin’s potential to enhance cancer therapy, but its most practical role is likely in combination regimens, where it sensitizes resistant tumors to standard chemotherapies or targeted agents. This strategy may be particularly relevant for malignancies with poor prognosis and high drug resistance, such as triple-negative breast cancer, GBM, and hepatocellular carcinoma.
Drug delivery innovation is central to overcoming pharmacological limitations. Nanoparticles, liposomes, and tumor-targeted carriers can improve solubility, bioavailability, and tumor selectivity, while reducing systemic toxicity. Organellar targeting, such as Golgi- or lysosome-directed systems, may further exploit monensin’s mechanistic strengths.
Looking forward, several priorities emerge:
Conduct detailed toxicological studies to define a safe therapeutic window in humans.
Optimize delivery strategies to enhance tumor specificity and minimize side effects.
Design clinical trials for cancer types with a clear biological rationale, such as those characterized by ionic imbalance or multidrug resistance.
Explore synergies with immunotherapies, as monensin’s signaling effects may enhance immune responsiveness.
To date, no registered clinical trials have tested monensin in oncology, underscoring the importance of translating preclinical promise into early-phase human studies.
7. Conclusion
Monensin exemplifies the potential of drug repurposing in oncology. Developed initially as a veterinary antibiotic, it exhibits broad anticancer activity through the disruption of ion gradients, induction of oxidative stress, modulation of oncogenic signaling pathways, and reversal of drug resistance—these mechanisms collectively position monensin as a promising candidate for therapeutic development.
Nevertheless, significant barriers—including toxicity, poor solubility, and the absence of clinical evaluation—must be addressed. Advances in targeted delivery systems and rational combination strategies offer practical solutions, but rigorous human studies are essential.
In summary, monensin holds genuine promise as an adjuvant or complementary therapy against resistant and aggressive cancers. Its transition from preclinical promise to clinical utility will depend on careful optimization and well-designed clinical trials.
Author Contributions
Ali Darabniya conceived the study, led the project, designed the framework, collected and analyzed the data, and prepared the initial draft. Sara Shahriari and Reza Fazeli contributed to the literature review, summarized the key findings, and assisted with the writing. Shahrzad Jalilpour Rezaei assisted with the literature review and writing, while Shakiba Sheikholeslami performed a thorough review and revision. All authors approved the final manuscript.
Funding
No funding was received for conducting this study or for the preparation of this manuscript.
Data Availability
No new data were generated or analyzed in this study. All data supporting the findings of this review are derived from previously published literature, which has been appropriately cited.
Statement of ethics
This article is a literature review and does not contain any studies involving human participants or animals performed by the author.
Consent to Participate
Not applicable. No human participants were involved in this study.
Consent for Publication
Not applicable.
Acknowledgements
The author has no acknowledgments to declare.
Competing Interests
The authors declare no competing interests.
References
- Ferlay J, Colombet M, Soerjomataram I, Parkin DM, Piñeros M, Znaor A, Bray F. Cancer statistics for the year 2020: An overview. Int J Cancer. [CrossRef] [PubMed]
- El-Sayes N, Vito A, Mossman K. Tumor Heterogeneity: A Great Barrier in the Age of Cancer Immunotherapy. Cancers (Basel). 2021, 13, 806. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Matĕjů J, Karnetová J, Nohýnek M, Vanĕk Z. Propionate and the production of monensins in Streptomyces cinnamonensis. Folia Microbiol (Praha). 1988, 33, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Muwalla MM, Abo-Shehada MN, Tawfiq F. Effects of monensin on daily gain and natural coccidial infection in Awassi lambs. Small Ruminant Research. 1994, 13, 205–209. [Google Scholar] [CrossRef]
- Nachliel E, Finkelstein Y, Gutman M. The mechanism of monensin-mediated cation exchange based on real time measurements. Biochim Biophys Acta. 1996, 1285, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Rajendran V, Ilamathi HS, Dutt S, Lakshminarayana TS, Ghosh PC. Chemotherapeutic potential of monensin as an anti-microbial agent. Current Topics in Medicinal Chemistry. 2018, 18, 1976–1986. [Google Scholar]
