Submitted:
09 September 2025
Posted:
09 September 2025
You are already at the latest version
Abstract
There is an urgent need for new alternative compounds with distinct modes of action due to the global rise in antibiotic resistance and the associated risks to public health. It is currently established that between 40 and 80% of bacterial biofilms cause antibiotic resistance. Furthermore, biofilm-forming bacteria are 1000 times more resistant to antibiotics than in their planktonic stages. Recently, the number of papers published in 2023 to find antibiofilm compounds from fungi has increased. Meanwhile, it has been proven that endophytic fungi can produce undiscovered compounds against bacterial biofilm. However, as shown in this review, there is still not enough attention given to highlight the relevance of intensifying studies amongst marine-derived fungi. This review summarizes the biologically active compounds isolated from marine-derived fungal extracts tested against bacterial biofilms published from 2015 to 2024. Moreover, this review discloses evidence on the scarcity of research on antibiofilm compounds from algal endophytic fungi. In addition, the primary approaches used in the hunt for bioactive secondary metabolites are covered in this review. Besides that, a few recent strategies have been mentioned to optimize the production of antibiofilm-active fungal metabolites by employing such techniques such as through media optimization, use of chemical elicitors, co-culture, and metabolic engineering.
Keywords:
Biofilm
; endophytic fungi
; metabolites
; anti-biofilm
; seaweeds
; elicitors
; co-culture
1. Introduction
The discovery of antibiotics accelerated after the penicillin breakthrough by Fleming. However, incorrect use and lack of monitoring by humans cause ineffectiveness or resistance to the antibiotic. The reason may also be due to the bacteria themselves, as they adapt to large doses due to their rapid reproduction, as some bacteria reproduce in less than 20 hours [1]. Recently, antibiotic resistance has emerged as a significant global health issue, and the World Health Organization (WHO) listed it as one of the top ten risks to public health worldwide in 2019 [2]. Moreover, the third disease that causes death in the world is untreated bacterial infection [1]. The global rise of antibiotic resistance will put up to 10 million people at risk yearly by 2050 [3,4]. In addition, multidrug-resistant bacterial infection is causing a burden cost amounting to more than 4.6 billion USD spent on the treatment of infectious diseases in 2017 in the United States alone, which has been only increasing since then [5]. As a result, these drive the need to identify the causes of antibiotic resistance and detect novel compounds for the antibiotic pipeline that effectively combat infections. There are various reasons behind antibiotic resistance, and the mechanisms are quite complex. In terms of molecular mechanisms, antibiotic resistance has been exhibited in the most commonly used types of antibiotics that includes: 1) decreased permeability in enterococcal bacterial resistance against low concentrations of aminoglycosides, 2) increased efflux pump in tetracycline resistance, 3) alteration of the antibiotic target as methicillin resistance, and 4) antibiotic hydrolysis by bacterial enzyme B-lactamase in penicillin and cephalosporin resistance [6,7,8,9]. Nowadays, bacterial biofilms are the cause of over 80% of bacterial illnesses, and about 40%-80% of bacterial biofilms lead to antibiotic resistance [10]. Bacterial biofilms have the potential to develop resistance and tolerance to antibiotics ranging from 10 to 1000 times more than planktonic bacteria [11]. Although bacterial biofilms have a positive effect as biological control agents against plant pathogens, it has been proven that bacterial biofilms had damaged human health, food safety, and the food industry [12,13,14]. Therefore, this mini review aims to focus on providing literature evidence from 2015 to 2024 on promising approaches and sources for the discovery of antibiofilm compounds that may help the antibiotic resistance crisis.
2. Materials and Methods
Using a certain phrase as the main search term in each year for ten years, a methodical search was carried out on Google Scholar to find relevant papers. Conference abstracts and review papers were not included in the search, which was limited to English-language publications. After initial retrieval, papers that did not contain the search keyword at all or only mentioned it in the conclusion or discussion sections were carefully eliminated manually. Furthermore, studies that did not specifically use the term "endophytic" were still included if the methodology explained techniques that were compatible with isolating endophytic or marine-derived/associated fungi (e.g., surface sterilization of plant or marine host tissues using 0.01% sodium hypochlorite to remove epiphytic microorganisms) as described in Table 1.
This search strategy was limited by several factors. The first search was conducted using only one term and no Boolean operators or alternative keywords, which might have limited the amount of material that could be found. Additionally, only Google Scholar database was searched; relevant research that was indexed elsewhere might have been missing because other databases were not included. This systematic search method was employed for literature quantification, but the compilation of fungal-derived chemicals in Table 2 depended on a more expansive and less standardized literature review, without uniform search keywords, exclusion criteria, or database limitations.
3. Bacterial Biofilm
A bacterial biofilm occurs when several bacteria aggregate on a biotic/abiotic surface and excrete extracellular polymeric substances (EPS) such as sugars, proteins, extracellular DNA, and water [15] . The stages of bacterial biofilm formation as shown in Figure 1, begin with planktonic bacterial cells when they adhere to a biotic or abiotic surface. When the bacteria aggregate together, they become sessile bacterial cells which can move away from the adhesion on the surface and return to their reversible planktonic state. After they form sessile bacteria, EPS is secreted which completes the fixed attachment of the bacteria, contributing to their being in an irreversible state. After that, the bacteria form colonies, which in turn secrete huge amounts of EPS, which act as a barrier to protect these colonies. In their maturity stage, the bacterial colonies are shaped like mushrooms, and they form a three-dimensional biofilm structure [16]. The shape of the bacterial biofilm allows the exchange of nutrients/water and genes between bacteria of same and different species [17]. The gene expression patterns of sessile cells in a bacterial biofilm differ significantly from those found in planktonic cells as in Pseudomonas aeruginosa (P. aeruginosa) colony [12]. Finally, these mature bacterial biofilms are destroyed, releasing microorganisms that may infect other parts of the host organism or further spread on their inert surface and form new biofilms [10]. In addition, dispersion of mature bacterial biofilms can be caused by their exposure to nitric oxide (NO) [18]. Bacteria communicate to each other or to other microorganisms, whether between same or different species in the biofilm matrix by a quorum sensing system (QS). This occurs through secreting an autoinducer (AI) in response to the density of bacterial cells. QS regulates gene expression to control their pathogenicity and allows microbes to produce virulent factors [12,19,20]. QS has been linked to a variety of bacterial functions, including motility, virulence, bioluminescence, pigment disposition, biofilm formation, and polysaccharide production [21]. Besides, EPS have a major impact on the development of these aggregations, the rate of gene exchange, and antimicrobial resistance [22]. There are various proposed reasons for bacterial biofilms to become resistant to antibiotics. Since bacterial biofilm entails EPS that is negatively charged, it is expected that positively charged antibiotics will interact with it, which may either reduce or prevent their bioactivity. Another point of view is restricting the diffusion of antibiotics by EPS and therefore, decreasing the absorption of antibiotics to inhibit the growth rate of bacteria within biofilms [17,23]. This antibiotic resistance was further demonstrated by Acinetobacter baumannii (A. baumannii) and Staphylococcus biofilm where the EPS matrix was shown to limit the diffusion of antibiotics while just keeping the antibiotic near the surface [24]. Generally, penetration of beta-lactams and glycopeptides is usually less than that of other antibiotics [25]. Additionally, bacteria within biofilms rapidly adapt to external changes through altered metabolism and gene expression, which may contribute to changes in target cells or hiding target sites and thus resulting to antibiotic resistance [13,17]. Meanwhile, new resistant pathogenic strains are currently being discovered in increasing frequency, demanding the need for a new pipeline of potentially beneficial compounds [19]. As the formation of a bacterial biofilm aggravated the problem of antibiotic resistance, this encourages the discovery of new antibiofilm compounds.
In the context of treatment strategies against bacterial biofilm formation or disruption, as was already mentioned above, QS contributes to the development of biofilms. Since QS are not necessary for bacterial growth, QS inhibitors (QSIs) cause less evolutionary pressure on the bacteria and thus, do not cause the development of resistance compared to the more common bactericidal or bacteriostatic mechanisms of antibiotic activity [21]. It seems that one beneficial method of controlling bacterial biofilm is employing QSIs to reduce or suppress biofilm formation as a treatment strategy [10]. QSIs have shown to increase the susceptibility of bacterial biofilms to antibiotics and thus, contribute to the success of the antibiotic treatment [26]. One study showed significant synergy effects in preventing biofilm in a rat model induced with Staphylococcal vascular graft infection when combining daptomycin antibiotic with a QS inhibitor [27]. Moreover, NO can also be used to disrupt biofilms, and its use in combination to existing antibiotics will likely increase their effectiveness against bacterial biofilms [18,28]. A published article showed that exposure to sodium nitroprusside as NO donor markedly improved the ability of some antimicrobial compounds such as hydrogen peroxide, sodium dodecyl sulphate, and tobramycin to effectively eliminate P. aeruginosa biofilms [29]. Given that most biofilm-dispersing drugs, do not kill bacterial cells, it is advantageous to combine them with an antibacterial agent [22]. Therefore, combination of novel anti-biofilms agent with biofilm-dispersing drugs as NO analogue or QSIs or conventional antibiotics could eradicate bacterial biofilm to treat microbial infections and help in drug resistance crises.
4. Natural Products for the Antibiotic Pipeline
Natural products (NPs) offer an appealing reservoir of chemicals with novel anti-microbial compounds. NPs have been a strong source of antibacterial compounds since ancient times, accounting for 78% of our current drugs in the market approved between 1983 and 1994 [30]. NPs afforded structural complexity and diverse chemical composition that make them important in the search for new drugs, especially for the treatment of new emerging infectious and cancer diseases [31,32]. These variations had also lead to differences in the QS inhibitory action of these compounds [19]. In addition, NPs are less prone to develop resistance against bacteria [5]. NPs are secondary metabolites that work as defensive molecules against existing pathogens [33]. In connection with anti-bacterial activity, NPs have been found effective against many pathogenic microbes, but they are distinct from synthetic antibiotics in that they have diverse modes of action against microbes [34,35]. This feature could help against bacterial resistance to antibiotics. As some NPs, most of which are phytochemicals or plant-derived such as 27 flavonoid, 23 alkaloids and 17 terpenes have been proven effective against MRSA (methicillin-resistant Staphylococcus aureus) by destroying the bacterial membrane and blocking the efflux pump [36]. In addition, plant-derived compounds have successfully inhibited the formation of new biofilms for Staphylococcus aureus (S. aureus) as well as Staphylococcus epidermidis (S. epidermidis) [37]. On the other hand, because oceans covering the earth contain 500,000 living species, it is beneficial to discover novel bioactive compounds from it, which could help challenge antibiotic resistance [38].
Marine-derived fungi have also shown to play a role in inhibiting S. aureus biofilm. Two compounds, secalonic acids B (70) and D (71) inhibited bacterial biofilms by more than 90% at 6.25 micrograms/mL without inhibiting cell growth [39]. Therefore, there is some evidence that support metabolites derived from the marine fungi may possess anti-quorum sensing and/or antibiofilm properties. For example, the marine fungus Penicillium chrysogenum DXY-1 afforded cyclo(L-Pro-L-Tyr) (68), a cyclic dipeptide or diketopiperazine that has been shown to inhibit biofilm formation and lowers QS gene expression in P. aeruginosa PA01 [40]. In addition, the fungal strain Blastobotrys parvus PPR3, which was isolated from the woods of the mangrove plant Avicennia marina, exhibited anti-QS activity and antibiofilm effects against P. aeruginosa PAO1 [41]. Meanwhile, there are huge untapped reservoirs for the isolation of new marine microbes that have a high potential to produce bioactive secondary metabolites [42]. From that point on, marine fungi are full of secondary bioactive metabolites, and the microorganisms derived from seaweeds are promising sources for the discovery of antibiofilm compounds and QSIs.
Seaweeds for instance are affected by changes in season, harvest time, geographic location, and ecological variables, which increases the probability of finding novel bioactive compounds [43]. Furthermore, seaweeds have been characterized by their chemical diversity that represents an opportunity to attract compounds with distinctive properties and uses with a wide range of biological activities [44]. The first record of antimicrobial activity from seaweed extracts was published in 1944 by Pratt et al. [45]. Moreover, the three extracts of brown, green, and red seaweeds have exhibited antibacterial activity as well [46,47]. In terms of antibiofilm activity, studies have shown the occurrence of compounds derived from seaweeds to possess antibiofilm activity against pathogenic microorganisms [48]. Fucoidan (sulfated polysaccharide), derived from the brown algae, Fucus vesiculosus, afforded antibiofilm activity against Streptococcus mutans and sobrinus [49]. Furthermore, phlorotannins derived from another brown seaweed, Hizikia fusiforme, were found to inhibit biofilm formation in P. aeruginosa as well as inhibit QS activity of the reporter strain Chromobacterium violaceum [50]. Nevertheless, some of the earlier reported bioactive compounds derived from algae are now attributed to their associated microorganisms [51]. In line with this, available reports have confirmed that these seaweed-associated microorganisms consisted of bacteria, fungi, and yeast [52,53,54]. Some of these microbes have antimicrobial properties as well as against nosocomial pathogens [55].
5. Seaweed-Derived Microorganisms
Seaweed-associated microorganisms have drawn a lot of attention lately as a potential new supply of secure and potent bioactive. It is worth mentioning that most macroalgae have a rich diversity of epiphytic and endophytic microorganisms [56]. Epiphytic microbes are the living microbes found on the surface of seaweed, and they are important in biogeochemical cycles [57,58,59]. Furthermore, epiphytic bacteria are the most studied microbes in algae due to their abundance and the convenience of isolating them from thinner fronds [60]. Epiphytic fungi associated seaweeds have shown to have antibacterial activity [61]. Regarding their antibiotic activity, 38 epiphytic bacterial strains from 5 algal species were found to be exhibit antibacterial activity [62]. On the other hand, when microorganisms colonize the internal tissues of the seaweeds without causing any infection to the host, they are called endophytic microorganisms [57]. They are primarily found in natural ecosystems that invade the intercellular spaces of the host organism and could modify the primary and secondary metabolism of the host, which affects several aspects of responses by the host organism [63,64]. The effectiveness of seaweed-associated endophytic bacteria against pathogenic microbes has been reported [65]. Furthermore, multiple studies have also proven the effectiveness of algal endophytic fungi against bacterial pathogens [66,67,68,69,70]. Recently most of the secondary biologically active metabolites derived from microbes are of fungal origin [71]. Generally, four categories of endophytic fungi were identified: Ascomycota, Basidiomycota, Zygomycota, and Oomycota [72]. The function of endophytic fungal metabolites is to strengthen the plant immune system, combat pathogen invasions, and lessen biotic and abiotic stresses [73]. Besides that, the endophytic fungi derived from seaweeds are thought to be a talented source for the synthesis of novel secondary metabolites with antibacterial activity [74].
5.1. Antibacterial Metabolites from Seaweed Endophytes and Other Marine-Derived Fungi
Secondary metabolites of endophytic and other host-associated fungi play a role in the discovery of new compounds, especially in the search for new antibacterials. Generally, the types of bioactive compounds found in abundance in endophytic fungi from seaweeds included lactones, peptides, polyketides, alkaloids, and terpenoids [75]. As examples of polyketides metabolites that have antibacterial activity include phenalenone and diketo-lactone ring derivatives derived from endophytic fungi (Coneutherium grains) associated with the green algae Enteromorpha sp [76]. Similarly, one of the five polyketide compounds obtained from the endophytic fungus (Paraconiothyrium sp.) derived from red seaweed (Chondrus ocellatus holmes) exhibited a moderate level of antibacterial activity [77]. The extracted an endophytic fungus derived from the green seaweed, Ulva sp., was capable of producing an antibacterial metabolite called ascosalipyrrolidinone, an alkaloid metabolite [78]. Likewise, the sesquiterpenoid metabolite albican-11,14-diol, isolated from the A. versicolor endophyte derived from Codium fragilis, showed antibacterial activity [79]. There are some examples of lactone metabolites that have antibacterial activity such as: 5-hydroxy-de-O-methyllasiodiplodin and de-O-methyllasiodiplodin. Subsequently, Fusarium oxysporum, Bacillus subtilis (B. subtilis), and S. aureus cannot grow in the presence of lasiodiplodin metabolites from the endophytic fungus associated with the brown algae, Sargassum sp. [80]. Therefore, lately there are many kinds of metabolites of algal endophytic fungi that have an antibacterial effect that makes researchers turn to them as a source of new antibiofilm compounds.
