
Submitted:
16 September 2025
Posted:
17 September 2025
You are already at the latest version
Abstract
Background/Objectives: Recently, epigenetic mechanisms have been recognized as crucial in atopic dermatitis development. The emphasis of this research was on ex-panding existing knowledge about the epigenetic aspects of atopic dermatitis, as well as identifying new molecules that could serve as disease biomarkers. Methods: The re-search was conducted as a prospective study examining two groups: the group with atopic dermatitis (50 patients) and the control group (50 healthy adults). The serum levels of total immunoglobulin E (IgE) and eosinophil count (Eos%) were done in rou-tine laboratory analyses, and the detection of microRNAs from peripheral blood was performed using RT-PCR. Results: Analysis of selected miRNA expression in patients with atopic dermatitis and controls revealed that only the expression and the relative expression of miRNA-146a were statistically significantly higher in patients with atopic dermatitis than in the control group (p=0.042 and p=0.021, respectively). There was a weak positive correlation between miRNA-146a expression and the eosinophilia / IgE level (r=0.22 and r=0.25, respectively). The miRNA-21, miRNA-29b, miRNA-143 and miRNA-223 were significantly upregulated in patients with higher SCORAD (p< 0.001, p< 0.001, p< 0.001 and p= 0.015, respectively). ROC curve analysis revealed the specificity of miRNA-146a as 82% and the sensitivity as 62%. The area under the ROC curve (AUC) was 0.7, confirming a valid diagnostic potential. Conclusions: Our find-ings imply that miRNA-146a may be a novel biomarker of atopic dermatitis and might play a specific role in the development of the disease, while miRNA-21, miRNA-29b and miRNA-223 may have an impact on the disease progression. Our study provides findings that can be utilized in future diagnostic applications, targeted personalized treatments and support the monitoring of atopic dermatitis treatment effectiveness.
Keywords:
atopic
; epigenetic
; biomarkers
; miRNA
1. Introduction
Atopic dermatitis (AD) is a multifactorial, inflammatory skin disease characterized by typical clinical manifestations, including itching and a chronic-relapsing course [1,2,3,4,5]. Atopic dermatitis has a wide range of clinical presentation depending on the stage of the disease, the patient’s age and the severity of the skin lesions [2,3,4]. In the acute phase, AD lesions occur as erythematous, exudative papules and plaques, with secretion, vesicles and blisters, leaving erosions and crusts [2,4,5]. In further progression, due to inflammation, scratching and rubbing, these lesions evolve into chronic thickened, dry and lichenified plaques resulting in skin dyspigmentation [2,4,5]. Characteristic adult forms of AD include ’head-and-neck’ and ’portrait dermatitis’ affecting periorificial region, face, neck and upper trunk, hand eczema susceptible to concomitant occupational contact dermatitis, extensive skin xerosis in elderly patients, or erythrodermic forms that must be distinguished from cutaneous T-cell lymphoma [2,3,4,5,6,7]. Some AD patients may experience seasonal exacerbations, accompanied by allergic rhinoconjunctivitis or allergic asthma [2]. This is consistent with the concept of ’atopic march’ in which patients with atopic dermatitis are prone to the production of allergen-specific IgE and the development of other forms of atopy [2,8,9,10,11].
The pathogenesis of AD is complex and involves interaction between numerous genetic factors, environmental agents and dysfunctional immunologic response [4,5,12]. Genetic research has revealed that the disruption of the skin barrier plays an essential role in the development of the disease [1,2,4,5,13,14,15,16,17]. Abnormalities in the production of filaggrin, ceramides and antimicrobial peptides result in skin barrier dysfunction, early onset of disease, more extensive lesions and a greater number of comorbidities [1,2,4,5,13,14,15,16,17]. On the other side, from an immunological aspect, it is widely accepted that the basis of atopic dermatitis lies in a type 2 inflammatory response [1,2,4,5,13,14,15,16,17]. Due to the epidermal impairment with increased susceptibility to pathogens and environmental substances, stimulated keratinocytes secrete TSLP, IL-25 and IL-33. These alarmins activate ILC2 cells, which then produce IL-4, IL-5and IL-13, that drive the differentiation and proliferation of inflammatory cells, Th2 and eosinophils [5,18,19,20,21]. Numerous studies have shown that inflammation mediated by Th2-lymphocytes is a key phenomenon in the development of immune dysfunction responsible for the occurrence of the acute phase of atopic dermatitis, while in chronic skin lesions, Th1-lymphocytes derived inflammation with gamma interferon (γIFN) and interleukin 12 (IL-12) is predominant [1,2,4,5,13,14,15,16,17]. In addition, some patients also express a Th17/Th22 immune response over time [1,2,4,5,13,14,15,16,17]. Therefore, elevated levels of Th-lymphocyte subpopulations (Th2, Th9, Th17, Th22) and specific interleukins (IL-4, IL-5, IL-9, IL-13, IL-22, IL-31) were observed in patients with atopic dermatitis [1,2,4,5,13,14,15,16,17].
Recent research has focused on specific types of epigenetic mechanisms that may contribute to AD disease development [13,15,18,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48]. Epigenetic modifications encompass reversible enzymatic processes of DNA-methylation, histone modification and posttranscriptional noncoding RNAs’ regulatory role [33,34]. These mechanisms exert epigenetic alterations on key pathogenetic events in AD, as activation of the immune response, T-cell polarization and keratinocytes dysfunction [33,34]. Micro RNAs (miRNAs) are short non-coding RNAs (19-25 nucleotides) e that regulate post-transcriptional gene expression [1,8,45,46,47,49]. According to available data, several miRNA molecules might be involved in epidermal regulation - increased levels of miRNA-203 and miRNA-184 are associated with dysregulated expression of filaggrin, involucrin and loricrin, while miRNA-194-5p is involved in the regulation of gene expression of structural proteins - LOR, KRT4, FLG2 and heparan sulfate - HS3ST2 [22,33,35]. Additionally, dysregulated levels are detected for miRNA-10a-5p and miRNA-26a-5p that are involved in the regulation of HAS3 (hyaluronan synthase 3), leading to inadequate keratinocyte differentiation and migration [18,33,34,40,41,44]. The epidermal dysfunction in AD may also be substantially due to miRNA-29b promoting keratinocyte apoptosis via inhibition of Bcl2L2 (Bcl-2-like protein 2) [27]. It was demonstrated that miRNAs play a regulatory role in immune response, especially in the pathogenesis of inflammatory diseases such as atopic dermatitis [1,8,15,24,25,26,28,29,30,31,32,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53]. In AD, certain miRNAs molecules exhibit potentially immunosuppressive activity, such as miRNA-124, miRNA-1294, miRNA-205 and miRNA-146a via regulating crucial parts of NF-κB pathway in keratinocytes and different immune cells, or miRNA-143 via targeting IL-13Rα1 in keratinocytes silencing IL-13-mediated decreased expression of filaggrin, involucrin and loricrin [18,24,25,26,34,36,40,43,44]. Concerning proinflammatory miRNAs, miRNA-144 may have been of high importance in AD by regulation of hBD-2 and SERPINB4 - NF-κB activators, while decreased levels of let-7a-5p in AD relate to overexpression of CCR7 on T-cells and DCs in atopic patients [34,40]. In the context of T-cell differentiation, several miRNAs are particularly important. For instance, miRNA-155 expression was found to positively correlate with the percentage of Th17 cells, levels of IL-17 and RORγ (retinoic acid-related orphan receptor), but negatively with SOCS1 (signaling inhibitor suppressor of cytokine signaling-1) and CTLA-4 (cytotoxic T lymphocyte-associated antigen 4) [28,29,30,34]. Additionally, miRNA-151a may have an inhibitory effect on IL12Rβ2/JAK/STAT included in Th1 differentiation, while miRNA-191 affects Treg cells activation by targeting SATB1 (AT-rich sequence-binding protein 1) [31,34,43]. Lastly, miRNA-302e and miRNA-135 showed the antiallergic effects via inhibiting NF-κB and GATA-3, respectively, resulting in suppression of Th2 differentiation and cytokine production. [39] In terms of the epigenetic profile of lesional AD, overexpression of miRNA-223 in AD lesions is associated with T-cell regulation and subsequent inflammation [18,40,44]. On the other hand, miRNA-21 presented a protective role through inhibition of the p38 pathway with subsequent suppression of mast cell degranulation, skin inflammation and scratching behavior in an animal model [39].
All the previously mentioned highlights the significant role of miRNAs in modifying allergic inflammation in atopic diseases, through regulation of the production of proinflammatory and anti-inflammatory molecules and cell biology, resulting in disruption of homeostasis and the eventual development of atopic dermatitis. With the recent introduction of novel targeted therapies in the treatment of atopic dermatitis, the importance of modern specific biomarkers and stratification of patients has been highlighted with the ultimate goal of the implementation of a personalized, medicine. 54, 55]. MiRNAs are molecules that are stable in various biological samples, serum or other body fluids, with exceptional potential as an ideal non-invasive biomarker [1,14,56,57,58,59]. The identification of miRNAs could enhance the accuracy of early diagnosis or, especially in cases with discordance between the clinical presentation and somewhat unreliable routine laboratory parameters, to reveal distinct endotypes of AD and/or predict treatment outcome [1,14,44,56,57,58,59]. It has been shown that certain miRNAs expressed highly dysregulated levels in atopic patients compared to healthy controls [1,8,14,18,40,41,42,43,44,45,46,47,56,57,59]. This brings a novel perspective of their further implementation in clinical practice and the creation of more sophisticated therapeutic modalities [40,44,58,60,61]. Addressed by the need for more precise diagnostics in atopic dermatitis, we managed to identify miRNA molecules that may serve as biomarkers for diagnosis or disease severity, independent of traditional laboratory parameters.
2. Materials and Methods
The research was conducted as a prospective study at the Clinic of Dermatovenereology, University Clinical Centre of Vojvodina, Serbia, with the recruitment period from April 28th, 2022 to December 6th, 2023. Two groups were formed: 1. The control group (C), consisted of 50 patients who fulfilled the required inclusion criteria (adults without any form of atopy such as atopic dermatitis, asthma, allergic rhinitis, food allergy, or any other systemic disease and have not been under any immunosuppressive therapy in the last two months); 2. The experimental group (AD), consisted of 50 patients who presented either as previously or newly discovered cases of atopic dermatitis and who fulfilled the inclusion criteria (atopic dermatitis with/without any other form of atopy but without any other systemic disease and have not been under any immunosuppressive therapy in the last two months).
The exclusion criteria were: patients younger than 18 years, patients who had been under any immunosuppressive therapy in the last two months, pregnant women, and patients with any kind of systemic disease.
All participants signed the informed consent and the research was approved by the Ethics Committee of the Clinical Centre of Vojvodina (protocol number #600-210).
The clinical examination of patients was performed independently by two dermatologists and the assessment of the extent and intensity of atopic dermatitis was done using the SCORAD index (SCORing Atopic Dermatitis). SCORAD index was interpreted as follows: final score less than 25 presented as mild AD, between 25 and 50 as moderate AD, and more than 50 as severe AD.
The serum levels of total immunoglobulin E (IgE) and eosinophil count (Eos%) were measured for each patient as routine laboratory analyses.
The detection of microRNAs was performed by a specific procedure: patients’ peripheral blood samples were collected in test tubes with a reagent for stabilization of cell RNAs (PAXgene® Blood RNA Tubes (Becton, Dickinson and Company, Eysins, Switzerland); extraction of total microRNA was performed using TaqMan® miRNA ABC Purification Kit (Thermo Fischer, Nerum, Denmark), followed by reverse transcription of microRNAs of interest using TaqMan® MicroRNA Reverse Transcription Kit and specific primers (Thermo Fischer, Nerum, Denmark), following the manufacturer's instructions. The exact primer sequences were proprietary and not provided by the manufacturer. According to previously published data, as explained in the introductory section, seven miRNAs of high importance in atopic diseases were selected. Amplification and analysis of selected microRNAs: 21, 26a, 29b, 203, 223, 143 and 146a was carried out using real-time PCR (ABI Prism 7000 Sequence Detection System, Applied Biosystems, Foster City, CA, USA) and TaqMan® MicroRNA Assay reagents (Thermo Fischer, Nerum, Denmark). All reactions were carried out according to the manufacturer’s protocols.
Data analysis was performed using SPSS software, version 23.0 (Chicago, IL, USA). Quantitative data were tested for normal distribution using the Kolmogorov–Smirnov test. Normally distributed data were compared using the Student’s t-test and nonparametric data were analyzed with the Kruskal-Wallis test. Parameters were further analyzed using binomial and multinomial logistic regression. The P-value of <0.05 indicated a statistically significant difference.
3. Results
3.1. Patient Dataset
The study included the total number of 100 participants - 71 (71%) female and 29 (29%) male participants with average age of 34.5± SD year (range 18-73 y), divided in the control group with 50 participants ( 35 (70%) female, 15 (30%) male) and experimental group with 50 participants ( 36 (72%) female, 14 (28%) male) with the disease duration (AD) of at least two weeks (average duration7.22 years): 13 patients <1y, 20 patients >1-5y, five patients >5-10y; 12 patients >10y among which eight patients have had AD since childhood. The experimental group was further divided in subgroups involving the patients with only atopic dermatitis - 22 (44%); atopic dermatitis and allergic rhinitis - 16 (32%); atopic dermatitis, allergic rhinitis and asthma - 7 (14%); atopic dermatitis, allergic rhinitis and food allergy – 4 (8%); atopic dermatitis and food allergy - 1 (2%).