- Aowicki D, Huczyński A. Structure and antimicrobial properties of monensin A and its derivatives: summary of the achievements. Biomed Res Int. 2013, 2013, 742149. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Singh M, Kalla NR, Sanyal SN. Effect of monensin on the enzymes of oxidative stress, thiamine pyrophosphatase and DNA integrity in rat testicular cells in vitro. Exp Toxicol Pathol. 2006, 58, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Souza AC, Machado FS, Celes MR, Faria G, Rocha LB, Silva JS, Rossi MA. Mitochondrial damage as an early event of monensin-induced cell injury in cultured fibroblasts L929. J Vet Med A Physiol Pathol Clin Med. 2005, 52, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Jia M, Shi M, Zhao Y, Li Y, Liu X, Zhao L. The role of ATF3 in the crosstalk between cellular stress response and ferroptosis in tumors. Crit Rev Oncol Hematol. 2025, 213, 104791. [Google Scholar] [CrossRef] [PubMed]
- Dayekh K, Johnson-Obaseki S, Corsten M, Villeneuve PJ, Sekhon HS, Weberpals JI, Dimitroulakos J. Monensin inhibits epidermal growth factor receptor trafficking and activation: synergistic cytotoxicity in combination with EGFR inhibitors. Mol Cancer Ther. 2014, 13, 2559–2571. [Google Scholar] [CrossRef] [PubMed]
- Yoon MJ, Kang YJ, Kim IY, Kim EH, Lee JA, Lim JH, Kwon TK, Choi KS. Monensin, a polyether ionophore antibiotic, overcomes TRAIL resistance in glioma cells via endoplasmic reticulum stress, DR5 upregulation and c-FLIP downregulation. Carcinogenesis. 2013, 34, 1918–1928. [Google Scholar] [CrossRef] [PubMed]
- Serter Kocoglu S, Secme M, Oy C, Korkusuz G, Elmas L. Monensin, an Antibiotic Isolated from Streptomyces Cinnamonensis, Regulates Human Neuroblastoma Cell Proliferation via the PI3K/AKT Signaling Pathway and Acts Synergistically with Rapamycin. Antibiotics (Basel). 2023, 12, 546. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fu B, Fang L, Wang R, Zhang X. Inhibition of Wnt/β-catenin signaling by monensin in cervical cancer. Korean J Physiol Pharmacol. 2024, 28, 21–30. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tumova L, Pombinho AR, Vojtechova M, Stancikova J, Gradl D, Krausova M, Sloncova E, Horazna M, Kriz V, Machonova O, Jindrich J, Zdrahal Z, Bartunek P, Korinek V. Monensin inhibits canonical Wnt signaling in human colorectal cancer cells and suppresses tumor growth in multiple intestinal neoplasia mice. Mol Cancer Ther. 2014, 13, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Verma SP, Das P. Monensin induces cell death by autophagy and inhibits matrix metalloproteinase 7 (MMP7) in UOK146 renal cell carcinoma cell line. In Vitro Cell Dev Biol Anim. 2018, 54, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Nakaya H, Kamoi K, Tran AM, Cochran DL. Effects of Monensin on the Immunocytochemical Detection of Matrix Metalloproteinase 3 in Human Periodontal Ligament Cells. Nihon Shishubyo Gakkai Kaishi (Journal of the Japanese Society of Periodontology). 1996, 38, 48–53. [Google Scholar] [CrossRef]
- Vanneste M, Huang Q, Li M, Moose D, Zhao L, Stamnes MA, Schultz M, Wu M, Henry MD. High content screening identifies monensin as an EMT-selective cytotoxic compound. Sci Rep. 2019, 9, 1200. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tan X, Cardin DL, Wang S, Xu Y, Russell WK. Monensin suppresses EMT-driven cancer cell motility by inducing Golgi pH-dependent exocytosis of GOLIM4. Proc Natl Acad Sci U S A. 2025, 122, e2501347122. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ling Y, Priebe W, Perezsoler R. Intrinsic cytotoxicity and reversal of multidrug-resistance by monensin in kb parent and mdr cells. Int J Oncol. 1993, 3, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Fang T, Hu S, Song X, Wang J, Zuo R, Yun S, Jiang S, Guo D. Combination of monensin and erlotinib synergistically inhibited the growth and cancer stem cell properties of triple-negative breast cancer by simultaneously inhibiting EGFR and PI3K signaling pathways. Breast Cancer Research and Treatment. 2024, 207, 435–451. [Google Scholar] [CrossRef]
- Blain M, Garrard A, Poppenga R, Chen B, Valento M, Halliday Gittinger M. Survival after severe rhabdomyolysis following monensin ingestion. Journal of Medical Toxicology. 2017, 13, 259–262. [Google Scholar] [CrossRef]
- Butaye P, Devriese LA, Haesebrouck F. Antimicrobial growth promoters used in animal feed: effects of less well-known antibiotics on gram-positive bacteria. Clin Microbiol Rev. 2003, 16, 175–188. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang M, Xu N, Xu W, Ling G, Zhang P. Potential therapies and diagnosis based on Golgi-targeted nano drug delivery systems. Pharmacol Res. 2022, 175, 105861. [Google Scholar] [CrossRef] [PubMed]
