
Submitted:
20 June 2023
Posted:
21 June 2023
You are already at the latest version
Abstract
Atopic dermatitis (AD) has been extensively investigated for genetic associations utilizing both candidate gene approaches and genome-wide scans. Here, we comprehensively evaluated the available literature to determine the association of candidate genes in AD to gain additional insight in the etiopathogenesis of the disease. We systematically screened all studies that explored the association between polymorphisms and AD risks in cases of European and Asian ancestry and synthesized the available evidence through random-effects meta-analysis. We identified 97 studies that met our inclusion/exclusion criteria that examined 17 candidate loci in Europeans and 14 candidate genes in Asians. We confirmed the significant associations between FLG variants in both European and Asian populations and AD risk, while additional synthesis of available data revealed novel loci mapped to IL18 and TGFB1 genes in Europeans and IL12RB1 and MIF in Asians, that have not yet been identified by genome-wide association studies. Our findings provide comprehensive evidence for AD risk loci in cases of both European and Asian ancestries, validating previous associations as well as revealing novel loci that could imply previously unexplored biological pathways.
Keywords:
Atopic Dermatitis
; Meta-analysis
; Eczema
; Association study
; polymorphism
; SNP
; candidate gene
1. Introduction
Atopic dermatitis (AD) is a common, chronic inflammatory cutaneous disease characterized by the development of recurrent eczematous lesions and intense itch [1]. AD affects about 20% of the worldwide infant population, displaying a lower prevalence during adulthood, incorporating both persistent as well as new clinical cases [1]. The increasing prevalence, nevertheless, of AD in the industrialized societies depicts the complex interactions of the stable genetic background with the constant perturbations of the environmental factors; AD is considered as a multifactorial disorder with a strong genetic background, accounting for approximately 75% of the total heritability [2]. Numerous attempts have been conducted in order to decipher the polygenic etiology of AD, examining both the inter-individual variation of AD cases [3,4] as well as under the allergic disease spectrum [5], including asthma, hay fever and AD. The majority of the associated loci lie in the central pathophysiological features of AD, characterized by the epidermal barrier dysfunction and the increased T helper (TH) 2 cell-like inflammatory pattern in the skin [1]. In specific, loss of function mutations in the filaggrin (FLG) gene including the rs558269137 (2282del4) and rs61816761 (R501X) [6] Single Nucleotide Polymorphisms (SNPs), which facilitates the formation of the cornified envelope, have been consistently associated with an increased risk for AD in various ethnic populations, due to their implication in the reduced granular layer [7]. Disease progression is additionally promoted by the participation of microbial alterations and physical damage [1], leading to the increased epithelial permeability and initiation of a type 2 immune response [1], mediated by the TSLP cytokine [8]. Genetic variants mapped in the type 2 cytokine cluster, such as the IL4 rs2243248 [9] and the IL13 rs1800925 [10] SNPs, regulate the secretion of several TH2-related interleukins (ILs), with the major examples of the IL4 and IL13 that have been further targeted by modern therapeutic approaches [11].
Accumulating evidence from genome wide association studies (GWASs) in participants of European ancestry (n=116,863, ncases=21,399) has identified 31 genetic variants that account for less than 20% of the general heritability [3], while extended rare variant approaches have extended the explained variation by 12.56% [4]. Contrastingly, the latest Asian GWAS reported 4 novel Asian-specific AD susceptibility loci from a total of 118,287 participants (ncases=2,639) [12]. However, discrepancies observed between the number of genome-wide scans conducted in both populations [13] limit the representation of diverse ethnic backgrounds in GWAS. In light of these high-throughput approaches, re-evaluation of the candidate gene studies and gene-by-interaction hypotheses is of paramount importance to complement the above results and further characterize the genetic architecture of complex disorders through genotyping of targeted biological pathways. Nevertheless, the relatively small sample size, lack of statistical power and potentially biased reportage of the derived results hinder the conducting of such approaches despite their established efficacy [14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110].
To address this issue, we assessed the inter-individual variability present in the genetic predisposition of AD, incorporating a conservative random effects meta-analysis (REM) approach to synthesize the evidence derived from available studies and thus unveil putative risk loci associated with the disease onset in participants of European [14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64] and Asian [65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110] ancestry.
2. Materials and Methods
2.1. Search strategy and selection criteria
We conducted a systematic literature search to identify studies that examined the inter-individual genetic variability in patients with AD. Searches in the Medline database (through PubMed) using keywords referring to the association (‘association’, ‘susceptibility’) of genetic variants (‘polymorphism’, ‘variant’, ‘SNP’ and related terms) with the AD trait (‘atopic dermatitis’, ‘eczema’) from inception to 18th of September 2022. Screening was performed by two independent authors in a standardized manner, following relevant titles and abstract reading before full-text reviewing. We considered all candidate-gene studies that examined the genetic predisposition of atopic dermatitis through targeted sequencing, regardless of the age of onset and the allergic disease spectrum that includes hay fever, rhinitis and asthma. Studies included in our analysis should be published in peer reviewed journals in the English language; variants incorporated in our meta-analysis should be investigated by at least two eligible studies. We excluded case reports, abstracts, animal studies, duplicate reported data and reports that did not provide the appropriate data for calculation of effect size and confidence intervals.
2.2. Data extraction
Two authors extracted concurrently and independently the eligible studies and extracted the appropriate data; discrepancies were resolved through re-examination of the respective publications until consensus was reached. Data extracted from the eligible studies referred to the first author, publication year, total sample size, and the reported genotypes for further effect size computations.
Odds Ratios (ORs) alongst with their 95% Confidence Intervals (95% CIs) were calculated for the allelic model of inheritance. To avoid the parallel assessment of non-independent SNPs in the same locus, we measured linkage disequilibrium (LD) using the LDpop tool provided by the National Institute of Health (NIH; https://ldlink.nci.nih.gov/?tab=ldpop). R-squared values greater than 0.9 highlighted SNPs in high LD and were therefore considered as a single locus in the meta-analysis.
2.3. Statistical analysis
We calculated OR and 95% CI for each respective heritance pattern by synthesizing the available study-specific evidence through the random effects model. Presence of heterogeneity was assessed with the Cochran’s Q test (considered significant at P-value < 0.1) and quantified via the I2 metric, with 0%< I2<25% reporting small, 25%< I2<50% reporting moderate, 50%< I2<75% reporting high and I2>75% reporting very high heterogeneity. The predominant role of the FLG variants in the AD predisposition led us to further evaluate their combined effect due to their similar biological effects. To explore the inter-study variability, we employed the Harbord’s modified test, a robust statistical analysis for the assessment of small study effects in each meta-analysis. All statistical analyses were performed with the Stata 13.1 software (Stata Corp, TX, USA). The metan plugin was incorporated for our REMs, while the metabias plugin was utilized for the Harbord’s modified test. Statistical significance threshold was set at P-value ≤ 0.05, while the P-value < 0.1 threshold was used for the Harbord’s modified test.
3. Results
3.1. Studies included in our analysis
Our systematic search identified 3,772 studies in the PubMed database. During our screening process, 3,082 were excluded according to both the abstract and inclusion/exclusion criteria, leaving 690 to be thoroughly assessed. Finally, a total of 97 eligible studies met our predefined criteria and were included in the meta-analysis. A brief overview of the information flow is depicted in Figure 1.
Selected characteristics of the available literature are provided at Table 1 and Table 2. In particular, 51 out of 97 studies (53%) examined participants of European ancestry [14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64] (Table 1) and 46 (47%) of Asian ancestry [65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110] (Table 2). Studies conducted in European participants focused on the FLG [14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38] (n=25, 26%), TLR2 [39,40,41,42,43,44,45] (n=7, 7%), IL10 [46,47,48,49,50,51,52] (n=7, 7%), 11q13.5 [23,38,53,54,55] (n=5, 5%), IL13 [18,47,50,56] (n=4, 4%), IL4 [47,48,49] (n=3, 3%), HRNR [23,54,57] (n=3, 3%) and IL6 [48,49,51] (n=3, 3%) loci, while evidence for possible association for relevant genes were examined by a total of 2 studies [18,40,42,48,49,51,52,58,59,60,61,62,63,64] (Table 1). Similarly, Asian studies primarily investigated SNPs mapped in the FLG [65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82] (n=18, 19%), IL4 [83,84,85,86,87,88,89] (n=7, 7%) and SPINK5 [65,90,91,92] (n=4, 4%) genes, with the rest of the explored loci assessed by 3 or 2 studies [65,83,85,86,88,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110] (Table 2).
3.2. Candidate gene approaches in cases of European ancestry
3.2.1. FLG gene
We confirmed all significant associations between the FLG loss of function (LOF) variants and increased AD risk from a total of 25 studies, incorporating 16,965 participants. Specifically, both FLG rs558269137 (OR (95% CI): 0.28 (0.21-0.36); I2 = 70.5%) and FLG rs61816761 (OR (95% CI): 0.25 (0.19-0.33); I2 = 66.3%) common alleles were associated with reduced AD risk from a total of 23 studies showing, however, a high heterogeneity (Figure 2; Supplementary Table S1). In addition, the rare alleles of the FLG rs138726443 (11 studies; n=10,794), FLG rs150597413 (7 studies; n=8,126) and FLG rs397507563 (3 studies, n=2,780) SNPs yielded significant associations with higher risk of developing AD (OR (95% CI): 0.27 (0.18-0.40); I2 = 13.2%; OR (95% CI): 0.32 (0.21-0.50); I2 = 0.0%; OR (95% CI): 0.14 (0.03-0.69); I2 = 0.0%) (Figure 2; Supplementary Table S1).
The predominant role of the FLG locus in the disease onset as well as the similar, trans-acting biological mechanism of the above variants [32] prompted us to investigate their effect under the spectrum of combined genotypes. Three combined genotypes (CGs) were identified from the included studies (CG1: rs558269137 and rs61816761; CG2: rs558269137, rs61816761 and rs138726443; CG3: rs558269137, rs61816761, rs138726443 and rs150597413); CG1 incorporated 5,263 participants from 9 studies, CG2 included 2,264 AD cases from 3 studies and CG3 was assessed in 5,391 eczema patients from 4 studies. As expected, all CGs yielded significant associations (OR (95% CI): 0.19 (0.13-0.27); I2 = 68.7%; OR (95% CI): 0.33 (0.17-0.63); I2 = 36.8%; OR (95% CI): 0.23 (0.15-0.36); I2 = 81.0%) highlighting thus the contribution of the FLG locus in the disease predisposition, nevertheless reporting a high heterogeneity metric (Figure 2; Supplementary Table S1).
3.2.2. TLR2 gene
Our REM did not reveal a significant association between the TLR2 rs5743708 polymorphism and disease onset from a total of 1,257 participants (OR (95% CI): 0.61 (0.29-1.30); I2 = 30.7%). Similarly, synthesis of evidence from 3 studies evaluating the TLR2 rs4696480 SNP in 741 participants failed to establish a significant association with the risk of developing AD (OR (95% CI): 0.81 (0.62-1.06); I2 = 40.2%) (Figure 2; Supplementary Table S1).