The experimental group (AD) included six patients with SCORAD<25 (12%), 26 patients with SCORAD >25-50 (52%), and 18 patients with severe atopic dermatitis and SCORAD>50 (36%). IgE levels ≤100 IU/ml were detected in 12 patients (24%), IgE levels <100-500 IU/ml were found in 18 patients (36%), while the remaining 20 patients expressed IgE ≥500 IU/ml (40%). In this group, moderate hypereosinophilia was found only in one case (values 1.5–5.0 × 10^9 /L) (2%), while mild eosinophilia was detected in eight patients (values 0.5–1.49 × 10^9 /L) (16%).
In the control group, IgE levels of ≤100 IU/ml were predominantly detected, in 39 participants (78%). IgE levels <100-500 IU/ml were detected in 11 participants (22%), while higher levels of IgE (>500 IU/ml) were not registered. A mild absolute eosinophilia was assessed in two participants (0,51 and 0,54 × 10^9 /L) (4%), while the absolute eosinophil count in the rest of the group was in the physiological reference interval.
Descriptive table detailing the characteristics of the 100 patients included in the study is given below (Table 1).
3.2. MiRNAs, Serum IgE Levels and Eosinophils in the Experimental - AD and Control Group – C
The seven microRNAs (miRNA – 21, 26a, 29b, 143, 146a, 203, 223) were analyzed in the whole blood samples from 50 patients with AD and 50 controls by real-time PCR (Figure 1). The statistics was performed using SPSS software version 23.0 (Chicago, IL, USA). p-value <0.05 indicated a statistically significant difference.
The quantitative data were presented as means, with 95% confidence intervals (CI) calculated for each group (Figure 1), checked for normal distribution using Kolmogorov – Smirnov test, and further analyzed by two-tailed Student’s t-test.
The expression of analyzed miRNA in patients with AD and those from the control group were compared, and only miRNA-146a was found to be significantly upregulated in AD patients compared to controls (p=0.042), while miRNA-26a, miRNA-21, miRNA-29b, miRNA-223, miRNA-143 and miRNA-203 did not show significantly different expression between groups (p= 0.585; p= 0.573; p= 0.665; p =0.717; p= 0.630; p= 0.739, respectively). Figure 1 presents the mean Ct values for each miRNA across the AD and control groups, with error bars indicating the 95% confidence intervals.
The relative expression of each miRNA was expressed as ΔCt, calculated as the difference in the threshold cycle (Ct) between the miRNA of interest and the miRNA-26a. miRNA-26a was used for the normalization of expression data because of the previously demonstrated similar expression among groups. Comparing all given values in both research groups revealed that, although all analyzed miRNAs were upregulated in the AD group, only the relative expression of miRNA146a was noticeably higher (p= 0.021) (Figure 2).
Given the interpretation that Ct values of miRNA above 30 are classified as weak positive, and below 30 as strong positive, and after comparing of those results by the implementation of the Kruskal-Wallis test, again, only miRNA-146a was identified with considerably higher expression in the AD group than in control group (p= 0.046) and therefore selected for further analysis. miRNA-146a was found to be significantly upregulated in AD patients compared to controls (p=0.046), while no other miRNAs showed statistically significant differences between the groups.
The level of IgE in the blood was substantially higher among patients with AD than in the control group (p< 0.001). Measured parameters in both research groups were additionally evaluated using binomial logistic regression, resulting in the following: the percentage of eosinophils was significantly higher in the AD group than among controls (p= 0.012) (Figure 3); the IgE level was markedly higher in the AD group than in the control group (p< 0.001) (Figure 4) and the miRNA-146a was considerably more expressed in the AD group than in the control group (p= 0.041). There was a significant, but weak positive correlation between miRNA-146a expression and the eosinophilia, and between miRNA-146a expression and IgE level (r= 0.22; p= 0.025 and r= 0.25; p= 0.013, respectively).
Assessing the correlation between measured parameters in the blood (IgE and Eos) and SCORAD resulted in the following: the eosinophil percentage was in significant correlation with SCORAD (r= 0.36; p< 0.001); as was the IgE level (r= 0.32; p= 0.025). There was also a moderate positive correlation between miRNA-223 expression and SCORAD (r= 0.342, p= 0.015) as well as between miRNA-146a expression and SCORAD (r= 0.191, p= 0.05). (Figure 5).
Additionally, multinomial logistic regression was performed to adjust for confounding factors (age, gender, disease duration, presence of allergic rhinitis (AR), asthma and food allergy), resulting in the following findings: the percentage of eosinophils was considerably higher in patients with higher SCORAD (p< 0.001); the miRNA-21, miRNA-29b and miRNA-143 were significantly more expressed in patients with higher SCORAD (p< 0.001 for all three miRNAs; Figure 6; Figure 7 and Figure 8, respectively)



MiRNA-146a diagnostic potential was evaluated based on ROC curve analysis (receiver operating characteristic curve). The area under the ROC curve (AUC) was used to establish the diagnostic applicability of miRNA-146a in AD patients. The optimal diagnostical result was estimated by comparing given values to borderline values with the highest Youden’s index (sensitivity +1, specificity -1). ROC curve analysis revealed that the specificity and sensitivity of miRNA-146a were 82% and 62%, respectively. The area under the ROC curve (AUC) was 0.7, which confirms that miRNA-146a has a valid diagnostic potential in AD (Figure 9). Therefore, miRNA-146a may be an essential biomarker of AD.
4. Discussion
4.1. Atopic Dermatitis
Atopic dermatitis (AD) is a skin inflammatory disease characterized by pruritus and typical eczematous cutaneous lesions that vary through acute, subacute and chronic phases. It affects approximately 20% of children and 2-10% of adults [1,56]. Additionally, 40-60% of patients with atopic dermatitis also have another form of atopy, such as asthma, allergic rhinitis or food allergies [1]. Our cohort showed a similar distribution of atopic comorbidities: more than half of the AD patients presented at least one of the other atopic manifestations, with allergic rhinitis being the most prevalent among them (54%).
AD might be classified in a mild, moderate or severe form due to the SCORAD index, interpreting SCORAD <25, SCORAD <25-50, and SCORAD >50, respectively [62,63,64]. Diagnostic criteria of AD established by Hanifin and Rajka include increased total serum IgE level, but with variable data in different studies. Some research has identified IgE levels as a reliable indicator of AD, while in others, the opposite was observed [4,63,65,66,67,68]. In our study, the level of total IgE in the blood among the AD group was statistically significantly higher than in the control group (p<0.001), and IgE level was positively correlated with SCORAD (p= 0.025). Another laboratory parameter that can be used in differentiating extrinsic and intrinsic AD, or may correlate with the degree of AD severity, is the blood eosinophil count [69]. A peripheral eosinophilia, either relative or absolute, might be seen in diverse allergic conditions, including atopic dermatitis. In our study, the percentage of eosinophils was significantly higher in the AD group than in healthy controls (0.012) and in significant correlation with SCORAD (r= 0.36; p< 0.001), as it was also considerably higher in patients with higher SCORAD (p< 0.001), which is in the accordance with literature data.
4.2. miRNAs
MiRNAs are small molecules consisting of 19-25 nucleotides that belong to non-coding RNAs [1,8,45,46,47,49]. Their exceptional role in the regulation of posttranscriptional gene expression has been recognized as an essential factor in immunity and pathogenesis of various inflammatory diseases, such as atopic dermatitis [1,8,13,18,40,43,44,45,50,51,52,53]. It was established that certain microRNAs were present in patients with atopy, or their levels significantly deviated from the values obtained in subjects who did not have any form of atopic diseases [1,8,18,44,45,47]. For our study, we selected seven miRNAs for further investigation based on previous publications [1,8,14,18,34,40,41,42,43,44,45,46,47,50,51,52]. Our results revealed that only miRNA-146a relative expression was statistically significantly higher in AD in comparison to the C group values (p=0.021). After performing ROC curve analysis, the diagnostic potential of miRNA-146a was validated (Figure 9). Recent studies on atopic dermatitis established the overexpression of miRNA-146a in the skin lesions and blood of AD patients [8,23,25,26,40,41,43,44,46,52,75]. Our results support available data that miRNA-146a might be a promising diagnostic biomarker for atopic dermatitis (p= 0.021, AUC= 0.7), indicating its role in the pathogenesis of the disease. Experimental studies revealed potential mechanisms through which miRNA-146a may regulate innate and adaptive immunity, antibody production, inflammation, haemopoiesis, differentiation and proliferation of keratinocytes [25,26,40,43,52,71,76,77,78]. Thus, miRNA-146a is involved in oncogenesis, pathogenesis of autoimmune disease and allergic response [78]. The studies on miRNA-146a expression revealed its clinical implications, serving as a valuable diagnostic and prognostic biomarker, while different models demonstrated molecular mechanisms underlying the development of various diseases [78]. It was found that miRNA-146a controls suppression of TNF-mediated osteoclastogenesis in rheumatoid arthritis (RA), while the decreased expression of miRNA-146a due to the rs2431697 variant in the miRNA-146a gene in systemic lupus erythematosus (SLE) can lead to altered IFN signaling and progression of the disease [78]. Conversely, one study detected a protective role of single-nucleotide polymorphism (rs2910164) in the gene encoding precursor of miRNA-146a in the early onset of psoriasis [77]. The authors proposed that miRNA-146a overexpression resulted in ameliorating IL-17-mediated inflammation, decreased neutrophil influx and regulation of keratinocyte hyperproliferation in psoriasis [77]. While EGFR has been verified target for miRNA-146a, another study identified FERMT1 as a psoriasis-related miRNA-146a target gene involved in keratinocyte proliferation [71,77]. About miRNA-146a regulatory role in cell biology and differentiation, available data indicate that Treg cells exert the capacity of suppressing CD4+CD25- T cells by the change of their intracellular miRNA-146a levels, while the Th2-differentitation can be inhibited via miRNA146a targeting SERPINB2 (serpin family B member 2) [26,71,79,80]. Additionally, miRNA-146a may be responsible for B cell class switching and increased production of IgE antibodies via upregulation of 14-3-3σ expression in the asthma animal model [76]. Research conducted on the miRNA-146a-deficient mice suggested the existence of a link between serum IgE level and the expression of miRNA-146a, while patients’ serum miRNA-146a levels were independently associated with IgE levels in AD, without correlations to the severity of AD [75]. In our research, a weak correlation was observed between miRNA-146a and IgE levels (r= 0.25, p= 0.013) in the AD cohort, while its correlation with SCORAD was moderate (r= 0.191, p= 0.05). Conversely, in a study on allergic conjunctivitis, the authors indicated the inverse correlation between miRNA-146a and serum IgE levels [79].
MiRNA-146a inhibits the production of proinflammatory molecules ubiquitin D, CCL5 and CCL8, and suppresses their expression in epidermal keratinocytes, but also adjusts the activity of B-cells, T-lymphocytes, mast cells, monocytes and dendritic cells [14,40,41,44,46,81,82]. Both tissue culture and animal models demonstrated that miRNA-146a is involved in the NF-κB pathway, targeting the upstream mediators- IRAK1 (IL-1 receptor-associated kinase 1), CARD10 (caspase recruitment domain containing protein 10) and TRAF6 (tumor necrosis factor receptor-associated factor 6) ameliorating the chronic inflammation [14,18,25,34,42,43,46]. Downregulation of adapter proteins IRAK1 and TRAF6 causes reduced activation of NLRP3, inhibition of TLR-signaling and in the final step leads to the significant reduction of IL-6, IL-18, IL1β and αTNF release [78,83]. To our knowledge, miRNA profile of eosinophils does not express significant miRNA-146a levels nor the correlation to blood eosinophil number was detected [84,85,86,87,88], as it was demonstrated for miRNA-21 and miRNA-223 to be involved in eosinophil biology: lower expression of miRNA-223 correlates to increased levels of eosinophil progenitors and eosinophil differentiation via targeting IGF1R [18,40,44,89], while miRNA-21 also play a role in IL-5-mediated eosinophil differentiation via direct regulation on eosinophil progenitors [39,90]. In our study, a weak but significant positive association between the miRNA-146a expression and eosinophils was detected (r= 0.22; p= 0.025), suggesting the potential indirect biological relation between upregulated miRNA-146a and eosinophilia. Several allergic models demonstrated miRNA-146a's direct impact on ILC2 cells' function and proliferation via IL-33/ST2 signaling through the negative regulation of IRAK1 and TRAF6, leading to a decreased release of IL-5 and IL-13 and subsequent amelioration of eosinophil inflammation [91,92]. Similarly to previous, in the allergic rhinitis model (AR) and β-lactoglobulin-induced food allergy, miRNA-146a showed anti-inflammatory potential by inhibiting the TLR4/TRAF6/NF-κB pathway [93,94]. For instance, in AR study model, administration of miRNA-146a mimic into nostril resulted in decreased levels of OVA-specific IgE, LTC4, PGD2, ECP, IL-4, IL-5 and IL-13, as well as decreased number of mast cells and reduced infiltration of eosinophils, lymphocytes and neutrophils, while converting Th1/Th2 imbalance and preventing IgE-mediated response [93]. Another experiment proposed that miRNA-146a inhibitors might lessen asthmatic inflammation via upregulation of the TLR2-signaling pathway in monocytes [95]. Lastly, in animal models of allergic conjunctivitis, pollen induced downregulation of miRNA-146a was followed by enhanced expression of TSLP/TSLPR/OX40L/CD11C molecules, while the other investigation revealed that overexpression of miRNA-146a induced upregulation of FOXP3, one of the essential transcription factors in promoting Treg cells, and further suppression of inflammation through targeting HIPK3 (homeodomain-interacting protein kinase 3) and reduced phosphorylation of STAT3 [96,97].
