Submitted:
20 September 2025
Posted:
23 September 2025
You are already at the latest version
Abstract
The urgent need for decentralized, rapid, and affordable point-of-care (POC) diagnostic tools has been starkly highlighted by recent global health crises, underscoring their critical role in achieving health equity. Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-Cas systems, repurposed from their revolutionary gene-editing origins, have emerged as a transformative platform technology for molecular diagnostics, offering unparalleled programmability, specificity, and sensitivity for nucleic acid detection. The core of these diagnostics lies in the unique catalytic mechanisms of specific Cas effectors, notably the target-activated, non-specific trans-cleavage activity of Cas12 and Cas13 proteins. This collateral cleavage of reporter molecules provides a powerful intrinsic mechanism for signal amplification, enabling sensitive detection. This review critically examines the rapid evolution of CRISPR-based diagnostics, from their fundamental molecular principles to their real-world applications. Key applications in infectious disease surveillance and management are summarized, including the rapid development of assays for SARS-CoV-2, HIV, and tuberculosis, with a particular focus on adaptations for field-deployable formats such as lateral flow assays. As the technology continues to mature, overcoming remaining hurdles in sample preparation, multiplexing, and reagent stability will be paramount. Ultimately, CRISPR-based diagnostics are poised to become an indispensable and democratizing component of the global health toolkit, empowering frontline healthcare workers and transforming disease control on a global scale.
Keywords:
CRISPR
; diagnostics
; point-of-care testing (POCT)
; Cas12
; Cas13
; SHERLOCK
; DETECTR
; global health
; molecular diagnostics
; nucleic acid detection
1. Introduction
The established paradigm of molecular diagnostics has long been anchored in centralized laboratories, relying on sophisticated infrastructure and highly trained personnel to perform gold-standard techniques like the polymerase chain reaction (PCR). While powerful, this model is defined by inherent limitations: high costs, complex instrumentation, long turnaround times, and logistical dependence on a robust cold chain for reagent transport and storage. These constraints are particularly acute in low-resource settings and become system-wide bottlenecks during public health emergencies, severely hampering effective disease surveillance and containment. The COVID-19 pandemic exposed the brittleness of this centralized model on a global scale, creating an unprecedented demand for diagnostic tools that could be deployed at the point of care (POC). This has intensified the pursuit of technologies that meet the World Health Organization’s (WHO) “ASSURED” criteria for POC tests: Affordable, Sensitive, Specific, User-friendly, Rapid and robust, Equipment-free, and Deliverable to end-users.
Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and CRISPR-associated (Cas) protein systems have emerged as a uniquely suited solution to the POC challenge. Originally discovered as an adaptive immune system in bacteria and archaea, their utility has expanded far beyond the Nobel Prize-winning application of gene editing. The diagnostic potential of CRISPR-Cas systems stems from a combination of powerful intrinsic properties. First is their exquisite programmability; the specificity of a Cas enzyme is directed by a small, easy-to-synthesize guide RNA (gRNA), allowing a diagnostic test to be rapidly designed or re-tasked for virtually any nucleic acid target. Second is their remarkable specificity, which enables the discrimination of closely related viral strains or even single-nucleotide polymorphisms (SNPs), a critical feature for identifying drug-resistance mutations or viral variants of concern. Third is their compatibility with isothermal operation; when coupled with upstream isothermal amplification methods, CRISPR-based assays can achieve attomolar sensitivity without the need for the complex thermal cyclers required for PCR, making them highly adaptable for field use.
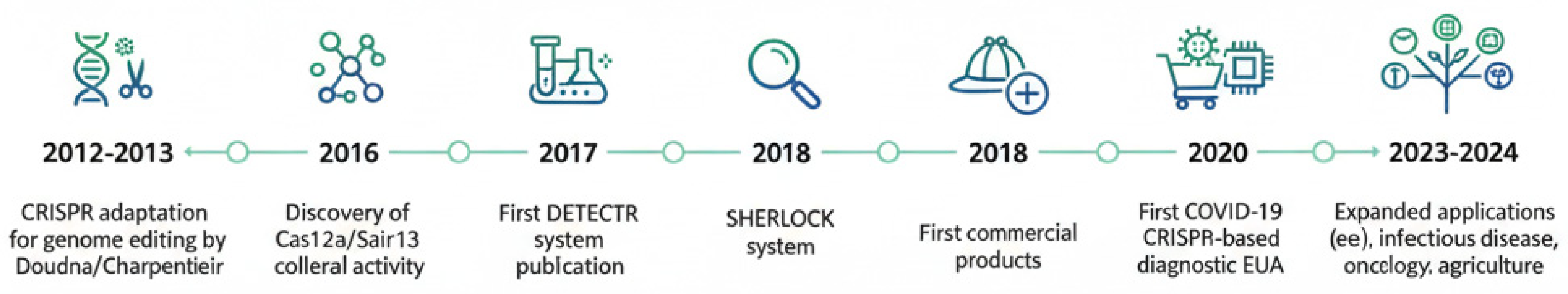
Figure 1.
Timeline of key milestones in the development of CRISPR-based diagnostics. This timeline highlights the rapid evolution of CRISPR technology from its Nobel Prize-winning application in gene editing to a versatile diagnostic platform. Key events include the initial description of CRISPR-Cas9 for genome editing (2012), the discovery of the RNA-targeting Cas13 enzyme (2017), and the subsequent development of the foundational diagnostic platforms SHERLOCK (Cas13-based) and DETECTR (Cas12-based) in 2018. The urgent need for decentralized testing during the COVID-19 pandemic (2020) served as a major catalyst, accelerating innovation and the development of commercial assays. More recent advances focus on enhancing multiplexing capabilities and streamlining workflows for broader applications.
Figure 1.
Timeline of key milestones in the development of CRISPR-based diagnostics. This timeline highlights the rapid evolution of CRISPR technology from its Nobel Prize-winning application in gene editing to a versatile diagnostic platform. Key events include the initial description of CRISPR-Cas9 for genome editing (2012), the discovery of the RNA-targeting Cas13 enzyme (2017), and the subsequent development of the foundational diagnostic platforms SHERLOCK (Cas13-based) and DETECTR (Cas12-based) in 2018. The urgent need for decentralized testing during the COVID-19 pandemic (2020) served as a major catalyst, accelerating innovation and the development of commercial assays. More recent advances focus on enhancing multiplexing capabilities and streamlining workflows for broader applications.
This review critically evaluates the rapid evolution of CRISPR-based diagnostics (CRISPR-Dx), from their fundamental molecular mechanisms to their translation into field-deployable tools for global health. It will analyze the core biochemical principles that enable detection, survey key applications in the management of pandemic, endemic, and drug-resistant pathogens, and provide a balanced assessment of the significant hurdles—technical, logistical, and regulatory—that remain on the path to widespread adoption. The central thesis is that CRISPR-Dx represents a democratizing technology with the potential to shift the diagnostic landscape, moving molecular testing from the lab to life and empowering a new era of decentralized disease management.
2. Core Mechanisms of CRISPR-Based Detection
The power of CRISPR-Dx is rooted in the unique biochemical activities of specific Class 2 Cas effector proteins, particularly those from the Cas12 and Cas13 families. Upon recognizing their designated nucleic acid target, these enzymes unleash a secondary, non-specific nuclease activity. This “collateral cleavage” is the cornerstone of signal amplification in the most prominent CRISPR diagnostic platforms.
