
Submitted:
18 September 2025
Posted:
19 September 2025
You are already at the latest version
Abstract
The intestinal microbiota and immune system are separated by an epithelial layer, whose barrier function is crucial for the healthy state of the host, while its impairment, usually associated with inflammation, leads to a variety of pathologies in humans. Different probiotics targeting the preservation of epithelial barrier homeostasis are thus currently being developed. Their selection and validation are a complex process involving in-vitro and in-vivo models. While the former lacks the complexity of the physiological system, the latter often fails to reflect the effect in humans.
We used the commercial microfluidic high-throughput device OrganoPlate to set up a Gut-on-Chip model comprising human epithelial and peripheral blood mononuclear cells (PBMC). As a proof-of-concept, we co-cultured several intestinal anaerobic bacteria species in direct contact with the epithelial monolayer for two days to investigate their impact on epithelial barrier integrity and immune response upon induction of inflammation. We demonstrate here that Bacteroides thetaiotaomicron protects the epithelial barrier and dampens CCL2 secretion provoked by LPS-stimulated PBMC.
Recapitulating the key features of intestinal inflammation, we suggest that the current Gut-on-Chip model enables an easy-to-use screen of next-generation probiotics and live biotherapeutics with homeostatic and immunomodulatory properties.
Keywords:
immunomodulation
; organ-on-chip
; probiotics
; epithelial barrier
; inflammation
; hypoxia
; anaerobe
; Bacteroides thetaiotaomicron
; Clostridium scindens
; CCL2
Introduction
The intestinal mucosa surface separates the host from the myriads of microorganisms found in the intestinal lumen. This barrier consists primarily of epithelial cells tightly connected thanks to different junction proteins, thus restricting the infiltration of external antigens between the cells [1,2]. Impairment of this barrier, a phenomenon called the “leaky gut”, and the associated inflammation underline and/or aggravate a variety of pathologies in humans starting from those manifested mainly within the gastrointestinal (GI) tract such as inflammatory bowel diseases (IBD), irritable bowel syndrome and celiac disease, and ending with those affecting distant organs throughout the human body, for instance rheumatoid arthritis, non-alcoholic fatty liver disease, diabetes, Parkinson’s disease and others [3,4]. Intensive research in the last decades unraveled the key role of the intestinal microbiota in the maintenance of the epithelial barrier homeostasis, while changes in the microbiome (also called “dysbiosis”) are often associated with a variety of autoimmune, metabolic and neurodegenerative diseases [5,6,7]. On the other hand, some microbial structural components and metabolites were demonstrated to strengthen the intestinal epithelial barrier through modification of the expression and spatial organization of junction proteins of intestinal epithelial cells [8,9] either directly or via the modulation of cytokines secretion (e.g. IFNγ, TNFα, IL-1, IL-6 and others) by epithelial and immune cells [10,11,12,13]. Therefore, “classical” probiotics and postbiotics, as well as next-generation probiotics such as live biotherapeutic products (LPBs), are extensively developed to aid in maintaining the intestinal epithelial barrier and dampening excessive and chronic inflammation [14,15]. However, while numerous product candidates pass through pre-clinical development that includes both in-vitro and in-vivo models, only ~15% of these candidates in the field of inflammatory diseases succeed in clinical trials and get approved [16]. Specifically, for IBDs, the use of adequate pre-clinical models that take into account microbiota–host interactions was identified as one of the major targets to improve the success rate of novel treatment development (Ciorba 2024). In this light, microfluidics-based models offer several advantages over “standard” (one cell-type based) in-vitro assays as well as over animal models including:
The use of flow, which mimics the physiological conditions in the intestine and also exerts shear stress on epithelial cells, promoting their differentiation [17,18]
- The possibility of combining several cell types in one assay (e.g. epithelial and immune cells)
- The 3D spatial organization of the tissues
- The use of human-derived host cells
Here, we co-cultured human PBMC and several intestinal bacteria in a previously described high-throughput Gut-on-Chip model [19,20,21], which allows rapid maturation of the epithelial monolayer and easy monitoring of its barrier integrity as well as cytokine sampling and microscopic analysis. Using E. coli LPS to stimulate PBMC, we induced inflammation-mediated epithelial damage and then tested the capacity of two bacterial strains to reduce the inflammation and protect the epithelial barrier. We suggest that the current Gut-on-Chip model recapitulates the key features of intestinal inflammation and enables an easy-to-use screen of novel (bio) therapeutics in a high-throughput manner.
Materials and Methods
Mammalian Cells
Human colon adenocarcinoma cell line Caco-2/TC-7 (Caco2) (Sigma-Aldrich) at passages between 2 and 12, were cultured in MEM medium (Gibco) supplemented with 10% FBS (Gibco), 1% Sodium Pyruvate (Gibco), 1% Non-essential Amino Acids (NEAA, Gibco) and 1% penicillin/streptomycin (Sigma-Aldrich). Cells were cultured at 37 °C in a humidified atmosphere with 5% CO2. PBMCs from human blood were isolated using standard procedure using Ficoll-Paque Plus (Cytiva). Briefly, the whole blood was diluted 1:1 with PBS and centrifuged at room temperature (RT) for 30 min at 400 g without a break on top of the Ficoll layer. The interphase containing PBMC was then recovered, and the cells were washed in PBS by centrifugation at 600 g and then again at 450 g, 10 minutes each time. PBMC were set to 106 cells/mL in RPMI medium (Sigma-Aldrich) supplemented with 10% FBS (Gibco) and 1% penicillin/streptomycin (Sigma-Aldrich). The blood from healthy donors was supplied by the French Blood Establishment (EFS), which is responsible for the collection and distribution of blood in France. All donors provided informed consent to EFS. This sample collection does not require any ethical committee submission.
