Submitted:
17 September 2025
Posted:
18 September 2025
You are already at the latest version
Abstract
Azaspiracids (AZAs) are marine polyether biotoxins produced by dinoflagellates that accumulate in filter-feeding organisms and pose a threat to human health and seafood safety. This study presents the first comprehensive analysis of azaspiracid analogs in shellfish from the Adriatic Sea including use of high-resolution mass spectrometry. AZA-2 was quantified in samples collected from Šibenik Bay between January and May 2024, with the highest concentrations observed in early January. In addition to AZA-2, several known analogs (AZA-6, AZA-9, AZA-10, AZA-19, AZA-41, AZA-43) and a potentially new analog (m/z 884.6066) were also detected. The fragmentation patterns of this new analog indicate a structural similarity to AZA-19 with a possible double bond modification. Potential pitfalls regarding misinterpretation of spectra derived from molecules containing 13C atoms were recognized. The presence of multiple analogs, some of which have high toxic potential, suggests that regulatory practice should consider including more than three analogs in the monitoring program.
Keywords:
LC-MS/MS
; LC-QTOF
; CID spectrum
; Adriatic Sea
; shellfish biotoxins
1. Introduction
Azaspiracids (AZAs) are a group of marine polyether biotoxins that were first identified in 1995 following a case of poisoning in the Netherlands in which people exhibited symptoms resembling diarrheic shellfish poisoning (DSP) after eating mussels harvested in Ireland [1]. Initially, toxin was known as “Killary toxin”, but after its structural characterization it was renamed azapiracid-1, AZA-1, to reflect its major structural features: a cyclic amino group, three spiro bonds and a carboxylic acid group [2]. Different AZA toxin analogs were discovered in 1997 and named AZA-2 and AZA-3, followed by the discovery of numerous analogs with a common structural backbone modified by the introduction of double bonds and methyl, hydroxy, and carboxy substituents at characteristic positions. At the moment, the total number of analogs appears to be 68 [3]. Some of these compounds are produced by phytoplankton and accumulate in shellfish, where they undergo structural changes as a result of shellfish metabolism. Given their potential impact on human health and the shellfish farming industry, azaspiracids have become the subject of intense research and ongoing monitoring in various regions of the world, including the Adriatic Sea.
Initially, it was believed that heterotrophic dinoflagellates of the genus Protoperidinium produced the azaspiracids. However, it was found that they only accumulate these toxins by feeding on phototrophic dinoflagellates [4]. This was confirmed in 2007 when the phototrophic dinoflagellate Azadinium spinosum was found and isolated in the northern Atlantic Ocean. A. spinosum was subsequently shown to produce the toxins AZA-1, AZA-2 and AZA-33 [5]. In addition to these toxins, A. spinosum is also known to synthesize AZA-11, AZA-34, AZA-35, AZA-50 and AZA-51 [6]. After the discovery of A. spinosum in the late 2000s and early 2010s, it was confirmed that other dinoflagellates from the Amphidomataceae family also produce azaspiracids. Research soon showed that the genera Azadinium and Amphidoma are more widespread than initially recorded and have been detected in many coastal waters of Europe, Asia and South America [5]. The dinoflagellate Azadinium poporum is a well-documented producer of azaspiracids (AZAs), with different strains having different AZA profiles depending on their geographical origin. This species has been found to produce AZA-2 as the main analog in strains from Argentina and the Mediterranean, while strains from Chilean coastal waters predominantly synthesize AZA-11 [7]. Other analogs such as AZA-36, AZA-37, AZA-40, AZA-41, AZA-42, AZA-59 and AZA-62 have been identified in different strains, further illustrating the biosynthetic variability of this species [6]. Another closely related species, Amphidoma languida, has been identified as a producer of azaspiracids, in particular AZA-2, AZA-38, AZA-39, AZA-43, AZA-52 and AZA-53 [8,9,10]. A recently described species, Azadinium dexteroporum, has been shown to synthesize several AZAs, including epi-AZA-7, AZA-35, AZA-54, AZA-55, AZA-56, AZA-57 and AZA-58 [11,12]. A comprehensive compilation of AZA produced by dinophlagelates from 2019 lists 26 variants, including two newly described variants AZA-42 and AZA-62 [6]. It should be mentioned that due to the extremely small cells of dinoflagellates, reliable detection methods must be used, which usually include PCR analysis of plankton and shellfish samples [13,14].
A number of studies have looked at the mechanisms of toxicity and have shown that azaspiracids can modulate the permeability of various ion channels (potassium, sodium, chloride and calcium). However, it is not yet entirely clear whether they bind directly to target proteins or act via intermediates [15]. Gastrointestinal symptoms (diarrhea, nausea, vomiting, abdominal pain) may also occur due to changes in the intestinal glial system and a weakening of the integrity of the intestinal barrier [16].
The structural diversity of AZAs arises from modifications such as methylation and hydroxylation, which occur predominantly at the carboxyl end of the molecule [6]. However, structural variations extend beyond this region, including oxidation, reduction, carboxylation and shortening of the carbon chain, which have been observed in analogs such as AZA-33, AZA-34 and AZA-35 [17]. Additionally, modifications in the nitrogen-containing I-ring contribute to this diversity. Most AZAs have a saturated I-ring that is methylated at the 39-position, resulting in a characteristic m/z 362 fragment. In some cases, the I-ring is unsaturated (m/z 360) or demethylated (m/z 348) [6].
Apart from the structural diversity due to differences in biosynthetic pathways, the potential biotransformation in shellfish presents a major challenge for AZA research. Primary forms such as AZA-1 and AZA-2 are enzymatically transformed to produce hydroxylated or methylated derivatives with altered polarity, toxicity and metabolic stability in marine organisms [18]. Despite this structural diversity, routine monitoring programs focus only on the sum of AZA-1, AZA-2, and AZA-3 as regulatory markers for seafood safety. The European Union and the United States have implemented a limit of 160 µg AZA eq/kg shellfish for the sum of AZA-1, AZA-2, and AZA-3 [19,20], as these are the most common analogs found in the human diet. However, emerging evidence suggests that other analogs, including AZA-6, AZA-11 and AZA-19, may also have significant toxic potential [21]. Understanding the distinct fragmentation patterns of AZAs is crucial for the accurate toxin identification and monitoring, particularly in the context of regulations that focus on the most toxic and prevalent analogs [4,6].
Monitoring of mussels from the Adriatic Sea for biotoxins is carried out in several key locations, including the Šibenik area and surrounding waters. While azaspiracids have been detected at lower levels in previous studies, the new research presented in this study provides an in-depth analysis of shellfish samples for the presence of various azaspiracid analogs, as well as the temporal distribution of regulated analogs following the January 2024 outbreak in Šibenik bay waters. Another focus is to use the identified suite of azaspiracid analogs to eventually draw a conclusion on a causative species behind produced toxins. Through the use of high-resolution mass spectrometry, this research aims to provide a detailed examination of the fragmentation patterns of detected analogues and potentially identify previously uncharacterized analogs of azaspiracids.