3.2.3. IL10 gene
Comparably to the TLR2 locus, our meta-analysis did not detect significant associations between the IL10 rs1800896 variant and AD predisposition (OR (95% CI): 1.06 (0.89-1.27); I2 = 29.6%), using data from 7 studies with 1,703 participants (Figure 1). Notably, the IL10 rs1800871 and IL10 rs1800872 SNP were found in high LD SNPs (r2 > 0.9) and were consequently assessed as a single genotype; we did not observe significant associations between these variants and disease risk from a total of 3 studies (n=1,332; OR (95% CI): 1.08 (0.72-1.64); I2 = 76.7%) (Figure 2; Supplementary Table S1).
3.2.4. 11. q13.5 locus
The rs7927894 variant mapped to the 11q13.5 locus demonstrated a significant association with AD risk, based on 5 studies with 5,506 participants (OR (95% CI): 0.78 (0.71-0.85); I2 = 0.0%) (Figure 2; Supplementary Table S1).
3.2.5. IL13 gene
Our systematic search identified 4 studies examining the IL13 rs1800925 SNP (n=931); synthesis of the available data depicted the association of the common rs1800925 SNP with reduced AD risk (OR (95% CI): 0.60 (0.48-0.75); I2 = 0.0%) (Figure 2; Supplementary Table S1).
3.2.6. SNPs in IL4, IL18 and TGFB1 genes
The common allele of IL4 rs2243248 was significantly associated with higher risk of developing AD, including data from 2 studies with a total of 503 participants (OR (95% CI): 2.67 (1.53- 4.65); I2 = 0.0%). Contrastingly, the IL4 rs2243250, assessed in 3 studies (n=639), did not yield a significant outcome (OR (95% CI): 0.81 (0.59-1.13); I2 = 3.5%). Two studies further assessed SNPs mapped to the IL18 (rs187238; n=388) and TGFB1 (rs1800471, rs1800470; n=461) genes, providing significant associations with the exception of the TGFB1 rs1800470 variant (Figure 2; Supplementary Table S1).
3.2.7. SNPs in IL6, HRNR, VDR, IL1B, TNF, SPINK5, IL4RA, TLR4 and GSTP1 genes
Two variants (IL6 rs1800795 and rs1800797) were found in high LD and were therefore considered together during data synthesis; meta-analysis from 3 studies (n=1,432) assessing these variants did not demonstrate significant association between the common allele and AD risk (OR (95% CI): 1.08 (0.89-1.31); I2 = 16.7%). Similarly, the HRNR rs877776 common allele was not associated with disease onset from a total of 3 studies incorporating 2,158 participants (OR (95%CI): 0.91 (0.62-1.34); I2 = 72.9%). Additional investigation of putative risk loci through meta-analysis in two studies, including VDR rs2228570, rs7975232, VDR rs1544410 and rs731236 (r2 > 0.9), IL1B rs16944, IL1B rs1143634, TNF rs1800629, TNF rs361525, SPINK5 rs2303067, IL4RA rs1801275, TLR4 rs4986790 and rs4986791 (r2 > 0.9) and GSTP1 rs1695 gave null results (Figure 2; Supplementary Table S1).
3.3. Candidate gene approaches in cases of Asian ancestry
3.3.1. FLG gene
Similarly to the studies assessing population of European descent, we validated significant associations between FLG LOF variants and AD risk in participants of Asian ancestry. In particular, the FLG rs200519781 frameshift variant was associated with increased AD risk from a total of 14 studies (n=7,704; OR (95% CI): 0.32; (0.25-0.42); I2 = 0.0%). As far as FLG rs121909626 (9 studies; n=4,054) and FLG rs761212672 (6 studies; n=2,092) SNPs are concerned, both yielded significant associations with higher eczema risk (OR (95% CI): 0.19 (0.11-0.34); I2 = 0.0%; OR (95% CI): 0.14 (0.04-0.44); I2 = 0.0%). Our REM highlighted 2 additional FLG variants (rs145738429, 5 studies, n=1,857; S2889X, 6 studies, n=1,736) associated with increased AD risk (OR (95% CI): 0.23 (0.06-0.89); I2 = 0.0%; OR (95% CI): 0.14 (0.07-0.27); I2 = 0.0%). Notably, the FLG rs558269137 variant was further associated in participants of Asian ancestry from a total of 2 studies including 1,382 cases (OR (95% CI): 0.43 (0.24-0.79); I2 = 0.0%) (Figure 3; Supplementary Table S2).
Despite the abundance of statistically significant associations between FLG variants and increased AD risk in both Europeans and Asians, several variants including FLG rs61816761 (7 studies; n=3,085), FLG rs146466242 (5 studies; n=1,930), FLG rs772851618 (5 studies; n=1,857) and FLG rs11584340 (2 studies; n=855) failed to provide significant signals (OR (95% CI): 0.45 (0.11-1.74); I2 = 0.0%; OR (95% CI): 0.54 (0.21-1.38); I2 = 1.7%; OR (95% CI): 2.39 (0.54-10.53); I2 = 0.0%; OR (95% CI): 0.80 (0.44-1.47); I2 = 86.5%) (Figure 3; Supplementary Table S2).
Regarding Asians, we identified one CG from the included studies (CG: rs200519781, rs121909626, rs761212672, rs145738429, S2889X, rs61816761, rs146466242 and rs772851618); CG incorporated 877 participants from 3 studies and yielded a significant association (OR (95% CI): 0.11 (0.06-0.19); I2 = 0.0%) (Figure 3; Supplementary Table S2).
3.3.2. IL4 gene
Two variants mapped to the IL4 gene (rs2243250, rs2070874) were found in high LD (r2 > 0.9) and were thus examined together from a total of 7 studies (n=2,454). Our REM did not demonstrate a significant association (OR (95% CI): 0.97 (0.64-1.48); I2 = 88.0%) (Figure 3; Supplementary Table S2).
3.3.3. SPINK5 gene
Comparably to IL4 variants, 2 SPINK5 variants (rs2303063, rs2303067) were found in high LD (r2 > 0.9) and were thus examined together from a total of 3 studies (n=1,280). However, we found that the above locus was significantly associated with AD risk (OR (95% CI): 1.26 (1.06-1.48); I2 = 0.0%). Relevant, independent variants mapped to the SPINK5 gene yielded additional significant signals, with the exemplars of SPINK5 rs2303070 (3 studies, n=741; OR (95% CI): 0.65 (0.51- 0.83); I2 = 0.0%) and SPINK5 rs2303065 (two studies, n=546; OR (95% CI: 0.74 (0.57- 0.96); I2 = 0.0%). Nevertheless, data synthesis from 4 studies (n=930) regarding the SPINK5 rs2303064 SNP showed no evidence for association with AD predisposition in the Asian population (OR (95% CI): 0.97 (0.79-1.19); I2 = 0.0%) (Figure 3; Supplementary Table S2).
3.3.4. SNPs in IL10, IL4RA and IL13 genes
Three studies assessed the association of IL10 rs1800896, IL10 rs1800871 rs1800872 (r2 > 0.9), IL4RA rs1801275, IL4RA rs1805010 and IL13 rs20541, while 2 studies assessed the IL13 rs1800925 in AD Asians. Our REM gave null results in all the above cases (Figure 3; Supplementary Table S2).
3.3.5. SNPs in IL12RB1, IL9 and MIF genes
Considering the IL12RB1 rs393548 and rs436857 variants, two studies assessed their association with disease onset (n=1,339, n=1,326 respectively). Our meta-analysis yielded statistically significant associations in both cases (OR (95% CI): 0.80 (0.66-0.96); I2 = 0.0%; OR (95% CI): 0.79 (0.65-0.95); I2 = 0.0%) (Figure 3; Supplementary Table S2). In addition, synthesis of the available data derived from 2 studies regarding the IL9 rs31563 (n=1,391) and MIF rs755622 (n=649) SNPs provided significant associations with AD risk (OR (95% CI): 1.32 (1.07- 1.64); I2 = 0.0%; OR (95% CI): 0.65 (0.49-0.86); I2 = 0.0%) (Figure 3; Supplementary Table S2).
3.3.6. SNPs in IL5RA, IL18, TNF, FCER1A and TARC genes
Lastly, 2 studies examined the association between variants mapped to the IL5RA (rs334809), TNF (rs1800629), FCER1A (rs2427837) and TARC (431C>T) genes, generating non-significant results (Figure 3; Supplementary Table S2). Remarkably, despite the significant association in cases of European descent, the IL18 rs187238 was not associated with AD risk in the Asian population (OR (95% CI): 1.52 (0.96-2.42); I2 = 0.0%) (Figure 3; Supplementary Table S2).
Harbord’s Modified test identified small study effect biases in 6 cases. In specific, we found a small study effect bias in the REM of FLG rs150597413 (P-value = 0.088), TLR2 rs4696480 (P-value = 0.006), 11q13.5 rs7927894 (P-value = 0.062) and TLR4 rs4986790, rs4986791 (P-value = 0.019) in European cases (Figure 2), while significant biases were found in the FLG rs121909626 (P-value = 0.049) and FLG rs772851618 (P-value = 0.021) in Asians (Figure 3).
4. Discussion
Here, we performed a systematic review and meta-analysis of all available evidence considering the genetic predisposition to AD in participants of both European and Asian ancestry. Despite the expanding list of risk loci identified through genome-wide scans in AD, our REM shed light upon new risk loci mapped to the IL18 and TGFB1 genes in Europeans (Figure 2), while the contribution of IL12RB1 and MIF loci in AD predisposition in Asians was additionally characterized through our analysis, results not previously reported by genome-wide scans (Figure 3). We thus provide novel insights into the genetic architecture of the disease, highlighting perturbed biological pathways that could be further implicated in the development of new therapeutic mechanisms.
The present meta-analysis confirms the pivotal role of the FLG locus in disease susceptibility, as LOF mutations disrupt the production of the filaggrin protein in the stratum corneum. The impaired barrier function facilitates loss of water, pH increase as well as the entry of irritants and allergens, contributing to the onset and exacerbation of allergic reactions [111]. The consistency of these findings across European and Asian populations highlights the universal importance of FLG in maintaining skin homeostasis and its relevance in AD [112]. The presence of significant heterogeneity concerning FLG rs558269137 and rs61816761 in European population was also observed in a prior meta-analysis of FLG polymorphisms in AD [113], which could be attributed to several demographic factors and environmental exposures that mediate the increased atopic eczema risk [111]. We further validate the 11q13.5 rs7927894 SNP as a risk locus, a variant localized within a non-coding genomic region. Notably, the above variant has been additionally implicated in Crohn's disease and deregulated epithelial function, thus potentially contributing to the pathogenic mechanisms underlying AD [53]. The cytokine cluster located at 5q31, harboring both IL4 and IL13, represents a prominent risk locus implicated in the pathogenesis of the disease and has been extensively linked with AD through several GWAS-derived variants. Here, we identified the IL13 rs1800925 SNP, a regulatory variant that enhances the activity of the IL13 promoter in human Th2 lymphocytes, thereby augmenting the risk of allergic disorders [10]. Remarkably, this particular variant further exhibits a significant association with cutaneous T-cell lymphoma [114], a malignancy that has been observed to manifest in some cases of AD patients following the administration of dupilumab [115], an anti-IL4RA biological drug that inhibits both IL4 and IL13 signaling [11]. Another significantly associated variant that belongs to the 5q31 locus is the IL4 rs2243248 SNP, where our REM reported the protective effect of the minor rs2243248 G allele in AD risk, similarly to asthma [9]. Relevant, previously identified risk variants refer to the SPINK5 nonsynonymous variants that disrupt the function of serine protease inhibitors in the integrity of the epidermal barrier, as well as the IL9 rs31563 variant that exhibits a regulatory role in IL9 expression, a cytokine known to contribute to B cell isotype switching from IgM to IgE synthesis [100].