OrganoPlate Seeding
Epithelial tubule formation by Caco-2 cells in the 3-lane OrganoPlate (Mimetas) was performed as previously reported [20]. In short, 2*104 Caco-2 cells were seeded against a pH-buffered 4 mg/mL Collagen I gel (Collagen-I rat tail, R&D systems) in a 3-lane OrganoPlate and allowed to attach against the gel for 4 h (37 °C, 5% CO2). Caco-2 medium was then added to the inlets and outlets and bidirectional flow was created by placing the plate on an interval rocker (Perfusion Rocker, Mimetas) switching between a +7° and −7° inclination every 8 min (37 °C, 5% CO2). Medium was changed 4 days after the seeding. When indicated, 5 days after Caco-2 seeding, the medium in the opposite channel was exchanged with 105 PBMCs in supplemented RPMI, with (or without) LPS mix of three previously communicated strains of E. coli (Sigma-Aldrich, cat. nos. L2637, 3012 and 3137, [22]) at 100 ng/mL each.
Bacteria co-Culture in OrganoPlate
Bacterial strains (Table 1) were cultivated first on previously described mGAM-CRIM agar plates [23] for 48 hours in the BACTRON600 (Sheldon) anaerobic chamber at 37 °C. Then a single colony of each strain was inoculated in 5 mL mGAM-CRIM liquid broth and incubated in the chamber for 24 additional hours. The next day, bacterial cultures were centrifuged at 10000 g for 10 minutes and the supernatants were filtered through a 0.22µm filter (Millex-GS, Millipore). Bacteria pellets were washed twice in PBS by repeating resuspension and centrifugation steps and then set to an optical density of 1 in a MEM medium supplemented with 1% Sodium Pyruvate (Gibco), 1% NEAA (Gibco), 1X Insulin-Transferrin-Selenium-Ethanolamine solution (Gibco), called here “MEM-co-culture medium”, to support mammalian cell viability in the absence of serum. On day 5, upon Caco-2 seeding and simultaneously with PBMC (+/- E. coli LPS) administration in the bottom channel, the medium in the top channel was replaced by bacteria suspension in MEM-co-culture medium or with sterile medium supplemented with filtered bacteria culture at 30% final concentration. OrganoPlates were then incubated in hypoxic conditions: 4% O2, 5% CO2 at 37 °C. At indicated time-points, 10µl samples of medium from the top channels were taken, serially diluted in PBS, plated on mGAM-CRIM plates and incubated for 48h in the anaerobic chamber for colony-forming units (CFU) determination.
Cytokine Secretion
Media were collected separately from Lumen and Basal channels and stored at −20 °C until further assessment by Luminex method. The concentrations of CCL2, CCL20, CCL28, IFNγ, IL-2, IL-8, TNFα, CCL3, CCL25, CXCL10, IL-1β, IL-6 and IL-10 were quantified using a human multiplex assay (R&D Systems) according to manufacturer protocol using a BIOPLEX 200 reader (BIORAD). Only the results of quantitatively detected cytokines are presented. IL-8 concentration was assessed using ELISA method according to the manufacturer protocol (R&D Systems).
TEER Measurements
Transepithelial electrical resistance (TEER) was measured at different time points using an automated multichannel impedance spectrometer designed for use with the OrganoPlate (OrganoTEER, Mimetas) as described by Beaurivage et al. [20]. OrganoPlates were equilibrated at room temperature for 30 minutes before each measurement. Data was analyzed using OrganoTEER software, which automatically extracts the TEER contribution and reports values normalized to Ohm*cm2.
Immunohistochemistry and Microscopy
At the end point of experiments, all cell cultures were fixed in 4% paraformaldehyde (VWR 100504-858) in DPBS (+Ca/+Mg, Gibco 14040-083) for 30 min. Cell membranes were permeabilized with a 0.1% Triton-X-100 solution in DPBS for 20 min and blocked with a 2% BSA solution in DPBS (Sigma A2153) for over 1h. Primary antibodies against ZO-1 (Invitrogen 61-7300, AB_138452) were then diluted in a 2% BSA solution in DPBS and incubated overnight at 4°C. Secondary antibodies (Invitrogen A-21429, AB_2535850), DAPI (Invitrogen, D1306), and Phalloidin (Sigma, P5282) were then also diluted in 2% BSA and incubated for 2h. Cell cultures were rinsed with DPBS prior to imaging. All steps, except when specified otherwise, were performed on chip and at room temperature. Confocal immunostaining images were acquired using a spinning disk microscope (Andor BC34 CF) using a 20X magnification air objective (NA 0.8). Z-stacks were acquired over a range of 200 µm with a Z resolution of 0.3 µm.
Statistical Analysis
TEER measurements: repeated measures (RM) two-way or one-way Anova analysis was performed comparing each condition to a control conditioned decided for each type of experiment. Only statistically significant different results were considered when the p-value was lower than 0.05 and presented with asterisks.
IL-8 ELISA: ratio paired T test was performed comparing each condition to control condition. Only statistically significant different results were considered when the p-value was lower than 0.05 and presented with asterisks.
Multiplex cytokine analysis (Luminex): for each comparison considered in each dataset of interest, each assay was analyzed individually, comparing its expression in one condition compared to another using a non-parametric Wilcoxon test (the null hypothesis being that the mean expression of the considered assay is not significantly different between the two investigated conditions). To correct for the multiple testing effect, p-values were adjusted using the Benjamini-Hochberg approach [24]. The log2-fold-change associated with the comparison was calculated as the log2 ratio of mean expression levels between the two conditions. We considered that the expression of an assay was significantly different in the two investigated conditions when the adjusted p-value was lower than 0.05.