2. Results
2.1. Quantitative Determination of the AZA 2 Content
The AZA content was determined using standard method with reference material containing AZA-1, AZA-2 and AZA-3 as standards using a triple quadrupole mass spectrometer coupled to HPLC. The presence of AZA-1 and AZA-3 was not confirmed in any of the samples analyzed. AZA-2 was detected for the first time in the samples from S1, S2 and S3 on 02.01.2024. The highest AZA-2 content was detected on 02.01.2024 at sampling site S1 and on 07.01.2024 at sampling sites S2 and S3, followed by a gradual decrease in AZA-2 levels until 20.05.2024. Latter no AZAs could be determined in the samples from these sites (Figure 1).
2.2. Variability of Azaspiracids at Sampling Sites – Toxin Profiles
High resolution mass spectrometer Sciex 6600+ were used to detect AZAs with known precursor mass, structure and fragmentation pattern other than AZAs present in the standard material and quantitatively determined by triple quadrupole instrument. The TOF-MS method was used to screen the samples for all known precursors found in the literature. Extracted ion chromatograms (XIC) were prepared using precursors m/z found in the literature. Mass spectra of acquired XIC were manually investigated for m/z within 10 ppm of AZA precursors. The detected precursors were then fragmented using the SRM method to investigate the fragmentation pattern of the potential azaspiracids. Several analogs beside most dominant AZA-2 were detected. A total of 8 analogs were separately detected, differing in precursor mass, fragmentation pattern and RT. After analysis of their fragmentation pattern and comparison with data from the literature [2,6,8,9,12,17,22,23,24,25,26,27,28] seven of them were recognized as AZA-2, AZA-6, AZA-9, AZA-10, AZA-19, AZA-41 and AZA-43. A comparison of their exact and calculated masses is shown in Table 1 along with their RT [8,24].
Fragmentation of two analogs set to m/z 856.5 and 858.5 resulted in spectra containing clusters of three peaks differing by one m/z, indicating fragmentation of a compound containing two 13C atoms. These spectra were analyzed and assigned to the known analogs.
The analog with fragmentation sets to a precursor m/z of 884.5 had fragmentation pattern that was, to our knowledge, not recorded yet. The only analog recorded with such a precursor m/z was AZA-56 with an exact precursor m/z of 884.5535 [12] and fragments 866, 848, 830, 812 consistent with precursor water losses, group 2 fragments 690, 672, and 654, group 3 fragment 462, group 4 fragment 362, and group 5 fragment 262. Analog, we recorded, with RT 3.89 had exact m/z of 884.4928, fragments 866.4739 (precursor water loss), 840.4946 (precursor -CO2), 822.4769, 804.4679, 786.4602 (precursor-1-3H2O, -CO2), group 2 fragments starting at 700.3698, decarboxylated at 656.3778, followed by one and two water losses at 638.3677 and 620.3558. In group 3, fragment 490.2801 is detected with low intensity, while 446.2908, the product of its decarboxylation, is clearly seen. Group 4 and 5 ions 360.2521 and 260.1644 are detected, with the group 4 ion followed by a signal at 342.2416, due to the loss of one water. The group 6 ion with a m/z of 166.1220 is also detected with low intensity. The characteristic spectra of all analogs are presented in Figure 2. Due to a limited size of a pictures in a main text, presenting all relevant ions in legible size would result in a cluttered picture so spectra with larger pictures are also presented in a supplement Figure S1-S8 along with a table containing m/z of detected fragment ions, their elemental composition and deviation of detected ions from theoretical values (Table S1).
2.3. Temporal Dynamics of an Analog Occurrence
Selected samples were analyzed in detail to define the toxin profiles at the beginning of the outbreak, as determined by the amount of AZA-2 detected with the triple-quad instrument, and later, in the declining stages of the toxic episode. All detected analogs were present in all samples characterized by high toxin levels (samples from 02.01., 07.01. and 10.01.). In the samples with low AZA-2 toxicity from 24.01.2024 only AZA-6 was detected in the sample from site S3 in addition to AZA-2. In the samples with slightly increased toxicity collected on 29.01., 05.02. and 14.02. along with AZA-2, AZA-6 was detected in all samples, while AZA-19 was detected in station S1 on all dates, in station S2 on 14.02. and in station S3 on 05.02. and 14.02. AZA-41 was detected in station S2 on 29.01. Among samples with low AZA-2 content collected on 19.02., only the S3 sample contained also AZA-6 and AZA-19. In samples collected on later dates, AZA-6 was only determined on 10.04., while AZA-2 was also not detected in any samples from 20.05. and in samples from stations S1 and S2 from 06.05.

Table 2.
AZA analogs detected in various stages of toxic episode with an analog XIC area relative to AZA-2.
Table 2.
AZA analogs detected in various stages of toxic episode with an analog XIC area relative to AZA-2.