It is noteworthy to mention the latest findings on type-2-inflammation in allergic diseases as they relate to the unique miRNA profile [98,99]. For instance, in an allergic asthma model due to exposure to house dust mites or in another one, with a mouse infected with Heligmosomoides polygyrus, miRNA-15a-5p, miRNA-20b-5p, miRNA146a-5p, miRNA-155-5p and miRNA-200c-3p successfully differentiate Th2 cells (IL-4+) profile [99].
The need for reliable and accurate biomarkers of disease is an imperative of modern medicine. Regarding allergic diseases, upregulated levels of miRNA-146a in asthma patients may be a part of the unique profile of miRNAs serving as a biomarker characteristic for asthma, while the miRNA profile in allergic rhinitis is quite specific and differentially expressed than in asthma patients [1,8,100,101,102]. On the other hand, specifically miRNA-146a was proven as significantly involved in the effects of allergen-specific immunotherapy in children with allergic rhinitis, highlighting this miRNA as a potential target for future therapies [1,103]. Additionally, examining miRNA expression in tissue biopsies of eosinophilic esophagitis, as well as patients’ plasma, miRNA-146a was identified as a valuable biomarker in monitoring the effectiveness of glucocorticoid treatment [46,104]. With regard to skin diseases, upregulated levels of miRNA-146a were validated as an accurate indicator of active psoriasis [40,43,70,71,77,105]. Other skin diseases, in which miRNA-146a might be involved in their development, are oral lichen planus, vitiligo, hidradenitis suppurativa and cutaneous lupus erythematosus [40,43,50,51,53].
Evaluating the relationship between laboratory findings, clinical severity of AD and miRNAs, it has been shown that miRNA-21, miRNA-29b, miRNA-143 and miRNA-223 were significantly more expressed in patients with higher SCORAD (p< 0.001, p< 0.001, p< 0.001, p= 0.015, respectively). MiRNA-21 is mostly studied in oncology, cardiovascular diseases, inflammatory skin diseases and asthma [40,43,106,107,108]. Its role in the stimulation of keratinocyte proliferation, apoptosis inhibition, angiogenesis, activation of T-cells regardless of the subtype, and relation to the IL-22 axis in maintaining inflammation in psoriatic lesions, was recognized through different research [40,43,53]. In the allergic murine model, the upregulation of miRNA-21 promoted Th2 polarization, targeting IL-12p35 [8,109]. Overexpressed miRNA-21 was found in the skin lesions of allergic contact dermatitis in humans and mice, skin lesions of atopic eczema, and mucosal biopsies of eosinophilic esophagitis [42,44,45,52,104]. However, there is currently limited data on the relationship between atopic dermatitis and miRNA-21 expression. According to the investigation conducted to establish the association between AD in infants and particular miRNAs in breast milk, the authors hypothesized that exposure to miRNA-21 in predisposed infants may affect their immune system and result in the early development of AD [1,110]. We did not identify miRNA-21 as statistically significantly dysregulated in AD patients (p= 0.573), but after applying multinomial logistic regression, we found that the miRNA-21 was significantly more expressed in patients with higher SCORAD (p< 0.001), indicating the potential of miRNA-21 as a severity biomarker and its role in the disease progression. A few research demonstrated that miRNA-29b was overexpressed in the lesional skin of AD patients [14,25,27,41,44,52]. Also, a research showed higher miRNA-29b expression in sera of AD patients and its positive correlation with SCORAD, suggesting that miRNA-29b may be a potential biomarker of AD [27]. Our results confirmed this finding, as the miRNA-29b was significantly more expressed in patients with higher SCORAD in our study (p< 0.001), but not significantly more expressed in sera of AD patients compared with healthy controls (p= 0.665). Therefore, we could confidently imply that miRNA-29b serves as a reliable severity biomarker in AD patients, and may be responsible for the progression of atopic eczema. An experimental part of the previously mentioned study proved that miRNA29b overexpression was involved in the epidermal barrier dysfunction in AD, based on mediating γIFN-induced keratinocyte apoptosis through targeting BcL2L2 [27].
Given the fact that miRNA-143 is one of the most expressed miRNAs in the skin, it was quite expected that this miRNA played a crucial role in the pathogenesis of cutaneous diseases [44]. Exploring the atopic lesional skin, scientists detected the downregulation of miRNA-143 [28]. Further experimental studies revealed that miRNA-143 mediated Th2-driven inflammation by targeting IL-13Rα1 in keratinocytes, resulting in IL-13 suppression and, presumably, epidermal barrier restoration via regulation of filaggrin, loricrin and involucrin [14,41,44,45]. It was also shown that miRNA-143, via targeting IL-13Rα1, suppressed mast cell activation and allergic response, as well as overall IL-13-mediated inflammation in AD [37,39,43,111]. The authors emphasized the tremendous therapeutic potential of miRNA-143 in AD due to its effects on the IL-13 pathway [18,40,43,112,113]. To evaluate the capacity of miRNA-143 as a valid biomarker for skin inflammatory diseases, a few studies proposed that the upregulated miRNA-143 levels in PBMCs could have a diagnostic purpose both in infant AD and psoriasis patients, while in the cases of psoriasis, miRNA-143 may serve as a severity biomarker and in monitoring the treatment response as well [40,56,114]. The reliability of miRNA-143 expression as a diagnostic biomarker for adult AD is questionable, as the results in our AD cohort were found to be statistically insignificant (p= 0.630). However, our results indicate that miRNA-143 might be a valuable severity biomarker as its overexpression was detected in patients with higher SCORAD (p< 0.001).
Investigations on allergic contact dermatitis identified the upregulated miRNA-223 levels in human skin lesions, proposing its involvement in T cell activation and consequent cutaneous inflammation [44,53]. Apart from that, the overexpression of miRNA-223 was found in the skin changes of atopic eczema [28]. A group of authors examined the association between miRNA-223 expression and Treg-cells following prenatal tobacco exposure and its effects on children in their first three years of life. It was demonstrated that the upregulation of miRNA-223 was associated with a decreased Treg-cells number in both cord and maternal blood, accompanied by a higher risk of AD development [44]. A recent study established that plasma levels of miRNA-223 in severe AD patients (EASI>20) were significantly increased, correlating positively with serum TARC levels as well, pointing out the association with Th2 cell activation [43,115]. Additionally, another research conducted in AD patients and OVA-induced AD mice model, revealed similar findings: miRNA-223 was significantly elevated in the blood from AD patients and mice model, with clinical potential to become a valuable diagnostic biomarker, while the miRNA-223 overexpression was also associated with more extensive eczema lesions in mice model [44,116]. Although miRNA-223 has been reported as upregulated in several studies of AD, our study found no significant difference in expression between AD patients and controls (p=0.717). This discrepancy may be due to differences in patient populations, experimental methods, or disease severity across studies. There was also no correlation between miRNA-223 levels and IgE (p= 0.67) and miRNA-223 levels and eosinophil count (p= 0.707) in our cohort. Additionally, a moderate positive correlation was observed between miRNA-223 levels and SCORAD (r= 0.342, p= 0.015).
Among other miRNAs examined in our research, we would like to refer to miRNA-203 and miRNA-26a. MiRNA-203 is highly specific for skin, almost exclusively found upregulated in keratinocytes in psoriatic lesions [52]. It was shown that miRNA-203 is involved in inflammation, keratinocyte differentiation and angiogenesis, by targeting SOCS-3, LXRα/PPARγ axis, regulating αTNF, IL-8 and IL-24, and also in JAK2/STAT3 pathway activation, promoting further VEGF secretion [40,43,52,70]. Downregulated miRNA-203 was also identified in early mycosis fungoides, distinguishing it from AD [50,117]. Assessing the clinical potential of miRNA-203 in pediatric AD, scientists found that miRNA-203 serum levels were significantly elevated and in a positive correlation with the increase of serum TNFRI and TNFRII, whilst they detected the decreased miRNA-203 expression in urine and the inverse relation to serum IgE levels, suggesting miRNA-203 as an appropriate disease biomarker [14,40,44,57]. However, the miRNA-203 expression was not significantly altered in our study group compared to healthy controls (p= 0.739), nor the association with the measured blood parameters (Eos and IgE) or SCORAD was established (p= 0.79, 0.454 and 0.784, respectively). The observed discrepancy in our results and available data might be due to the age-dependent existence of distinct miRNA profiles among the AD patients.
The clinical importance of miRNA-26 was established in tumorigenesis and metabolic diseases [118,119]. It was found that lower levels of miRNA-26b contribute to enhancing inflammation via cyclooxygenase-2, but also miRNA-26a/b exhibited tumor-suppressive features and had an anti-inflammatory function in type-2 diseases such as atopic dermatitis and asthma [50,99]. Bioinformatic analysis, particularly in atopic dermatitis, identified the essential genetic targets of miRNA-26a, such as hyaluronan synthase 3, while in asthma, miRNA-26a influenced SMAD2,3 and TGF-β-related signaling pathways [1,8,34,41,120]. The decreased expression of miRNA-26 was identified in several studies regarding inflammatory skin lesions [28,50,53]. Furthermore, the downregulated miRNA-26a/b levels were detected in urine in the children with AD, while the other study found the upregulated plasma levels of miRNA-26a-1-3p in the pediatric AD [22,40,42,57]. Our study did not reveal significantly differentially expressed miRNA-26a levels in the AD group compared to controls (p= 0.585).
Summarizing, our findings imply that miRNA-146a may be a novel diagnostic biomarker of AD and might play a specific role in the development of AD, while miRNA-21, miRNA-29b and miRNA-223 may serve as severity biomarkers with an impact on the disease progression.