2.1. The Cas12 Family (e.g., DETECTR): DNA-Targeted Collateral Cleavage
The Cas12 family of enzymes, particularly Cas12a (formerly Cpf1), are RNA-guided DNA endonucleases that form the basis of platforms like DETECTR (DNA Endonuclease-Targeted CRISPR Trans Reporter). The detection mechanism begins with the formation of a ribonucleoprotein (RNP) complex between the Cas12a protein and a CRISPR RNA (crRNA). For target recognition, the Cas12a RNP scans double-stranded DNA (dsDNA) for a specific, short sequence known as a protospacer adjacent motif (PAM), which is typically T-rich (e.g., 5′-TTTV-3′) for commonly used orthologs like LbCas12a. Upon binding to the PAM, the crRNA’s spacer sequence hybridizes with the complementary target DNA strand, creating an R-loop structure. This binding event induces a significant conformational change in the Cas12a protein, which activates its single RuvC nuclease domain. This activation results in two distinct catalytic activities:
- Cis-cleavage: The specific, staggered cleavage of both strands of the target dsDNA molecule.
- Trans-cleavage (collateral activity): Once activated, the Cas12a RNP becomes a non-specific DNase, indiscriminately cleaving any single-stranded DNA (ssDNA) molecules in its vicinity. This collateral activity is a high-turnover process, with a single target recognition event triggering the cleavage of thousands of bystander ssDNA molecules.
This trans-cleavage is the key to signal generation in DETECTR-based assays. It is important to note that the reported kinetic parameters, such as the turnover rate (kcat), can vary dramatically (from 0.02 to 17 s−1) depending on the specific Cas12 homolog, crRNA design, target sequence, and reaction conditions, making careful optimization essential for robust assay development.
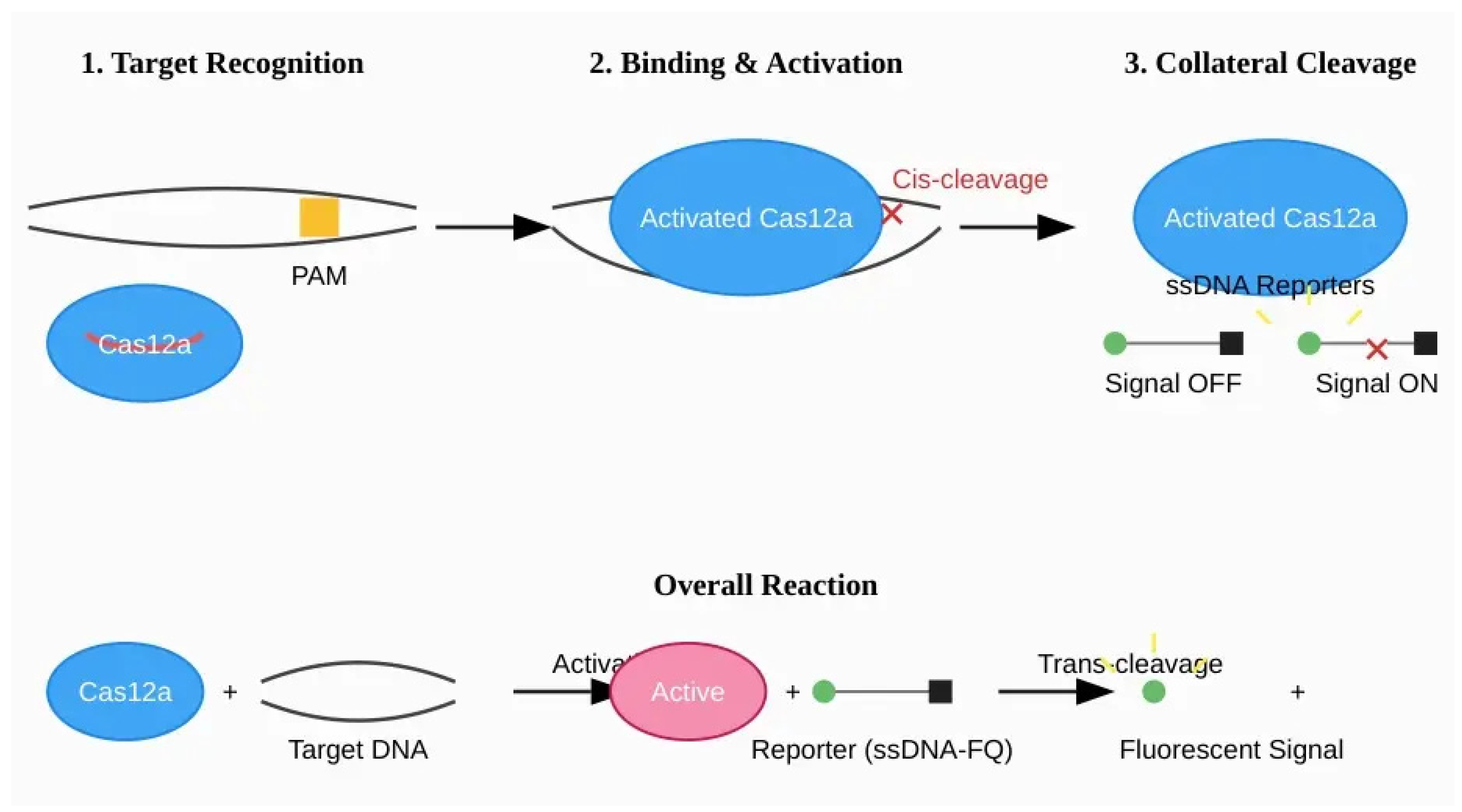
Figure 2.
Mechanism of DETECTR (Cas12a-based) diagnostics. (1) The Cas12a-crRNA ribonucleoprotein (RNP) complex scans dsDNA for a specific Protospacer Adjacent Motif (PAM). (2) Upon PAM recognition and target DNA binding, the complex undergoes a conformational change, activating its nuclease domain. This leads to specific cis-cleavage of the target DNA. (3) The activated enzyme then exhibits non-specific trans-cleavage (collateral) activity, indiscriminately cleaving nearby single-stranded DNA (ssDNA) reporter molecules. Cleavage of these ssDNA-FQ reporters separates a fluorophore (F) from a quencher (Q), generating a detectable fluorescent signal.
Figure 2.
Mechanism of DETECTR (Cas12a-based) diagnostics. (1) The Cas12a-crRNA ribonucleoprotein (RNP) complex scans dsDNA for a specific Protospacer Adjacent Motif (PAM). (2) Upon PAM recognition and target DNA binding, the complex undergoes a conformational change, activating its nuclease domain. This leads to specific cis-cleavage of the target DNA. (3) The activated enzyme then exhibits non-specific trans-cleavage (collateral) activity, indiscriminately cleaving nearby single-stranded DNA (ssDNA) reporter molecules. Cleavage of these ssDNA-FQ reporters separates a fluorophore (F) from a quencher (Q), generating a detectable fluorescent signal.
2.2. The Cas13 Family (e.g., SHERLOCK): RNA-Targeted Collateral Cleavage
The Cas13 family of enzymes are RNA-guided RNA-targeting ribonucleases that underpin platforms like SHERLOCK (Specific High-sensitivity Enzymatic Reporter unLOCKing). Unlike Cas12, Cas13 enzymes directly target single-stranded RNA (ssRNA). The Cas13-crRNA complex binds to its complementary ssRNA target without a strict PAM requirement, although some orthologs exhibit a preference for a protospacer flanking site (PFS). Target RNA binding activates the enzyme’s two Higher Eukaryotes and Prokaryotes Nucleotide-binding (HEPN) domains, which come together to form a single, active catalytic site for RNase activity. This activation unleashes a potent collateral cleavage effect, whereby the enzyme begins to indiscriminately degrade any nearby, non-target ssRNA molecules. This promiscuous RNase activity is the engine of signal amplification in SHERLOCK-based diagnostics. The reported turnover number for Cas13 trans-cleavage can be very high, ranging from 1 to 700 s−1, enabling substantial signal generation from a small number of target recognition events.
Figure 3.