Results
A tubule-shaped monolayer of Caco2 cells within one of the channels (“Lumen” channel) was prepared within each chip of the microfluidic device called OrganoPlate®, as it was previously demonstrated by Beaurivage et al. [20]. Once the epithelial barrier (assessed by TEER) was established, 6 bacterial species (Table 1) commonly inhabiting the human intestine were introduced within the Lumen channel of the chips to survive in co-culture with the epithelial cells (Figure 1A). Since these bacteria are anaerobic, experiments were conducted at 4% oxygen, which better reflects the physiological values ranging from ∼6% in the vascularized submucosa to 1 - 2% near the crypt-lumen interface [25]. Except for Roseburia intestinalis, all 5 other species maintained a concentration of viable units above 106 CFU/chip (Figure 1B) for 48 hours. At the same time, no decrease in epithelial barrier efficiency (Figure 1D) and no significant increase in secretion of the main pro-inflammatory cytokine IL-8 [26,27] was detected (Figure 1C), suggesting that epithelial cells suffered no damage or inflammatory stress.
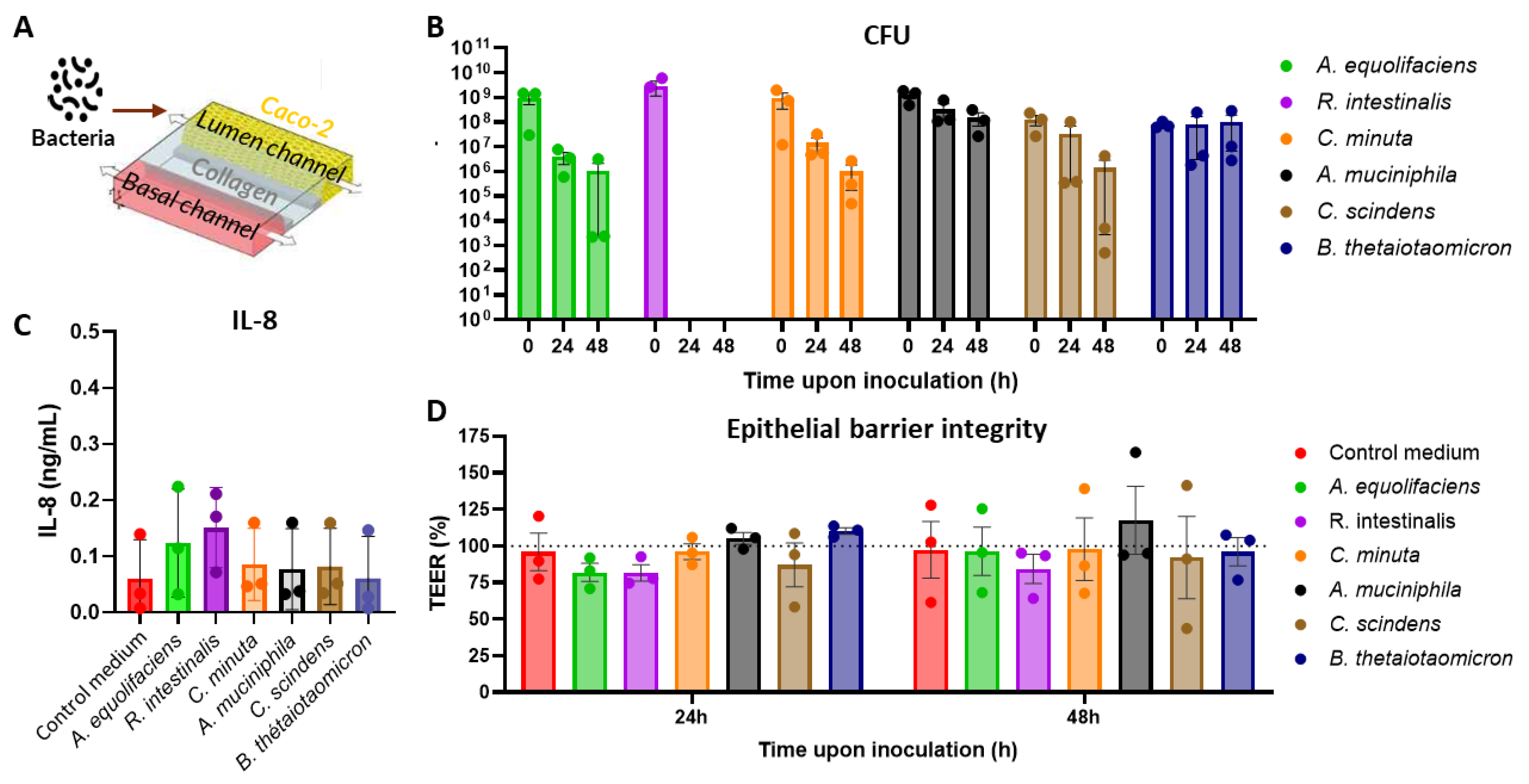
Encouraged by these observations, we tested the capacity of some of these bacterial species to mitigate inflammation-induced epithelial damage. We decided to proceed with two species that were proposed for live biotherapeutic development. The first is B. thetaiotaomicron, whose anti-inflammatory and epithelial barrier supporting features were demonstrated both in vitro and in vivo [28,29,30,31]. The second is C. scindens, which was not reported to possess such abilities but is capable of secondary bile acid production, preventing the emergence of important human pathogen Clostridioides (Clostridium) difficile [32,33] and playing a significant role in steroid metabolism in humans [34]. To mimic epithelial damage mediated by the pro-inflammatory activity of immune cells, we introduced human peripheral blood mononuclear cells (PBMC) into the opposite (“Basal”) channel of each chip and stimulated them with a lipopolysaccharide (LPS) from E.coli as we reported previously [19]. Simultaneously with PBMC addition to the Basal channel, washed B. thetaiotaomicron, C. scindens or their filtered spent culture medium from overnight cultures at 30% vol/vol were co-incubated in the Lumen channels (Figure 2A). Similarly to the experiment without PBMC, B. thetaiotaomicron viability remained roughly unchanged during the two days of the co-culture experiment, while the CFU of C. scindens rapidly declined (Figure 2B). Intriguingly, while LPS stimulation of PBMC provoked a decrease in TEER as previously described [19], co-culture with B. thetaiotaomicron or co-incubation with its supernatant protected the Caco-2 cells from the LPS-induced damage (Figure 2C). On the other hand, the spent medium of C. scindens, but not bacteria themselves, demonstrated only a (not statistically significant) tendency to mitigate the epithelial barrier damage.
