Submitted:
15 September 2025
Posted:
17 September 2025
You are already at the latest version
Abstract
The effect of temperature (T) on filtration rate (F) in filter-feeding bivalves is controversial. Some studies show a clear increase in F with increase in T, in agreement with decrease of kinematic viscosity with increasing temperature and simultaneous increase of beat frequency of the water-pumping lateral cilia. But other studies show little or no effect. Here, we present our understanding of the effect of T on F in the blue mussel, Mytilus edulis, primarily based on own previously published experimental studies. We point out the possible pitfalls for little or no effect of T on F, most likely caused by erroneous use of the method for measuring F and failure of separating the factors that may affect F, e.g., the temperature-tolerance interval, and the algal-concentration interval within which the mussel is optimally filtering at a specified T.
Keywords:
pumping rate
; clearance rate
; temperature-tolerance
; viscosity
; valve-opening degree
1. Introduction
There are many publications dealing with the effect of temperature on filtration rate in the blue mussel, Mytilus edulis, and other ciliary filter-feeding bivalves. The older literature has been reviewed by Kittner & Riisgård (2005) who found that the subject was controversial due to often inconsistent results, likely caused by different experimental conditions and methodological shortcomings.
When dealing with the filtration rate of mussels, it is essential to differentiate between the effect of temperature on the filtration rate measured on optimally filtering mussels and the effect of both high and low temperature on the valve-opening state, which may significantly affect the filtration (or pumping) rate (Jørgensen et al. 1990, Riisgård et al. 2003, Kittner & Riisgård 2005). Therefore, it is important to identify the temperature-tolerance interval for optimally filtering. When mussels are exposed to a change in temperature outside the tolerance interval they may after some time adjust and become tolerant to the new temperature, which is called acclimatization. One of the most important environmental parameters affecting the filtration rate of mussels is the phytoplankton (algal) concentration. Under optimal conditions, M. edulis exploit its filtration capacity, but under sub-optimal conditions, including low algal concentrations, the filtration rate is being reduced by reduction or closure of the valve-gape (Riisgård et al. 2011). The valve closing/opening response has been studied in-depth. Thus, the critical phytoplankton biomass below which the mussel closes its valves is about 0.9 chl a l-1 (Riisgård et al. 2003, 2006). During the winter starvation period in temperate Danish waters M. edulis reduces its valve gape and thereby the filtration rate (Lassen et al. 2006) which reduces the oxygen uptake (respiration) (Tang & Riisgård 2016) and thus the body-weight loss (Riisgård & Larsen 2015). The algal (phytoplankton) biomass in the sea is typically between 1 and 5 µg chl a l-1 (Riisgård et al. 2014a), but because mussels are often living in dense beds their grazing impact on the phytoplankton may become pronounced, and likewise, starvation may occur during winter.
Filtration rate (= pumping rate) is sometimes referred to as clearance rate because it may be measured by the so-called clearance method (Riisgård 2001). The “clearance method” is a reliable and precise method for measuring the filtration rate at known temperatures when correctly used (Figure 1). Here, the clearance rate (Cl) of a group of mussels (n) is estimated from the exponential decline in algal concentration in a well-mixed, strongly aerated aquarium as Cl = aV/n where a = slope of regression line in a semi-ln plot of algal concentration versus time, and V = volume of seawater. A prerequisite for using the clearance method is immediate mixing of once filtered water in all the water, which is ensured by strong air-mixing. However, other methods as reviewed by Riisgård (2001) may be used to measure the filtration rate in mussels, especially the “flow-through chamber method” has frequently been used. In the present account, we present our understanding of the effect of temperature on filtration rate in the blue mussel, Mytilus edulis, primarily based on our own previously published experimental studies of filtration rate and beat frequency of water-pumping lateral cilia. We use the following definitions: clearance rate (Cl) = volume of water cleared of suspended particles per unit of time. If the algal cells are retained with 100% efficiency, then Cl = F (= pumping rate).
2. Our present Understanding
In mussels, the water-pumping lateral cilia in the gills create the feeding current (Riisgård et al. 2015), and because the viscosity of seawater is inversely related to temperature, the increase of filtration rate with temperature in mussels cannot only be accounted for by increased biological activity. The early study of Jørgensen et al. (1990) tried to explain in detail whether alteration in filtration rate with temperature in mussels is under chiefly biological/physiological or physical/mechanical control. They showed that the filtration rate of Mytilus edulis increased with temperature (Figure 2) and decreased linearly with the temperature-determined increase in viscosity of the seawater (Figure 3). The effects of changes in temperature on filtration rate in mussels acclimatized to the seasonally varying temperatures at the collecting site showed that the temperature ranges used in the experiments were those tolerated by the mussels indicating that they were optimally filtering. From Figure 2 it may be seen that in the cold-adapted mussels in February the upper temperature tolerated was 16 °C, compared with 22 °C in June. In the lower range, the cold-adapted mussels tolerated temperatures approaching zero, whereas the 12 °C adapted mussels began to reduce or close the valve gape at 4 to 5 °C. Kittner & Riisgård (2005) studied the effect of temperature on filtration rate in M. edulis and likewise found a linear relationship in the temperature-tolerance interval and acclimatation ranges (Figure 4). For example, in the lower part of the temperature interval, the cold-adapted (11°C) mussels tolerated temperatures down to at least 4.1°C without closing their valves hence maintaining the maximum filtration rate. From Figure 4B it appears that the filtration rate (F, ml min-1) versus temperature (T, °C) for a 50.5 mm shell length M. edulis was found to be F = 3.27T + 38.1 (r2 = 0.953), which implies F = 54.5 and 103.5 ml min-1 at 5 and 20 °C, respectively, thus indicating an 1.9 times increase of the filtration rate.