| Date and station | AZA-2 XIC area | Analog XIC area relative to AZA-2 (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| AZA-2 | AZA-6 | AZA-9 | AZA-10 | AZA-19 | AZA-41 | AZA-43 | 884,5 | |
| 02.01.2024. S1 | 1,30E+05 | 56,50 | 27,4 | 18,1 | 73,5 | 41,6 | 52,7 | 26,0 |
| 02.01.2024. S2 | 3,86E+05 | 29,20 | 13,7 | 8,9 | 32,8 | 60,8 | 51,5 | 11,1 |
| 02.01.2024. S3 | 4,30E+05 | 36,50 | 17,7 | 10,8 | 35,2 | 3,2 | 52,6 | 15,8 |
| 07.01.2024. S1 | 3,51E+05 | 19,90 | 12,5 | 7,9 | 25,2 | 65,1 | 50,5 | 6,0 |
| 07.01.2024. S2 | 6,04E+05 | 15,50 | 9,2 | 6,4 | 24,3 | 62,5 | 46,7 | 7,9 |
| 07.01.2024. S3 | 1,68E+05 | 28,40 | 13,7 | 10,1 | 42,6 | 60,4 | 48,6 | 9,5 |
| 10.01.2024. S1 | 2,26E+05 | 22,00 | 27,6 | 10,6 | 42,4 | 55,8 | 48,1 | 13,4 |
| 10.01.2024. S2 | 1,55E+05 | 15,30 | 17,5 | 7,6 | 52,8 | 77,2 | 69,9 | 22,6 |
| 10.01.2024. S3 | 1,46E+05 | 18,80 | 22,3 | 8,7 | 52,2 | 75,5 | 61,1 | 21,1 |
| 24.01.2024. S1 | - | - | - | - | - | - | - | - |
| 24.01.2024. S2 | - | - | - | - | - | - | - | - |
| 24.01.2024. S3 | - | - | - | - | - | - | - | - |
| 29.01.2024. S1 | 6336,45 | 130,30 | - | - | - | - | - | - |
| 29.01.2024. S2 | 16014,53 | 60,30 | - | - | - | 84,6 | - | - |
| 29.01.2024. S3 | 5318,2 | 146,60 | - | - | 99,3 | - | - | - |
| 05.02.2024. S1 | 10289 | 60,40 | - | - | 137,3 | - | - | - |
| 05.02.2024. S2 | 16049,5 | 32,90 | - | - | - | - | - | - |
| 05.02.2024. S3 | 16783,49 | 35,90 | - | - | 59,7 | - | - | - |
| 14.02.2024. S1 | 3128,26 | 174,70 | - | - | 241,0 | - | - | - |
| 14.02.2024. S2 | 3533,74 | 95,90 | - | - | 186,4 | - | - | - |
| 14.02.2024. S3 | 11010,7 | 28,30 | - | - | 56,5 | - | - | - |
| 19.02.2024. S1 | 1597,98 | 150,90 | - | - | 137,9 | - | - | - |
| 19.02.2024. S2 | 1989,23 | - | - | - | - | - | - | - |
| 19.02.2024. S3 | 1759,86 | - | - | - | - | - | - | - |
| 10.04.2024. S1 | 7349,2 | 100,60 | - | - | - | - | - | - |
| 10.04.2024. S2 | 7963,33 | 86,30 | - | - | - | - | - | - |
| 10.04.2024. S3 | 8438,44 | 92,70 | - | - | - | - | - | - |
| 15.04.2024. S1 | 5562,55 | - | - | - | - | - | - | - |
| 15.04.2024. S2 | 7380,33 | - | - | - | - | - | - | - |
| 15.04.2024. S3 | 7141,97 | - | - | - | - | - | - | - |
| 24.04.2024. S1 | 7174,9 | - | - | - | - | - | - | - |
| 24.04.2024. S2 | 12457,55 | - | - | - | - | - | - | - |
| 24.04.2024. S3 | 7901,95 | - | - | - | - | - | - | - |
| 28.04.2024. S1 | 5572,41 | - | - | - | - | - | - | - |
| 28.04.2024. S2 | 10633,13 | - | - | - | - | - | - | - |
| 28.04.2024. S3 | 6835,4 | - | - | - | - | - | - | - |
| 06.05.2024. S1 | 3958,01 | - | - | - | - | - | - | - |
| 06.05.2024. S2 | - | - | - | - | - | - | - | - |
| 06.05.2024. S3 | - | - | - | - | - | - | - | - |
| 20.05.2024. S1 | - | - | - | - | - | - | - | - |
| 20.05.2024. S2 | - | - | - | - | - | - | - | - |
| 20.05.2024. S3 | - | - | - | - | - | - | - | - |
3. Discussion
3.1. Variability of Azaspiracids at Sampling Sites – Toxin Profiles
The AZA-2 toxin was detected and quantified in samples from Šibenik Bay in January 2024 and its presence was followed until May, when no more toxin could be detected in the shellfish samples. High-resolution mass spectrometry was used to establish presence of other known analogs in addition to the regulated AZA-2. Eight different analogs were detected, seven of which were already known as AZA-2, AZA-6, AZA-9, AZA-10, AZA-19, AZA-41, and AZA-43, while eight that fragmented at a precursor set at 884.5 exhibited fragmentation spectra that, to our knowledge, could not be assigned to any known analog. All detected peaks are similar to the peaks of AZA-19 analog, only shifted by two Daltons to a lower m/z, indicating the introduction of a double bond in the smallest fragment region (rings H or I) of AZA-19.
Among large number of AZA analogs some differs in m/z for two Daltons resulting in possible misidentification of an analog with two 13C as a molecule with a two Dalton larger m/z as noted by Krock at al. 2014 [25]. Such a fragmentation spectrum resulting from 13C containing molecules could be recognized by a typical clusters of three peaks that differ by one m/z, a consequence of the distribution of 13C atoms in the fragments formed. In our case AZA-41 (m/z 854.5) with two heavy C atoms fragmented in the window of AZA-2 (m/z 856.5) and heavy AZA-2 fragmented in the window of AZA-9 and AZA-10 (m/z 858.5). Their characteristic spectra containing typical clusters of three peaks are presented in supplement Figure S9.
3.2. Origin of Detected Analogs
Among analogs detected, only AZA-2, AZA-41 and AZA-43 are of phytoplanktonic origin, the other analogs are products of shellfish metabolism [6,8]. Since no causative dinoflagellate has been confirmed in the water samples from the affected sites, the origin of the toxicity outbreak could only be speculated on the basis of the published data. Azaspiracids were detected in the Adriatic Sea for the first time in 2015 with no causative phytoplankton species being defined [29]. The only AZA producers detected so far in the Mediterranean Sea were A. dexteroporum in the Gulf of Naples and A. poporum in the waters of Corsica [12,30] and in the Ionian Sea [31]. AZAs that have so far been detected as toxins produced by A. dexteroporum include AZA-35, AZA-54 to AZA-58 and 3-epiAZA-7, none of which were detected in our samples. Different strains of A. poporum at different geographical locations produce AZA-2, AZA-11, AZA-36, AZA-37, AZA-40 to AZA-42, AZA-59 and AZA-62 [6,30]. Among them, AZA-2 and AZA-41 were detected in our samples. AZA-43 has so far only been detected as a product of Am. languida [8], but it cannot be excluded that it can be produced by a strain of A. poporum or be a product of shellfish metabolism. Other analogs detected in this study could be formed by biotransformations of AZA-2 and AZA-41. AZA-19 could be formed from AZA-2 by C-22 Me oxidation, it can be transformed to AZA-6 by C-22 decarboxylation. AZA-6 can be C-23 hydroxylated to form AZA-10, or it can be C-3 hydroxylated to form AZA-9 [32]. Among the toxins formed by shellfish transformation of AZA-2 in our samples, the most prominent form pertained in samples with less than higher AZA-2 content was AZA-6 and to a lesser extent AZA-19.