Comparative analysis of key Cas effector enzymes for diagnostics. The table compares the distinct biochemical properties of Cas12a, Cas13a, and the more recently characterized Cas14. These differences, including target preference (dsDNA, ssRNA, or ssDNA), the requirement for a Protospacer Adjacent Motif (PAM) sequence for target recognition, and collateral cleavage activity, make each enzyme uniquely suited for different diagnostic applications. For example, Cas13a’s ability to directly target ssRNA is ideal for detecting RNA viruses, while Cas12a’s dsDNA targeting is used for DNA pathogens or DNA amplicons.
Figure 3.
Comparative analysis of key Cas effector enzymes for diagnostics. The table compares the distinct biochemical properties of Cas12a, Cas13a, and the more recently characterized Cas14. These differences, including target preference (dsDNA, ssRNA, or ssDNA), the requirement for a Protospacer Adjacent Motif (PAM) sequence for target recognition, and collateral cleavage activity, make each enzyme uniquely suited for different diagnostic applications. For example, Cas13a’s ability to directly target ssRNA is ideal for detecting RNA viruses, while Cas12a’s dsDNA targeting is used for DNA pathogens or DNA amplicons.
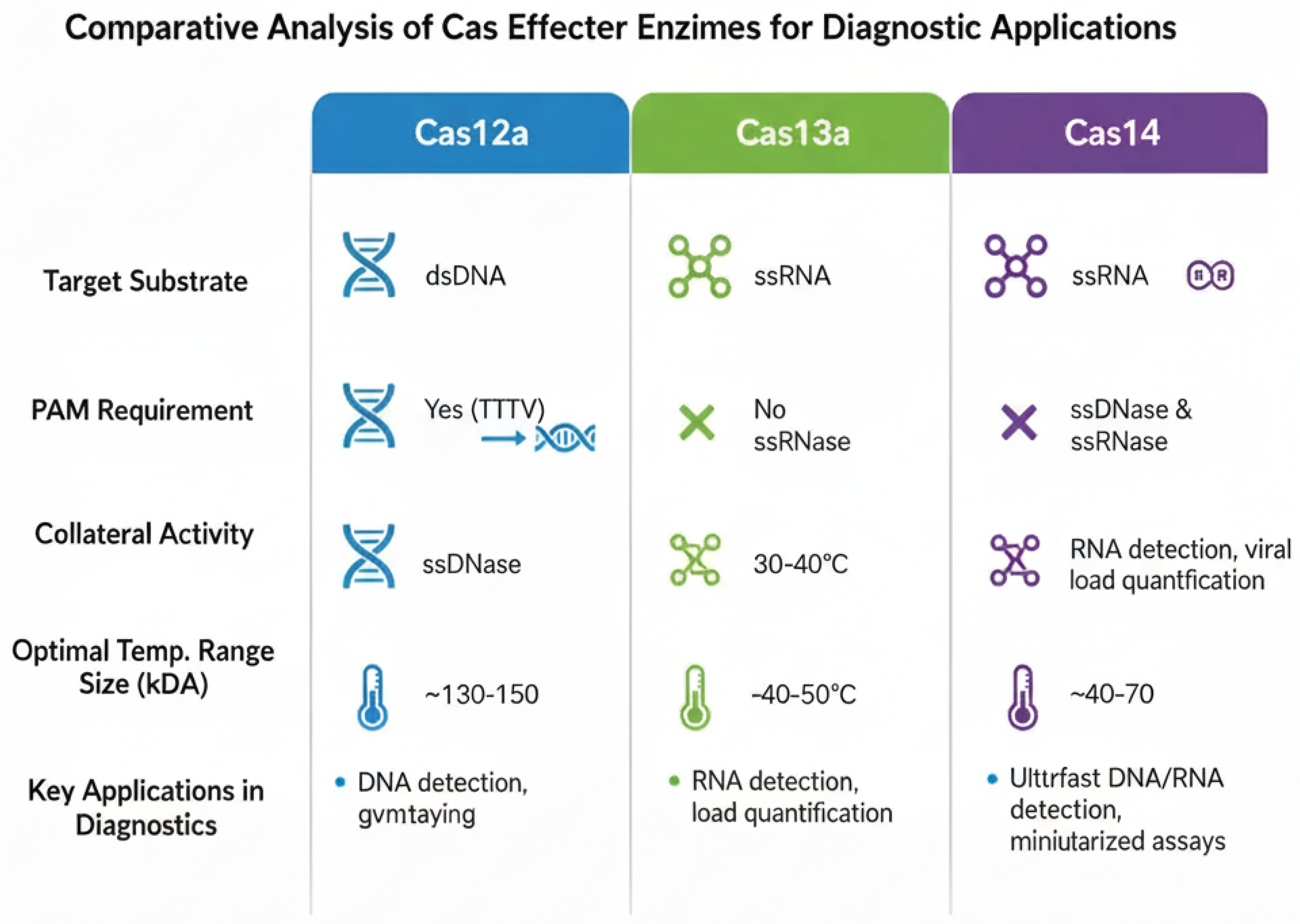
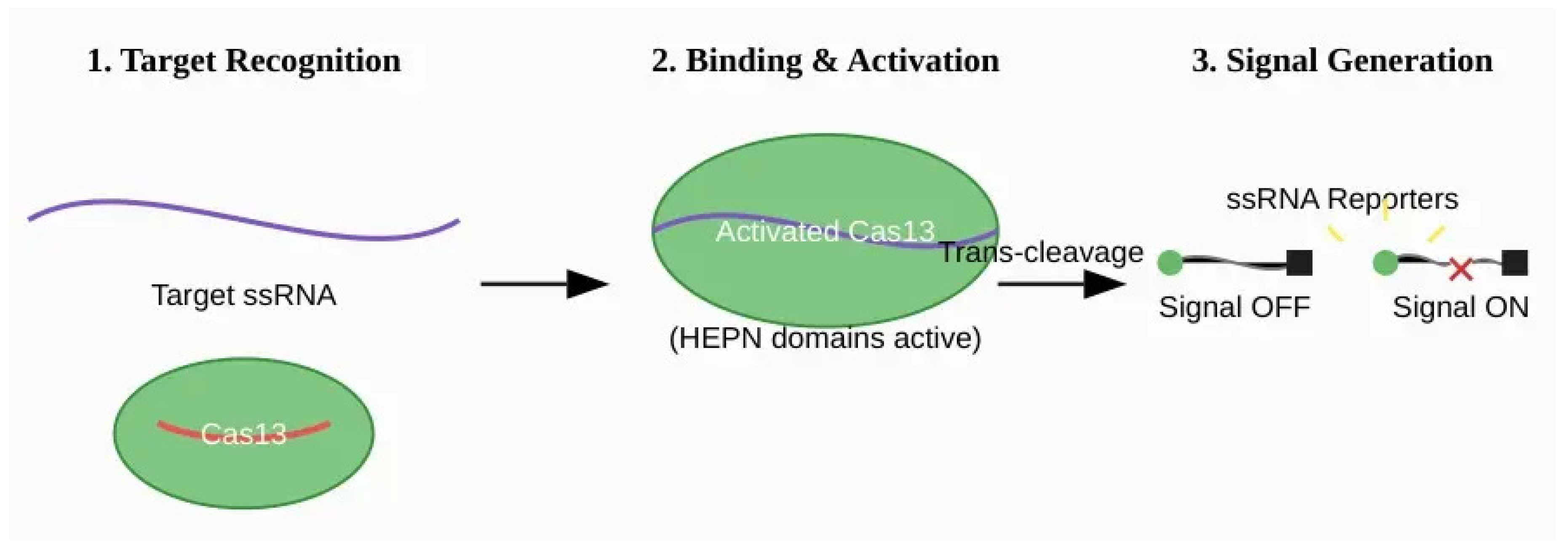
Figure 4.
