Submitted:
19 September 2025
Posted:
22 September 2025
You are already at the latest version
Abstract
Mosquito-borne diseases such as malaria, dengue, chikungunya, and filariasis continue to pose a serious public health challenge in India. Vector control remains central to managing these diseases, with insecticides forming the backbone of control programs. However, the effectiveness of these chemicals has declined as mosquito populations have developed resistance, while drug resistance has also emerged in pathogens. Resistance in mosquitoes is driven by multiple mechanisms, including enhanced detoxification processes and mutations that reduce insecticide sensitivity. This review traces the history of insecticide use in India, focusing on the five major classes - organochlorines, organophosphates, pyrethroids, carbamates, and neonicotinoids. Prolonged use has led to widespread resistance in key species such as Anopheles culicifacies, An. stephensi, Culex quinquefasciatus, and Aedes aegypti. The review highlights target-site mutations (such as kdr L1014F/S, F1534C), metabolic resistance (CYP450s, GSTs, esterases), behavioral changes, and cuticular thickening. Resistance patterns vary across regions and often involve multiple classes of insecticides. Recent molecular findings offer important insights for surveillance and provide direction for integrated and sustainable resistance management strategies in India.
Keywords:
mosquito vectors
; mosquito-borne disease
; indoor residual spraying
; insecticide resistance
; knockdown resistance (kdr)
; metabolic detoxification
Introduction
Vector-borne diseases continue to be a major public health concern in India, contributing significantly to morbidity and mortality. Mosquitoes belonging to the groups Anopheles, Aedes, and Culex are the main carriers that transmit malaria, dengue, chikungunya, Zika virus, and lymphatic filariasis. (Raghavendra et al., 2011). For decades, vector control in India has relied heavily on chemical insecticides, particularly organochlorines, organophosphates, carbamates, and pyrethroids. However, the extensive and often indiscriminate use of these compounds has led to the emergence and spread of insecticide resistance, posing a significant challenge to vector management and disease control programs (Hemingway et al., 2016).
Insecticide resistance refers to the heritable decrease in the sensitivity of mosquito populations to chemicals that were previously effective at controlling them. This is an evolutionary response facilitated by the selection pressure exerted by sustained exposure to insecticides. Resistance mechanisms are broadly categorized into four types: metabolic resistance (enhanced detoxification), target site insensitivity (mutations in genes such as kdr for pyrethroids), behavioral resistance, and cuticular resistance (Liu, 2015).
In India, resistance has been widely reported in vector mosquitoes in multiple states. For example, An. culicifacies, the major malaria vector in rural India, has shown resistance to DDT and pyrethroids in various regions (Raghavendra et al., 2011). Aedes aegypti, the main mosquito responsible for spreading dengue and chikungunya, has developed resistance to temephos and pyrethroids, raising concerns regarding the efficacy of current control strategies (Kushwah et al., 2015). Similarly, Cx. quinquefasciatus, the vector of filariasis, has exhibited widespread resistance to organophosphates and pyrethroids, complicating efforts to eliminate lymphatic filariasis (Thomas et al., 2013).
The challenge of managing insecticide resistance is compounded by the lack of alternative tools and the limited development of new insecticidal compounds. The World Health Organization’s Global Plan for Insecticide Resistance Management (GPIRM) has emphasized the need for integrated vector management (IVM), rotation of insecticides, and the use of combination interventions to delay the evolution of resistance (WHO, 2020). In India, the National Centre for Vector Borne Diseases Control (NCVBDC) has initiated periodic resistance surveillance to inform and optimize vector-control policies.
This literature review aims to synthesize the knowledge on the status, mechanisms, and patterns of insecticide resistance in Indian vector mosquitoes.
Historical Overview of Insecticide Use for Vector Control in India
Since the mid-20th century, insecticides have been a cornerstone of vector control strategies aimed at managing vector-borne diseases. The introduction of synthetic insecticides has revolutionized vector control programs by providing potent and relatively affordable tools for suppressing vector populations. Based on their chemical composition, insecticides are generally categorized into five major groups. a) Organochlorines, b) Organophosphates, c) Pyrethroids, d) Carbamates, e) Neonicotinoids (Nayak and Solanki, 2021).
Organochlorines: In India, commonly used organochlorine insecticides for vector control include DDT, BHC (banned since 1997), and dieldrin (banned since 2001) (NIMR, 2006). Among these, DDT is the principal agent in the fight against malaria. Owing to its excito-repellent properties, DDT not only kills mosquitoes but also drives them away upon contact (NCVBDC, 2016).
DDT was first introduced in India in 1944 by American and British soldiers in Odisha and Karnataka. In 1953, the Government of India mandated the use of DDT in Indoor Residual Spraying (IRS) at a dosage of 1 g/m² under the National Malaria Control Programme (NMCP). At the time, malaria accounted for approximately 75 million cases and 800,000 deaths annually, but by 1960, cases had dropped to 108394 with zero fatalities. Despite its initial effectiveness, malaria resurgence, particularly due to Plasmodium falciparum, was observed in the 1990s (Sharma, 2003; Betne and Rajankar, 2011). Between 2000 and 2008, although nearly 6 million kilograms of DDT were distributed annually across Indian states, the malaria incidence plateaued, indicating the emergence of insecticide resistance among vectors (Betne and Rajankar, 2011).
Currently, India has significantly reduced DDT usage. It is restricted to a maximum of 10,000 metric tons annually and is employed only in public health programs, particularly during outbreaks or epidemics. According to the WHO recommendations, DDT use is strictly limited to Indoor Residual Spraying (IRS) (NCVBDC, 2016). Notably, DDT has been banned for agricultural purposes since 1989 (Betne and Rajankar, 2011).
Organophosphates: Common organophosphate insecticides used for vector control in India include Malathion, Fenthion, Fenitrothion, Pirimiphos-methyl, and Temephos (NIMR, 2006). Malathion is primarily used for mosquito and fruit fly control.Temephos (1%) granules are widely used to manage dengue and chikungunya vectors. Its 50% emulsifiable concentrate formulation is noted for its low toxicity to mammals and non-lethality to aquatic organisms, such as fish (NCVBDC, 2016).
Organophosphates were first introduced in India in the 1960s (Kumar et al., 2016). In 1969, malathion was deployed in regions where mosquitoes had developed resistance to DDT (WHO, 2015). However, the current use of organophosphates remains limited because of their toxicological risks and the development of resistance among vector species (Kumar et al., 2016).
Pyrethroids: Synthetic pyrethroids are among the newer insecticides used for vector control in India. The Central Insecticide Board has recommended deltamethrin (2.5% WP), cyfluthrin (10% WP), alphacypermethrin (5% WP), lambdacyhalothrin (10% WP), and bifenthrin (10% WP) as adulticides (NCVBDC, 2016). The use of pyrethroids in India commenced in the 1980s (Kumar et al., 2016). These compounds are favored for Long-Lasting Insecticidal Nets (LLINs) and IRS due to their rapid knockdown effect and low toxicity to mammals. However, widespread resistance has become a significant concern (Hemingway et al., 2016; WHO, 2015).
Carbamates: The most commonly used carbamate insecticide for vector control in India is propoxur. It was initially introduced by the Union Carbide Corporation in 1956 for agricultural use (Paul, 2018), and its application in mosquito control began in the 1970s (Kumar et al., 2016), particularly in areas with resistance to organochlorines such as DDT. Resistance to propoxur has been reported in major vector species, such as Cx. quinquefasciatus (vector of filariasis) (Shetty et al., 2013) and An. stephensi (a key malaria vector) (Mukhopadhyay et al., 1996).
Neonicotinoids: Neonicotinoids are being explored as novel alternatives to conventional insecticides for vector control. Compounds such as dinotefuran, thiamethoxam, and imidacloprid exhibit a high binding affinity for nicotinic acetylcholine receptors in mosquitoes, making them promising candidates for effective vector management (Natarajan et al., 2014). Due to increasing resistance, a neonicotinoid called clothianidin is being explored for IRS as a pyrethroid replacement (Kumar et al., 2024).
Common insecticides and their recommended dosages are presented in Table 1.
Insecticide Resistance Among Mosquito Vectors in India
Insecticide resistance is the heritable reduction in sensitivity to an insecticide in a pest population, such that normal dosages fail to achieve expected control. It arises through genetic changes (e.g., mutations, enhanced detoxification) driven by repeated exposure. Resistance can be specific, cross, or multiple, depending on mechanisms and insecticide classes (IRAC, 2023).
Anopheles culicifacies, the primary malaria vector in India, has shown widespread resistance. Recent studies reported a 39% mortality rate and confirmed resistance to DDT (4%) across all surveyed regions. Mortality rates generally range between 49% and 78%, with resistance also observed to deltamethrin (0.05%) and malathion (5%). Notably, An. culicifacies populations are triple-resistant (to DDT, malathion, and deltamethrin) in nine Indian states (Rahi et al., 2024).
Anopheles stephensi, another major malaria vector, exhibits resistance to DDT and malathion in seven regions and to deltamethrin in one area (Rahi et al., 2024). Anopheles fluviatilis shows variable susceptibility patterns, with resistance differing across several states in India (Rahi et al., 2024).
Aedes aegypti and Aedes albopictus, important vectors of dengue, chikungunya, and Zika, are generally resistant to DDT but remain susceptible to certain pyrethroids (Kumar et al., 2024).