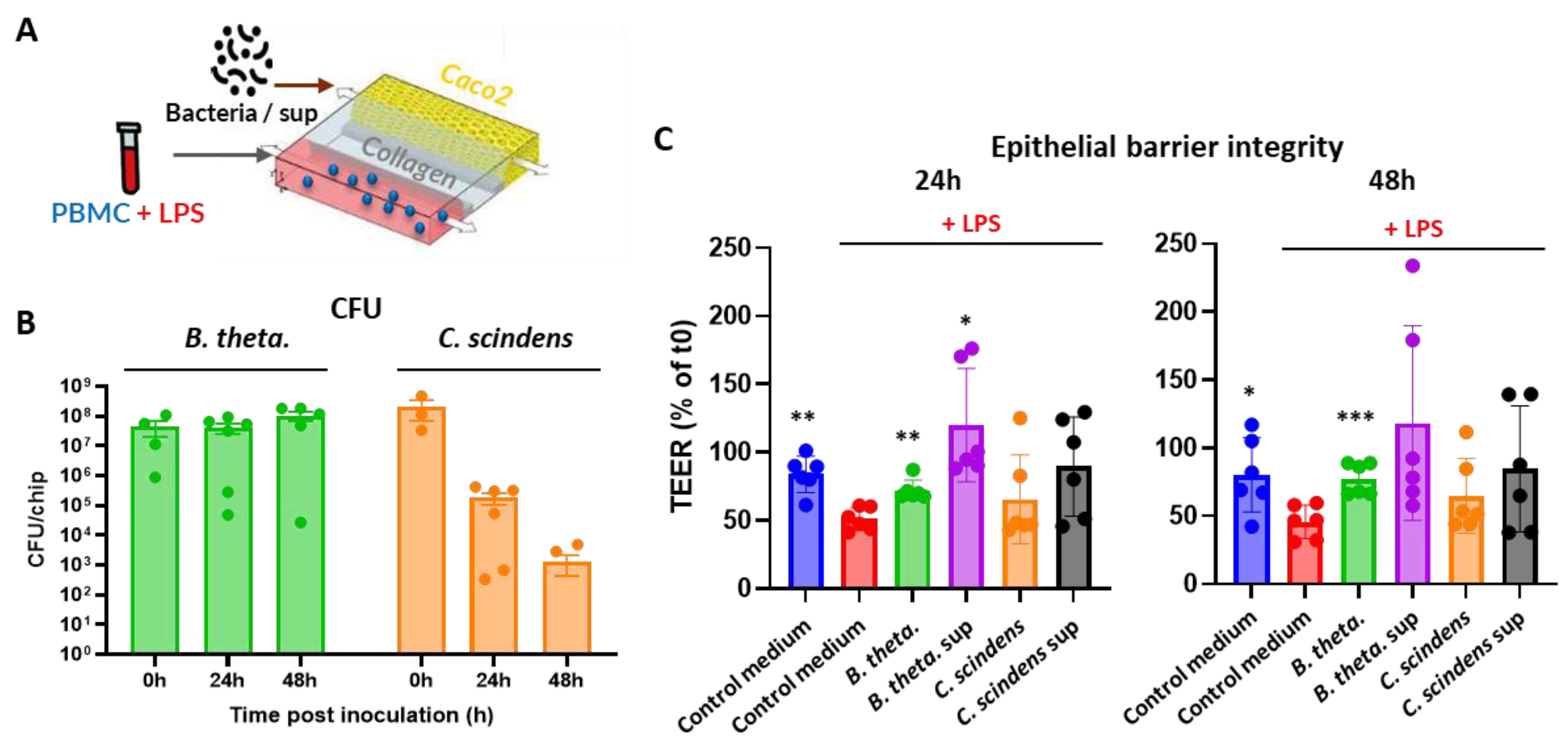
To test whether the protective effect is associated with immunomodulation, we analyzed the cytokine profile in both channels at the end of the 48 hours of the experiment. As expected, LPS stimulation of PBMC induced a sharp increase in concentration of most tested cytokines, which was more profound in the Basal channel where the PBMC resided (Figure 3A and Figure 3B). Interestingly, bacteria and their supernatants had very limited and mostly statistically insignificant effect on the cytokine profiles in the Basal channel (Figure 3C). On the other hand, the concentrations of CCL2 and CCL20 chemokines in the Lumen channel was strongly altered (Figure 3D). Specifically, B. thetaiotaomicron and its spent medium strongly decreased the CCL2 concentration, while only the bacteria, but not the supernatant, reduced the concentration of CCL20. Co-culture of C. scindens with the Caco-2 cells provoked even more profound decrease (about 17 folds) in the concentration of CCL20, however this tendency was reverted when only the spend medium of C. scindens was used. Accordingly, the concentration of two pro-inflammatory cytokines IL-6 and IL-8 also showed moderate, but statistically significant, increase when only the supernatant of C. scindens was added to the Lumen channel.
Since the efficiency of the epithelial barrier is maintained mainly by tight junction proteins [1,2], we investigated the spatial organization of the key component of this complex – Zonula Occludin-1 (ZO-1) – using immunofluorescence microscopy. Indeed, LPS stimulation of PBMC altered the homogeneous distribution of ZO-1 along Caco-2 cell edges and also impacted the general morphology of the epithelial monolayer as evidenced by the filamentous actin (F-Actin) staining (Figure 4). In accordance with TEER measurements, co-culturing B. thetaiotaomicron in the Lumen channel mitigated the alteration of ZO-1 and F-actin spatial distribution. The spent medium of B. thetaiotaomicron, on the other hand, provoked a dramatic effect on the organization of both markers: ZO-1 often appeared in patches along the cell edges, while F-actin seemed to be concentrated almost exclusively at the cell edges (cortex) in contrast to a more homogeneous distribution seen in other experimental conditions. The morphology of epithelial cells upon incubation with C. scindens or its supernatant looked similar to the condition with LPS-stimulated PBMC without any additional treatment.