Mechanism of SHERLOCK (Cas13-based) diagnostics. (1) The Cas13-crRNA RNP complex recognizes and binds to a complementary target single-stranded RNA (ssRNA) molecule. This binding does not require a strict PAM sequence. (2) Target binding activates the two HEPN nuclease domains of the Cas13 enzyme. (3) The activated Cas13 becomes a promiscuous RNase, engaging in potent trans-cleavage of any nearby non-target ssRNA molecules, including synthetic ssRNA-FQ reporters. This collateral activity cleaves the reporters, separating the fluorophore from the quencher and producing a fluorescent signal.
Figure 4.
Mechanism of SHERLOCK (Cas13-based) diagnostics. (1) The Cas13-crRNA RNP complex recognizes and binds to a complementary target single-stranded RNA (ssRNA) molecule. This binding does not require a strict PAM sequence. (2) Target binding activates the two HEPN nuclease domains of the Cas13 enzyme. (3) The activated Cas13 becomes a promiscuous RNase, engaging in potent trans-cleavage of any nearby non-target ssRNA molecules, including synthetic ssRNA-FQ reporters. This collateral activity cleaves the reporters, separating the fluorophore from the quencher and producing a fluorescent signal.
2.3. Signal Amplification and Readout
CRISPR diagnostic systems harness the collateral cleavage activity of Cas12 and Cas13 by introducing synthetic reporter molecules into the reaction. For Cas12-based assays, these are typically short ssDNA molecules labeled with a fluorophore on one end and a quencher on the other (ssDNA-FQ). For Cas13, the reporters are analogous ssRNA-FQ molecules. In their intact state, the quencher suppresses the fluorophore’s signal. When the Cas enzyme is activated by the target nucleic acid, its collateral activity cleaves these reporters, separating the fluorophore from the quencher and producing a measurable fluorescent signal that indicates a positive result.
For POC applications, this mechanism has been ingeniously coupled with simple, instrument-free visual readouts, most notably lateral flow assays (LFAs). In a common LFA design, the ssDNA reporter is labeled with two different molecules (haptens), such as fluorescein (FAM) at one end and biotin at the other. The LFA strip contains two lines: a control line with streptavidin (which binds biotin) and a test line with anti-FAM antibodies. If the target is absent, the intact FAM-biotin reporters are captured at the first (control) line. If the target is present, Cas12a cleaves the reporters, separating FAM from biotin. The FAM-labeled fragments are no longer captured at the control line and instead flow to the test line, where they are captured by the anti-FAM antibodies, producing a visible colored band that signifies a positive result. While effective, early LFA designs were susceptible to false positives, leading to the development of improved formats that enhance specificity and reliability, such as those based on competitive assays or chain hybridization strategies [15].
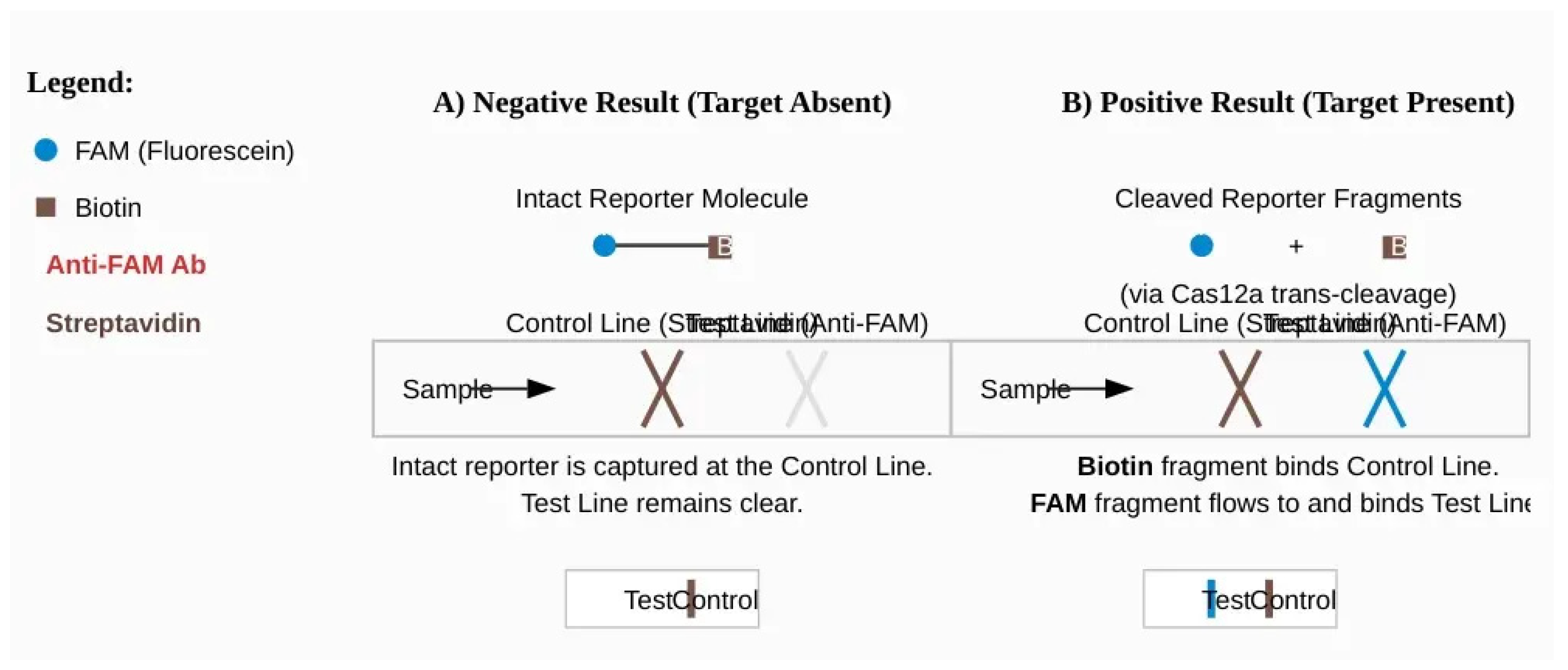
Figure 5.
Principle of a CRISPR-based Lateral Flow Assay (LFA) for visual readout. The assay uses a dual-labeled ssDNA reporter with FAM (F) and Biotin (B). The LFA strip has a Control Line coated with streptavidin (binds Biotin) and a Test Line coated with anti-FAM antibodies. (A) Negative Result: In the absence of the target, the Cas enzyme is inactive, and the F-B reporters remain intact. They are captured by streptavidin at the Control Line, resulting in a single visible band. (B) Positive Result: In the presence of the target, the activated Cas enzyme cleaves the reporters. The separated Biotin-containing fragments are captured at the Control Line, while the FAM-containing fragments flow past and are captured by the anti-FAM antibodies at the Test Line. The appearance of two bands (Test and Control) indicates a positive result.
Figure 5.
Principle of a CRISPR-based Lateral Flow Assay (LFA) for visual readout. The assay uses a dual-labeled ssDNA reporter with FAM (F) and Biotin (B). The LFA strip has a Control Line coated with streptavidin (binds Biotin) and a Test Line coated with anti-FAM antibodies. (A) Negative Result: In the absence of the target, the Cas enzyme is inactive, and the F-B reporters remain intact. They are captured by streptavidin at the Control Line, resulting in a single visible band. (B) Positive Result: In the presence of the target, the activated Cas enzyme cleaves the reporters. The separated Biotin-containing fragments are captured at the Control Line, while the FAM-containing fragments flow past and are captured by the anti-FAM antibodies at the Test Line. The appearance of two bands (Test and Control) indicates a positive result.
2.4. Isothermal Pre-Amplification: Achieving Attomolar Sensitivity
While the collateral activity of Cas enzymes provides signal amplification, the initial concentration of pathogen nucleic acids in clinical samples (e.g., blood, saliva, swabs) is often too low for direct detection. To bridge this sensitivity gap, CRISPR-Dx platforms almost universally incorporate an upstream target amplification step. To maintain suitability for POC use, this is achieved using isothermal amplification techniques that operate at a single, constant temperature, thereby avoiding the need for bulky and expensive thermal cyclers. Two methods are predominant, with distinct operational trade-offs:

Table 1.