- Kerr CL, Pennington TH. The effect of monensin on virion production and protein secretion in pseudorabies virus-infected cells. J Gen Virol. 1984, 65 Pt 6, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Melroy D, Jones RL. The effect of monensin on intracellular transport and secretion of α-amylase isoenzymes in barley aleurone. Planta. 1986, 167, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Mollenhauer HH, Rowe LD, Witzel DA. Effect of monensin on the morphology of mitochondria in rodent and equine striated muscle. Vet Hum Toxicol. 1984, 26, 15–19. [Google Scholar] [PubMed]
- Henn D, Lensink AV, Botha CJ. Ultrastructural changes in cardiac and skeletal myoblasts following in vitro exposure to monensin, salinomycin, and lasalocid. PLoS One. 2024, 19, e0311046. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Deng Y, Zhang J, Wang Z, Yan Z, Qiao M, Ye J, Wei Q, Wang J, Wang X, Zhao L, Lu S, Tang S, Mohammed MK, Liu H, Fan J, Zhang F, Zou Y, Liao J, Qi H, Haydon RC, Luu HH, He TC, Tang L. Antibiotic monensin synergizes with EGFR inhibitors and oxaliplatin to suppress the proliferation of human ovarian cancer cells. Sci Rep. 2015, 5, 17523. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhao X, Ma Y, Luo J, Xu K, Tian P, Lu C, Song J. Blocking the WNT/β-catenin pathway in cancer treatment:pharmacological targets and drug therapeutic potential. Heliyon. 2024, 10, e35989. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang X, Wu X, Zhang Z, Ma C, Wu T, Tang S, Zeng Z, Huang S, Gong C, Yuan C, Zhang L, Feng Y, Huang B, Liu W, Zhang B, Shen Y, Luo W, Wang X, Liu B, Lei Y, Ye Z, Zhao L, Cao D, Yang L, Chen X, Haydon RC, Luu HH, Peng B, Liu X, He TC. Monensin inhibits cell proliferation and tumor growth of chemo-resistant pancreatic cancer cells by targeting the EGFR signaling pathway. Sci Rep. 2018, 8, 17914. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Vraila M, Asp E, Melo FR, Grujic M, Rollman O, Pejler G, Lampinen M. Monensin induces secretory granule-mediated cell death in eosinophils. J Allergy Clin Immunol. 2023, 152, 1312–1320.e3. [Google Scholar] [CrossRef] [PubMed]
- Tumova L, Pombinho AR, Vojtechova M, Stancikova J, Gradl D, Krausova M, Sloncova E, Horazna M, Kriz V, Machonova O, Jindrich J, Zdrahal Z, Bartunek P, Korinek V. Monensin inhibits canonical Wnt signaling in human colorectal cancer cells and suppresses tumor growth in multiple intestinal neoplasia mice. Mol Cancer Ther. 2014, 13, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Yoon MJ, Kang YJ, Kim IY, Kim EH, Lee JA, Lim JH, Kwon TK, Choi KS. Monensin, a polyether ionophore antibiotic, overcomes TRAIL resistance in glioma cells via endoplasmic reticulum stress, DR5 upregulation and c-FLIP downregulation. Carcinogenesis. 2013, 34, 1918–1928. [Google Scholar] [CrossRef] [PubMed]
- Charvat RA, Arrizabalaga G. Oxidative stress generated during monensin treatment contributes to altered Toxoplasma gondii mitochondrial function. Sci Rep. 2016, 6, 22997. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bharti H, Singal A, Raza M, Ghosh PC, Nag A. Ionophores as Potent Anti-malarials: A Miracle in the Making. Curr Top Med Chem. 2019, 18, 2029–2041. [Google Scholar] [CrossRef] [PubMed]
- Ivanova J, Kamenova K, Petrova E, Vladov I, Gluhcheva Y, Dorkov P. Comparative study on the effects of salinomycin, monensin and meso-2,3-dimercaptosuccinic acid on the concentrations of lead, calcium, copper, iron and zinc in lungs and heart in lead-exposed mice. J Trace Elem Med Biol. 2020, 58, 126429. [Google Scholar] [CrossRef] [PubMed]
- Dragwidge JM, Scholl S, Schumacher K, Gendall AR. NHX-type Na+(K+)/H+ antiporters are required for TGN/EE trafficking and endosomal ion homeostasis in Arabidopsis thaliana. J Cell Sci. 2019, 132, jcs226472. [Google Scholar] [CrossRef] [PubMed]
- Song X, Akasaka H, Wang H, Abbasgholizadeh R, Shin JH, Zang F, Chen J, Logsdon CD, Maitra A, Bean AJ, Wang H. Hematopoietic progenitor kinase 1 down-regulates the oncogenic receptor tyrosine kinase AXL in pancreatic cancer. J Biol Chem. 2020, 295, 2348–2358. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kawano M, Ueno A, Ashida Y, Matsumoto N, Inoue H. Effects of sialagogues on ornithine decarboxylase induction and proto-oncogene expression in murine parotid gland. J Dent Res. 1992, 71, 1885–1890. [Google Scholar] [CrossRef] [PubMed]
- Li Y, Sun Q, Chen S, Yu X, Jing H. Monensin Inhibits Anaplastic Thyroid Cancer via Disrupting Mitochondrial Respiration and AMPK/mTOR Signaling. Anticancer Agents Med Chem. 2022, 22, 2539–2547. [Google Scholar] [CrossRef] [PubMed]
- Tan X, Cardin DL, Wang S, Xu Y, Russell WK. Monensin suppresses EMT-driven cancer cell motility by inducing Golgi pH-dependent exocytosis of GOLIM4. Proc Natl Acad Sci U S A. 2025, 122, e2501347122. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang Y, Wang Y, Read E, Fu M, Pei Y, Wu L, Wang R, Yang G. Golgi Stress Response, Hydrogen Sulfide Metabolism, and Intracellular Calcium Homeostasis. Antioxid Redox Signal. 2020, 32, 583–601. [Google Scholar] [CrossRef] [PubMed]
