
Submitted:
10 September 2025
Posted:
12 September 2025
You are already at the latest version
Abstract
Background: The frequency of necrotizing cutaneous and soft tissue infections caused by the Mucorales fungi Apophysomyces and Sakasenaea is increasing. The absence of sophis-ticated diagnostic technologies in low- and middle-income countries (LMIC), means that detection of cutaneous mucormycosis continues to rely on culture of the infecting path-ogens from biopsy, and their differentiation based on morphological characteristics. However, Apophysomyces and Sakasenaea are notorious for their failure to sporulate on standard mycological media used for the identification of human pathogenic fungi. Differentiation of these pathogens and their discrimination from Aspergillus fumigatus, the most common mold pathogen of humans, is essential due to their differing sensitivities to the antifungal drugs used to treat mucormycosis. Methods: A murine IgG1 monoclonal antibody, JD4, has been developed which is specific to Apophysomyces species. In western blotting and enzyme-linked immunosorbent assay (ELISA), mAb JD4 is shown to bind to an extracellular 15 kDa protein, readily detectable in crude antigen extracts from non-sporulating cultures of Apophysomyces. Results: When combined with a Mu-corales-specific lateral-flow immunoassay (LFIA), mAb JD4 allows the differentiation of Apophysomyces from Saksenaea species, and discrimination from Aspergillus fumigatus. Conclusions: Monoclonal antibody JD4 enables the detection and differentiation of Apophysomyces species from other fungal pathogens of humans which cause rapidly progressive cutaneous and soft tissue mycoses of humans. When combined with a rapid LFIA, improvements are offered in the sensitivity and specificity of Mucorales detection based on mycological culture which remains a gold standard procedure for mucormycosis detection in LMIC lacking access to more sophisticated diagnostic procedures.
Keywords:
Mucorales
; Apophysomyces
; Saksenaea
; mucormycosis
; monoclonal antibody
; antigen
; lateral-flow immunoassay
; ELISA
1. Introduction
Mucormycosis is a lethal angio-invasive disease of humans caused by fungi in the class Zygomycetes, order Mucorales, recently identified as a high priority pathogen group [1,2,3]. The Mucorales fungi most commonly reported as human pathogens are species of Lichtheimia, Mucor, and Rhizopus, followed by Apophysomyces, Cunninghamella, Rhizomucor and Saksenaea [4,5]. Apophysomyces spp. are the second most common agents of rhino-orbital-cerebral mucormycosis (ROCM) in India after Rhizopus arrhizus [6,7,8] and are responsible, along with Saksenaea spp., for the majority of cases of cutaneous and soft tissue mucormycosis in the form of necrotizing fasciitis [9,10,11,12,13,14]. As emerging pathogens of humans [15,16], reports of Apophysomyces spp. in necrotizing cutaneous and soft tissue infections of immunocompetent individuals following penetrating trauma and burns [4,13,17,18,19,20,21,22,23,24,25] have increased over recent years, particularly following natural disasters such as tornadoes and tsunamis, and following combat-related blast injuries [26,27,28,29,30].
The prompt detection and differentiation of Mucorales spp. from other mold pathogens such as Aspergillus fumigatus is critical to patient survival, enabling timely treatment with surgery and Mucorales-active antifungal drugs [4]. Detection of Mucorales pathogens has traditionally relied on fungal culture from a biopsy sample and identification by microscopy of characteristic morphological structures. However, Apophysomyces and Saksenaea species are notorious for their failure to sporulate on routine isolation media used for the cultivation of filamentous fungi [9,11,31,32], requiring instead lengthy and laborious methods to induce sporulation for identification by specialist medical mycologists [33,34,35,36,37]. For this reason, more sophisticated and costly diagnostic modalities such as DNA sequencing or matrix assisted laser desorption ionization time of flight (MALDI-TOF) are used for identification following culture in vitro [11,38,39], but widespread access to such technology is limited, especially in low- and middle-income countries (LMIC) which continue to rely on culture for the detection of fungal infections, and which remains a cornerstone for definitive diagnosis of mucormycosis worldwide [4,14]. Indeed, the majority of cases of mucormycosis during the second wave of the COVID-19 pandemic in India were diagnosed by culture of the infecting fungi [40,41].
Antigens that can act as biomarkers for Mucorales fungi have recently been reported, including extracellular polysaccharides (EPS) detectable using species-specific [42] and pan-Mucorales-specific [43] monoclonal antibodies (mAbs) in lateral-flow immunoassay (LFIA) and in immunohistochemistry [42,43,44,45].
In this paper, we report the development of a murine IgG1 mAb, JD4, which binds to a major 15-kDa antigen specific to the genus Apophysomyces. We show how the mAb can be employed in an enzyme-linked immunosorbent assay (ELISA) to differentiate Apophysomyces spp. from Saksenaea spp., following rapid discrimination of Mucorales pathogens from Aspergillus fumigatus using a simple culture-based antigen extraction procedure and Mucorales-specific LFIA.
2. Materials and Methods
2.1. Ethics Statement
Hybridoma work described in this study was conducted under a UK Home Office Project License, and was reviewed by the institution’s Animal Welfare Ethical Review Board (AWERB) for approval. The work was carried out in accordance with The Animals (Scientific Procedures) Act 1986 Directive 2010/63/EU, and followed all the Codes of Practice which reinforce this law, including all elements of housing, care, and euthanasia of the animals.
2.2. Fungal Culture
Fungi (Table 1) were routinely cultured on malt extract agar (MEA; 70145, Sigma). The medium was autoclaved at 121 oC for 15 min prior to use, and fungi were grown at 37 oC. To induce sporulation in Apophysomyces spp., the fungi were grown on autoclaved Czapek Dox agar (70185, Sigma) at 37 oC. To induce sporulation of Saksenaea spp., the method of Padhye and Ajello [34] was used.
2.3. Production of Hybridomas and Screening by ELISA
Extracellular polysaccharides (EPS) were prepared according to Davies and Thornton [42]. For hybridoma production, the immunogen comprised a 1 mg/mL solution of EPS from Apophysomyces variabilis CBS658.93, with six-week-old BALB/c white mice each given four intra-peritoneal injections (300 µL per injection) of immunogen at 2-wk intervals, and a single booster injection 5 d before fusion. Hybridoma cells were produced by the method described elsewhere [46], and monoclonal antibody (mAb)-producing clones identified in indirect ELISA tests by using 20 μg EPS/mL phosphate-buffered saline (PBS; 137 mM NaCl, 2.7 mM KCl, 8 mM Na2HPO4, 1.5 mM KH2PO4 [pH 7.2]) immobilized to the wells of Maxisorp microtiter plates (Nunc) at 50 µL/well. Wells containing immobilized antigen were incubated with 50 µL of mAb hybridoma tissue culture supernatant (TCS) for 1 h, after which wells were washed three times, for 5 min each, with PBST (PBS containing 0.05% (vol:vol) Tween-20). Goat anti-mouse polyvalent immunoglobulin (G, A, M) peroxidase conjugate (PA1-84388; Invitrogen), diluted 1:5000 (vol:vol) in PBST, was added to the wells and incubated for a further hour. The plates were washed with PBST as described, given a final 5 min wash with PBS, and bound antibody visualised by incubating wells with tetramethyl benzidine (TMB) substrate solution [46] for 30 min, after which reactions were stopped by the addition of 3 M H2SO4. Absorbance values were determined at 450 nm using a microplate reader (infinite F50, Tecan Austria GmbH). Control wells were incubated with tissue culture medium (TCM) containing 10% (vol:vol) foetal bovine serum (F7254, Sigma) only. All incubation steps were performed at 23 °C in sealed plastic bags. The threshold for detection of the antigen in indirect ELISA was determined from control means (2 x TCM absorbance values). These values were consistently in the range of 0.050-0.100. Consequently, absorbance values ≥0.100 were considered as positive for the detection of antigen.
2.4. Determination of Ig Class and Sub-Cloning Procedure
The Ig class of mAbs was determined by using antigen-mediated ELISA [46]. Wells of microtiter plates coated with 20 mg EPS/mL PBS were incubated successively with hybridoma TCS for 1 h, followed by goat anti-mouse IgG1, IgG2a, IgG2b, IgG3, IgM or IgA-specific antiserum (ISO-2, Sigma) diluted 1:3000 (vol:vol) in PBST for 30 min, and rabbit anti-goat peroxidase conjugate (A5420, Sigma) diluted 1:1000 (vol:vol) for a further 30 min. Bound antibody was visualised with TMB substrate as described. Hybridoma cell lines were sub-cloned three times by limiting dilution, and cell lines were grown in bulk in a non-selective medium, preserved by slowly freezing in FBS/dimethyl sulfoxide (92:8 vol:vol), and stored in liquid N2.
2.5. Antibody Purification and Enzyme Conjugation
Hybridoma TCS of mAb JD4 was harvested by centrifugation at 2,147 x g for 40 min at 4 °C, followed by filtration through a 0.8 μM cellulose acetate filter (10462240, GE Healthcare Life Sciences, UK). Culture supernatant was loaded onto a HiTrap Protein A column (17-0402-01, GE Healthcare Life Sciences) using a peristaltic pump P-1 (18-1110-91, GE Healthcare Life Sciences) with a low pulsation flow of 1 mL/min. Columns were equilibrated with 10 mL of PBS, and column-bound antibody was eluted with 5 mL of 0.1 M glycine-HCl buffer (pH 2.5) with a flow rate of 0.5 mL/min. The buffer of the purified antibody was exchanged to PBS using a disposable PD-10 desalting column (17-0851-01, GE Healthcare Life Sciences). Following purification, the antibody was sterile filtered with a 0.24 µm syringe filter (85037-574-44, Sartorius) and stored at 4 °C. Protein concentrations were determined using a Nanodrop spectrophotometer with protein concentration calculated using the mass extinction coefficient of 13.7 at 280 nm for a 1% (10 mg/mL) IgG solution. Antibody purity was confirmed by SDS-PAGE and gel staining using Coomassie Brilliant Blue R-250 dye (Thermo Fisher Scientific). Protein A-purified mAb JD4 was conjugated to horseradish peroxidase (HRP) for ELISA studies using a Lightning-Link horseradish peroxidase conjugation kit (701-0000, Bio-Techne Ltd.), or to alkaline phosphatase (AKP) for western blotting studies using a Lightning-Link alkaline phosphatase conjugation kit (702-0010, Bio-Techne Ltd.).