On the other hand, from marine invertebrates, which are a more common source of -derived fungi, three new peptides known as asperversiamides, were isolated from the fungus A. versicolor, which was derived from a gorgonian soft coral. These substances demonstrated potent inhibitory action against Mycobacterium marinum [75]. Additionally, Penicillium brocae, an endophytic fungus derived from the marine mangrove plant Avicennia marina provided four new sulphide diketopiperazine derivatives showed antimicrobial activity [64]. Although, 47% of the peptides isolated from marine fungi have no biological activity and about 53% of them have cytotoxic effects, consistently needs intensive evaluation [81]. One of the bioactivities of the cyclic dipeptide, cis-cyclo (Leucyl-Tyrosyl) derived from a marine sponge-associated Penicillium sp has remarkable ability to inhibit up to 85% of the formation of biofilms from S. epidermidis [82].
5.2. Antibiofilm Compounds from Marine-Derived, Endophytic and Other Fungi
Figure 2 shows a paucity of studies directly pertaining to ‘Seaweed endophytic fungi antibiofilm compounds’, with only two identified from 1,010 articles between 2015 and 2024 [83,84]. Conversely, the more general phrase resulted in 12 articles throughout the same timeframe, suggesting insufficient focused research on this domain [41,83,85,86,87,88,89,90,91,92,93]. All these articles discussed in depth below. Nonetheless, this shortage indicates that although general characteristics of antibiofilm compounds from marine endophytic fungus have been investigated, the specialized area of antibiofilm compounds produced from seaweed remains insufficiently examined. Addressing this gap is crucial for advancing algal endophytic fungi, since it directly influences the discovery of novel antibiotics.
Fungi represent a source of antibiofilm-active secondary metabolites for a promising drug pipeline against bacterial biofilms. In this section, we are considering other sources of antibiofilm fungal metabolites because a very small percentage of these compounds have been described in comparison to the larger number of described antibacterial compounds [94]. The classification of bacterial strains identified as weak, moderate, and strong biofilm producers, include Escherichia coli (E. coli), S. aureus, B. subtilis, and P. aeruginosa, respectively [95]. In this review, we will focus on the formation of robust bacterial biofilm. There are two phenotypic types of P. aeruginosa biofilm: mucoid and non-mucoid. The mucoid variant is more challenging to eliminate compared to non-mucoid biofilms [96]. Nonetheless, the mucoid strains of P. aeruginosa exhibit greater sensitivity to antibiotics compared to non-mucoid isolates [97]. The non-mucoid strains of P. aeruginosa, including PAO1 and ATCC 27853, are commonly the focus of investigation [98]. There are numerous antibiofilm activity of the extracts of fungi against bacteria have been published.
Dendryphiella salina (D. salina) a fungi derived from the seaweed Laminaria hyperborea displayed 100 % inhibition activity against P. aeruginosa (ATCC 27853) biofilm at 100 μg/mL [84]. Differences in antibacterial and antibiofilm bioactivity have been described in literature depending on the type of fungus, media used for cultivation, and the extraction solvent employed. As in the investigation of non-marine fungal extracts, Attia EZ and co-workers used several media, also known as OSMAC approach (One-Strain-Many-Compounds) and got different percentages of biofilm inhibition, however, the modified potato dextrose broth (MPDB) medium gave the highest percentage of inhibition [99]. Whereas, the potato dextrose agar (PDA) media effects positively on the production of secondary metabolites [100,101]. Indeed, the biosynthesis of secondary metabolites is significantly influenced by the composition of the media [102]. Similarly, Jaber [103] also did an OSMAC approach with malt extract with and without sea salt, Wickersham media with and without sea salt, marine broth, rice media with and without sea salt, and oat with and without sea salt. For the scale-up, the malt extract broth with sea salt was also chosen to grow D. salina for 30 days as it provided the most potent antimicrobial activity and afforded a different chemical profile to that of D. salina grown on oat media with MIC of 20.0 and 17.5 µg/ml and MBEC (Minimum Biofilm Eradication Concentration) of 21.8 and 18.8 µg/ml against both S. aureus and P. aeruginosa, respectively. The optimum growth of D. salina with the sea salt requirement was evident that the fungus is a “true” marine algal endophyte.
Furthermore, the solvent employed for extraction will influence the activity as well. The ethyl acetate (EtOAc) extract had superior antibiofilm activity compared to both methanol and acetone extracts, achieving complete biofilm formation inhibition (100%) against non-mucoid strains of P. aeruginosa, whereas the acetone and methanol extracts exhibited lower activities of 40% and 50%, respectively [104,105,106]. Furthermore, the EtOAc extracts derived from the endophytic fungus Neocosmospora sp. MFLUCC 17-0253, obtained from the mangrove plant Rhizophora apiculata, demonstrated inhibition of biofilm formation in Acidovorax citrulli JT-0003 at 44 to 77%, when applied at concentrations ranging from 12.5 to 100 µg/mL [87]. Similarly, EtOAc extracts demonstrated higher potency of antibiofilm activity compared to their chloroform extracts, as evidenced by its greater percentage of biofilm inhibition against the tested S. aureus as demonstrated by several studies [91,107,108,109].
In terms of the activity of fungal crude extracts and their purified metabolites, the crude extract of Alternaria alternata has slightly increased its antibiofilm activity against P. aeruginosa than their purified metabolites [90,110]. A study likewise demonstrated that a fungal extract that afforded weak biofilm inhibition against P. aeruginosa at 36%, totally loss its bioactivity when the respective metabolites were purified [111]. The crude extract could have contained several metabolites that when their modes of action were combined would lead to the extract’s bioactivity causing a synergistic effect as interpreted by Njateng and his colleagues [102,112]. Alternatively, another study showed that the crude extract of a marine-derived Penicillium sp. inhibited biofilm formation in S. aureus at only 19% while their purified metabolites, particularly β-sitosterol (77), exhibited antibiofilm activity at 64% [113]. In this case, bioactivity required specificity and concentration dependent on a single metabolite. It is also worth mentioning that the bioactivity of the host organisms is totally independent from that of the endophyte. The plant extract of Phragmites australis lacks antibacterial properties, while the metabolites of its endophytic fungi associated with the plant showed moderate bioactivity against gram-positive strains, with some achieving 80% biofilm inhibition of S. aureus [90,114] .
Table 2 lists the various studies on the antibiofilm activities of fungal metabolites against several bacterial test strains. The fungal metabolites were categorized under three groups of sources: 1) non-endophytic-terrestrial, 2) terrestrial endophytes, and 3) marine symbionts. The chemical structures of these metabolites are shown in Figure 3. Polyketides are the most common type of bioactive metabolites found in literature and have a major role in the inhibition of biofilm activities. Steroids and terpenoids as secondary metabolites have been reported from both marine- and soil-derived fungi against various harmful bacteria.
5.2.1. Terpenoids and Steroids
Soil-derived fungi demonstrated superior efficacy, with cholest-5-en-3-ol (4), exhibiting 78% biofilm inhibition against B. subtilis, 59% against S. aureus, and 59% against E. coli [111]. In contrast, two steroidal compounds from a marine-derived Penicillium sp. revealed that β-sitosterol (77) achieved 28% inhibition of biofilm formation in B. subtilis and 64% in S. aureus, while ergosterol (80) displayed 40-55% antibiofilm activity solely against E. coli [113]. Diterpenes such as aszonapyrone A (72) from marine-derived fungus Neosartorya siamensis isolated from a sea fan exhibited 72% efficacy at 9 μg/mL and 94% efficacy at 6.25 μg/mL against S. aureus ATCC 29213 and S. aureus 272123, respectively [115]. However, the Diplodia corticola fungal phytotoxin inhibited biofilm formation at 53% at 1/4 MIC of 1.56 μg/mL against a clinical strain of MRSA and 62% at a 1/4 MIC of 3.12 μg/mL against a clinical strain of P. aeruginosa [116]. A recently described terpenoid from Epicoccum nigrum of the Phaeurus antarcticus seaweed showed significant activity against MRSA with a MBEC of 25 μg/mL and ruptured formed biofilm at 100 μg/mL [83]. Penicillium erubescens KUFA0220, isolated from the marine sponge Neopetrosia sp, showed significant biofilm formation inhibition against Enterococcus faecalis ATCC29212 at 8 μg/mL and 16 μg/mL [89]. This revealed that the terpenoid compounds derived from marine fungi exhibited greater activity against gram-positive bacteria, aligning with a recent study on marine-derived terpenoids showcasing their antibacterial properties [117].
5.2.2. Alkaloids and Peptides
Table 2 reveals that most active alkaloids extracted from fungi are the indole type that includes epifiscalin C (9), fiscalin C (10), gibellamine A (18), two prenylated indole carbaldehydes (62) and (63), neofiscalin A (69), aszonalenin (73), and meleagrin (76). Marine-derived indole alkaloids exhibit significant antibacterial activity against various pathogens, as outlined in the preceding review [118]. In addition, synthetic indole derivatives exhibited antibiofilm activity against Serratia marcescens and interfere with QS [119]. While a non-marine fungal source derived from the Canadian thistle Circium arvense afforded alkaloids, macrocidin A (28), macrocidin Z (29), macrooxazole B (30), macrooxazole C (31) exhibiting no more than 80% inhibition of biofilm formation in S. aureus DSM 1104 [92,120] .
Eight non-ribosomal peptide compounds exhibited significant biofilm inhibition, with seven isolated from marine sources such as phragamide A (55), tenuazonic acid (57), epicorazines A-C (63) and (64), epicotripeptin (66), cyclo(L-Pro-L-Ile) (67), cyclo(L-Pro-L-Tyr) (68) in figure 3. The three secondary metabolites, epicotripeptin (66), cyclo(L-Pro-L-Ile) (67), cyclo(L-Pro-L-Tyr) (68)) were isolated from the endophytic fungus Epicoccum nigrum M13, derived from seagrass and displayed moderate bioactivity against positive bacterial strains [90].
5.2.3. Flavonoids, Phenolics and Polyketide Compounds
Two studies identified fungal flavonoid compounds that exhibited antibiofilm activity. Tricin (8) was isolated from the soil fungus Sarocladium kiliense SDA20, and exhibited weak inhibition of biofilm formation in E. coli and S. aureus [111]. Flavonoids isolated from the endophytic fungus Aspergillus candidus T1219W1, derived from Pittosporum mannii Hook f. exhibited significant biofilm inhibition (exceeding 60%) in E. coli and S. aureus [121]. Other phenolic metabolites are also listed in Table 2. This includes 5[3E,5E)-nona-3,5-dien-1-yl]benzene-1,3-diol (81) isolated from the marine sponge-derived Aspergillus stellatus KUFA 2017 exhibited 100% inhibition of biofilm formation in S. aureus and E. faecalis. In terms of low MWs phenolic compound derived from fungi, one example is 3'-hydroxylbenzyl alcohol (27), from Aspergillus nidulans isolated from a forest soil, could inhibit a broader range of bacterial biofilms that included S. aureus, B. subtilis, Micrococcus luteus (M. luteus), Actinomyces viscosus (Act. viscosus), C. violaceum, E. coli, Klebsiella planticola (K. planticola), and P. aeruginosa. While the compound isolated from the endopyte Daldinia eschscholtzii showed 49% inhibition against P. aeruginosa [122,123] . Three studies found antibiofilm active metabolites against S. aureus DSM 1104 such as the cytochalasin polyketides (13 to 17), phenolic sclerin (24 ) and its diacide (25), along with unsaturated fatty acids (33 to 38) isolated from Hypoxylon fragiforme fungus derived from the Harz Mountains [124,125,126].
Antibiofilm phenolic compounds biosynthesized from the polyketide pathway have been afforded by marine sponge-derived fungi. These include aspulvinones (82 to 87) isolated from Aspergillus flavipes KUFA1152 [85], tenellic acid C (88) and neospinosic acid (89) from Neosartorya spinosa KUFA 1047[86], and bacillisporins (90 and 91) from Talaromyces pinophilus KUFA 1767 [93].
Primary metabolites such as fatty acids (5, 6, 33 to 38) do play a role as well in biofilm inhibition [111,126] , particularly in their absorbance or crossing through the EPS matrix to disrupt the biofilm. Amino acids from endophytic fungi, such as Rhizopus oryzae and Aspergillus tubingensis, have significant biofilm inhibition activity as well [127,128] . Primary metabolites are crucial for microbial growth and exhibit similarities among microbial species; consequently, researchers often investigate secondary metabolites to discover novel antibiofilm compounds that have not been thoroughly examined. Furthermore, climate change and salinity alteration in cases of marine resources affect the production of these secondary metabolites, suggesting fungi or other microorganisms could be good reservoirs of various active substances [129]. However, as shown in Table 2, only two investigation that has extracted antibiofilm compounds from endophytic fungi from seaweeds [84]. Maybe because working with endophytic fungi of seaweeds can be challenging requiring specific sterilization methods to avoid epiphytic contaminants [130]. Then again, there is a trend for researchers to find secondary metabolites from endophytic fungi that have antibiofilm activity, but the challenge arises in increasing the production of these metabolites.

Table 2.