4.3. Therapeutic Potential of miRNAs
Apart from the role of miRNAs as biomarkers in early diagnosis or disease prognosis, their therapeutic potential has been recognized widely [1,18,40,44,60,61]. The research on this subject across diverse diseases is developing rapidly. The findings from such studies open new possibilities for the treatment of various disorders, given that epigenetic deviations are relatively reversible and susceptible to modification [1,18,40,44,60,61]. There are two main approaches to miRNAs administration, either by substitution or inhibition of their activity [40]. The replacement of downregulated miRNAs may be reached via application of miRNAs mimics (agomiRs), while conversely, miRNAs antagonists (antagomiRs) may be potent therapeutical agents in cases of aberrantly upregulated miRNAs across different diseases [40,60,61]. Additionally, a concept of 'organ-specific' rather than 'disease-specific' miRNA profile proposed by one group of authors through their comprehensive review on the usefulness of antagomiRs in lung diseases should be kept in mind. [60] They pointed out that miRNA-21 was identified as the most frequently dysregulated and involved in the underlying pathogenetic mechanisms of lung diseases [60]. For instance, with regard to allergic conditions, several studies demonstrated successful inflammatory modulation via administration of anti-miR-21, resulting in reduced PI3K activity, restored HDAC2 (Histone deacetylase 2) and suppressed airway hyper-responsiveness, or reduced eosinophil count, IL-5 and IL-13 levels, depending on the study [60]. According to these authors, the most critical point may be the proper selection of essential target miRNAs that will yield the best outcome [60]. MiRNAs serve as master regulators of different biological processes, expressed in different tissues, having a wide impact on cellular mechanisms and immune modulation [1,18,40,44,60,61]. Thus, the implementation of miRNAs agonists or antagonists should be wisely optimized due to their cell-specific or organ-specific expression [18]. Notably, some miRNAs are constitutionally found in the skin, among which are miRNA-21, miRNA-143 and miRNA-203, implying their exclusive influence on both the maintenance of skin homeostasis and pathogenetic contribution to disease development [44]. Concerning skin diseases, miRNA-146a, miRNA-155 and miRNA-223 expressed altered levels in the skin of AD and allergic contact dermatitis, indicating an overlap in pathogenetic processes between these conditions [44]. Studies conducted on psoriasis similarly showed upregulated miRNA-146a and miRNA-155 levels in psoriatic and atopic lesions [40,43,52,53,70,71]. Other skin diseases in which dysregulated miRNA-146a might be involved in disease development are oral lichen planus, vitiligo, hidradenitis suppurativa and cutaneous lupus erythematosus [40,43,50]. Additionally, miRNA-21 was found upregulated in the lesions of hidradenitis suppurativa and psoriasis, while one study showed that targeting miRNA-21 may offer a promising approach to treating psoriasis [40,43,72]. A few experimental models demonstrated significant amelioration of psoriasis resulting in less hyperplasia after administration of anti-miRNA-21, similarly to successful challenge with anti-miRNA-31 or anti-miRNA-210, which led to alleviation of inflammation, epidermal changes and severity of psoriasis [40,72]. Furthermore, application of miRNA-146, miRNA-145-5p, or miRNA-310 mimics also efficiently reduced inflammation from psoriasis [40,72]. Several studies investigated the role of miRNA-29 in collagen synthesis and fibrosis. MiRNA-29 levels were dysregulated in scars, keloids and in patients with scleroderma [40,42]. Given the fact that miRNA-29 has an antifibrotic role, it was proposed that agonist-miRNA-29 may serve as a novel treatment option in cutaneous fibrotic disorders and excessive scarring [40]. In the field of oncology, miRNA-29a/b/c was defined as one of the tumor-suppressive miRNAs in squamous cell carcinoma [50]. It is worth mentioning that miRNA-29a/b is involved in the Nrf2-desmocollin-2 axis, resulting in structural abnormalities of desmosomes in the pathogenesis of bullous diseases [50]. Increased levels of miRNA-223 were found in skin changes of toxic epidermal necrolysis, psoriatic plaques and skin lesions of hidradenitis suppurativa due to abundant inflammatory infiltrate, while the lower miRNA-203 expression was also found in lichen planus, indicating its additional role in apoptosis and inflammation in the disease progression [43,51,53,73]. Regarding the anti-miRNA-203 implementation in the treatment, experimental studies revealed the positive effects in wound healing, reduced fibrosing and scar formation, and the topical nanodelivery via elastic liposomes might successfully modulate miRNAs levels in lesional skin and therefore could potentially be used in the treatment of psoriasis [42]. In research on the AD model, remarkable improvement of atopic inflammation was reached after the introduction of anti-miRNA-155-5p, suggesting great potential of this antagomiR in treating AD. [40] Comparably, a few studies managed to show significant improvement in AD models after administration of anti-inflammatory miRNAs mimics [61]. Hence, a favorable outcome was demonstrated for agomiR-10a-5p targeting hyaluronan synthase-3 that resulted in reduced IL-8, CCL5 and keratinocyte proliferation rate, or in a case of agomiR-124 transfection into keratinocytes that led to the decrease in IL-8, CCL5 and CCL8 levels via targeting p65 [61]. Comparably, several other studies described silencing of IL-13-mediated inflammation via applying miRNA-143 mimic, or suppression of TLR2-induced generation of IL-8, CCL20 and αTNF with miRNA-146a mimic directly targeting TRAF6 and IRAK1 [61]. Additionally, administration of agomiR-375-3p or agomiR-1294 showed inhibitory activity in the inflammatory response, ultimately causing barrier restoration in AD models [61]. In support of the hypothesis of miRNAs being involved in possible treatment mechanisms, administration of belinostat, the pan-HDAC inhibitor, resulted in restoration of epidermal barrier by upregulating miRNA-335, while promoting terminal keratinocyte differentiation [34]. For the role of TLR, another study demonstrated that inhibition of TLR8 may play a protective role in AD [39]. In an experimental model, authors showed that TLR8 induced NF-κB/MyD88 signaling pathway with further production of IL-1, IL-6, IL-12, αTNF, γIFN, while its suppression resulted in reduced levels of chemokines, IgE and IL-4 [74]. The authors managed to modulate the TLR8 gene via vector miRNA derived from Salmonella enterica subsp. enterica serovar Typhimurium, suggesting the therapeutic potential of epigenetic alteration of TLR8 in severe chronic AD [39,74]. Taking all of the above into account, exploring the therapeutic potential of miRNAs could rely on a new classification of medicines, known as 'epidrugs', that exert epi-modulatory and epi-regulatory functions [33,34,61].
5. Conclusions
The broad area of research and identification of specific microRNAs offers results that can be further applied for creating new disease biomarkers, targeted individual therapies, as well as monitoring the effectiveness of treatment.
Considering that many years of clinical practice have demonstrated that current laboratory parameters in atopic dermatitis do not always correlate the severity of the clinical picture and patients' complaints, which significantly impair their quality of life and the overall health, it is crucial to expand existing knowledge, determinate novel biomarkers of atopy and implement them in routine clinical practice.
Additionally, there is still a need for more specific and safe therapy options, with special reference to the treatment of quite challenging patient subpopulations, such as those who do not respond to conventional therapies, refractory cases and patients with contraindications or severe adverse effects.
This research advances the field by establishing the specific miRNA profile in atopic patients, emphasizing miRNA-146a, miRNA-21, miRNA-223 and miRNA-29b, while providing a suitable base for future investigation. We believe this study makes a significant contribution to the scientific audience by addressing the need for more precise diagnostic strategies and the urgent identification of novel molecules that could serve as potential targets for future treatment options. Apart from dermatology, this multidisciplinary topic permeates allergology, immunology and molecular biology. Also, results imply a novel role and possible future implementation in clinical practice guidelines and stepping further, in pharmaceutical science and drug production.
However, the study has a few potential limitations. In view of the number of patients, larger cohorts from multiple health centers encompassing all age and ethnic groups would be more preferable to generalize results. Also, a more diverse patient cohort would be advisable, with special respect to a variety in AD forms (e.g. intrinsic vs. extrinsic AD) or subgrouping AD patients dependent on clinical phenotypes (e.g. classic flexural AD, 'head-and-neck' dermatitis, hand eczema, erythrodermic forms, etc.) and miRNAs profile in creating novel pheno-endotypes of AD.
Future work should be aimed at exploring definitive biologic mechanisms by which miRNA-146a may be involved in eosinophil regulation and atopic inflammation in humans, or focusing on monitoring miRNA-146a, miRNA-21, miRNA-223 and miRNA-29b during the administration of different therapeutic modalities, to establish their potential as treatment response biomarkers. Further research is needed to elucidate the contribution of combinatorial miRNAs in allergic inflammation, forming a major basis for creating precise, individually tailored treatment options.
Author Contributions
Conceptualization, S.J., Lj.V. and I.B..; methodology, S.J., Lj.V. and I.B.; validation, I.B. and O.L.; formal analysis, I.B.; investigation, S.J. and Lj.V.; resources, D.J.; data curation, I.B. and O.L.; writing—original draft preparation, S.J.; writing—review and editing, O.L. and D.J.; visualization, I.B. and D.J.; supervision, Lj.V.; funding acquisition, S.J. and Lj.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NAOS-Bioderma donation for investigation, conducted as a part of a PhD dissertation at the University Clinical Centre of Vojvodina, Serbia.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by Ethics Committee of the Clinical Centre of Vojvodina (protocol number #600-210).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
We hereby declare competing interests: this research was supported by NAOS-Bioderma donation. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Specjalski K, Jassem E. MicroRNAs: Potential Biomarkers and Targets of Therapy in Allergic Diseases? Arch Immunol Ther Exp (Warsz). 2019 Aug;67(4):213-23. Epub 2019 May 28.
- Lugović-Mihić L, Meštrović-Štefekov J, Potočnjak I, Cindrić T, Ilić I, Lovrić I, et al. Atopic Dermatitis: Disease Features, Therapeutic Options, and a Multidisciplinary Approach. Life. 2023; 13(6):1419. [CrossRef]
- Lobefaro F, Gualdi G, Di Nuzzo S, Amerio P. Atopic Dermatitis: Clinical Aspects and Unmet Needs. Biomedicines. 2022; 10(11):2927. [CrossRef]
- Plewig G, French L, Ruzicka T, Kaufmann R, Hertl M, editors. Braun-Falcos Dermatology. Berlin, Germany: Springer; 2022.
- Bolognia JL, Jorizzo JL, Schaffer JV, editors. Dermatology. 4thed. Philadelphia: Elsevier Saunders; 2018.
- Raimondo A, Lembo S. Atopic Dermatitis: Epidemiology and Clinical Phenotypes. Dermatol Pract Concept [Internet]. 2021 Oct. 29 [cited 2025 Mar. 17];:e2021146. Available from: https://dpcj.org/index.php/dpc/article/view/dermatol-pract-concept-articleid-dp1104a146.
- Thyssen JP, Halling AS, Schmid-Grendelmeier P, Guttman-Yassky E, Silverberg JI. Comorbidities of atopic dermatitis-what does the evidence say? J Allergy Clin Immunol. 2023 May;151(5):1155-1162. Epub 2023 Jan 6. [CrossRef] [PubMed]
- Weidner J, Bartel S, Kılıç A, Zissler UM, Renz H, Schwarze J, et al. Spotlight on microRNAs in allergy and asthma. Allergy. 2021 Jun;76(6):1661-78. Epub 2020 Nov 20.
- Mrkić Kobal I, Plavec D, Vlašić Lončarić Ž, Jerković I, Turkalj M. Atopic March or Atopic Multimorbidity-Overview of Current Research. Medicina (Kaunas). 2023 Dec 22;60(1):21. [CrossRef] [PubMed] [PubMed Central]
- Tham EH, Leung DY. Mechanisms by Which Atopic Dermatitis Predisposes to Food Allergy and the Atopic March. Allergy Asthma Immunol Res. 2019 Jan;11(1):4-15. [CrossRef] [PubMed] [PubMed Central]
- Yaneva M, Darlenski R. The link between atopic dermatitis and asthma- immunological imbalance and beyond. Asthma Res Pract. 2021 Dec 15;7(1):16. [CrossRef] [PubMed] [PubMed Central]
- de Las Vecillas L, Quirce S. The Multiple Trajectories of the Allergic March. J Investig Allergol Clin Immunol. 2024 Apr 12;34(2):75-84. Epub 2023 Dec 19. [CrossRef] [PubMed]
- Bin L, Leung DY. Genetic and epigenetic studies of atopic dermatitis. Allergy Asthma Clin Immunol. 2016 Oct 19;12:52. [CrossRef] [PubMed] [PubMed Central]
- Hernández-Rodríguez RT, Amezcua-Guerra LM. The potential role of microRNAs as biomarkers in atopic dermatitis: a systematic review. Eur Rev Med Pharmacol Sci. 2020 Nov;24(22):11804-11809. [CrossRef] [PubMed]
- Bao L, Chau C, Bao J, Tsoukas MM, Chan LS. IL-4 dysregulates microRNAs involved in inflammation, angiogenesis and apoptosis in epidermal keratinocytes. Microbiol Immunol. 2018 Nov;62(11):732-736. [CrossRef] [PubMed]
- Jadali Z. Th9 Cells as a New Player in Inflammatory Skin Disorders. Iran J Allergy Asthma Immunol. 2019 Apr 1;18(2):120-130. PMID: 31066248.
- Pescitelli L, Rosi E, Ricceri F, Pimpinelli N, Prignano F. Novel Therapeutic Approaches and Targets for the Treatment of Atopic Dermatitis. Curr Pharm Biotechnol. 2021;22(1):73-84. [CrossRef]
- Khosrojerdi M, Azad FJ, Yadegari Y, Ahanchian H, Azimian A. The role of microRNAs in atopic dermatitis. Noncoding RNA Res. 2024 May 28;9(4):1033-39.
- Tsuji G, Yamamura K, Kawamura K, Kido-Nakahara M, Ito T, Nakahara T. Novel Therapeutic Targets for the Treatment of Atopic Dermatitis. Biomedicines. 2023 Apr 27;11(5):1303. [CrossRef] [PubMed] [PubMed Central]
- Gatmaitan JG, Lee JH. Challenges and Future Trends in Atopic Dermatitis. International Journal of Molecular Sciences. 2023; 24(14):11380. [CrossRef]
- Otsuka A, Nomura T, Rerknimitr P, Seidel JA, Honda T, Kabashima K. The interplay between genetic and environmental factors in the pathogenesis of atopic dermatitis. Immunol Rev. 2017 Jul;278(1):246-262. [CrossRef] [PubMed]
- Meng L, Li M, Gao Z, Ren H, Chen J, Liu X, et al. Possible role of hsa-miR-194-5p, via regulation of HS3ST2, in the pathogenesis of atopic dermatitis in children. Eur J Dermatol. 2019 Dec 1;29(6):603-13.
- Schmidt AD, de Guzman Strong C. Current understanding of epigenetics in atopic dermatitis. Exp Dermatol. 2021 Aug;30(8):1150-5.
- Lee YS, Han SB, Ham HJ, Park JH, Lee JS, Hwang DY, et al. IL-32γ suppressed atopic dermatitis through inhibition of miR-205 expression via inactivation of nuclear factor-kappa B. J Allergy Clin Immunol. 2020 Jul;146(1):156-68. Epub 2020 Jan 10.
- Rebane A, Runnel T, Aab A, Maslovskaja J, Rückert B, Zimmermann M, et al. MicroRNA-146a alleviates chronic skin inflammation in atopic dermatitis through suppression of innate immune responses in keratinocytes. J Allergy Clin Immunol. 2014 Oct;134(4):836-47.e11. Epub 2014 Jul 2.
- Yan F, Meng W, Ye S, Zhang X, Mo X, Liu J, et al. MicroRNA 146a as a potential regulator involved in the pathogenesis of atopic dermatitis. Mol Med Rep. 2019 Nov;20(5):4645-53. Epub 2019 Sep 20.
- Gu C, Li Y, Wu J, Xu J. IFN-γ-induced microRNA-29b up-regulation contributes tokeratinocyte apoptosis in atopic dermatitis through inhibiting Bcl2L2. Int J Clin Exp Pathol. 2017 Sep 1;10(9):10117-26.