Despite the prominence of GWAS outcomes as the gold standard in genetic association studies, there are still certain risk loci that have not been yet identified. We further report four novel loci, two belonging to Europeans (Figure 2) and two Asian-specific (Figure 3). In particular, we detected a significant association between the IL18 rs187238 polymorphism and AD risk in European cases (Figure 2); this variant, residing within the IL18 promoter region, supports the modulation of the binding affinity of transcription factors and thus participates in the dysregulated Th2 response [116]. Our second novel addition in Europeans is the TGFB1 rs1800471 missense variant (Figure 2) that significantly alters the inhibitory action of TGFB1 protein molecule during allergic reactions [117].
Additionally, the promoter IL12RB1 rs393548 and rs436857 variants were significantly associated with AD risk in Asians (Figure 3); the IL12RB1 receptor subunit is involved in the modulation of the IL12-dependent inhibition of IgE synthesis and Th2 cell function. Consequently, potential decrease of the IL12RB1 expression perturbs the above inhibitory activity [99], thus leading to the dysregulation of downstream immune responses and potentially contributing to allergic reactions. Finally, our findings indicate a significant association between the MIF rs755622 C allele and increased AD susceptibility (Figure 3); the rare C allele exhibits enhanced promoter activity relatively to the common G allele, leading to differential MIF expression levels [101]. Studies conducted on mouse models of AD have demonstrated that MIF holds a significant role in the development of AD-related immune dysregulation through the induction of a type-2 immune response, as well as fostering the recruitment of eosinophils in the cutaneous inflammation [118].
However, our study displays some constraints. Despite the assessment of small study effect bias through the Harbord’s modified test and identification of 6 significant cases, the majority of studies included in our synthesis incorporate a relatively small sample size and are thus susceptible to biases [119]. In addition, discrepancies between the available data derived from the included studies, with the exemplars of disease activity, age of onset and relevant clinical information restricted further stratification and subgroup examination in our analysis. The predominant role of the exposome in the AD predisposition [111], and inclusion of such environmental risk factors in multivariate analyses is of paramount importance to gain a holistic understanding of AD etiopathology and thus unveil novel therapeutic targets.
In conclusion, we conducted a systematic review and meta-analysis of all available data regarding the genetic predisposition to AD in participants of both European and Asian ancestry. We report four novel risk loci that have not been previously shown by GWAS, partially contributing thus to the elucidation of the genetic architecture of the disease. By exploring data from both ethnic groups, our study endeavors to provide a broader understanding and enhance the applicability of our findings across diverse populations. High-throughput investigation of putative risk loci in AD [120] could further unveil their functional role in the disease predisposition. In addition, incorporation of clinical information for each included participant, as well as the vast amount of environmental exposures associated with increased AD risk shall facilitate the interpretation of disease-associated interactions and form the framework for precision medicine.
Supplementary Materials
Supplementary Table S1. Association between single nucleotide polymorphisms and atopic dermatitis susceptibility in patients of European descent. Supplementary Table S2. Association between single nucleotide polymorphisms and atopic dermatitis susceptibility in patients of Asian descent.
Author Contributions
Conceptualization, C.A. and Y.V.; methodology, A.P., C.A., E.E. and Y.V.; software, formal analysis, A.P., C.A.; data curation, A.P., C.A.; writing—original draft preparation, A.P.; writing—review and editing, A.P., C.A., E.E. and Y.V.; visualization, A.P.; supervision, Y.V. All authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Langan, S.M.; Irvine, A.D.; Weidinger, S. Atopic Dermatitis. Lancet 2020, 396, 345–360. [Google Scholar] [CrossRef]
- Thomsen, S.F.; Ulrik, C.S.; Kyvik, K.O.; Hjelmborg, J. v B.; Skadhauge, L.R.; Steffensen, I.; Backer, V. Importance of Genetic Factors in the Etiology of Atopic Dermatitis: A Twin Study. Allergy Asthma Proc 2007, 28, 535–539. [CrossRef]
- Paternoster, L.; Standl, M.; Waage, J.; Baurecht, H.; Hotze, M.; Strachan, D.P.; Curtin, J.A.; Bønnelykke, K.; Tian, C.; Takahashi, A.; et al. Multi-Ancestry Genome-Wide Association Study of 21,000 Cases and 95,000 Controls Identifies New Risk Loci for Atopic Dermatitis. Nat Genet 2015, 47, 1449–1456. [Google Scholar] [CrossRef] [PubMed]
- Mucha, S.; Baurecht, H.; Novak, N.; Rodríguez, E.; Bej, S.; Mayr, G.; Emmert, H.; Stölzl, D.; Gerdes, S.; Jung, E.S.; et al. J Allergy Clin Immunol 2020, 145, 1208–1218. [CrossRef] [PubMed]
- Ferreira, M.A.R.; Vonk, J.M.; Baurecht, H.; Marenholz, I.; Tian, C.; Hoffman, J.D.; Helmer, Q.; Tillander, A.; Ullemar, V.; Lu, Y.; et al. Eleven Loci with New Reproducible Genetic Associations with Allergic Disease Risk. J Allergy Clin Immunol 2019, 143, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Hoober, J.K.; Eggink, L.L. The Discovery and Function of Filaggrin. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Kypriotou, M.; Huber, M.; Hohl, D. The Human Epidermal Differentiation Complex: Cornified Envelope Precursors, S100 Proteins and the “fused Genes” Family. Exp Dermatol 2012, 21, 643–649. [Google Scholar] [CrossRef]
- Wynn, T.A. Type 2 Cytokines: Mechanisms and Therapeutic Strategies. Nat Rev Immunol 2015, 15, 271–282. [Google Scholar] [CrossRef]
- Amirzargar, A.A.; Movahedi, M.; Rezaei, N.; Moradi, B.; Dorkhosh, S.; Mahloji, M.; Mahdaviani, S.A. Polymorphisms in IL4 and ILARA Confer Susceptibility to Asthma. J Investig Allergol Clin Immunol 2009, 19, 433–438. [Google Scholar]
- Cameron, L.; Webster, R.B.; Strempel, J.M.; Kiesler, P.; Kabesch, M.; Ramachandran, H.; Yu, L.; Stern, D.A.; Graves, P.E.; Lohman, I.C.; et al. Th2 Cell-Selective Enhancement of Human IL13 Transcription by IL13-1112C>T, a Polymorphism Associated with Allergic Inflammation. J Immunol 2006, 177, 8633–8642. [Google Scholar] [CrossRef]
- Gooderham, M.J.; Hong, H.C.-H.; Eshtiaghi, P.; Papp, K.A. Dupilumab: A Review of Its Use in the Treatment of Atopic Dermatitis. J Am Acad Dermatol 2018, 78, S28–S36. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Koido, M.; Suzuki, A.; Otomo, N.; Suetsugu, H.; Kochi, Y.; Tomizuka, K.; Momozawa, Y.; Kamatani, Y.; Ikegawa, S.; et al. Eight Novel Susceptibility Loci and Putative Causal Variants in Atopic Dermatitis. J Allergy Clin Immunol 2021, 148, 1293–1306. [Google Scholar] [CrossRef]
- Brown, S.J. What Have We Learned from GWAS for Atopic Dermatitis? J Invest Dermatol 2021, 141, 19–22. [Google Scholar] [CrossRef]
- Vardar Acar, N.; Cavkaytar, Ö.; Arik Yilmaz, E.; Büyüktiryaki, A.B.; Uysal Soyer, Ö.; Şahiner, Ü.M.; Şekerel, B.E.; Karaaslan, I.Ç.; Saçkesen, C. Rare Occurrence of Common Filaggrin Mutations in Turkish Children with Food Allergy and Atopic Dermatitis. Turk J Med Sci 2020, 50, 1865–1871. [Google Scholar] [CrossRef]
- Jurakic Toncic, R.; Kezic, S.; Jakasa, I.; Ljubojevic Hadzavdic, S.; Balic, A.; Petkovic, M.; Pavicic, B.; Zuzul, K.; Marinovic, B. Filaggrin Loss-of-Function Mutations and Levels of Filaggrin Degradation Products in Adult Patients with Atopic Dermatitis in Croatia. J Eur Acad Dermatol Venereol 2020, 34, 1789–1794. [Google Scholar] [CrossRef]