Anti-biofilm activity of reported fungal metabolites.
|
Bioactive compounds |
Fungal species |
Fungal source |
Antibiofilm activity |
Test Bacteria used | Reference |
| Non marine and non-endophytic fungal source | |||||
| sphaeropsidin A (1) |
Diplodia. corticola |
fungal phytotoxin |
Biofilm formation inhibition: Reference MRSA strains: 60% at 3. 12 μg/mL. MRSA clinical: 53% at 1/4 MIC (1.56 μg/mL) Preformed biofilms: (adhesion) Reference P. aeruginosa strains: 50% at a 1/4 MIC: 3.12 μg/mL. clinical P. aeruginosa: 62% at a 1/4 MIC: 3.12 μg/mL. |
MRSA ATCC 43300 reference strains MRSA 1118-116 P. aeruginosa PAO1 as reference strains extended-spectrum beta- lactamase (ESBL) producing P. aeruginosa 0418-925 P. aeruginosa 0418-925. |
[116] |
| pigmentosin A (2) and pigmentosin B (3) | Gibellula sp. | spider |
Biofilm formation inhibition: MIC values: 1.9 and 15.6 μg/mL |
S. aureus DSM1104 |
[131] |
| astucin composed of 92 amino acid residues. | Aspergillus tubingensis | soil |
Biofilm formation inhibition: 99.9%: MBIC: 2 μg/mL against S. aureus MBIC: 8 μg/mL against MRSA |
S. aureus & MRSA |
[128] |
| cholest-5-en-3-ol (4), palmitic acid (5), oleic acid (6), nicotinamide (7), tricin (8) |
Sarocladium kiliense SDA20 | soil |
Biofilm formation inhibition: B. subtilis: Compound 4: 78 %, Compound 5: 71 %, Compound 6: 52% S. aureus: Compounds 4 and 5: 59 %, Compound 8: low inhibition activity around 35%. E coli: Compound 4: 59% Compound 6: 53% Compound 7: 40% Compound 8: ca. 42% P. aeruginosa: No activity |
S. aureus B. subtilis P. aeruginosa E. coli |
[111] |
| epifiscalin C (9), fiscalin C (10), aspernolide B (11), and butyrolactone IV (12) |
fungal extract | NA |
Biofilm formation inhibition: Compounds 9, 10, and 11: Above 60%: 16 μg/mL. Compound 12: Above 50%: 64 μg/mL |
MRSA (ATCC 43300) | [132] |
| cytochalasin A (13), cytochalasin C (14), L-696,474 (15), 19,20-epoxy-cytochalasin C (16), and phenochalasin D (17) |
Hypoxylon fragiforme |
Harz mountains |
Biofilm formation inhibition: Compound 13: 91%: 16 μg/mL, Compound 14: 42%: 256 μg/mL, Compound 15: 44%: 16 μg/mL Compounds 16 and 17: 20 to 40%. |
S. aureus DSM 1104 |
[124] |
| gibellamine A (18) |
Gibellula gamsii strain BCC47868 |
spider |
Biofilm formation inhibition: MIC value of 62.5 μg/mL It significantly inhibited biofilm formation. |
S. aureus DSM1104 (ATCC 25923) |
[133] |
| pretenellin C (19) prototenellin D (20), pretenellin B (21), tenellin (22) and pyridovericin (23) |
Entomopatho-genic fungus Beauveria neobassiana |
Coleoptera |
Biofilm formation inhibition: Compound 19: 46 ± 9% at 31.3 µg/mL Compound 20: 53 ± 7% at 7.8 µg/mL Compound 21: 37 ± 7% at 7.8 µg/mL Compound 22: 36 ± 13% at 7.8 µg/mL. Compound 23: 48 ± 5 % at 62.5 µg/mL. |
S. aureus DSM 346 |
[134] |
| sclerin (24) and its diacid (25) |
Hypoxylon fragiforme |
Harz mountains |
Biofilm formation inhibition: Compound 24 86% at 256 μg/mL Compound 25 80% at 256 μg/mL |
S. aureus DSM 1104 |
[125] |
| harzianic acid (26) |
Trichoderma harzianum, two strains E45 and ET45 |
soil |
Biofilm formation inhibition: at 16 μg/mL: Significant inhibition of biofilm formation in both MRSP and MSSP Pre-formed biofilms: at 32 and 64 μg/mL: Statistically significant disaggregation of pre-formed biofilm produced by MSSP. |
Staphylo- coccus pseudinter- medius (S. pseudinter- medius) methicillin-resistant (MRSP) methicillin-susceptible (MSSP) strains. |
[135] |
| Exopolysaccharide (EPS) | Fomitopsis meliae AGDP-2 | mushrooms |
Biofilm formation inhibition: at 10,000 µg/mL: P. aeruginosa: 86.01% S. typhi: 17.64% |
P. aeruginosa Salmonella typhi (S. typhi) ATCC 6539 |
[136] |
| 3-hydroxyl-benzyl alcohol (27) |
Aspergillus nidulans strain KZR-132 |
forest soil |
Biofilm formation inhibition: at 18.75 μg/mL: S. aureus MTCC 96: 89.2% S. aureus MLS16 MTCC 2940: 88.2 % S. aureus ATCC 6538P: 85.6% B. subtilis MTCC 121: 80% M. luteus MTCC 2470: 90.2% Act. viscosus ATCC 15987: 80.6% C. violaceum: 88% at 37.5 μg/mL: E. coli: 75% K. planticola: 70.9% P. aeruginosa: 65.5% |
S. aureus MTCC 96, S. aureus MLS16 MTCC 2940, S. aureus ATCC 6538P, B. subtilis MTCC 121, M. luteus MTCC 2470, Act. viscosus ATCC 15987 C. violaceum E. coli, K. planticola P. aeruginosa |
[122] |
| macrocidin A (28), macrocidin Z (29), macrooxazole B (30), macrooxazole C (31) |
Phoma macrostoma DAOMC 175,940 |
Circium arvense |
Biofilm formation inhibition: at 250 µg/mL: Compound 30: 65%, Compound 31: 75% Compound 28: 79% Compound 29: 76% Pre-formed biofilms: at 250 µg/mL: Compound 28: 75% Compound 29: 73% |
S. aureus DSM 1104 |
[120] |
| 5-butyl-2-pyridine carboxylic acid (32) |
Aspergillus fumigatus nHF-01 |
NA |
Biofilm formation inhibition: 22.30%: 4 µg/mL against B. cereus 129 µg/mL against E. coli |
Bacillus cereus (B. cereus) MTCC 1272, E. coli MTCC 723, E. coli ATCC DH5α, |
[137] |
| palmitoleic acid (33), palmitelaidic acid (34), 7Z,10Z-hexadecadienoic acid (35), linoleic acid (36), γ-linolenic acid (37), arachidonic acid (38) |
Hypoxylon fragiforme |
Harz mountains |
Biofilm formation inhibition: Compound 33: Sub-MIC: at 64 µg/mL: 54 ± 2% against S. aureus at 120 µg/mL: 49 ± 2% against S. aureus Compound 34: Sub-MIC: at 256 µg/mL: 25 ± 4% against E. coli at 16 µg/mL: 21± 4 % against S. aureus Compound 35: Sub-MIC: at 128 µg/mL: 62 ± 8% against E. coli at 8 µg/mL: 60± 9 % against S. aureus at 8 µg/mL: 22± 14 % against S. epidermidis at 128 µg/mL: 38 ± 12 % against S. mutans Compound 36: Sub-MIC: at 16 µg/mL: 35 ± 15 % against B. cereus at 8 µg/mL: 54 ± 4 % against S. aureus Compound 37: at 4 µg/mL: 58 ± 1 % against S. aureus at 128 µg/mL: 44 ± 19 % against S. epidermidis Compound 38: Sub-MIC: at 64 µg/mL: 36 ± 2 % against B. cereus 35 ± 11 % against S. aureus |
B. cereus DSM 626, E. coli MT102, P. aeruginosa PA14, S. aureus DSM 1104, S. epidermidis ATCC 35984, S. mutans UA59 |
[126] |
| extracellular protein |
Aspergillus oryzae | NA |
Biofilm formation inhibition: Significantly inhibited at 1/4 MIC (75 μg/mL) and 1/2 MIC (150 μg/mL). |
Klebsiella pneumoniae (K. pneumoniae) ESBL |
[138] |
| 2) Endophytic Fungal Metabolites | |||||
| vulculic acid (39), curvulol (40) | Chaetosphaeronema achilleae | Taxus baccata |
Biofilm formation inhibition: Both compounds: 100% at 256 μg/mL. |
S. aureus | [139] |
| 5-(1-hydroxybutyl)-4-methoxy-3- methyl-2H-pyran-2-one (41) | Colletotrichum acutatum | Angelica sinensis | NA |
S. aureus K. pneumonia |
[140] |
| fractions (AF1 and AF2) |
Alternaria destruens (AKL-3) |
Calotropis gigantea |
Biofilm formation inhibition: AF1: 22.5 μg/mL: P. aeruginosa: (58.15%) E. coli (around 49%) S. enterica (around 42%). AF2: 22.5 μg/mL: S. enterica (23.2%). Pre-formed biofilms inhibition: AF1: 22.5 μg/mL: P. aeruginosa (35.58%) E. coli (21.84%) AF2: 22.5 μg/mL: P. aeruginosa 19.2% |
P. aeruginosa E. coli Salmonella enterica (S. enterica) |
[141] |
| 1,3,6-trihydroxy-7-methoxy-9H-xanthen -9-one (42) |
Penicillium citrinum-314 |
Halocnemum strobilaceum |
Biofilm formation inhibition: 100 %: MBIC value of 62.5 μg/mL. |
P. aeruginosa | [142] |
| dihydroauroglaucin (43) |
Aspergillus amstelodami (MK215708) |
Ammi majus L. Fruits |
Biofilm formation inhibition: S. aureus and E. coli: MBIC = 7.81 μg/mL S.mutans: MBIC = 15.63 μg/mL P. aeruginosa: MBIC = 31.25 μg/mL |
S. aureus E. coli S. mutans P. aeruginosa |
[143] |
|
(±)-periplanamide B (44), butyrolactone I (45) |
Aspergillus terreus AH1 |
Ipomoea carnea |
Biofilm formation inhibition: Compound 44: S. aureus 90.65%: B. subtilis 85.09% E. coli 63.144% Compound 45: P. aeruginosa. 71.81% |
P. aeruginosa (ATCC 27853) S. aureus (ATCC 6538-P), E. coli (ATCC 25955) B. subtilis (ATCC 6633) |
[144] |
| 2,4-di-tert-butylphenol (46) |
Daldinia eschscholtzii (TP2-6) |
Tridax procumbens |
Biofilm formation inhibition: 49% at 80 μg/mL |
P. aeruginosa PAO1 | [123] |
| L-tyrosine (47) | Rhizopus oryzae AUMC14899 | Opuntia ficus-indica (L.) |
Biofilm formation inhibition: PA-02: 83% (3 μg/mL) SA-04: 87% (7.5 μg/mL) |
P. aeruginosa PA-02, S. aureus SA-04 |
[127] |
| chlorflavonin (48) and chlorflavonin A (49) |
Aspergillus candidus T1219W1 |
Pittosporum mannii Hook f. |
Biofilm formation inhibition: S. aureus: MBEC50: 256 μg/mL Compound 48: 72.15% Compound 49: 75.32% E. coli MBEC50: 128 μg/mL Compound 48: 80.12% Compound 49: 81.22% |
S. aureus E. coli |
[121] |
| terricoxanthone F (50), aspergillusone A (51), 4-hydroxy vertixanthone (52) alatinone (53) emodin (54) |
Neurospora terricola HDF-Br-2 |
vulnerable
conifer Pseudotsuga gaussenii |
Biofilm formation inhibition: (MBIC) Compounds 50 and 51: 128 μg/mL Compounds 52 and 53: 32 μg/mL Compound 54: 16 μg/mL Pre-formed biofilms: (MBIC) Compound 50: 256 μg/mL. Compounds 51, 52 and 53: 128 μg/mL Compound 54: 32 μg/mL |
S. aureus | [145] |
| Mixture of fatty acids | Arthrographis kalrae |
Coriandrum sativum |
Biofilm formation inhibition: Minimal biofilm inhibitory concentration (MBIC) of 31.3 μg/mL completely inhibited S. mutans biofilm |
S. mutans ATCC 25175 |
[146] |
| 3) Marine fungal metabolites | |||||
| phragamides A (55) and B (56), tenuazonic acid (57) and altechromone (58) altenusin (59) |
A. alternata 13A |
Phragmites australis |
Biofilm formation inhibition: Gram-positive strains: 70 to 80% Gram-negative strains: 40 to 60%. Compound 59 exhibited moderate biofilm formation inhibition only against B. subtilis. |
S. aureus B. subtilis E. coli P. areuginosa |
[90] |
| emodin (54) and physcion (60), and two prenylated indole carbaldehydes (61) and (62) |
Eurotium chevalieri KUFA 0006 | Rhizophora mucronata |
Biofilm formation inhibition: Compounds 54, 60, 61 and 62 showed inhibition of biofilm production in S. aureus ATCC 25923 significantly (*p<0.05). Compound 61: at 64 μg/mL. nearly 80% reduction of S. aureus. |
S. aureus ATCC 25923 E. coli ATCC 25922 |
[88] |
| epicorazines A (63) and C (64) as well as a new terpenoid (65) |
Epicoccum nigrum |
Phaeurus antarcticus (seaweed) |
Biofilm formation inhibition: MBEC: Compound 63: 50 μg/mL Compound 64: 25 μg/mL Compound 65: 25 μg/mL Post- biofilms Inhibition: Compound 65: 100 μg/mL. |
MRSA | [83] |
| epicotripeptin (66) cyclo(L-Pro-L-Ile) (67), cyclo(L-Pro-L-Tyr) (68) |
Epicoccum nigrum M13 (marine endophytes) |
Thalassia hemprichii leaves seagrass |
Biofilm formation inhibition: Compound 66: Gram-positive strains (55 to 70% inhibition) Gram-negative strains (20 to 30% inhibition) Compounds 67 and 68: moderate inhibition of biofilm formation in both Gram-positive strains but were not active against the tested Gram-negative strains. |
S. aureus B. subtilis E. coli P. areuginosa |
[90] |
| neofiscalin A (69) |
Neosartorya siamensis (KUFA 0017) |
marine sponge |
Biofilm formation inhibition: Compound 69 against: MRSA: 96 μg/mL VRE: 80 μg/mL at a concentration of 200 μg/mL, it was able to reduce the metabolic activity of the biofilms by 50%. |
MRSA Vancomycin -resistant E. faecalis (VRE) |
[147] |
| secalonic acid B (70) and D (71) | Penicillium sp. SCSGAF 0023 CCTCCM 2012507 | marine |
Biofilm formation inhibition: Both Inhibited by >90% at 6.25 μg/mL |
S. aureus |
[39] |
| aszonapyrone A (72), aszonalenin (73), methyl ester derivative (74), xanthomegnin (75) |
Neosartory siamensis Neosartorya takakii Aspergillus elegans |
marine |
Biofilm formation inhibition: Compound 72: S. aureus ATCC 29213 at 9 μg/mL: 72% S. aureus 272123 6.25 μg/mL: 94% Compound 73: S. aureus ATCC 29213 at 100 μg/mL: 63% S. aureus 272123 at 6.25 μg/mL: 93% Compound 74: S. aureus ATCC 29213 at 10 μg/mL: 88% S. aureus 272123 at 25 μg/mL: 98% Compound 75: S. aureus ATCC 29213 at 100 μg/mL: 96% S. aureus 272123 at 50 μg/mL: (84%) |
S.aureus ATCC 29213 S. aureus 272123 |
[115] |
|
meleagrin (76) |
Emericella dentata Nq45 |
marine |
Biofilm formation inhibition: 250 μg/mL: 87.1% |
S. aureus ATCC 29213 |
[148] |
| β-sitosterol (77), veridicatol (78), aurantiomide C (79), ergosterol (80) |
Penicillium sp. MMA | marine |
Biofilm formation inhibition: Compound 77: B. subtilis 28% S. aureus 64% Compound 78: B. subtilis 35% Compounds 78, 79, 80: E. coli from 40 -55% |
S. aureus E. coli B. subtilis |
[113] |
| 5[(3E,5E)-nona-3,5-dien-1-yl]benzene-1,3-diol (81) | Aspergillus stellatus KUFA 2017 | marine sponge Mycale sp. |
Biofilm formation inhibition: 100 % at E. faecalis: MIC (16 μg/mL) S. aureus: 2xMIC (32 μg/mL). |
S. aureus ATCC 29213, E. faecalis ATCC 29212 |
[92] |
| Fraction AW1011 |
Aspergillus welwitschiae FMPV 28 |
marine sponge
Taedania sp. |
Biofilm formation inhibition: remarkable decrease in biofilm formation, in a dose-dependent antibiofilm activity. |
S. aureus ATCC 25904 |
[149] |
| Extracellular thermostable antibacterial peptide designated as MFAP9 |
Aspergillus fumigatus BTMF9 |
marine |
Biofilm formation inhibition: - > 85% against all test bacteria. |
B. cereus (NCIM 2155), Bacillus circulans (B. circulans) (NCIM 2107), Bacillus coagulans (NCIM 2030), Bacillus pumilus (NCIM 2189) S. aureus (NCIM 2127) |
[150] |
| aspulvinones R (82), S (83), and U (84) aspulvinones A (85), B’ (86), H (87) |
Aspergillus flavipes KUFA1152 |
marine sponge
Mycale sp |
Biofilm formation inhibition: Compound 87: at MIC (32μg/ mL) and 2xMIC for both strains Compound 86 at ½ MIC (16 μg/ mL). Compounds 82 and 83, all concentrations tested 2xMIC (16μg/ mL), MIC (8 μg/ mL), ½ MIC (4 μg/ mL), including ¼ MIC (2μg/ mL) Mixture of 84 and 85 E. faecalis at MIC (32μg/ mL) and 2xMIC (64 μg/ mL). |
E. faecalis ATCC 29212 S. aureus ATCC 29213 |
[85] |
| tenellic acid C (88), neospinosic acid (89) |
Neosartorya spinosa KUFA 1047 |
marine sponge |
Biofilm formation inhibition: Compound 88: at 64 μg/ mL: E. coli (11.61± 0.09%) E. faecalis (24.11 ± 0.1%) S. aureus (15.54 ± 0.1%) Compound 89: at 64 μg/ mL: E. coli (16.11 ± 0.19%) S. aureus (44 ± 0.06 %) |
E. coli ATCC 25922 E. faecalis ATCC 29212 S. aureus ATCC 29213 |
[86] |
| bacillisporins A (90) and B (91) |
Talaromyces pinophilus KUFA 1767 |
marine sponge |
Biofilm formation inhibition: Compound 90: at 8 μg/ mL (2xMIC): 99.92 ± 0.03% 4 μg/ mL (MIC): 99.81 ± 0.17% Compound 91: at 16 μg/ mL (2xMIC): 99.87 ± 0.05% 8 μg/ mL (MIC): 99.71 ± 0.13% |
S. aureus ATCC 29213 | [93] |
| GKK1032B (92) | Penicillium erubescens KUFA0220 |
marine sponge
Neopetrosia sp |
Biofilm formation inhibition: at: 8 μg/ mL (MIC) and 16 μg/ mL (2xMIC) displayed significant activities. |
E. faecalis ATCC29212 | [89] |
6. Future Directions
6.1. Inducing the Production of Fungal Metabolites
Strategies aimed at increasing the production of fungal metabolites have been currently being developed and are continuously progressing for the purpose of biotechnological scale up. Primarily, the metabolism of fungal strains are being improved to enhance metabolite production by identifying genes involved in the biosynthesis of specific secondary or specialized metabolites while metabolic engineering techniques are being employed to understand these pathways involved in their biosynthesis [151]. The biosynthetic gene cluster (BGC) in the filamentous fungus Curvularia clavata, that is engaged in the synthesis of a potent antifungal cyclic peptide KK-1 have been discovered [151]. Cyclic peptide KK-1 consists of 10-amino acids particularly against the plant pathogen Botrytis cinerea that causes gray mold affecting more than 200 dicotyledonous plant species including essential agricultural crops. Another common practice is employing chemicals, such as 5-azacytidine and suberoylanilide hydroxamic acid, to increase fungal secondary metabolite production [152,153].