Culex mosquitoes, including Cx. quinquefasciatus, have been reported to be resistant to multiple classes of insecticides, including pyrethroids, carbamates, DDT, and organophosphates (Bharadwaj et al., 2025).
Historical Perspective: India has one of the earliest records of mosquito insecticide resistance. Cx. quinquefasciatus (formerly Cx. fatigans) was the first species reported resistant to DDT in 1952 in Bombay and Uttar Pradesh (Mehrotra, 1989; Sharma and Moin, 2021). An. stephensi from Tamil Nadu was reported DDT-resistant in 1956, followed by An. culicifacies in Gujarat in 1957. In 1964, An. fluviatilis populations from Mumbai and Mysore also developed DDT resistance. Among Aedes species, A. aegypti in Calcutta showed DDT resistance in 1963, while A. vittatus in Baroda and A. albopictus in Lucknow demonstrated resistance in 1964 and 1965, respectively. Resistance of Aedes mosquitoes to BHC and DDT is now well documented (Mehrotra, 1989).
Mechanisms of Insecticide Resistance in Indian Vector Mosquitoes
Insecticide resistance in vector mosquitoes (e.g. Anopheles, Aedes, Culex) arises through biochemical, genetic, and behavioral adaptations that allow them to survive exposure, undermining control efforts. The major mechanisms are target-site resistance, metabolic resistance, behavioral resistance, and cuticular resistance (Elanga Ndille et al., 2023).
Target Site Resistance
Structural alterations or point mutations in genes encoding target proteins that interact with insecticides cause target-site resistance. Pyrethroids and dichlorodiphenyltrichloroethane (DDT) are insecticides that target sodium channels. Both organophosphate and carbamate insecticides target acetylcholinesterase (AChE). γ-Aminobutyric acid (GABA) receptors are the targets of insecticides such as cyclodiene and fipronil (Liu, 2015).
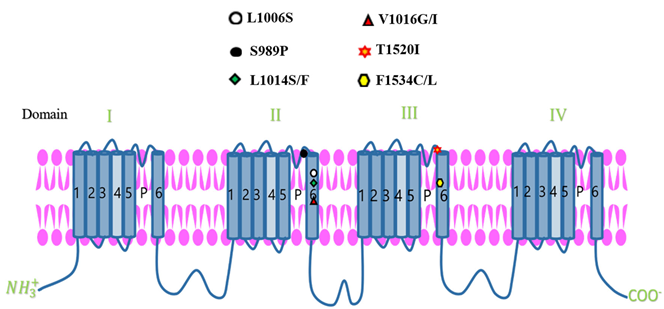
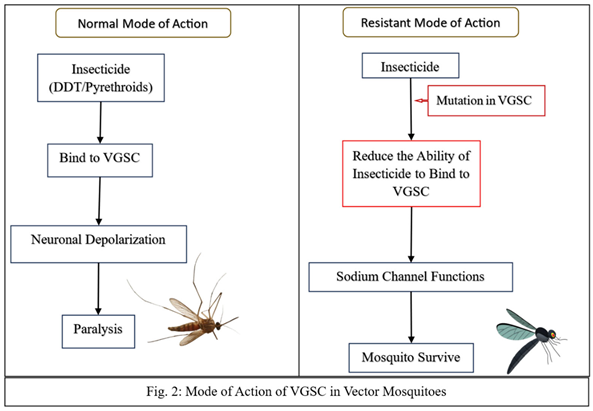
Voltage-gated sodium channel (VGSC): Mutations in the voltage-gated sodium channel (VGSC), commonly referred to as knockdown resistance (kdr), are a well-established mechanism of insecticide resistance in mosquitoes. These mutations, mainly concentrated in the IS6, IIS6, and IIIS6 domains of the VGSC, reduce sensitivity to pyrethroids and DDT. The most critical substitution occurs at codon 1014 in domain IIS6, where leucine (L) is replaced by phenylalanine (F), serine (S), or histidine (H), conferring resistance (Gan et al., 2021).
In Anopheles species, kdr mutations have been widely documented. An. culicifacies populations carry L1014F and L1014S mutations, which are associated with resistance to pyrethroids and DDT (Singh et al., 2009; Dykes et al., 2015). An. stephensi populations in northern India carry both L1014F and L1014S mutations (Dykes et al., 2016), and An. subpictus has been found with the L1014F mutation, exhibiting notable geographic variation in its prevalence (Sindhania et al., 2023).
In Aedes species, several distinct kdr mutations have been identified. In A. albopictus, the T1520I and F1534C mutations in the VGSC gene are linked to resistance to DDT and pyrethroids, with F1534C first reported in northern West Bengal (Modak et al., 2022). A. aegypti frequently displays kdr mutations such as S989P, V1016G, T1520I, F1534C, and F1534L, which collectively reduce susceptibility to these insecticides (Kumawat et al., 2021). The V1016G + F1534C double mutation in A. aegypti is associated with high resistance and genetic polymorphism in West Bengal (Saha et al., 2019). The Delhi population of A. aegypti has exhibited resistance to DDT, deltamethrin, and permethrin, with both F1534C and a novel T1520I mutation detected (Kushwah et al., 2015). Further studies from Punjab have identified V1016G and L1006S mutations in A. aegypti (Kaura et al., 2022).
In Cx. quinquefasciatus populations from West Bengal, both L1014F and L1014S mutations in the VGSC gene have been detected, conferring resistance to DDT and pyrethroids. Notably, L1014S was reported for the first time in this species from the region (Rai & Saha, 2022; retracted 2023). The L1014F mutation was also confirmed in Cx. quinquefasciatus from the sub-Himalayan areas of West Bengal (Modak et al., 2024).
Kdr mutations in the VGSC gene (Figure 1) are widespread in many mosquito species and reduce their sensitivity to insecticides such as pyrethroids and DDT. A word diagram (Figure 2) is provided to explain how these mutations affect the VGSC.
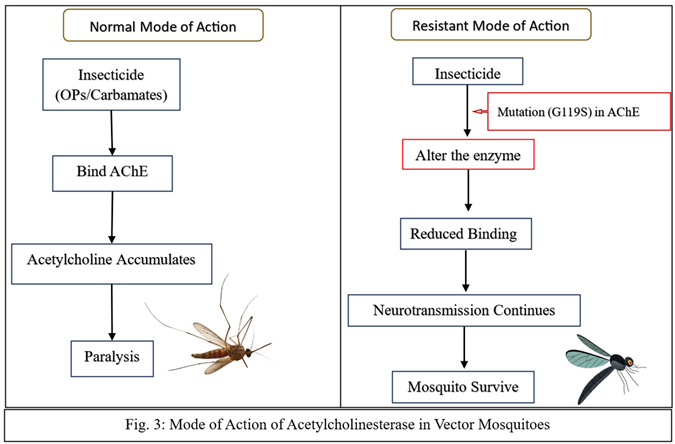
Acetylcholinesterase (AChE): Carbamate and organophosphate pesticides primarily target acetylcholinesterase (AChE), preventing nerve transmission at cholinergic synapses. Resistance arises from amino acid substitutions in the ace-1/ace-2 gene, rendering AChE insensitive to inhibition (Gan et al., 2021). Numerous mosquito species, including An. gambiae, An. albimanus, Cx. vishnui, Cx. pipiens, and Cx. quinquefasciatus, have been found to exhibit the G119S substitution (Liu, 2015).
Acetylcholinesterase (ace-1) mutations have been documented in several Indian vector mosquitoes and are associated with resistance to organophosphates and carbamates. In Culex quinquefasciatus, the G119S mutation in the ace-1 gene has been reported from eastern Uttar Pradesh, where it was linked to malathion resistance. The same study also identified the F331W substitution in Culex tritaeniorhynchus, along with evidence of altered ace gene activity that indicated an insensitive AChE mechanism (Misra and Gore 2015). In Aedes aegypti, populations from Tamil Nadu (Namakkal district) were found to carry the G119S mutation at a frequency of about 0.24. This mutation, together with elevated esterases, contributed to larval resistance against the organophosphate temephos (Muthusamy and Shivakumar 2015). These findings confirm that ace-1 mutations play a significant role in insecticide resistance in Indian mosquito vectors, although their prevalence appears to vary across species and regions. A word diagram (Figure 3) illustrating the mode of action of Acetylcholinesterase is shown below.