2.6. Determination of Antibody Specificity
For antibody specificity tests, wells were coated with purified EPS antigens at a concentration 20 μg/mL PBS from different Mucorales fungi and from unrelated molds of clinical importance (Table 1). The wells were assayed by Direct-ELISA using the JD4-HRP conjugate diluted 1:5000 (vol:vol) in PBST for 1 h, followed by TMB substrate solution for 30 min.
2.7. Epitope Characterisation by Heat
Heat stability of the JD4 epitope was determined by heating EPS from Apophysomyces variabilis strain CBS658.93 at a concentration of 20 μg/mL PBS in a boiling water bath. At 10 min intervals, 50 μL volumes were removed and, after cooling, were transferred to the wells of microtiter plates for assay by Direct-ELISA using the JD4-HRP conjugate as described.
2.8. Polyacrylamide Gel Electrophoresis and Western Blotting
Sodium-dodecyl-sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) was carried out using 4–20% gradient polyacrylamide gels (161-1159, Bio-Rad) under denaturing conditions. Antigens were separated electrophoretically at 165 V and pre-stained markers (161-0318, Bio-Rad) were used for molecular weight determinations. For western blotting, separated antigens were transferred electrophoretically onto a PVDF membrane (162-0175, Bio-Rad) for 2 h at 75 V, and the membrane was blocked for 16 h at 4 °C in PBS containing 1% (wt:vol) BSA. Blocked membranes were incubated with JD4-AKP diluted 1:15,000 (vol:vol) in PBS containing 0.5% (wt:vol) BSA (PBSA) for 2 h at 23 °C. Membranes were washed three times with PBS, once with PBST and bound antibody visualised by incubation in substrate solution. Reactions were stopped by immersing membranes in dH2O, and membranes were then air dried between sheets of Whatman filter paper.
2.9. Antigen Production In Vitro
For antigen production studies, A. variabilis CBS658.93 was grown in YNB+G medium [42,43]. Three replicate 250 mL conical flasks containing 100 mL of autoclaved medium were inoculated with spores to a final concentration of 103 spores/mL, and the cultures incubated at 37 oC with shaking (120 RPM) in a New Brunswick orbital shaker. At 24 h intervals, replicate flasks were harvested and culture fluids separated from mycelium by filtration through Miracloth (475855-1R, Merck Millipore, Germany). Mycelial biomass was dried for 4 d at 80 oC and weighed. Protein precipitates were prepared by mixing culture filtrates with ethanol at a ratio of 1:4 (vol:vol) in 50 mL Falcon tubes (62.547.254, Sarstedt AG, Germany) and chilling at -20 oC for 16 h. After centrifugation for 10 min at 3202 g, the clear supernatants were aspirated, the pellets re-suspended in 1 mL PBS and the suspensions then stored at -20 oC. On thawing, any insoluble material was removed by centrifugation for 5 min at 16,000 g, and then assayed by Direct-ELISA with JD4-HRP conjugate as described.
2.10. Culture-Based Detection and Differentiation of Mucorales Fungi
Three replicate 6 cm diameter culture plates (628161, Greiner) containing MEA were inoculated centrally with 5 mm diameter discs of mycelium generated, using a #2 cork-borer, from MEA cultures of Mucorales spp. and the non-sporulating mutant (strain ΔAfbrlA7 [47]) of Aspergillus fumigatus (Table 2). The culture plates were incubated at 37 oC for 24 h after which five discs of mycelium, generated using the cork borer (Figure 1, Step 1 and Supplementary Figure S1A), were transferred to three replicate 25 mL sterile Universal tubes each containing 10 mL of YNB+G medium. The tubes were incubated at 37 oC for 24 h with horizontal shaking (60 RPM) after which the culture fluids were separated from the fungal microcolonies (Supplementary Figure S1B) by filtering through Miracloth. Protein precipitates were prepared by mixing the culture fluids with ethanol at a ratio of 1:4 (vol:vol) in 50 mL Falcon tubes and chilling at -20 oC for 7 h. The mixtures were centrifuged at 3,202 x g for 10 min, the pelleted material resuspended in 1 mL PBS, and 50 μL volumes of suspension used to coat the wells of microtiter plates for assay by Direct-ELISA using the JD4-HRP conjugate as described. For the detection of Aspergillus antigen in Direct-ELISA, the Aspergillus-specific mAb JF5 [48] was directly conjugated to HRP as described and used at a concentration of 1:5000 (vol:vol) in PBST. Hyphal biomass retained on Miracloth filters was heated in an oven at 4 d at 80 oC and dry weights determined.
For lateral-flow immunoassay, solubilized antigens were prepared from the 24-h-old MEA cultures following removal of the mycelial discs (Figure 1, Step 1). Sterile disposable sampling (nasal) swabs (Type G-015, Jiangsu HanHeng Medical Technology Co. Ltd., Changzhou, China) were wetted in 250 μL of TG11-LFD running buffer (RB; PBS containing 0.05% (vol:vol) Tween-20 and 0.05% (vol:vol) Octoxinol-10) contained in 1.5 mL microcentrifuge tubes. The wetted swabs were used to rub the surfaces of the 24-h-old MEA cultures (Figure 1, Step 1) and then returned to the tubes containing the RB for recovery of soluble antigen (Figure 1, Step 2). Antigen extracts were then centrifuged for 5 min at 17,000 x g (Figure 1, Step 3) and 100 μL of supernatant applied to TG11-LFD tests (Figure 1, Step 4). After 30 min, the intensities of the test (T) and control (C) lines were determined as artificial units (a.u.) using a Cube reader [42,43].
2.11. Statistical Analysis
Numerical data were analysed using the statistical programme Minitab (Minitab 16; Minitab, Coventry, UK). Analysis of variance (ANOVA) was used to compare means, and post hoc Tukey–Kramer analysis was then performed to determine statistical significance.
3. Results
3.1. Production of Hybridomas and mAb Isotyping
A single hybridoma fusion was performed, and 327 hybridoma cell lines were tested in indirect ELISA tests for recognition of the immunogen. Seven cell lines produced EPS-reactive antibodies, 5 of which produced mAbs of the immunoglobulin class, G1 (IgG1). The remaining 2 cell lines produced mAbs of the class, G2b (IgG2b). The cell line JD4 (an IgG1) was selected for further evaluation due to its specificity for Apophysomyces species, and lack of cross-reactivity with non-Mucorales fungi.
3.2. Specificity of mAb JD4 and Epitope Characterisation
In Direct-ELISA tests of purified EPS from a range of clinically-relevant Mucorales and non-Mucorales mold pathogens, mAb JD4 was found to be specific to fungi in the Mucorales genus, Apophysomyces (Table 1). No cross-reactivity was found with the other Mucorales pathogens tested (Cunninghamella, Lichtheimia, Mucor, Rhizomucor, Rhizopus, Saksenaea) or with unrelated molds of clinical importance (Aspergillus, Fusarium, Lomentospora, and Scedosporium spp.) [49]. Direct-ELISA tests showed that mAb JD4 binds to a heat-labile epitope (Figure 1A), present in a single 15 kDa Apophysomyces-specific protein in western immunoblots, but absent in purified EPS preparations from other Mucorales fungi and from Aspergillus fumigatus (Figure 1B,C).
3.3. Antigen Production In Vitro
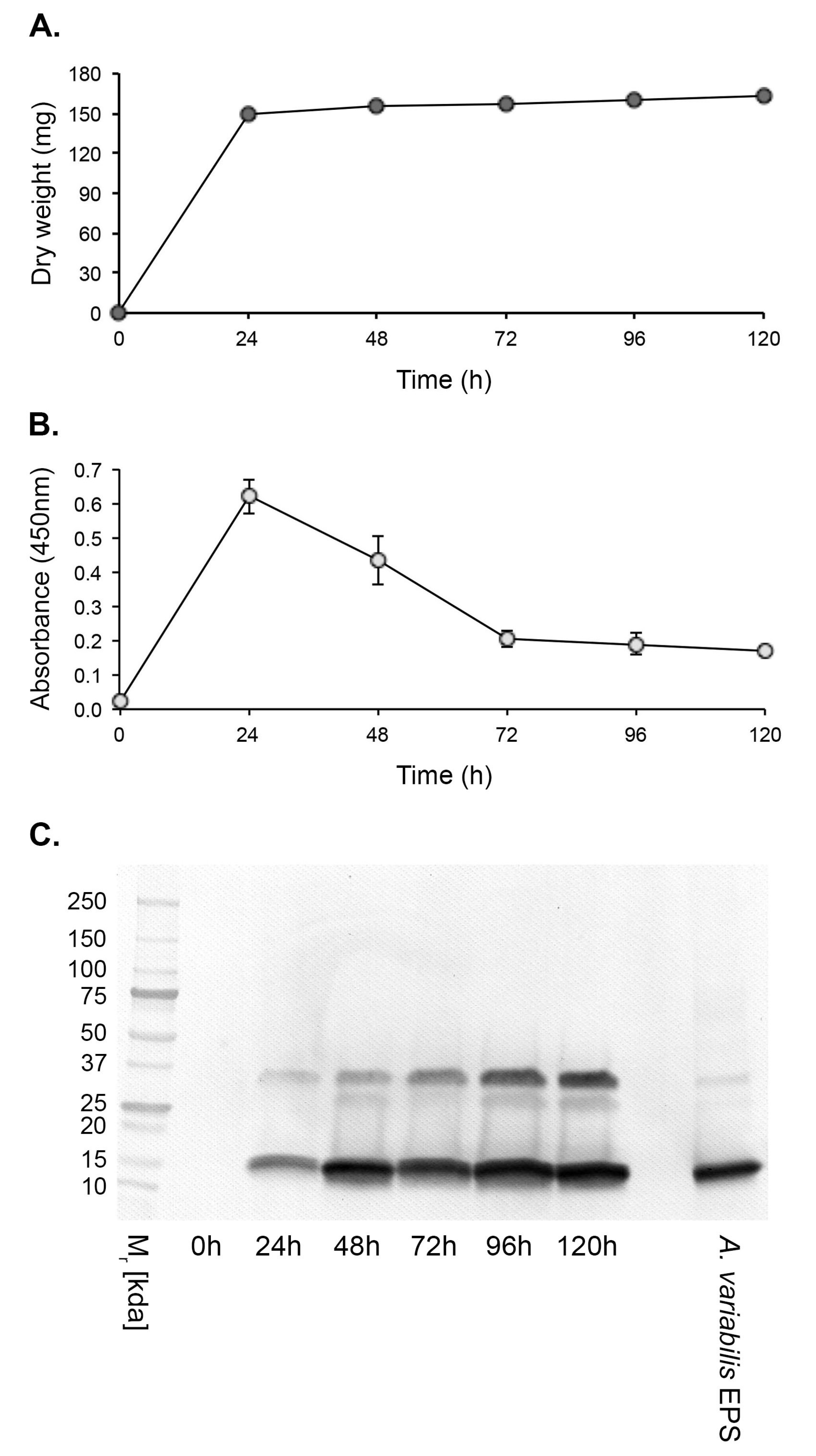
Hyphal growth of A. variabilis CBS658.93 in YNB+G shake culture plateaued at 24 h post-inoculation with a mean dry weight of 148.93 mg ± 0.020 mg (Figure 3A). Antigen production, determined by Direct-ELISA, peaked after 24 h and declined thereafter until the end of the experimental period at 120 h (Figure 3B). In western immunoblots, a 15 kDa immunoreactive antigen was first discernible at 24 h and was detectable throughout the experimental period (Figure 3 C). A second immunoreactive band of ~35 kDa was also evident, possibly as a dimer of the 15 kDa protein.