As mentioned above, the employment of various media affects the biosynthesis of respective secondary metabolites, thus media optimization must be used for enhancing the production of target bioactive secondary metabolites in fungi as well. Changing the culture conditions by utilizing various media and incubation periods can have an impact on the variety and number of metabolites [154]. Similarly, some fermentation environmental factors such as temperature, aeration, and media composition as high salt stress could either regress or improve the metabolites production [155,156]. Finally, using a variety of combinations of microorganisms as co-cultures is an effective method for inducing the production of secondary metabolites [157,158]. A study demonstrated that interactions with neighboring organisms in the same media can significantly affect the production of fungal secondary metabolites [159].
6.2. Metabolomic Approach
Overall, metabolomics makes it possible to comprehend the metabolic reactions of fungi in detail. The metabolomics approach has brought about a significant expansion in the field of metabolite fingerprinting and profiling, along with the identification and selection of marker metabolites [160]. The analysis of small metabolites (Mr ≤ 1 kDa) produced in cells and organisms in a sample is known as metabolomics. The metabolomics approach is a method that reflects a biological process at a systemic level using statistical techniques and equipment’s [161]. Either targeted or untargeted metabolomics is chosen depending on the aim of the study. As for a non-targeted approach, it is holistic method to profile the metabolites in the sample and detect the presence of new biologically active compounds based on high-resolution mass spectrometry (HRMS) and nuclear magnetic resonance spectroscopy (NMR) coupled to a database for dereplication purposes [162]. Whereas, the targeted metabolomics are quantitative analysis by using mass spectrometry to quantify a specific class of metabolites [163]. Sometimes targeted metabolomics are used after non-targeted metabolomics to verify the validity of results and perform quantitative analysis [161]. Metabolomics studies typically begin with screening metabolites from the target source and isolating the target features that could discriminate respective variables under certain experimental conditions, by acquiring spectral datasets using analytical tools such as HRMS coupled with liquid or gas chromatography (LC or GC) and NMR, followed by processing/analyzing these data, and finally identifying these metabolites using databases [164,165,166,167,168,169]. The potential for future pharmaceutical applications of secondary metabolites is then determined through biological assays [71]. Coupling the metabolomics profile of the spectral dataset and the biological assay results through multivariate analysis helps visualizes the distribution of metabolites between two or more experimental conditions (i.e active versus inactive or between various fermentation conditions) [168,169,170]. Moreover, it will clarify the relationship between metabolites and their behavior in biological processes and displays the metabolites with the assistance of simpler visual plots (i.e. scatter and S-plots) [164,171]. The most common multivariate analyses used along with metabolomics studies are Principal Component Analysis (PCA), Partial Least Square-Discriminant Analysis (PLS-DA), and Orthogonal Partial Least Square-Discriminant Analysis (OPLS-DA) [170].
6.3. Detecting Antibiofim Compounds
In terms of biological assays, the bioassay screens the activity of respective metabolites against the biofilm of specific bacteria. That is done by testing the viability of biofilm and inhibit the bacterial growth through the Alamar blue assay and planktonic assay along with the Minimum inhibitory concentration (MIC) and Minimum biofilm eradication bacteria (MBEC) assay, respectively, as the two metrics used to assess the bioactive antibiofilm compounds [96,169,172,173,174] . QS is another mechanism involved in the development of thin microbial biofilms and regulates bacterial motility and enzymes, inhibiting QS would decrease EPS production. Therefore, the anti-QS activity can also be detected by measuring the motility inhibition activity (i.e. swimming and swarming) of the test bacteria. In addition, it is commended to focus on the virulence factors associated with biofilm-forming bacteria and monitor QS regulation through various mechanisms such as pyocyanin activity, chitinase activity, LasA protease activity, LasA staphylolytic activity, LasB elastase activity, and HCN production [175].
7. Summary and Conclusions
In conclusion, fungal natural products have been successfully offering bioactive antibiofilm compounds which might help in antibiotic resistance crisis. From cited papers, it was shown that fungal-derived natural products can produce secondary metabolites with antibiofilm activity albeit less investigated in comparison to their antimicrobial capability. Moreover, we demonstrated that seaweed endophytes have not been comprehensively explored for their bioactive secondary metabolites. Therefore, further exploring antibiofilm compounds from endophytic fungi associated with seaweed could be found effective in combination with existing antibiotics and prevent multi-resistance. In addition, the synergistic effects of the discovered compounds against bacterial biofilms can be increased by using proven elicitors such as NO-donors and QSIs. So currently, there is an urgent need to turn to this area of research, and we are looking to discover new antibiofilm compounds from seaweed endophytes. At the same time, there is a compulsion for vital effort to accelerate the production of these metabolites and expand our current antibiotic pipeline. Since seaweed metabolites are influenced by geographical location and variable climate changes, the occurrence of endophytic fungal metabolites is expected to fluctuate and vary as well, which further encourages the study of seaweed endophytic fungal metabolites as diverse but sustainable sources to increase the chance of discovering new anti-biofilm compounds. Furthermore, the development of metabolomics approaches employing high resolution instrumentation to afford more reliable spectral datasets that can be coupled with the biological assay results improves the efficiency of detecting and isolating novel antibiofilm compounds.
The challenge of bacterial resistance remains a critical concern that necessitates collaborative efforts across various fields and innovative technological developments. The investigation of marine endophytic fungal strains exhibiting distinctive metabolic adaptations, especially in extreme marine environments, offers an exciting opportunity in drug discovery. Targeting underexplored genera of seaweed endophytic fungi and their metabolites may provide a partial solution to bacterial resistance. Recent technological advancements have facilitated the isolation of new compounds via metabolomics methodologies. Utilizing bioactivity-guided metabolomics alongside LC-MS/MS-based untargeted metabolomics facilitates rapid identification of these compounds. The metabolomics approach supported by multivariate analysis enables the tracking of anti-biofilm compounds and the examination of responses to environmental stressors. Challenges include low product yield and difficulties in compound purification. To address the increasing demand, emphasis must be placed on improving the production and yield of bioactive molecules. Metabolomics-transcriptomics pipelines and synthetic biology tools should establish connections between compound production and BGC expression.
Author Contributions
All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| WHO | World Health Organization |
| EPS | Extracellular polymeric substances |
| NO | Nitric oxide |
| QS | Quorum sensing system |
| AI | Auto inducer |
| SNP | Sodium nitroprusside |
| NPs | Natural products |
| EtOAc | Ethyl acetate |
| sp | species |
| BGC | Biosynthetic gene cluster |
| Mr | Molecular weight |
| KDa | KiloDaltons |
| MS | Mass spectrometry |
| NMR | Nuclear magnetic resonance spectroscopy |
| LC | Liquid chromatography |
| MIC | Minimum inhibitory concentration |
| MBEC | Minimum biofilm eradication bacteria |
| MBIC | Minimum biofilm inhibitory concentration |
| PCA | Principal Component Analysis |
| PLS-DA | Partial Least Square-Discriminant Analysis |
| OPLS-DA | Orthogonal Partial Least Square-Discriminant Analysis |
References
- Talebi Bezmin Abadi, A.; Rizvanov, A. A.; Haertlé, T.; Blatt, N. L. , World Health Organization report: current crisis of antibiotic resistance. BioNanoScience 2019, 9, 778–788. [Google Scholar] [CrossRef]
- Organization, W. H. , Global antimicrobial resistance and use surveillance system (GLASS) report: 2021. 2021.
- Kurt Yilmaz, N.; Schiffer, C. A. , Introduction: drug resistance. Chemical reviews 2021, 121, 3235–3237. [Google Scholar] [CrossRef]
- Murray, C. J.; Ikuta, K. S.; Sharara, F.; Swetschinski, L.; Aguilar, G. R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E. , Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. The Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Nelson, R. E.; Hatfield, K. M.; Wolford, H.; Samore, M. H.; Scott, R. D.; Reddy, S. C.; Olubajo, B.; Paul, P.; Jernigan, J. A.; Baggs, J. National estimates of healthcare costs associated with multidrug-resistant bacterial infections among hospitalized patients in the United States. Clinical Infectious Diseases 2021, 72, (Supplement_1), S17-S26.
- Blair, J. M.; Webber, M. A.; Baylay, A. J.; Ogbolu, D. O.; Piddock, L. J. , Molecular mechanisms of antibiotic resistance. Nature reviews microbiology 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Urban-Chmiel, R.; Marek, A.; Stępień-Pyśniak, D.; Wieczorek, K.; Dec, M.; Nowaczek, A.; Osek, J. , Antibiotic resistance in bacteria—A review. Antibiotics 2022, 11, 1079. [Google Scholar] [CrossRef]
- Sharifi, S.; Bakhshi, B.; Najar-Peerayeh, S. , Significant contribution of the CmeABC Efflux pump in high-level resistance to ciprofloxacin and tetracycline in Campylobacter jejuni and Campylobacter coli clinical isolates. Annals of Clinical Microbiology and Antimicrobials 2021, 20, 1–9. [Google Scholar] [CrossRef]
- Dever, L. A.; Dermody, T. S. , Mechanisms of bacterial resistance to antibiotics. Archives of internal medicine 1991, 151, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Zhao, A.; Sun, J.; Liu, Y. , Understanding bacterial biofilms: From definition to treatment strategies. Frontiers in Cellular and Infection Microbiology 2023, 13, 1137947. [Google Scholar] [CrossRef]
- Wu, H.; Moser, C.; Wang, H.-Z.; Høiby, N.; Song, Z.-J. , Strategies for combating bacterial biofilm infections. International journal of oral science 2015, 7, 1–7. [Google Scholar] [CrossRef]
- Muhammad, M. H.; Idris, A. L.; Fan, X.; Guo, Y.; Yu, Y.; Jin, X.; Qiu, J.; Guan, X.; Huang, T. , Beyond risk: bacterial biofilms and their regulating approaches. Frontiers in microbiology 2020, 11, 928. [Google Scholar] [CrossRef] [PubMed]
- Vestby, L. K.; Grønseth, T.; Simm, R.; Nesse, L. L. , Bacterial biofilm and its role in the pathogenesis of disease. Antibiotics 2020, 9, 59. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wuertz, S. , Bacteria and archaea on Earth and their abundance in biofilms. Nature Reviews Microbiology 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Singh, S.; Singh, S. K.; Chowdhury, I.; Singh, R. , Understanding the mechanism of bacterial biofilms resistance to antimicrobial agents. The open microbiology journal 2017, 11, 53. [Google Scholar] [CrossRef]
- Dufour, D.; Leung, V.; Lévesque, C. M. , Bacterial biofilm: structure, function, and antimicrobial resistance. Endodontic Topics 2010, 22, 2–16. [Google Scholar] [CrossRef]
- Abebe, G. M. , The role of bacterial biofilm in antibiotic resistance and food contamination. International journal of microbiology 2020, 2020. [Google Scholar] [CrossRef]
- Barraud, N.; J Kelso, M.; A Rice, S.; Kjelleberg, S. , Nitric oxide: a key mediator of biofilm dispersal with applications in infectious diseases. Current pharmaceutical design 2015, 21, 31–42. [Google Scholar] [CrossRef]
- Martín-Martín, R. P.; Carcedo-Forés, M.; Camacho-Bolós, P.; García-Aljaro, C.; Angulo-Preckler, C.; Avila, C.; Lluch, J. R.; Garreta, A. G. , Experimental evidence of antimicrobial activity in Antarctic seaweeds: ecological role and antibiotic potential. Polar Biology 2022, 45, 923–936. [Google Scholar] [CrossRef]
- Estrela, A. B.; Abraham, W.-R. , Fungal metabolites for the control of biofilm infections. Agriculture 2016, 6, 37. [Google Scholar] [CrossRef]
- Joo, H.-S.; Deyrup, S. T.; Shim, S. H. , Endophyte-produced antimicrobials: a review of potential lead compounds with a focus on quorum-sensing disruptors. Phytochemistry Reviews 2021, 20, 543–568. [Google Scholar] [CrossRef]
- Gebreyohannes, G.; Nyerere, A.; Bii, C.; Sbhatu, D. B. Challenges of intervention, treatment, and antibiotic resistance of biofilm-forming microorganisms. Heliyon 2019, 5, (8).