- González-Tarancón, R.; Sanmartín, R.; Lorente, F.; Salvador-Rupérez, E.; Hernández-Martín, A.; Rello, L.; Puzo, J.; Gilaberte, Y. Prevalence of FLG Loss-of-Function Mutations R501X, 2282del4, and R2447X in Spanish Children with Atopic Dermatitis. Pediatr Dermatol 2020, 37, 98–102. [Google Scholar] [CrossRef]
- Woźniak, M.; Kaczmarek-Skamira, E.; Romańska-Gocka, K.; Czajkowski, R.; Kałużna, L.; Zegarska, B. The Prevalence of Mutations in the Gene Encoding Filaggrin in the Population of Polish Patients with Atopic Dermatitis. Postepy Dermatol Alergol 2016, 33, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Trzeciak, M.; Gleń, J.; Rębała, K.; Bandurski, T.; Sikorska, M.; Nowicki, R. Coexistence of 2282del4 FLG Gene Mutation and IL-18 -137G/C Gene Polymorphism Enhances the Risk of Atopic Dermatitis. Postepy Dermatol Alergol 2016, 33, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Trzeciak, M.; Wesserling, M.; Bandurski, T.; Glen, J.; Nowicki, R.; Pawelczyk, T. Association of a Single Nucleotide Polymorphism in a Late Cornified Envelope-like Proline-Rich 1 Gene (LELP1) with Atopic Dermatitis. Acta Derm Venereol 2016, 96, 459–463. [Google Scholar] [CrossRef]
- Ballardini, N.; Kull, I.; Söderhäll, C.; Lilja, G.; Wickman, M.; Wahlgren, C.F. Eczema Severity in Preadolescent Children and Its Relation to Sex, Filaggrin Mutations, Asthma, Rhinitis, Aggravating Factors and Topical Treatment: A Report from the BAMSE Birth Cohort. Br J Dermatol 2013, 168, 588–594. [Google Scholar] [CrossRef]
- Ercan, H.; Ispir, T.; Kirac, D.; Baris, S.; Ozen, A.; Oztezcan, S.; Cengizlier, M.R. Predictors of Atopic Dermatitis Phenotypes and Severity: Roles of Serum Immunoglobulins and Filaggrin Gene Mutation R501X. Allergol Immunopathol (Madr) 2013, 41, 86–93. [Google Scholar] [CrossRef]
- Mlitz, V.; Latreille, J.; Gardinier, S.; Jdid, R.; Drouault, Y.; Hufnagl, P.; Eckhart, L.; Guinot, C.; Tschachler, E. Impact of Filaggrin Mutations on Raman Spectra and Biophysical Properties of the Stratum Corneum in Mild to Moderate Atopic Dermatitis. J Eur Acad Dermatol Venereol 2012, 26, 983–990. [Google Scholar] [CrossRef] [PubMed]
- O’Regan, G.M.; Campbell, L.E.; Cordell, H.J.; Irvine, A.D.; McLean, W.H.I.; Brown, S.J. Chromosome 11q13.5 Variant Associated with Childhood Eczema: An Effect Supplementary to Filaggrin Mutations. J Allergy Clin Immunol 2010, 125, 170–174.e1-2. [Google Scholar] [CrossRef] [PubMed]
- Greisenegger, E.; Novak, N.; Maintz, L.; Bieber, T.; Zimprich, F.; Haubenberger, D.; Gleiss, A.; Stingl, G.; Kopp, T.; Zimprich, A. Analysis of Four Prevalent Filaggrin Mutations (R501X, 2282del4, R2447X and S3247X) in Austrian and German Patients with Atopic Dermatitis. J Eur Acad Dermatol Venereol 2010, 24, 607–610. [Google Scholar] [CrossRef]
- Gao, P.-S.; Rafaels, N.M.; Hand, T.; Murray, T.; Boguniewicz, M.; Hata, T.; Schneider, L.; Hanifin, J.M.; Gallo, R.L.; Gao, L.; et al. Filaggrin Mutations That Confer Risk of Atopic Dermatitis Confer Greater Risk for Eczema Herpeticum. J Allergy Clin Immunol 2009, 124, 507–513, 513.e1-7. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.J.; Relton, C.L.; Liao, H.; Zhao, Y.; Sandilands, A.; Wilson, I.J.; Burn, J.; Reynolds, N.J.; McLean, W.H.I.; Cordell, H.J. Filaggrin Null Mutations and Childhood Atopic Eczema: A Population-Based Case-Control Study. J Allergy Clin Immunol 2008, 121, 940–946.e3. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.J.; Sandilands, A.; Zhao, Y.; Liao, H.; Relton, C.L.; Meggitt, S.J.; Trembath, R.C.; Barker, J.N.W.N.; Reynolds, N.J.; Cordell, H.J.; et al. Prevalent and Low-Frequency Null Mutations in the Filaggrin Gene Are Associated with Early-Onset and Persistent Atopic Eczema. J Invest Dermatol 2008, 128, 1591–1594. [Google Scholar] [CrossRef]
- Giardina, E.; Paolillo, N.; Sinibaldi, C.; Novelli, G. R501X and 2282del4 Filaggrin Mutations Do Not Confer Susceptibility to Psoriasis and Atopic Dermatitis in Italian Patients. Dermatology 2008, 216, 83–84. [Google Scholar] [CrossRef]
- Weidinger, S.; O’Sullivan, M.; Illig, T.; Baurecht, H.; Depner, M.; Rodriguez, E.; Ruether, A.; Klopp, N.; Vogelberg, C.; Weiland, S.K.; et al. Filaggrin Mutations, Atopic Eczema, Hay Fever, and Asthma in Children. J Allergy Clin Immunol 2008, 121, 1203–1209.e1. [Google Scholar] [CrossRef]
- Rogers, A.J.; Celedón, J.C.; Lasky-Su, J.A.; Weiss, S.T.; Raby, B.A. Filaggrin Mutations Confer Susceptibility to Atopic Dermatitis but Not to Asthma. J Allergy Clin Immunol 2007, 120, 1332–1337. [Google Scholar] [CrossRef]
- Lerbaek, A.; Bisgaard, H.; Agner, T.; Ohm Kyvik, K.; Palmer, C.N.A.; Menné, T. Filaggrin Null Alleles Are Not Associated with Hand Eczema or Contact Allergy. Br J Dermatol 2007, 157, 1199–1204. [Google Scholar] [CrossRef] [PubMed]
- Sandilands, A.; Terron-Kwiatkowski, A.; Hull, P.R.; O’Regan, G.M.; Clayton, T.H.; Watson, R.M.; Carrick, T.; Evans, A.T.; Liao, H.; Zhao, Y.; et al. Comprehensive Analysis of the Gene Encoding Filaggrin Uncovers Prevalent and Rare Mutations in Ichthyosis Vulgaris and Atopic Eczema. Nat Genet 2007, 39, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, S.; Rodríguez, E.; Stahl, C.; Wagenpfeil, S.; Klopp, N.; Illig, T.; Novak, N. Filaggrin Mutations Strongly Predispose to Early-Onset and Extrinsic Atopic Dermatitis. J Invest Dermatol 2007, 127, 724–726. [Google Scholar] [CrossRef] [PubMed]
- Marenholz, I.; Nickel, R.; Rüschendorf, F.; Schulz, F.; Esparza-Gordillo, J.; Kerscher, T.; Grüber, C.; Lau, S.; Worm, M.; Keil, T.; et al. Filaggrin Loss-of-Function Mutations Predispose to Phenotypes Involved in the Atopic March. J Allergy Clin Immunol 2006, 118, 866–871. [Google Scholar] [CrossRef]
- Stemmler, S.; Parwez, Q.; Petrasch-Parwez, E.; Epplen, J.T.; Hoffjan, S. Two Common Loss-of-Function Mutations within the Filaggrin Gene Predispose for Early Onset of Atopic Dermatitis. J Invest Dermatol 2007, 127, 722–724. [Google Scholar] [CrossRef]
- Barker, J.N.W.N.; Palmer, C.N.A.; Zhao, Y.; Liao, H.; Hull, P.R.; Lee, S.P.; Allen, M.H.; Meggitt, S.J.; Reynolds, N.J.; Trembath, R.C.; et al. Null Mutations in the Filaggrin Gene (FLG) Determine Major Susceptibility to Early-Onset Atopic Dermatitis That Persists into Adulthood. J Invest Dermatol 2007, 127, 564–567. [Google Scholar] [CrossRef]
- Palmer, C.N.A.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.D.; et al. Common Loss-of-Function Variants of the Epidermal Barrier Protein Filaggrin Are a Major Predisposing Factor for Atopic Dermatitis. Nat Genet 2006, 38, 441–446. [Google Scholar] [CrossRef]
- Dêbiñska, A.; Danielewicz, H.; Drabik-Chamerska, A.; Kalita, D.; Boznañski, A. Chromosome 11q13.5 Variant as a Risk Factor for Atopic Dermatitis in Children. Postepy Dermatol Alergol 2020, 37, 103–110. [Google Scholar] [CrossRef]
- Can, C.; Yazıcıoğlu, M.; Gürkan, H.; Tozkır, H.; Görgülü, A.; Süt, N.H. Lack of Association Between Toll-like Receptor 2 Polymorphisms (R753Q and A-16934T) and Atopic Dermatitis in Children from Thrace Region of Turkey. Balkan Med J 2017, 34, 232–238. [Google Scholar] [CrossRef]
- Salpietro, C.; Rigoli, L.; Miraglia Del Giudice, M.; Cuppari, C.; Di Bella, C.; Salpietro, A.; Maiello, N.; La Rosa, M.; Marseglia, G.L.; Leonardi, S.; et al. TLR2 and TLR4 Gene Polymorphisms and Atopic Dermatitis in Italian Children: A Multicenter Study. Int J Immunopathol Pharmacol 2011, 24, 33–40. [Google Scholar] [CrossRef]