- Sonkoly E, Janson P, Majuri ML, Savinko T, Fyhrquist N, Eidsmo L, et al. MiR-155 is overexpressed in patients with atopic dermatitis and modulates T-cell proliferative responses by targeting cytotoxic T lymphocyte-associated antigen 4. J Allergy Clin Immunol. 2010 Sep;126(3):581-9.e1-20. Epub 2010 Jul 31.
- Bergallo M, Accorinti M, Galliano I, Coppo P, Montanari P, Quaglino P, et al. Expression of miRNA 155, FOXP3 and ROR gamma, in children with moderate and severe atopic dermatitis. G Ital Dermatol Venereol. 2020 Apr;155(2):168-72.
- Ma L, Xue HB, Wang F, Shu CM, Zhang JH. MicroRNA-155 may be involved in the pathogenesis of atopic dermatitis by modulating the differentiation and function of T helper type 17 (Th17) cells. Clin Exp Immunol. 2015 Jul;181(1):142-9. Epub 2015 May 25.
- Chen XF, Zhang LJ, Zhang J, Dou X, Shao Y, Jia XJ, et al. MiR-151a is involved in the pathogenesis of atopic dermatitis by regulating interleukin-12 receptor β2. Exp Dermatol. 2018 Apr;27(4):427-32. Epub 2017 Apr 11.
- Acevedo N, Benfeitas R, Katayama S, Bruhn S, Andersson A, Wikberg G, et al. Epigenetic alterations in skin homing CD4+CLA+ T cells of atopic dermatitis patients. Sci Rep. 2020 Oct 22;10(1):18020. [CrossRef]
- Moltrasio C, Romagnuolo M, Marzano AV. Epigenetic Mechanisms of Epidermal Differentiation. Int J Mol Sci. 2022 Apr 28;23(9):4874. [CrossRef] [PubMed] [PubMed Central]
- Livshits G, Kalinkovich A. Resolution of Chronic Inflammation, Restoration of Epigenetic Disturbances and Correction of Dysbiosis as an Adjunctive Approach to the Treatment of Atopic Dermatitis. Cells. 2024 Nov 18;13(22):1899. [CrossRef] [PubMed] [PubMed Central]
- Lee, AY. The Role of MicroRNAs in Epidermal Barrier. Int J Mol Sci. 2020 Aug 12;21(16):5781. [CrossRef] [PubMed] [PubMed Central]
- Yang Z, Zeng B, Wang C, Wang H, Huang P, Pan Y. MicroRNA-124 alleviates chronic skin inflammation in atopic eczema via suppressing innate immune responses in keratinocytes. Cell Immunol. 2017 Sep;319:53-60. Epub 2017 Aug 25. [CrossRef] [PubMed]
- Zeng YP, Nguyen GH, Jin HZ. MicroRNA-143 inhibits IL-13-induced dysregulation of the epidermal barrier-related proteins in skin keratinocytes via targeting to IL-13Rα1. Mol Cell Biochem. 2016 May;416(1-2):63-70. Epub 2016 Apr 5. [CrossRef] [PubMed]
- Sun Y, Li XQ, Sahbaie P, Shi XY, Li WW, Liang DY, et al. miR-203 regulates nociceptive sensitization after incision by controlling phospholipase A2 activating protein expression. Anesthesiology. 2012 Sep;117(3):626-38. [CrossRef] [PubMed] [PubMed Central]
- Deng Q, Yao X, Fang S, Sun Y, Liu L, Li C, et al. Mast cell-mediated microRNA functioning in immune regulation and disease pathophysiology. Clin Exp Med. 2025 Jan 15;25(1):38. [CrossRef] [PubMed] [PubMed Central]
- Domingo S, Solé C, Moliné T, Ferrer B, Cortés-Hernández J. MicroRNAs in Several Cutaneous Autoimmune Diseases: Psoriasis, Cutaneous Lupus Erythematosus and Atopic Dermatitis. Cells. 2020 Dec 10;9(12):2656.
- Yu X, Wang M, Li L, Zhang L, Chan MTV, Wu WKK. MicroRNAs in atopic dermatitis: A systematic review. J Cell Mol Med. 2020 Jun;24(11):5966-72. Epub 2020 Apr 30.
- Rożalski M, Rudnicka L, Samochocki Z. MiRNA in atopic dermatitis. Postepy Dermatol Alergol. 2016 Jun;33(3):157-62. Epub 2016 Jun 17.
- Dopytalska K, Czaplicka A, Szymańska E, Walecka I. The Essential Role of microRNAs in Inflammatory and Autoimmune Skin Diseases-A Review. Int J Mol Sci. 2023 May 23;24(11):9130.
- Brancaccio R, Murdaca G, Casella R, Loverre T, Bonzano L, Nettis E, et al. miRNAs' Cross-Involvement in Skin Allergies: A New Horizon for the Pathogenesis, Diagnosis and Therapy of Atopic Dermatitis, Allergic Contact Dermatitis and Chronic Spontaneous Urticaria. Biomedicines. 2023 Apr 24;11(5):1266.
- Dissanayake E, Inoue Y. MicroRNAs in Allergic Disease. Curr Allergy Asthma Rep. 2016 Sep;16(9):67. [CrossRef]
- Rebane A. microRNA and Allergy. In: Gaetano S, editor. microRNA: Medical Evidence: From Molecular Biology to Clinical Practice. Cham: Springer International Publishing; 2015.
- Rebane A, Akdis CA. MicroRNAs in allergy and asthma. Curr Allergy Asthma Rep. 2014 Apr;14(4):424.
- Chen L, Zhong JL. MicroRNA and heme oxygenase-1 in allergic disease. Int Immunopharmacol. 2020 Mar;80:106132. Epub 2020 Jan 22.
- Lu TX, Rothenberg ME. MicroRNA. J Allergy Clin Immunol. 2018 Apr;141(4):1202-07. Epub 2017 Oct 23.
- Ruksha TG, Komina AV, Palkina NV. MicroRNA in skin diseases. Eur J Dermatol. 2017 Aug 1;27(4):343-352. [CrossRef] [PubMed]
- Mannucci C, Casciaro M, Minciullo PL, Calapai G, Navarra M, Gangemi S. Involvement of microRNAs in skin disorders: A literature review. Allergy Asthma Proc. 2017 Jan 1;38(1):9-15. [CrossRef] [PubMed]
- Sonkoly E, Wei T, Janson PC, Sääf A, Lundeberg L, Tengvall-Linder M, et al. MicroRNAs: novel regulators involved in the pathogenesis of psoriasis? PLoS One. 2007 Jul 11;2(7):e610. [CrossRef] [PubMed] [PubMed Central]
- Løvendorf MB, Skov L. miRNAs in inflammatory skin diseases and their clinical implications. Expert Rev Clin Immunol. 2015 Apr;11(4):467-77. Epub 2015 Feb 26. [CrossRef] [PubMed]
- Bakker D, de Bruin-Weller M, Drylewicz J, van Wijk F, Thijs J. Biomarkers in atopic dermatitis. J Allergy Clin Immunol. 2023 May;151(5):1163-1168. Epub 2023 Feb 14. [CrossRef] [PubMed]
- Mastraftsi S, Vrioni G, Bakakis M, Nicolaidou E, Rigopoulos D, Stratigos AJ, et al. Atopic Dermatitis: Striving for Reliable Biomarkers. J Clin Med. 2022 Aug 9;11(16):4639. [CrossRef] [PubMed] [PubMed Central]
- Nousbeck J, McAleer MA, Hurault G, Kenny E, Harte K, Kezic S, et al. MicroRNA analysis of childhood atopic dermatitis reveals a role for miR-451a. Br J Dermatol. 2021 Mar;184(3):514-23. [CrossRef]
- Lv Y, Qi R, Xu J, Di Z, Zheng H, Huo W, et al. Profiling of serum and urinary microRNAs in children with atopic dermatitis. PLoS One. 2014 Dec 22;9(12):e115448. [CrossRef]
- Lu TX, Rothenberg ME. Diagnostic, functional, and therapeutic roles of microRNA in allergic diseases. J Allergy Clin Immunol. 2013 Jul;132(1):3-13; quiz 14. Epub 2013 Jun 2. [CrossRef] [PubMed] [PubMed Central]
- Yasuike R, Tamagawa-Mineoka R, Nakamura N, Masuda K, Katoh N. Plasma miR223 is a possible biomarker for diagnosing patients with severe atopic dermatitis. Allergol Int. 2021 Jan;70(1):153-155. Epub 2020 Sep 1. [CrossRef] [PubMed]
- Murdaca G, Tonacci A, Negrini S, Greco M, Borro M, Puppo F, et al. Effects of AntagomiRs on Different Lung Diseases in Human, Cellular, and Animal Models. Int J Mol Sci. 2019 Aug 13;20(16):3938. [CrossRef] [PubMed] [PubMed Central]
- Yang SC, Alalaiwe A, Lin ZC, Lin YC, Aljuffali IA, Fang JY. Anti-Inflammatory microRNAs for Treating Inflammatory Skin Diseases. Biomolecules. 2022 Aug 3;12(8):1072. [CrossRef] [PubMed] [PubMed Central]
- Rullo VE, Segato A, Kirsh A, Sole D. Severity scoring of atopic dermatitis: a comparison of two scoring systems. Allergol Immunopathol (Madr). 2008 Jul-Aug;36(4):205-11.
- Ricci G, Dondi A, Patrizi A. Useful tools for the management of atopic dermatitis. Am J Clin Dermatol. 2009;10(5):287-300.
- Severity scoring of atopic dermatitis: the SCORAD index. Consensus Report of the European Task Force on Atopic Dermatitis. Dermatology. 1993;186(1):23-31. [CrossRef]
- Eichenfield LF, Tom WL, Chamlin SL, Feldman SR, Hanifin JM, Simpson EL, et al. Guidelines of care for the management of atopic dermatitis: section 1. Diagnosis and assessment of atopic dermatitis. J Am Acad Dermatol. 2014 Feb;70(2):338-51. Epub 2013 Nov 27.
- Kortekaas Krohn I, Badloe FMS, Herrmann N, Maintz L, De Vriese S, Ring J, et al. Immunoglobulin E autoantibodies in atopic dermatitis associate with Type-2 comorbidities and the atopic march. Allergy. 2023 Dec;78(12):3178-92. Epub 2023 Jul 24.
- Vaneckova J, Bukač J. The severity of atopic dermatitis and the.
- relation to the level of total IgE, onset of atopic dermatitis and family history about atopy. Food Agric. Immunol. 2016 May; 27(5):1-8.
- Furue M, Chiba T, Tsuji G, Ulzii D, Kido-Nakahara M, Nakahara T, et al. Atopic dermatitis: immune deviation, barrier dysfunction, IgE autoreactivity and new therapies. Allergol Int. 2017 Jul;66(3):398-403. Epub 2017 Jan 2.
- Simon D, Braathen LR, Simon HU. Eosinophils and atopic dermatitis. Allergy. 2004 Jun;59(6):561-70.
- Timis TL, Orasan RI. Understanding psoriasis: Role of miRNAs. Biomed Rep. 2018 Nov;9(5):367-374. Epub 2018 Sep 11. [CrossRef] [PubMed] [PubMed Central]
- Hermann H, Runnel T, Aab A, Baurecht H, Rodriguez E, Magilnick N, et al. miR-146b Probably Assists miRNA-146a in the Suppression of Keratinocyte Proliferation and Inflammatory Responses in Psoriasis. J Invest Dermatol. 2017 Sep;137(9):1945-1954. Epub 2017 Jun 6. [CrossRef] [PubMed] [PubMed Central]
- Guinea-Viniegra J, Jiménez M, Schonthaler HB, Navarro R, Delgado Y, Concha-Garzón MJ, et al. Targeting miR-21 to treat psoriasis. Sci Transl Med. 2014 Feb 26;6(225):225re1. [CrossRef] [PubMed]
- Ichihara A, Wang Z, Jinnin M, Izuno Y, Shimozono N, Yamane K, et al. Upregulation of miR-18a-5p contributes to epidermal necrolysis in severe drug eruptions. J Allergy Clin Immunol. 2014 Apr;133(4):1065-74. Epub 2013 Nov 1. [CrossRef] [PubMed]
- Yoon W, Kim EJ, Park Y, Kim S, Park YK, Yoo Y. Bacterially Delivered miRNA-Mediated Toll-like Receptor 8 Gene Silencing for Combined Therapy in a Murine Model of Atopic Dermatitis: Therapeutic Effect of miRTLR8 in AD. Microorganisms. 2021 Aug 12;9(8):1715. [CrossRef] [PubMed] [PubMed Central]
- Carreras-Badosa G, Runnel T, Plaas M, Kärner J, Rückert B, et al. microRNA-146a is linked to the production of IgE in mice but not in atopic dermatitis patients. Allergy. 2018 Dec;73(12):2400-3. Epub 2018 Aug 21.