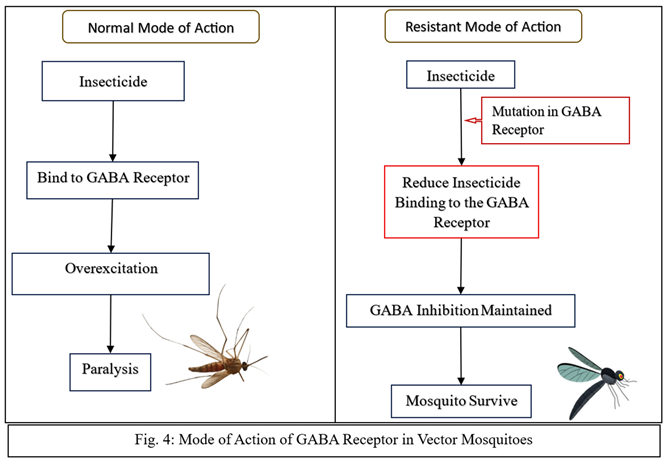
GABA Receptor (Rdl Gene): Neuronal signalling involves the resistance to dieldrin (RDL) gene, which encodes the GABA receptor. The rdl receptor belongs to the Cys-loop ligand-gated ion channel superfamily and has an N-terminal extracellular domain for GABA binding. Each of the five subunits of this receptor has four transmembrane domains and an external cysteine loop (M1 - M4). Numerous pesticides, including cyclodiene, fipronil, and pyrethroids, target rdl, and their effects are influenced by post-translational changes. GABA receptors, integral chloride channels targeted by cyclodiene insecticides (dieldrin) and phenyl pyrazoles (fipronil), display mutations such as A296S/G that confer resistance (Liu, 2015). There is no specific evidence confirming the presence of these mutations in Indian vectors like Anopheles stephensi, Aedes aegypti, or Culex quinquefasciatus. The mode of action of the GABA receptor is shown in the word diagram (Figure 4) below.
Below is a list of all target site mutations reported in Indian mosquitoes.

Table 5.
Major target-site mutations observed in different mosquito vectors in India.
| Mosquito species | Mutation | Transmembrane Domain / gene | Reference(s) |
| A. Aegypti | V1016G, S989P, L1006S | ΙΙ | Kumawat et al., 2021; Saha et al., 2019; Kaura et al., 2022 |
| F1534C, F1534L, T1520I, | ΙΙΙ | Kumawat et al., 2021; Kushwah et al., 2015; Saha et al., 2019 |
|
| G119S | Ace-1 | Muthusamy and Shivakumar, 2015 | |
| A. albopictus | T1520I, F1534C | ΙΙΙ | Modak et al., 2022 |
| An. culicifacies | L1014F, L1014S | ΙΙ | Singh et al., 2009; Dykes et al., 2015 |
| An. subpictus | L1014F | ΙΙ | Sindhania et al., 2023 |
| An. stephensi | L1014F, L1014S | ΙΙ | Dykes et al., 2016 |
| Cx. quinquefasciatus | G119S | Ace-1 | Misra and Gore 2015 |
| L1014F, L1014S | ΙΙ | Rai & Saha, 2022; Modak et al., 2022 | |
| Culex tritaeniorhynchus | F331W | Ace-1 | Misra and Gore 2015 |
Metabolic Resistance
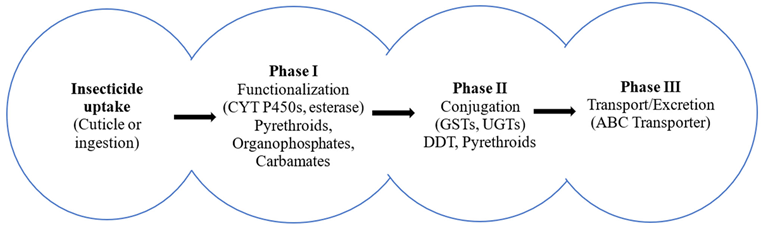
As a result of point mutations in the cis/trans loci of the enzymes, metabolically resistant strains detoxify toxins or insecticides by overexpressing the enzymes or changing their conformation. Three main enzymatic processes are typically linked to metabolic detoxification: cytochrome P450 monooxygenases (P450s), carboxylesterases and glutathione S-transferases (GSTs) (Gan et al., 2021). Insecticide detoxification in mosquitoes involves a multi-step biochemical process (Figure 5). These are-
- Uptake: Insecticides penetrate the cuticle or are ingested.
- Phase I (functionalization): Oxidation, reduction, or hydrolysis introduces or exposes polar groups to the substrate. The key actors are cytochrome P450 monooxygenases (P450s) and carboxyl/cholinesterases (esterases) (David et al., 2013).
- Phase II (conjugation): Conjugating enzymes (notably glutathione S-transferases, GSTs) attach polar groups (e.g., glutathione) to Phase I products, increasing their solubility (Ranson and Hemingway, 2005).
- Phase III (transport/excretion): Transporter proteins (ATP-binding cassette (ABC) transporters and other efflux systems) move metabolites out of cells and across barriers for excretion (Dermauw and Van Leeuwen, 2014).
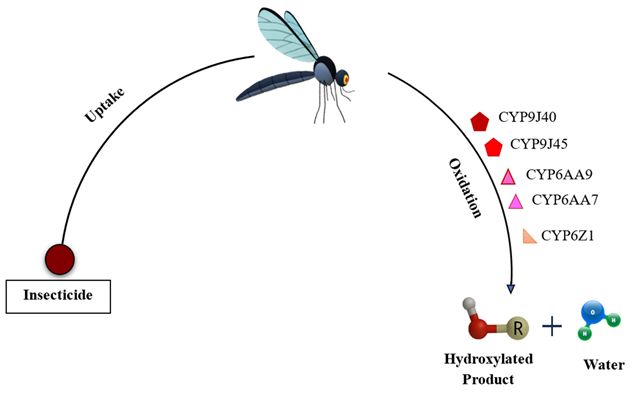
Cytochrome P450 Monooxygenases (CytP450s): The overexpression of cytochrome P450 (CYP) genes, particularly those belonging to the CYP6 and CYP9 families, is a well-established marker of metabolic resistance in mosquitoes (Bharadwaj et al., 2025). CYP450 enzymes play a crucial role in broad-spectrum detoxification, including the metabolism of pyrethroids and carbamates (Ballav et al., 2022).
In Indian vector mosquitoes, cytochrome P450s (CYPs) are consistently implicated in insecticide resistance, particularly against pyrethroids and organochlorines. In Anopheles culicifacies, metabolic resistance has been linked to the strong overexpression of CYP6Z1, which is associated with resistance to deltamethrin (Kareemi et al., 2022).
In Culex quinquefasciatus, permethrin-resistant populations from India showed significant upregulation of multiple CYP genes. Notably, CYP6AA7 was upregulated about 10-fold in both larvae and adults, CYP9J34 nearly 9-fold, and CYP6Z2 approximately 5-fold compared to susceptible strains (Ramkumar et al., 2023).
Field studies from northern West Bengal further confirmed these trends. Wild Culex quinquefasciatus larval populations from the sub-Himalayan region exhibited widespread resistance to cyphenothrin. Biochemical assays revealed high monooxygenase activity, while gene expression analysis showed elevated levels of CYP6AA7 and CYP9J40 (Saha et al., 2025). P450-mediated resistance has been documented against a number of pesticide classes, including as organophosphates and pyrethroids. Below is a general mechanism of this resistance (Figure 6).
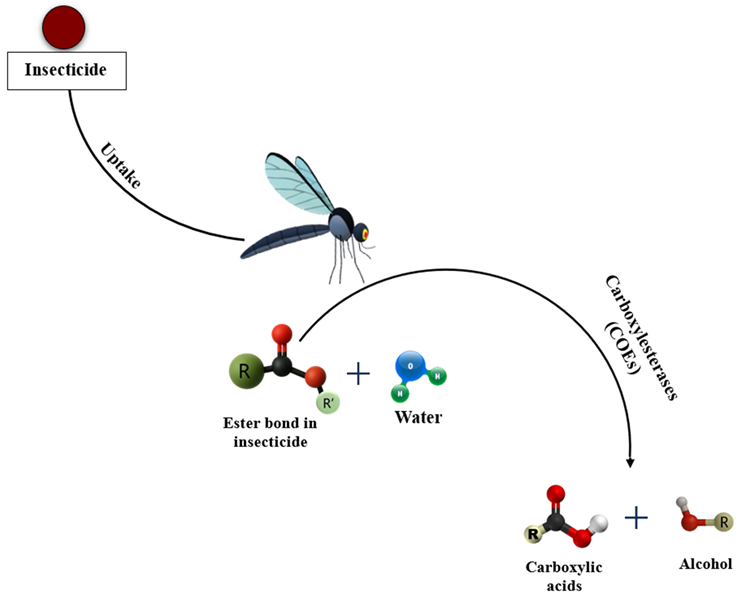
Carboxylesterases (esterases): Enzymes that hydrolyse ester bonds, such as esterases, contribute to insecticide resistance through gene duplication and overexpression, thereby amplifying detoxification capacity (Liu, 2015).
Esterase activity has been widely implicated in insecticide resistance among Indian vector mosquitoes. In Culex quinquefasciatus from the sub-Himalayan districts of West Bengal, wild larval populations resistant to cyphenothrin and temephos demonstrated high levels of carboxylesterase activity, with gene expression analysis confirming the overexpression of esterase genes (Saha et al., 2025).
In Aedes aegypti, α- and β-esterase activities were reported to be 1.2–3.1-fold and 2.0–23.0-fold higher, respectively, in field populations from the Dooars and Terai regions of West Bengal compared with a susceptible laboratory strain. Isozyme profiles further revealed notable population variation (Bharati et al., 2018). Similarly, in Pondicherry, A. aegypti collected from Lawspet showed elevated β-esterase activity associated with malathion resistance, likely driven by repeated thermal fogging, whereas populations from Abishegapakkam displayed lower activity (Ramesh et al., 2023).
More than 50% and up to 80% of all four Japanese encephalitis (JE) vector species (Cx. tritaeniorhynchus, Cx. vishnui, Cx. pseudovishnui, and Cx. gelidus) showed elevated levels of both α- and β-esterases (Ballav et al., 2022). Furthermore, A. aegypti populations in West Bengal and Assam have demonstrated esterase-mediated resistance (Kumawat et al., 2021).