Discussion
To co-culture anaerobic bacteria with host cells in vitro, researchers usually use sophisticated models involving pumps and/or anaerobic chambers that allow control of oxygen tension in different compartments of the microfluidic device [35,36,37,38,39,40]. The current model demonstrates the feasibility of prolonged co-culturing of some anaerobic bacteria in an easy and high-throughput mode simply by reducing the oxygen concentration to the physiologically relevant value of 4%. It is also possible that the rapid oxygen consumption by epithelial cells creates a niche in their close vicinity with even lower oxygen concentration, allowing anaerobic bacteria to survive.
LPS stimulation of PBMC in the current Gut-on-chip model upregulated several cytokines that play a crucial role in modulating the intestinal epithelial barrier (Figure 3). These include chemokines that control the recruitment of immune cells to the inflamed tissue, such as CCL-20, CCL2 (MCP-1) and CCL3 (MIP-1α), as well as cytokines regulating the activation status of these cells: IL-1β, IL-8, IL-6, TNFα and IL-10 [11,41]. Administration of live bacteria or their filtered supernatants in the Lumen channel mostly did not modulate the increase in cytokine concentration in opposite Basal channel, whose main source is PBMC (Figure 3C). In the Lumen channel, however, the increase in CCL2 concentration was reverted by B. thetaiotaomicron and its supernatant, and the increase in CCL20 was reverted by both bacterial strains (Figure 3D). As no modulation of these two chemokines was observed in the Basal channel, the impact of bacteria is most probably mediated via modulation of cytokine secretion from the Caco-2 cells. The decrease in CCL2, but not in CCL20, correlates with the improvement in epithelial barrier function assessed by TEER (Figure 2B), suggesting that CCL2 plays a key role in epithelial barrier damage in this model. In agreement with this observation, elevated CCL2 concentrations were detected in intestinal specimens of patients suffering from inflammatory bowel diseases (IBD) in several studies [42,43,44]. While C. scindens did not significantly improve epithelial barrier integrity, its pronounced impact on CCL20 secretion, a central chemokine in intestinal inflammation, underscores its relevance for future studies as a biotherapeutic in the autoimmune disease area [45,46]. However, the opposite effect of C. scindens supernatant on CCL20 as well as increase in IL-6 and IL-8 concentrations in the Lumen channel suggest ambivalent interaction of this species with intestinal epithelium.
On the cell morphology level, B. thetaiotaomicron co-culture with epithelial cells seems to prevent the LPS-induced decrease of ZO-1 localization at the cell edges (Figure 4), which aligns well with a previous communication about the positive effect of this species on ZO-1 mRNA expression [30]. The fact that the bacterial supernatant alone also promoted the protection of epithelial barrier function suggests that the effect of B. thetaiotaomicron is mediated by a secreted factor. Indeed, tryptophan metabolites, including indole, produced by this bacterium, were demonstrated to enhance epithelial barrier function [29]. However, the effect of the supernatant on F-actin and ZO-1 organization looked strikingly different from the one provoked by the B. thetaiotaomicron co-culture, although both treatments protected the epithelial barrier from inflammation-induced deterioration. While the patchy ZO-1 staining is not usually associated with a strong epithelial barrier, we observed a noticeable enhancement of cortical actin cytoskeleton, which is critical for the integrity of the gut barrier [47] and might explain the positive effect of B. thetaiotaomicron supernatant after all. In general, the different results obtained with live bacteria and its supernatant suggest that B. thetaiotaomicron has more than one way of impacting epithelial homeostasis. In fact, at least one cell-associated factor of B. thetaiotaomicron – pirin-like protein – was demonstrated to reduce pro-inflammatory NF-κB signaling in Caco-2 cells [28]. More investigations are needed to elucidate the complete picture of the effect of this species.
We thus conclude that the current Gut-on-Chip model recapitulates the key features of intestinal inflammation – epithelial barrier damage and pro-inflammatory cytokine secretion – while supporting the co-culture of anaerobic bacteria. Established in the commercially available OrganoPlate platform, the model is easily standardizable and scalable to higher-throughput applications. As a proof of concept, we screened six anaerobic bacteria and demonstrated the protective effect of B. thetaiotaomicron on the epithelial barrier against inflammation-mediated damage, associated with CCL2 secretion prevention and modulation of ZO-1 and F-actin spatial organization, while also flagging C. scindens as a promising candidate for future studies.
Figures and legends

Table 1.
Bacterial strains used in the study.
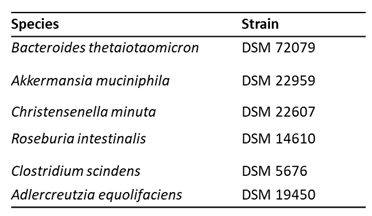 |
Figure 1.
Co-culture of anaerobic bacteria with Caco-2 in Gut-on-chip model. (A) Graphical representation of the experimental system with one microfluidic module (chip) within the OrganoPlate. (B) Colony forming units (CFU) of each bacterial species retrieved from the Lumen channel at the indicated time points. (C) IL-8 concentrations in the Lumen channel after 48 h co-culture with the different bacteria. Ratio paired T test was performed comparing each condition to Control medium and no condition obtained p-values <0.05. (D) TEER values along the experiment normalized to the T=0 h measured just prior to PBMC introduction and defined as 100%. RM two-way Anova analysis was performed comparing each condition to Control medium for each time point and no condition obtained p-values <0.05.
Figure 1.