- Li F, Huang Y, Huang YY, Kuang YS, Wei YJ, Xiang L, et al. MicroRNA-146a promotes IgE class switch in B cells via upregulating 14-3-3σ expression. Mol Immunol. 2017 Dec;92:180-189. Epub 2017 Nov 2. [CrossRef] [PubMed]
- Srivastava A, Nikamo P, Lohcharoenkal W, Li D, Meisgen F, Xu Landén N, et al. MicroRNA-146a suppresses IL-17-mediated skin inflammation and is genetically associated with psoriasis. J Allergy Clin Immunol. 2017 Feb;139(2):550-561. Epub 2016 Aug 24. [CrossRef]
- Gilyazova I, Asadullina D, Kagirova E, Sikka R, Mustafin A, Ivanova E, et al. MiRNA-146a-A Key Player in Immunity and Diseases. Int J Mol Sci. 2023 Aug 14;24(16):12767. [CrossRef] [PubMed] [PubMed Central]
- Yang Y, Yin X, Yi J, Peng X. MiR-146a overexpression effectively improves experimental allergic conjunctivitis through regulating CD4+CD25-T cells. Biomed Pharmacother. 2017 Oct;94:937-943. Epub 2017 Aug 12. [CrossRef] [PubMed]
- Zhou J, Lu Y, Wu W, Feng Y. HMSC-Derived Exosome Inhibited Th2 Cell Differentiation via Regulating miR-146a-5p/SERPINB2 Pathway. J Immunol Res. 2021 May 14;2021:6696525. [CrossRef] [PubMed] [PubMed Central]
- Shefler I, Salamon P, Mekori YA. MicroRNA Involvement in Allergic and Non-Allergic Mast Cell Activation. Int J Mol Sci. 2019 Apr 30;20(9):2145. [CrossRef] [PubMed] [PubMed Central]
- Luo X, Han M, Liu J, Wang Y, Luo X, Zheng J, et al. Epithelial cell-derived micro RNA-146a generates interleukin-10-producing monocytes to inhibit nasal allergy. Sci Rep. 2015 Nov 3;5:15937. [CrossRef] [PubMed] [PubMed Central]
- Liu J, Xu Z, Yu J, Zang X, Jiang S, Xu S, et al. MiR-146a-5p engineered hucMSC-derived extracellular vesicles attenuate Dermatophagoides farinae-induced allergic airway epithelial cell inflammation. Front Immunol. 2024 Sep 19;15:1443166. [CrossRef] [PubMed] [PubMed Central]
- Bélanger É, Laprise C. Could the Epigenetics of Eosinophils in Asthma and Allergy Solve Parts of the Puzzle? Int J Mol Sci. 2021 Aug 19;22(16):8921. [CrossRef] [PubMed] [PubMed Central]
- Rodrigo-Muñoz JM, Cañas JA, Sastre B, Rego N, Greif G, Rial M, et al. Asthma diagnosis using integrated analysis of eosinophil microRNAs. Allergy. 2019 Mar;74(3):507-517. Epub 2018 Oct 11. [CrossRef] [PubMed]
- Bélanger É, Madore AM, Boucher-Lafleur AM, Simon MM, Kwan T, Pastinen T, et al. Eosinophil microRNAs Play a Regulatory Role in Allergic Diseases Included in the Atopic March. Int J Mol Sci. 2020 Nov 27;21(23):9011. [CrossRef] [PubMed] [PubMed Central]
- Allantaz F, Cheng DT, Bergauer T, Ravindran P, Rossier MF, Ebeling M, et al. Expression profiling of human immune cell subsets identifies miRNA-mRNA regulatory relationships correlated with cell type specific expression. PLoS One. 2012;7(1):e29979. Epub 2012 Jan 20. [CrossRef] [PubMed] [PubMed Central]
- Weidner J, Ekerljung L, Malmhäll C, Miron N, Rådinger M. Circulating microRNAs correlate to clinical parameters in individuals with allergic and non-allergic asthma. Respir Res. 2020 May 7;21(1):107. [CrossRef] [PubMed] [PubMed Central]
- Lu TX, Lim EJ, Besse JA, Itskovich S, Plassard AJ, Fulkerson PC, et al. MiR-223 deficiency increases eosinophil progenitor proliferation. J Immunol. 2013 Feb 15;190(4):1576-82. Epub 2013 Jan 16. [CrossRef] [PubMed] [PubMed Central]
- Lu TX, Lim EJ, Itskovich S, Besse JA, Plassard AJ, Mingler MK, et al. Targeted ablation of miR-21 decreases murine eosinophil progenitor cell growth. PLoS One. 2013;8(3):e59397. Epub 2013 Mar 22. [CrossRef] [PubMed] [PubMed Central]
- Lyu B, Wei Z, Jiang L, Ma C, Yang G, Han S. MicroRNA-146a negatively regulates IL-33 in activated group 2 innate lymphoid cells by inhibiting IRAK1 and TRAF6. Genes Immun. 2020 Jan;21(1):37-44. Epub 2019 Aug 22. [CrossRef] [PubMed]
- Fang SB, Zhang HY, Wang C, He BX, Liu XQ, Meng XC, et al. Small extracellular vesicles derived from human mesenchymal stromal cells prevent group 2 innate lymphoid cell-dominant allergic airway inflammation through delivery of miR-146a-5p. J Extracell Vesicles. 2020 Feb 10;9(1):1723260. [CrossRef] [PubMed] [PubMed Central]
- Wang J, Cui Z, Liu L, Zhang S, Zhang Y, Zhang Y, et al. MiR-146a mimic attenuates murine allergic rhinitis by downregulating TLR4/TRAF6/NF-κB pathway. Immunotherapy. 2019 Sep;11(13):1095-1105. Epub 2019 Jul 30. [CrossRef] [PubMed]
- Li A, Li Y, Zhang X, Zhang C, Li T, Zhang J, et al. The human milk oligosaccharide 2'-fucosyllactose attenuates β-lactoglobulin-induced food allergy through the miR-146a-mediated toll-like receptor 4/nuclear factor-κB signaling pathway. J Dairy Sci. 2021 Oct;104(10):10473-10484. Epub 2021 Jul 30. [CrossRef] [PubMed]
- Wang X, Lu X, Ma C, Ma L, Han S. Combination of TLR agonist and miR146a mimics attenuates ovalbumin-induced asthma. Mol Med. 2020 Jun 29;26(1):65. [CrossRef] [PubMed] [PubMed Central]
- Sun W, Sheng Y, Chen J, Xu D, Gu Y. Down-Regulation of miR-146a Expression Induces Allergic Conjunctivitis in Mice by Increasing TSLP Level. Med Sci Monit. 2015 Jul 11;21:2000-7. [CrossRef] [PubMed] [PubMed Central]
- Guo H, Zhang Y, Liao Z, Zhan W, Wang Y, Peng Y, et al. MiR-146a upregulates FOXP3 and suppresses inflammation by targeting HIPK3/STAT3 in allergic conjunctivitis. Ann Transl Med. 2022 Mar;10(6):344. [CrossRef] [PubMed] [PubMed Central]
- Kılıç A, Santolini M, Nakano T, Schiller M, Teranishi M, Gellert P, et al. A systems immunology approach identifies the collective impact of 5 miRs in Th2 inflammation. JCI Insight. 2018 Jun 7;3(11):e97503. [CrossRef] [PubMed] [PubMed Central]
- Guidi R, Wedeles CJ, Wilson MS. ncRNAs in Type-2 Immunity. Noncoding RNA. 2020 Mar 6;6(1):10. [CrossRef]
- Panganiban RP, Wang Y, Howrylak J, Chinchilli VM, Craig TJ, August A, et al. Circulating microRNAs as biomarkers in patients with allergic rhinitis and asthma. J Allergy Clin Immunol. 2016 May;137(5):1423-32. Epub 2016 Mar 26. [CrossRef] [PubMed]
- Sharma R, Tiwari A, McGeachie MJ. Recent miRNA Research in Asthma. Curr Allergy Asthma Rep. 2022 Dec;22(12):231-258. Epub 2022 Dec 2. [CrossRef] [PubMed] [PubMed Central]
- Hammad Mahmoud Hammad R, Hamed DHED, Eldosoky MAER, Ahmad AAES, Osman HM, Abd Elgalil HM, et al. Plasma microRNA-21, microRNA-146a and IL-13 expression in asthmatic children. Innate Immun. 2018 Apr;24(3):171-179. [CrossRef] [PubMed] [PubMed Central]
- Luo X, Hong H, Tang J, Wu X, Lin Z, Ma R, et al. Increased Expression of miR-146a in Children With Allergic Rhinitis After Allergen-Specific Immunotherapy. Allergy Asthma Immunol Res. 2016 Mar;8(2):132-40. Epub 2015 Oct 22. [CrossRef] [PubMed] [PubMed Central]
- Lu TX, Sherrill JD, Wen T, Plassard AJ, Besse JA, Abonia JP, et al. MicroRNA signature in patients with eosinophilic esophagitis, reversibility with glucocorticoids, and assessment as disease biomarkers. J Allergy Clin Immunol. 2012 Apr;129(4):1064-75.e9. Epub 2012 Mar 3. [CrossRef] [PubMed] [PubMed Central]
- Leal B, Carvalho C, Ferreira AM, Nogueira M, Brás S, Silva BM, et al. Serum Levels of miR-146a in Patients with Psoriasis. Mol Diagn Ther. 2021 Jul;25(4):475-485. Epub 2021 May 2. [CrossRef] [PubMed]
- Kumarswamy R, Volkmann I, Thum T. Regulation and function of miRNA-21 in health and disease. RNA Biol. 2011 Sep-Oct;8(5):706-13. Epub 2011 Jul 7.
- Silvia Lima RQD, Vasconcelos CFM, Gomes JPA, Bezerra de Menezes EDS, de Oliveira Silva B, Montenegro C, et al. miRNA-21, an oncomiR that regulates cell proliferation, migration, invasion and therapy response in lung cancer. Pathol Res Pract. 2024 Nov;263:155601. Epub 2024 Oct 3.
- Jayawardena E, Medzikovic L, Ruffenach G, Eghbali M. Role of miRNA-1 and miRNA-21 in Acute Myocardial Ischemia-Reperfusion Injury and Their Potential as Therapeutic Strategy. Int J Mol Sci. 2022 Jan 28;23(3):1512.
- Lu TX, Munitz A, Rothenberg ME. MicroRNA-21 is up-regulated in allergic airway inflammation and regulates IL-12p35 expression. J Immunol. 2009 Apr 15;182(8):4994-5002.
- Simpson MR, Brede G, Johansen J, Johnsen R, Storrø O, Sætrom P, et al. Human Breast Milk miRNA, Maternal Probiotic Supplementation and Atopic Dermatitis in Offspring. PLoS One. 2015 Dec 14;10(12):e0143496.
- Yu S, Zhang R, Zhu C, Cheng J, Wang H, Wu J. MicroRNA-143 downregulates interleukin-13 receptor alpha1 in human mast cells. Int J Mol Sci. 2013 Aug 19;14(8):16958-69. [CrossRef] [PubMed] [PubMed Central]
- Jia QN, Zeng YP. Rapamycin blocks the IL-13-induced deficiency of Epidermal Barrier Related Proteins via upregulation of miR-143 in HaCaT Keratinocytes. Int J Med Sci. 2020 Jul 25;17(14):2087-94.
- Zu Y, Chen XF, Li Q, Zhang ST. CYT387, a Novel JAK2 Inhibitor, Suppresses IL-13-Induced Epidermal Barrier Dysfunction Via miR-143 Targeting IL-13Rα1 and STAT3. Biochem Genet. 2021 Apr;59(2):531-46. Epub 2020 Nov 15.
- Løvendorf MB, Zibert JR, Gyldenløve M, Røpke MA, Skov L. MicroRNA-223 and miR-143 are important systemic biomarkers for disease activity in psoriasis. J Dermatol Sci. 2014 Aug;75(2):133-9. Epub 2014 May 21.
- Yasuike R, Tamagawa-Mineoka R, Nakamura N, Masuda K, Katoh N. Plasma miR223 is a possible biomarker for diagnosing patients with severe atopic dermatitis. Allergol Int. 2021 Jan;70(1):153-5. Epub 2020 Sep 1.
- Jia HZ, Liu SL, Zou YF, Chen XF, Yu L, Wan J, et al. MicroRNA-223 is involved in the pathogenesis of atopic dermatitis by affecting histamine-N-methyltransferase. Cell Mol Biol (Noisy-le-grand). 2018 Feb 28;64(3):103-7.
- Ralfkiaer U, Lindahl LM, Litman T, Gjerdrum LM, Ahler CB, Gniadecki R, et al. MicroRNA expression in early mycosis fungoides is distinctly different from atopic dermatitis and advanced cutaneous T-cell lymphoma. Anticancer Res. 2014 Dec;34(12):7207-17. Erratum in: Anticancer Res. 2015 Feb;35(2):1219.
- Li C, Li Y, Lu Y, Niu Z, Zhao H, Peng Y, et al. miR-26 family and its target genes in tumorigenesis and development. Crit Rev Oncol Hematol. 2021 Jan;157:103124. Epub 2020 Oct 20.
- Dandare A, Khan MJ, Naeem A, Liaquat A. Clinical relevance of circulating non-coding RNAs in metabolic diseases: Emphasis on obesity, diabetes, cardiovascular diseases and metabolic syndrome. Genes Dis. 2022 Jun 3;10(6):2393-413.
- Kärner J, Wawrzyniak M, Tankov S, Runnel T, Aints A, Kisand K, et al. Increased microRNA-323-3p in IL-22/IL-17-producing T cells and asthma: a role in the regulation of the TGF-β pathway and IL-22 production. Allergy. 2017 Jan;72(1):55-65. Epub 2016 Apr 29.