Comparative studies in Mysore highlighted species-specific trends: An. stephensi exhibited higher α-esterase activity, while An. culicifacies showed elevated β-esterase levels, correlating with differences in tolerance to deltamethrin and permethrin (Ganesh et al., 2003). Consequently, the reduced efficacy of the insecticide enables insect survival. The general pathway of this resistance is presented below (Figure 7).
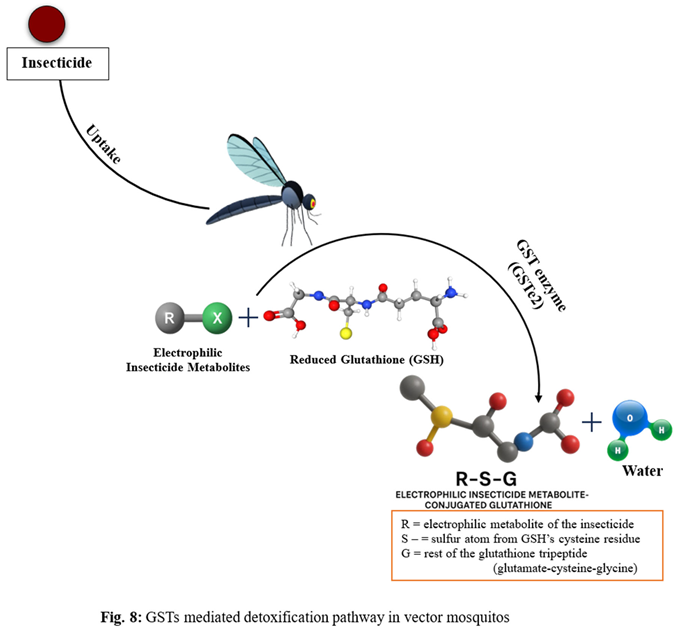
Glutathione-S-transferases (GSTs): Enzymes that conjugate reactive intermediates and are recognized mechanisms of DDT resistance that contribute to pyrethroid metabolism (Dykes et al., 2022). Glutathione S-transferases (GSTs) have been repeatedly implicated in metabolic insecticide resistance across Indian vector mosquitoes, with biochemical, molecular and genomic evidence reported from multiple species and regions. A study showed significantly elevated GST activity in DDT-resistant Anopheles culicifacies and An. annularis from Malkangiri and Koraput (Odisha), with median GST activities in resistant populations roughly three times those of a susceptible An. fluviatilis comparator, supporting a GST-mediated DDT detoxification mechanism (Gunasekaran et al., 2011). More recently, molecular and genomic work has directly tied specific GST genes to resistance in Indian An. Stephensi, documented a tandem duplication of a genomic region containing GSTe2 and GSTe4 in a laboratory-colonized DDT-resistant strain derived from Alwar, India, a structural change likely to increase GST gene dose and enzyme levels and to contribute to DDT resistance (Dykes et al., 2022). Transcriptomic and resistance-mechanism studies in Indian An. culicifacies populations have also noted co-upregulation of GSTe2 alongside cytochrome P450s in pyrethroid- and DDT-resistant samples, further implicating GST epsilon class enzymes in field resistance (Kareemi et al., 2022). Approximately 70–95% of Culex mosquitoes across all species exhibited Glutathione S-transferase (GST) activity levels above the threshold, with significant variation observed across sites in northern West Bengal, indicating a strong association with DDT resistance (Bharati et al., 2018). Similarly, Aedes aegypti populations from Maharashtra displayed elevated GST activity, which has been linked to organochlorine resistance (Kumawat et al., 2021). Below is a general mechanism of this resistance (Figure 8).
Transport Protein: Transporter proteins, mainly ATP-binding cassette (ABC) family, are membrane pumps that expel insecticide metabolites during Phase III detoxification, contributing to metabolic resistance in mosquitoes
Studies on Anopheles mosquitoes have shown that ABC transporters play a role in permethrin tolerance in these species. In Anopheles stephensi larvae, mortality increased when permethrin was combined with the ABC inhibitor verapamil, and the ABCG4 transporter gene was upregulated following exposure (Epis et al., 2014). In adults of the same species, several ABC genes, including ABCB2, ABCB member 6, and ABCG4, were upregulated in both males and females after permethrin treatment (Adedeji et al., 2020). Similarly, in Anopheles gambiae larvae, exposure to permethrin in combination with an ABC inhibitor resulted in approximately a 15-fold increase in mortality, further confirming the involvement of these transporters in detoxification (Mastrantonio et al., 2019).
The table displays the various metabolic resistance categories found in Indian vector mosquitoes by species.
Table 6.
Major Metabolic Resistance observed in different mosquito vectors in India.
| Vector species | Metabolic mechanisms | Insecticide affected | Ref. |
| Anopheles stephensi | GSTe2 GSTe4 duplication, P450 overexpression, esterase | DDT, Pyrethroids |
Dykes et al., 2022 |
| Anopheles culicifacies | P450s (CYP6Z1), esterases, GSTs (GSTe2) | DDT, Pyrethroids, Organophosphates | Sahu et al., 2015; Kareemi et al., 2022 |
| Culex quinquefasciatus | P450s (CYP6/9 families), esterases | Pyrethroids, Organophosphates, Carbamates | Ramkumar et al., 2023 |
| Cx. pipiens | P450s (CYP6AA9) | Bharadwaj et al., 2025 |
Behavioral Resistance
Recent entomological investigations have revealed important insights into the resting behavior, host preference, seasonal abundance, and insecticide response of major mosquito vector species in India.
Abdominal Condition and Resting Behavior: Abdominal condition analysis revealed that more than 60% of Anopheles culicifacies, An. stephensi, and An. subpictus were semi-gravid or gravid, suggesting a strong endophilic resting behavior, as these mosquitoes were primarily found resting indoors (Kumar et al., 2024).
Blood Meal Analysis and Host Preference: Blood meal source identification using multiplex PCR indicated that An. stephensi exhibited a Human Blood Index (HBI) of 9.09%, with 21.81% of specimens also testing positive for bovine blood, highlighting its zoophilic tendency. In contrast, An. culicifacies had a lower HBI of 6.66% and showed no evidence of bovine feeding, suggesting a more anthropophilic feeding behavior (Kumar et al., 2024).
Seasonal Abundance Patterns: Seasonal distribution analysis showed that An. subpictus populations peaked during the monsoon season, whereas An. culicifacies was more prevalent in the post-monsoon and winter periods, indicating species-specific seasonal preferences (Kumar et al., 2024).
Excito-repellency and Delayed Mortality: Exposure to synthetic pyrethroids and organophosphates in formulated insecticides induced strong excito-repellency responses in Aedes aegypti, An. stephensi and Culex quinquefasciatus, in addition to causing delayed mortality, suggesting behavioral avoidance mechanisms alongside toxic effects (Dhiman et al., 2021).
Cuticular Resistance
Cuticular resistance refers to the structural and biochemical modifications in the insect cuticle that reduce insecticide penetration. A primary mechanism involves cuticle thickening, often attributed to increased deposition of cuticular hydrocarbons (CHCs) and chitin. This phenomenon has been observed in resistant populations of Anopheles gambiae, Culex pipiens pallens, and Bactrocera dorsalis. The overexpression of cytochrome P450 genes, particularly CYP4G16 and CYP4G17, in oenocytes enhances hydrocarbon biosynthesis, whereas the upregulation of cuticular protein genes, such as CPLCG3 and CPR127, contributes to the structural thickening of all cuticular layers (Balabanidou et al., 2018).
In India, Aedes aegypti mosquitoes selected for resistance to deltamethrin (a pyrethroid) and acetamiprid (a neonicotinoid) exhibited significantly thicker cuticles than susceptible strains, further supporting the role of cuticular thickening in resistance mechanisms (Samal et al., 2021).
Alterations in cuticle composition also contribute to this resistance. Two major biochemical pathways are involved: (1) the overexpression of laccase 2, which enhances cuticle sclerotization and hardness, and (2) the upregulation of ATP-Binding Cassette (ABC) transporters, which facilitate the transport of CHCs and lipids to the cuticle surface, thereby increasing cuticular impermeability. ABC transporters, particularly those of subfamily G, have been found to be enriched in the legs of resistant mosquitoes and are implicated in lipid export from epidermal cells (Balabanidou et al., 2018).
Conclusions
The historical and current use of insecticides in India for vector control highlights a complex trajectory shaped by efficacy, resistance evolution, and public health requirements. Initially, organochlorines such as DDT were instrumental in reducing malaria prevalence during the 1950s. However, over time, the efficacy of DDT and similar compounds has declined due to widespread resistance, particularly in species such as An. culicifacies, A. aegypti, and Cx. quinquefasciatus.
India has incorporated other chemical groups, such as organophosphates (e.g., malathion and temephos), synthetic pyrethroids (e.g., deltamethrin and permethrin), carbamates (e.g., propoxur and bendiocarb), and more recently, neonicotinoids. Despite the rotation and combination of these insecticide classes, resistance has become increasingly prevalent in recent years. For example, triple resistance (to DDT, malathion, and pyrethroids) is now observed in several states for major vector species.