- Xu F, Zhong H, Chang Y, Li D, Jin H, Zhang M, Wang H, Jiang C, Shen Y, Huang Y. Targeting death receptors for drug-resistant cancer therapy: Codelivery of pTRAIL and monensin using dual-targeting and stimuli-responsive self-assembling nanocomposites. Biomaterials. 2018, 158, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak A, Heflin B, Siegel E, Reed MR, Nix JS, Yee EU, Jędrzejczyk M, Klejborowska G, Stępczyńska N, Huczyński A, Nagalo MB, Chambers TC, Post S, Eoff RL, MacNicol MC, Tiwari AK, Kelly T, Tackett AJ, MacNicol AM. Monensin and Its Analogs Exhibit Activity Against Breast Cancer Stem-Like Cells in an Organoid Model. bioRxiv [Preprint]. 2025 Jul 9:2025.07.05.663311. [CrossRef] [PubMed] [PubMed Central]
- Fiorilla S, Tasso F, Clemente N, Trisciuoglio T, Boldorini R, Carini R. Monensin Inhibits Triple-Negative Breast Cancer in Mice by a Na+-Dependent Cytotoxic Action Unrelated to Cytostatic Effects. Cells. 2025, 14, 185. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yang J, Dong Y, Xu J, Qian X, Cai Y, Chen Y, Zhang P, Gao D, Cui Z, Cui Y. TM9SF3 is a Golgi-resident ATG8-binding protein essential for Golgi-selective autophagy. Dev Cell. S: 27, 1534. [CrossRef] [PubMed]
- Yang J, Cui Y. TM9SF3 is a mammalian Golgiphagy receptor that safeguards Golgi integrity and glycosylation fidelity. Autophagy. 1: 28. [CrossRef] [PubMed]
- Fang T, Hu S, Song X, Wang J, Zuo R, Yun S, Jiang S, Guo D. Combination of monensin and erlotinib synergistically inhibited the growth and cancer stem cell properties of triple-negative breast cancer by simultaneously inhibiting EGFR and PI3K signaling pathways. Breast Cancer Res Treat. 2024, 207, 435–451. [Google Scholar] [CrossRef] [PubMed]
- Chen L, Jiang P, Shen X, Lyu J, Liu C, Li L, Huang Y. Cascade Delivery to Golgi Apparatus and On-Site Formation of Subcellular Drug Reservoir for Cancer Metastasis Suppression. Small. 2023, 19, e2204747. [Google Scholar] [CrossRef] [PubMed]
- Marjanović M, Mikecin Dražić AM, Mioč M, Paradžik M, Kliček F, Novokmet M, Lauc G, Kralj M. Salinomycin disturbs Golgi function and specifically affects cells in epithelial-to-mesenchymal transition. J Cell Sci. 2023, 136, jcs260934. [Google Scholar] [CrossRef] [PubMed]
- Silva SV, Lima MA, Hodgson L, Freitas VM, Rodríguez-Manzaneque JC. ADAMTS-1 has nuclear localization in cells with epithelial origin and leads to decreased cell migration. Exp Cell Res. 2023, 433, 113852. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hansen MB, Postol M, Tvingsholm S, Nielsen IØ, Dietrich TN, Puustinen P, Maeda K, Dinant C, Strauss R, Egan D, Jäättelä M, Kallunki T. Identification of lysosome-targeting drugs with anti-inflammatory activity as potential invasion inhibitors of treatment resistant HER2 positive cancers. Cell Oncol (Dordr). 2021, 44, 805–820. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gu J, Huang L, Zhang Y. Monensin inhibits proliferation, migration, and promotes apoptosis of breast cancer cells via downregulating UBA2. Drug Dev Res. 2020, 81, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Russo-Abrahão T, Lacerda-Abreu MA, Gomes T, Cosentino-Gomes D, Carvalho-de-Araújo AD, Rodrigues MF, Oliveira ACL, Rumjanek FD, Monteiro RQ, Meyer-Fernandes JR. Characterization of inorganic phosphate transport in the triple-negative breast cancer cell line, MDA-MB-231. PLoS One. 2018, 13, e0191270. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bareford LM, Phelps MA, Foraker AB, Swaan PW. Intracellular processing of riboflavin in human breast cancer cells. Mol Pharm. 2008, 5, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Ketola K, Vainio P, Fey V, Kallioniemi O, Iljin K. Monensin is a potent inducer of oxidative stress and inhibitor of androgen signaling leading to apoptosis in prostate cancer cells. Mol Cancer Ther. 2010, 9, 3175–3185. [Google Scholar] [CrossRef] [PubMed]
- Kim SH, Kim KY, Yu SN, Park SG, Yu HS, Seo YK, Ahn SC. Monensin induces PC-3 prostate cancer cell apoptosis via ROS production and Ca2+ homeostasis disruption. Anticancer research. 2016, 36, 5835–5843. [Google Scholar] [CrossRef]
- He Z, Zhang Y, Mehta SK, Pierson DL, Wu H, Rohde LH. Expression profile of apoptosis-related genes and radio-sensitivity of prostate cancer cells. J Radiat Res. 2011, 52, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Ye Z, Rohde LH, Wu H. Radio-sensitization of Prostate Cancer Cells by Monensin Treatment and its associated Gene Expression Profiling Changes. In54th Annual Meeting of the Radiation Research Society 2008 Jan 1.