3.3. Temporal Dynamics of an Analog Occurrence
In attempt to quantitatively access contribution of analogs to overall amount of toxins present in shellfish sample their appearance and areas of survey scan extracted ion chromatograms of detected analogs were determined and compared in Table 2. Although, due to their different molar responses, precise comparison is not possible, it is informative to note that in the samples with high toxicity (AZA-2 content) no analog is present in amount that exceeds 77% of AZA-2. In high toxicity samples the most abundant analogs beside AZA-2, were AZA-19, AZA-41 and AZA-43. In low toxicity samples AZA-6 and to a less extent AZA-19 are present in amount that in some samples reach higher values than AZA-2. Both, AZA-6 and AZA-19, are product of biotransformation of AZA-2. This difference in the occurrence of the various analogs could reflect differences in the dynamics of toxin production or in the rates of bioaccumulation and depuration in shellfish or merely some toxins are present in lower quantities and fall below the detection limit earlier. Clarifying the dynamics of analog occurrence would require a method suitable for quantifying a range of different analogs.
3.4. Contribution of Unregulated Analogues to Total Toxicity
Different analogs vary in their toxicity, so that the transformation modifies toxicity for humans. Since only AZA-1, AZA-2 and AZA-3 are monitored as directed by regulatory authorities, some level of toxicity could remain undetected due to transformation. The contribution of the various analogs to the total toxicity is an important and as yet unresolved issue. Assessing the contribution of individual toxins to overall toxicity is complicated due to their different molar responses in the LC-MS/MS quantification method and their widely varying toxicity. There are only commercially available standards for AZA 1 – 3 that are regulated by the regulatory authorities. Using any of these standards to estimate the amount of other analogs is hampered by their different molar responses of either the parent ions or the fragments used to determine the amount of the compound in the extract. The relative response factors of AZA-4 to 10 differed significantly from AZA-1 under both isocratic and gradient conditions, and this difference was even more pronounced for the hydroxylated analogs [33]. The toxicity of the analogs could be compared using several methods, adding complexity to the assessment of overall toxicity. Using the Jurkat T-lymphocyte cytotoxicity assay for AZAs 1 - 10, relative toxicities were determined based on the EC50 in the range of 0.1 to 3.1 mM. It is worth noting that although AZA-2 was the most toxic, the analogs AZA-6 and AZA-8 were judged to be more toxic than AZA-3 and AZA-1, which are used by EFSA and FDA to determine AZA toxicity [33]. In another study, the same test was used to show that the analog AZA-34 with a shorter side chain at the carboxy end also have 5.5 times higher toxicity than AZA-1 [17]. The first in vivo study with intraperitoneal administration to mice showed toxicity equivalency factors (TEFs) of 1.0, 1.8 and 1.4 for AZA-1, AZA-2 and AZA-3, respectively, while another study yielded different TEFs of 1.0, 0.6 and 0.5 for the same toxins [2,22,34]. Orally administered toxins showed lower potency with TEFs of 1.0, 0.7 and 0.5, similar to more recent intraperitoneal tests [34]. Therefore, the overall toxicity and contribution of the detected analogs could only be roughly estimated. However, the presence of multiple analogs in our samples, as well as in most other samples, including some (AZA-6 in our samples) with significant toxic potential, suggests that the assessment of the toxic potential of azaspiracids may involve the quantification of more than three analogs.
4. Materials and Methods
4.1. Samples
The mussel samples (Mytilus galloprovincialis) were collected from January to May 2024 at three aquaculture sites, with a total of 60 samples collected during the study period. Samples were collected at regular weekly intervals, with one sample per site each week.
4.1.1. Sample Preparation for LC-MS/MS Analysis
The samples were prepared for analysis each week immediately upon arrival at the laboratory using liquid chromatography-tandem mass spectrometry (LC-MS/MS) method. Approximately 150 g of wet soft tissue was separated per sample and homogenized with a blender (22,000 rpm; 3 min). From the resulting homogenate, 2.00 ± 0.05 g of tissue was weighed and extracted with 9 mL of 100% methanol (MeOH). The mixture was vortexed for 2 min and centrifuged at 4500 x g for 10 min. The supernatant was transferred to a clean tube and the extraction was repeated by adding an additional 9 mL of 100% MeOH to the pellet. The supernatant from the second extraction was combined with the first and the final volume was adjusted to 20 mL with MeOH. The combined extract was then filtered through a 0.22 μm nylon syringe filter into vials for further analysis.
4.1.2. Certified Reference Materials
Certified reference materials (CRMs) for lipophilic toxins were obtained from the Institute of Marine Sciences, National Research Council (Halifax, Canada) for LC-MS/MS analysis. These included okadaic acid (OA), dinophysistoxin-1 (DTX-1), dinophysistoxin-2 (DTX-2), yessotoxin (YTX), 1-homoyessotoxin (hYTX), pectenotoxin-2 (PTX-2), azaspiracid-1 (AZA-1), azaspiracid-2 (AZA-2), azaspiracid-3 (AZA-3), gymnodimine A (GYM), 13-desmethylspirolide C (SPX-1) and freeze-dried mussel tissue (FDMT1) as reference material to verify method accuracy. To match the sample matrix, 0.35 g FDMT1 was weighed and 1.65 mL of water was added, whereupon the extraction procedure was identical to that described for the samples under “Sample preparation for LC-MS/MS analysis".
4.1.3. Detection of Lipophilic Toxins Using the LC-MS/MS Method
The regulated AZAs were detected and quantified using an LC (1100, Altium, Zagreb, Croatia) coupled to a triple quadrupole mass spectrometer (6410, Altium, Zagreb, Croatia) equipped with an electrospray interface. Chromatographic separation was performed on a C18 Poroshell 120 column (50 mm × 2.1 mm, particle size 2.7 μm, Altium, Zagreb, Croatia) coupled with a Poroshell 120 EC-C18 guard precolumn (5 mm × 2.1 mm, particle size 2.7 μm, Altium, Zagreb, Croatia).
The mobile phases, water (phase A) and 95% acetonitrile (ACN) (phase B), contained 2 mM ammonium formate (AF) and 50 mM formic acid (FA). Before mixing with ACN, AF was dissolved in 1 mL of water. A linear elution gradient was applied, progressing from 10% to 80% phase B over 4 minutes, held for 2 minutes, reduced to 10% phase B over 0.5 minutes and held for an additional 4.5 minutes to equilibrate the column for subsequent runs. The flow rate of the mobile phase was 0.3 mL/min, and the injection volume was 2 μL. The mass spectrometer operated in positive ionization mode for the detection of AZA-1, AZA-2 and AZA-3 with a spray voltage of 3.5 kV. The capillary temperature was maintained at 350 °C. Regulated AZAs were quantified by analyzing predetermined qualification and quantification fragments (dependent scan) listed in Table 3.