Molecular analyses have revealed numerous genetic mutations that confer resistance. Notably, kdr (knockdown resistance) mutations, such as L1014F/S, in An. and Cx. and F1534C, V1016G, and S989P in A. aegypti interferes with the voltage-gated sodium channel targets of DDT and pyrethroids. In addition, resistance to organophosphates and carbamates is associated with mutations in acetylcholinesterase (e.g., G119S), whereas resistance to cyclodienes is linked to mutations in the GABA receptor (e.g., A296G/S).
Metabolic resistance is also widespread and is characterized by the elevated activity of detoxification enzymes, such as cytochrome P450 monooxygenases (e.g., CYP6Z1 and CYP6M2), glutathione S-transferases (e.g., GSTe2), and carboxylesterases. These mechanisms are particularly pronounced in Cx. populations inhabiting polluted environments, and A. aegypti from urban centers.
Behavioral and cuticular resistance further complicate pest control efforts. Excito-repellency and altered host-feeding behaviors reduce insecticide contact, whereas cuticle thickening and altered hydrocarbon composition hinder chemical penetration. These adaptations diminish the overall effectiveness of insecticide-based interventions, such as indoor residual spraying (IRS) and long-lasting insecticidal nets (LLINs).
In conclusion, although insecticides have historically played a critical role in vector control in India, their long-term utility is threatened by the rapid evolution of resistance across all major mosquito vectors. A multifaceted, evidence-based, and adaptive approach is essential to sustain gains in vector-borne disease control.
References
- Adedeji, E. O., Ogunlana, O. O., Fatumo, S., Beder, T., Ajamma, Y., Koenig, R., and Adebiyi, E. (2020). Anopheles metabolic proteins in malaria transmission, prevention and control: a review. Parasites and vectors, 13(1), 465. [CrossRef]
- Baig, M. M., Panigrahi, D. K., Patnaik, M. K., Acharya, P., Choudhury, D., Vijayakumar, B., Sahu, S. S., and Kumar, A. (2021). Susceptibility status of A. aegypti (Linnaeus) and A. albopictus (Skuse) (Diptera: Culicidae) to insecticides in southern Odisha, India. International Journal of Mosquito Research, 8(3), 10–15.
- Balabanidou, V., Grigoraki, L., and Vontas, J. (2018). Insect cuticle: A critical determinant of insecticide resistance. Current Opinion in Insect Science, 27, 68–74. [CrossRef]
- Ballav, S., Chatterjee, M., Sardar, A. A., et al. (2022). Biochemical characterization of insecticide resistance in field population of major JE vectors from northern districts of West Bengal, India. International Journal of Tropical Insect Science, 42, 661–675. [CrossRef]
- Betne, R., and Rajankar, P. (2011, May). Factsheet 38: DDT losing grounds in India – Alternatives imminent. Toxics Link. https://toxicslink.org/wp-content/uploads/2022/08/Factsheet_38_DDT.pdf.
- Bharadwaj, N., Sharma, R., Subramanian, M., Ragini, G., Nagarajan, S. A., and Rahi, M. (2025). Omics approaches in understanding insecticide resistance in mosquito vectors. International Journal of Molecular Sciences, 26(5), 1854. [CrossRef]
- Bharati, M., Saha, P., and Saha, D. (2018). Variation in esterase activity among different A. aegypti L. populations from the Dooars and Terai regions of West Bengal, India. Proceedings of the Zoological Society, 71, 239–247. [CrossRef]
- David, J. P., Ismail, H. M., Chandor-Proust, A., and Paine, M. J. (2013). Role of cytochrome P450s in insecticide resistance: impact on the control of mosquito-borne diseases and use of insecticides on Earth. Philosophical transactions of the Royal Society of London. Series B, Biological sciences, 368(1612), 20120429. [CrossRef]
- Dermauw, W., and Van Leeuwen, T. (2014). The ABC gene family in arthropods: comparative genomics and role in insecticide transport and resistance. Insect biochemistry and molecular biology, 45, 89–110. [CrossRef]
- Dhiman, S., Rabha, B., Talukdar, P. K., Das, N. G., Yadav, K., Baruah, I., Singh, L., and Veer, V. (2013). DDT and deltamethrin resistance status of known Japanese encephalitis vectors in Assam, India. The Indian Journal of Medical Research, 138(6), 988–994.
- Dhiman, S., Yadav, K., Acharya, B. N., et al. (2021). Behavioural response of mosquito vectors A. aegypti, An. stephensi and Cx. quinquefasciatus to synthetic pyrethroid and organophosphorus-based slow-release insecticidal paint. Parasites and Vectors, 14, 259. [CrossRef]
- Dykes, C. L., Das, M. K., Eapen, A., Batra, C. P., Ghosh, S. K., Vijayan, V. A., Mishra, S., and Singh, O. P. (2016). Knockdown resistance (kdr) mutations in Indian An. stephensi (Diptera: Culicidae) populations. Journal of Medical Entomology, 53(2), 315–320. [CrossRef]
- Dykes, C. L., Kushwah, R. B., Das, M. K., Sharma, S. N., Bhatt, R. M., Veer, V., Agrawal, O. P., Adak, T., and Singh, O. P. (2015). Knockdown resistance (kdr) mutations in Indian An. culicifacies populations. Parasites and Vectors, 8, 333. [CrossRef]
- Dykes, C. L., Sharma, G., Behera, A. K., Kapoor, N., Paine, M. J. I., Donnelly, M. J., and Singh, O. P. (2022). Tandem duplication of a genomic region encoding glutathione S-transferase epsilon-2 and -4 genes in DDT-resistant Anopheles stephensi strain from India. Scientific reports, 12(1), 17872. [CrossRef]
- Elanga-Ndille, E., Tchouakui, M., et al. (2023). Impact of insecticide resistance on malaria vector competence: a literature review. Malaria Journal, 22, Article 19. [CrossRef]
- Epis, S., Porretta, D., Mastrantonio, V., Comandatore, F., Sassera, D., Rossi, P., Cafarchia, C., Otranto, D., Favia, G., Genchi, C., Bandi, C., and Urbanelli, S. (2014). ABC transporters are involved in defense against permethrin insecticide in the malaria vector Anopheles stephensi. Parasites and vectors, 7, 349. [CrossRef]
- Gan, S. C., Leong, Y. Q., Barhanuddin, M. F. A., Wong, S. T., Wong, S. F., Mak, J. W., and Ahmad, R. (2021). Dengue fever and insecticide resistance in Aedes mosquitoes in Southeast Asia: A review. Parasites and Vectors, 14, Article 315. [CrossRef]
- Ganesh, K. N., Urmila, J., & Vijayan, V. A. (2003). Pyrethroid susceptibility & enzyme activity in two malaria vectors, Anopheles stephensi (Liston) &. A. culicifacies (Giles) from Mysore, India. The Indian journal of medical research, 117, 30–38.
- Gunasekaran, K., Muthukumaravel, S., Sahu, S. S., Vijayakumar, T., & Jambulingam, P. (2011). Glutathione S transferase activity in Indian vectors of malaria: A defense mechanism against DDT. Journal of medical entomology, 48(3), 561–569. [CrossRef]
- Hemingway, J., Ranson, H., Magill, A., Kolaczinski, J., Fornadel, C., Gimnig, J., Coetzee, M., Simard, F., Roch, D. K., Hinzoumbe, C. K., Pickett, J., Schellenberg, D., Gething, P., Hoppé, M., and Hamon, N. (2016). Averting a malaria disaster: Will insecticide resistance derail malaria control? The Lancet, 387(10029), 1785–1788. [CrossRef]
- Insecticide Resistance Action Committee. (2023, August 29). Introduction to resistance. https://irac-online.org/training-centre/resistance/.
- Kareemi, T. I., Mishra, A. K., Chand, S. K., Nirankar, J. K., Vishwakarma, A. K., Tiwari, A., and Bharti, P. K. (2022). Analysis of the insecticide resistance mechanism in An. culicifacies sensu lato from a malaria-endemic state in India. Transactions of the Royal Society of Tropical Medicine and Hygiene, 116(3), 252–260. [CrossRef]
- Kaura, T., Devi, S., Mewara, A., Kaur, J., Singh, N. I., Lovleen, Sharma, S. K., Ratho, R. K., Sehgal, R., and Grover, G. S. (2022). Detection of insecticide susceptibility status and kdr mutation in field-collected A. aegypti from different districts of Punjab, India. Journal of Communicable Diseases, 54(4), 21–28. [CrossRef]
- Krishnan, J., Anbalagan, R., Shukla, A., Subramanian, V., and Srivastava, P. K. (2021). Monitoring of insecticide resistance and exploring the presence of virus in field populations of Cx. gelidus at Thiruvarur District of Tamil Nadu, India. Journal of Communicable Diseases, 53(4), 76–83. [CrossRef]
- Kumar, G., Gupta, S., Kaur, J., Pasi, S., Baharia, R., Mohanty, A. K., Goel, P., Sharma, A., and Rahi, M. (2024). Mapping malaria vectors and insecticide resistance in a high-endemic district of Haryana, India: Implications for vector control strategies. Malaria Journal, 23(1), 107. [CrossRef]
- Kumar, S., Kaushik, G., and Villarreal-Chiu, J. F. (2016). Scenario of organophosphate pollution and toxicity in India: A review. Environmental Science and Pollution Research, 23(10), 9480–9491. [CrossRef]
- Kumawat, N., Meena, S., and Kumari, V. (2021). Insecticide resistance status of Aedes mosquito vector in India: A review. International Journal of Mosquito Research, 8(4), 20–26.