- Galli, E.; Ciucci, A.; Cersosimo, S.; Pagnini, C.; Avitabile, S.; Mancino, G.; Delle Fave, G.; Corleto, V.D. Eczema and Food Allergy in an Italian Pediatric Cohort: No Association with TLR-2 and TLR-4 Polymorphisms. Int J Immunopathol Pharmacol 2010, 23, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.-Y.; Schumann, R.R.; Hamann, L.; Neumann, K.; Worm, M.; Heine, G. Association of the Toll-like Receptor 2 A-16934T Promoter Polymorphism with Severe Atopic Dermatitis. Allergy 2009, 64, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Niebuhr, M.; Langnickel, J.; Draing, C.; Renz, H.; Kapp, A.; Werfel, T. Dysregulation of Toll-like Receptor-2 (TLR-2)-Induced Effects in Monocytes from Patients with Atopic Dermatitis: Impact of the TLR-2 R753Q Polymorphism. Allergy 2008, 63, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Levchenko, L.Yu.; Izmailova, O.V.; Shlykova, O.A.; Kaidashev, I.P. TLR4 896A/G Gene Polymorphism, Rather than the TLR4 1196C/T and TLR2 2258G/A Gene Polymorphisms, Determines the Severe and Aggravated Course of Atopic Dermatitis in Children. Cytology and Genetics 2013, 47, 167–173. [Google Scholar] [CrossRef]
- Ahmad-Nejad, P.; Mrabet-Dahbi, S.; Breuer, K.; Klotz, M.; Werfel, T.; Herz, U.; Heeg, K.; Neumaier, M.; Renz, H. The Toll-like Receptor 2 R753Q Polymorphism Defines a Subgroup of Patients with Atopic Dermatitis Having Severe Phenotype. J Allergy Clin Immunol 2004, 113, 565–567. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Patria, M.F.; Spena, S.; Codecà, C.; Tagliabue, C.; Zampiero, A.; Lelii, M.; Montinaro, V.; Pelucchi, C.; Principi, N. Impact of Genetic Polymorphisms on Paediatric Atopic Dermatitis. Int J Immunopathol Pharmacol 2015, 28, 286–295. [Google Scholar] [CrossRef]
- Lesiak, A.; Zakrzewski, M.; Przybyłowska, K.; Rogowski-Tylman, M.; Wozniacka, A.; Narbutt, J. Atopic Dermatitis Patients Carrying G Allele in -1082 G/A IL-10 Polymorphism Are Predisposed to Higher Serum Concentration of IL-10. Arch Med Sci 2014, 10, 1239–1243. [Google Scholar] [CrossRef]
- Kayserova, J.; Sismova, K.; Zentsova-Jaresova, I.; Katina, S.; Vernerova, E.; Polouckova, A.; Capkova, S.; Malinova, V.; Striz, I.; Sediva, A. A Prospective Study in Children with a Severe Form of Atopic Dermatitis: Clinical Outcome in Relation to Cytokine Gene Polymorphisms. J Investig Allergol Clin Immunol 2012, 22, 92–101. [Google Scholar]
- Stavric, K.; Peova, S.; Trajkov, D.; Spiroski, M. Gene Polymorphisms of 22 Cytokines in Macedonian Children with Atopic Dermatitis. Iran J Allergy Asthma Immunol 2012, 11, 37–50. [Google Scholar]
- Lesiak, A.; Kuna, P.; Zakrzewski, M.; van Geel, M.; Bladergroen, R.S.; Przybylowska, K.; Stelmach, I.; Majak, P.; Hawro, T.; Sysa-Jedrzejowska, A.; et al. Combined Occurrence of Filaggrin Mutations and IL-10 or IL-13 Polymorphisms Predisposes to Atopic Dermatitis. Exp Dermatol 2011, 20, 491–495. [Google Scholar] [CrossRef]
- Reich, K.; Westphal, G.; König, I.R.; Mössner, R.; Schupp, P.; Gutgesell, C.; Hallier, E.; Ziegler, A.; Neumann, C. Cytokine Gene Polymorphisms in Atopic Dermatitis. Br J Dermatol 2003, 148, 1237–1241. [Google Scholar] [CrossRef]
- Arkwright, P.D.; Chase, J.M.; Babbage, S.; Pravica, V.; David, T.J.; Hutchinson, I.V. Atopic Dermatitis Is Associated with a Low-Producer Transforming Growth Factor Beta(1) Cytokine Genotype. J Allergy Clin Immunol 2001, 108, 281–284. [Google Scholar] [CrossRef]
- Ponińska, J.K.; Samoliński, B.; Tomaszewska, A.; Raciborski, F.; Samel-Kowalik, P.; Walkiewicz, A.; Lipiec, A.; Piekarska, B.; Krzych-Fałta, E.; Namysłowski, A.; et al. Haplotype Dependent Association of Rs7927894 (11q13.5) with Atopic Dermatitis and Chronic Allergic Rhinitis: A Study in ECAP Cohort. PLoS One 2017, 12, e0183922. [Google Scholar] [CrossRef]
- Greisenegger, E.K.; Zimprich, F.; Zimprich, A.; Gleiss, A.; Kopp, T. Association of the Chromosome 11q13.5 Variant with Atopic Dermatitis in Austrian Patients. Eur J Dermatol 2013, 23, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Marenholz, I.; Bauerfeind, A.; Esparza-Gordillo, J.; Kerscher, T.; Granell, R.; Nickel, R.; Lau, S.; Henderson, J.; Lee, Y.-A. The Eczema Risk Variant on Chromosome 11q13 (Rs7927894) in the Population-Based ALSPAC Cohort: A Novel Susceptibility Factor for Asthma and Hay Fever. Hum Mol Genet 2011, 20, 2443–2449. [Google Scholar] [CrossRef] [PubMed]
- Hummelshoj, T.; Bodtger, U.; Datta, P.; Malling, H.J.; Oturai, A.; Poulsen, L.K.; Ryder, L.P.; Sorensen, P.S.; Svejgaard, E.; Svejgaard, A. Association between an Interleukin-13 Promoter Polymorphism and Atopy. Eur J Immunogenet 2003, 30, 355–359. [Google Scholar] [CrossRef]
- Dębińska, A.; Danielewicz, H.; Sozańska, B. Genetic Variants in Epidermal Differentiation Complex Genes as Predictive Biomarkers for Atopic Eczema, Allergic Sensitization, and Eczema-Associated Asthma in a 6-Year Follow-Up Case-Control Study in Children. J Clin Med 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Trzeciak, M.; Gleń, J.; Roszkiewicz, J.; Nedoszytko, B. Association of Single Nucleotide Polymorphism of Interleukin-18 with Atopic Dermatitis. J Eur Acad Dermatol Venereol 2010, 24, 78–79. [Google Scholar] [CrossRef] [PubMed]
- Kılıç, S.; Sılan, F.; Hız, M.M.; Işık, S.; Ögretmen, Z.; Özdemir, Ö. Vitamin D Receptor Gene BSMI, FOKI, APAI, and TAQI Polymorphisms and the Risk of Atopic Dermatitis. J Investig Allergol Clin Immunol 2016, 26, 106–110. [Google Scholar] [CrossRef]
- Heine, G.; Hoefer, N.; Franke, A.; Nöthling, U.; Schumann, R.R.; Hamann, L.; Worm, M. Association of Vitamin D Receptor Gene Polymorphisms with Severe Atopic Dermatitis in Adults. Br J Dermatol 2013, 168, 855–858. [Google Scholar] [CrossRef]
- Dežman, K.; Korošec, P.; Rupnik, H.; Rijavec, M. SPINK5 Is Associated with Early-Onset and CHI3L1 with Late-Onset Atopic Dermatitis. Int J Immunogenet 2017, 44, 212–218. [Google Scholar] [CrossRef]
- Fölster-Holst, R.; Stoll, M.; Koch, W.A.; Hampe, J.; Christophers, E.; Schreiber, S. Lack of Association of SPINK5 Polymorphisms with Nonsyndromic Atopic Dermatitis in the Population of Northern Germany. Br J Dermatol 2005, 152, 1365–1367. [Google Scholar] [CrossRef]
- Vavilin, V.A.; Safronova, O.G.; Lyapunova, A.A.; Lyakhovich, V.V.; Kaznacheeva, L.F.; Manankin, N.A.; Molokova, A.V. Interaction of GSTM1, GSTT1, and GSTP1 Genotypes in Determination of Predisposition to Atopic Dermatitis. Bull Exp Biol Med 2003, 136, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Safronova, O.G.; Vavilin, V.A.; Lyapunova, A.A.; Makarova, S.I.; Lyakhovich, V.V.; Kaznacheeva, L.F.; Manankin, N.A.; Batychko, O.A.; Gavalov, S.M. Relationship between Glutathione S-Transferase P1 Polymorphism and Bronchial Asthma and Atopic Dermatitis. Bull Exp Biol Med 2003, 136, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Wang, H.-Y.; Lee, H.; Lee, S.-Y.; Hong, S.-J.; Choi, E.H. Clinical Characteristics and Genetic Variations in Early-Onset Atopic Dermatitis Patients. Ann Dermatol 2019, 31, 286–293. [Google Scholar] [CrossRef]
- Sasaki, T.; Furusyo, N.; Shiohama, A.; Takeuchi, S.; Nakahara, T.; Uchi, H.; Hirota, T.; Tamari, M.; Shimizu, N.; Ebihara, T.; et al. Filaggrin Loss-of-Function Mutations Are Not a Predisposing Factor for Atopic Dermatitis in an Ishigaki Island under Subtropical Climate. J Dermatol Sci 2014, 76, 10–15. [Google Scholar] [CrossRef]
- Meng, L.; Wang, L.; Tang, H.; Tang, X.; Jiang, X.; Zhao, J.; Gao, J.; Li, B.; Fu, X.; Chen, Y.; et al. Filaggrin Gene Mutation c.3321delA Is Associated with Various Clinical Features of Atopic Dermatitis in the Chinese Han Population. PLoS One 2014, 9, e98235. [Google Scholar] [CrossRef]