4.1.4. Characterization of AZA Toxins Using High Resolution Mass Spectrometry
An ultra-high performance liquid chromatography (UHPLC) (ExionLC AD series, Sciex, Framingham, MA, U.S.A.) was coupled to a high-resolution mass spectrometer (TripleTOF 6600+, Sciex, Framingham, MA, U.S.A.) equipped with a DuoSpray Ion Source operated in ESI mode and a Quadrupole Time-of-Flight mass analyzer controlled by Analyst TF 1.8.1. software. A Kinetex Core-Shell C18 column (100 × 2.1 mm, 2.6 µm, Phenomenex) coupled to SecurityGuard ULTRA Cartige C18, (Phenomenex) was used with a mobile phase A H2O and B ACN:H2O (95:5), both containing 50 mM FA and 2 mM AF. In the beginning (0 minutes) the mobile phase contained 15% phase B. During the first 2 minutes the composition changed to 65% phase B. This composition was maintained for up to 9 minutes. At 10 minutes, the composition returned to its initial state of 15% phase B and remained constant until the end of the run after 13 minutes. The flow rate was 0.3 mL/min and remained constant throughout the run. Samples were first analyzed in positive ionization mode using the Time-of-Flight Mass Spectrometry (TOF-MS) method with no fragmentation of precursor ions. The total ion chromatogram was then searched for all known AZA precursor ions available from the literature. The detected precursor ions were then analyzed with the Selected Reaction Monitoring High Resolution (SRMHR) method and manually examined for typical AZA fragmentation patterns. The column temperature was 30 °C and the injection volume was 5 µL. The source conditions were: Ion Spray Volatge Front (ISVF) = 5500 V, capillary temperature 425 °C, GS1 at 40, GS2 at 50 and Curtain Gas at 30 (arbitrary units), Collision Energy (CE) = 10 eV for MS experiments and CE = 65 eV for MS/MS experiments, Declustering Potential (DP) = 80 eV. The accumulation time for the TOF-MS method, MS experiment only, was 750 ms with a resulting cycle time of 775 ms, while the accumulation time for the SRMHR method was 100 ms for the TOF-MS scan and 150 ms for the MS/MS of selected ions. If necessary, to reduce cycle time, the precursor ions were grouped into three periods according to their RT as required, so that the cycle time for all experiments was approximately 2000 ms.
4.2. Study Area
The presence of marine biotoxins in farmed mussels is continuously monitored at several locations along the Adriatic coast. Monitored biotoxins include azaspiracids, with analogs AZA-1, AZA-2 and AZA-3 included in the monitoring program. Azaspiracids were detected in samples from shellfish farms near Šibenik in a sample collected on 02.01.2024. The level of azaspiracids was subsequently followed until it reached a very low level. The samples were collected from shellfish farms in three locations near Šibenik (Figure 3).
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1, Figure S1: title; Table S1: title; Video S1: title.
Author Contributions
“Conceptualization, S.O. and I.U.; methodology, A.B. and I.P.; validation, A.B., and T.B.; formal analysis, A.B. and I.P.; investigation, I.U.; resources, S.O. I.U. and T.B.; writing—original draft preparation, S.O. and I.P.; writing—review and editing, I.U. A.B. and T.B.; funding acquisition, I.U. All authors have read and agreed to the published version of the manuscript.”.
Funding
This work was supported by the Croatian Science Foundation under the project number HRZZ-IP-2024-05-9464. The work of doctoral student Antonija Bulić has been supported in part by the “Young researchers’ career development project – training of doctoral students” of the Croatian Science Foundation (DOK -2021-02-7058).
Institutional Review Board Statement
“Not applicable”.
Data Availability Statement
Dataset available on request from the authors.
Conflicts of Interest
“The authors declare no conflicts of interest.” “The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
Abbreviations
The following abbreviations are used in this manuscript:
| AZAs | Azaspiracids |
| DSP | Diarrheic Shellfish Poisoning |
| LC-MS/MS | Liquid Chromatography-tandem Mass Spectrometry |
| MeOH | Methanol |
| ACN | Acetonitrile |
| FA | Formic Acid |
| AF | Ammonium Formate |
| LOD | Limit of Detection |
| LOQ | Limit of Quantification |
| S/N | Signal to noise |
| CRMs | Certified Reference Materials |
| OA | Okadaic Acid |
| DTX-1 | Dinophysistoxin-1 |
| DTX-2 | Dinophysistoxin-2 |
| hYTX | 1-homoYessotoxin |
| PTX-2 | Pectenotoxin-2 |
| YTX | Yessotoxin |
| GYM | Gymnodimine A |
| SPX-1 | 13-desmethylspirolide C |
| FDMT1 | Freeze-Dried Mussel Tissue |
| RT | Retention Time |
| UHPLC | Ultra-High-Performance Liquid Chromatography |
| HPLC | High-Performance Liquid Chromatography |
| ESI | Electrospray Ionisation |
| TOF-MS | Time-of-Flight Mass Spectrometry |
| SRMHR | Selected Reaction Monitoring High Resolution |
| ISVF | Ion Spray Voltage Front |
| CE | Collision Energy |
| DP | Declustering Potential |
| EFSA | European Food Safety Authority |
| FDA | Food and Drug Administration |
| TEFs | Toxicity Equivalency Factors |
References
- McMahon, T.; Silke, J. Winter toxicity of unknown aetiology in mussels. Harmful Algae News, 1996, 14, 2. [Google Scholar]
- Satake, M.; Ofuji, K.; Naoki, H.; James, K.J.; Furey, A.; McMahon, T.; Silke, J.; Yasumoto, T. Azaspiracid, a new marine toxin having unique spiro ring assemblies, isolated from Irish mussels, Mytilus edulis. J. Am. Chem. Soc., 1998, 120, 9967–9968. [Google Scholar] [CrossRef]
- Sandvik, M.; Miles, C.O.; Løvberg, K.L.E.; Kryuchkov, F.; Wright, E.J.; Mudge, E.M.; Kilcoyne, J.; Samdal, I.A. In Vitro Metabolism of Azaspiracids 1-3 with a Hepatopancreatic Fraction from Blue Mussels (Mytilus edulis). J. Agric. Food Chem., 2021, 69, 11322–11335. [Google Scholar] [CrossRef] [PubMed]
- James, K. J.; Moroney, C.; Roden, C.; Satake, M. , Yasumoto, T.; Lehane, M.; Furey, A. Ubiquitous ‘benign’ alga emerges as the cause of shellfish contamination responsible for the human toxic syndrome, azaspiracid poisoning. Toxicon, 2003, 41, 145–51. [Google Scholar] [CrossRef] [PubMed]
- Tillmann, U.; Elbrächter, M.; Krock, B.; John, U.; Cembella, A.D. Azadinium spinosum gen. et sp nov (Dinophyceae) identified as a primary producer ofazaspiracid toxins. Eur. J. Phycol. 2009, 44, 63–79. [Google Scholar] [CrossRef]
- Krock, B.; Tillmann, U.; Tebben, J.; Trefault, N.; Gu, H. Two novel azaspiracids from Azadinium poporum, and a comprehensive compilation of azaspiracids produced by Amphidomataceae, (Dinophyceae). Harmful Algae 2019, 82, 1–8. [Google Scholar] [CrossRef]
- Tillmann, U.; Elbrächter, M.; Gottschling, M.; Gu, H.; Jeong, H.J.; Krock, B.; Nézan, E.; Potvin, E.; Salas, R.; Soehner, S. The dinophycean genus Azadinium and related species – morphological and molecular characterization, biogeography, and toxins. In: Kim, H.G., B. Reguera, G.M. Hallegraeff, C.K. Lee, M.S. Han and J.K. Choi. (eds). "Harmful Algae 2012, Proceedings of the 15th International Conference on Harmful Algae". International Society for the Study of Harmful Algae. 2014, 149-152.