Figure 2.
Epitope characterization by heat treatment and specificity of mAb JD4. (A) JD4 Direct-ELISA of the Apophysomyces variabilis CBS658.93 EPS antigen following heating at 100 oC for 10, 20, 30, 40, 50 and, 60 min. Bars are the means of three replicates ±SE, and bars with the same letter are not significantly different at p < 0.05. After 10 min of heat treatment, there was a significant (p < 0.05) decrease in ELISA absorbance value compared to the control (no heat treatment) indicating that the mAb binds to a heat labile epitope. (B) Western immunoblot of EPS antigens from Mucorales fungi of clinical importance, showing a major JD4 immunoreactive protein of 15 kDa specific to Apophysomyces variabilis. (C) Western immunoblot of purified EPS prepared from species of the Mucorales fungi, Apophysomyces and Saksenaea, and from the fungus Aspergillus fumigatus. Immunoreactive proteins of 15 kDa are present in the purified EPS preparations of Apophysomyces spp., but absent in Saksenaea spp. and Aspergillus fumigatus preparations.
Figure 2.
Epitope characterization by heat treatment and specificity of mAb JD4. (A) JD4 Direct-ELISA of the Apophysomyces variabilis CBS658.93 EPS antigen following heating at 100 oC for 10, 20, 30, 40, 50 and, 60 min. Bars are the means of three replicates ±SE, and bars with the same letter are not significantly different at p < 0.05. After 10 min of heat treatment, there was a significant (p < 0.05) decrease in ELISA absorbance value compared to the control (no heat treatment) indicating that the mAb binds to a heat labile epitope. (B) Western immunoblot of EPS antigens from Mucorales fungi of clinical importance, showing a major JD4 immunoreactive protein of 15 kDa specific to Apophysomyces variabilis. (C) Western immunoblot of purified EPS prepared from species of the Mucorales fungi, Apophysomyces and Saksenaea, and from the fungus Aspergillus fumigatus. Immunoreactive proteins of 15 kDa are present in the purified EPS preparations of Apophysomyces spp., but absent in Saksenaea spp. and Aspergillus fumigatus preparations.
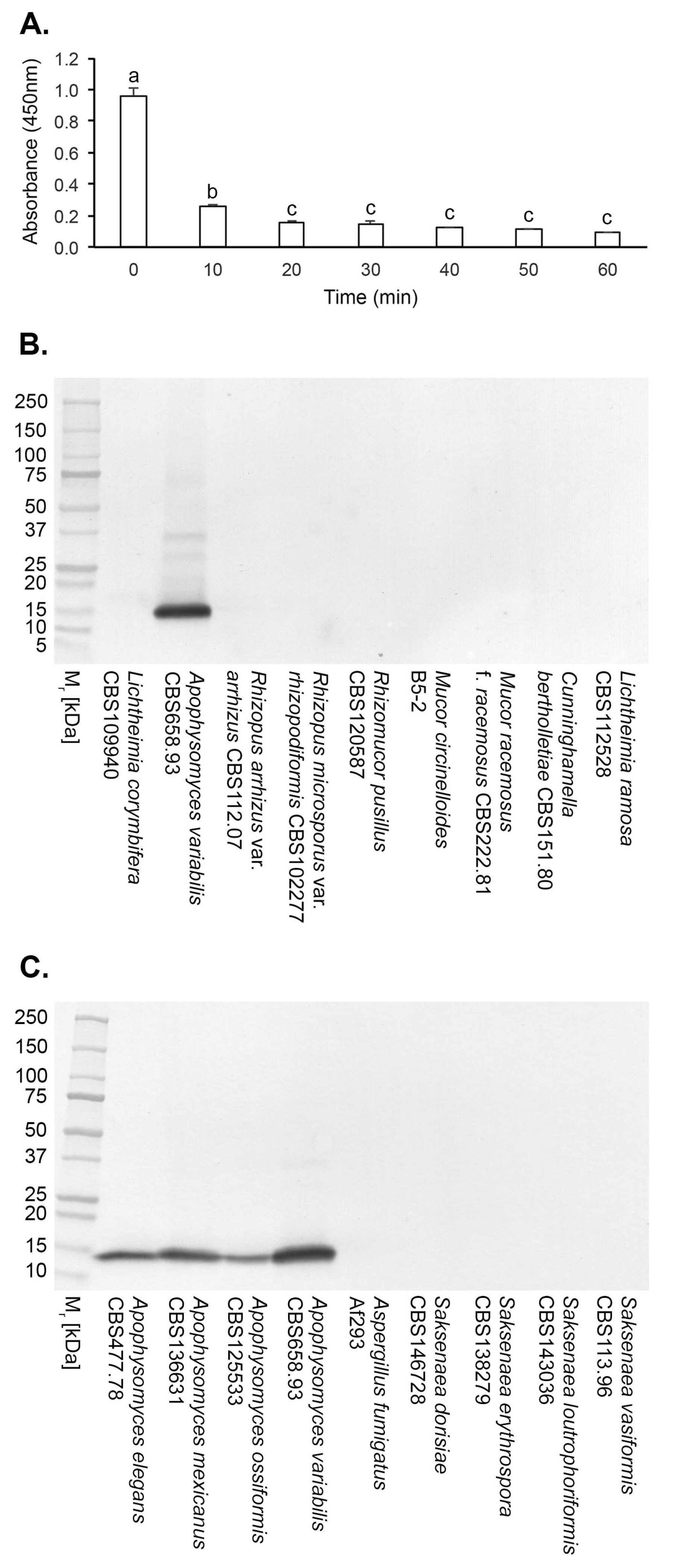
Figure 3.
In vitro production of the JD4 antigen by A. variabilis CBS658.93. (A) Dry weights of the pathogen over the 120-h experimental period. (B) Direct-ELISA of YNB+G protein precipitates using mAb JD4. Each data point is the mean of three replicates ±SE (B,C). (C) Western immunoblots of protein precipitates, showing the presence of a 15 kDa immunoreactive protein, and a higher molecular weight species of ~35 kDa. The positive control consisted of purified A. variabilis EPS antigen at a concentration of 80 ng/mL.
Figure 3.
In vitro production of the JD4 antigen by A. variabilis CBS658.93. (A) Dry weights of the pathogen over the 120-h experimental period. (B) Direct-ELISA of YNB+G protein precipitates using mAb JD4. Each data point is the mean of three replicates ±SE (B,C). (C) Western immunoblots of protein precipitates, showing the presence of a 15 kDa immunoreactive protein, and a higher molecular weight species of ~35 kDa. The positive control consisted of purified A. variabilis EPS antigen at a concentration of 80 ng/mL.
3.4. Culture-Based Detection and Differentiation of Mucorales Fungi
3.4.1. Lateral-flow immunoassay
Using crude antigen extracts from swabs of 24-h-old MEA cultures of the fungi, the TG11-LFD test was shown to be rapid and specific, detecting the non-sporulating Mucorales species of Apophysomyces and Saksenaea, and a strain of Lichtheimia ramosa (DRH226533346) recovered from a biopsy of cutaneous mucormycosis, only. Antigen extracts from these Mucorales fungi gave mean test (T) line Cube reader values of <40 a.u. (Table 2), below the threshold a.u. for test positivity (≤400 a.u.). Extracts from the non-sporulating mutant of Aspergillus fumigatus (strain ΔAfbrlA7) gave a mean T line a.u. value of 474.0±19.0, showing lack of detection of this pathogen. The negative controls (RB only and purified EPS from A. fumigatus Af293 at a concentration of 100 μg/mL RB), were similarly negative with mean T line a.u. values >400 a.u.. The positive control comprising purified EPS from the Lichtheimia corymbifera strain CBS109940 at a concentration of 100 μg/mL RB, gave a positive test result, with mean T line values of 29.5±2.2.
3.4.2. Direct-ELISA
In Direct-ELISA tests of protein precipitates from fungi grown for 24-h-old in YNB+G shake cultures (Figure 4A; Supplementary Figure S1B), mAb JD4 was shown to be genus-specific, reacting with Apophysomyces species only (Figure 4B). The Aspergillus-specific mAb, JF5, acting as an immunoglobulin G (IgG) control, reacted with A. fumigatus Af293 only (Figure 4C).