- Lee, D.-E.; Park, S.-Y.; Han, J.-Y.; Ryu, H.-M.; Lee, H.-C.; Han, Y.S. Association between Filaggrin Mutations and Atopic Dermatitis in Korean Pregnant Women. Int J Dermatol 2013, 52, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Common, J.E.A.; Haines, R.L.; Balakrishnan, A.; Brown, S.J.; Goh, C.S.M.; Cordell, H.J.; Sandilands, A.; Campbell, L.E.; Kroboth, K.; et al. Wide Spectrum of Filaggrin-Null Mutations in Atopic Dermatitis Highlights Differences between Singaporean Chinese and European Populations. Br J Dermatol 2011, 165, 106–114. [Google Scholar] [CrossRef]
- Zhang, H.; Guo, Y.; Wang, W.; Shi, M.; Chen, X.; Yao, Z. Mutations in the Filaggrin Gene in Han Chinese Patients with Atopic Dermatitis. Allergy 2011, 66, 420–427. [Google Scholar] [CrossRef]
- Osawa, R.; Konno, S.; Akiyama, M.; Nemoto-Hasebe, I.; Nomura, T.; Nomura, Y.; Abe, R.; Sandilands, A.; McLean, W.H.I.; Hizawa, N.; et al. Japanese-Specific Filaggrin Gene Mutations in Japanese Patients Suffering from Atopic Eczema and Asthma. J Invest Dermatol 2010, 130, 2834–2836. [Google Scholar] [CrossRef]
- Nomura, Y.; Akiyama, M.; Nomura, T.; Nemoto-Hasebe, I.; Abe, R.; McLean, W.H.I.; Shimizu, H. Chromosome 11q13.5 Variant: No Association with Atopic Eczema in the Japanese Population. J Dermatol Sci 2010, 59, 210–212. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, L.; Di, Z.-H.; Zhao, L.-P.; Lu, Y.-N.; Xu, J.; Chen, H.-D.; Gao, X.-H. Association Analysis of Filaggrin Gene Mutations and Atopic Dermatitis in Northern China. Br J Dermatol 2010, 162, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Nemoto-Hasebe, I.; Akiyama, M.; Nomura, T.; Sandilands, A.; McLean, W.H.I.; Shimizu, H. FLG Mutation p.Lys4021X in the C-Terminal Imperfect Filaggrin Repeat in Japanese Patients with Atopic Eczema. Br J Dermatol 2009, 161, 1387–1390. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Akiyama, M.; Sandilands, A.; Nemoto-Hasebe, I.; Sakai, K.; Nagasaki, A.; Palmer, C.N.A.; Smith, F.J.D.; McLean, W.H.I.; Shimizu, H. Prevalent and Rare Mutations in the Gene Encoding Filaggrin in Japanese Patients with Ichthyosis Vulgaris and Atopic Dermatitis. J Invest Dermatol 2009, 129, 1302–1305. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Akiyama, M.; Sandilands, A.; Nemoto-Hasebe, I.; Sakai, K.; Nagasaki, A.; Ota, M.; Hata, H.; Evans, A.T.; Palmer, C.N.A.; et al. Specific Filaggrin Mutations Cause Ichthyosis Vulgaris and Are Significantly Associated with Atopic Dermatitis in Japan. J Invest Dermatol 2008, 128, 1436–1441. [Google Scholar] [CrossRef]
- Enomoto, H.; Hirata, K.; Otsuka, K.; Kawai, T.; Takahashi, T.; Hirota, T.; Suzuki, Y.; Tamari, M.; Otsuka, F.; Fujieda, S.; et al. Filaggrin Null Mutations Are Associated with Atopic Dermatitis and Elevated Levels of IgE in the Japanese Population: A Family and Case-Control Study. J Hum Genet 2008, 53, 615. [Google Scholar] [CrossRef]
- Nomura, T.; Sandilands, A.; Akiyama, M.; Liao, H.; Evans, A.T.; Sakai, K.; Ota, M.; Sugiura, H.; Yamamoto, K.; Sato, H.; et al. Unique Mutations in the Filaggrin Gene in Japanese Patients with Ichthyosis Vulgaris and Atopic Dermatitis. J Allergy Clin Immunol 2007, 119, 434–440. [Google Scholar] [CrossRef]
- Ching, G.K.; Hon, K.-L.; Ng, P.-C.; Leung, T.-F. Filaggrin Null Mutations in Childhood Atopic Dermatitis among the Chinese. Int J Immunogenet 2009, 36, 251–254. [Google Scholar] [CrossRef]
- Zhong, W.-L.; Wu, X.; Yu, B.; Zhang, J.; Zhang, W.; Xu, N.; Zhou, J.; Zheng, J.-C.; Chen, X.-F.; Dou, X. Filaggrin Gene Mutation c.3321delA Is Associated with Dry Phenotypes of Atopic Dermatitis in the Chinese Han Population. Chin Med J (Engl) 2016, 129, 1498–1500. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Yang, S.W.; Kim, H.-L.; Kim, S.-H.; Kim, S.J.; Park, S.-M.; Son, M.; Ryu, S.; Pyo, Y.-S.; Lee, J.-S.; et al. Association between P478S Polymorphism of the Filaggrin Gene & Atopic Dermatitis. Indian J Med Res 2013, 138, 922–927. [Google Scholar] [PubMed]
- Wang, I.J.; Lin, T.J.; Kuo, C.F.; Lin, S.L.; Lee, Y.L.; Chen, P.C. Filaggrin Polymorphism P478S, IgE Level, and Atopic Phenotypes. Br J Dermatol 2011, 164, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Hua, L.; Chen, Q.; Liu, Q.-H.; Guo, Y.-F.; Cheng, R.-H.; Zhang, J.; Zhang, J.-H.; Wang, L.-W.; Ji, R.-X. Interaction between Antibiotic Use and MS4A2 Gene Polymorphism on Childhood Eczema: A Prospective Birth Cohort Study. BMC Pediatr 2021, 21, 314. [Google Scholar] [CrossRef] [PubMed]
- Shang, H.; Cao, X.-L.; Wan, Y.-J.; Meng, J.; Guo, L.-H. IL-4 Gene Polymorphism May Contribute to an Increased Risk of Atopic Dermatitis in Children. Dis Markers 2016, 2016, 1021942. [Google Scholar] [CrossRef]
- Hussein, Y.M.; Alzahrani, S.S.; Alharthi, A.A.; Alhazmi, A.S.; Ghonaim, M.M.; Alghamdy, A.A.N.; El Askary, A. Gene Polymorphism of Interleukin-4, Interleukin-4 Receptor and STAT6 in Children with Atopic Dermatitis in Taif, Saudi Arabia. Immunol Invest 2016, 45, 223–234. [Google Scholar] [CrossRef]
- Gharagozlou, M.; Behniafard, N.; Amirzargar, A.A.; Hosseinverdi, S.; Sotoudeh, S.; Farhadi, E.; Khaledi, M.; Aryan, Z.; Moghaddam, Z.G.; Mahmoudi, M.; et al. Association between Single Nucleotide Polymorphisms of the Interleukin-4 Gene and Atopic Dermatitis. Acta Dermatovenerol Croat 2015, 23, 96–100. [Google Scholar]
- Hussein, Y.M.; Shalaby, S.M.; Nassar, A.; Alzahrani, S.S.; Alharbi, A.S.; Nouh, M. Association between Genes Encoding Components of the IL-4/IL-4 Receptor Pathway and Dermatitis in Children. Gene 2014, 545, 276–281. [Google Scholar] [CrossRef]
- Tanaka, K.; Sugiura, H.; Uehara, M.; Hashimoto, Y.; Donnelly, C.; Montgomery, D.S. Lack of Association between Atopic Eczema and the Genetic Variants of Interleukin-4 and the Interleukin-4 Receptor Alpha Chain Gene: Heterogeneity of Genetic Backgrounds on Immunoglobulin E Production in Atopic Eczema Patients. Clin Exp Allergy 2001, 31, 1522–1527. [Google Scholar] [CrossRef]
- Kawashima, T.; Noguchi, E.; Arinami, T.; Yamakawa-Kobayashi, K.; Nakagawa, H.; Otsuka, F.; Hamaguchi, H. Linkage and Association of an Interleukin 4 Gene Polymorphism with Atopic Dermatitis in Japanese Families. J Med Genet 1998, 35, 502–504. [Google Scholar] [CrossRef]
- Morizane, S.; Ouchida, M.; Sunagawa, K.; Sugimoto, S.; Kobashi, M.; Sugihara, S.; Nomura, H.; Tsuji, K.; Sato, A.; Miura, Y.; et al. Analysis of All 34 Exons of the SPINK5 Gene in Japanese Atopic Dermatitis Patients. Acta Med Okayama 2018, 72, 275–282. [Google Scholar] [CrossRef]
- Zhao, L.P.; Di, Z.; Zhang, L.; Wang, L.; Ma, L.; Lv, Y.; Hong, Y.; Wei, H.; Chen, H.D.; Gao, X.H. Association of SPINK5 Gene Polymorphisms with Atopic Dermatitis in Northeast China. J Eur Acad Dermatol Venereol 2012, 26, 572–577. [Google Scholar] [CrossRef]
- Kato, A.; Fukai, K.; Oiso, N.; Hosomi, N.; Murakami, T.; Ishii, M. Association of SPINK5 Gene Polymorphisms with Atopic Dermatitis in the Japanese Population. Br J Dermatol 2003, 148, 665–669. [Google Scholar] [CrossRef]
- Bin Huraib, G.; Al Harthi, F.; Arfin, M.; Al-Sugheyr, M.; Rizvi, S.; Al-Asmari, A. Cytokine Gene Polymorphisms in Saudi Patients With Atopic Dermatitis: A Case-Control Study. Biomark Insights 2018, 13, 1177271918777760. [Google Scholar] [CrossRef] [PubMed]
- Behniafard, N.; Amirzargar, A.A.; Gharagozlou, M.; Delavari, F.; Hosseinverdi, S.; Sotoudeh, S.; Farhadi, E.; Mahmoudi, M.; Khaledi, M.; Moghaddam, Z.G.; et al. Single Nucleotide Polymorphisms of the Genes Encoding IL-10 and TGF-Β1 in Iranian Children with Atopic Dermatitis. Allergol Immunopathol (Madr) 2018, 46, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Sohn, M.H.; Song, J.S.; Kim, K.-W.; Kim, E.-S.; Kim, K.-E.; Lee, J.M. Association of Interleukin-10 Gene Promoter Polymorphism in Children with Atopic Dermatitis. J Pediatr 2007, 150, 106–108. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Tanaka, K.; Arakawa, M. Case-Control Study of Eczema in Relation to IL4Rα Genetic Polymorphisms in Japanese Women: The Kyushu Okinawa Maternal and Child Health Study. Scand J Immunol 2013, 77, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Tanaka, K.; Arakawa, M. IL13 Genetic Polymorphisms, Smoking, and Eczema in Women: A Case-Control Study in Japan. BMC Med Genet 2011, 12, 142. [Google Scholar] [CrossRef]