- Tillmann, U.; Jaén, D.; Fernández, L.; Gottschling, M.; Witt, M.; Blanco, J.; Krock, B. Amphidoma languida (Amphidomatacea, Dinophyceae) with a novel azaspiracid toxin profile identified as the cause of molluscan contamination at the Atlantic coast of southern Spain. Harmful Algae 2017, 62, 113–126. [Google Scholar] [CrossRef]
- Krock, B.; Tillmann, U.; Voß, D.; Koch, B.P.; Salas, R.; Witt, M.; Potvin, E.; Jeong, H.J. New azaspiracids in Amphidomataceae (Dynophyceae). Toxicon 2012, 60, 830–839. [Google Scholar] [CrossRef]
- Tillmann, U.; Edvardsen, B.; Krock, B.; Smith, K.F.; Paterson, R.F.; Voß, D. Diversity, distribution, and azaspiracids of Amphidomataceae (Dinophyceae) along the Norwegian coast. Harmful Algae 2018, 80, 15–34. [Google Scholar] [CrossRef]
- Percopo, I.; Siano, R.; Rossi, R.; Soprano, V.; Sarno, D.; Zingone, A. A new potentially toxic Azadinium species (Dinophyceae) from the Mediterranean Sea, A. dexteroporum sp. nov. J. Phycol. 2013, 49, 950–966. [Google Scholar] [CrossRef]
- Rossi, R.; Dell’Aversano, C.; Krock, B.; Ciminiello, P.; Percopo, I.; Tillmann, U.; Soprano, V.; Zingone, A. Mediterranean Azadinium dexteroporum (Dinophyceae) produces six novel azaspiracids and azaspiracid-35: a structural study by a multi-platform mass spectrometry approach. Anal. Bioanal. Chem. 2017, 409, 1121–1134. [Google Scholar] [CrossRef]
- Toebe, K.; Joshi, A.R.; Messtorff, P. , Tillmann U., Cembella A., Uwe John U., Molecular discrimination of taxa within the dinoflagellate genus Azadinium, the source of azaspiracid toxins. J. Plankton Res. 2013, 35, 225–230. [Google Scholar] [CrossRef]
- Wietkamp, S.; Tillmann, U.; Clarke, D.; Toebe, K. Molecular detection and quantification of the azaspiracid-producing dinoflagellate Amphidoma languida (Amphidomataceae, Dinophyceae). J. Plankton Res. 2019, 41, 363–367. [Google Scholar] [CrossRef]
- Yang, J.; Sun, W.; Sun, M.; Cui, Y.; Wang, L. ; Current Research Status of Azaspiracids. Mar. Drugs 2024, 22, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Reale, O.; Huguet, A.; Fessard, V. Novel Insights on the Toxicity of Phycotoxins on the Gut through the Targeting of Enteric Glial Cells. Mar. Drugs 2019, 17, 429–453. [Google Scholar] [CrossRef]
- Kilcoyne, J.; Nulty, C.; Jauffrais, T.; McCarron, P.; Herve, F.; Foley, B.; Rise, F.; Crain, S.; Wilkins, A.L.; Twiner, M.J.; Hess, P.; Miles, C.O. Isolation, Structure Elucidation, Relative LC-MS Response, and in Vitro Toxicity of Azaspiracids from the Dinoflagellate Azadinium spinosum. J. Nat. Prod. 2014, 77, 2465–2474. [Google Scholar] [CrossRef] [PubMed]
- Hess, P.; McCarron, P.; Krock, B. Azaspiracids: chemistry, biosynthesis, metabolism, and detection. In Botana, L. M. (Ed.), Marine and Freshwater Toxins 2015, 763-788. CRC Press.
- Regulation (EC) No 853/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific hygiene rules for food of animal origin. OJ L 139, 30.4.2004, p. 55–205.
- Fish and Fishery Products Hazards and Controls Guidance. U.S. Department of Health and Human Services Food and Drug Administration Center for Food Safety and Applied Nutrition, 2022, (240) 402-2300.
- Botana, L. M.; Vilariño, N.; Alfonso, A. New insights into azaspiracid poisoning syndrome: toxicity and molecular targets. Toxins 2019, 11, 132. [Google Scholar]
- Ofuji, K.; Satake, M.; McMahon, T.; Silke, J.; James, K.J.; Naoki, H.; Oshima, Y.; Yasumoto, T. Two Analogs of Azaspiracid Isolated from Mussels,Mytilus edulis, Involved in Human Intoxication in Ireland. Nat. Toxins 1999, 7, 99–102. [Google Scholar] [CrossRef]
- Brombacher, S.; Edmonds, S.; Volmer, D.A. Studies on azaspiracid biotoxins. II. Mass spectral behavior and structural elucidation of azaspiracid analogs. Rapid Commun Mass Spectrom. 2002, 16, 2306–16. [Google Scholar] [CrossRef]
- Rehmann, N.; Hess, P.; Quilliam, M. A. Discovery of new analogs of the marine biotoxin azaspiracid in blue mussels (Mytilus edulis) by ultra-performance liquid chromatography/tandem mass spectrometry. Rapid Communications in Mass Spectrometry, 2008, 22, 549-558.