4. Discussion
Mucormycosis is an aggressive angio-invasive disease of humans, manifesting as life-threatening rhino-orbital-cerebral, pulmonary, gastro-intestinal, cutaneous and disseminated infections [41]. Despite best-practice treatment, overall all-cause mortality remains high at approximately 50% [50]. While cutaneous disease is associated with lower mortality of around 30%, extensive surgery including limb amputation is frequently required to control the infection. Due to the rapid progression and destructive nature of mucormycosis, early diagnosis is crucial to improving mortality and limiting tissue loss and disfigurement [4,13].
Options for the treatment of mucormycosis with antifungal drugs are limited [51,52]. It is imperative that Mucorales pathogens are differentiated from Aspergillus fumigatus due to intrinsic resistance of Mucorales species to short-tailed azoles, such as voriconazole, used as a front-line antifungal drug for aspergillosis [53], the most frequent differential diagnosis associated with mucormycosis [54]. Furthermore, discrimination of Apophysomyces from Sakasenaea species is required, since Apophysomyces spp. display decreased sensitivity to amphotericin B [55,56], the antifungal drug most effective for treatment of mucormycosis [4,57].
Currently, diagnosis of cutaneous mucormycosis requires histochemistry and culture from a tissue biopsy, with identification of characteristic morphological features (broad ribbon-like aseptate hyphae and spore-bearing structures) remaining the gold standard procedure for detection and differentiation of infecting species. However, Apophysomyces and Saksenaea species are notorious in their failure to sporulate on standard mycological media, preventing accurate identification and delaying appropriate treatment. To mitigate this, we report here the development of simple culture-based detection methods that incorporate Mucorales-specific immunoassays for the detection and discrimination of these fastidious pathogens. Rapid discrimination of Mucorales species from A. fumigatus is achieved with lateral-flow immunoassay, with subsequent differentiation of non-sporulating species of Apophysomyces and Saksenaea using a newly developed mAb, JD4, in ELISA.
The pan-Mucorales-specific monoclonal antibody (mAb), TG11, used in the lateral-flow immunoassay has been used previously to detect Mucorales invasive hyphae in lung samples using immunohistochemistry [44], and in a lateral-flow device (LFD) to detect the Mucorales EPS antigen in bronchoalveolar lavage (BAL) samples from patients with pulmonary mucormycosis [45]. In a competitive LFD format such as the pan-Mucorales-specific TG11-LFD [43], the test (T) line signal intensity is inversely proportional to the analyte concentration [58]. Consequently, a strong test line signal means that little or no of the target analyte (Mucorales EPS) is present in the sample. TG11-LFD tests using MEA culture swabs of the Mucorales pathogens resulted in test line “whiteouts” (complete absence of visible tests line signals) indicating positive test results (presence of the EPS analyte in swab samples). This equated to Cube reader values in the range of 15 to 40 artificial units (a.u.). Cube reader values for swabs from the non-sporulating mutant of A. fumigatus, ΔAfbrlA7 [47], and for A. fumigatus purified EPS, exceeded the threshold value for test positivity of ≥400 a.u. [43], further demonstrating the specificity of the TG11-LFD for Mucorales fungi. The simple and quick LFD swab test was therefore able to accurately differentiate non-sporulating Apophysomyes and Saksenaea species from the most common mold pathogen of humans, A. fumigatus. This is especially important for skin and soft tissue infections where the symptoms of cutaneous aspergillosis can overlap with those of cutaneous mucormycosis [59], or where mixed Mucorales and Aspergillus infections are present [14,60]. The TG11-LFD swab test was also able to rapidly detect a strain of Lichtheimia ramosa recovered from culture of a tissue biopsy from a patient with cutaneous mucormycosis. Consequently, this swab test is applicable to other Mucorales fungi capable of causing infections in humans, including those similarly capable of causing necrotizing fasciitis such as Lichtheimia [61,62]. The LFD test conforms to the ASSURED (affordable, sensitive, specific, user-friendly, rapid, equipment-free, delivered) criteria for diagnostics for the developing world [63], where access to more sophisticated and costly diagnostic tests such as qPCR [64], MALDI-TOF [38] or metagenomic next-generation sequencing (NGS) [65] is limited or absent.
The genera Apophysomyces and Saksenaea are ascribed to the family Saksenaeaceae within the order Mucorales, with both considered as species complexes [66]. Despite their close molecular taxonomic relatedness [67], mAb JD4 was able to discriminate between the two complexes, binding to a major 15 kDa antigen specific to the genus Apophysomyces. The JD4 antigen was readily detected in protein precipitates from all four Apophysomyces species reported as human pathogens [13], comprising A. elegans [68], A. mexicanus [69], A. ossiformis [70,71,72], and A. variabilis [8,16,19,22,24,25,39,73,74], but did not cross-react with species of Saksenaeae, notably the human pathogens S. dorisiae [36], S. erythrospora [75,76,77,78], and S. vasiformis [9,11,23,79]. We were unable to determine the reactivity of mAb JD4 with A. trapeziformis [26,80] due to the lack of availability of this species for testing.
Enzyme-Linked Immunosorbent Assay (ELISA) is a well-established and commonly used immunoassay performed in most diagnostic laboratories worldwide. The commercial availability of mAb JD4 means that diagnostic facilities equipped for this technique can use the mAb-based ELISA to detect and differentiate Apophysomyces species from other fungal pathogens of humans which cause rapidly progressive cutaneous and soft tissue mycoses of humans. When combined with the rapid TG11-LFD test, improvements are offered in the speed, sensitivity and specificity of Mucorales detection based on mycological culture, a gold standard procedure for mucormycosis detection [4,52].
Author Contributions
Conceptualisation, CRT and GD; methodology, CRT and GD; investigation, CRT and GD; formal analysis, CRT and GD; resources, CRT; data curation, CRT and GD; writing - original draft preparation, CRT; writing, review and editing, CRT and GD; project administration, CRT; funding acquisition, CRT.
Funding
This work was supported by ISCA Diagnostics Ltd. (project title: Development of Monoclonal Antibodies for Mucorales Detection).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author, but are not publicly available due to commercial confidentialities.
Conflicts of Interest
CRT is a Director of ISCA Diagnostics Ltd. This manuscript does not have any potential conflict of interest with ISCA Diagnostics Ltd.
References
- WHO (2022). WHO fungal priority pathogens list to guide research, development and public health action (Geneva: World Health Organization). License: CC BY-NC-SA 3.0 IGO.
- Brown, G.D.; Ballou, E.R.; Bates, S.; Bignell, E.M.; Borman, A.M.; Brand, A.C.; Brown, A.J.P.; Coelho, C.; Cook, P.C.; Farrer, R.A.; et al. The pathobiology of human fungal infections. Nat. Rev. Microbiol. 2024, 22, 687–704. [Google Scholar] [CrossRef]
- Morrissey, C.O.; Kim, H.Y.; Granham, K.; Dao, A.; Chakrabarti, A.; Perfect, J.R.; Alastruey-Izquierdo, A.; Harrison, T.S.; Bongomin, F.; Galas, M.; et al. Mucorales: a systematic review to inform the World Health Organization priority list of fungal pathogens. Med. Mycol. 2024, 62, myad130. [Google Scholar] [CrossRef] [PubMed]
- Cornely, O.A.; Alastruey-Izquierdo, A.; Arenz, D.; Chen, S.C.A.; Dannaoui, E.; Hochhegger, B.; Hoenigl, M.; Jensen, H.E.; Lagrou, K.; Lewis, R.E.; et al. Global guidelines for the diagnosis and management of mucormycosis: an initiative of the European Confederation of Medical Mycology in cooperation with the Mycoses Study Group Education and Research Consortium. Lancet Infect. Dis. 2019, 19, e405–e421. [Google Scholar] [CrossRef] [PubMed]
- Muthu, V.; Rudramurthy, S.; Chakrabarti, A.; Agarwal, R. Epidemiology and pathophysiology of COVID-19-associated mucormycosis: India versus the rest of the world. Mycopathologia 2021, 186, 739–754. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Singh, R. Mucormycosis in India: unique features. Mycoses 2014, 57, 85–90. [Google Scholar] [CrossRef]
- Patel, A.; Kaur, H.; Xess, I.; Michael, J.S.; Savio, J.; Rudramurthy, S.; Singh, R.; Shastri, P.; Umabala, P.; Sardana, R.; et al. A multicentre observational study on the epidemiology, risk factors, management and outcomes of mucormycosis in India. Clin. Microbiol. Infect. 2020, 26, 944e9–944e15. [Google Scholar] [CrossRef]
- Erami, M.; Mirhendi, H.; Momen-Heravi, M.; Hezaveh, S.J.H.; Ahsaniarani, A.H.; Sabet, A.S.; Aboutalebian, S. A case of COVID-19-associated rhino-orbito-cerebral mucormycosis caused by Apophysomyces variabilis with a review of the literature. Front. Cell. Infect. Microbiol. 2022, 12, 898477. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Kumar, P.; Padhye, A.A.; Chatha, L.; Singh, S.K.; Das, A.; Wig, J.D.; Kataria, R.N. Primary cutaneous zygomycosis due to Saksenaea vasiformis and Apophysomyces elegans. Clin. Infect. Dis. 1997, 24, 580–582. [Google Scholar] [CrossRef]
- Pamidimukkala, U.; Sudhaharan, S.; Kancharia, A.; Vemu, L.; Challa, S.; Karanam, S.D.; Chavali, P.; Prakash, H.; Ghosh, A.K.; Gupta, S.; et al. Mucormycosis due to Apophysomyces species complex – 25 years’ experience at a tertiary care hospital in southern India. Med. Mycol. 2020, 58, 425–433. [Google Scholar] [CrossRef]
- Singh, S.; Kanaujia, R.; Kumar, M.B.; Naga Santhosh Irrinki, R.N.; Satish, S.N.; Choudhary, H.; Kaur, H.; Rudramurthy, S.M. Saksenaea vasiformis infection: extensive abdominal necrotizing fasciitis with systemic review and analysis of 65 cases. Mycoses 2023, 66, 697–704. [Google Scholar] [CrossRef]
- Sykes, B.A.; Krause, H.; Lamparelli, M.; Austin, D. Saksenaea mucormycosis: a rare and dangerous cause of necotising fasciitis. BMJ Case Rep. 2023, 16, e254183. [Google Scholar] [CrossRef] [PubMed]
- Landré, V.; Klingebiel, F.K-L.; van Niftrik, C.H.B.; Goetze, E.; Speck, R.F.; Hübner, C.T.; Pape, H-C.; Schäfer, F.P. Mucormycosis caused by Apophysomyces elegans—a case report and systematic review of the literature of rhino-orbital-cerebral cases of the genus Apophysomyces. J. Fungi 2025, 11, 368.