- Miyake, Y.; Kiyohara, C.; Koyanagi, M.; Fujimoto, T.; Shirasawa, S.; Tanaka, K.; Sasaki, S.; Hirota, Y. Case-Control Study of Eczema Associated with IL13 Genetic Polymorphisms in Japanese Children. Int Arch Allergy Immunol 2011, 154, 328–335. [Google Scholar] [CrossRef]
- Takahashi, N.; Akahoshi, M.; Matsuda, A.; Ebe, K.; Inomata, N.; Obara, K.; Hirota, T.; Nakashima, K.; Shimizu, M.; Tamari, M.; et al. Association of the IL12RB1 Promoter Polymorphisms with Increased Risk of Atopic Dermatitis and Other Allergic Phenotypes. Hum Mol Genet 2005, 14, 3149–3159. [Google Scholar] [CrossRef]
- Namkung, J.-H.; Lee, J.-E.; Kim, E.; Park, G.T.; Yang, H.S.; Jang, H.Y.; Shin, E.-S.; Cho, E.-Y.; Yang, J.-M. An Association between IL-9 and IL-9 Receptor Gene Polymorphisms and Atopic Dermatitis in a Korean Population. J Dermatol Sci 2011, 62, 16–21. [Google Scholar] [CrossRef]
- Kim, J.S.; Choi, J.; Hahn, H.-J.; Lee, Y.-B.; Yu, D.-S.; Kim, J.-W. Association of Macrophage Migration Inhibitory Factor Polymorphisms with Total Plasma IgE Levels in Patients with Atopic Dermatitis in Korea. PLoS One 2016, 11, e0162477. [Google Scholar] [CrossRef]
- Ma, L.; Xue, H.-B.; Guan, X.-H.; Qi, R.-Q.; Liu, Y.-B. Macrophage Migration Inhibitory Factor Promoter 173G/C Polymorphism Is Associated with Atopic Dermatitis Risk. Int J Dermatol 2014, 53, e75–77. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Tanaka, K.; Arakawa, M. IL5RA Polymorphisms, Smoking and Eczema in Japanese Women: The Kyushu Okinawa Maternal and Child Health Study. Int J Immunogenet 2015, 42, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Tsunemi, Y.; Saeki, H.; Shibata, S.; Sekiya, T.; Nakamura, K.; Kakinuma, T.; Kagami, S.; Fujita, H.; Tada, Y.; et al. Interferon-18 Gene Polymorphism -137 G/C Is Associated with Susceptibility to Psoriasis Vulgaris but Not with Atopic Dermatitis in Japanese Patients. J Dermatol Sci 2009, 53, 162–163. [Google Scholar] [CrossRef] [PubMed]
- Osawa, K.; Etoh, T.; Ariyoshi, N.; Ishii, I.; Ohtani, M.; Kariya, S.; Uchino, K.; Kitada, M. Relationship between Kaposi’s Varicelliform Eruption in Japanese Patients with Atopic Dermatitis Treated with Tacrolimus Ointment and Genetic Polymorphisms in the IL-18 Gene Promoter Region. J Dermatol 2007, 34, 531–536. [Google Scholar] [CrossRef]
- Behniafard, N.; Gharagozlou, M.; Farhadi, E.; Khaledi, M.; Sotoudeh, S.; Darabi, B.; Fathi, S.M.; Gholizadeh Moghaddam, Z.; Mahmoudi, M.; Aghamohammadi, A.; et al. TNF-Alpha Single Nucleotide Polymorphisms in Atopic Dermatitis. Eur Cytokine Netw 2012, 23, 163–165. [Google Scholar] [CrossRef]
- Zhou, J.; Zhou, Y.; Lin, L.; Wang, J.; Peng, X.; Li, J.; Li, L. Association of Polymorphisms in the Promoter Region of FCER1A Gene with Atopic Dermatitis, Chronic Uticaria, Asthma, and Serum Immunoglobulin E Levels in a Han Chinese Population. Hum Immunol 2012, 73, 301–305. [Google Scholar] [CrossRef]
- Park, K.Y.; Park, M.K.; Kim, E.J.; Lee, M.-K.; Seo, S.J. FCεRI Gene Promoter Polymorphisms and Total IgE Levels in Susceptibility to Atopic Dermatitis in Korea. J Korean Med Sci 2011, 26, 870–874. [Google Scholar] [CrossRef] [PubMed]
- Tsunemi, Y.; Komine, M.; Sekiya, T.; Saeki, H.; Nakamura, K.; Hirai, K.; Kakinuma, T.; Kagami, S.; Fujita, H.; Asano, N.; et al. The -431C>T Polymorphism of Thymus and Activation-Regulated Chemokine Increases the Promoter Activity but Is Not Associated with Susceptibility to Atopic Dermatitis in Japanese Patients. Exp Dermatol 2004, 13, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, T.; Tsunemi, Y.; Miyamasu, M.; Ohta, K.; Morita, A.; Saeki, H.; Matsushima, K.; Yoshie, O.; Tsuchiya, N.; Yamaguchi, M.; et al. Variations in the Human Th2-Specific Chemokine TARC Gene. Immunogenetics 2003, 54, 742–745. [Google Scholar] [CrossRef] [PubMed]
- Grafanaki, K.; Antonatos, C.; Maniatis, A.; Petropoulou, A.; Vryzaki, E.; Vasilopoulos, Y.; Georgiou, S.; Gregoriou, S. Intrinsic Effects of Exposome in Atopic Dermatitis: Genomics, Epigenomics and Regulatory Layers. Journal of Clinical Medicine 2023, 12. [Google Scholar] [CrossRef]
- McAleer, M.A.; Irvine, A.D. The Multifunctional Role of Filaggrin in Allergic Skin Disease. J Allergy Clin Immunol 2013, 131, 280–291. [Google Scholar] [CrossRef]
- Rodríguez, E.; Baurecht, H.; Herberich, E.; Wagenpfeil, S.; Brown, S.J.; Cordell, H.J.; Irvine, A.D.; Weidinger, S. Meta-Analysis of Filaggrin Polymorphisms in Eczema and Asthma: Robust Risk Factors in Atopic Disease. J Allergy Clin Immunol 2009, 123, 1361–1370.e7. [Google Scholar] [CrossRef] [PubMed]
- Nedoszytko, B.; Olszewska, B.; Roszkiewicz, J.; Glen, J.; Zabłotna, M.; Ługowska-Umer, H.; Nowicki, R.; Sokołowska-Wojdyło, M. The Role of Polymorphism of Interleukin-2, -10, -13 and TNF-α Genes in Cutaneous T-Cell Lymphoma Pathogenesis. Postepy Dermatol Alergol 2016, 33, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Park, A.; Wong, L.; Lang, A.; Kraus, C.; Anderson, N.; Elsensohn, A. Cutaneous T-Cell Lymphoma Following Dupilumab Use: A Systematic Review. Int J Dermatol 2022. [Google Scholar] [CrossRef] [PubMed]
- Imboden, M.; Nieters, A.; Bircher, A.J.; Brutsche, M.; Becker, N.; Wjst, M.; Ackermann-Liebrich, U.; Berger, W.; Probst-Hensch, N.M. Cytokine Gene Polymorphisms and Atopic Disease in Two European Cohorts. (ECRHS-Basel and SAPALDIA). Clin Mol Allergy 2006, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Weissler, K.A.; Frischmeyer-Guerrerio, P.A. Genetic Evidence for the Role of Transforming Growth Factor-β in Atopic Phenotypes. Curr Opin Immunol 2019, 60, 54–62. [Google Scholar] [CrossRef]
- Bozza, M.T.; Lintomen, L.; Kitoko, J.Z.; Paiva, C.N.; Olsen, P.C. The Role of MIF on Eosinophil Biology and Eosinophilic Inflammation. Clin Rev Allergy Immunol 2020, 58, 15–24. [Google Scholar] [CrossRef]
- Egger, M.; Smith, G.D. Bias in Location and Selection of Studies. BMJ 1998, 316, 61–66. [Google Scholar] [CrossRef]
- Lou, C.; Mitra, N.; Wubbenhorst, B.; D’Andrea, K.; Hoffstad, O.; Kim, B.S.; Yan, A.; Zaenglein, A.L.; Fuxench, Z.C.; Nathanson, K.L.; et al. Association between Fine Mapping Thymic Stromal Lymphopoietin and Atopic Dermatitis Onset and Persistence. Ann Allergy Asthma Immunol 2019, 123, 595–601.e1. [Google Scholar] [CrossRef]
Figure 1.
Flow diagram of our included studies.
Figure 2.
Association between single nucleotide polymorphisms and atopic dermatitis susceptibility in patients of European descent. Dotted diamonds represent statistically significant small-study effects.
Figure 2.
Association between single nucleotide polymorphisms and atopic dermatitis susceptibility in patients of European descent. Dotted diamonds represent statistically significant small-study effects.
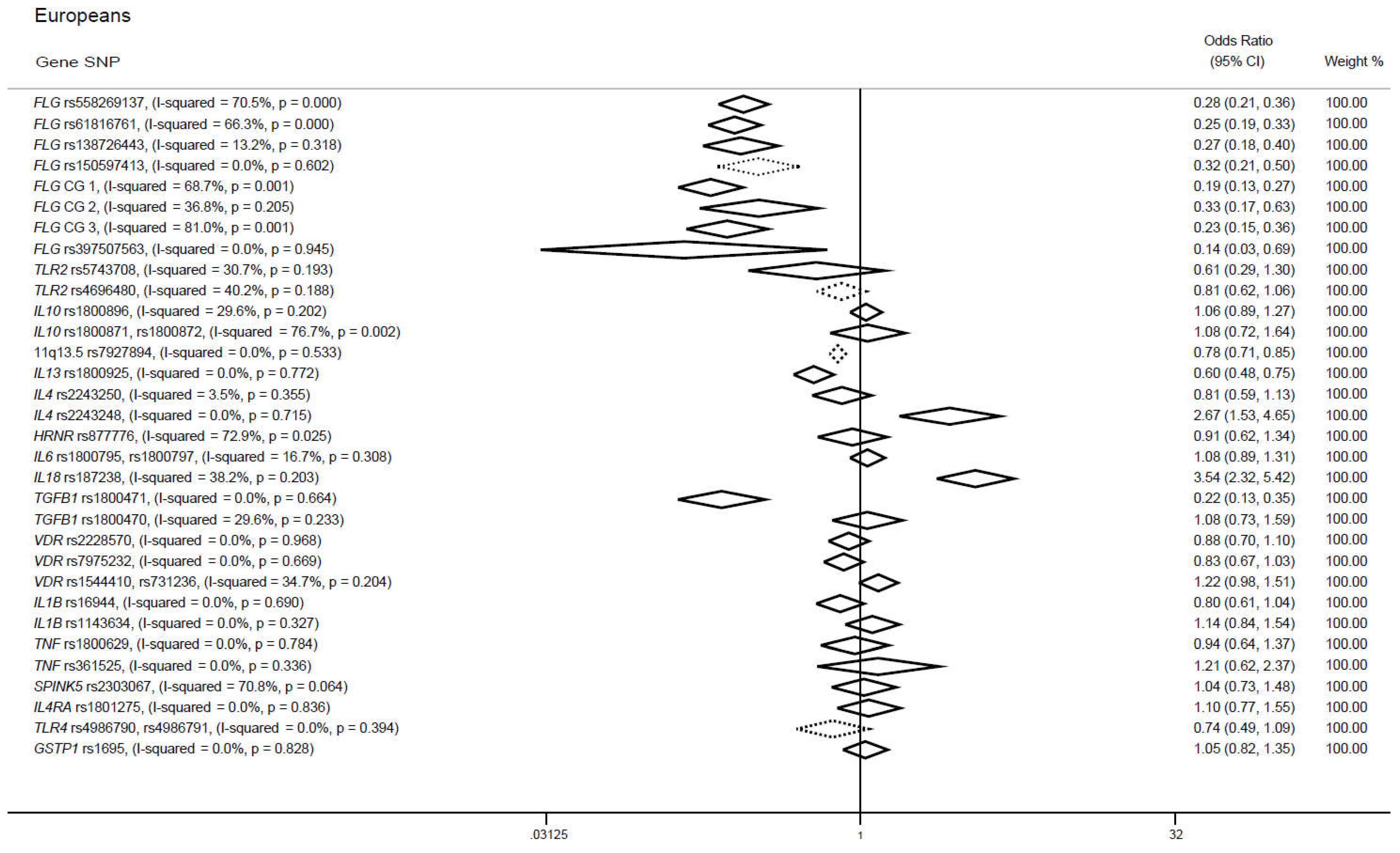
Figure 3.
Association between single nucleotide polymorphisms and atopic dermatitis susceptibility in patients of Asian descent. Dotted diamonds represent statistically significant small-study effects.
Figure 3.
Association between single nucleotide polymorphisms and atopic dermatitis susceptibility in patients of Asian descent. Dotted diamonds represent statistically significant small-study effects.
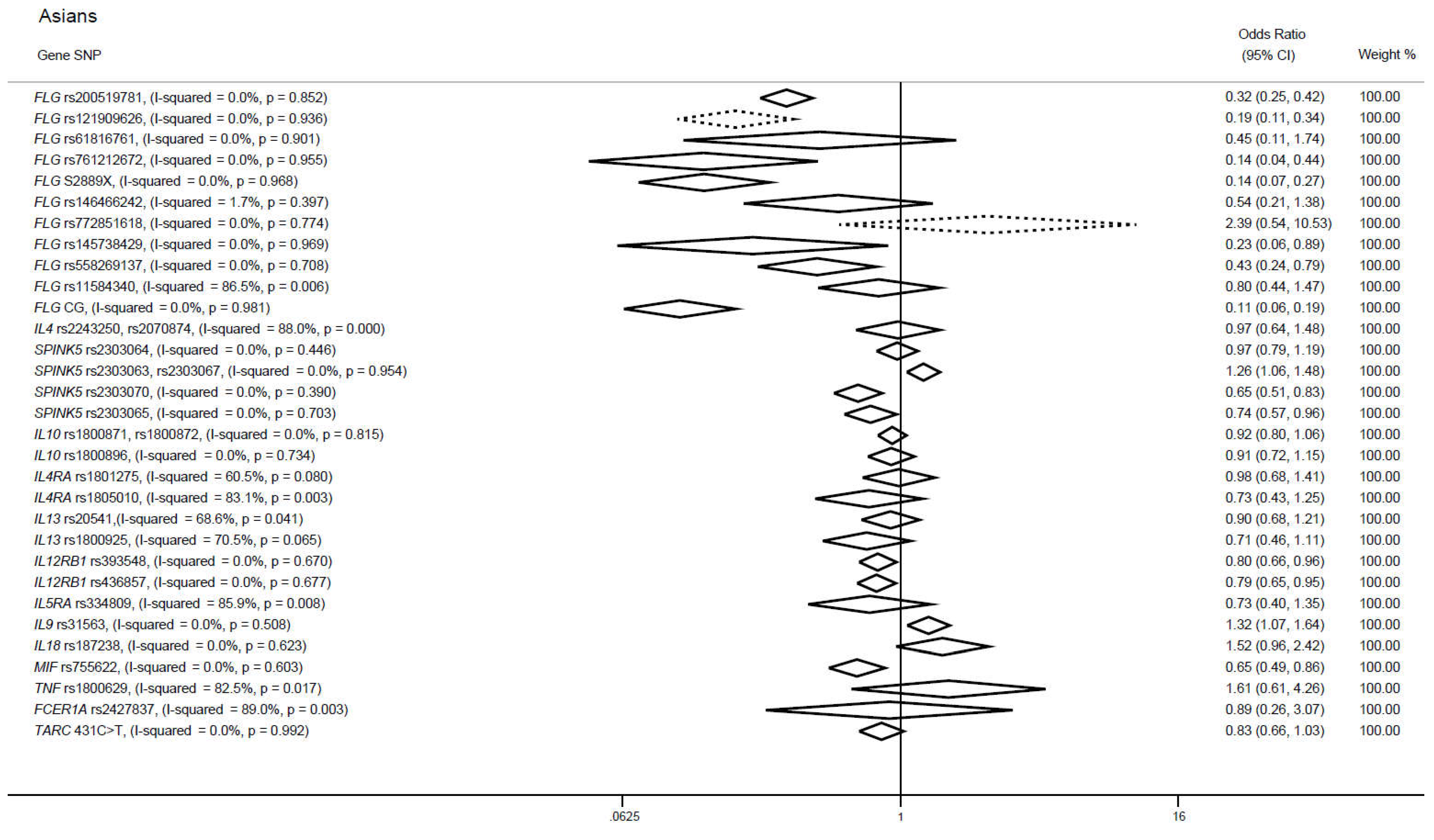

Table 1.