- Hall, C. W.; Mah, T.-F. , Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS microbiology reviews 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed]
- Davenport, E. K.; Call, D. R.; Beyenal, H. , Differential protection from tobramycin by extracellular polymeric substances from Acinetobacter baumannii and Staphylococcus aureus biofilms. Antimicrobial agents and chemotherapy 2014, 58, 4755–4761. [Google Scholar] [CrossRef]
- Singh, R.; Ray, P.; Das, A.; Sharma, M. , Penetration of antibiotics through Staphylococcus aureus and Staphylococcus epidermidis biofilms. Journal of antimicrobial chemotherapy 2010, 65, 1955–1958. [Google Scholar] [CrossRef]
- Brackman, G.; Cos, P.; Maes, L.; Nelis, H. J.; Coenye, T. , Quorum sensing inhibitors increase the susceptibility of bacterial biofilms to antibiotics in vitro and in vivo. Antimicrobial agents and chemotherapy 2011, 55, 2655–2661. [Google Scholar] [CrossRef]
- Cirioni, O.; Mocchegiani, F.; Cacciatore, I.; Vecchiet, J.; Silvestri, C.; Baldassarre, L.; Ucciferri, C.; Orsetti, E.; Castelli, P.; Provinciali, M. , Quorum sensing inhibitor FS3-coated vascular graft enhances daptomycin efficacy in a rat model of staphylococcal infection. Peptides 2013, 40, 77–81. [Google Scholar] [CrossRef]
- Grayton, Q. E.; Nguyen, H. K.; Broberg, C. A.; Ocampo, J.; Nagy, S. G.; Schoenfisch, M. H. , Biofilm Dispersal, Reduced Viscoelasticity, and Antibiotic Sensitization via Nitric Oxide-Releasing Biopolymers. ACS Infectious Diseases 2023, 9, 1730–1741. [Google Scholar] [CrossRef]
- Barraud, N.; Hassett, D. J.; Hwang, S.-H.; Rice, S. A.; Kjelleberg, S.; Webb, J. S. , Involvement of nitric oxide in biofilm dispersal of Pseudomonas aeruginosa. Journal of bacteriology 2006, 188, 7344–7353. [Google Scholar] [CrossRef]
- Cragg, G. M.; Newman, D. J.; Snader, K. M. , Natural products in drug discovery and development. Journal of natural products 1997, 60, 52–60. [Google Scholar] [CrossRef]
- Atanasov, A. G.; Zotchev, S. B.; Dirsch, V. M.; Supuran, C. T. , Natural products in drug discovery: Advances and opportunities. Nature reviews Drug discovery 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Dehelean, C. A.; Marcovici, I.; Soica, C.; Mioc, M.; Coricovac, D.; Iurciuc, S.; Cretu, O. M.; Pinzaru, I. , Plant-derived anticancer compounds as new perspectives in drug discovery and alternative therapy. Molecules 2021, 26, 1109. [Google Scholar] [CrossRef]
- Sadeek, A.; Abdallah, E. M. Phytochemical compounds as antibacterial agents a mini review. Saudi Arabia Glob J Pharmaceu Sci 2019, 53, (4).
- Barbieri, R.; Coppo, E.; Marchese, A.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S. F.; Nabavi, S. M. , Phytochemicals for human disease: An update on plant-derived compounds antibacterial activity. Microbiological research 2017, 196, 44–68. [Google Scholar] [CrossRef]
- Borges, A.; J Saavedra, M.; Simoes, M. , Insights on antimicrobial resistance, biofilms and the use of phytochemicals as new antimicrobial agents. Current medicinal chemistry 2015, 22, 2590–2614. [Google Scholar] [CrossRef]
- Liang, M.; Ge, X.; Xua, H.; Ma, K.; Zhang, W.; Zan, Y.; Efferth, T.; Xue, Z.; Hua, X. , Phytochemicals with activity against methicillin-resistant Staphylococcus aureus. Phytomedicine 2022, 100, 154073. [Google Scholar] [CrossRef]
- Artini, M.; Papa, R.; Barbato, G.; Scoarughi, G.; Cellini, A.; Morazzoni, P.; Bombardelli, E.; Selan, L. Bacterial biofilm formation inhibitory activity revealed for plant derived natural compounds. Bioorganic & medicinal chemistry 2012, 20, 920-926.
- Doshi, G.; Aggarwal, G.; Martis, E.; Shanbhag, P. , Novel antibiotics from marine sources. International journal of Pharmaceutical sciences and Nanotechnology 2011, 4, 1446–1461. [Google Scholar]
- Wang, J.; Nong, X.-H.; Zhang, X.-Y.; Xu, X.-Y.; Amin, M.; Qi, S.-H. , Screening of anti-biofilm compounds from marine-derived fungi and the effects of secalonic acid D on Staphylococcus aureus biofilm. 2017.
- Yu, X.; Li, L.; Sun, S.; Chang, A.; Dai, X.; Li, H.; Wang, Y.; Zhu, H. , A cyclic dipeptide from marine fungus Penicillium chrysogenum DXY-1 exhibits anti-quorum sensing activity. ACS omega 2021, 6, 7693–7700. [Google Scholar] [CrossRef]
- Parasuraman, P.; Devadatha, B.; Sarma, V. V.; Ranganathan, S.; Ampasala, D. R.; Siddhardha, B. , Anti-quorum sensing and antibiofilm activities of Blastobotrys parvus PPR3 against Pseudomonas aeruginosa PAO1. Microbial Pathogenesis 2020, 138, 103811. [Google Scholar] [CrossRef]
- Vladkova, T. G.; Martinov, B. L.; Gospodinova, D. N. , Anti-biofilm agents from marine biota. Journal of Chemical Technology and Metallurgy 2023, 58, 825–839. [Google Scholar] [CrossRef]
- Polat, S.; Trif, M.; Rusu, A.; Šimat, V.; Čagalj, M.; Alak, G.; Meral, R.; Özogul, Y.; Polat, A.; Özogul, F. , Recent advances in industrial applications of seaweeds. Critical reviews in food science and nutrition 2023, 63, 4979–5008. [Google Scholar] [CrossRef]
- Rosa, G. P.; Tavares, W. R.; Sousa, P. M.; Pagès, A. K.; Seca, A. M.; Pinto, D. C. , Seaweed secondary metabolites with beneficial health effects: An overview of successes in in vivo studies and clinical trials. Marine drugs 2019, 18, 8. [Google Scholar] [CrossRef]
- Mansuya, P.; Aruna, P.; Sridhar, S.; Kumar, J. S.; Babu, S. Antibacterial activity and qualitative phytochemical analysis of selected seaweeds from Gulf of Mannar region. Journal of Experimental Sciences 2010, 1, (8).
- Lu, W.-J.; Lin, H.-J.; Hsu, P.-H.; Lai, M.; Chiu, J.-Y.; Lin, H.-T. V. Brown and red seaweeds serve as potential efflux pump inhibitors for drug-resistant Escherichia coli. Evidence-Based Complementary and Alternative Medicine 2019, 2019.
- Rima, M.; Trognon, J.; Latapie, L.; Chbani, A.; Roques, C.; El Garah, F. , Seaweed extracts: A promising source of antibiofilm agents with distinct mechanisms of action against Pseudomonas aeruginosa. Marine Drugs 2022, 20, 92. [Google Scholar] [CrossRef]
- Danquah, C. A.; Minkah, P. A. B.; Agana, T. A.; Moyo, P.; Tetteh, M.; Junior, I. O. D.; Amankwah, K. B.; Somuah, S. O.; Ofori, M.; Maharaj, V. J. , Natural Products as Antibiofilm Agents. 2022.
- Jun, J.-Y.; Jung, M.-J.; Jeong, I.-H.; Yamazaki, K.; Kawai, Y.; Kim, B.-M. , Antimicrobial and antibiofilm activities of sulfated polysaccharides from marine algae against dental plaque bacteria. Marine drugs 2018, 16, 301. [Google Scholar] [CrossRef]
- Tang, J.; Wang, W.; Chu, W. , Antimicrobial and anti-quorum sensing activities of phlorotannins from seaweed (Hizikia fusiforme). Frontiers in Cellular and Infection Microbiology 2020, 10, 586750. [Google Scholar] [CrossRef]
- Menaa, F.; Wijesinghe, P.; Thiripuranathar, G.; Uzair, B.; Iqbal, H.; Khan, B. A.; Menaa, B. , Ecological and industrial implications of dynamic seaweed-associated microbiota interactions. Marine drugs 2020, 18, 641. [Google Scholar] [CrossRef]
- Picon, A.; Del Olmo, A.; Nuñez, M. , Bacterial diversity in six species of fresh edible seaweeds submitted to high pressure processing and long-term refrigerated storage. Food Microbiology 2021, 94, 103646. [Google Scholar] [CrossRef] [PubMed]
- Genilloud, O.; Pelaez, F.; Gonzalez, I.; Diez, M. Diversity of actinomycetes and fungi on seaweeds from the Iberian coasts. Microbiologia (Madrid, Spain) 1994, 10, 413-422.
- Francis, M.; Webb, V.; Zuccarello, G. , Marine yeast biodiversity on seaweeds in New Zealand waters. New Zealand Journal of Botany 2016, 54, 30–47. [Google Scholar] [CrossRef]
- Asharaf, S.; Chakraborty, K.; Chakraborty, R. D. , Seaweed-associated heterotrophic bacteria: are they future novel sources of antimicrobial agents against drug-resistant pathogens? Archives of Microbiology 2022, 204, 232. [Google Scholar] [CrossRef]
- Potin, P. , Intimate associations between epiphytes, endophytes, and parasites of seaweeds. In Seaweed biology: novel insights into ecophysiology, ecology and utilization, Springer: 2012; pp 203-234.
- Kumar, J.; Singh, D.; Ghosh, P.; Kumar, A. Endophytic and epiphytic modes of microbial interactions and benefits. Plant-Microbe Interactions in Agro-Ecological Perspectives: Volume 1: Fundamental Mechanisms, Methods and Functions 2017, 227-253.
- Silver, W. L.; Brown, S.; Lugo, A. E. In Biodiversity and biogeochemical cycles, Biodiversity and ecosystem processes in tropical forests, 1996; Springer: pp 49-67.
- Chen, J.; Zang, Y.; Yang, Z.; Qu, T.; Sun, T.; Liang, S.; Zhu, M.; Wang, Y.; Tang, X. , Composition and functional diversity of epiphytic bacterial and fungal communities on marine macrophytes in an intertidal zone. Frontiers in Microbiology 2022, 13, 839465. [Google Scholar] [CrossRef]
- Kaur, M.; Saini, K. C.; Mallick, A.; Bast, F. , Seaweed-associated epiphytic bacteria: diversity, ecological and economic implications. Aquatic Botany 2023, 103698. [Google Scholar] [CrossRef]
- Yuexin, M.; Tian, S.; Haiyan, W.; Yan, W.; Ying, Z. , Antimicrobial activity in the epiphytic fungi on sea weed species in intertidal zone of Dalian. Journal of Dalian Fisheries College 2004, 19, 6–9. [Google Scholar]
- Lemos, M. L.; Toranzo, A. E.; Barja, J. L. , Antibiotic activity of epiphytic bacteria isolated from intertidal seaweeds. Microbial Ecology 1985, 11, 149–163. [Google Scholar] [CrossRef]
- Pratiwy, F. M.; Arifah, F. N. , The potentiality of endophytes bacterial in red algae as anti-microbial agents in aquaculture: A review. 2021.
- Mishra, S.; Sharma, S. , Metabolomic insights into endophyte-derived bioactive compounds. Frontiers in Microbiology 2022, 13, 835931. [Google Scholar] [CrossRef]
- Sugrani, A.; Ahmad, A.; Djide, M. N.; Natsir, H. Toxicological evaluation and antibacterial activity of crude protein extract from endophytic bacteria associated with Algae Eucheuma spinosum. Journal of Physics: Conference Series 2019, 1341, 032006.
- Kandou, F. E. F.; Mangindaan, R. E. P.; Rompas, R. M.; Simbala, H. I. Molecular identification and antibacterial activity of marine-endophytic fungi isolated from sea fan Annella sp. from Bunaken waters, Manado, North Sulawesi, Indonesia. Aquaculture, Aquarium, Conservation & Legislation 2021, 14, 317-327.
- Felício, R. d.; Pavão, G. B.; Oliveira, A. L. L. d.; Erbert, C.; Conti, R.; Pupo, M. T.; Furtado, N. A.; Ferreira, E. G.; Costa-Lotufo, L. V.; Young, M. C. M. , Antibacterial, antifungal and cytotoxic activities exhibited by endophytic fungi from the Brazilian marine red alga Bostrychia tenella (Ceramiales). Revista Brasileira de Farmacognosia 2015, 25, 641–650. [Google Scholar] [CrossRef]
- Handayani, D.; Ananda, N.; Artasasta, M. A.; Ruslan, R.; Fadriyanti, O.; Tallei, T. E. , Antimicrobial activity screening of endophytic fungi extracts isolated from brown algae Padina sp. Journal of Applied Pharmaceutical Science 2019, 9, 009-013. Journal of Applied Pharmaceutical Science 2019, 9, 009–013. [Google Scholar]
- Flewelling, A. J.; Johnson, J. A.; Gray, C. A. , Isolation and bioassay screening of fungal endophytes from North Atlantic marine macroalgae. Botanica Marina 2013, 56, 287–297. [Google Scholar] [CrossRef]
- Parthasarathy, R.; Chandrika, M.; Rao, H. Y.; Kamalraj, S.; Jayabaskaran, C.; Pugazhendhi, A. , Molecular profiling of marine endophytic fungi from green algae: Assessment of antibacterial and anticancer activities. Process Biochemistry 2020, 96, 11–20. [Google Scholar] [CrossRef]
- Conrado, R.; Gomes, T. C.; Roque, G. S. C.; De Souza, A. O. , Overview of bioactive fungal secondary metabolites: Cytotoxic and antimicrobial compounds. Antibiotics 2022, 11, 1604. [Google Scholar] [CrossRef] [PubMed]
- Jindal, A. B.; Singh, B. P.; Paul, A. T. , Plant-associated endophytic fungi and its secondary metabolites against drug-resistant pathogenic microbes. In Antimicrobial Resistance, Springer: 2022; pp 253-288.
- Jha, P.; Kaur, T.; Panja, A.; Paul, S.; Kumar, V.; Malik, T. , Endophytic fungi: Hidden treasure chest of antimicrobial metabolites interrelationship of endophytes and metabolites. Frontiers in Microbiology 2023, 14, 1227830. [Google Scholar] [CrossRef] [PubMed]
- Al-Rajhi, A. M.; Mashraqi, A.; Al Abboud, M. A.; Shater, A.-R. M.; Al Jaouni, S. K.; Selim, S.; Abdelghany, T. M. , Screening of bioactive compounds from endophytic marine-derived fungi in Saudi Arabia: Antimicrobial and anticancer potential. Life 2022, 12, 1182. [Google Scholar] [CrossRef] [PubMed]
- El-Bondkly, E. A. M.; El-Bondkly, A. A. M.; El-Bondkly, A. A. M. Marine endophytic fungal metabolites: A whole new world of pharmaceutical therapy exploration. Heliyon 2021, 7, (3).
- Elsebai, M. F.; Kehraus, S.; Lindequist, U.; Sasse, F.; Shaaban, S.; Gütschow, M.; Josten, M.; Sahl, H.-G.; König, G. M. Antimicrobial phenalenone derivatives from the marine-derived fungus Coniothyrium cereale. Organic & Biomolecular Chemistry 2011, 9, 802-808.
- Suzuki, T.; Ariefta, N. R.; Koseki, T.; Furuno, H.; Kwon, E.; Momma, H.; Harneti, D.; Maharani, R.; Supratman, U.; Kimura, K.-i. New polyketides, paralactonic acids A–E produced by Paraconiothyrium sp. SW-B-1, an endophytic fungus associated with a seaweed, Chondrus ocellatus Holmes. Fitoterapia 2019, 132, 75-81.
- Osterhage, C.; Kaminsky, R.; König, G. M.; Wright, A. D. , Ascosalipyrrolidinone a, an antimicrobial alkaloid, from the obligate marine fungus Ascochyta s alicorniae. The Journal of Organic Chemistry 2000, 65, 6412–6417. [Google Scholar] [CrossRef]
- Liu, X.-H.; Miao, F.-P.; Li, X.-D.; Yin, X.-L.; Ji, N.-Y. , A new sesquiterpene from an endophytic Aspergillus versicolor strain. Natural Product Communications 2012, 7, 1934578X1200700702. [Google Scholar] [CrossRef]
- Yang, R.-y.; Li, C.-y.; Lin, Y.-c.; Peng, G.-t.; She, Z.-g.; Zhou, S.-n. Lactones from a brown alga endophytic fungus (No. ZZF36) from the South China Sea and their antimicrobial activities. Bioorganic & medicinal chemistry letters 2006, 16, 4205-4208.
- Youssef, F. S.; Ashour, M. L.; Singab, A. N. B.; Wink, M. , A comprehensive review of bioactive peptides from marine fungi and their biological significance. Marine drugs 2019, 17, 559. [Google Scholar] [CrossRef]
- Scopel, M.; Abraham, W.-R.; Henriques, A. T.; Macedo, A. J. Dipeptide cis-cyclo (Leucyl-Tyrosyl) produced by sponge associated Penicillium sp. F37 inhibits biofilm formation of the pathogenic Staphylococcus epidermidis. Bioorganic & medicinal chemistry letters 2013, 23, 624-626.