- Kushwah, R. B. S., Kaur, T., Dykes, C. L., et al. (2020). A new knockdown resistance (kdr) mutation, F1534L, in the voltage-gated sodium channel of A. aegypti, co-occurring with F1534C, S989P and V1016G. Parasites and Vectors, 13, 327. [CrossRef]
- Kushwah, R. B., Dykes, C. L., Kapoor, N., Adak, T., and Singh, O. P. (2015). Pyrethroid-resistance and presence of two knockdown resistance (kdr) mutations, F1534C and a novel mutation T1520I, in Indian A. aegypti. PLoS Neglected Tropical Diseases, 9(1), e3332. [CrossRef]
- Liu, N. (2015). Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annual Review of Entomology, 60, 537–559. [CrossRef]
- Mastrantonio, V., Ferrari, M., Negri, A., Sturmo, T., Favia, G., Porretta, D., Epis, S., and Urbanelli, S. (2019). Insecticide Exposure Triggers a Modulated Expression of ABC Transporter Genes in Larvae of Anopheles gambiae s.s. Insects, 10(3), 66. [CrossRef]
- Mehrotra, K. N. (1989). Pesticide resistance in insect-pests Indian scenario. Pesticide Research Journal, 1(2), 95–103. http://www.spsindia.org.in/images/files/2.Mehorotra.pdf.
- Misra, B. R., and Gore, M. (2015). Malathion Resistance Status and Mutations in Acetylcholinesterase Gene (Ace) in Japanese Encephalitis and Filariasis Vectors from Endemic Area in India. Journal of medical entomology, 52(3), 442–446. [CrossRef]
- Modak, M. P., and Saha, D. (2022). First report of F1534C kdr mutation in deltamethrin resistant A. albopictus from northern part of West Bengal, India. Scientific Reports, 12(1), 13653. [CrossRef]
- Modak, P. M., Das, S., Saha, A., and Saha, D. (2024). Insecticide resistance and occurrence of L1014F kdr mutation in wild Cx. quinquefasciatus populations from sub-Himalayan region of West Bengal, India. Journal of Communicable Diseases, 56(3), 81–90. [CrossRef]
- Mukhopadhyay, A. K., Chakraborty, S., Karmakar, P., and Banerjee, P. L. (1996). Insecticidal susceptibility status of An. stephensi (Liston) in selected areas of Calcutta (West Bengal). Indian Journal of Public Health, 40(4), 130–133.
- Mukhopadhyay, A. K., Sinha, S. N., Yadav, R. L., and Narasimham, M. V. (1993). Susceptibility status of Cx. quinquefasciatus in Patna to insecticides. Indian Journal of Public Health, 37(2), 57–60.
- Muthusamy, R., and Shivakumar, M. S. (2015). Susceptibility status of Aedes aegypti (L.) (Diptera: Culicidae) to temephos from three districts of Tamil Nadu, India. Journal of vector borne diseases, 52(2), 159–165.
- Natarajan, E., Verma, V., Velamuri, P., Sreehari, U., and Kamaraju, R. (2014). Neonicotinoids in vector control: In silico approach. Asian Journal of Biomedical and Pharmaceutical Sciences, 4(39), 25–29. [CrossRef]
- National Center for Vector Borne Diseases Control. (2022, December 9). Insecticides: Formulations and dosage (IRS and larvicide). https://ncvbdc.mohfw.gov.in/Doc/tech-specification/Insecticides-Formulations_Dosage(IRS%20and%20Larvicide).pdf.
- National Institute of Malaria Research. (2006). Environmental management plan for the Vector Borne Disease Control Project India. Indian Council of Medical Research. https://nvbdcp.gov.in/Doc/EMP_final.pdf.
- Nayak, P., and Solanki, H. (2021). Pesticides and Indian agriculture – A review. International Journal of Research – Granthaalayah, 9(5), 250–263. [CrossRef]
- Nihad, P. P. M., Subramanian, M., Gunasekaran, K., & Kumar, A. (2023). Response to insecticides and underlying mechanisms of resistance in the field populations of Aedes aegypti Linnaeus (Diptera: Culicidae) in Puducherry, India. Journal of Vector Borne Diseases, 60(4), 393–400. [CrossRef]
- Paul, R. (2018, February 20). List of chemicals used for pest control: Agriculture. Essays, Research Papers and Articles on Agriculture in India. https://www.agricultureinindia.net/pest-control/chemicals-pest-control/list-of-chemicals-used-for-pest-control-agriculture.
- Raghavendra, K., Barik, T. K., Reddy, B. P., Sharma, P., and Dash, A. P. (2011). Malaria vector control: From past to future. Parasitology Research, 108(4), 757–779. [CrossRef]
- Raghavendra, K., Rahi, M., Verma, V., Velamuri, P. S., Kamaraju, D., Baruah, K., Chhibber-Goel, J., and Sharma, A. (2022). Insecticide resistance status of malaria vectors in the malaria endemic states of India: Implications and way forward for malaria elimination. Heliyon, 8(12), e11902. [CrossRef]
- Rahi, M., Mishra, A., Chand, G., Baharia, R., Hazara, R., Singh, S., Khan, S., Sreehari, U., Kamaraju, D., Kumar, G., Gupta, S., Sharma, A., Raghavendra, K., Gunasekaran, K., Singh, O., and Subbarao, S. (2024). Malaria vector bionomics: Countrywide surveillance study on implications for malaria elimination in India. JMIR Public Health and Surveillance, 10, e42050. https://publichealth.jmir.org/2024/1/e42050.
- Rai, P., and Saha, D. (2022). Occurrence of L1014F and L1014S mutations in insecticide resistant Cx. quinquefasciatus from filariasis endemic districts of West Bengal, India [Retracted]. PLoS Neglected Tropical Diseases, 16(1), e0010000. [CrossRef]
- Rai, P., and Saha, D. (2023). Retraction: Occurrence of L1014F and L1014S mutations in insecticide resistant Cx. quinquefasciatus from filariasis endemic districts of West Bengal, India. PLoS Neglected Tropical Diseases, 17(6), e0011417. [CrossRef]
- Rai, P., Bharati, M., Subba, A., and Saha, D. (2019). Insecticide resistance mapping in the vector of lymphatic filariasis, Cx. quinquefasciatus Say from northern region of West Bengal, India. PLoS ONE, 14(5), e0217706. [CrossRef]
- Ramkumar, G., Muthusamy, R., Narayanan, M., Shivakumar, M. S., and Kweka, E. J. (2023). Overexpression of cytochrome P450 and esterase genes involved in permethrin resistance in larvae and adults of Culex quinquefasciatus. Parasitology research, 122(12), 3205–3212. [CrossRef]
- Ranson, H., and Hemingway, J. (2005). Mosquito glutathione transferases. Methods in enzymology, 401, 226–241. [CrossRef]
- Rath, A., Mohanty, I., and Hazra, R. K. (2018). Insecticide susceptibility status of invasive A. albopictus across dengue endemic districts of Odisha, India. Pest Management Science, 74(6), 1431–1440. [CrossRef]
- Saha, A., Das, P., Das, S., and Saha, D. (2025). Overexpression of cytochrome P450 and esterase genes in pyrethroid-resistant wild Culex quinquefasciatus populations from sub-Himalayan West Bengal, India. Parasitology research, 124(6), 68. [CrossRef]
- Saha, P., Ballav, S., Chatterjee, M., Ganguly, S., Sarker, M., Biswas, A. K., Pramanik, T., Basu, N., and Maji, A. K. (2018). The status of susceptibility of Japanese encephalitis vectors to insecticides in endemic areas of northern districts of West Bengal, India. Japanese Journal of Infectious Diseases, 71(2), 91–98. [CrossRef]
- Saha, P., Chatterjee, M., Ballav, S., Chowdhury, A., Basu, N., and Maji, A. K. (2019). Prevalence of kdr mutations and insecticide susceptibility among natural population of A. aegypti in West Bengal. PLoS ONE, 14(4), e0215541. [CrossRef]
- Sahu, S. S., Gunasekaran, K., Vijayakumar, T., and Jambulingam, P. (2015). Triple insecticide resistance in Anopheles culicifacies: A practical impediment for malaria control in Odisha State, India. The Indian journal of medical research, 142 Suppl(Suppl 1), S59–S63. [CrossRef]
- Sahu, S. S., Sonia, T., Dash, S., Gunasekaran, K., and Jambulingam, P. (2019). Insecticide resistance status of three vectors of Japanese encephalitis in east central India. Medical and Veterinary Entomology, 33(2), 213–219. [CrossRef]
- Samal, R. R., and Kumar, S. (2021). Cuticular thickening associated with insecticide resistance in dengue vector, A. aegypti L. International Journal of Tropical Insect Science, 41, 809–820. [CrossRef]
- Sarkar, M., Bhattacharyya, I. K., Borkotoki, A., Goswami, D., Rabha, B., Baruah, I., and Srivastava, R. B. (2009). Insecticide resistance and detoxifying enzyme activity in the principal bancroftian filariasis vector, Cx. quinquefasciatus, in northeastern India. Medical and Veterinary Entomology, 23(2), 122–131. [CrossRef]
- Sharma, N., and Moin, S. (2021). Insecticide resistance, its origin and historical purview in reference to the mosquito vector. Journal of Environment, Science and Technology, 7(2). https://www.gyanvihar.org/journals/wp-content/uploads/2021/09/3.-Nitesh-Review-16-25.pdf.