Studies of European descent identified through our systematic search.
| Study, Year [REF] | Rs ID | Sample Size |
|---|---|---|
| Vardar Acar N et al., 2020 [14] | rs558269137, rs61816761, rs138726443, rs150597413 | 189 |
| Jurakic Toncic R et al., 2020 [15] | rs558269137, rs61816761, rs138726443 | 150 |
| González-Tarancón R et al., 2020 [16] | rs558269137, rs61816761, rs138726443, CG2 | 214 |
| Woźniak M et al., 2016 [17] | rs558269137, rs61816761, CG1 | 121 |
| Trzeciak M et al. (a), 2016 [18] | rs558269137, rs61816761, rs1800925, rs187238 | 275 |
| Trzeciak M et al. (b), 2016 [19] | rs558269137 | 256 |
| Ballardini N et al., 2013 [20] | rs558269137, rs61816761, rs138726443, CG2 | 1,854 |
| Ercan H et al., 2013 [21] | rs61816761 | 99 |
| Mlitz V et al., 2012 [22] | rs558269137, rs61816761, rs138726443, CG2 | 196 |
| O'Regan GM et al., 2010 [23] | rs558269137, rs61816761, rs138726443, rs150597413, CG3, rs7927894, rs877776 | 1,511 |
| Greisenegger E et al., 2010 [24] | rs558269137, rs61816761, rs138726443, rs150597413, CG3 | 864 |
| Gao PS et al., 2009 [25] | rs558269137, rs61816761, CG1 | 435 |
| Brown SJ et al. (a), 2008 [26] | rs558269137, rs61816761, rs138726443, rs150597413, rs397507563 | 811 |
| Brown SJ et al. (b), 2008 [27] | rs558269137, rs61816761, rs138726443, rs150597413, rs397507563 | 1,221 |
| Giardina E et al., 2008 [28] | rs558269137, rs61816761 | 388 |
| Weidinger S et al., 2008 [29] | rs558269137, rs61816761, rs138726443, rs150597413, CG3 | 3,099 |
| Rogers AJ et al., 2007 [30] | rs558269137, rs61816761, CG1 | 646 |
| Lerbaek A et al., 2007 [31] | rs558269137, rs61816761, CG1 | 215 |
| Sandilands A et al., 2007 [32] | rs558269137, rs61816761, rs138726443, rs150597413, rs397507563 | 924 |
| Weidinger S et al., 2007 [33] | rs558269137, rs61816761, CG1 | 526 |
| Marenholz I et al., 2006 [34] | rs558269137, rs61816761, CG1 | 507 |
| Stemmler S et al., 2007 [35] | rs558269137, rs61816761, CG1 | 1,078 |
| Barker JN et al., 2007 [36] | rs558269137, rs61816761, CG1 | 1,626 |
| Palmer CN et al., 2006 [37] | rs558269137, rs61816761, CG1 | 241 |
| Dêbiñska A et al., 2020 [38] | CG3, rs7927894 | 188 |
| Can C et al., 2017 [39] | rs5743708, rs4696480 | 139 |
| Salpietro C et al., 2011 [40] | rs5743708, rs4696480, rs4986790, rs4986791 | 337 |
| Galli E et al., 2010 [41] | rs5743708 | 249 |
| Oh DY et al., 2009 [42] | rs5743708, rs4696480, rs4986790, rs4986791 | 265 |
| Niebuhr M et al., 2008 [43] | rs5743708 | 19 |
| Levchenko L Yu et al., 2013 [44] | rs5743708 | 131 |
| Ahmad-Nejad et al., 2003 [45] | rs5743708 | 117 |
| Esposito S et al., 2015 [46] | rs1800896, rs1800872 | 223 |
| Lesiak A et al., 2014 [47] | rs1800896, rs1800925, rs2243250 | 136 |
| Kayserova J et al., 2012 [48] | rs1800896, rs1800871, rs1800872, rs2243250, rs2243248, rs1800795, rs1800797, rs1801275 | 197 |
| Stavric K et al., 2012 [49] | rs1800896, rs1800871, rs1800872, rs2243250, rs2243248, rs1800471, rs1800470, rs1800795, rs1800797, rs16944, rs1143634, rs1800629, rs361525, rs1801275 | 367 |
| Lesiak A et al., 2011 [50] | rs1800896, rs1800925 | 367 |
| Reich K et al., 2003 [51] | rs1800896, rs1800795, rs16944, rs1143634, rs1800629, rs361525 | 308 |
| Arkwright PD et al., 2001 [52] | rs1800896, rs1800471, rs1800470 | 118 |
| Ponińska JK et al., 2017 [53] | rs7927894 | 810 |
| Greisenegger EK et al., 2013 [54] | rs7927894, rs877776 | 518 |
| Marenholz I et al., 2011 [55] | rs7927894 | 2,485 |
| Hummelshoj T et al., 2003 [56] | rs1800925 | 159 |
| Dêbiñska A et al., 2022 [57] | rs877776 | 188 |
| Trzeciak M et al., 2010 [58] | rs187238 | 113 |
| Kılıç S et al., 2016 [59] | rs2228570, rs7975232, rs1544410, rs731236 | 138 |
| Heine G et al., 2013 [60] | rs2228570, rs7975232, rs1544410, rs731236 | 530 |
| Dežman K et al., 2017 [61] | rs2303067 | 405 |
| Fölster-Holst R et al., 2005 [62] | rs2303067 | 569 |
| Vavilin VA et al., 2003 [63] | rs1695 | 325 |
| Safronova OG et al., 2003 [64] | rs1695 | 274 |
| Abbreviations: CG, Combined Genotype. | ||
Table 2.
Studies of Asian descent identified through our systematic search.
| Study, Year [REF] | Rs ID | Sample Size |
|---|---|---|
| Kim BJ et al., 2019 [65] | rs200519781, rs146466242, rs2303064, rs2303070, rs2303065, rs393548, rs436857, rs31563, rs334809 | 325 |
| Sasaki T et al., 2014 [66] | rs200519781, rs121909626, rs761212672, rs145738429, rs61816761, rs146466242, rs772851618 | 721 |
| Meng L et al., 2014 [67] | rs200519781 | 1,988 |
| Lee DE et al., 2013 [68] | rs200519781 | 175 |
| Chen H et al., 2011 [69] | rs200519781, rs61816761 | 865 |
| Zhang H et al., 2011 [70] | rs200519781 | 353 |
| Osawa R et al., 2010 [71] | rs200519781, rs121909626, rs761212672, rs145738429, S2889X, rs61816761, rs146466242, rs772851618, CG | 306 |
| Nomura Y et al., 2010 [72] | rs200519781, rs121909626, rs761212672, rs145738429, S2889X, rs61816761, rs146466242, rs772851618, CG | 307 |
| Ma L et al., 2010 [73] | rs200519781 | 329 |
| Nemoto-Hasebe I et al., 2009 [74] | rs200519781, rs121909626, rs761212672, rs145738429, S2889X, rs61816761, rs146466242, rs772851618, CG | 271 |
| Nomura T et al., 2009 [75] | rs200519781, rs121909626, rs761212672, rs145738429, S2889X, rs61816761, rs772851618 | 252 |
| Nomura T et al., 2008 [76] | rs200519781, rs121909626, rs761212672, S2889X | 235 |
| Enomoto H et al., 2008 [77] | rs200519781, rs121909626 | 1,299 |
| Nomura T et al., 2007 [78] | rs200519781, rs121909626 | 299 |
| Ching GK et al., 2009 [79] | rs121909626, S2889X, rs558269137, rs61816761 | 365 |
| Zhong WL et al., 2016 [80] | rs558269137 | 1,017 |
| Kim SY et al., 2013 [81] | rs11584340 | 527 |
| Wang IJ et al., 2011 [82] | rs11584340 | 328 |
| Hua L et al., 2021 [83] | rs2243250, rs2070874, rs1801275, rs1805010, rs20541 | 597 |
| Shang H et al., 2016 [84] | rs2243250, rs2070874 | 182 |
| Hussein YM et al., 2016 [85] | rs2243250, rs2070874, rs1805010 | 100 |
| Gharagozlou M et al., 2015 [86] | rs2243250, rs2070874, rs1801275 | 228 |
| Hussein YM et al., 2014 [87] | rs2243250, rs2070874 | 206 |
| Tanaka K et al., 2001 [88] | rs2243250, rs2070874, rs1805010 | 424 |
| Kawashima T et al., 1998 [89] | rs2243250, rs2070874 | 425 |
| Morizane S et al., 2018 [90] | rs2303064, rs2303063, rs2303067, rs2303070 | 107 |
| Zhao LP et al., 2012 [91] | rs2303064, rs2303063, rs2303067, rs2303070 | 341 |
| Kato A et al., 2003 [92] | rs2303064, rs2303063, rs2303067, rs2303065 | 234 |
| Bin Huraib G et al., 2018 [93] | rs1800871, rs1800872, rs1800896, rs1800629 | 315 |
| Behniafard N et al., 2018 [94] | rs1800871, rs1800872, rs1800896 | 229 |
| Sohn MH et al., 2007 [95] | rs1800871, rs1800872, rs1800896 | 416 |
| Miyake Y et al., 2013 [96] | rs1801275 | 823 |
| Miyake Y et al. (a), 2011 [97] | rs20541, rs1800925 | 1,270 |
| Miyake Y et al. (b), 2011 [98] | rs20541, rs1800925 | 533 |
| Takahashi N et al., 2005 [99] | rs393548, rs436857 | 1,040 |
| Namkung JH et al., 2011 [100] | rs31563 | 1,090 |
| Kim JS et al., 2016 [101] | rs755622 | 258 |
| Ma L et al., 2013 [102] | rs755622 | 391 |
| Miyake Y et al., 2015 [103] | rs334809 | 1,318 |
| Kato T et al., 2009 [104] | rs187238 | 264 |
| Osawa K et al., 2007 [105] | rs187238 | 121 |
| Behniafard N et al., 2012 [106] | rs1800629 | 226 |
| Zhou J et al, 2012 [107] | rs2427837 | 380 |
| Park KY et al., 2011 [108] | rs2427837 | 231 |
| Tsunemi Y et al., 2004 [109] | 431C>T | 351 |
| Sekiya T et al., 2003 [110] | 431C>T | 306 |
| Abbreviations: CG, Combined Genotype. | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