Figure 1.
Mean Ct values for selected miRNA in both groups (mean ± 95% CI) AD – atopic dermatitis, C – controls.
Figure 1.
Mean Ct values for selected miRNA in both groups (mean ± 95% CI) AD – atopic dermatitis, C – controls.
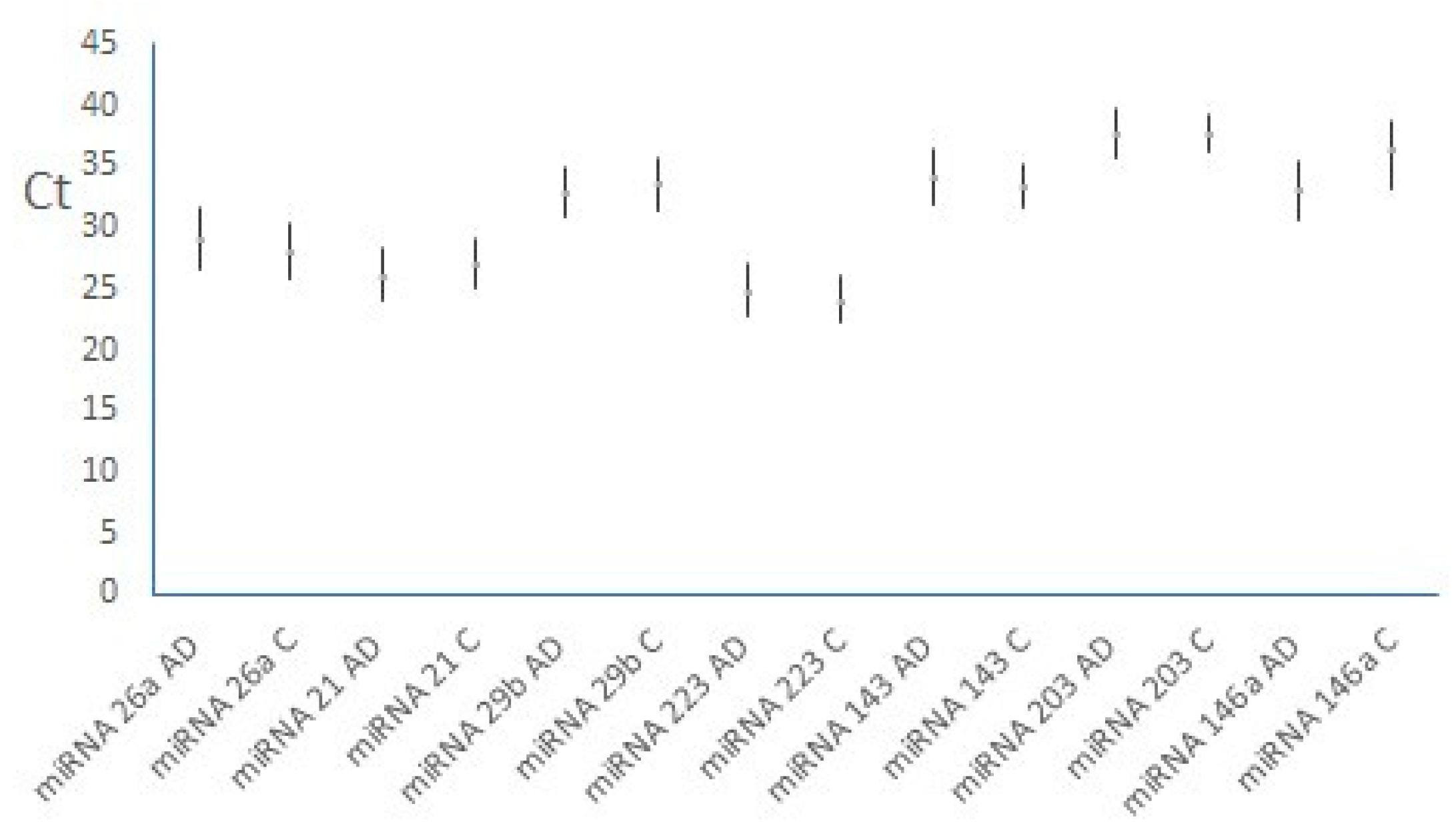
Figure 2.
Mean ΔCt values for selected miRNAs in both groups (mean ± 95% CI). AD – atopic dermatitis, C – controls.
Figure 2.
Mean ΔCt values for selected miRNAs in both groups (mean ± 95% CI). AD – atopic dermatitis, C – controls.
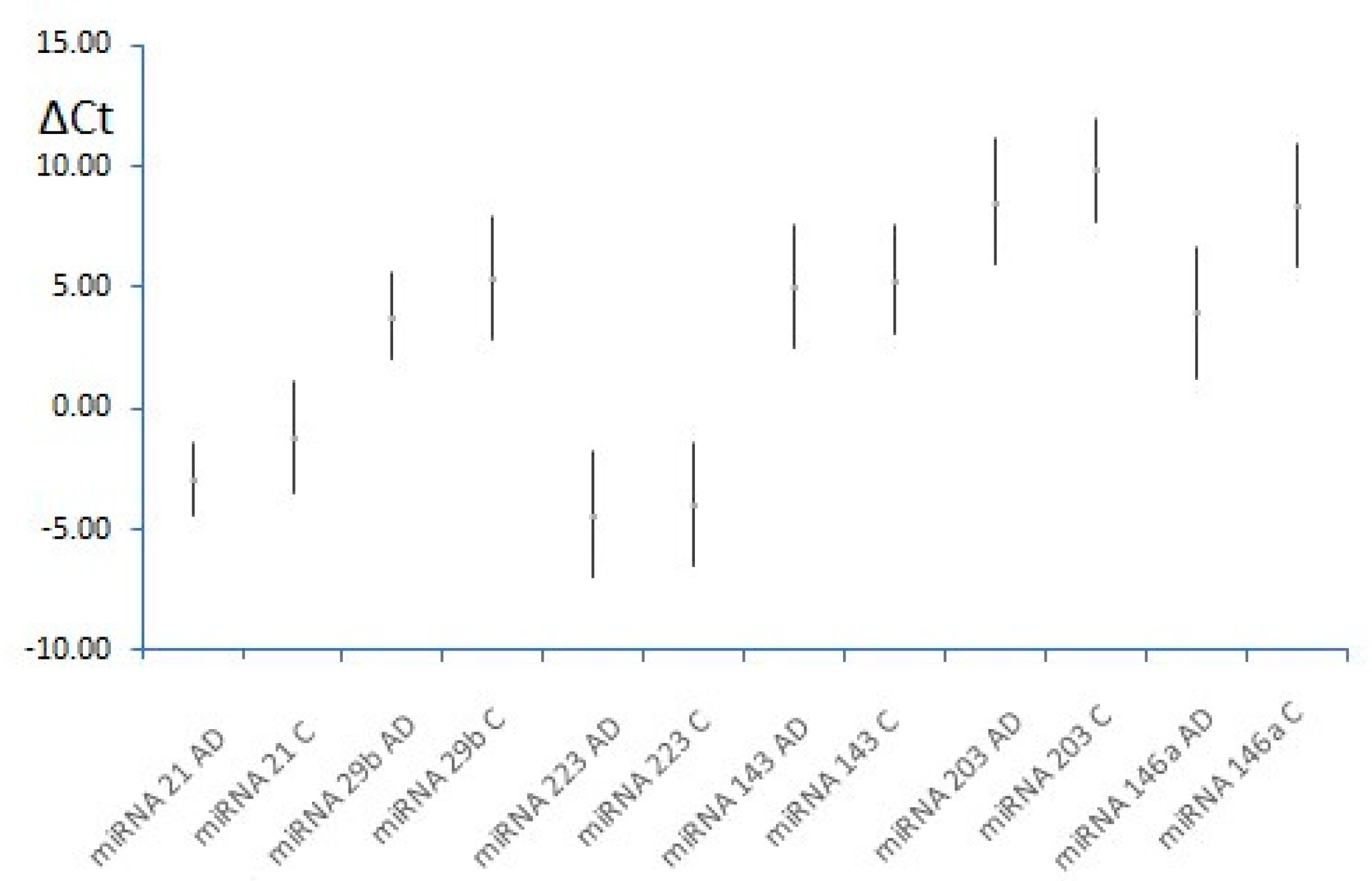
Figure 3.
The comparison of the percentage of eosinophils between the AD and C groups (p= 0.012).

Figure 4.
The comparison of the IgE level between the AD and C groups (p< 0.001).
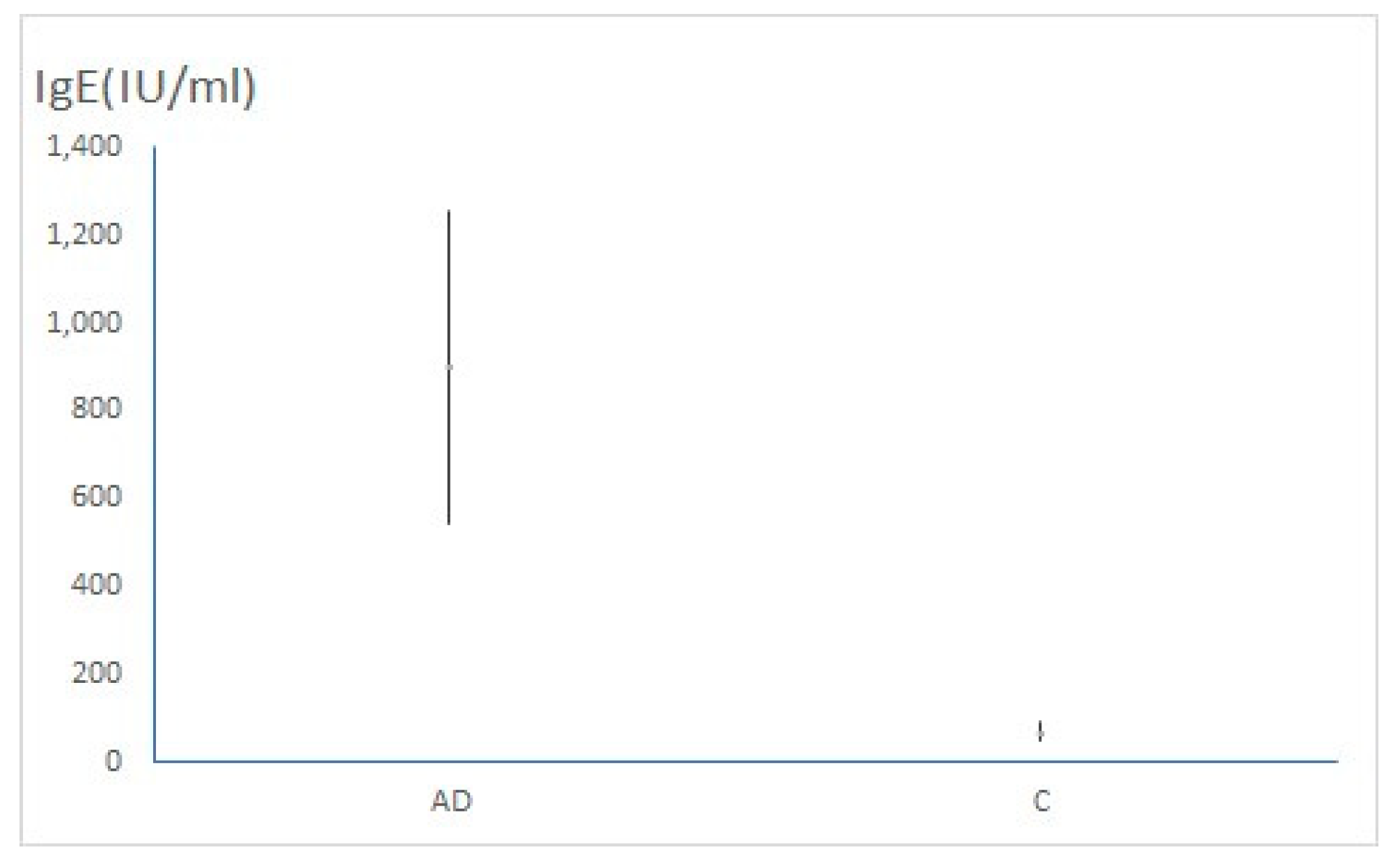
Figure 5.
Correlation between miRNA-223 expression and disease severity (r= 0.342, p= 0.015).