- Santos, G. S. d. Phaeurus antarcticus and its endophytic fungi: chemical diversity of a hidden pharmacy underneath the Antarctic Ocean. Universidade de São Paulo, 2022.
- Jaber, S. A. M. F.; Young, L.; Black, K.; Tate, R.; Edrada-Ebel, R. In Implementing Metabolomics Tools to Optimise the Production of Anti-Biofilm Metabolites of Endophytic Fungi from Scottish Seaweed, 10th European Marine Natural Products Conference, 2019.
- Machado, F. P.; Kumla, D.; Pereira, J. A.; Sousa, E.; Dethoup, T.; Freitas-Silva, J.; Costa, P. M.; Mistry, S.; Silva, A. M.; Kijjoa, A. , Prenylated phenylbutyrolactones from cultures of a marine sponge-associated fungus Aspergillus flavipes KUFA1152. Phytochemistry 2021, 185, 112709. [Google Scholar] [CrossRef]
- de Sá, J. D.; Pereira, J. A.; Dethoup, T.; Cidade, H.; Sousa, M. E.; Rodrigues, I. C.; Costa, P. M.; Mistry, S.; Silva, A. M.; Kijjoa, A. , Anthraquinones, diphenyl ethers, and their derivatives from the culture of the marine sponge-associated fungus Neosartorya spinosa KUFA 1047. Marine Drugs 2021, 19, 457. [Google Scholar] [CrossRef]
- Klomchit, A.; Calderin, J. D.; Jaidee, W.; Watla-Iad, K.; Brooks, S. Napthoquinones from Neocosmospora sp.—Antibiotic activity against acidovorax citrulli, the causative agent of bacterial fruit blotch in watermelon and melon. Journal of Fungi 2021, 7, 370.
- Zin, W. W. M.; Buttachon, S.; Dethoup, T.; Pereira, J. A.; Gales, L.; Inácio, Â.; Costa, P. M.; Lee, M.; Sekeroglu, N.; Silva, A. M. , Antibacterial and antibiofilm activities of the metabolites isolated from the culture of the mangrove-derived endophytic fungus Eurotium chevalieri KUFA 0006. Phytochemistry 2017, 141, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Kumla, D.; Pereira, J. A.; Dethoup, T.; Gales, L.; Freitas-Silva, J.; Costa, P. M.; Lee, M.; Silva, A. M.; Sekeroglu, N.; Pinto, M. M. , Chromone derivatives and other constituents from cultures of the marine sponge-associated fungus Penicillium erubescens KUFA0220 and their antibacterial activity. Marine drugs 2018, 16, 289. [Google Scholar] [CrossRef] [PubMed]
- Qader, M. M.; Hamed, A. A.; Soldatou, S.; Abdelraof, M.; Elawady, M. E.; Hassane, A. S.; Belbahri, L.; Ebel, R.; Rateb, M. E. , Antimicrobial and antibiofilm activities of the fungal metabolites isolated from the marine endophytes Epicoccum nigrum M13 and Alternaria alternata 13A. Marine Drugs 2021, 19, 232. [Google Scholar] [CrossRef] [PubMed]
- Abdelgawad, M. A.; Hamed, A. A.; Nayl, A. A.; Badawy, M. S. E.; Ghoneim, M. M.; Sayed, A. M.; Hassan, H. M.; Gamaleldin, N. M. , The chemical profiling, docking study, and antimicrobial and antibiofilm activities of the endophytic fungi Aspergillus sp. AP5. Molecules 2022, 27, 1704. [Google Scholar] [CrossRef]
- Machado, F. P.; Rodrigues, I. C.; Gales, L.; Pereira, J. A.; Costa, P. M.; Dethoup, T.; Mistry, S.; Silva, A. M.; Vasconcelos, V.; Kijjoa, A. , New Alkylpyridinium Anthraquinone, Isocoumarin, C-Glucosyl Resorcinol Derivative and Prenylated Pyranoxanthones from the Culture of a Marine Sponge-Associated Fungus, Aspergillus stellatus KUFA 2017. Marine Drugs 2022, 20, 672. [Google Scholar] [CrossRef]
- Machado, F. P.; Rodrigues, I. C.; Georgopolou, A.; Gales, L.; Pereira, J. A.; Costa, P. M.; Mistry, S.; Hafez Ghoran, S.; Silva, A. M.; Dethoup, T. , New hybrid phenalenone dimer, highly conjugated dihydroxylated C28 steroid and azaphilone from the culture extract of a marine sponge-associated fungus, Talaromyces pinophilus KUFA 1767. Marine Drugs 2023, 21, 194. [Google Scholar] [CrossRef]
- Edrada-Ebel, R.; Michael, A.; Alsaleh, F.; Zaharuddin, H. , Penicillium, and Fusarium for the Antibiotic Pipeline. In Fungi Bioactive Metabolites: Integration of Pharmaceutical Applications, Deshmukh, S. K.; Takahashi, J. A., Ed.; Saxena, S., Eds. Springer Nature Singapore: Singapore, 2024. [Google Scholar]
- Salem, S. H.; El-Maraghy, S. S.; Abdel-Mallek, A. Y.; Abdel-Rahman, M. A.; Hassanein, E. H.; Al-Bedak, O. A.; El-Aziz, F. E.-Z. A. A. , The antimicrobial, antibiofilm, and wound healing properties of ethyl acetate crude extract of an endophytic fungus Paecilomyces sp.(AUMC 15510) in earthworm model. Scientific Reports 2022, 12, 19239. [Google Scholar] [CrossRef]
- Hengzhuang, W.; Wu, H.; Ciofu, O.; Song, Z.; Høiby, N. , Pharmacokinetics/pharmacodynamics of colistin and imipenem on mucoid and nonmucoid Pseudomonas aeruginosa biofilms. Antimicrobial agents and chemotherapy 2011, 55, 4469–4474. [Google Scholar] [CrossRef]
- Owlia, P.; Nosrati, R.; Alaghehbandan, R.; Lari, A. R. Antimicrobial susceptibility differences among mucoid and non-mucoid Pseudomonas aeruginosa isolates. GMS hygiene and infection control 2014, 9, (2).
- Kapoor, P.; Murphy, P. Combination antibiotics against Pseudomonas aeruginosa, representing common and rare cystic fibrosis strains from different Irish clinics. Heliyon 2018, 4, (3).
- Attia, E. Z.; Khalifa, B. A.; Shaban, G. M.; Abdelraheem, W. M.; Mustafa, M.; Abdelmohsen, U. R.; Mo'men, H. , Discovering the chemical profile, antimicrobial and antibiofilm potentials of the endophytic fungus Penicillium chrysogenum isolated from Artemisia judaica L. assisted with docking studies. South African Journal of Botany 2022, 151, 218–227. [Google Scholar] [CrossRef]
- Wong Chin, J. M.; Puchooa, D.; Bahorun, T.; Jeewon, R. , Antimicrobial properties of marine fungi from sponges and brown algae of Mauritius. Mycology 2021, 12, 231–244. [Google Scholar] [CrossRef]
- Westphal, K. R.; Heidelbach, S.; Zeuner, E. J.; Riisgaard-Jensen, M.; Nielsen, M. E.; Vestergaard, S. Z.; Bekker, N. S.; Skovmark, J.; Olesen, C. K.; Thomsen, K. H. , The effects of different potato dextrose agar media on secondary metabolite production in Fusarium. International Journal of Food Microbiology 2021, 347, 109171. [Google Scholar] [CrossRef]
- Charria-Girón, E.; Espinosa, M. C.; Zapata-Montoya, A.; Méndez, M. J.; Caicedo, J. P.; Dávalos, A. F.; Ferro, B. E.; Vasco-Palacios, A. M.; Caicedo, N. H. , Evaluation of the antibacterial activity of crude extracts obtained from cultivation of native endophytic fungi belonging to a tropical montane rainforest in Colombia. Frontiers in Microbiology 2021, 12, 2515. [Google Scholar] [CrossRef] [PubMed]
- Jaber, S. A. M. F. Metabolomic profiling of antibiofilm compounds from fungal endophytes derived from Scottish seaweeds. http://purl.org/coar/resource_type/c_db06, University of Strathclyde, 2021.
- Doreswamy, K.; Shenoy, P.; Bhaskar, S.; Kini, R. K.; Sekhar, S. Woodfordia fruticosa (Linn.) Kurz’s fungal endophyte Mucor souzae’s secondary metabolites, kaempferol and quercetin, bestow biological activities. Journal of Applied Biology and Biotechnology 2022, 10, 44-53.
- Bajpai, R.; Yusuf, M. A.; Upreti, D. K.; Gupta, V. K.; Singh, B. N. , Endolichenic fungus, Aspergillus quandricinctus of Usnea longissima inhibits quorum sensing and biofilm formation of Pseudomonas aeruginosa PAO1. Microbial pathogenesis 2020, 140, 103933. [Google Scholar]
- İrez, E. İ.; Doğru, N. H.; Demir, N. Fomes fomentarius (L.) Fr. extracts as sources of an antioxidant, antimicrobial and antibiofilm agents. Biologica Nyssana 2021, 12, (1).
- Kaur, N.; Arora, D. S. , Prospecting the antimicrobial and antibiofilm potential of Chaetomium globosum an endophytic fungus from Moringa oleifera. AMB Express 2020, 10, 206. [Google Scholar] [CrossRef]
- Kaur, N.; Arora, D. S.; Kalia, N.; Kaur, M. , Antibiofilm, antiproliferative, antioxidant and antimutagenic activities of an endophytic fungus Aspergillus fumigatus from Moringa oleifera. Molecular biology reports 2020, 47, 2901–2911. [Google Scholar] [CrossRef]
- Caruso, D. J.; Palombo, E. A.; Moulton, S. E.; Duggan, P. J.; Zaferanloo, B. , Antibacterial and Antibiofilm Activity of Endophytic Alternaria sp. Isolated from Eremophila longifolia. Antibiotics 2023, 12, 1459. [Google Scholar] [CrossRef]
- Rashmi, M.; Meena, H.; Meena, C.; Kushveer, J.; Busi, S.; Murali, A.; Sarma, V. , Anti-quorum sensing and antibiofilm potential of Alternaria alternata, a foliar endophyte of Carica papaya, evidenced by QS assays and in-silico analysis. Fungal biology 2018, 122, 998–1012. [Google Scholar] [CrossRef]
- Eskander, D. M.; Atalla, S. M.; Hamed, A. A.; El-Khrisy, E.-D. A. Investigation of secondary metabolites and its bioactivity from Sarocladium kiliense SDA20 using shrimp shell wastes. Pharmacognosy Journal 2020, 12, (3).
- Njateng, G. S. S.; Du, Z.; Gatsing, D.; Mouokeu, R. S.; Liu, Y.; Zang, H.-X.; Gu, J.; Luo, X.; Kuiate, J.-R. , Antibacterial and antioxidant properties of crude extract, fractions and compounds from the stem bark of Polyscias fulva Hiern (Araliaceae). BMC complementary and alternative medicine 2017, 17, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Boulis, A. G.; Hamed, A. A.; El-Awady, M. E.; Mohamed, A. R.; Eliwa, E. M.; Asker, M. M.; Shaaban, M. Diverse bioactive metabolites from Penicillium sp. MMA derived from the red sea: structure identification and biological activity studies. Archives of Microbiology 2020, 202, 1985-1996.
- Chikowe, G. R.; Mpala, L. N.; Cock, I. E. Phragmites australis (Cav.) Trin. Ex Steud. Leaf Extracts Lack Anti-bacterial Activity and are Non-toxic in vitro. Pharmacognosy Communications 2023, 13, (4).
- Durães, F.; Szemerédi, N.; Kumla, D.; Pinto, M.; Kijjoa, A.; Spengler, G.; Sousa, E. , Metabolites from marine-derived fungi as potential antimicrobial adjuvants. Marine drugs 2021, 19, 475. [Google Scholar] [CrossRef] [PubMed]
- Roscetto, E.; Masi, M.; Esposito, M.; Di Lecce, R.; Delicato, A.; Maddau, L.; Calabrò, V.; Evidente, A.; Catania, M. R. , Anti-biofilm activity of the fungal phytotoxin sphaeropsidin A against clinical isolates of antibiotic-resistant bacteria. Toxins 2020, 12, 444. [Google Scholar] [CrossRef]
- Liu, X.; Xin, J.; Sun, Y.; Zhao, F.; Niu, C.; Liu, S. , Terpenoids from Marine Sources: A Promising Avenue for New Antimicrobial Drugs. Marine Drugs 2024, 22, 347. [Google Scholar] [CrossRef]
- França, P. H.; Barbosa, D. P.; da Silva, D. L.; Ribeiro, Ê. A.; Santana, A. E.; Santos, B. V.; Barbosa-Filho, J. M.; Quintans, J. S.; Barreto, R. S.; Quintans-Júnior, L. J. , Indole alkaloids from marine sources as potential leads against infectious diseases. BioMed research international 2014, 2014, 375423. [Google Scholar] [CrossRef]
- Sethupathy, S.; Sathiyamoorthi, E.; Kim, Y.-G.; Lee, J.-H.; Lee, J. , Antibiofilm and antivirulence properties of indoles against Serratia marcescens. Frontiers in microbiology 2020, 11, 584812. [Google Scholar] [CrossRef]
- Kemkuignou, B. M.; Treiber, L.; Zeng, H.; Schrey, H.; Schobert, R.; Stadler, M. Macrooxazoles a–d, new 2, 5-disubstituted oxazole-4-carboxylic acid derivatives from the plant pathogenic fungus Phoma macrostoma. Molecules 2020, 25, (23).