Later, Riisgård & Larsen (2007) found that the effect of temperature on the beat frequency of the water-pumping lateral cilia in Mytilus edulis gill preparations is controlled by the viscosity of seawater, see Figure 5. Recognizing that filtration rate of mussels and viscosity depend on temperature may have motivated the development of pumping models as done by Jørgensen et al. (1990), Riisgård & Larsen (2007), and Zhang et al. (2024). Common to these studies, the pump head is equated to the sum of frictional losses in the flow system through a mussel, accounting also for kinetic energy contributions at inhalant and exhalant openings. Since viscosity decreases with increasing temperature, friction decreases, and pumping rate increases as it also does due to increasing cilia beat frequency. This is true within the temperature-tolerance interval, which however seems to have been exceeded in the study by Zhang et al. (2024, Figure 4 therein) who found a decrease in filtration rate at 26 and 28 °C, and at 2 °C, very likely due to reduction of the valve gape.
Water pumping cilia are not only important in bivalves, but beating cilia are also important in other aquatic organisms. Thus, viscosity controls or strongly affects bio-mechanical activity such as the swimming of ciliates and micro- and meso-scale aquatic organisms using cilia (Riisgård & Larsen 2009, Larsen & Riisgård 2009). In Mytilus edulis, Jørgensen et al. (1990) and Kittner & Riisgård (2005) found a linear relationship between filtration rate and temperature in the temperature-tolerance interval (Figs. 2 & 4) as also found in the soft clam Mya arenaria by Riisgård & Seerup (2003), and in Arctic clams Hiatella arctica and Mya sp. by Petersen et al. (2003).
2.1. Recommendations, Critical Remarks and Pitfalls
As it appears from the preceding sections, the valve-opening degree of a mussel ultimately controls the filtration rate. To eliminate this variable from the effect of temperature on the water-pumping lateral cilia it is necessary to ensure that the mussel is wide open and exploiting its filtration capacity when the temperature is being varied. When a mussel experiences very low algal concentrations, this eventually leads to reduced valve-gape, or complete closure, along with cessation of the filtration activity, which lasts until the algal concentration is elevated above the lower threshold level.
The opening-closing phenomenon and response times in Mytilus edulis in the presence and absence of algal cells were studied by Riisgård et al. (2003) in controlled laboratory experiments. When algal cells were added to unfed mussels they soon after opened their valves along with a strong increase of the filtration rate. Later on, when the algal concentration decreased under a certain level, the mussels reduced their valve-opening degree and eventually stopped filtering. However, new algal additions rapidly restored maximum filtration rate (Riisgård et al. 2003, Figure 9 therein).
Another study by Riisgård et al. 2006) of valve-gape responses of Mytilus edulis to presence or absence of algal cells in the ambient water, revealed that the critical algal concentration (Cmin) below which the mussel closes its valves is about 700 Rhodomonas salina cells ml-1, equivalent to 0.9 µg chl a l-1. But high algal concentrations may also result in reduced filtration rate due to saturation of the digestive system, Riisgård et al. (2013). Therefore, controlled studies on the effect of temperature on filtration rate should be made on optimally filtering mussels exposed to ambient algal concentrations within this interval.
When using the clearance method, this implies that algal cells should be added repeatedly to ensure that the mussels after some time open their valves and filter at maximum rate, which should be ensured by observation of parallel linear regression lines in a semi-ln plot after every new algal addition (Figure 1) (Marroni et al. 2021, Figure 1 therein). Only one initial algal cell addition and measurement of only the start and end concentration after a certain (arbitrary) time without noting the valve-opening degree will inevitably result in uncertain and uninterpretable data. This seems to be the case in the study by Pestana et al. (2009) who found that the filtration rate was “largely unaffected by temperature” for the golden mussel Limnoperna fortune while Zhang et al. (2024) found a skewed unimodal pattern of increase below and decrease above about 15 oC for this species. Further, Gopalakrishnan & Kashian (2020, Figure 1 therein) found that the filtration rate of the quagga mussel Dreissena rostriformis burgensis increased linearly with temperature from 2 to 25 oC followed by a “decline” at 30 oC, which was outside the mussel’s temperature-tolerance interval.
Cranford et al. (2011) claimed that temperature has “not been identified as an important control on feeding behavior” under natural conditions for mussels, and the authors referred to a figure (Cranford et al. 2011, Figure 4.6 therein, based on unpublished data) showing the clearance rates of Mytilus edulis measured in the field over a wide range of water temperatures using the flow-through chamber method which “revealed little dependence of feeding on temperature”. However, the same shortcomings as mentioned below when using this method are likely. Cranford et al. (2011) criticized the study by e.g. Kittner & Riisgård (2005) for being limited to “an artificial condition that does not exist in nature” where the mussels are not “stimulated by a controlled artificial diet”. This criticism is unwarranted. To separate the various factors that may affect the filtration rate, it is necessary to identify 1) the temperature-tolerance interval within which the valves remain fully open, and further, 2) to identify the algal-concentration interval within which the valves are fully open, The separate effect of temperature and viscosity on the water-pumping lateral cilia can only be precisely measured in fully open and optimally filtering mussels. Finally, the use of “artificial diet” (cultivated algal cells) is also justified. Thus, Lüskow & Riisgård (2018) found in situ filtration rate measurements of M. edulis using ambient natural phytoplankton were comparable to filtration rates obtained in laboratory studies using several methods and controlled diets of cultivated algal cells.