Comparison of Recombinase Polymerase Amplification (RPA) and Loop-Mediated Isothermal Amplification (LAMP). The table outlines key features of the two predominant isothermal amplification methods used in CRISPR-Dx, including operating temperature, reaction time, primer design, specificity, and advantages.
Table 1.
Comparison of Recombinase Polymerase Amplification (RPA) and Loop-Mediated Isothermal Amplification (LAMP). The table outlines key features of the two predominant isothermal amplification methods used in CRISPR-Dx, including operating temperature, reaction time, primer design, specificity, and advantages.
| Feature | Recombinase Polymerase Amplification (RPA) | Loop-Mediated Isothermal Amplification (LAMP) |
| Operating Temperature | 37–42°C (low, body temperature range) | 60–65°C (higher, requires simple heating block) |
| Reaction Time | Fast (typically <20 minutes) | Rapid (15-60 minutes) |
| Primer Design | Moderate complexity (one pair, 30-35 nt) | High complexity (4-6 specialized primers) |
| Specificity | High (based on 2 primers) | Very High (based on 6-8 target regions) |
| Key Advantage | Speed and low temperature operation | High efficiency and robustness against inhibitors |
This system architecture can be conceptualized as a two-tiered amplification engine. The first stage, isothermal amplification, serves to amplify the target nucleic acid, increasing its concentration from potentially just a few copies per microliter (attomolar levels) to a level that is readily detectable. This step provides the raw sensitivity. However, isothermal methods can sometimes generate non-specific amplicons, which could lead to false positives. The second stage, CRISPR-Cas detection, acts as a highly specific verifier. The Cas RNP will only be activated by the correct amplicon sequence, ignoring any spurious amplification products. Its subsequent collateral cleavage of thousands of reporter molecules then amplifies the signal, providing a final, robust, and highly specific readout. This elegant two-stage process is what endows CRISPR-Dx with its hallmark combination of ultra-high sensitivity and specificity, but it also introduces complexities in reagent compatibility and reaction workflow that are the focus of ongoing innovation, such as the development of single-pot assays.
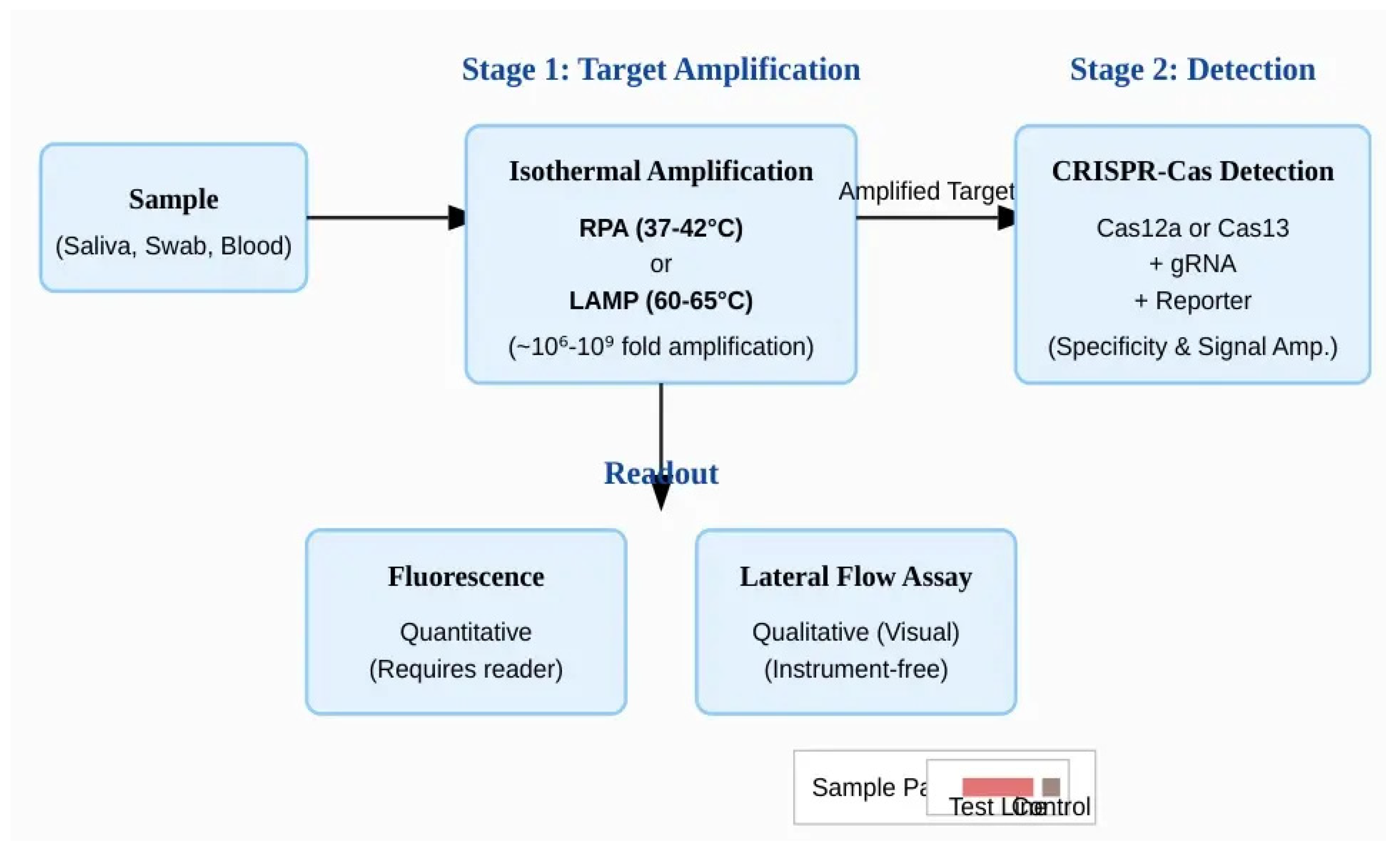
Figure 6.
The two-stage architecture of a typical CRISPR-Dx assay. The workflow begins with a clinical sample. In Stage 1 (Target Amplification), the target nucleic acid concentration is increased millions- to billions-fold using an isothermal method like RPA or LAMP. This provides the necessary sensitivity for detection. In Stage 2 (Detection), the amplified product is introduced to the CRISPR-Cas system. The Cas enzyme, guided by a specific gRNA, recognizes the correct target sequence and activates its collateral cleavage activity, which in turn acts on reporter molecules. This step provides high specificity and signal amplification. The final result is visualized through a readout method, such as a quantitative fluorescence measurement or a qualitative, instrument-free lateral flow assay.
Figure 6.
The two-stage architecture of a typical CRISPR-Dx assay. The workflow begins with a clinical sample. In Stage 1 (Target Amplification), the target nucleic acid concentration is increased millions- to billions-fold using an isothermal method like RPA or LAMP. This provides the necessary sensitivity for detection. In Stage 2 (Detection), the amplified product is introduced to the CRISPR-Cas system. The Cas enzyme, guided by a specific gRNA, recognizes the correct target sequence and activates its collateral cleavage activity, which in turn acts on reporter molecules. This step provides high specificity and signal amplification. The final result is visualized through a readout method, such as a quantitative fluorescence measurement or a qualitative, instrument-free lateral flow assay.
3. Key Applications in Global Health and Infectious Disease
The programmability and field-deployable nature of CRISPR-Dx have enabled its application to a wide spectrum of global health challenges, from pandemic response to the management of long-standing endemic diseases.