Co-culture of anaerobic bacteria with Caco-2 in Gut-on-chip model. (A) Graphical representation of the experimental system with one microfluidic module (chip) within the OrganoPlate. (B) Colony forming units (CFU) of each bacterial species retrieved from the Lumen channel at the indicated time points. (C) IL-8 concentrations in the Lumen channel after 48 h co-culture with the different bacteria. Ratio paired T test was performed comparing each condition to Control medium and no condition obtained p-values <0.05. (D) TEER values along the experiment normalized to the T=0 h measured just prior to PBMC introduction and defined as 100%. RM two-way Anova analysis was performed comparing each condition to Control medium for each time point and no condition obtained p-values <0.05.
Each experiment was repeated three times with three technical replicates. Means and standard deviations are presented except for CFU data for which the standard error of the mean (SEM) is used.
Figure 2.
Co-culture of B. thetaiotaomicron (B. theta.) and C. scindens or incubation of their supernatants (sup) in the Gut-on-chip model with LPS-stimulated PBMC. (A) Graphical representation of the experimental system. (B) Colony forming units (CFU) of each bacterial species retrieved from the Lumen channel at the indicated time points. (C) TEER values along the experiment normalized T=0 h measured just prior to PBMC introduction and defined as 100%. RM one-way Anova analysis was performed with *, ** and *** representing statistical significance with p-values < 0.05, 0.01, and 0.001, respectively, when comparing each treatment to “Control medium” + LPS.
Figure 2.
Co-culture of B. thetaiotaomicron (B. theta.) and C. scindens or incubation of their supernatants (sup) in the Gut-on-chip model with LPS-stimulated PBMC. (A) Graphical representation of the experimental system. (B) Colony forming units (CFU) of each bacterial species retrieved from the Lumen channel at the indicated time points. (C) TEER values along the experiment normalized T=0 h measured just prior to PBMC introduction and defined as 100%. RM one-way Anova analysis was performed with *, ** and *** representing statistical significance with p-values < 0.05, 0.01, and 0.001, respectively, when comparing each treatment to “Control medium” + LPS.
Each experiment was repeated three times with PBMC from two donors each time (6 PBM donors in total) and at least three technical replicates. Means and standard deviations are presented except for CFU data for which SEM is used.
Figure 3.
Cytokine secretion in immunocompetent Gut-on-Chip model after 48h of the experiment start. (A and B) Relative change in cytokines concentration in Lumen (A) and Basal (B) channels after LPS-stimulated (or non-stimulated) PBMC administration normalized to “PBMC, No LPS” group using non-parametric Wilcoxon test. (C and D) Heat-map of relative change in cytokine concentration in Lumen (C) and Basal (D) channels comparing each treatment to the control (fresh) medium in the presence of LPS-stimulated PBMC in the Basal channel using non-parametric Wilcoxon test. The concentration of IL-8 in basal channel upon LPS stimulation was too high for quantitative assessment and therefore could not be plotted in panels A and C. *, ** and *** represent statistical significance with p-value < 0.05, 0.01, and 0.001, respectively. Each experiment was repeated three times with PBMC from two donors each time (6 PBM donors in total) and at least three technical replicates.
Figure 3.
Cytokine secretion in immunocompetent Gut-on-Chip model after 48h of the experiment start. (A and B) Relative change in cytokines concentration in Lumen (A) and Basal (B) channels after LPS-stimulated (or non-stimulated) PBMC administration normalized to “PBMC, No LPS” group using non-parametric Wilcoxon test. (C and D) Heat-map of relative change in cytokine concentration in Lumen (C) and Basal (D) channels comparing each treatment to the control (fresh) medium in the presence of LPS-stimulated PBMC in the Basal channel using non-parametric Wilcoxon test. The concentration of IL-8 in basal channel upon LPS stimulation was too high for quantitative assessment and therefore could not be plotted in panels A and C. *, ** and *** represent statistical significance with p-value < 0.05, 0.01, and 0.001, respectively. Each experiment was repeated three times with PBMC from two donors each time (6 PBM donors in total) and at least three technical replicates.
Figure 4.
Effect of B. thetaiotaomicron, C. scindens and their supernatants on Caco-2 morphology. Representative immunofluorescence images. Shown images are the mean projection of 5 consecutive slices of Z-stacks, centered on the bottom cell layer of the Lumen channel, acquired by confocal microscopy. Individual channels are shown separately in grayscale. A color image with all channels merged shown on the right with the following color code: nuclei (cyan), F-actin (magenta) and ZO-1 (orange). Scale bars represent 100 µm.
Figure 4.
Effect of B. thetaiotaomicron, C. scindens and their supernatants on Caco-2 morphology. Representative immunofluorescence images. Shown images are the mean projection of 5 consecutive slices of Z-stacks, centered on the bottom cell layer of the Lumen channel, acquired by confocal microscopy. Individual channels are shown separately in grayscale. A color image with all channels merged shown on the right with the following color code: nuclei (cyan), F-actin (magenta) and ZO-1 (orange). Scale bars represent 100 µm.