In a recent paper by Rosa et al. (2024) the authors conclude that their experimental data indicate that changes in water temperature and the associated changes in water viscosity have no effect on the filtration rate of mussels. This statement is controversial, and we disagree. To perform precise and reliable laboratory filtration rate measurements on mussels with the ‘flow-through chamber method’ used by Rosa et al. (2024) the design of the chamber must ensure that only inflow water reaches the bivalve’s inhalant aperture and that exit flow is fully mixed. These prerequisites can be checked by a plot of clearance rate versus increasing through-flow to reach a plateau, which is the true clearance rate (Riisgård 2001, Larsen & Riisgård 2012). This requirement was not fulfilled by Rosa et al. (2024) who measured the clearance rate of 55.9 mm shell length Mytilus edulis in chambers of undescribed size and shape (possibly allowing bypass of some water and/or recirculation of once filtered water). They used a constant through-flow of 150 ml min-1, which resulted in strongly varying clearance rates ranging between 28 to 56 ml min-1 and with “no effect of temperature” on the clearance rate. The maximum filtration rate (F, l h-1) in M. edulis may be estimated from the shell length (L, mm) as (Riisgård et al., 2014b): F = 0.00135L2.088, which for a 55.9 mm mussel gives 6 l h-1 = 100 ml min-1, or about 2 to 4 times higher than reported by Rosa et al. (2024). More precise measurements using a chamber with proper shape and size and checked for optimal flow-through rate to ensure a clearance plateau would probably have resulted in a close correlation between temperature and filtration rate as found in M. edulis by Jørgensen et al. (1990) and Kittner & Riisgård (2005).
Author Contributions
H.U.R. and P.S.L. equally contributed with input and text writing. Both authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
[to come]
Conflicts of Interest
The authors declare no conflict of interest
References
- Cranford, P. J., Ward, J. E., Shumway, S. E. (2011). Bivalve filter feeding: Variability and limits of the aquaculture biofilter. In S. E. Shumway (Ed.), Shellfish aquaculture and the environment (pp. 81–124). John Wiley & Sons Publ. [CrossRef]
- Gopalakrishnan, K. K., Kashian, D. R. (2020). Identification of optimal calcium and temperature conditions for quagga mussel filtration rates as a potential predictor of invasion. Environmental Toxicology and Chemistry 39, 410–418.
- Jørgensen, C. B., Larsen, P. S., Riisgård, H. U. (1990). Effects of temperature on the mussel pump. Marine Ecology Progress Series, 64, 89–97.
- Kittner, C., Riisgård, H. U. (2005). Effect of temperature on filtration rate in the mussel Mytilus edulis: No evidence for temperature compensation. Marine Ecology Progress Series, 305, 147-152.
- Larsen, P. S., Riisgård, H. U. (2009). Viscosity and not biological mechanisms often controls the effects of temperature on ciliary activity and swimming velocity of small aquatic organisms. Journal of Experimental Marine Biology and Ecology, 381, 67-73.
- Larsen, P. S, Riisgård, H. U. (2012). Validation of the flow-through chamber (FTC) and steady-state (SS) methods for clearance rate measurements in bivalves. Biology Open. 2012 Jan 15;1(1):6-11. [CrossRef]
- Lassen, J., Kortegård, M., Riisgård, H. U., Friedrichs, M., Graff, G., Larsen, P. S. (2006). Down-mixing of phytoplankton above filter-feeding mussels - interplay between water flow and biomixing. Marine Ecology Progress Series, 314, 77-88.
- Lüskow, F., Riisgård H. U. (2018). In situ filtration rates of blue mussels (Mytilus edulis) measured by an open-top chamber method. Open Journal of Marine Science, 8, 395-406.
- Marroni, S., Mezzeo, N., Iglesias, C. (2021). Effects of temperature and food availability on the filtration and excretion rates of Diplodon parallelopipedon (Hyriidae). International Review of Hydrobiology 106, 249-258. [CrossRef]
- Pestana, D., Ostrensky, A., Boeger, W. A. P., Pie, M. R. (2009). The effect of temperature and body size on filtration rates of Limnoperna fortunei (Bivalvia, Mytilidae) under laboratory conditions. Brazilian Archives of Biology and Technology, 52, 135-144.
- Petersen, J. K., Sejr, M, K., Larsen, J, E. N, (2003). Clearance rates in the Arctic bivalves Hiatella arctica and Mya sp. Polar Biology 26:334–341.
- Riisgård, H. U. (2001). On measurement of filtration rate in bivalves - the stony road to reliable data, review and interpretation. Marine Ecology Progress Series, 211, 275-291.
- Riisgård, H. U., Larsen, P. S. (2007). Viscosity of seawater controls beat frequency of water-pumping cilia and filtration rate of mussels Mytilus edulis. Marine Ecology Progress Series, 343, 141–150.