3.1. Viral Pandemics: A Proving Ground for Rapid Response
The COVID-19 pandemic served as an unprecedented proving ground for CRISPR-Dx, demonstrating its capacity for rapid development and deployment. Within weeks of the SARS-CoV-2 genome being published, researchers had adapted both DETECTR (Cas12-based) and SHERLOCK (Cas13-based) platforms to detect the virus from patient respiratory swabs. A seminal study on a DETECTR-based assay for SARS-CoV-2 demonstrated remarkable clinical performance, achieving 95% positive predictive agreement and 100% negative predictive agreement when compared to the gold-standard RT-qPCR test. This work, and others like it, highlighted the technology’s potential as a faster and more accessible alternative to lab-based PCR. Key innovations during this period included the development of one-pot assays, such as STOPCovid, which combine amplification and detection in a single tube to simplify the workflow, and extraction-free protocols that lyse the virus directly in the sample, bypassing the need for laborious nucleic acid purification.
The utility of CRISPR-Dx extends to other viruses with pandemic potential. Assays have been developed to detect Zika virus (ZIKV) RNA in clinical samples, providing a specific tool to manage outbreaks of the mosquito-borne illness. For influenza, CRISPR-based platforms like SHINE (Streamlined Highlighting of Infections to Navigate Epidemics) have demonstrated the ability not only to differentiate between influenza A and B species but also to distinguish clinically relevant subtypes like H1N1 and H3N2 with 100% concordance to RT-qPCR. Critically, the high sequence specificity of CRISPR allows for the rapid adaptation of these assays to new and emerging viral threats, a crucial capability for future pandemic preparedness.
3.2. Endemic Diseases
Beyond pandemic response, CRISPR-Dx offers significant promise for managing endemic diseases that disproportionately affect low-resource settings. For tuberculosis (TB), a leading cause of death globally, CRISPR-based assays are being developed to rapidly detect Mycobacterium tuberculosis and identify drug-resistance mutations directly from sputum samples, which traditionally require lengthy culture-based methods. Similarly, in the fight against HIV, CRISPR-Dx platforms are being explored for viral load monitoring, offering a simpler and more affordable alternative to conventional PCR-based methods, which is crucial for effective treatment management in remote areas. The technology also holds potential for malaria diagnosis, enabling sensitive detection of Plasmodium parasites, including those with drug-resistant mutations, directly from blood samples, which could significantly aid in surveillance and control efforts.
3.3. Antimicrobial Resistance (AMR)
Antimicrobial Resistance (AMR) represents a growing global health crisis, demanding rapid and accurate diagnostic tools to guide appropriate antibiotic stewardship. CRISPR-Dx is uniquely positioned to address this challenge by rapidly identifying bacterial species and their associated AMR genes directly from clinical samples. The ability to quickly detect resistance markers can inform treatment decisions, helping to preserve the efficacy of existing antibiotics and prevent the spread of drug-resistant infections.
3.4. Other Applications
Beyond infectious diseases, the utility of CRISPR-Dx is expanding into diverse fields. In oncology, it enables the early detection of cancer-associated mutations and circulating tumor DNA (ctDNA) in liquid biopsies [30]. In genetics, it is used for identifying markers for inherited disorders and single-nucleotide polymorphisms (SNPs). Its application also extends to environmental monitoring for pathogens and contaminants in water, soil, and air [33], ensuring food safety through the rapid identification of foodborne pathogens and allergens [34], and diagnosing plant and animal diseases in agriculture [35].
4. Bridging the Gap: Overcoming Hurdles to Widespread Adoption
Despite the immense potential of CRISPR-Dx, several significant hurdles must be overcome to translate these technologies from laboratory prototypes to widely adopted clinical and public health tools. These challenges span technical, logistical, and regulatory domains.
4.1. Technical and Logistical Challenges
One of the most significant bottlenecks for true point-of-care application is sample preparation. While extraction-free protocols have been developed, their performance can be inconsistent across different sample types (e.g., blood, saliva, sputum), which often contain inhibitors that can interfere with both the amplification and CRISPR-Cas reactions. Developing robust, universal, and simple methods for sample lysis and nucleic acid release that work reliably across diverse clinical matrices is a critical area of ongoing research.
Multiplexing—the ability to detect multiple targets in a single test—is another key challenge. While some CRISPR-based assays have demonstrated the ability to detect a handful of targets simultaneously, achieving high-level multiplexing (e.g., for comprehensive respiratory panels or AMR gene panels) remains difficult. This is due to the potential for cross-reactivity between different guide RNAs and the complexity of designing orthogonal reporter systems that can be read out simultaneously. Furthermore, the stability of reagents, particularly the enzymes (polymerases, reverse transcriptases, Cas proteins), is a major logistical concern. Many of these proteins require a continuous cold chain for transport and storage, which is often unavailable in low-resource settings. Lyophilization (freeze-drying) of reagents into stable, room-temperature-compatible formats is a promising solution, but it requires careful optimization to ensure that the enzymes retain their activity upon rehydration.
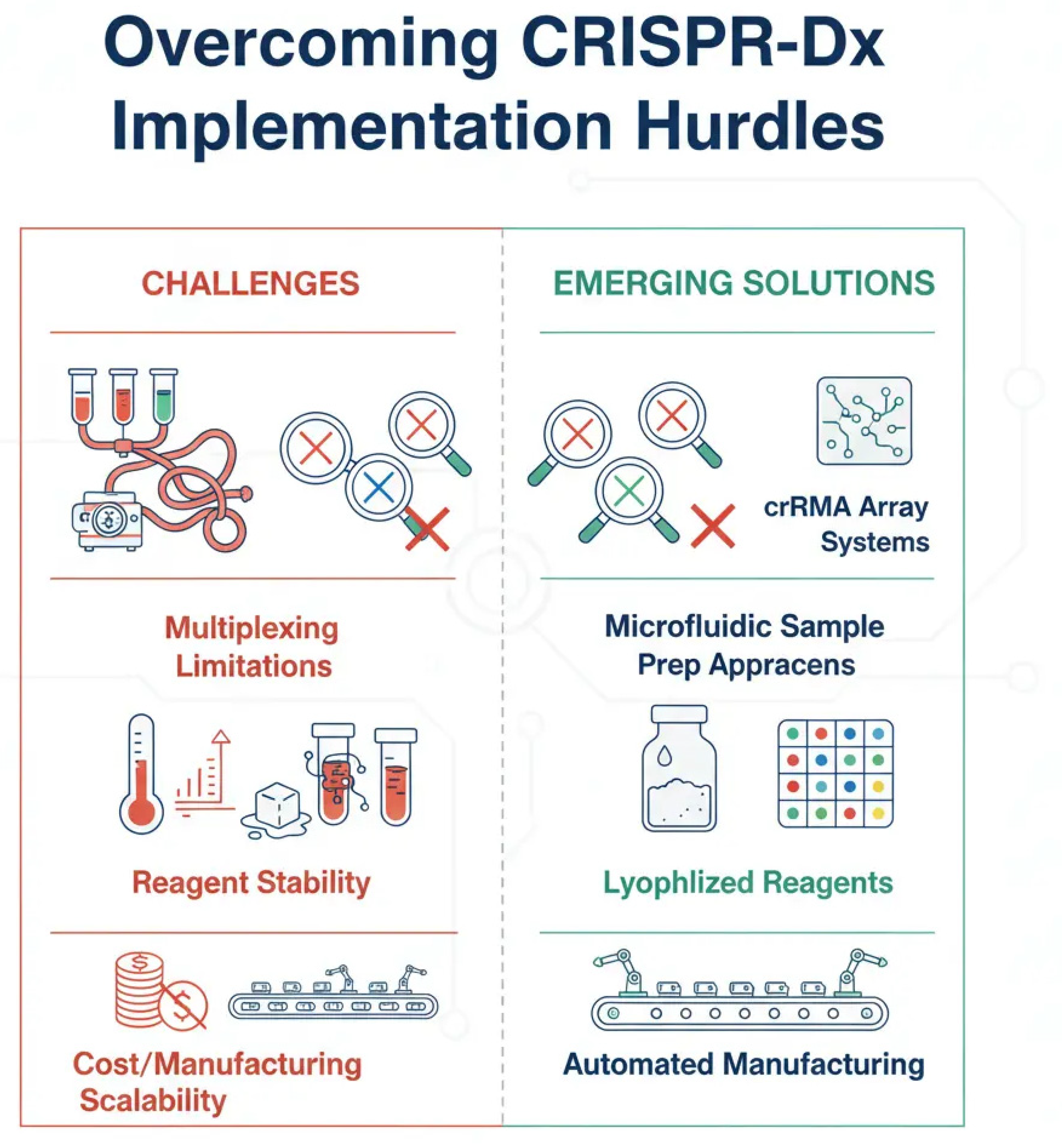
Figure 7.