- Skiada, A.; Drogari-Apiranthitou, M.; Roilides, E.; Chander, J.; Khostelidi, S.; Klimko, N.; Hamal, P.; Chrenkova, V.; Kanj, S.S.; El Zein, S.; et al. A global analysis of cases of mucormycosis recorded in the European Confederation of Medical Mycology / International Society for Human and Animal Mycology (ECMM / ISHAM) Zygomyco.net registry from 2009 to 2022. Mycopathologia 2025, 190, 53. [Google Scholar] [CrossRef]
- Prakash, H.; Ghosh, A.K.; Rudramurthy, S.M.; Paul, R.A.; Guota, S.; Negi, V.; Chakrabarti, A. The environmental source of emerging Apophysomyces variabilis infection in India. Med. Mycol. 2016, 54, 567–575. [Google Scholar] [CrossRef]
- Li, X-Y.; Wang, S-J.; Huang, G-M.; Zheng, D-Y.; Al-Odaini, N.; Pan, K-S.; Zheng, Y-Q.; Cao, C-W. Apophysomyces variabilis as an emerging pathogen in mainland China. Mycoses 2023, 66, 795-800.
- Liang, K.P.; Tleyjeh, I.M.; Wilson, W.R.; Roberts, G.D.; Temesgen, Z. Rhino-orbitocerebral mucormycosis caused by Apophysomyces elegans. J. Clin. Microbiol. 2006, 44, 892–898. [Google Scholar] [CrossRef]
- Vitrat-Hincky, V.; Lebeau, B.; Bozonnet, E.; Falcon, D.; Pradel, P.; Faure, O.; Aubert, A.; Piolat, C.; Grillot, R.; Pelloux, H. Severe filamentous fungal infections after widespread tissue damage due to traumatic injury: six cases and review of the literature. Scand. J. Infect. Dis. 2009, 41, 491–500. [Google Scholar] [CrossRef]
- DeLa Cruz, W.P.; Calvano, T.P.; Griffith, M.E.; White, C.E.; Kim, S.H.; Sutton, D.A.; Thompson, E.H.; Fu, J.; Wickes, B.L.; Guarro, J.; et al. Invasive Apophysomyces variabilis infection in a burn patient. J. Clin. Microbiol. 2012, 50, 2814–2817. [Google Scholar] [CrossRef]
- Lelievre, L.; Garcia-Hermosa, D.; Abdoul, H.; Hivelin, M.; Chouaki, T.; Toubas, D.; Mamez, A-C.; Lantieri, L.; Lortholary, O.; Lanternier, F. Posttraumatic mucormycosis. Medicine 2014, 93, 395-404.
- Narayanan, S.; Narayanan, C.D.; Kindon, A.J.; Arora, A.; Haridas, P.A. Fatal fungal infection: the living dead. J. Surg. Case Rep. 2014, 2014, rju104. [Google Scholar] [CrossRef]
- Shankarappa, V.K.G.; Shivappa, S.G.; Puttaswamy, M.C.; Ramakrishna, R.; Ramalingaiah, R.; Puranik, V. Apophysomyces variabilis a flesh-eating fungus – a case report. IOSR J. Dent. Med. Sci. 2014, 13, 86–90. [Google Scholar] [CrossRef]
- Coronel-Pérez, I.; Rodríguez-Rey, E.; Castilla-Guerra, L.; Dominguez, M. Primary cutaneous mucormycosis due to Saksenaea vasiformis in an immunocompetent patient. Actas Dermosifiliogr. 2015, 106, 516–518. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.Y.; Morales-López, S.E.; Rodríguez, G.J.; Álvarez-Moreno, C.A.; Ocampo, W.; Cepeda, M.L.; Mora-Valderrama, M.A. Necrotizing fasciitis caused by Apophysomyces variabilis in an immunocompetent patient. Med. Mycol. Case Rep. 2018, 20, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Samaddar, A.; Sharma, A.; Maurya, V.K.; Tak, V. Necrotizing fasciitis caused by Apophysomyces variabilis. IDCases 2019, 18, e00660. [Google Scholar] [CrossRef]
- Etienne, K.A.; Gillece, J.; Hilsabeck, R.; Schupp, J.M.; Colman, R.; Lockhart, S.R.; Gade, L.; Thompson, E.H.; Sutton, D.A.; Neblett-Fanfair, R.; et al. Whole genome sequence typing to investigate the Apophysomyces outbreak following a tornado in Joplin, Missouri, 2011. PLoS ONE 2012, 7, e49989. [Google Scholar] [CrossRef] [PubMed]
- Warketien, T.; Rodriguez, C.; Lloyd, B.; Wells, J.; Weintrob, A.; Dunne, J.R.; Ganesan, A.; Li, P.; Bradley, W.; Gaskins, L.J.; et al. Invasive mold infections following combat-related injuries. Clin. Infect. Dis. 2012, 55, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Austin, C.L.; Finley, P.J.; Mikkleson, D.R.; Tibbs, B. Mucormycosis: a rare fungal infection in tornado victims. J. Burn Care Res. 2014, 35, e164–e171. [Google Scholar] [CrossRef] [PubMed]
- Kronen, R.; Liang, S.Y.; Bochicchio, G.; Bochicchio, K.; Powderly, W.G.; Spec, A. Invasive fungal infections secondary to traumatic injury. Int. J. Infect. Dis. 2017, 62, 102–111. [Google Scholar] [CrossRef]
- Walsh, T.J.; Hospenthal, D.R.; Petraitis, V.; Kontoyiannis, D.P. Necrotizing mucormycosis of wounds following combat injuries, natural disasters, burns, and other trauma. J. Fungi 2019, 5, 57. [Google Scholar] [CrossRef]
- Echaiz, J.F.; Burnham, C-A.; Bailey, T.C. A case of Apophysomyces trapeziformis nectrotizing soft tissue infection. Int. J. Infect. Dis. 2013, 17, e1240-e1242.
- Bertumen, J.B.; Schell, W.A.; Joyce, M.; Alley, C.; Woods, C.W. Diagnostic difficulty identifying Apophysomyces trapeziformis septic arthritis in a patient with multiple myeloma. JMM Case Rep. 2016, 3, e005075. [Google Scholar] [CrossRef]
- Ellis, J.J.; Ajello, L. An unusual source for Apophysomyces elegans and a method for stimulating sporulation of Saksenaea vasiformis. Mycologia 74, 144-145.
- Padhye, A.A.; Ajello, L. Simple method of inducing sporulation by Apophysomyces elegans and Saksenaea vasiformis. J. Clin. Microbiol. 1988, 26, 1861–1863. [Google Scholar] [CrossRef]
- Chander, J.; Stchigel, A.M.; Alastruey-Izquierdo, A.; Jayant, M.; Bala, K.; Rani, H.; Handa, U.; Punia, R.S.; Dalal, U.; Attri, A.K.; et al. Fungal necrotizing fasciitis, an emerging infectious disease caused by Apophysomyces (Mucorales). Rev. Iberoam. Micol. 2015, 32, 93–98. [Google Scholar] [CrossRef]
- Labuda, R.; Bernreiter, A.; Hochenauer, D.; Schüller, C.; Kubátová, A.; Strauss, J.; Wagner, M. Saksenaeae dorisiae sp. nov., a new opportunistic pathogenic fungus from Europe. Int. J. Microbiol. 2019, 6253829.