- Riisgård, H. U., Larsen, P. S. (2009). Ciliary-propelling mechanism, effect of temperature and viscosity on swimming speed, and adaptive significance of the 'jumping' ciliate Mesodinium rubrum. Marine Biology Research, 5, 585-595.
- Riisgård, H. U., Larsen, P. S. (2015). Physiologically regulated valve-closure makes mussels long-term starvation survivors: test of hypothesis. Journal of Molluscan Studies, 81, 303-307.
- Riisgård, H. U., Larsen, P. S. (2018). Thermal and viscous effects on ciliary suspension-feeding bivalves - no need for a new explanation: Comment on Specht & Fuchs (2018). Marine Ecology Progress Series, 596, 263-265.
- Riisgård, H. U., Seerup, D. F. (2003). Filtration rates in the soft clam Mya arenaria: Effects of temperature and body size. Sarsia, 88, 416-428.
- Riisgård, H. U., Kittner, C., Seerup, D.F. (2003). Regulation of opening state and filtration rate in filter-feeding bivalves (Cardium edule, Mytilus edulis, Mya arenaria) in response to low algal concentration. Journal of Experimental Marine Biology and Ecology, 284, 105-127.
- Riisgård, H. U., Lassen, J., Kittner, C. (2006). Valve-gape response times in mussels (Mytilus edulis) - effects of laboratory preceding-feeding conditions and in situ tidally induced variation in phytoplankton biomass. Journal of Shellfish Research, 25, 901-913.
- Riisgård, H. U., Egede, P. P., Saavedra, I. B. (2011). Feeding behaviour of mussels, Mytilus edulis, with a mini-review of current knowledge. Journal of Marine Biology, Volume 2011, Article ID 312459, 13 pages. [CrossRef]
- Riisgård, H. U., Pleissner, D., Lundgreen, K., Larsen P. S. (2013). Growth of mussels Mytilus edulis at algal (Rhodomonas salina) concentrations below and above saturation levels for reduced filtration rate. Mar. Biol. Res. 9(10): 1005-1017.
- Riisgård, H. U., Lundgreen, K., Larsen, P. S. (2014a). Potential for production of 'mini-mussels' in Great Belt (Denmark) evaluated on basis of actual growth of young mussels Mytilus edulis. Aquaculture International, 22, 859-885.
- Riisgård, H. U., Larsen, P. S., Pleissner, D. (2014b). Allometric equations for maximum filtration rate in blue mussels Mytilus edulis and importance of condition index. Helgoland Marine Research, 68, 193-198.
- Riisgård, H. U., Funch, P., Larsen, P. S. (2015). The mussel filter–pump - Present understanding, with a re-examination of gill preparations. Acta Zoologica, 96(3), 273–282.
- Rosa, M., Capriotti, M., Austin, K., Shumway, S. E., Ward, J. E. (2024). Effect of seasonal changes in temperature on capture efficiency in the blue mussel, Mytilus edulis, fed seston and microplastics. Invertebrate Biology, e12446. [CrossRef]
- Tang, B., Riisgård, H. U. (2016). Physiological regulation of valve-opening degree enables mussels Mytilus edulis to overcome starvation periods by reducing the oxygen uptake. Open Journal of Marine Science, 6, 341-352.
- Zhang, J. H., Xu, M. Z., Yang, Y. (2024). Assessing the effects of environmental factors on filtration rates of golden mussel (Limnoperna fortunei). Ecological Indicators, 158, 111544. [CrossRef]
Figure 1.
Measurement of the filtration rate in Mytilus edulis using the clearance method. Exponential decrease in algal cell concentration as a function of time due to fully open and actively filtering mussels (length 50.54 ± 5.5 mm, n = 5) during a clearance experiment at 20.3 °C in an aquarium with well-mixed seawater. Regression lines and estimated filtration rates (l h-1 ind.-1) are shown. Arrows indicate addition of algal cells. From Kittner & Riisgård (2005).
Figure 1.
Measurement of the filtration rate in Mytilus edulis using the clearance method. Exponential decrease in algal cell concentration as a function of time due to fully open and actively filtering mussels (length 50.54 ± 5.5 mm, n = 5) during a clearance experiment at 20.3 °C in an aquarium with well-mixed seawater. Regression lines and estimated filtration rates (l h-1 ind.-1) are shown. Arrows indicate addition of algal cells. From Kittner & Riisgård (2005).