Conflict of interest statement
The authors declare no conflict of interest
References
- Buckley A, Turner JR: Cell Biology of Tight Junction Barrier Regulation and Mucosal Disease. Cold Spring Harbor perspectives in biology 2018, 10(1). [CrossRef]
- Moonwiriyakit A, Pathomthongtaweechai N, Steinhagen PR, Chantawichitwong P, Satianrapapong W, Pongkorpsakol P: Tight junctions: from molecules to gastrointestinal diseases. Tissue barriers 2023, 11(2):2077620. [CrossRef]
- Di Tommaso N, Gasbarrini A, Ponziani FR: Intestinal Barrier in Human Health and Disease. Int J Environ Res Public Health 2021, 18(23). [CrossRef]
- Stolfi C, Maresca C, Monteleone G, Laudisi F: Implication of Intestinal Barrier Dysfunction in Gut Dysbiosis and Diseases. Biomedicines 2022, 10(2). [CrossRef]
- Kinashi Y, Hase K: Partners in Leaky Gut Syndrome: Intestinal Dysbiosis and Autoimmunity. Front Immunol 2021, 12:673708. [CrossRef]
- Régnier M, Van Hul M, Knauf C, Cani PD: Gut microbiome, endocrine control of gut barrier function and metabolic diseases. The Journal of endocrinology 2021, 248(2):R67-r82. [CrossRef]
- Mou Y, Du Y, Zhou L, Yue J, Hu X, Liu Y, Chen S, Lin X, Zhang G, Xiao H et al: Gut Microbiota Interact With the Brain Through Systemic Chronic Inflammation: Implications on Neuroinflammation, Neurodegeneration, and Aging. Front Immunol 2022, 13:796288. [CrossRef]
- Kayama H, Okumura R, Takeda K: Interaction Between the Microbiota, Epithelia, and Immune Cells in the Intestine. Annual review of immunology 2020, 38:23-48. [CrossRef]
- Ghosh S, Whitley CS, Haribabu B, Jala VR: Regulation of Intestinal Barrier Function by Microbial Metabolites. Cellular and molecular gastroenterology and hepatology 2021, 11(5):1463-1482. [CrossRef]
- Neurath MF: Cytokines in inflammatory bowel disease. Nature Reviews Immunology 2014, 14(5):329-342.
- Barbara G, Barbaro MR, Fuschi D, Palombo M, Falangone F, Cremon C, Marasco G, Stanghellini V: Inflammatory and Microbiota-Related Regulation of the Intestinal Epithelial Barrier. Frontiers in nutrition 2021, 8:718356. [CrossRef]
- Chelakkot C, Ghim J, Ryu SH: Mechanisms regulating intestinal barrier integrity and its pathological implications. Experimental & molecular medicine 2018, 50(8):1-9. [CrossRef]
- Capaldo CT, Nusrat A: Cytokine regulation of tight junctions. Biochimica et biophysica acta 2009, 1788(4):864-871.
- Martyniak A, Medyńska-Przęczek A, Wędrychowicz A, Skoczeń S, Tomasik PJ: Prebiotics, Probiotics, Synbiotics, Paraprobiotics and Postbiotic Compounds in IBD. Biomolecules 2021, 11(12). [CrossRef]
- Tsai YL, Lin TL, Chang CJ, Wu TR, Lai WF, Lu CC, Lai HC: Probiotics, prebiotics and amelioration of diseases. Journal of biomedical science 2019, 26(1):3. [CrossRef]
- Wong CH, Siah KW, Lo AW: Estimation of clinical trial success rates and related parameters. Biostatistics (Oxford, England) 2019, 20(2):273-286.
- Pocock K, Delon L, Bala V, Rao S, Priest C, Prestidge C, Thierry B: Intestine-on-a-Chip Microfluidic Model for Efficient in Vitro Screening of Oral Chemotherapeutic Uptake. ACS biomaterials science & engineering 2017, 3(6):951-959. [CrossRef]
- Chi M, Yi B, Oh S, Park DJ, Sung JH, Park S: A microfluidic cell culture device (μFCCD) to culture epithelial cells with physiological and morphological properties that mimic those of the human intestine. Biomedical microdevices 2015, 17(3):9966. [CrossRef]
- Belotserkovsky I VC, Beitz B, Ben Abdallah B, Poissonier S, Bellais S, Hesketh A, Roelens M, Meza Torres J, Mouharib M, Sunshine J, Shaffer M, Parrino J, Silverman J, COSIPOP Study group, Daillere R, Vedrine C: Bifidobacterium longum subsp. infantis and Lacticaseibacillus rhamnosus GG protect intestinal epithelium against inflammation-mediated damage in an immunocompetent in-vitro model Preprint 2025.
- Beaurivage C, Naumovska E, Chang YX, Elstak ED, Nicolas A, Wouters H, van Moolenbroek G, Lanz HL, Trietsch SJ, Joore J et al: Development of a Gut-On-A-Chip Model for High Throughput Disease Modeling and Drug Discovery. Int J Mol Sci 2019, 20(22). [CrossRef]
- Gijzen L, Marescotti D, Raineri E, Nicolas A, Lanz HL, Guerrera D, van Vught R, Joore J, Vulto P, Peitsch MC et al: An Intestine-on-a-Chip Model of Plug-and-Play Modularity to Study Inflammatory Processes. SLAS TECHNOLOGY: Translating Life Sciences Innovation 2020:247263032092499. [CrossRef]
- Bounab Y, Eyer K, Dixneuf S, Rybczynska M, Chauvel C, Mistretta M, Tran T, Aymerich N, Chenon G, Llitjos JF et al: Dynamic single-cell phenotyping of immune cells using the microfluidic platform DropMap. Nature protocols 2020, 15(9):2920-2955. [CrossRef]
- Meslier V, Plaza Oñate F, Ania M, Nehlich M, Belotserkovsky I, Bellais S, Thomas V: Draft Genome Sequence of Isolate POC01, a Novel Anaerobic Member of the Oscillospiraceae Family, Isolated from Human Feces. Microbiology resource announcements 2022, 11(1):e0113421. [CrossRef]
- Benjamini Y, Hochberg Y: Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. Journal of the Royal Statistical Society: Series B (Methodological) 1995, 57(1):289-300. [CrossRef]
- Singhal R, Shah YM: Oxygen battle in the gut: Hypoxia and hypoxia-inducible factors in metabolic and inflammatory responses in the intestine. Journal of Biological Chemistry 2020, 295(30):10493-10505. [CrossRef]
- van Deventer SJ: Review article: Chemokine production by intestinal epithelial cells: a therapeutic target in inflammatory bowel disease? Aliment Pharmacol Ther 1997, 11 Suppl 3:116-120; discussion 120-111.