- Planegger, A.; Uyulmaz, S.; Poskevicius, A.; Zbinden, A.; Muller, N.J.; Clacagni, M. Cutaneous invasive fungal infections with Saksenaea species in immunocompetent patients in Europe: a systematic review and case report. Plast. Reconstr. Surg. Glob. Open. 2022, 10, e4230. [Google Scholar] [CrossRef]
- Shao, J.; Wan, Z.; Li, R.; Yu, J. Species identification and delineation of pathogenic Mucorales by Matrix-Assisted Laser Desorption Ionisation-Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2018, 56, e01886–17. [Google Scholar] [CrossRef]
- Chander, J.; Singla, N.; Kaur, M.; Punia, R.P.S.; Attri, A.K.; Alastruey-Izquierdo, A.; Stchigel, A.; Cano-Lira, J.F.; Guarro, J. Apophysomyces variabilis, an emerging and worrisome cause of primary cutaneous necrotizing infections in India. J. Med. Mycol. 2021, 31, 101197. [Google Scholar] [CrossRef]
- Jain, D.; Kumar, Y.; Vasishta, R.K.; Rajesh, L.; Pattari, S.K.; Chakrabarti, A. Zygomycotic necrotizing fasciitis in immunocompetent patients: a series of 18. Mod. Pathol. 2006, 19, 1221–1226. [Google Scholar] [CrossRef]
- Thornton, C.R. The potential for rapid antigen testing for mucormycosis in the context of COVID-19. Expert Rev. Mol. Diagn. 2024, 3, 161–167. [Google Scholar] [CrossRef]
- Davies, G.E.; Thornton, C.R. Development of a monoclonal antibody and a serodiagnostic lateral-flow device specific to Rhizopus arrhizus (Syn. R. oryzae), the principal global agent of mucormycosis in humans. J. Fungi 2022, 8, 756. [Google Scholar]
- Thornton, C.R.; Davies, G.E.; Dougherty, L. Development of a monoclonal antibody and a lateral-flow device for the rapid detection of a Mucorales-specific biomarker. Front. Cell. Infect. Microbiol. 2023, 13, 1305662. [Google Scholar] [CrossRef] [PubMed]
- Hudson, A.C.; Corzo-Léon, D.E.; Kalinina, I.; Wilson, D.; Thornton, C.R.; Warris, A.; Ballou, E.R. Characterization of the spatiotemporal localization of a pan-Mucorales-specific antigen during germination and immunohistochemistry. J. Infect. Dis. 2025, 231, e244–e253. [Google Scholar] [CrossRef]
- Rousselot, J.; Millon, L.; Scherer, E.; Bourgeois, N.; Imbert, S.; Dupont, D.; Debourgogne, A.; Maubon, D.; Bellanger, A.P.; Thornton, C.R. Detection of Mucorales antigen in bronchoalveolar lavage samples using a newly developed lateral-flow device. J. Clin. Microbiol. 2025, 63, e00226–25. [Google Scholar] [CrossRef]
- Thornton, C.R. Immunological Methods for Fungi. In Molecular and Cellular Biology of Filamentous Fungi. In Molecular and Cellular Biology of Filamentous Fungi: A Practical Approach, 1st ed.; Talbot, N.J., Ed.; Oxford University Press: Oxford, UK, 2001; pp. 227–256. [Google Scholar]
- Mah, J.-E.; Yu, J.-H. Upstream and downstream regulation of asexual development in Aspergillus fumigatus. Euk. Cell 2006, 5, 1585–1595. [Google Scholar] [CrossRef]
- Thornton, C.R. Development of an immunochromatographic lateral-flow device for rapid serodiagnosis of invasive aspergillosis. Clin. Vaccine Immunol. 2008, 15, 1095–1105. [Google Scholar] [CrossRef]
- Thornton, C.R. Detection of the ‘big five’ mold killers of humans: Aspergillus, Fusarium, Lomentospora, Scedosporium and Mucormycetes. Adv. Appl. Microbiol. 2020, 110, 1–61. [Google Scholar]
- Roden, M.M.; Zaoutis, T.E.; Buchanan, W.I.; Knudsen, T.A.; Sarkisova, T.A.; Schaufer, R.L.; et al. Epidemiology and outcome of zygomycosis: a review of 929 reported cases. Clin. Infect. Dis. 2005, 41, 634–653. [Google Scholar] [CrossRef]
- Graves, B.; Morrissey, C.O.; Wei, A.; Coutsouvelis, J.; Ellis, S.; Pham, A.; Gooi, J.; Ananda-Rajah, M. Isavuconazole as salvage therapy for mucormycosis. Med. Mycol. Case Rep. 2016, 11, 36–39. [Google Scholar] [CrossRef]
- Pham, D.; Howard-Jones, A.R.; Sparks, R.; Stefani, M.; Sivalingham, V.; Halliday, C.L.; Beardsley, J.; Chen, C.-A. Epidemiology, modern diagnostics, and the management of Mucorales infections. J. Fungi 2023, 9, 659. [Google Scholar] [CrossRef] [PubMed]
- Caramalho, R.; Tyndall, J.D.A.; Monk, B.C.; Larentis, T.; Lass-Flörl, C.; Lackner, M. Intrinsic short-tailed azole resistance in mucormycetes is due to an evolutionary conserved aminoacid substitution of the lanosterol 14α-demethylase. Sci. Rep. 2017, 7, 15898. [Google Scholar] [CrossRef] [PubMed]
- Skiada, A.; Lanternier, F.; Groll, A.H.; Pagano, L.; Zimmerli, S.; Herbrecht, R.; Lortholary, O.; Petrikkos, G.L. Diagnosis and treatment of mucormycosis in patients with hematological malignancies: Guidelines from the 3rd European Conference on Infections in Leukemia (ECIL 3). Haematologica 2013, 98, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Alastruey-Izquierdo, A.; Castelli, M.V.; Cuesta, I.; Monzon, A.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Activity of Posaconazole and other antifungal agents against Mucorales strains identified by sequencing of internal transcribed spacers. Antimicrob. Agents Chemotherap. 2009, 53, 1686–1689. [Google Scholar] [CrossRef]
- Skiada, A.; Pavleas, I.; Drogari-Apiranthitou, M. Rare fungal infectious agents: a lurking enemy. F1000Res. 2017, 6, 1917. [Google Scholar] [CrossRef]
- Bonifaz, A.; Tirado-Sánchez, A.; Hernández-Medel, M.L.; Kassak, J.J.; Araiza, J.; González, G.M. Mucormycosis with cutaneous involvement. A retrospective study of 115 cases at a tertiary care hospital in Mexico. Australas. J. Dermatol. 2021, 62, 162–167. [Google Scholar] [CrossRef]
- Pedreira-Rincón, J.; Rivas, L.; Comenge, J.; Skouridou, V.; Camprubí-Ferrer, D.; Muñoz, J.; O’Sullivan, C.K.; Chamorro-Garcia, A.; Parolo, C. A comprehensive review of competitive lateral flow assays over the past decade. Lab Chip 2025, 25, 2578–2608. [Google Scholar] [CrossRef]
- Castrejón-Pérez, A.D.; Welsh, E.C.; Miranda, I.; Ocampo-Candiani, J.; Welsh, O. Cutaneous mucormycosis. An. Bras. Dermatol. 2017, 92, 304–311. [Google Scholar] [CrossRef]
- Murray, J.; Lu, Z.A.; Miller, K.; Meadows, A.; Totten, M.; Zhang, S.X. Disseminated aspergillosis and mucormycosis diagnosed at autopsy: a report of two cases of coinfection and a review of the literature. J. Fungi 2023, 9, 357. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Keighley, C.; Wolfe, R.; Lee, W.L.; Slavin, M.A.; Kong, D.C.M.; Chen, S.C.-A. The Epidemiology and clinical manifestations of Mucormycosis: A systematic review and meta-analysis of case reports. Clin. Microbiol. Infect. 2019, 25, 26–34. [Google Scholar] [CrossRef] [PubMed]
- William, A.; Kaur, R.; Rawat, D.; Kandir, N.S.S.; Sharma, A. Necrotizing fasciitis in neonate by Lichtheimia ramosa: a case study. Access Microbiol. 2022, 4, 000327. [Google Scholar] [CrossRef] [PubMed]
- Land, K.J.; Boeras, D.I.; Chen, X.-S. ; Ramsay, A.; Peeling, R.W. REASSURED diagnostics to inform disease control strategies, strengthen health systems and improve patient outcomes. Nat. Microbiol. 2019, 4, 46–54. [Google Scholar] [CrossRef]
- Millon, L.; Herbrecht, R.; Grenouillet, F.; Morio, F.; Alanio, A.; Letscher-Bru, V.; Cassaing, S.; Chouaki, T.; Kauffman-Lacroix, C.; Poirier, P.; et al. Early diagnosis and monitoring of mucormycosis by detection of circulating DNA in serum: retrospective analysis of 44 cases collected through the French Surveillence Network of Invasive Fungal Infections (RESSIF). Clin. Microbiol. Infect. 2016, 22, 810. [Google Scholar] [CrossRef]
- Shen, H.; Cai, X.; Liu, J.; Yan, G.; Ye, Y.; Ding, R.; Wu, J.; Li, L.; Shen, Q.; Ma, Y.; et al. Case report: the clinical utility of metagenomic next-generation sequencing in mucormycosis diagnosis caused by fatal Lichtheimia ramosa infection in pediatric neuroblastoma. Front. Pediatr. 2023, 11, 1130775. [Google Scholar] [CrossRef]
- Gomes, M.Z.R.; Lewis, R.E.; Kontoyiannis, D.P. Mucormycosis caused by unusual Mucormycetes, non-Rhizopus, -Mucor, and -Lichtheimia species. Clin. Microbiol. Rev. 2011, 24, 411–445. [Google Scholar] [CrossRef]
- Alvarez, E.; Stchigel, A.M.; Cano, J.; Sutton, D.A.; Fothergill, A.W.; Chander, J.; Salas, V.; Rinaldi, M.G.; Guarro, J. Molecular phylogenetic diversity of the emerging mucoralean fungus Apophysomyces: proposal of three new species. Rev. Iberoam. Micol. 2010, 27, 80–89. [Google Scholar] [CrossRef]
- Snell, B.; Tavakoli, K. Necrotizing fasciitis caused by Apophysomyces elegans complicating soft tissue and pelvic injuries in a tsunami survivor from Thailand. Plast. Reconst. Surg. 2007, 119, 448–449. [Google Scholar] [CrossRef]
- Bonifaz, A.; Stchigel, A.M.; Guarro, J.; Guevara, E.; Pintos, L.; Sanchis, M.; Cano-Lira, J.F. Primary cutaneous mucormycosis produced by the new species Apophysomyces mexicanus. J. Clin. Microbiol. 2014, 52, 4428–4431. [Google Scholar] [CrossRef]
- Miller, T.J.; Scheckter, C.C.; Watt, A.J. Humeral osteomyelitis with Apophysomyces ossiformis in an immunocompetent patient: a case report. Surg. Case Rep. 2019, 2, 3–3. [Google Scholar]
- Martínez-Herrera, E.; Frías-De-León, M.G.; Julián-Castrejón, A.; Cruz-Benítez, L.; Xicohtencatl-Cortes, J.; Hernández-Castro, R. Rhino-orbital mucormycosis due to Apophysomyces ossiformis in a patient with diabetes mellitus: a case report. BMC Infect. Dis. 2020, 20, 614. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, B.; Laínez-Arteaga, I.; Galindo-Morales, C.; Acevedo-Almendárez, L.; Aguilar, K.; Valladares, D.; López, M.; Fontecha, G. First molecular identification of three clinical isolates of fungi causing mucormycosis in Honduras. Infect. Dis. Rep. 2022, 14, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Wolkow, N.; Jakobiec, F.A.; Stagner, A.M.; Piantadosi, A.L.; Basgoz, N.; Lefebvre, D. Chronic orbital and calvarial fungal infection with Apophysomyces variabilis in an immunocompetent patient. Clin. Pathol. Rev. 2017, 62, P70–82. [Google Scholar] [CrossRef] [PubMed]
- Pamidimukkala, U.; Sudhaharan, S.; Kancharia, A.; Vemu, L.; Challa, S.; Karanam, S.D.; Chavali, P.; Prakash, H.; Ghosh, A.K.; Gupta, S.; et al. Mucormycosis due to Apophysomyces species complex – 25 years’ experience at a tertiary care hospital in southern India. Med. Mycol. 2020, 58, 425–433. [Google Scholar] [CrossRef]
- Hospental, D.R.; Chung, K.K.; Lairet, K.; Thompson, E.H.; Guarro, J.; Renz, E.M.; Sutton, D.A. Saksenaea erythrospora infection following combat trauma. J. Clin. Microbiol. 2011, 49, 3707–3709. [Google Scholar] [CrossRef]
- Relloso, S.; Romano, V.; Landaburu, M.F.; Herrera, F.; Smayevsky, J.; Veciño, C. , Mujica, M.T. Saksenaea erythrospora infection following a serious sailing accident. J. Med. Microbiol. 2014, 63, 317–321. [Google Scholar]
- Rodríguez, J.Y.; Rodríguez, G.J.; Morales-López, S.E.; Cantillo, C.E.; Le Pape, P.; Álvarez-Moreno, C.A. Saksenaea erythropora infection after medical tourism for esthetic breast augmentation surgery. Int. J. Infect. Dis. 2016, 49, 107–110. [Google Scholar] [CrossRef]
- Chander, J.; Singla, N.; Kaur, M. , Punia, R.S.; Attri, A.; Alastruey-Izquierdo, A.; Cano-Lira, J.F.; Stchigel, A.M.; Guarro, J. Saksenaea erythrospora, an emerging mucoralean fungus causing severe necrotizing skin and soft tissue infections – a study from a tertiary care hospital in north India. Infect. Dis. 2017, 49, 170–177. [Google Scholar]
- Tendolkar, U.; Baradkar, V.; Baveja, S.; Gore, M.; Bankar.; Gore, S. Cutaneous zygomycosis due to Saksenaea vasiformis in a patient with paraparesis, burns and pressure ulcer. Int. J. Inf. Dis. 2011, 10, 6.