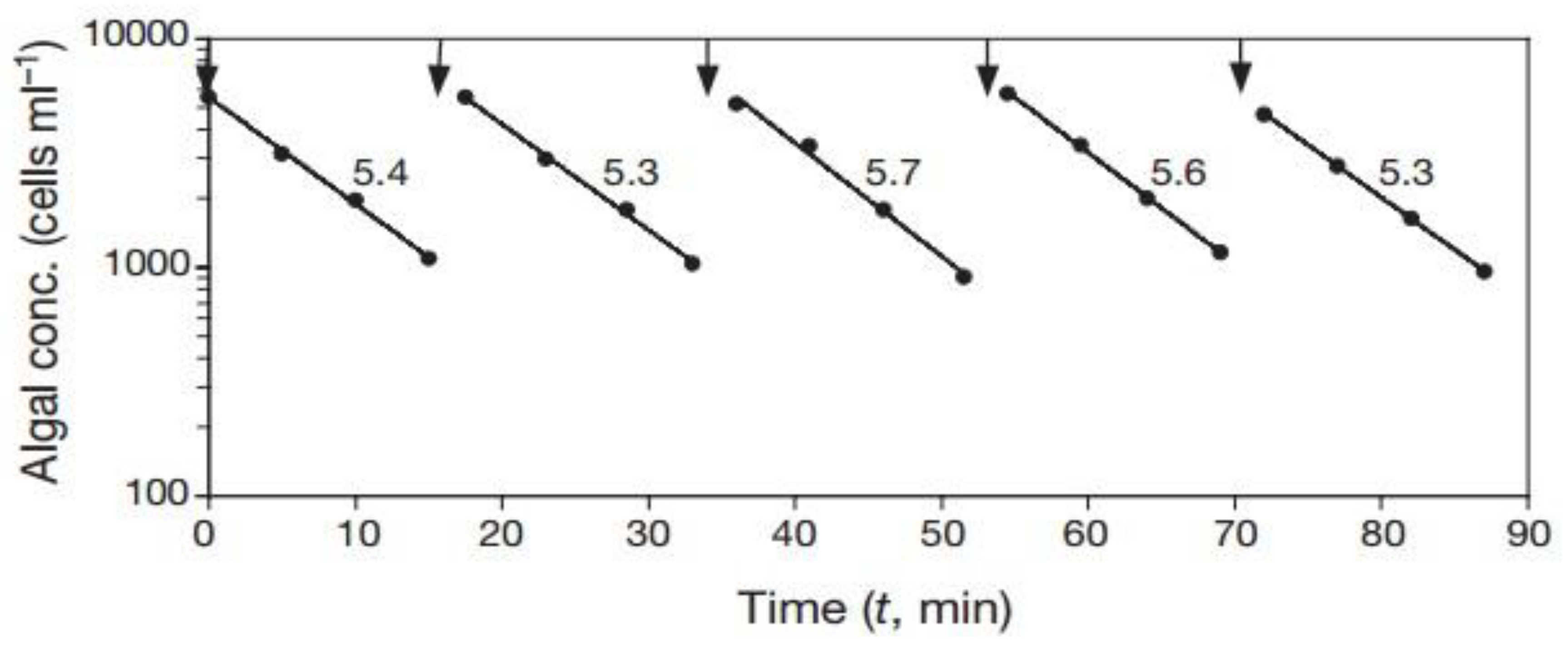
Figure 2.
Mytilus edulis. Relationship between temperature and pumping rate (= filtration rate) in groups of acclimated mussels of different size at different times of the year. (○) February, 25 mussels, 31 ± 0.7 (SD) mm in length, acclimated to 6 °C; (x) May, 20 mussels, 39 ± 2.0 mm, acclimatized to 7.5 °C; (●) June, 30 mussels, 29 ± 2.6 mm, acclimatized to 12 °C. From Jørgensen et al. (1990).
Figure 2.
Mytilus edulis. Relationship between temperature and pumping rate (= filtration rate) in groups of acclimated mussels of different size at different times of the year. (○) February, 25 mussels, 31 ± 0.7 (SD) mm in