- Ketola K, Vuoristo A, Oresic M, Kallioniemi O, Iljin K. Abstract A62: Monensin-induced oxidative stress reduces prostate cancer cell motility and cancer stem cell markers. Molecular Cancer Therapeutics. 2011, 10 (Suppl. 11). [Google Scholar]
- Kaneski CR, Constantopoulos G, Brady RO. Effect of dimethylsulfoxide on the proliferation and glycosaminoglycan synthesis of rat prostate adenocarcinoma cells (PAIII) in vitro: isolation and characterization of DMSO-resistant cells. Prostate. 1991, 18, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Seçme M, Kocoglu SS. Investigation of the TLR4 and IRF3 signaling pathway-mediated effects of monensin in colorectal cancer cells. Med Oncol. 2023, 40, 187. [Google Scholar] [CrossRef] [PubMed]
- Zhou Y, Deng Y, Wang J, Yan Z, Wei Q, Ye J, Zhang J, He TC, Qiao M. Effect of antibiotic monensin on cell proliferation and IGF1R signaling pathway in human colorectal cancer cells. Ann Med. 2023, 55, 954–964. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tumova L, Pombinho AR, Vojtechova M, Stancikova J, Gradl D, Krausova M, Sloncova E, Horazna M, Kriz V, Machonova O, Jindrich J, Zdrahal Z, Bartunek P, Korinek V. Monensin inhibits canonical Wnt signaling in human colorectal cancer cells and suppresses tumor growth in multiple intestinal neoplasia mice. Mol Cancer Ther. 2014, 13, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Park WH, Kim ES, Jung CW, Kim BK, Lee YY. Monensin-mediated growth inhibition of SNU-C1 colon cancer cells via cell cycle arrest and apoptosis. Int J Oncol. 2003, 22, 377–382. [Google Scholar] [PubMed]
- Owatari S, Akune S, Komatsu M, Ikeda R, Firth SD, Che XF, Yamamoto M, Tsujikawa K, Kitazono M, Ishizawa T, Takeuchi T, Aikou T, Mercer JF, Akiyama S, Furukawa T. Copper-transporting P-type ATPase, ATP7A, confers multidrug resistance and its expression is related to resistance to SN-38 in clinical colon cancer. Cancer Res. 2007, 67, 4860–4868. [Google Scholar] [CrossRef] [PubMed]
- Paduch R, Kandefer-Szerszeń M. Expression and activation of proteases in co-cultures. Exp Toxicol Pathol. 2011, 63, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Miraglia E, Viarisio D, Riganti C, Costamagna C, Ghigo D, Bosia A. Na+/H+ exchanger activity is increased in doxorubicin-resistant human colon cancer cells and its modulation modifies the sensitivity of the cells to doxorubicin. Int J Cancer. 2005, 115, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Griffin T, Rybak ME, Recht L, Singh M, Salimi A, Raso V. Potentiation of antitumor immunotoxins by liposomal monensin. J Natl Cancer Inst. 1993, 85, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Singh M, Griffin T, Salimi A, Micetich RG, Atwal H. Potentiation of ricin A immunotoxin by monoclonal antibody targeted monensin containing small unilamellar vesicles. Cancer Lett. 1994, 84, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Griffin TW, Pagnini PG, McGrath JJ, McCann JC, Houston LL. In vitro cytotoxicity of recombinant ricin A chain-antitransferrin receptor immunotoxin against human adenocarcinomas of the colon and pancreas. J Biol Response Mod. 1988, 7, 559–567. [Google Scholar] [PubMed]
- Griffin TW, Childs LR, FitzGerald DJ, Levin LV. Enhancement of the cytotoxic effect of anti-carcinoembryonic antigen immunotoxins by adenovirus and carboxylic ionophores. J Natl Cancer Inst. 1987, 79, 679–685. [Google Scholar] [PubMed]
- Batra S, Alenfall J. Effect of diverse categories of drugs on human colon tumour cell proliferation. Anticancer Res. 1991, 11, 1221–1224. [Google Scholar] [PubMed]
- Clemente N, Baroni S, Fiorilla S, Tasso F, Reano S, Borsotti C, Ruggiero MR, Alchera E, Corrazzari M, Walker G, Follenzi A, Crich SG, Carini R. Boosting intracellular sodium selectively kills hepatocarcinoma cells and induces hepatocellular carcinoma tumor shrinkage in mice. Commun Biol. 2023, 6, 574. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhu Q, Liao S, Wei T, Liu S, Yang C, Tang J. Development of a novel prognostic signature based on cytotoxic T lymphocyte-evasion genes for hepatocellular carcinoma patient management. Discov Oncol. 2025, 16, 144. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Park WH, Lee MS, Park K, Kim ES, Kim BK, Lee YY. Monensin-mediated growth inhibition in acute myelogenous leukemia cells via cell cycle arrest and apoptosis. International journal of cancer. 2002, 101, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Park WH, Kim ES, Kim BK, Lee YY. Monensin-mediated growth inhibition in NCI-H929 myeloma cells via cell cycle arrest and apoptosis. International journal of oncology. 2003, 23, 197–204. [Google Scholar]
- Yusenko MV, Trentmann A, Andersson MK, Ghani LA, Jakobs A, Paz MF, Mikesch JH, von Kries JP, Stenman G, Klempnauer KH. Monensin, a novel potent MYB inhibitor, suppresses proliferation of acute myeloid leukemia and adenoid cystic carcinoma cells. Cancer letters. 2020, 479, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Srinivas RV, Melsen LR, Compans RW. Effects of monensin on morphogenesis and infectivity of Friend murine leukemia virus. Journal of Virology. 1982, 42, 1067–1075. [Google Scholar] [CrossRef]