- Krock, B.; Tillmann, U.; Witt, M.; Gu, H. Azaspiracid variability of Azadinium poporum (Dinophyceae) from the China Sea. Harmful Algae 2014, 36, 22–8. [Google Scholar] [CrossRef]
- Krock, B.; Tillmann, U.; Potvin, É.; Jeong, H.J.; Drebing, W.; Kilcoyne, J.; Al-Jorani, A.; Twiner, M.J.; Göthel, Q.; Köck, M. ; Structure elucidation and in vitro toxicity of new azaspiracids isolated from the marine dinoflagellate Azadinium poporum. MarDrugs 2015, 13, 6687–702. [Google Scholar] [CrossRef]
- Tebben, J.; Zurhelle, C.; Tubaro, A.; Samdal, I.A.; Krock, B.; Kilcoyne, J.; Sosa, S.; Trainer, V.L.; Deeds, J.R.; Tillmann, U. Structure and toxicity of AZA-59, an azaspiracid shellfish poisoning toxin produced by Azadinium poporum (Dinophyceae). Harmful Algae 2023, 124, 102388. [Google Scholar] [CrossRef]
- James, K.J.; Sierra, M.D.; Lehane, M.; Braña Magdalena, A.; Furey, A. Detection of five new hydroxyl analogues of azaspiracids in shellfish using multiple tandem mass spectrometry. Toxicon 2003, 41, 277–283. [Google Scholar] [CrossRef]
- Bacchiocchi, S.; Siracusa, M.; Ruzzi, A.; Gorbi, S.; Ercolessi, M.; Cosentino, MA.; Ammazzalorso, P.; Orletti, R. Two-year study of lipophilic marine toxin profile in mussels of the North-central Adriatic Sea: First report of azaspiracids in Mediterranean seafood. Toxicon 2015, 108, 115–125. [Google Scholar] [CrossRef]
- Luo, Z.; Krock, B.; Mertens, K.N.; Nézan, E.; Chomérat, N.; Bilien, G.; Tillmann, U.; Gu, H. Adding new pieces to the Azadinium(Dinophyceae) diversity and biogeography puzzle: Non-toxigenic Azadinium zhuanum sp. nov. from China, toxigenic A. poporum from the Mediterranean, and a non-toxigenic A. dalianense from the French Atlantic. Harmful Algae 2017, 66, 65–78. [Google Scholar] [CrossRef]
- Luo, Z.; Krock, B.; Giannakourou, A.; Venetsanopoulou, A.; Pagou, K.; Tillmann, U.; Gu, H. Sympatric occurrence of two Azadinium poporum ribotypes in the Eastern Mediterranean Sea. Harmful Algae 2018, 78, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Kilcoyne, J.; McCarron, P.; Twiner, M.J.; Rise, F.; Hess, P.; Wilkins, A.L.; Miles, C.O. Identification of 21,22-dehydroazaspiracids in mussels (Mytilus edulis) and in vitro toxicity of Azaspiracid-26. J. Mol. Evol. 2018, 81, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Kilcoyne, J.; Twiner, M.J.; McCarron, P.; Crain, S.; Giddings, S.D.; Foley, B.; Rise, F.; Hess, P.; Wilkins, A.L.; Miles, C.O. Structure Elucidation, Relative LC-MS Response and In Vitro Toxicity of Azaspiracids 7-10 Isolated from Mussels (Mytilus edulis). J. Agric. Food Chem. 2015, 63, 5083–5091. [Google Scholar] [CrossRef] [PubMed]
- Kilcoyne, J.; Jauffrais, T.; Twiner, M.J.; Doucette, G.J.; Aasen Bunes, J.A.; Sosa, S.; Krock, B.; Séhet, V.; Nulty, C.; Salas, R.; Clarke, D.; Geraghty, J.; Duffy, C.; Foley, B.; John, U.; Quilliam, M.A.; McCarron, P.; Miles, C.O.; Silke, J.; Cembella, A.; Tillmann, U.; Hess, P. AZASPIRACIDS – toxicological evaluation, test methods and identifcation of the source organisms (ASTOX II) Technical Report Marine Institutem Galway 2014, p. 188 Ireland (2014) (ISSN:2009-3195).
Figure 1.
AZA-2 content in samples from Šibenik bay from January to May 2024 determined using the LC-MS/MS method, red S1, green S2 and orange S3.
Figure 1.
AZA-2 content in samples from Šibenik bay from January to May 2024 determined using the LC-MS/MS method, red S1, green S2 and orange S3.
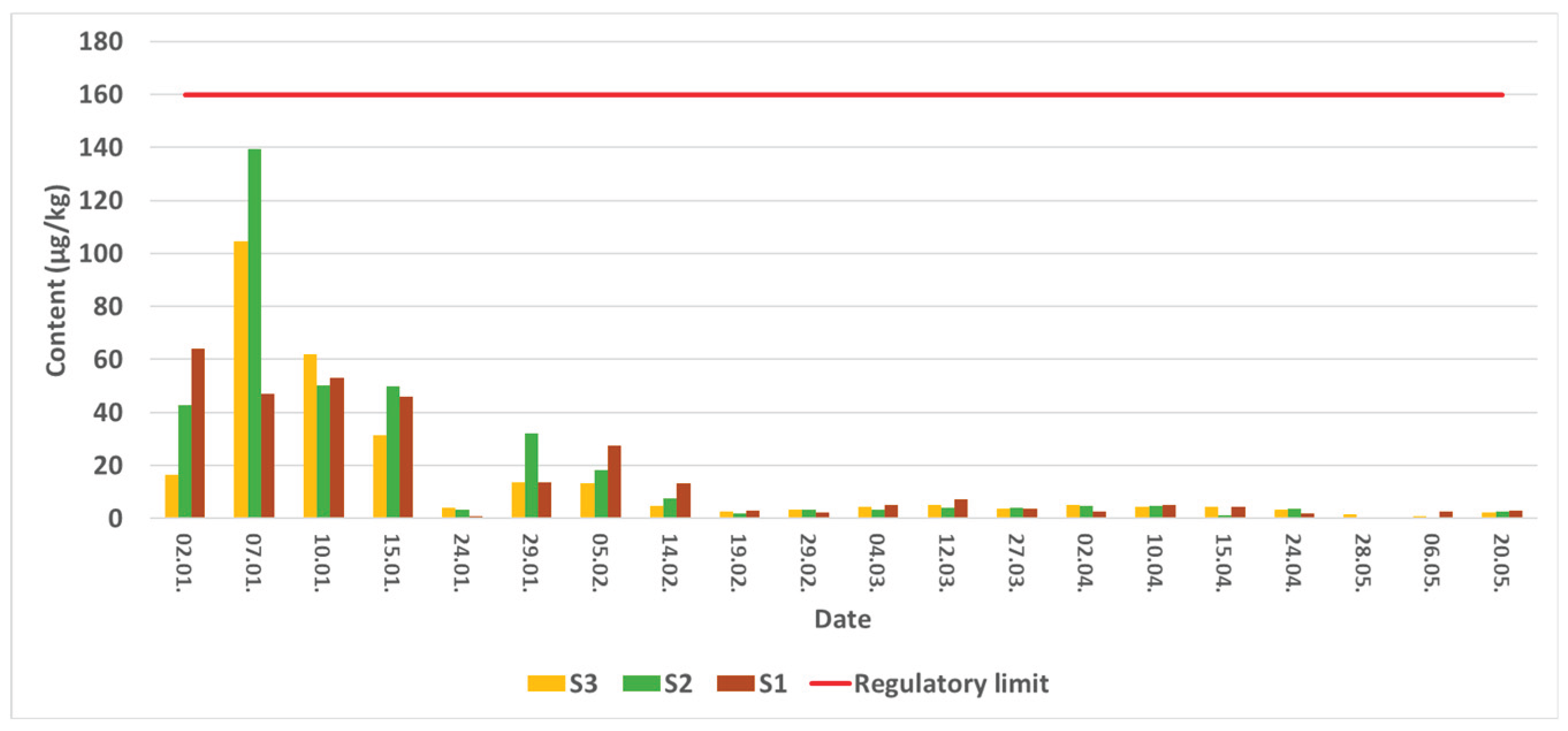
Figure 2.