Key implementation hurdles for CRISPR-Dx and emerging solutions. This diagram illustrates the primary challenges hindering the widespread, point-of-care adoption of CRISPR diagnostics and the innovative strategies being developed to overcome them. (A) Multiplexing: The challenge of detecting multiple targets simultaneously is being addressed by designing orthogonal crRNA arrays and integrating assays with microfluidic devices. (B) Reagent Stability: The need for a cold chain is a major logistical barrier, which is being solved by lyophilizing (freeze-drying) enzymatic reagents into stable, room-temperature formats. (C) Cost & Accessibility: High costs and complex workflows are being reduced through automation and the development of simplified, single-pot reaction protocols.
Figure 7.
Key implementation hurdles for CRISPR-Dx and emerging solutions. This diagram illustrates the primary challenges hindering the widespread, point-of-care adoption of CRISPR diagnostics and the innovative strategies being developed to overcome them. (A) Multiplexing: The challenge of detecting multiple targets simultaneously is being addressed by designing orthogonal crRNA arrays and integrating assays with microfluidic devices. (B) Reagent Stability: The need for a cold chain is a major logistical barrier, which is being solved by lyophilizing (freeze-drying) enzymatic reagents into stable, room-temperature formats. (C) Cost & Accessibility: High costs and complex workflows are being reduced through automation and the development of simplified, single-pot reaction protocols.
4.2. Regulatory and Commercial Landscape.
Navigating the regulatory landscape is a crucial step for the clinical implementation of any new diagnostic technology. While the FDA has granted Emergency Use Authorization (EUA) to several CRISPR-based tests for SARS-CoV-2, the path to full regulatory approval (e.g., 510(k) clearance or De Novo classification) is more stringent and requires extensive clinical validation to demonstrate safety and efficacy. Establishing clear regulatory pathways and harmonizing standards across different countries will be essential for global deployment.
Finally, cost and manufacturing at scale are significant considerations. While the per-test cost of CRISPR-Dx is projected to be low (often cited as a few dollars), this depends on the ability to manufacture the core components—particularly the Cas enzymes and guide RNAs—at high volume and low cost. Ensuring quality control and batch-to-batch consistency during large-scale production is a non-trivial manufacturing challenge that must be addressed for the technology to be economically viable and accessible in the markets that need it most.
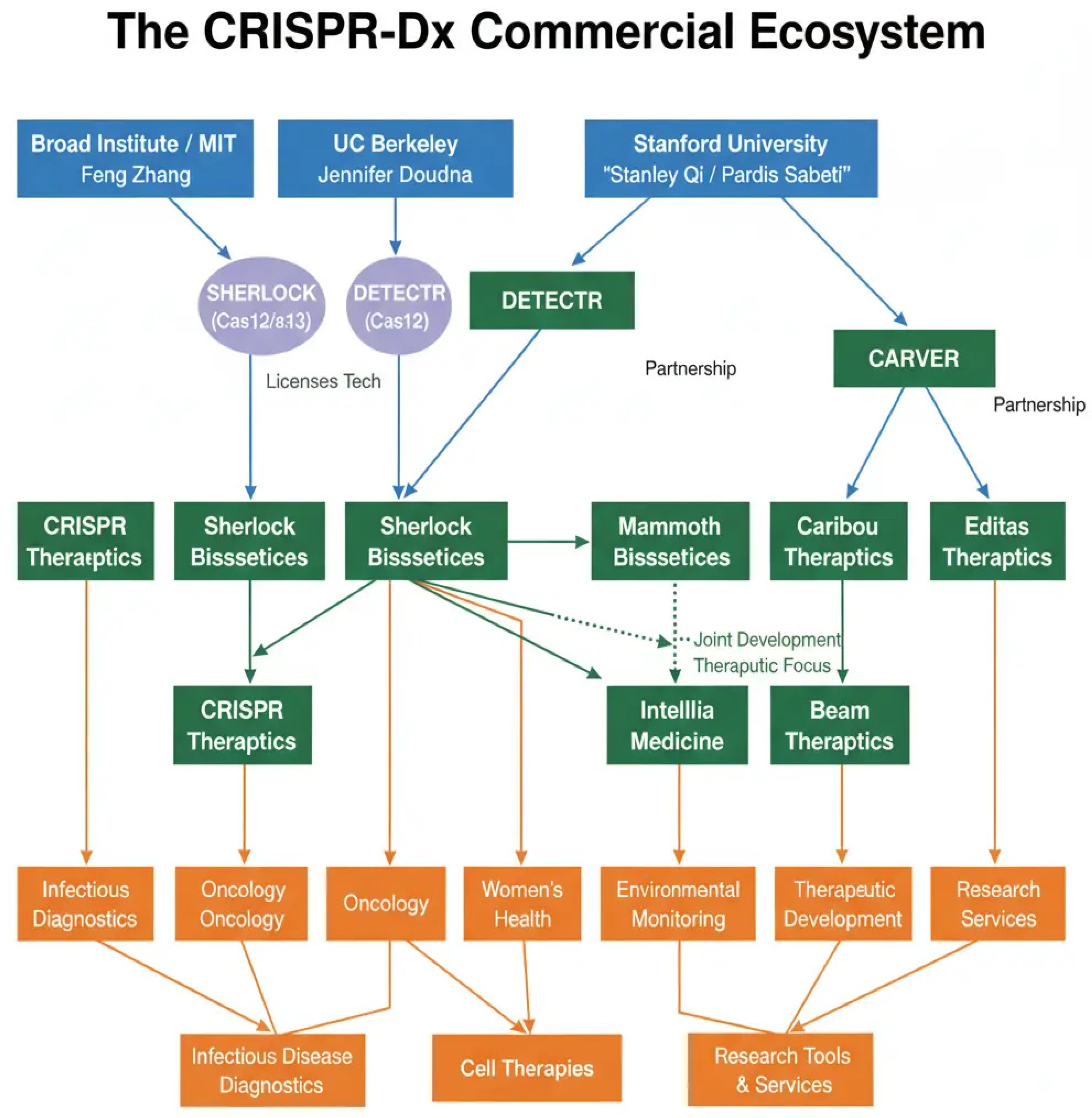
Figure 8.
The commercial ecosystem of CRISPR-based diagnostics. The figure maps the translation of CRISPR technology from foundational academic research to commercial enterprise. Pioneering work at institutions like the Broad Institute of MIT and Harvard, the University of California, Berkeley, and Stanford University led to the creation of spin-out companies such as Sherlock Biosciences, Mammoth Biosciences, and Caribou Biosciences. These companies, along with others, are now at the forefront of developing and scaling FDA-authorized diagnostic tests, driving the technology’s transition from the laboratory to clinical and public health use.
Figure 8.
The commercial ecosystem of CRISPR-based diagnostics. The figure maps the translation of CRISPR technology from foundational academic research to commercial enterprise. Pioneering work at institutions like the Broad Institute of MIT and Harvard, the University of California, Berkeley, and Stanford University led to the creation of spin-out companies such as Sherlock Biosciences, Mammoth Biosciences, and Caribou Biosciences. These companies, along with others, are now at the forefront of developing and scaling FDA-authorized diagnostic tests, driving the technology’s transition from the laboratory to clinical and public health use.