- Neblett-Fanfair, R.; Benedict, K.; Bos, J.; Bennett, S.D.; Lo, Y-C.; Adebanjo, T.; Etienne, K.; Deak, E.; Derado, G.; Shieh, W-J.; et al. Necrotizing cutaneous mucormycosis after a tornado in Joplin, Missouri, in 2011. N. Engl. J. Med. 2012, 367, 2214-2225.
Figure 1.
Procedure for the detection of pan-Mucorales-specific antigen by lateral-flow immunoassay (LFIA). Step 1. Fungus is grown at 37 oC for 24 h on MEA and five discs of mycelium, prepared using a cork borer, transferred to YNB+G shake culture. The remaining MEA culture is then swabbed for 10 sec with a disposable sampling swab wetted with TG11-LFD running buffer (RB). Step 2. Soluble antigen is recovered from swab by rotation for 10 sec in 250 μL RB contained in a 1.5 mL micro-centrifuge tube. Step 3. The tube is centrifuged for 5 min at 17, 000 x g and the supernatant removed for testing by TG11-LFD. Step 4. One hundred μL of supernatant is added to the sample pad of the LFD and, after 30 min, the intensity of the test (T) and internal control (C) line determined as artificial units (a.u.) using a Cube reader [42,43]. Representative Test Results show complete loss of T lines for swab samples from Av (A. variabilis CBS658.93), Sv (Saksenaea vasiformis CBS113.96), and Lr (L. ramosa DRH226533346), while there is no loss of the T line for Af (Aspergillus fumigatus, strain ΔAfbrlA7). The negative control shown here consists of RB only, while the positive control (Lc) consists of purified EPS from Lichtheimia corymbifera CBS109940 at a concentration of 100 μg/mL RB. The corresponding Cube reader a.u. values for T lines are shown in Table 2.
Figure 1.
Procedure for the detection of pan-Mucorales-specific antigen by lateral-flow immunoassay (LFIA). Step 1. Fungus is grown at 37 oC for 24 h on MEA and five discs of mycelium, prepared using a cork borer, transferred to YNB+G shake culture. The remaining MEA culture is then swabbed for 10 sec with a disposable sampling swab wetted with TG11-LFD running buffer (RB). Step 2. Soluble antigen is recovered from swab by rotation for 10 sec in 250 μL RB contained in a 1.5 mL micro-centrifuge tube. Step 3. The tube is centrifuged for 5 min at 17, 000 x g and the supernatant removed for testing by TG11-LFD. Step 4. One hundred μL of supernatant is added to the sample pad of the LFD and, after 30 min, the intensity of the test (T) and internal control (C) line determined as artificial units (a.u.) using a Cube reader [42,43]. Representative Test Results show complete loss of T lines for swab samples from Av (A. variabilis CBS658.93), Sv (Saksenaea vasiformis CBS113.96), and Lr (L. ramosa DRH226533346), while there is no loss of the T line for Af (Aspergillus fumigatus, strain ΔAfbrlA7). The negative control shown here consists of RB only, while the positive control (Lc) consists of purified EPS from Lichtheimia corymbifera CBS109940 at a concentration of 100 μg/mL RB. The corresponding Cube reader a.u. values for T lines are shown in Table 2.
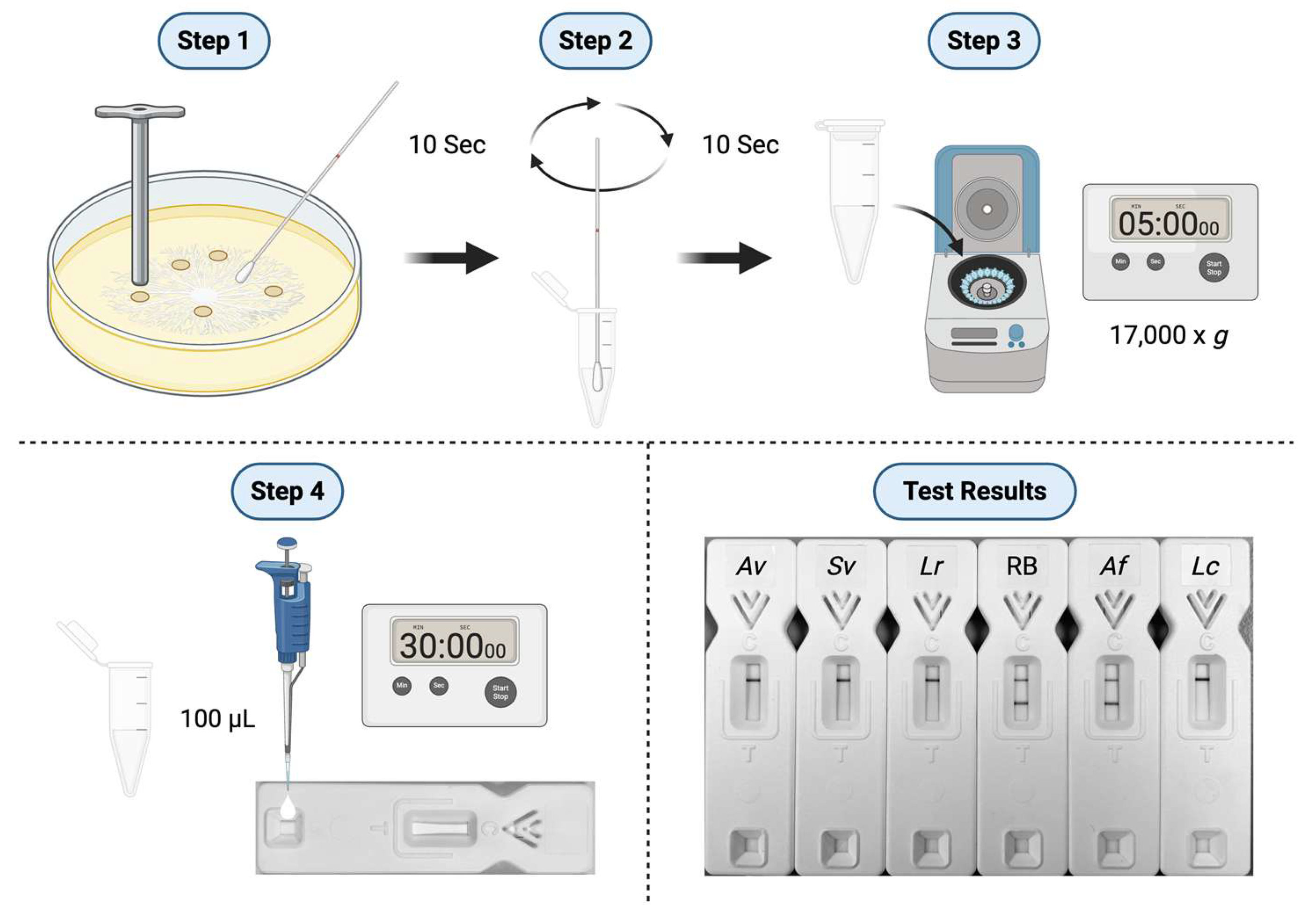
Figure 4.
In vitro production of the JD4 antigen by species of Apophysomyces, Saksenaea and Aspergillus; Ae (Apophysomyces elegans), Am (Apophysomyces mexicanus), Ao (Apophysomyces ossiformis), Av (Apophysomyces variabilis), Af (Aspergillus fumigatus, strain ΔAfbrlA7), Sd (Saksenaea dorisiae), Se (Saksenaea erythrospora), Sl (Saksenaea loutrophoriformis), Sv (Saksenaea vasiformis). (A) Dry weights of hyphal microcolonies after 24 h growth in YNB+G shake cultures. (B) Direct-ELISA of protein precipitates using the Apophysomyces-specific mAb JD4. The threshold ELISA absorbance value for immunoassay positivity is ≥0.2, with values above this threshold positive for detection of Apophysomyces antigen. (C) Direct-ELISA of protein precipitates using the Aspergillus-specific mAb JF5 [49]. The threshold ELISA absorbance value for immunoassay positivity is ≥0.2, with values above this threshold positive for detection of Aspergillus antigen. Each bar is the mean of three replicates ±SE (A,B,C).
Figure 4.