Mass spectra acquired using high resolution QqTOF method of (a) AZA-2, (b) AZA-6, (c) AZA-9, (d) AZA-10, (e) AZA-19, (f) AZA-41, (g) AZA-43, (h) novel AZA 884.
Figure 2.
Mass spectra acquired using high resolution QqTOF method of (a) AZA-2, (b) AZA-6, (c) AZA-9, (d) AZA-10, (e) AZA-19, (f) AZA-41, (g) AZA-43, (h) novel AZA 884.
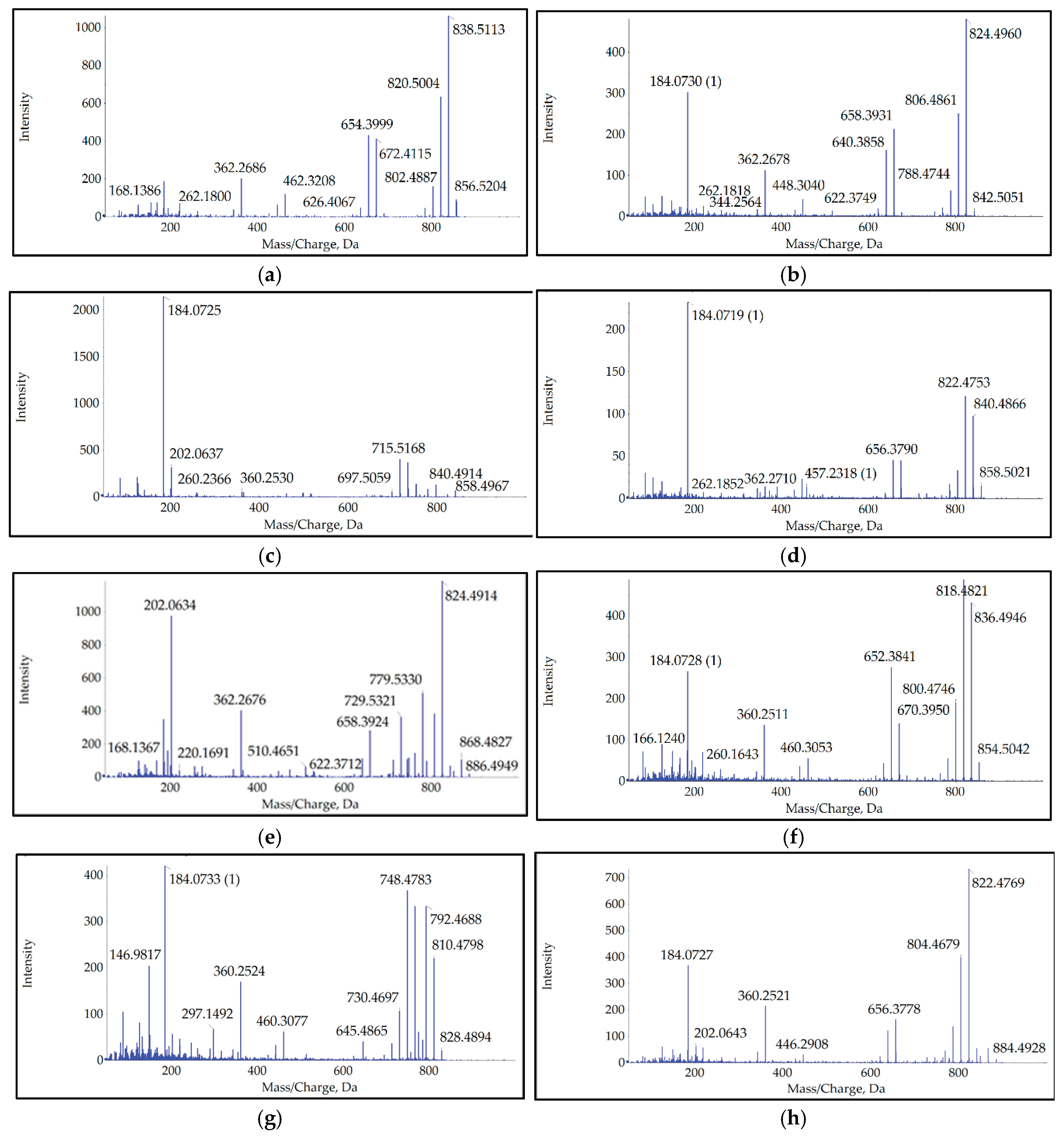
Figure 3.
Sampling stations in Šibenik bay, S1-Strmica, S2-Šibenik, S3-St Antony's Channel.

Table 1.
Exact mass data of detected analogues.
| Name | Exact mass | Calculated mass | Δ, ppm | RT, min |
|---|---|---|---|---|
| AZA-2 | 856.5205 | 856.5211 | -0.7 | 5.78 |
| AZA-6 | 842.5051 | 842.5055 | -0.5 | 4.83 |
| AZA-9 | 858.4967 | 858.5004 | -4.3 | 3.56 |
| AZA-10 | 858.5021 | 858.5004 | 2.0 | 3.91 |
| AZA-19 | 886.4949 | 886.4953 | -0.5 | 4.09 |
| AZA-41 | 854.5044 | 854.5050 | -0.7 | 5.36 |
| AZA-43 | 828.4894 | 828.4892 | 0.2 | 4.63 |
| 884.5 | 884.4928 | 884.4791 | 15.5 | 3.89 |
Table 3.
Precursor ions, product ions and method parameters for quantification of regulated AZAs.
| Compound name | Precursor ion (m/z) | Product ions (m/z) | Adduct | Collision energy (eV) | Fragmentor voltage (V) |
|---|---|---|---|---|---|
| AZA-1 | 842.5 | 824.5; 806.5 | [M+H]+ | 30; 42 | 230 |
| AZA-2 | 856.5 | 838.5; 820.5 | [M+H]+ | 30;42 | 210 |
| AZA-3 | 828.5 | 810.5; 792.5 | [M+H]+ | 28;42 | 210 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