- Akin DT, Kinkade Jr JM, Parmley RT. Biochemical and ultrastructural effects of monensin on the processing, intracellular transport, and packaging of myeloperoxidase into low and high density compartments of human leukemia (HL-60) cells. Archives of biochemistry and biophysics. 1987, 257, 451–463. [Google Scholar] [CrossRef]
- Zeng C, Long M, Lu Y. Monensin synergizes with chemotherapy in uveal melanoma through suppressing RhoA. Immunopharmacology and Immunotoxicology. 2023, 45, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Hertler AA, Schlossman DM, Borowitz MJ, Blythman HE, Casellas P, Frankel AE. An anti-CD5 immunotoxin for chronic lymphocytic leukemia: enhancement of cytotoxicity with human serum albumin-monensin. International journal of cancer. 1989, 43, 215–219. [Google Scholar] [CrossRef]
- Park WH, Seol JG, Kim ES, Kang WK, Im YH, Jung CW, Kim BK, Lee YY. Monensin-mediated growth inhibition in human lymphoma cells through cell cycle arrest and apoptosis. Br J Haematol. 2002, 119, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Torun A, Zdanowicz A, Miazek-Zapala N, Zapala P, Pradhan B, Jedrzejczyk M, Ciechanowicz A, Pilch Z, Skorzynski M, Słabicki M, Rymkiewicz G, Barankiewicz J, Martines C, Laurenti L, Struga M, Winiarska M, Golab J, Kucia M, Ratajczak MZ, Huczynski A, Calado DP, Efremov DG, Zerrouqi A, Pyrzynska B. Potassium/sodium cation carriers robustly upregulate CD20 antigen by targeting MYC, and synergize with anti- CD20 immunotherapies to eliminate malignant B cells. Haematologica. 2025, 110, 1555–1572. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Di Virgilio S, Rampelberg M, Greimers R, Schnek G, Hooghe R. The effects of monensin on blood-borne arrest and glycosylation of BL/VL3 lymphoma cells. Cell Biochem Funct. 1992, 10, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Park WH, Kim ES, Kim BK, Lee YY. Monensin-mediated growth inhibition in NCI-H929 myeloma cells via cell cycle arrest and apoptosis. Int J Oncol. 2003, 23, 197–204. [Google Scholar] [PubMed]
- Park WH, Kim ES, Jung CW, Kim BK, Lee YY. Monensin-mediated growth inhibition of SNU-C1 colon cancer cells via cell cycle arrest and apoptosis. Int J Oncol. 2003, 22, 377–382. [Google Scholar] [PubMed]
- Steinmetz TD, Schlötzer-Schrehardt U, Hearne A, Schuh W, Wittner J, Schulz SR, Winkler TH, Jäck HM, Mielenz D. TFG is required for autophagy flux and to prevent endoplasmic reticulum stress in CH12 B lymphoma cells. Autophagy. 2021, 17, 2238–2256. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Augustine EC, Kensinger RS. Endocytosis and degradation of ovine prolactin by Nb2 lymphoma cells: characterization and effects of agents known to alter prolactin-induced mitogenesis. Endocrinology. 1990, 127, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Van Der Luit AH, Budde M, Verheij M, Van Blitterswijk WJ. Different modes of internalization of apoptotic alkyl-lysophospholipid and cell-rescuing lysophosphatidylcholine. Biochem J. 2003, 374 Pt 3, 747–753. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang X, Wu X, Zhang Z, Ma C, Wu T, Tang S, Zeng Z, Huang S, Gong C, Yuan C, Zhang L, Feng Y, Huang B, Liu W, Zhang B, Shen Y, Luo W, Wang X, Liu B, Lei Y, Ye Z, Zhao L, Cao D, Yang L, Chen X, Haydon RC, Luu HH, Peng B, Liu X, He TC. Monensin inhibits cell proliferation and tumor growth of chemo-resistant pancreatic cancer cells by targeting the EGFR signaling pathway. Sci Rep. 2018, 8, 17914. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Pádua D, Figueira P, Pombinho A, Monteiro I, Pereira CF, Almeida R, Mesquita P. HMGA1 stimulates cancer stem-like features and sensitivity to monensin in gastric cancer. Exp Cell Res. 2024, 442, 114257. [Google Scholar] [CrossRef] [PubMed]
- Pádua D, Barros R, Amaral AL, Mesquita P, Freire AF, Sousa M, Maia AF, Caiado I, Fernandes H, Pombinho A, Pereira CF, Almeida R. A SOX2 Reporter System Identifies Gastric Cancer Stem-Like Cells Sensitive to Monensin. Cancers (Basel). 2020, 12, 495. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tokui T, Takatori T, Shinozaki N, Ishigami M, Shiraishi A, Ikeda T, Tsuruo T. Delivery and cytotoxicity of RS-1541 in St-4 human gastric cancer cells in vitro by the low-density-lipoprotein pathway. Cancer Chemother Pharmacol. 1995, 36, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yao S, Wang W, Zhou B, Cui X, Yang H, Zhang S. Monensin suppresses cell proliferation and invasion in ovarian cancer by enhancing MEK1 SUMOylation. Exp Ther Med. 2021, 22, 1390. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Deng Y, Zhang J, Wang Z, Yan Z, Qiao M, Ye J, Wei Q, Wang J, Wang X, Zhao L, Lu S, Tang S, Mohammed MK, Liu H, Fan J, Zhang F, Zou Y, Liao J, Qi H, Haydon RC, Luu HH, He TC, Tang L. Antibiotic monensin synergizes with EGFR inhibitors and oxaliplatin to suppress the proliferation of human ovarian cancer cells. Sci Rep. 2015, 5, 17523. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hansen MB, Postol M, Tvingsholm S, Nielsen IØ, Dietrich TN, Puustinen P, Maeda K, Dinant C, Strauss R, Egan D, Jäättelä M, Kallunki T. Identification of lysosome-targeting drugs with anti-inflammatory activity as potential invasion inhibitors of treatment resistant HER2 positive cancers. Cell Oncol (Dordr). 