In vitro production of the JD4 antigen by species of Apophysomyces, Saksenaea and Aspergillus; Ae (Apophysomyces elegans), Am (Apophysomyces mexicanus), Ao (Apophysomyces ossiformis), Av (Apophysomyces variabilis), Af (Aspergillus fumigatus, strain ΔAfbrlA7), Sd (Saksenaea dorisiae), Se (Saksenaea erythrospora), Sl (Saksenaea loutrophoriformis), Sv (Saksenaea vasiformis). (A) Dry weights of hyphal microcolonies after 24 h growth in YNB+G shake cultures. (B) Direct-ELISA of protein precipitates using the Apophysomyces-specific mAb JD4. The threshold ELISA absorbance value for immunoassay positivity is ≥0.2, with values above this threshold positive for detection of Apophysomyces antigen. (C) Direct-ELISA of protein precipitates using the Aspergillus-specific mAb JF5 [49]. The threshold ELISA absorbance value for immunoassay positivity is ≥0.2, with values above this threshold positive for detection of Aspergillus antigen. Each bar is the mean of three replicates ±SE (A,B,C).
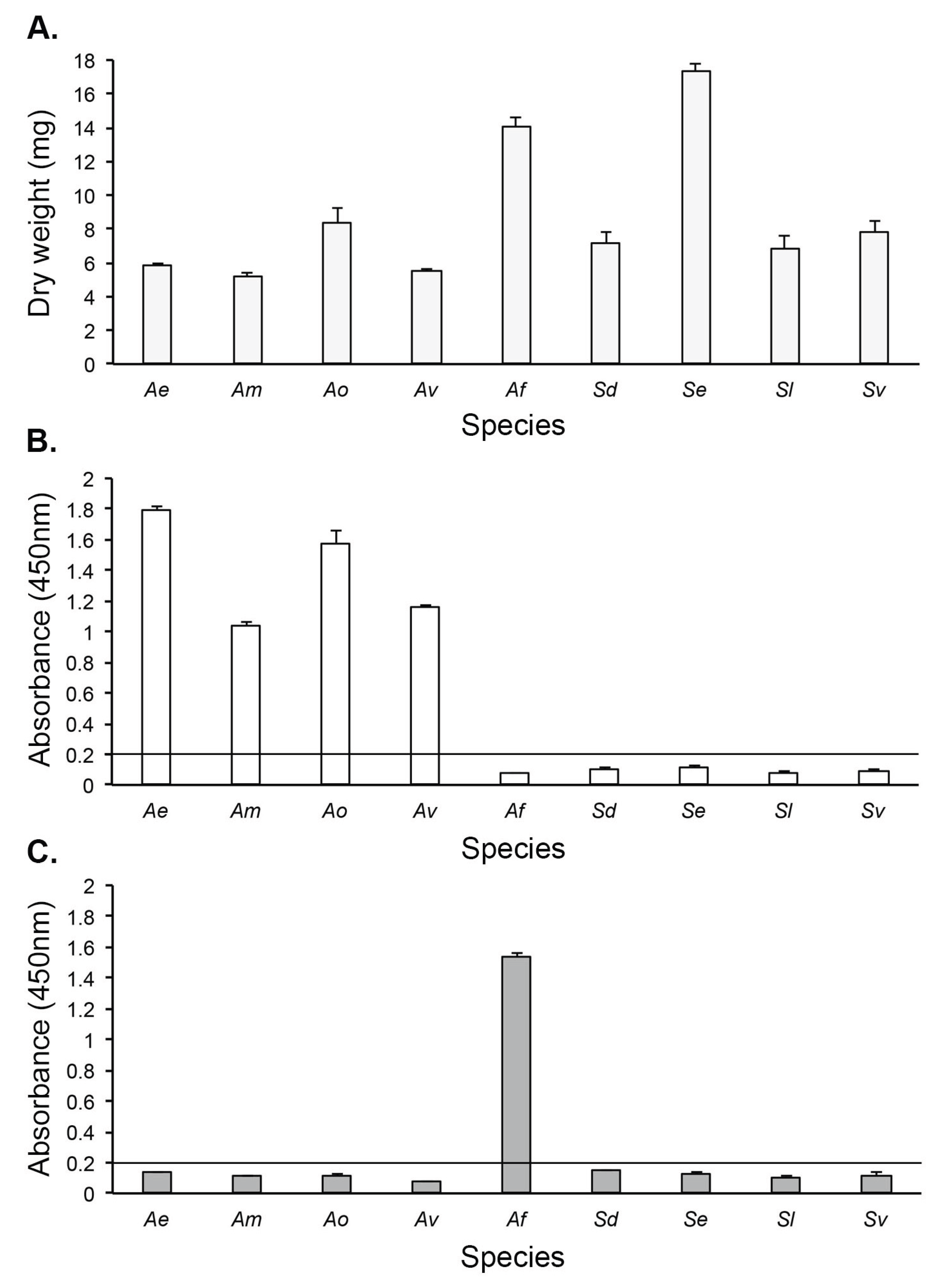

Table 1.
Details of human pathogenic fungi used to determine the specificity of mAb JD4, and results of Direct-ELISA tests against purified EPS antigens.
Table 1.
Details of human pathogenic fungi used to determine the specificity of mAb JD4, and results of Direct-ELISA tests against purified EPS antigens.
| Species | Isolate Number | Source1 |
JD4 Direct-ELISA Absorbance (450nm)2 |
|||
| Apophysomyces elegans | 477.78 | CBS | 1.466±0.005 | |||
| Apophysomyces mexicanus | 136361 | CBS | 1 1.224±0.006 | |||
| Apophysomyces ossiformis | 125533 | CBS | 0.674±0.018 | |||
| Apophysomyces variabilis | 658.93 | CBS | 1.646±0.022 | |||
| Aspergillus fumigatus | Af293 | FGSC | 0.082±0.003 | |||
| Cunninghamella bertholletiae | 151.80 | CBS | 0.084±0.011 | |||
| Fusarium solani | 224.34 | CBS | 0.049±0.000 | |||
| Lichtheimia corymbifera | 109940 | CBS | 0.093±0.002 | |||
| Lichtheimia ornata | 142195 | CBS | 0.083±0.000 | |||
| Lichtheimia hyalospora | 146576 | CBS | 0.073±0.002 | |||
| Lichtheimia ramosa | 112528 | CBS | 0.059±0.003 | |||
| Lichtheimia ramosa | 124197 | CBS | 0.071±0.005 | |||
| Lichtheimia ramosa | 2845 | NCPF | 0.067±0.002 | |||
| Lomentospora prolificans | 3.1 | CRT | 0.056±0.001 | |||
| Mucor circinelloides | B-52 | CRT | 0.050±0.001 | |||
| Mucor racemosus f. racemosus | 222.81 | CBS | 0.076±0.003 | |||
| Rhizomucor pusillus | 120587 | CBS | 0.073±0.002 | |||
| Rhizopus arrhizus | 111233 | CBS | 0.050±0.001 | |||
| Rhizopus arrhizus var. arrhizus | 112.07 | CBS | 0.044±0.002 | |||
| Rhizopus microsporus var. rhizopodiformis | 102277 | CBS | 0.039±0.005 | |||
| Saksenaea dorisiae | 146728 | CBS | 0.044±0.001 | |||
| Saksenaea erythrospora | 138279 | CBS | 0.044±0.002 | |||
| Saksenaea loutrophoriformis | 143036 | CBS | 0.045±0.006 | |||
| Saksenaea vasiformis | 113.96 | CBS | 0.056±0.001 | |||
| Scedosporium apiospermum | 8353 | CBS | 0.045±0.001 | |||
| Scedosporium aurantiacum | 121926 | CBS | 0.062±0.003 | |||
| Scedosporium boydii | 835.96 | CBS | 0.057±0.003 | |||
1CBS; Westerdijk Fungal Biodiversity Institute, The Netherlands. CRT; C.R. Thornton; FGSC; Fungal Genetics Stock Center, Kansas City University, USA. 2For Direct-ELISA tests with mAb JD4, the absorbance values are the means of two replicates ±SE, and the threshold absorbance value for immunoassay positivity is ≥0.100.
Table 2.
Details of human pathogenic fungi, and results of TG11-LFD tests using swabs from fungal cultures grown for 24 h on MEA medium.
Table 2.
Details of human pathogenic fungi, and results of TG11-LFD tests using swabs from fungal cultures grown for 24 h on MEA medium.
| Species | Isolate Number | Source1 |
TG11-LFD (a.u.)2 |
| Apophysomyces elegans | 477.78 | CBS | 34.5±1.5 |
| Apophysomyces mexicanus | 136361 | CBS | 27.8±0.1 |
| Apophysomyces ossiformis | 125533 | CBS | 30.8±0.3 |
| Apophysomyces variabilis | 658.93 | CBS | 15.5±4.5 |
| Aspergillus fumigatus ΔAfbrlA7 | A1176 | FGSC | 474.0±19.0 |
| Lichtheimia ramosa3 | 226533346 | DRH | 15.7±1.5 |
| Saksenaea dorisiae | 146728 | CBS | 38.9±0.5 |
| Saksenaea erythrospora | 138279 | CBS | 34.4±1.7 |
| Saksenaea loutrophoriformis | 143036 | CBS | 21.0±2.8 |
| Saksenaea vasiformis | 113.96 | CBS | 22.5±1.0 |
| Negative Control4 | Af293 | FGSC | 591.7±79.9 |
| Positive Control5 | 109940 | CBS | 29.5±2.2 |
| RB only | - | - | 506.9±20.8 |
1CBS; Westerdijk Fungal Biodiversity Institute, The Netherlands. CRT; C.R. Thornton. FGSC; Fungal Genetics Stock Center, Kansas City University, USA. 2For TG11-LFD tests, the T line intensity measured in artificial units (a.u.) is the mean of two replicate samples ±SE, and the threshold a.u. value for test positivity is ≤400 [43]. 3Isolate cultured from hand of patient with cutaneous mucormycosis. 4Negative control comprising purified EPS from A. fumigatus strain Af293 at 100 μg/mL running buffer (RB). 5Positive control comprising purified EPS from L. corymbifera strain CBS109940 at 100 μg/mL RB. All tests had internal control (C) line intensities of ≥400 a.u..
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