length, acclimated to 6 °C; (x) May, 20 mussels, 39 ± 2.0 mm, acclimatized to 7.5 °C; (●) June, 30 mussels, 29 ± 2.6 mm, acclimatized to 12 °C. From Jørgensen et al. (1990).
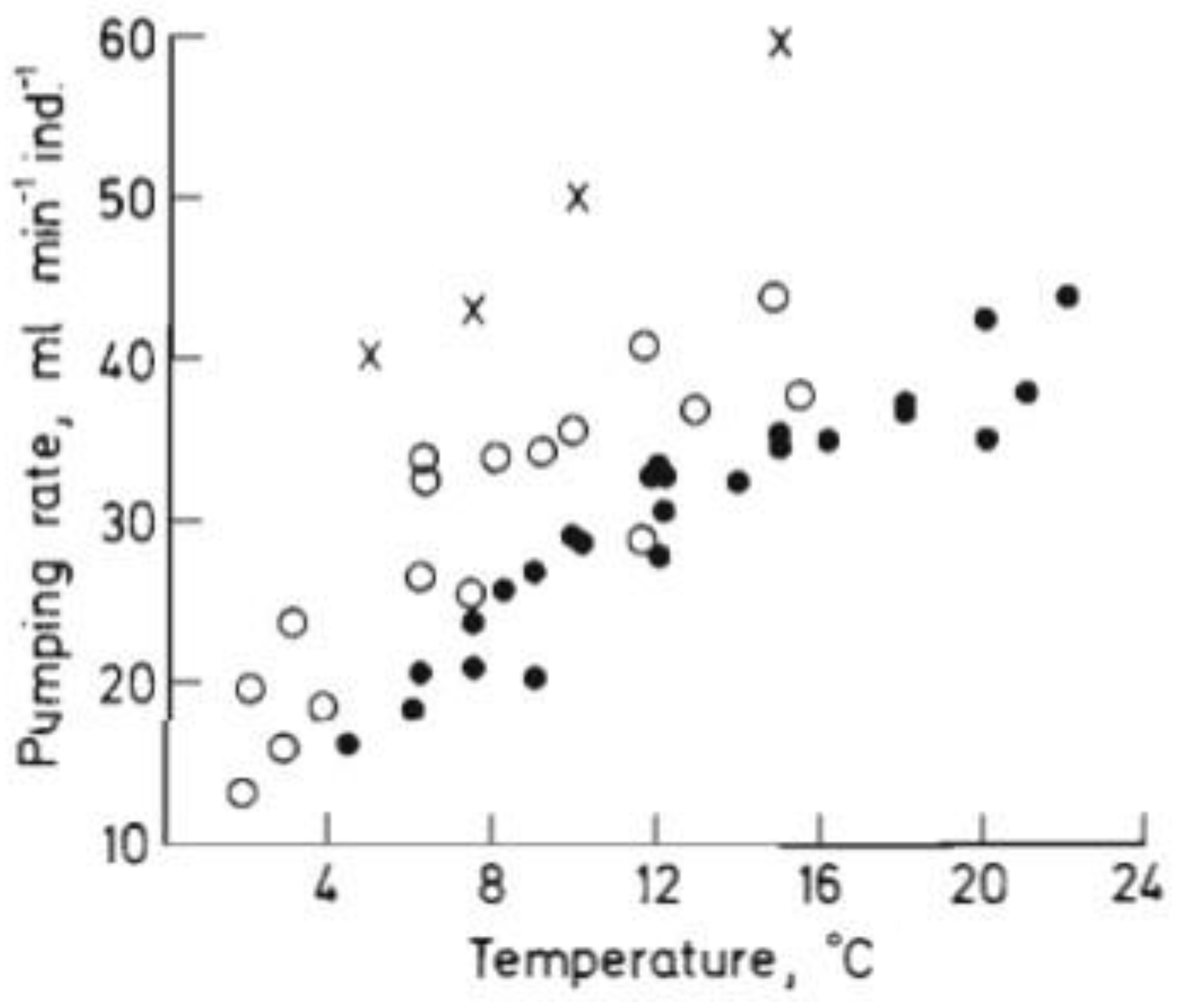
Figure 3.
Mytilus edulis. Relationship between pumping rate (= filtration rate) and temperature-dependent kinematic viscosity of seawater in the groups of mussels shown in Figure 2, using the same symbols.
Figure 3.
Mytilus edulis. Relationship between pumping rate (= filtration rate) and temperature-dependent kinematic viscosity of seawater in the groups of mussels shown in Figure 2, using the same symbols.
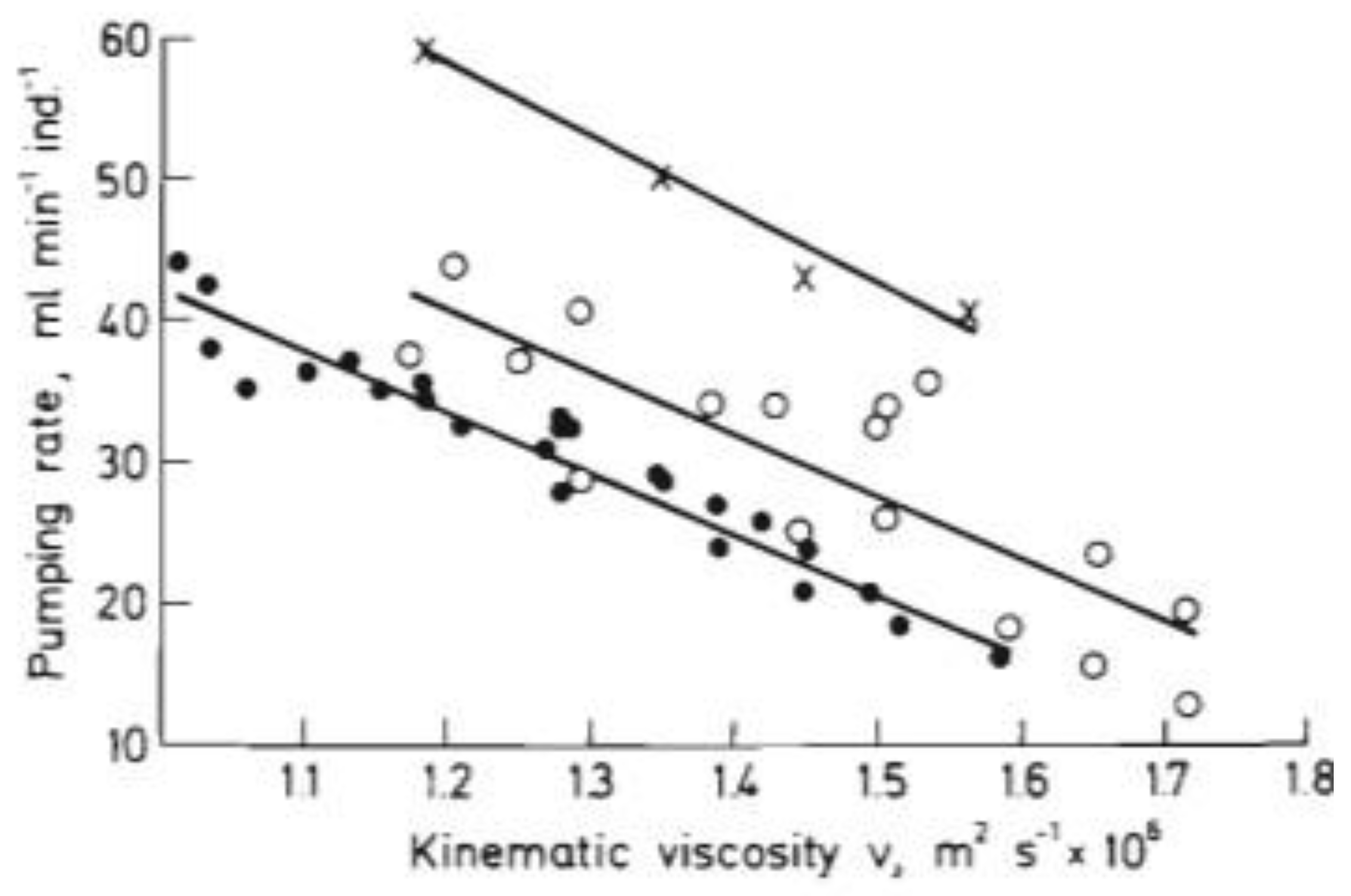
Figure 4.