2021, 44, 805–820. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fu B, Fang L, Wang R, Zhang X. Inhibition of Wnt/β-catenin signaling by monensin in cervical cancer. Korean J Physiol Pharmacol. 2024, 28, 21–30. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Choi HS, Jeong EH, Lee TG, Kim SY, Kim HR, Kim CH. Autophagy Inhibition with Monensin Enhances Cell Cycle Arrest and Apoptosis Induced by mTOR or Epidermal Growth Factor Receptor Inhibitors in Lung Cancer Cells. Tuberc Respir Dis (Seoul). 2013, 75, 9–17. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ochi K, Suzawa K, Tomida S, Shien K, Takano J, Miyauchi S, Takeda T, Miura A, Araki K, Nakata K, Yamamoto H, Okazaki M, Sugimoto S, Shien T, Yamane M, Azuma K, Okamoto Y, Toyooka S. Overcoming epithelial-mesenchymal transition-mediated drug resistance with monensin-based combined therapy in non-small cell lung cancer. Biochem Biophys Res Commun. 2020, 529, 760–765. [Google Scholar] [CrossRef] [PubMed]
- Tan X, Cardin DL, Wang S, Xu Y, Russell WK. Monensin suppresses EMT-driven cancer cell motility by inducing Golgi pH-dependent exocytosis of GOLIM4. Proc Natl Acad Sci U S A. 2025, 122, e2501347122. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chung BM, Raja SM, Clubb RJ, Tu C, George M, Band V, Band H. Aberrant trafficking of NSCLC-associated EGFR mutants through the endocytic recycling pathway promotes interaction with Src. BMC Cell Biol. 2009, 10, 84. [Google Scholar] [CrossRef]
- Cleary I, Doherty G, Moran E, Clynes M. The multidrug-resistant human lung tumour cell line, DLKP-A10, expresses novel drug accumulation and sequestration systems. Biochem Pharmacol. 1997, 53, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Zhou Y, Liu Q, Dai X, Yan Y, Yang Y, Li H, Zhou X, Gao W, Li X, Xi Z. Transdifferentiation of type II alveolar epithelial cells induces reactivation of dormant tumor cells by enhancing TGF-β1/SNAI2 signaling. Oncol Rep. 2018, 39, 1874–1882. [Google Scholar] [CrossRef] [PubMed]
- Mastroianni A, Invernizzi L, Tagliabue E, Fassina G, Albini A, Ménard S, Colnaghi MI. Alteration of laminin production in small-cell lung carcinoma: possible correlation with the absence of the basement membrane. Tumour Biol. 1993, 14, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Derbyshire EJ, Henry RV, Stahel RA, Wawrzynczak EJ. Potent cytotoxic action of the immunotoxin SWA11-ricin A chain against human small cell lung cancer cell lines. Br J Cancer. 1992, 66, 444–451. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Urbaniak A, Reed MR, Heflin B, Gaydos J, Piña-Oviedo S, Jędrzejczyk M, Klejborowska G, Stępczyńska N, Chambers TC, Tackett AJ, Rodriguez A, Huczyński A, Eoff RL, MacNicol AM. Anti-glioblastoma activity of monensin and its analogs in an organoid model of cancer. Biomed Pharmacother. 2022, 153, 113440. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wan W, Zhang X, Huang C, Chen L, Yang X, Bao K, Peng T. Monensin inhibits glioblastoma angiogenesis via targeting multiple growth factor receptor signaling. Biochem Biophys Res Commun. 2020, 530, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Ko M, Makena MR, Schiapparelli P, Suarez-Meade P, Mekile AX, Lal B, Lopez-Bertoni H, Kozielski KL, Green JJ, Laterra J, Quiñones-Hinojosa A, Rao R. The endosomal pH regulator NHE9 is a driver of stemness in glioblastoma. PNAS Nexus. 2022, 1, pgac013. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Schraen-Maschke S, Zanetta JP. Role of oligomannosidic N-glycans in the proliferation, adhesion and signalling of C6 glioblastoma cells. Biochimie. 2003, 85, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Chignola R, Foroni R, Candiani C, Franceschi A, Pasti M, Stevanoni G, Anselmi C, Tridente G, Colombatti M. Cytoreductive effects of anti-transferrin receptor immunotoxin in a multicellular tumor spheroid model. Int J Cancer. 1994, 57, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Griffin T, Rybak ME, Recht L, Singh M, Salimi A, Raso V. Potentiation of antitumor immunotoxins by liposomal monensin. J Natl Cancer Inst. 1993, 85, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Zovickian J, Johnson VG, Youle RJ. Potent and specific killing of human malignant brain tumor cells by an anti-transferrin receptor antibody-ricin immunotoxin. J Neurosurg. 1987, 66, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Griffin T, Rybak ME, Recht L, Singh M, Salimi A, Raso V. Potentiation of antitumor immunotoxins by liposomal monensin. J Natl Cancer Inst. 1993, 85, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Basque J, Martel M, Leduc R, Cantin AM. Lysosomotropic drugs inhibit maturation of transforming growth factor-beta. Can J Physiol Pharmacol. 2008, 86, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Businaro R, Fabrizi C, Fumagalli L, Lauro GM. Synthesis and secretion of alpha 2-macroglobulin by human glioma established cell lines. Exp Brain Res. 1992, 88, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Benya RV, Kusui T, Shikado F, Battey JF, Jensen RT. Desensitization of neuromedin B receptors (NMB-R) on native and NMB-R-transfected cells involves down-regulation and internalization. J Biol Chem. 1994, 269, 11721–11728. [Google Scholar] [CrossRef] [PubMed]