- Talle Juidzou, G.; Gisèle Mouafo Anoumedem, E.; Kehdinga Sema, D.; Flaure Tsague Tankeu, V.; Bosco Leutcha, P.; Yetendje Chimi, L.; Paul Dzoyem, J.; Kouam Fogue, S.; Sewald, N.; Choudhary, M. I. , A New Unsaturated Aliphatic Anhydride from Aspergillus candidus T 12 19W1, an Endophytic Fungus, from Pittosporum mannii Hook f. Journal of Chemistry 2023, 2023. [Google Scholar]
- Kumar, C. G.; Mongolla, P.; Pombala, S.; Bandi, S.; Babu, K.; Ramakrishna, K. , Biological evaluation of 3-hydroxybenzyl alcohol, an extrolite produced by Aspergillus nidulans strain KZR-132. Journal of applied microbiology 2017, 122, 1518–1528. [Google Scholar] [CrossRef]
- Mishra, R.; Kushveer, J. S.; Khan, M. I. K.; Pagal, S.; Meena, C. K.; Murali, A.; Dhayalan, A.; Venkateswara Sarma, V. , 2, 4-Di-tert-butylphenol isolated from an endophytic fungus, Daldinia eschscholtzii, reduces virulence and quorum sensing in Pseudomonas aeruginosa. Frontiers in Microbiology 2020, 11, 1668. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, K. T.; Wendt, L.; Surup, F.; Kretz, R.; Chepkirui, C.; Wittstein, K.; Boonlarppradab, C.; Wongkanoun, S.; Luangsa-Ard, J.; Stadler, M. , Cytochalasans act as inhibitors of biofilm formation of Staphylococcus aureus. Biomolecules 2018, 8, 129. [Google Scholar] [CrossRef]
- Yuyama, K. T.; Chepkirui, C.; Wendt, L.; Fortkamp, D.; Stadler, M.; Abraham, W.-R. , Bioactive compounds produced by Hypoxylon fragiforme against Staphylococcus aureus biofilms. Microorganisms 2017, 5, 80. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, K. T.; Rohde, M.; Molinari, G.; Stadler, M.; Abraham, W.-R. , Unsaturated fatty acids control biofilm formation of Staphylococcus aureus and other gram-positive bacteria. Antibiotics 2020, 9, 788. [Google Scholar] [CrossRef]
- El-Zawawy, N. A.; Ali, S. S.; Nouh, H. S. , Exploring the potential of Rhizopus oryzae AUMC14899 as a novel endophytic fungus for the production of l-tyrosine and its biomedical applications. Microbial Cell Factories 2023, 22, 31. [Google Scholar] [CrossRef]
- Yazici, A.; Örtücü, S.; Taşkin, M. , Screening and characterization of a novel Antibiofilm polypeptide derived from filamentous Fungi. Journal Of Proteomics 2021, 233, 104075. [Google Scholar] [CrossRef] [PubMed]
- Jones, E. G.; Ramakrishna, S.; Vikineswary, S.; Das, D.; Bahkali, A. H.; Guo, S.-Y.; Pang, K.-L. , How do fungi survive in the sea and respond to climate change? Journal of Fungi 2022, 8, 291. [Google Scholar] [CrossRef]
- Teixeira, T. R.; Santos, G. S. d.; Armstrong, L.; Colepicolo, P.; Debonsi, H. M. , Antitumor potential of seaweed derived-endophytic fungi. Antibiotics 2019, 8, 205. [Google Scholar] [CrossRef] [PubMed]
- Helaly, S. E.; Kuephadungphan, W.; Phainuphong, P.; Ibrahim, M. A.; Tasanathai, K.; Mongkolsamrit, S.; Luangsa-Ard, J. J.; Phongpaichit, S.; Rukachaisirikul, V.; Stadler, M. , Pigmentosins from Gibellula sp. as antibiofilm agents and a new glycosylated asperfuran from Cordyceps javanica. Beilstein Journal of Organic Chemistry 2019, 15, 2968–2981. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Rodríguez, O. P.; García-Contreras, R.; Aguayo-Ortiz, R.; Figueroa, M. , Antimicrobial and antibiofilm activity of fungal metabolites on methicillin-resistant Staphylococcus aureus (ATCC 43300) mediated by SarA and AgrA. Biofouling 2023, 39, 830–837. [Google Scholar] [CrossRef]
- Kuephadungphan, W.; Macabeo, A. P. G.; Luangsa-Ard, J. J.; Tasanathai, K.; Thanakitpipattana, D.; Phongpaichit, S.; Yuyama, K.; Stadler, M. , Studies on the biologically active secondary metabolites of the new spider parasitic fungus Gibellula gamsii. Mycological Progress 2019, 18, 135–146. [Google Scholar] [CrossRef]
- Toshe, R.; Charria-Girón, E.; Khonsanit, A.; Luangsa-Ard, J. J.; Khalid, S. J.; Schrey, H.; Ebada, S. S.; Stadler, M. , Bioprospection of Tenellins Produced by the Entomopathogenic Fungus Beauveria neobassiana. Journal of Fungi 2024, 10, 69. [Google Scholar] [CrossRef]
- De Filippis, A.; Nocera, F. P.; Tafuri, S.; Ciani, F.; Staropoli, A.; Comite, E.; Bottiglieri, A.; Gioia, L.; Lorito, M.; Woo, S. L. , Antimicrobial activity of harzianic acid against Staphylococcus pseudintermedius. Natural Product Research 2021, 35, 5440–5445. [Google Scholar] [CrossRef]
- Prajapati, D.; Bhatt, A.; Gupte, A. , Evaluation of Bioactive Attributes and Emulsification Potential of Exopolysaccharide Produced by a Brown-rot Fungus Fomitopsis meliae AGDP-2. Applied Biochemistry and Biotechnology 2023, 195, 2974–2992. [Google Scholar] [CrossRef]
- Mandal, V.; Ghosh, N. N.; Mitra, P. K.; Mandal, S.; Mandal, V. , Production and characterization of a broad-spectrum antimicrobial 5-butyl-2-pyridine carboxylic acid from Aspergillus fumigatus nHF-01. Scientific Reports 2022, 12, 6006. [Google Scholar] [CrossRef]
- Rachma, L. N.; Fitri, L. E.; Prawiro, S. R.; Mardining Raras, T. Y. , Aspergillus oryzae attenuates quorum sensing-associated virulence factors and biofilm formation in Klebsiella pneumoniae extended-spectrum beta-lactamases. F1000Research 2022, 11, 1148. [Google Scholar] [CrossRef]
- Narmani, A.; Teponno, R. B.; Helaly, S. E.; Arzanlou, M.; Stadler, M. , Cytotoxic, anti-biofilm and antimicrobial polyketides from the plant associated fungus Chaetosphaeronema achilleae. Fitoterapia 2019, 139, 104390. [Google Scholar] [CrossRef] [PubMed]
- Yehia, R. S. , Multi-function of a new bioactive secondary metabolite derived from endophytic fungus Colletotrichum acutatum of Angelica sinensis. Journal of Microbiology and Biotechnology 2023, 33, 806. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Sharma, P.; Kaur, R.; Kaur, S.; Kaur, A. , Assessment of alpha glucosidase inhibitors produced from endophytic fungus Alternaria destruens as antimicrobial and antibiofilm agents. Molecular Biology Reports 2020, 47, 423–432. [Google Scholar] [CrossRef]
- Abdel Razek, M. M.; Moussa, A. Y.; El-Shanawany, M. A.; Singab, A. N. B. A New Phenolic Alkaloid from Halocnemum strobilaceum endophytes: antimicrobial, antioxidant and biofilm Inhibitory activities. Chemistry & Biodiversity 2020, 17, e2000496.
- Fathallah, N.; Raafat, M. M.; Issa, M. Y.; Abdel-Aziz, M. M.; Bishr, M.; Abdelkawy, M. A.; Salama, O. Bio-guided fractionation of prenylated benzaldehyde derivatives as potent antimicrobial and antibiofilm from Ammi majus L. fruits-associated Aspergillus amstelodami. Molecules 2019, 24, 4118.
- Elkhouly, H. I.; Sidkey, N. M.; Ghareeb, M. A.; El Hosainy, A. M.; Hamed, A. A. , Bioactive secondary metabolites from endophytic Aspergillus terreus AH1 isolated from Ipomoea carnea growing in Egypt. Egyptian Journal of Chemistry 2021, 64, 7611–7620. [Google Scholar]
- Chen, H.-W.; Wu, X.-Y.; Zhao, Z.-Y.; Huang, Z.-Q.; Lei, X.-S.; Yang, G.-X.; Li, J.; Xiong, J.; Hu, J.-F. , Terricoxanthones A–E, unprecedented dihydropyran-containing dimeric xanthones from the endophytic fungus Neurospora terricola HDF-Br-2 associated with the vulnerable conifer Pseudotsuga gaussenii. Phytochemistry 2024, 219, 113963. [Google Scholar] [CrossRef]
- Abdel-Aziz, M. M.; M. Emam, T.; Raafat, M. M., Hindering of cariogenic streptococcus mutans biofilm by fatty acid array derived from an endophytic arthrographis kalrae strain. Biomolecules 2020, 10, 811. [Google Scholar] [CrossRef]
- Bessa, L. J.; Buttachon, S.; Dethoup, T.; Martins, R.; Vasconcelos, V.; Kijjoa, A.; Martins da Costa, P. , Neofiscalin A and fiscalin C are potential novel indole alkaloid alternatives for the treatment of multidrug-resistant Gram-positive bacterial infections. FEMS Microbiology Letters 2016, 363, fnw150. [Google Scholar] [CrossRef]
- Hamed, A.; Abdel-Razek, A. S.; Araby, M.; Abu-Elghait, M.; El-Hosari, D. G.; Frese, M.; Soliman, H. S.; Stammler, H. G.; Sewald, N.; Shaaban, M. , Meleagrin from marine fungus Emericella dentata Nq45: crystal structure and diverse biological activity studies. Natural product research 2021, 35, 3830–3838. [Google Scholar] [CrossRef]
- Loges, L. A.; Silva, D. B.; Paulino, G. V.; Landell, M. F.; Macedo, A. J. , Polyketides from marine-derived Aspergillus welwitschiae inhibit Staphylococcus aureus virulence factors and potentiate vancomycin antibacterial activity in vivo. Microbial pathogenesis 2020, 143, 104066. [Google Scholar] [CrossRef]
- Raghavan, R. M. K.; Pannippara, M. A.; Kesav, S.; Mathew, A.; Bhat, S. G.; Aa, M. H.; Elyas, K. , MFAP9: Characterization of an extracellular thermostable antibacterial peptide from marine fungus with biofilm eradication potential. Journal of Pharmaceutical and Biomedical Analysis 2021, 194, 113808. [Google Scholar]
- Yamaguchi, S.; Fujioka, T.; Yoshimi, A.; Kumagai, T.; Umemura, M.; Abe, K.; Machida, M.; Kawai, K. , Discovery of a gene cluster for the biosynthesis of novel cyclic peptide compound, KK-1, in Curvularia clavata. Frontiers in Fungal Biology 2023, 3, 1081179. [Google Scholar] [CrossRef] [PubMed]
- Munusamy, M.; Ching, K. C.; Yang, L. K.; Crasta, S.; Gakuubi, M. M.; Chee, Z. Y.; Wibowo, M.; Leong, C. Y.; Kanagasundaram, Y.; Ng, S. B. , Chemical elicitation as an avenue for discovery of bioactive compounds from fungal endophytes. Frontiers in Chemistry 2022, 10, 1024854. [Google Scholar] [CrossRef] [PubMed]
- Gakuubi, M. M.; Ching, K. C.; Munusamy, M.; Wibowo, M.; Liang, Z.-X.; Kanagasundaram, Y.; Ng, S. B. , Enhancing the discovery of bioactive secondary metabolites from fungal endophytes using chemical elicitation and variation of fermentation media. Frontiers in Microbiology 2022, 13, 898976. [Google Scholar] [CrossRef]
- VanderMolen, K. M.; Raja, H. A.; El-Elimat, T.; Oberlies, N. H. , Evaluation of culture media for the production of secondary metabolites in a natural products screening program. Amb Express 2013, 3, 1–7. [Google Scholar] [CrossRef]
- Shinta, D. Y.; Juliandi, M. D.; Widyastuti, W.; Sonata, H.; Saryono, S. , Microbial inhibition test and optimization of temperature, aeration fermentation of endophytic Fusarium sp LBKURCC 41 from Dahlia tuber (Dahlia variabilis). Bali Medical Journal 2023, 12, 77–82. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, Z.; Sun, K.; Zhu, W. , Effects of high salt stress on secondary metabolite production in the marine-derived fungus Spicaria elegans. Marine drugs 2011, 9, 535–542. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, W.-C.; Shi, X.; Zheng, H.-Z.; Zheng, Z.-H.; Lu, X.-H.; Xing, Y.; Ji, K.; Liu, M.; Dong, Y.-S. , Inducing secondary metabolite production of Aspergillus sydowii through microbial co-culture with Bacillus subtilis. Microbial Cell Factories 2021, 20, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Caudal, F.; Tapissier-Bontemps, N.; Edrada-Ebel, R. A. Impact of Co-Culture on the Metabolism of Marine Microorganisms. Mar Drugs 2022, 20, (2).
- Azzollini, A.; Boggia, L.; Boccard, J.; Sgorbini, B.; Allard, P.-M.; Rubiolo, P.; Rudaz, S.; Wolfender, J.-L. , Dynamics of metabolite induction in fungal co-cultures by metabolomics at both volatile and non-volatile levels. Frontiers in microbiology 2018, 9, 326597. [Google Scholar] [CrossRef]
- Okada, T.; Mochamad Afendi, F.; Altaf-Ul-Amin, M.; Takahashi, H.; Nakamura, K.; Kanaya, S. Metabolomics of medicinal plants: the importance of multivariate analysis of analytical chemistry data. Current Computer-Aided Drug Design 2010, 6, 179-196.
- Lajis, N.; Maulidiani, M.; Abas, F.; Ismail, I. , Metabolomics approach in pharmacognosy. In Pharmacognosy, Elsevier: 2017; pp 597-616.
- Naz, S.; Vallejo, M.; García, A.; Barbas, C. , Method validation strategies involved in non-targeted metabolomics. Journal of Chromatography A 2014, 1353, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, M.; Caterino, M.; Ruoppolo, M. , Targeted metabolomics. In Metabolomics Perspectives, Elsevier: 2022; pp 219-236.
- Nagarajan, K.; Ibrahim, B.; Ahmad Bawadikji, A.; Lim, J.-W.; Tong, W.-Y.; Leong, C.-R.; Khaw, K. Y.; Tan, W.-N. , Recent developments in metabolomics studies of endophytic fungi. Journal of Fungi 2021, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Tawfike, A. F.; Tate, R.; Abbott, G.; Young, L.; Viegelmann, C.; Schumacher, M.; Diederich, M.; Edrada-Ebel, R. Metabolomic Tools to Assess the Chemistry and Bioactivity of Endophytic Aspergillus Strain. Chem Biodivers 2017, 14, (10).
- Tawfike, A. F.; Romli, M.; Clements, C.; Abbott, G.; Young, L.; Schumacher, M.; Diederich, M.; Farag, M.; Edrada-Ebel, R. Isolation of anticancer and anti-trypanosome secondary metabolites from the endophytic fungus Aspergillus flocculus via bioactivity guided isolation and MS based metabolomics. J Chromatogr B Analyt Technol Biomed Life Sci 2019, 1106-1107, 71-83.
- Tawfike, A. F.; Viegelmann, C.; Edrada-Ebel, R. , Metabolomics and dereplication strategies in natural products. Methods Mol Biol 2013, 1055, 227–44. [Google Scholar]
- Mazlan, N. W.; Tate, R.; Yusoff, Y. M.; Clements, C.; Edrada-Ebel, R. , Metabolomics-Guided Isolation of Anti-Trypanosomal Compounds from Endophytic Fungi of the Mangrove plant Avicennia Lanata. Curr Med Chem 2020, 27, 1815–1835. [Google Scholar] [CrossRef]
- Singh, S.; Nwagwu, E.; Young, L.; Kumar, P.; Shinde, P. B.; Edrada-Ebel, R. Targeted Isolation of Antibiofilm Compounds from Halophytic Endophyte Bacillus velezensis 7NPB-3B Using LC-HR-MS-Based Metabolomics. Microorganisms 2024, 12, (2).
- Alhadrami, H. A.; Sayed, A. M.; El-Gendy, A. O.; Shamikh, Y. I.; Gaber, Y.; Bakeer, W.; Sheirf, N. H.; Attia, E. Z.; Shaban, G. M.; Khalifa, B. A. , A metabolomic approach to target antimalarial metabolites in the Artemisia annua fungal endophytes. Scientific Reports 2021, 11, 2770. [Google Scholar] [CrossRef]
- Saccenti, E.; Hoefsloot, H. C.; Smilde, A. K.; Westerhuis, J. A.; Hendriks, M. M. , Reflections on univariate and multivariate analysis of metabolomics data. Metabolomics 2014, 10, 361–374. [Google Scholar] [CrossRef]
- Dong, D.; Thomas, N.; Ramezanpour, M.; Psaltis, A. J.; Huang, S.; Zhao, Y.; Thierry, B.; Wormald, P.-J.; Prestidge, C. A.; Vreugde, S. , Inhibition of Staphylococcus aureus and Pseudomonas aeruginosa biofilms by quatsomes in low concentrations. Experimental Biology and Medicine 2020, 245, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Peeters, E.; Nelis, H. J.; Coenye, T. , Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. Journal of microbiological methods 2008, 72, 157–165. [Google Scholar] [CrossRef]
- Repp, K. K.; Menor, S. A.; Pettit, R. K. , Microplate Alamar blue assay for susceptibility testing of Candida albicans biofilms. Medical mycology 2007, 45, 603–607. [Google Scholar] [CrossRef]
- Preda, V. G.; Săndulescu, O. Communication is the key: biofilms, quorum sensing, formation and prevention. Discoveries 2019, 7, (3).