- Sharma, V. P. (2003). DDT: The fallen angel. Current Science, 85(11), 1532–1537. http://www.jstor.org/stable/24110014.
- Shetty, V., Sanil, D., and Shetty, N. J. (2013). Insecticide susceptibility status in three medically important species of mosquitoes, An. stephensi, A. aegypti and Cx. quinquefasciatus, from Bruhat Bengaluru Mahanagara Palike, Karnataka, India. Pest Management Science, 69(2), 257–267. [CrossRef]
- Sindhania, A., Lohani, H. P., Kona, M. P., Kaur, T., Kaushal, B. R., and Singh, O. P. (2023). Molecular forms of the Indian An. subpictus complex differ in their susceptibility to insecticides and the presence of knockdown resistance (kdr) mutations in the voltage-gated sodium channel. PLoS ONE, 18(2), e0280289. [CrossRef]
- Singh, O. P., Bali, P., Hemingway, J., Subbarao, S. K., Dash, A. P., and Adak, T. (2009). PCR-based methods for the detection of L1014 kdr mutation in An. culicifacies sensu lato. Malaria Journal, 8, 154. [CrossRef]
- Singh, R. K., Kumar, G., and Mittal, P. K. (2014). Insecticide susceptibility status of malaria vectors in India: A review. International Journal of Mosquito Research, 1(1), 5–9.
- Srivastava, P. K., and Dhariwal, A. C. (2016). Compendium on entomological surveillance and vector control in India. National Center for Vector Borne Diseases Control (NCVBDC). https://ncvbdc.mohfw.gov.in/WriteReadData/l892s/Compendium-Entomological-SurveillanceandVector-Control-India.pdf.
- Suman, D. S., Devi, N., Tikar, S. N., Parashar, B. D., and Prakash, S. (2010). Development of insecticide resistance in Cx. quinquefasciatus mosquito from different locations in India. Journal of Pesticide Science, 35(1), 27–32. [CrossRef]
- Thomas, T. G., Prakash, V., Singh, S., Mandal, A. K., and Chauhan, L. S. (2013). Insecticide susceptibility status of Cx. quinquefasciatus Say, the vector of bancroftian filariasis against temephos in Delhi and National Capital Region. Japanese Journal of Infectious Diseases, 66(3), 238–240. [CrossRef]
- Thomas, T. G., Sharma, S. K., Prakash, A., Sokhey, J., and Sharma, B. R. (2000). Insecticide susceptibility status of Cx. tritaeniorhynchus Giles, vector of Japanese encephalitis in Delhi. Japanese Journal of Infectious Diseases, 53(1), 11–14.
- Viswan, K. A., Pushapalatha, E., and Azhahianambi, P. (2016). Application of synthetic insecticide and change in detoxifying enzyme levels in Cx. quinquefasciatus Say. International Journal of Mosquito Research, 3(6), 31–35.
- World Health Organization. (2015). Global plan for insecticide resistance management in malaria vectors (GPIRM). https://www.who.int/publications/i/item/WHO-HTM-GMP-2012.5.
- World Health Organization. (2015). Indoor residual spraying: An operational manual for indoor residual spraying (IRS) for malaria transmission control and elimination (2nd ed.). https://iris.who.int/handle/10665/177242.
- World Health Organization. (2017, September 20). Report of the twentieth WHOPES working group meeting, WHO/HQ, Geneva, 20–24 March 2017. https://www.who.int/publications/i/item/who-htm-ntd-whopes-2017.04.
- World Health Organization. (2020). Global plan for insecticide resistance management (GPIRM). https://www.who.int/publications/i/item/9789241564472.
Table 1.
List of different insecticides with recommended doses against different mosquito vectors.
| Insecticide | Formulation | Dosage | Application method | Duration of action | Target vector | Ref. |
| Organochlorines | ||||||
| DDT | 50% WP* | 1.0-1.5 g/m² | Indoor Residual Spraying (IRS) | Up to 6 months | Anopheles (malaria), Phlebotomus | WHO, 2015; NCVBDC, 2022 |
| Organophosphates | ||||||
| Malathion | 25% WP* | 1-2 g/m² | IRS | 2-3 months | Anopheles, Aedes, Culex | WHO, 2015; NCVBDC, 2022 |
| Malathion | 50% EC* | 1:100 (fogging dilution) | Fogging | 2-3 months | Anopheles, Aedes, Culex | NIMR, 2006; NCVBDC, 2022 |
| Temephos | 50% EC* | 2.5 cc in 10 L | Larvicide in Water | 1 Week | Anopheles, Aedes, Culex | NIMR, 2006; NCVBDC, 2022 |
| Fenthion | 100% EC* | 5 cc in 10 L | Larvicide in Water | 1 Week | Mosquitoes, flies | NIMR, 2006 |
| Fenitrothion | 50% or 25% WP* | 2 g/m² | IRS | 3-6 months | Anopheles, Aedes | WHO, 2015 |
| Pirimiphos-methyl | 50% WP*, 50% EC*, | 1-2 g/m² | IRS | 2-3 months | Anopheles, Aedes, Culex | WHO, 2015 |
| Pyrethroids | ||||||
| Deltamethrin | 2.5% WP*, 62.5 SC* | 20-25 mg/m² | IRS, ITN | 6 months | Anopheles, Phlebotomus | WHO, 2015; NCVBDC, 2022 |
| Alpha-Cypermethrin | 5% WP*, 10% SC* |
25 mg/m² | IRS | 4-6 months | Anopheles, Phlebotomus | WHO, 2015; NCVBDC, 2022 |
| Lambda-Cyhalothrin | 10% WP* | 25 mg/m² | IRS, ITN | 3-6 months | Anopheles | NIMR, 2006; WHO, 2015; NCVBDC, 2022 |
| Cyfluthrin | 10% WP* | 20-50 mg/m² | IRS, ITN | 3-6 months | Mosquitoes, Houseflies | NIMR, 2006; WHO, 2015; NCVBDC, 2022 |
| Bifenthrin | 10% WP* | 20-50 mg/m² | IRS | 3-6 months | Mosquitoes | WHO, 2015; NCVBDC, 2022 |
| Etofenprox | 5% WP* | 100-300 mg/m² | IRS | 3-6 months | Mosquitoes, flies | WHO, 2015 |
| Permethrin | 10% EC* | 1:100 dilution for fogging | Space spraying, Fogging | 1 month | Aedes | NIMR, 2006 |
| Cyphenothrin | 5% EC* | 0.50 mg/m² for spraying 0.35 mg/m² for fogging |
Space spraying, Fogging | 1-3 months | Mosquitoes, flies | NCVBDC, 2022 |
| Carbamates | ||||||
| Propoxur | 20% WP* | 1-2 g/m² | IRS | 3-6 months | Mosquitoes, Houseflies | WHO, 2015 |
| Bendiocarb | 80% WP* | 100-400 mg/m² | IRS | 2-6 months | Mosquitoes, flies | WHO, 2015 |
| Neonicotinoids | ||||||
| Clothianidin | 50% WDG* | IRS | up to 6-9 months | Mosquitoes | WHO, 2017 | |
*WP = Wettable powder; EC = Emulsifiable concentrate; SC = Suspension concentrates; WDG = Water Dispersible Granules.
Table 2.
The status of insecticide resistance in Anopheles mosquito vectors in various Indian states.
Table 2.
The status of insecticide resistance in Anopheles mosquito vectors in various Indian states.