Mytilus edulis. Individual filtration rate of mussels (50 mm shell length, n = 5) as a function of stepwise increasing and decreasing temperature as indicated by the inserted numbers starting at (1). (A) Mussels seasonally acclimated to 18°C. (B) The same group of mussels after 1 week of acclimation to 11°C. The regression lines for filtration rate as a function of temperature in the temperature-tolerance interval and their equations are show. It is seen that there is a linear and reversible effect of temperature. From Kittner & Riisgård (2005).
Figure 4.
Mytilus edulis. Individual filtration rate of mussels (50 mm shell length, n = 5) as a function of stepwise increasing and decreasing temperature as indicated by the inserted numbers starting at (1). (A) Mussels seasonally acclimated to 18°C. (B) The same group of mussels after 1 week of acclimation to 11°C. The regression lines for filtration rate as a function of temperature in the temperature-tolerance interval and their equations are show. It is seen that there is a linear and reversible effect of temperature. From Kittner & Riisgård (2005).
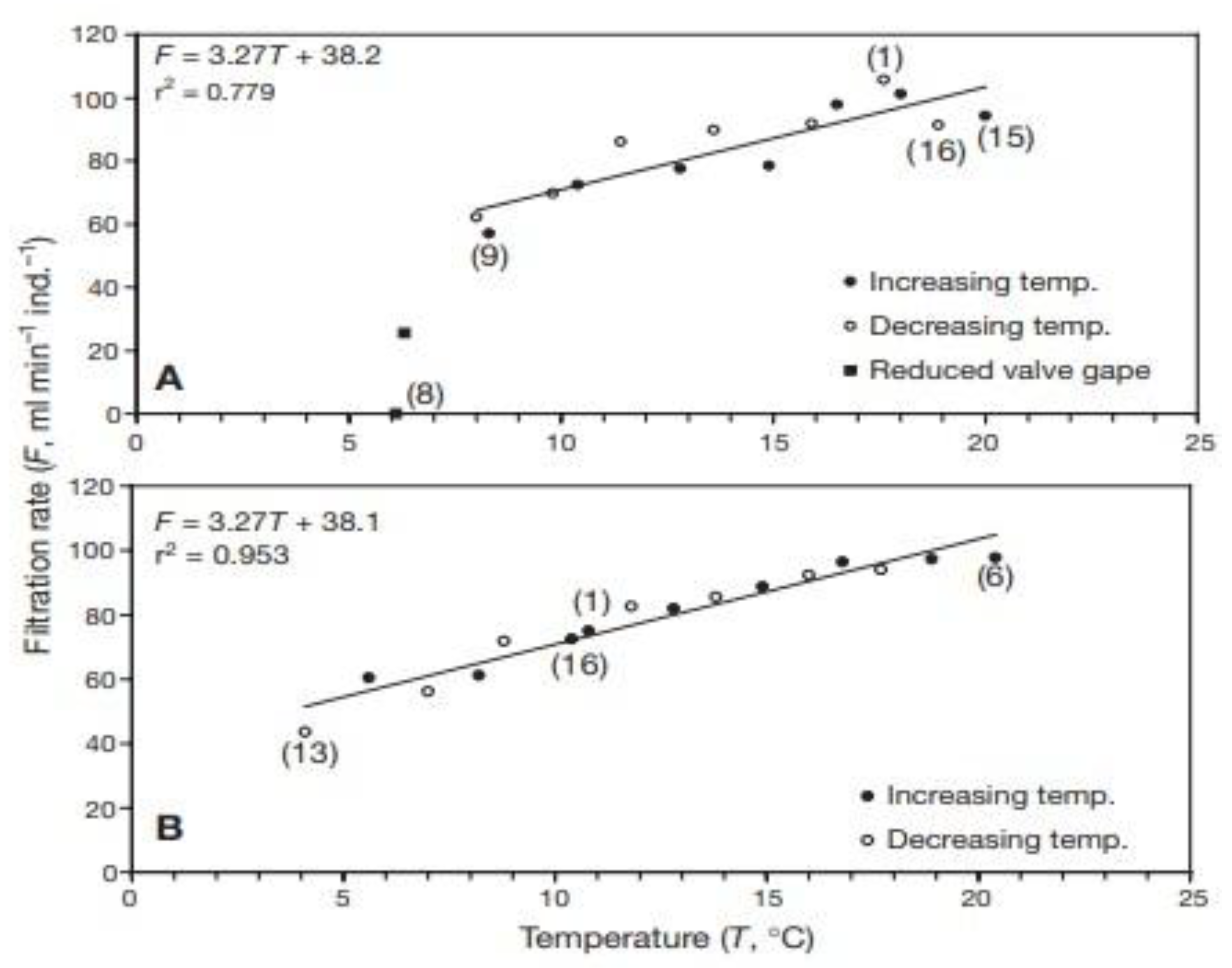
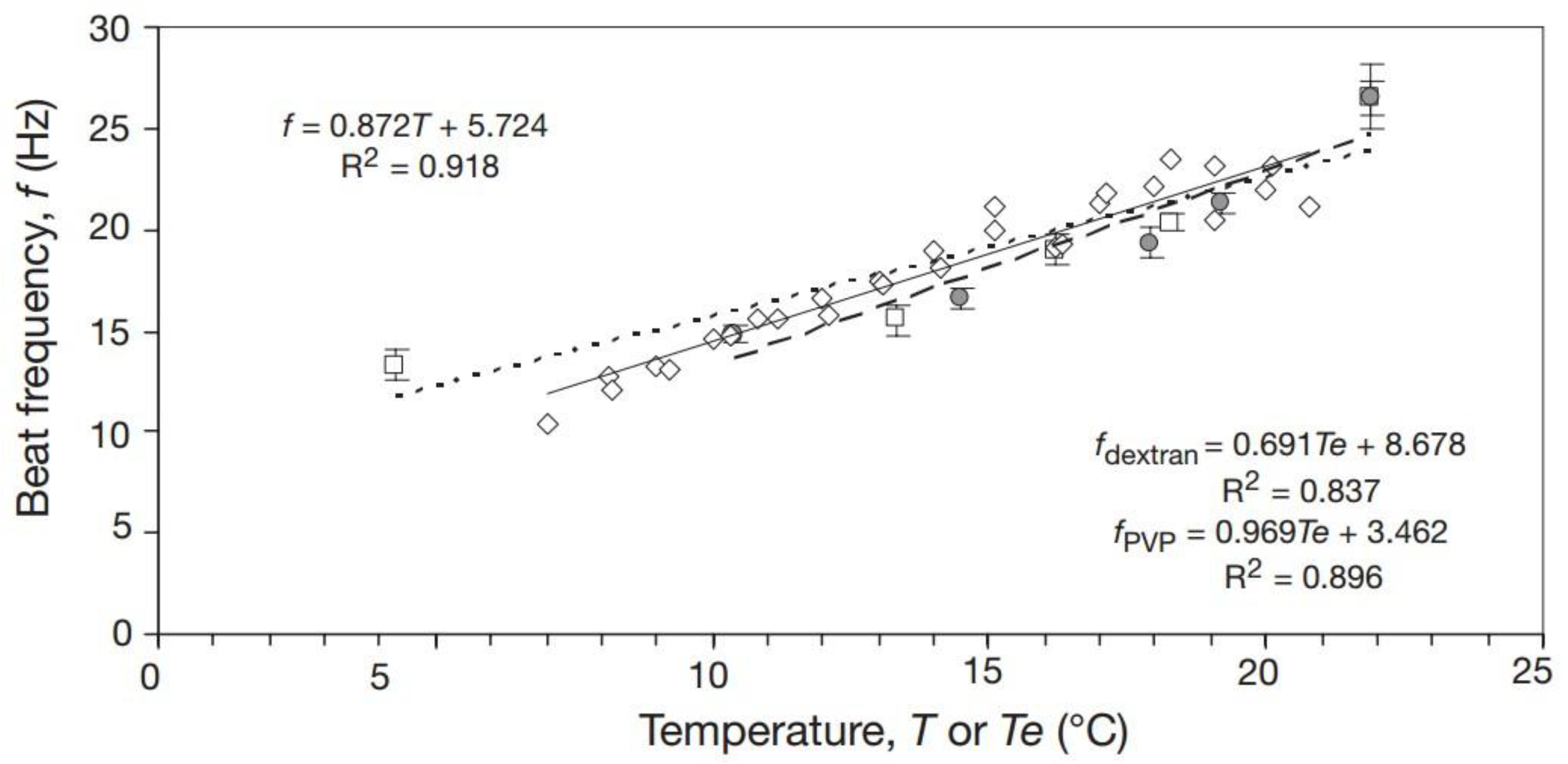
Figure 5.
Mytilus edulis. Beat frequency of water-pumping lateral cilia in mussel-gill preparations stimulated with serotonin (10-5 M) versus temperature (T) or temperature equivalents (Te); (◊) exposed to different temperatures of the seawater; (□) exposed to different dextran-manipulated viscosities of seawater (22°C, 20 psu); (○) exposed to different PVP-manipulated viscosities of seawater (22°C, 20 psu). Mean (±2 SD, n = 8) indicated for PVP and dextran. It is seen that the effect of temperature on lateral cilia activity is controlled by the viscosity of the ambient seawater. From Riisgård & Larsen (2007).
Figure 5.
Mytilus edulis. Beat frequency of water-pumping lateral cilia in mussel-gill preparations stimulated with serotonin (10-5 M) versus temperature (T) or temperature equivalents (Te); (◊) exposed to different temperatures of the seawater; (□) exposed to different dextran-manipulated viscosities of seawater (22°C, 20 psu); (○) exposed to different PVP-manipulated viscosities of seawater (22°C, 20 psu). Mean (±2 SD, n = 8) indicated for PVP and dextran. It is seen that the effect of temperature on lateral cilia activity is controlled by the viscosity of the ambient seawater. From Riisgård & Larsen (2007).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