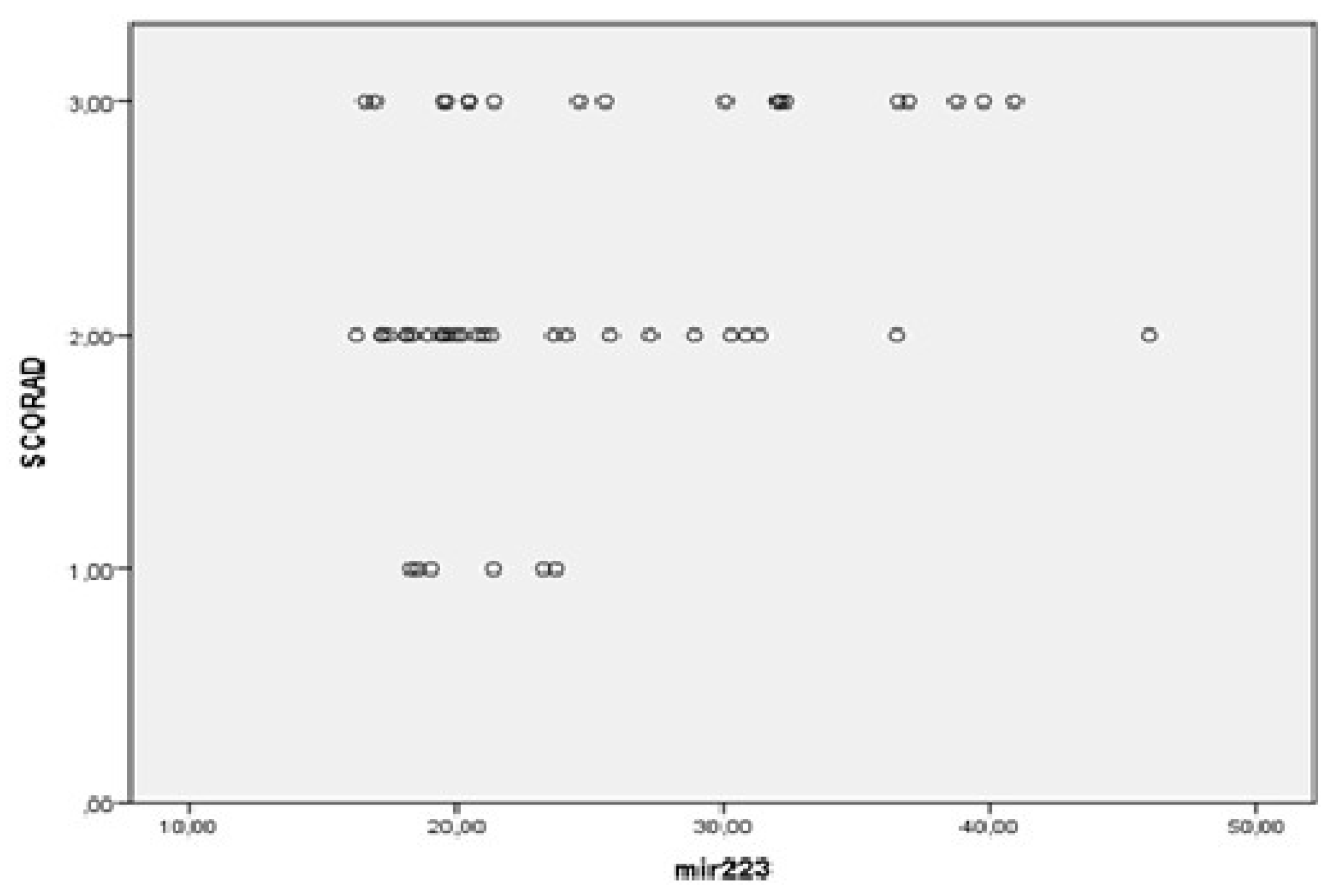
Figure 6.
Association of miRNA-21 expression and disease severity (p < 0.001).
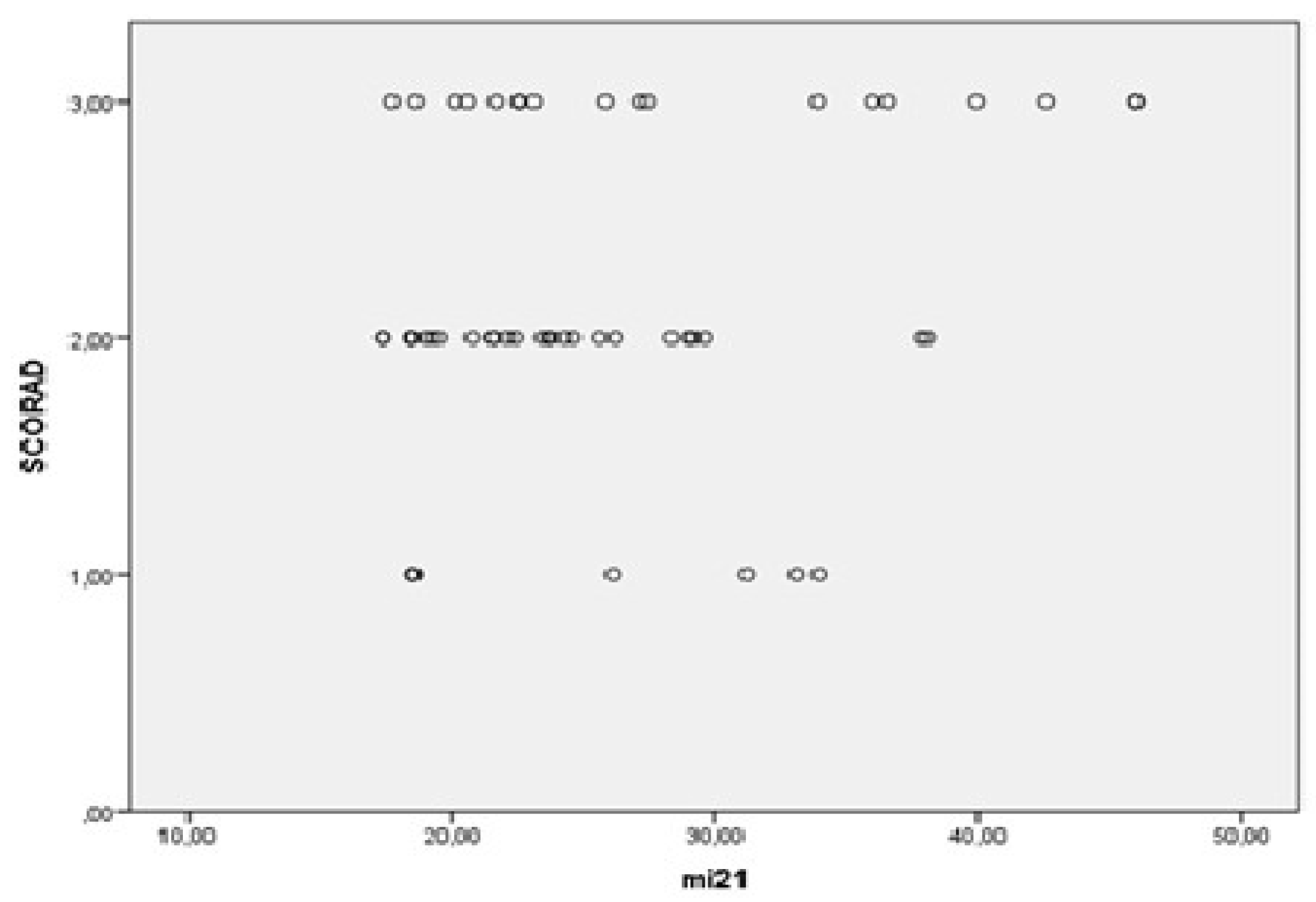
Figure 7.
Association of miRNA-29b expression and disease severity (p < 0.001).
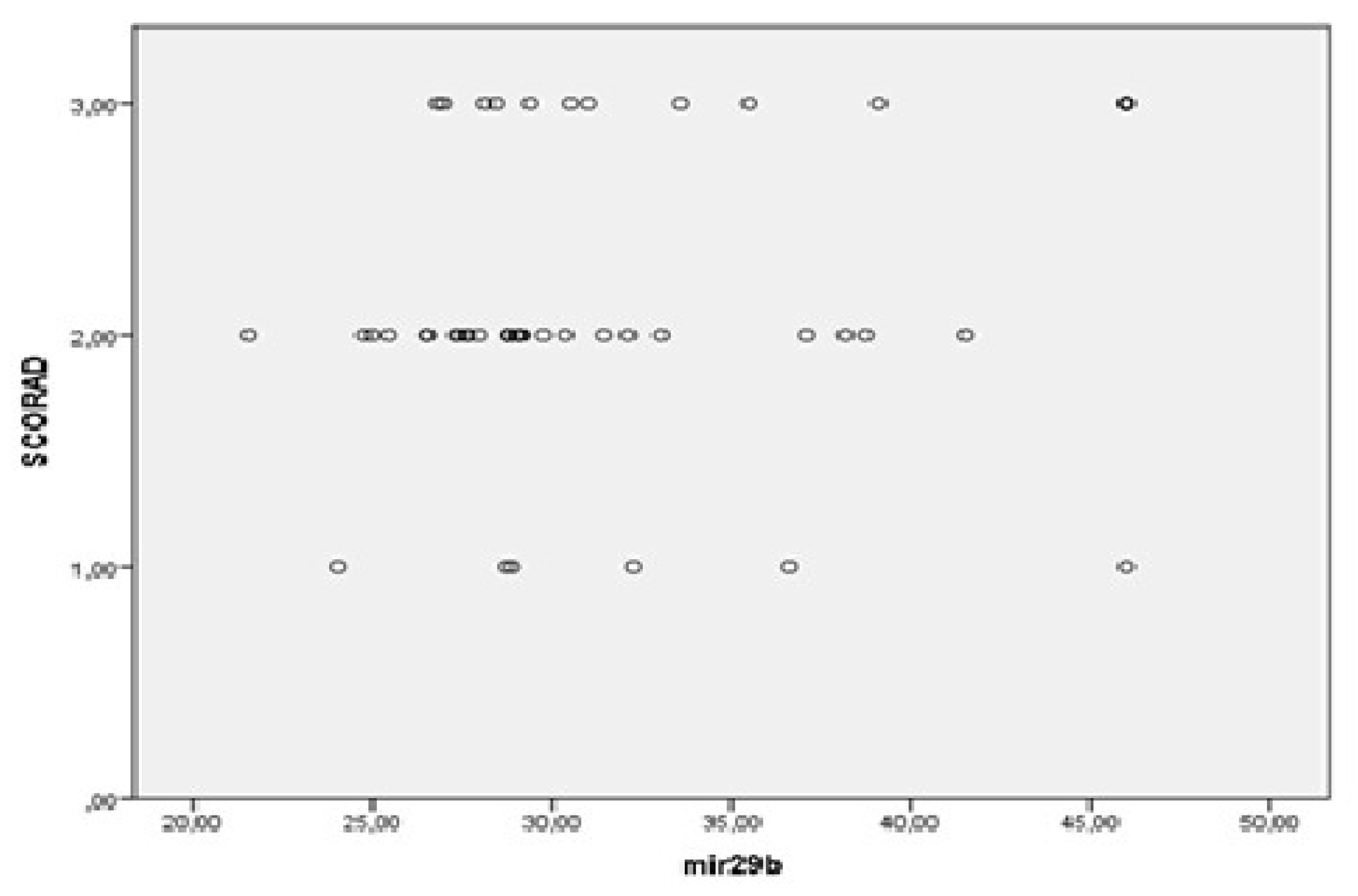
Figure 8.
Association of miRNA-143 expression and disease severity (p < 0.001).
Figure 9.
ROC curve analysis revealed that miRNA-146a is a good biomarker for AD (AUC= 0.7. p= 0.04).
Figure 9.
ROC curve analysis revealed that miRNA-146a is a good biomarker for AD (AUC= 0.7. p= 0.04).

Table 1.
Demographic and clinical information of the study population. AD-atopic dermatitis, AR- allergic rhinitis, As-asthma, FA-food allergy, SCORAD- SCORing Atopic Dermatitis, IgE- immunoglobulin E, Eos- eosinophils, y- year.
Table 1.
Demographic and clinical information of the study population. AD-atopic dermatitis, AR- allergic rhinitis, As-asthma, FA-food allergy, SCORAD- SCORing Atopic Dermatitis, IgE- immunoglobulin E, Eos- eosinophils, y- year.
| Variable | AD patients | Healthy controls | p-value |
|---|---|---|---|
| Total (n) | 50 | 50 | / |
| Gender Male (n, (%)) Female (n, (%)) |
14 (28) 36 (72) |
15 (30) 35 (70) |
0.83 |
| Age, years (average ± 95%CI) | 32.62 (29.22 – 36.02) | 36.4 (33.39 – 39.41) | 0.09 |
| Early onset in childhood (n, (%)) | 8 (16) | / | / |
| Disease duration (year) (average ±95%CI) | 7.22 (4.34 - 10.09) | / | / |
| < 1y (n, (%)) | 13 (26) | / | / |
| > 1-5y (n, (%)) | 20 (40) | / | / |
| > 5-10y (n, (%)) | 5 (10) | / | / |
| > 10y (n, (%)) | 12 (24) | / | / |
| SCORAD 0-24 Mild AD (n, (%)) 25-50 Moderate AD (n, (%)) 51-103 Severe AD (n, (%)) |
6 (12) 26 (52) 18 (36) |
/ / / |
/ |
| Total IgE (IU/ml) (average ± 95%CI) | 899.8 (544.45 – 1255.15) | 69.76 (47.53 – 91.99) | p< 0.001 |
| Eos (%) (average ± 95%CI) | 4.73 (3.51 – 5.96) | 2.68 (2.22 – 3.14) | p= 0.002 |
| Comorbidities AR, yes (n, (%)) As, yes (n, (%)) FA, yes (n, (%)) |
27 (54) 7 (14) 5 (1) |
/ / / |
/ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