- Xin H, Li J, Zhang H, Li Y, Zeng S, Wang Z, Zhang Z, Deng F. Monensin may inhibit melanoma by regulating the selection between differentiation and stemness of melanoma stem cells. PeerJ. 2019, 7, e7354. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zeng C, Long M, Lu Y. Monensin synergizes with chemotherapy in uveal melanoma through suppressing RhoA. Immunopharmacol Immunotoxicol. 2023, 45, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Bumol TF, Reisfeld RA. Biosynthesis and secretion of fibronectin in human melanoma cells. J Cell Biochem. 1983, 21, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Tumova L, Pombinho AR, Vojtechova M, Stancikova J, Gradl D, Krausova M, Sloncova E, Horazna M, Kriz V, Machonova O, Jindrich J, Zdrahal Z, Bartunek P, Korinek V. Monensin inhibits canonical Wnt signaling in human colorectal cancer cells and suppresses tumor growth in multiple intestinal neoplasia mice. Mol Cancer Ther. 2014, 13, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Fukao H, Ueshima S, Sakai T, Okada K, Matsuo O. Effect of monensin on secretion of t-PA from melanoma (Bowes). Cell Struct Funct. 1989, 14, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Negroiu G, Dwek RA, Petrescu SM. Tyrosinase-related protein-2 and -1 are trafficked on distinct routes in B16 melanoma cells. Biochem Biophys Res Commun. 2005, 328, 914–921. [Google Scholar] [CrossRef] [PubMed]
- Roth JA, Ames RS, Fry K, Lee HM, Scannon PJ. Mediation of reduction of spontaneous and experimental pulmonary metastases by ricin A-chain immunotoxin 45-2D9-RTA with potentiation by systemic monensin in mice. Cancer Res. 1988, 48, 3496–3501. [Google Scholar] [PubMed]
- Saeki H, Oikawa A. Stimulation by ionophores of tyrosinase activity of mouse melanoma cells in culture. J Invest Dermatol. 1985, 85, 423–425. [Google Scholar] [CrossRef] [PubMed]
- 27. Jara JR, Martinez-Liarte JH, Solano F. Transport of L-tyrosine by B16/F10 malignant melanocytes: characterization of the process. Pigment Cell Res. 1990, 3, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Verma SP, Das P. Monensin induces cell death by autophagy and inhibits matrix metalloproteinase 7 (MMP7) in UOK146 renal cell carcinoma cell line. In Vitro Cell Dev Biol Anim. 2018, 54, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Park WH, Jung CW, Park JO, Kim K, Kim WS, Im YH, Lee MH, Kang WK, Park K. Monensin inhibits the growth of renal cell carcinoma cells via cell cycle arrest or apoptosis. Int J Oncol. 2003, 22, 855–860. [Google Scholar] [PubMed]
- Xiao GF, Yan X, Chen Z, Zhang RJ, Liu TZ, Hu WL. Identification of a Novel Immune-Related Prognostic Biomarker and Small-Molecule Drugs in Clear Cell Renal Cell Carcinoma (ccRCC) by a Merged Microarray-Acquired Dataset and TCGA Database. Front Genet. 2020, 11, 810. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Park WH, Kim ES, Kim BK, Lee YY. Monensin-mediated growth inhibition in NCI-H929 myeloma cells via cell cycle arrest and apoptosis. Int J Oncol. 2003, 23, 197–204. [Google Scholar] [PubMed]
- Puvvula PK, Martinez-Medina L, Cinar M, Feng L, Pisarev A, Johnson A, Bernal-Mizrachi L. A retrotransposon-derived DNA zip code internalizes myeloma cells through Clathrin-Rab5a-mediated endocytosis. Front Oncol. 2024, 14, 1288724. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li Y, Sun Q, Chen S, Yu X, Jing H. Monensin Inhibits Anaplastic Thyroid Cancer via Disrupting Mitochondrial Respiration and AMPK/mTOR Signaling. Anticancer Agents Med Chem. 2022, 22, 2539–2547. [Google Scholar] [CrossRef] [PubMed]
- Pooley L, Shakur Y, Rena G, Houslay MD. Intracellular localization of the PDE4A cAMP-specific phosphodiesterase splice variant RD1 (RNPDE4A1A) in stably transfected human thyroid carcinoma FTC cell lines. Biochem J. 1997, 321 Pt 1, 177–85. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Plattner VE, Wagner M, Ratzinger G, Gabor F, Wirth M. Targeted drug delivery: binding and uptake of plant lectins using human 5637 bladder cancer cells. Eur J Pharm Biopharm. 2008, 70, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Gamou S, Shimizu N. Glycosylation of the epidermal growth factor receptor and its relationship to membrane transport and ligand binding. J Biochem. 1988, 104, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Gamou S, Hirai M, Rikimaru K, Enomoto S, Shimizu N. Biosynthesis of the epidermal growth factor receptor in human squamous cell carcinoma lines: secretion of the truncated receptor is not common to epidermal growth factor receptor-hyperproducing cells. Cell Struct Funct. 1988, 13, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Orr RB, Kamen BA. UMSCC38 cells amplified at 11q13 for the folate receptor synthesize a mutant nonfunctional folate receptor. Cancer Res. 1994, 54, 3905–3911. [Google Scholar] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