- Vebr M, Pomahačová R, Sýkora J, Schwarz J: A Narrative Review of Cytokine Networks: Pathophysiological and Therapeutic Implications for Inflammatory Bowel Disease Pathogenesis. Biomedicines 2023, 11(12). [CrossRef]
- Delday M, Mulder I, Logan ET, Grant G: Bacteroides thetaiotaomicron Ameliorates Colon Inflammation in Preclinical Models of Crohn's Disease. Inflammatory bowel diseases 2019, 25(1):85-96. [CrossRef]
- Luo Y, Lan C, Ren W, Wu A, Yu B, He J, Chen D: Bacteroides thetaiotaomicron: A symbiotic ally against diarrhea along with modulation of gut microbial ecological networks via tryptophan metabolism and AHR-Nrf2 signaling. Journal of advanced research 2025. [CrossRef]
- Pan M, Barua N, Ip M: Mucin-degrading gut commensals isolated from healthy faecal donor suppress intestinal epithelial inflammation and regulate tight junction barrier function. Front Immunol 2022, 13:1021094. [CrossRef]
- Li K, Hao Z, Du J, Gao Y, Yang S, Zhou Y: Bacteroides thetaiotaomicron relieves colon inflammation by activating aryl hydrocarbon receptor and modulating CD4(+)T cell homeostasis. International immunopharmacology 2021, 90:107183. [CrossRef]
- Buffie CG, Bucci V, Stein RR, McKenney PT, Ling L, Gobourne A, No D, Liu H, Kinnebrew M, Viale A et al: Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature 2015, 517(7533):205-208. [CrossRef]
- Abt MC, McKenney PT, Pamer EG: Clostridium difficile colitis: pathogenesis and host defence. Nature reviews Microbiology 2016, 14(10):609-620.
- Daniel SL, Ridlon JM: Clostridium scindens: history and current outlook for a keystone species in the mammalian gut involved in bile acid and steroid metabolism. FEMS microbiology reviews 2025, 49. [CrossRef]
- Zhang J, Huang YJ, Trapecar M, Wright C, Schneider K, Kemmitt J, Hernandez-Gordillo V, Yoon JY, Poyet M, Alm EJ et al: An immune-competent human gut microphysiological system enables inflammation-modulation by Faecalibacterium prausnitzii. NPJ Biofilms Microbiomes 2024, 10(1):31. [CrossRef]
- Jalili-Firoozinezhad S, Gazzaniga FS, Calamari EL, Camacho DM, Fadel CW, Bein A, Swenor B, Nestor B, Cronce MJ, Tovaglieri A et al: A complex human gut microbiome cultured in an anaerobic intestine-on-a-chip. Nature biomedical engineering 2019, 3(7):520-531.
- Marzorati M, Vanhoecke B, De Ryck T, Sadaghian Sadabad M, Pinheiro I, Possemiers S, Van den Abbeele P, Derycke L, Bracke M, Pieters J et al: The HMI module: a new tool to study the Host-Microbiota Interaction in the human gastrointestinal tract in vitro. BMC Microbiol 2014, 14:133. [CrossRef]
- Shah P, Fritz JV, Glaab E, Desai MS, Greenhalgh K, Frachet A, Niegowska M, Estes M, Jager C, Seguin-Devaux C et al: A microfluidics-based in vitro model of the gastrointestinal human-microbe interface. Nat Commun 2016, 7:11535. [CrossRef]
- Shin W, Wu A, Massidda MW, Foster C, Thomas N, Lee DW, Koh H, Ju Y, Kim J, Kim HJ: A Robust Longitudinal Co-culture of Obligate Anaerobic Gut Microbiome With Human Intestinal Epithelium in an Anoxic-Oxic Interface-on-a-Chip. Frontiers in bioengineering and biotechnology 2019, 7:13. [CrossRef]
- Fofanova TY, Karandikar UC, Auchtung JM, Wilson RL, Valentin AJ, Britton RA, Grande-Allen KJ, Estes MK, Hoffman K, Ramani S et al: A novel system to culture human intestinal organoids under physiological oxygen content to study microbial-host interaction. PLoS One 2024, 19(7):e0300666. [CrossRef]
- Singh UP, Singh NP, Murphy EA, Price RL, Fayad R, Nagarkatti M, Nagarkatti PS: Chemokine and cytokine levels in inflammatory bowel disease patients. Cytokine 2016, 77:44-49. [CrossRef]
- Reinecker HC, Loh EY, Ringler DJ, Mehta A, Rombeau JL, MacDermott RP: Monocyte-chemoattractant protein 1 gene expression in intestinal epithelial cells and inflammatory bowel disease mucosa. Gastroenterology 1995, 108(1):40-50. [CrossRef]
- MacDermott RP, Sanderson IR, Reinecker HC: The central role of chemokines (chemotactic cytokines) in the immunopathogenesis of ulcerative colitis and Crohn's disease. Inflammatory bowel diseases 1998, 4(1):54-67. [CrossRef]
- Banks C, Bateman A, Payne R, Johnson P, Sheron N: Chemokine expression in IBD. Mucosal chemokine expression is unselectively increased in both ulcerative colitis and Crohn's disease. The Journal of pathology 2003, 199(1):28-35. [CrossRef]
- Meitei HT, Jadhav N, Lal G: CCR6-CCL20 axis as a therapeutic target for autoimmune diseases. Autoimmunity reviews 2021, 20(7):102846. [CrossRef]
- Lee AY, Eri R, Lyons AB, Grimm MC, Korner H: CC Chemokine Ligand 20 and Its Cognate Receptor CCR6 in Mucosal T Cell Immunology and Inflammatory Bowel Disease: Odd Couple or Axis of Evil? Front Immunol 2013, 4:194. [CrossRef]
- Ivanov AI, Parkos CA, Nusrat A: Cytoskeletal Regulation of Epithelial Barrier Function During Inflammation. The American journal of pathology 2010, 177(2):512-524. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