| State | Mosquito Species | Insecticides Resistant | Reference(s) |
| Assam | An. minimus | Possible Resistant to Organochlorine (DDT) | Singh et al., 2014 |
| Chhattisgarh | An. culicifacies | Organochlorine (DDT), Organophosphate (Malathion), Pyrethroid (Deltamethrin) | Singh et al., 2014 |
| Gujarat | An. culicifacies | Organochlorine (DDT), Organophosphate (Malathion), Pyrethroid (Deltamethrin) | Raghavendra et al., 2022 |
| An. stephensi | Resistant to Organochlorine (DDT), Organophosphate (Malathion) and Possible Resistant to Pyrethroid (Deltamethrin) | Singh et al., 2014 | |
| Haryana | An. culicifacies | Organochlorine (DDT), Organophosphate (Malathion), Pyrethroid (Deltamethrin) | Raghavendra et al., 2022 |
| An. stephensi | Organochlorine (DDT) | Raghavendra et al., 2022 | |
| Jharkhand | An. culicifacies, An. annularis | Resistant to DDT (Organochlorine) and Possible Resistant to Organophosphate (Malathion) | Singh et al., 2014; Raghavendra et al., 2022 |
| An. fluviatilis, | DDT (Organochlorine) | Singh et al., 2014; Raghavendra et al., 2022 | |
| Karnataka | An. culicifacies | Organochlorine (DDT), Organophosphate (Malathion), Pyrethroid (Deltamethrin, alpha-cypermethrin) | Raghavendra et al., 2022 |
| An. stephensi | Organophosphate (Malathion) | Singh et al., 2014 | |
| Kerala | An. stephensi | DDT (Organochlorine) | Singh et al., 2014 |
| Madhya Pradesh | An. culicifacies | Resistant to Organochlorine (DDT), Organophosphate (Malathion) and Possible Resistant to Pyrethroid (Deltamethrin) | Singh et al., 2014; Raghavendra et al., 2022 |
| An. stephensi | Resistant to Organochlorine (DDT) and Possible Resistant to Organophosphate (Malathion) | Singh et al., 2014 | |
| Maharashtra | An. culicifacies | Resistant to Organochlorine (DDT), Organophosphate (Malathion), Pyrethroid (Deltamethrin, cyfluthrin, alpha-cypermethrin, and lambda-cyhalothrin) | Raghavendra et al., 2022 |
|
An. fluviatilis, An. annularis |
Organochlorine (DDT) | Singh et al., 2014 | |
| Odisha | An. culicifacies | Resistant to Organochlorine (DDT) and Possible Resistant to Pyrethroid (Deltamethrin) | Singh et al., 2014; Raghavendra et al., 2022 |
| An. fluviatilis | Possible Resistant to Organochlorine (DDT) | Singh et al., 2014; Raghavendra et al., 2022 | |
| An. minimus | Organochlorine (DDT) | Singh et al., 2014 | |
| Rajasthan | An. culicifacies, An. stephensi | Resistant to Organochlorine (DDT) and Possible Resistant to Organophosphate (Malathion) | Singh et al., 2014 |
| Tamil Nadu | An. culicifacies | Organochlorine (DDT), Pyrethroid (Deltamethrin) | Singh et al., 2014 |
| Tripura | An. minimus | Possible Resistant to Organochlorine (DDT) | Singh et al., 2014 |
| Uttar Pradesh | An. culicifacies | Resistant to Organochlorine (DDT) and Possible Resistant to Organophosphate (Malathion) | Singh et al., 2014 |
| West Bengal | An. stephensi | Organochlorine (DDT), Carbamate (Propoxur), Organophosphate (Malathion) | Mukhopadhyay et al., 1996 |
Table 3.
The status of insecticide resistance in Aedes mosquito vectors among different states of India.
Table 3.
The status of insecticide resistance in Aedes mosquito vectors among different states of India.
| State | Mosquito Species | Insecticides | Reference(s) |
| Andhra Pradesh | A. aegypti | DDT (Organochlorine) and Malathion (Organophosphate) | Kumawat et al., 2021 |
| Arunachal Pradesh |
A. Aegypti, A. albopictus |
DDT (Organochlorine) | Kumawat et al., 2021 |
| Assam |
A. aegypti, A. albopictus |
DDT (Organochlorine), Temephos (Organophosphate) | Kumawat et al., 2021 |
| A. albopictus (Guwahati) | Resistant to DDT (Organochlorine) and Incipient Resistant to Deltamethrin, Permethrin (Pyrethroids) | Kumawat et al., 2021 | |
|
A. Aegypti, A. albopictus |
DDT (Organochlorine) | Kumawat et al., 2021 | |
| Delhi | A. aegypti | Resistant to DDT and Dieldrin (Organochlorine) and Possible Resistant to Temephos (Organophosphate) | Kumawat et al., 2021 |
| A. albopictus | Resistant to DDT (Organochlorine) and Incipient Resistance to Deltamethrin, Permethrin (Pyrethroids) | Kumawat et al., 2021 | |
| Goa | A. aegypti | DDT (Organochlorine) | Kumawat et al., 2021 |
| Haryana (Gurgaon) | A. albopictus | Resistant to DDT (Organochlorine) and Incipient Resistance to Deltamethrin, Permethrin (Pyrethroids) | Kumawat et al., 2021 |
| Jharkhand |
A. Aegypti, A. albopictus |
DDT (Organochlorine) | Kumawat et al., 2021 |
| Karnataka | A. aegypti | DDT (Organochlorine), Propoxur (Carbamate), deltamethrin, and Permethrin (Pyrethroid) | Kumawat et al., 2021 |
| Kerala |
A. Aegypti, A. albopictus |
DDT and Dieldrin (Organochlorine) | Kumawat et al., 2021 |
| A. albopictus | Resistant to DDT (Organochlorine) and Incipient Resistance to Deltamethrin, Permethrin (Pyrethroids) | Kumawat et al., 2021 | |
| Maharashtra | A. albopictus | DDT (Organochlorine) | Kumawat et al., 2021 |
| A. aegypti | Temephos, Fenthion (Organophosphate) | Kumawat et al., 2021 | |
| Odisha | A. albopictus | DDT (Organochlorine) | Baig et al., 2021; Rath et al., 2018 |
| A. aegypti | Resistant to DDT (Organochlorine), Deltamethrin (Pyrethroids) and Possible Resistance Malathion (Organophosphate) | Baig et al., 2021; Rath et al., 2018 | |
| Rajasthan |
A. aegypti, A. vittatus, A. w-albus |
DDT and Dieldrin (Organochlorine) | Kumawat et al., 2021 |
| A. aegypti | Possible Resistant to Cypermethrin, Permethrin (Pyrethroids) | Kumawat et al., 2021 | |
| Tamil Nadu | A. aegypti | Permethrin (Pyrethroid) | Kumawat et al., 2021 |
| Uttarakhand (Haridwar) | A. albopictus | Resistant to DDT (Organochlorine) and Incipient Resistance to Deltamethrin, Permethrin (Pyrethroids) | Kumawat et al., 2021 |
| West Bengal | A. albopictus | DDT (Organochlorine), Permethrin (pyrethroid), propoxur (carbamate), and Temephos (Organophosphate) | Kumawat et al., 2021 |
| A. aegypti | Permethrin (Pyrethroid) and Propoxur (Carbamate) | Kumawat et al., 2021 |
Table 4.
The status of insecticide resistance in Culex mosquito vectors in various Indian states.
| State | Mosquito Species | Insecticides | Reference(s) |
| Assam | Cx. gelidus, Cx. vishnui |
Suspected Resistant to DDT (Organochlorine), Sensitive to Deltamethrin (Pyrethroid) |
Dhiman et al., 2013 |
| Cx. quinquefasciatus | Resistant to DDT (Organochlorine), Susceptible to Deltamethrin (Pyrethroid) |
Sarkar et al., 2009 | |
| Bihar (Patna) | Cx. quinquefasciatus | Organochlorine (DDT and dieldrin) | Mukhopadhyay et al., 1993 |
| Delhi | Cx. tritaeniorhynchus | Organochlorine (DDT), Organophosphate (Malathion, Fenitrothion), Carbamate (Propoxur) | Thomas et al., 2000 |
| Cx. quinquefasciatus | Temephos (Organophosphate) | Thomas et al., 2013 | |
| Gujarat (Jamnagar) | Cx. quinquefasciatus | Lower resistant to Pyrethroid (alpha-cypermethrin) and Organophosphate (fenthion) | Suman et al., 2010 |
| Karnataka (Bengaluru) | Cx. quinquefasciatus | Highly resistant to Propoxur (Carbamate), Susceptible to Temephos (Organophosphate) | Paul, 2018 |
| Odisha | Cx. vishnui, Cx. tritaeniorhynchus |
DDT (Organochlorine), Deltamethrin (pyrethroids), and malathion (Organophosphate) | Sahu et al., 2019 |
| Cx. bitaeniorhynchus | Remains susceptible to DDT (organochlorine), deltamethrin (pyrethroids), and malathion (organophosphate). | ||
| Punjab (Bathinda) | Cx. quinquefasciatus | High larval Resistant to Organophosphate (temephos and fenthion) and pyrethroids (Lambda Cyhalothrin, alpha cypermethrin, and Cypermethrin) | Suman et al., 2010 |
| Rajasthan (Jodhpur) | Cx. quinquefasciatus | High Resistant to Organophosphate (Temephos, Fenthion), Neemarin (a neem-based insecticide), pyrethroids (Cypermethrin) | Suman et al., 2010 |
| Rajasthan (Bikaner) | Cx. quinquefasciatus | Moderately resistant to organophosphates (temephos, fenthion), Neemarin (a neem-based insecticide), and pyrethroids (alpha-cypermethrin). | |
| Tamil Nadu (Thiruvarur) | Cx. gelidus | Possible Resistant to Deltamethrin (pyrethroids) and malathion (Organophosphate) | Krishnan et al., 2021 |
| West Bengal | Cx. quinquefasciatus | organophosphate (temephos and malathion), carbamate (propoxur), synthetic pyrethroids (deltamethrin, lambdacyhalothrin and permethrin) and organochlorine (DDT) |
Rai et al., 2019 |
| Cx. tritaeniorhynchus | organochlorine (DDT), Deltamethrin (Pyrethroids), malathion (Organophosphate) | Saha et al., 2018 | |
| Cx. pseudovishnui | organochlorine (DDT), Deltamethrin (Pyrethroids), | Saha et al., 2018 | |
| Cx. vishnui | organochlorine (DDT), Deltamethrin (Pyrethroids) | Saha et al., 2018 | |
| Cx. gelidus | Resistant to organochlorine (DDT) and Possible Resistant to Deltamethrin (Pyrethroids) | Saha et al., 2018 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