5. Concluding Remarks and Future Perspectives
CRISPR-based diagnostics have transitioned from a theoretical possibility to a tangible reality with remarkable speed. The technology’s inherent programmability, specificity, and compatibility with instrument-free readouts position it as a transformative force in molecular diagnostics, particularly for global health and point-of-care applications. The COVID-19 pandemic served as a powerful catalyst, accelerating innovation and demonstrating the real-world potential of CRISPR-Dx for rapid and decentralized disease surveillance.
The future of CRISPR-Dx is bright, with several exciting avenues of research poised to expand its capabilities. The discovery and characterization of novel Cas enzymes with different properties (e.g., enhanced thermal stability, different PAM requirements, or alternative collateral activities) will continue to broaden the diagnostic toolkit. Advances in protein engineering may allow for the creation of bespoke Cas enzymes with optimized kinetics or enhanced stability, further improving assay performance. The integration of CRISPR-Dx with microfluidics and lab-on-a-chip technologies promises to create fully automated, sample-to-answer devices that require minimal user intervention, bringing us closer to the ideal of a truly equipment-free diagnostic platform.
Furthermore, the application of CRISPR-Dx is expanding beyond infectious diseases to include the detection of cancer-associated mutations in liquid biopsies, the identification of genetic markers for inherited diseases, and the monitoring of environmental and agricultural pathogens [30,31,33,34,35]. As these technologies mature, they will not only empower healthcare providers in remote and low-resource settings but also enable a more proactive and personalized approach to medicine globally.
In conclusion, while significant challenges in sample preparation, multiplexing, and regulatory approval remain, the trajectory of CRISPR-based diagnostics is clear. By continuing to innovate and address these hurdles, the scientific community is poised to deliver on the promise of this revolutionary technology, moving molecular diagnostics from the lab to life and creating a more equitable and resilient global health landscape.
References
- Unemo M, van der Veen S, Rösth O, et al. The challenge of introducing and implementing new and rapid point-of-care tests for sexually transmitted infections. PLoS Med. 2021;18(6):e1003655.
- World Health Organization. (2021). Diagnostic testing for SARS-CoV-2. WHO.
- World Health Organization. (2006). Sexually Transmitted Diseases Diagnostics Initiative (SDI): ASSURED criteria. WHO.
- Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 2012;337(6096):816-821.
- Myhrvold C, Freije CA, Gootenberg JS, et al. Field-deployable viral diagnostics using CRISPR-Cas13. Science. 2018;360(6387):444-448.
- Chen JS, Ma E, Harrington LB, et al. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science. 2018;360(6387):436-439.
- Zetsche B, Gootenberg JS, Abudayyeh OO, et al. Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system. Cell. 2015;163(3):759-771.
- Li SY, Cheng QX, Wang JM, et al. CRISPR-Cas12a-assisted nucleic acid detection. Cell Discov. 2018;4:20.
- Ramachandran A, Huyke DA, Sharma E, Sotomayor M, Santiago JG. Enzyme kinetics of CRISPR molecular diagnostics. Anal Chem. 2021;93(10):4341-4349.
- Gootenberg JS, Abudayyeh OO, Lee JW, et al. Multiplexed and portable nucleic acid detection platform with Cas13, Cas12a, and Csm6. Science. 2018;360(6387):439-444.
- Abudayyeh OO, Gootenberg JS, Konermann S, et al. RNA targeting with CRISPR-Cas13. Nature. 2017;550(7675):280-284.
- Feng W, et al. Signal Amplification by the trans-Cleavage Activity of CRISPR-Cas Systems: Kinetics and Performance. Anal Chem. 2023;95(1):401-409.
- Kaminski MM, Abudayyeh OO, Gootenberg JS, et al. CRISPR-based diagnostics. Nat Biomed Eng. 2021;5(7):643-656.
- Broughton JP, et al. CRISPR–Cas12-based detection of SARS-CoV-2. Nat Biotechnol. 2020;38(7):871-875.
- Cheng M, et al. Chain hybridization-based CRISPR-lateral flow assay for sensitive and specific detection of nucleic acids. Anal Chim Acta. 2023;1277:341778.
- Kaminski MM, Abudayyeh OO, Gootenberg JS, et al. CRISPR-based diagnostics. Nat Biomed Eng. 2021;5(7):643-656.
- Lobato IM, O’Sullivan DM. Recombinase polymerase amplification: Basics and applications. Adv Clin Chem. 2017;80:1-35. https://www.sciencedirect.com/science/article/pii/S006524231730030X.
- Notomi T, et al. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000;28(12):e63.
- Chen L, Hu M, Zhou X. Trends in developing one-pot CRISPR diagnostics strategies. Trends Biotechnol. 2024;42(5):495-507. [CrossRef]
- Broughton JP, et al. CRISPR–Cas12-based detection of SARS-CoV-2. Nat Biotechnol. 2020;38(7):871-875.
- Joung J, et al. Detection of SARS-CoV-2 with SHERLOCK One-Pot Testing. N Engl J Med. 2020;383(15):1492-1494.
- Broughton JP, et al. CRISPR–Cas12-based detection of SARS-CoV-2. Nat Biotechnol. 2020;38(7):871-875. https://www.nature.com/articles/s41587-020-0502-2.
- Joung J, et al. Point-of-care testing for COVID-19 using SHERLOCK diagnostics. Med. 2020;1(1):101-106.
- Qiu X, et al. An extraction-free one-step CRISPR-assisted detection platform and a potential Streptococcus pneumoniae at-home self-testing kit. Biosens Bioelectron. 2023;223:115001.
- Pardee K, et al. Rapid, Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 2016;165(5):1255-1266.
- Zhang Y, et al. A one-pot method for universal Dengue virus detection by combining RT-RPA amplification and CRISPR/Cas12a assay. BMC Microbiol. 2024;24(1):1-13. [CrossRef]
- Li X, et al. Highly sensitive and rapid point-of-care testing for HIV-1 infection based on CRISPR-Cas13a system. BMC Infect Dis. 2023;23(1):577.
- Lee RA, et al. Ultrasensitive CRISPR-based diagnostic for field-applicable detection of Plasmodium species in symptomatic and asymptomatic malaria. Proc Natl Acad Sci U S A. 2020;117(40):24922-24931.
- Ai JW, et al. CRISPR-based rapid and ultra-sensitive diagnostic test for Mycobacterium tuberculosis. Emerg Microbes Infect. 2019;8(1):131-141.
- Di Carlo E, et al. State of the art CRISPR-based strategies for cancer diagnostics and treatment. Biomarker Research. 2024;12(1):1-19.
- Kaminski MM, Abudayyeh OO, Gootenberg JS, et al. CRISPR-based diagnostics. Nat Biomed Eng. 2021;5(7):643-656.
- Tao S, et al. The Application of the CRISPR-Cas System in Antibiotic Resistance. Infect Drug Resist. 2022;15:4591-4602.
- Wani AK, et al. CRISPR/Cas12a-based biosensors for environmental monitoring and diagnostics. Environ Technol Innov. 2024;34:103625.
- Li Y, et al. CRISPR-Cas-based detection for food safety problems: Current status, challenges, and opportunities. Compr Rev Food Sci Food Saf. 2022;21(4):3521-3543.
- Tanny T, et al. CRISPR/Cas-Based Diagnostics in Agricultural Applications. J Agric Food Chem. 2023;71(30):11087-11099.
- Ghouneimy A, et al. CRISPR-Based Diagnostics: Challenges and Potential Solutions Toward Point-of-Care Applications. ACS Synth Biol. 2022;11(1):11-29.
- Ghouneimy A, et al. CRISPR-Based Diagnostics: Challenges and Potential Solutions Toward Point-of-Care Applications. ACS Synth Biol. 2022;11(1):11-29.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