Figure 1.
Bacterial biofilm development. A) Reversible adherence, B) Micro-colony formation (irreversible attachment). C) Biofilm maturation. D) Biofilm Dispersion.
Figure 1.
Bacterial biofilm development. A) Reversible adherence, B) Micro-colony formation (irreversible attachment). C) Biofilm maturation. D) Biofilm Dispersion.
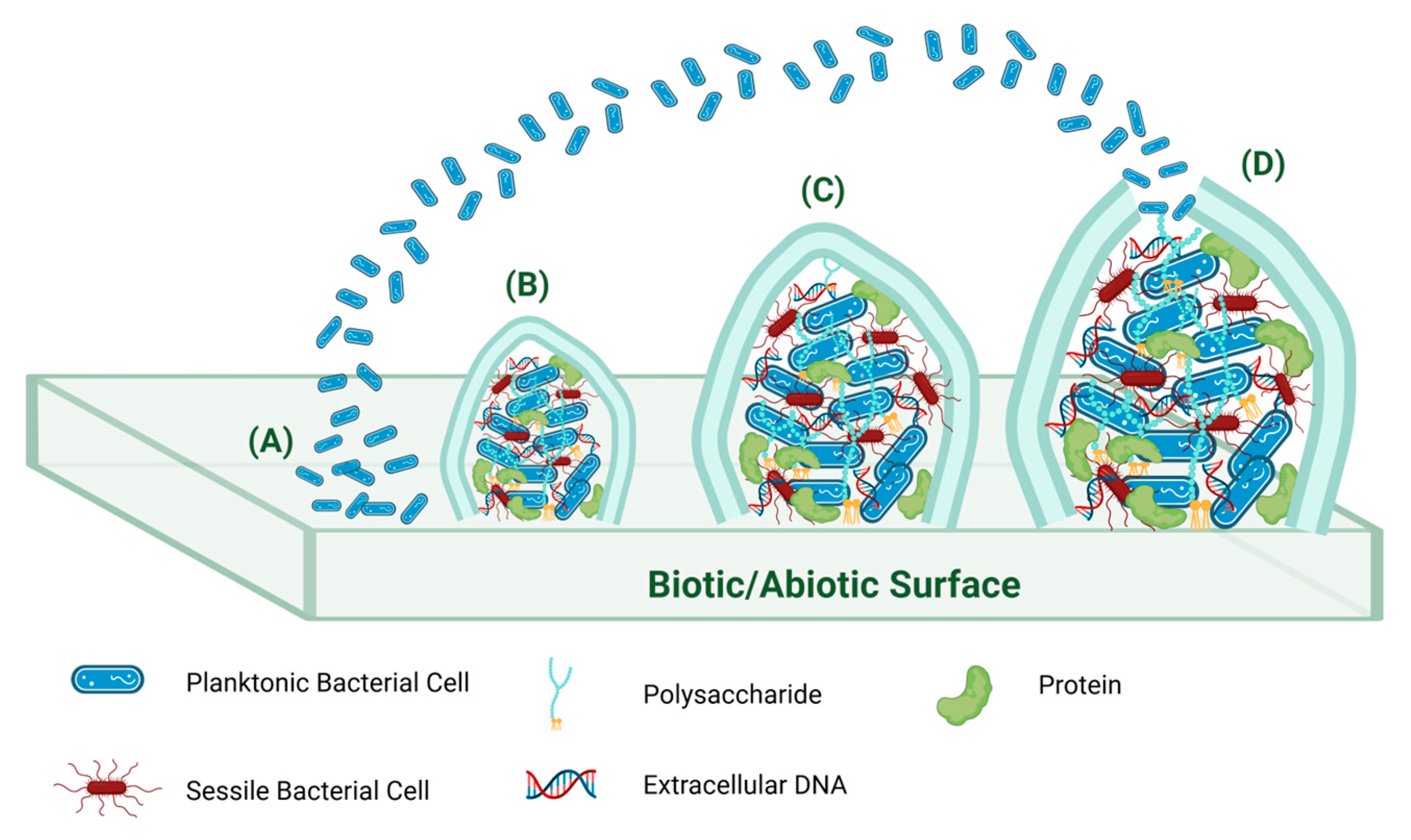
Figure 2.
Comparative data about the number of publications on antibiofilm compounds from marine endophytic fungi sources in Google Scholar from 2015 till 2024.
Figure 2.
Comparative data about the number of publications on antibiofilm compounds from marine endophytic fungi sources in Google Scholar from 2015 till 2024.
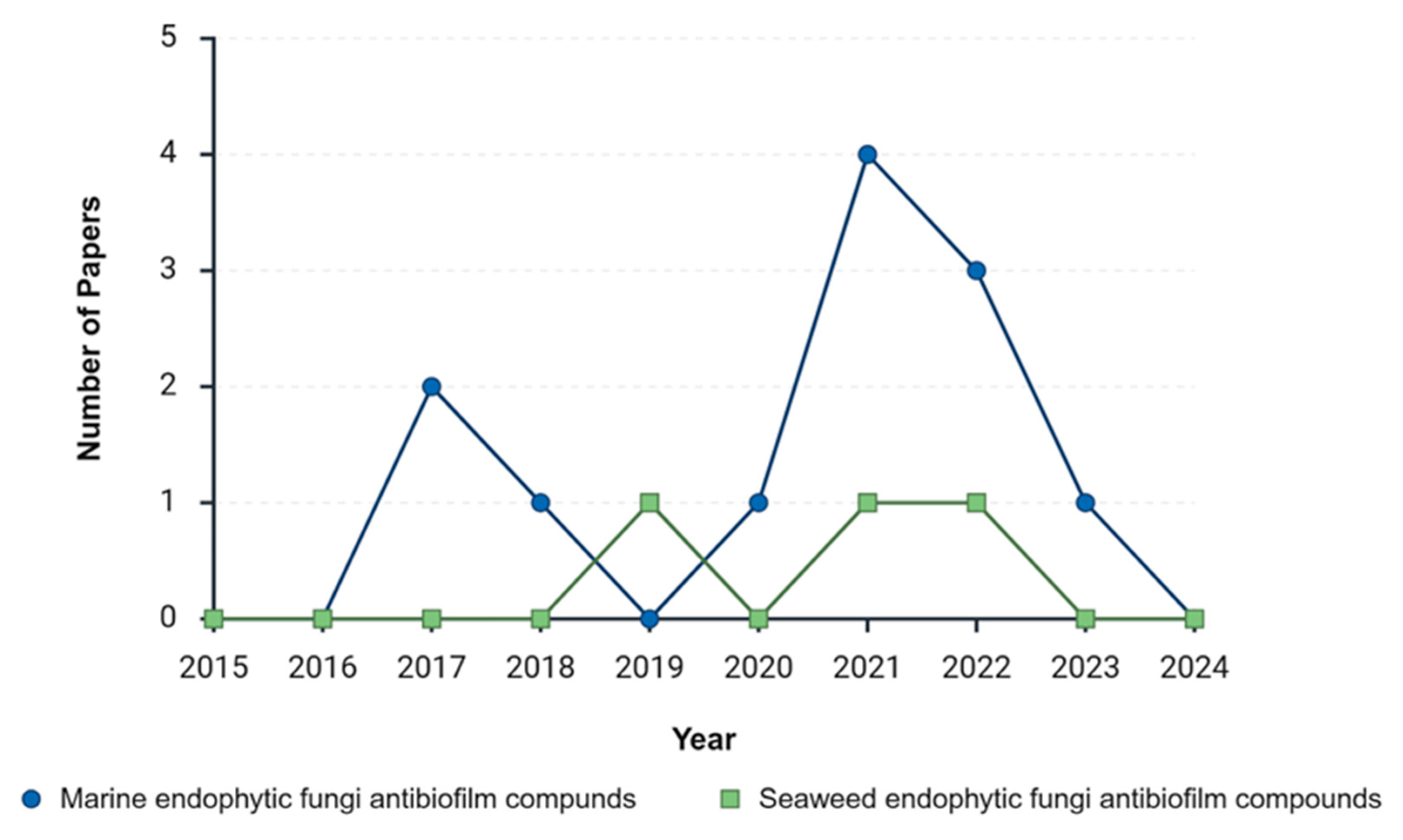
Figure 3.
Chemical structures of antibiofilm-active fungal secondary metabolites.
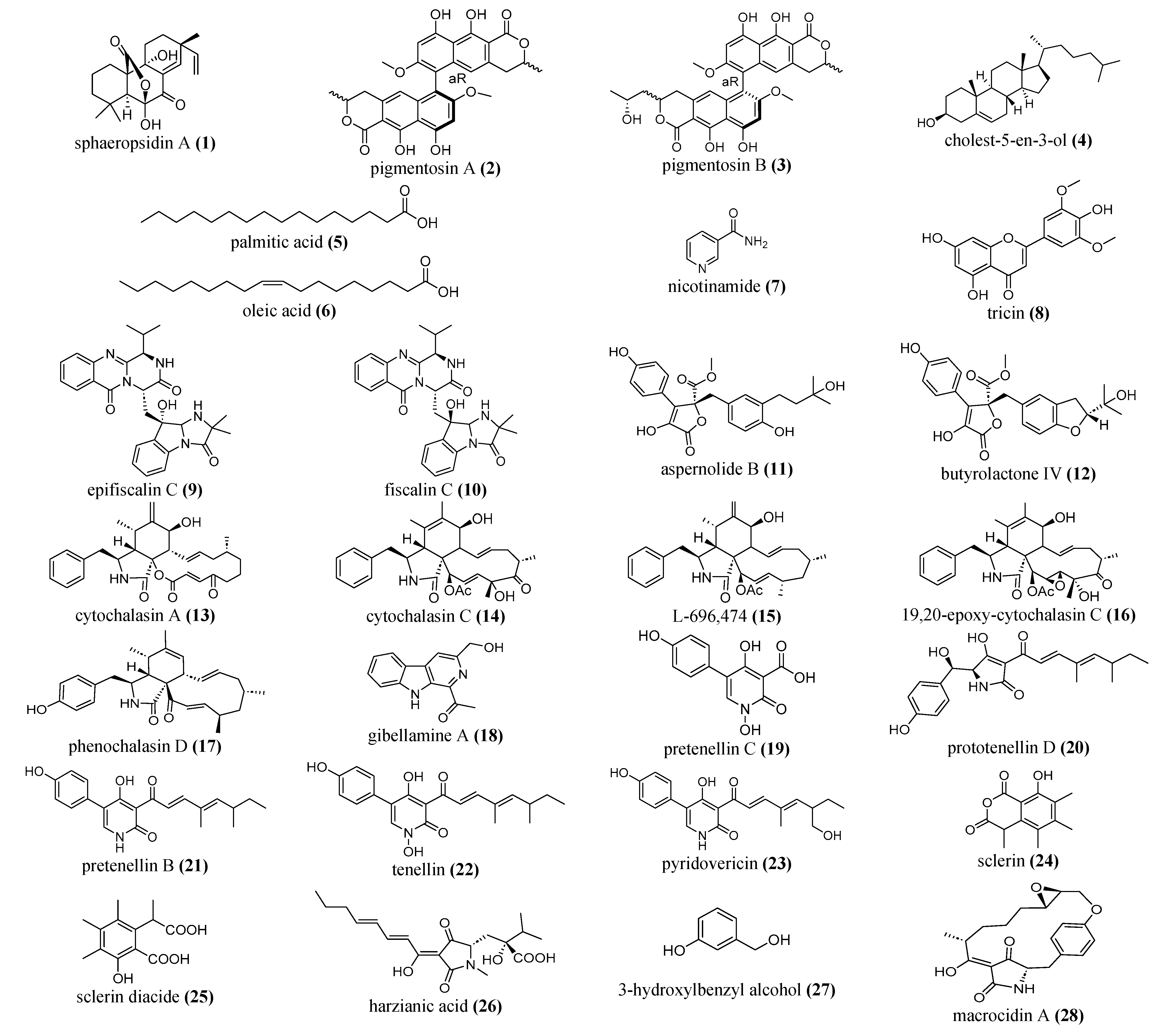
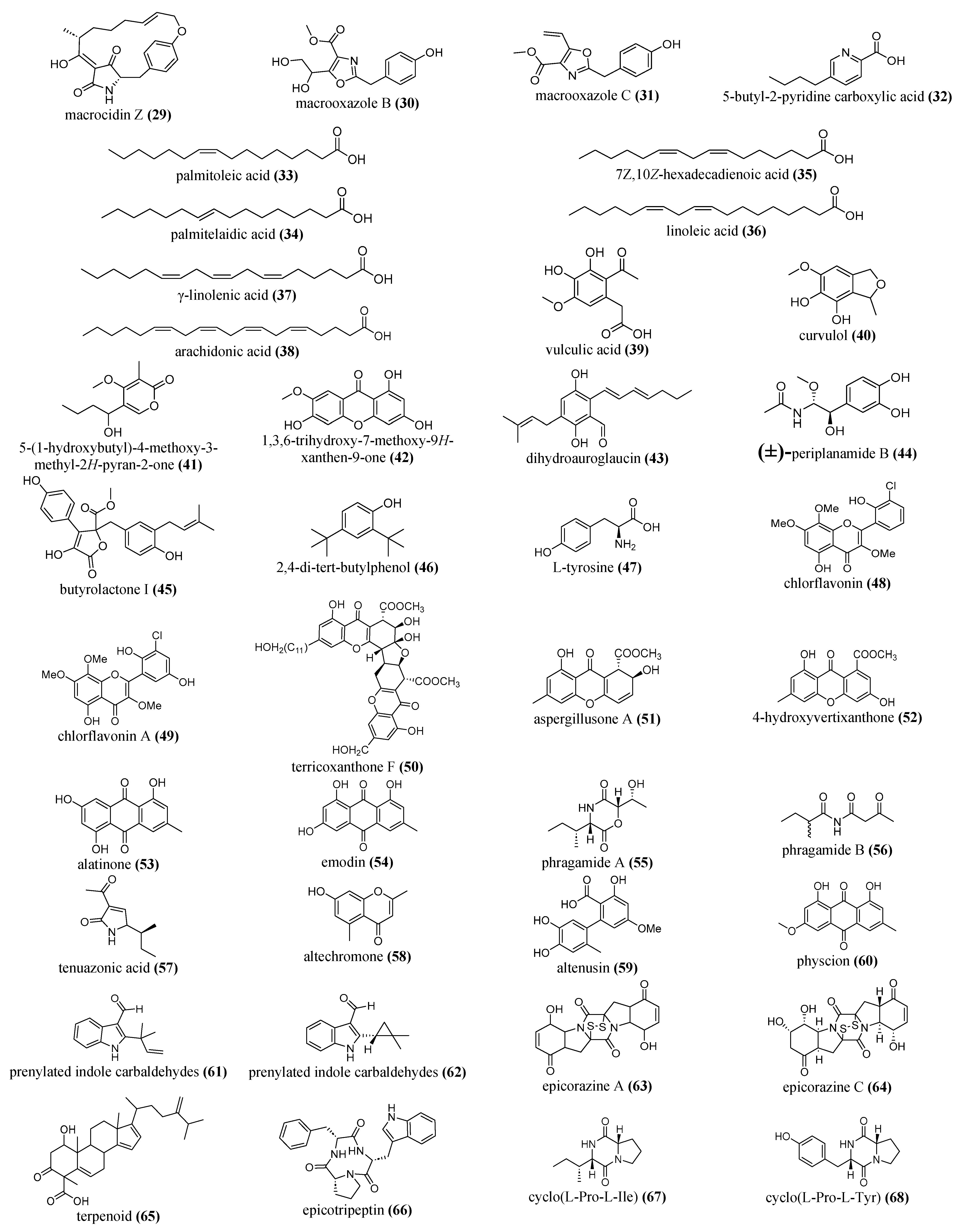
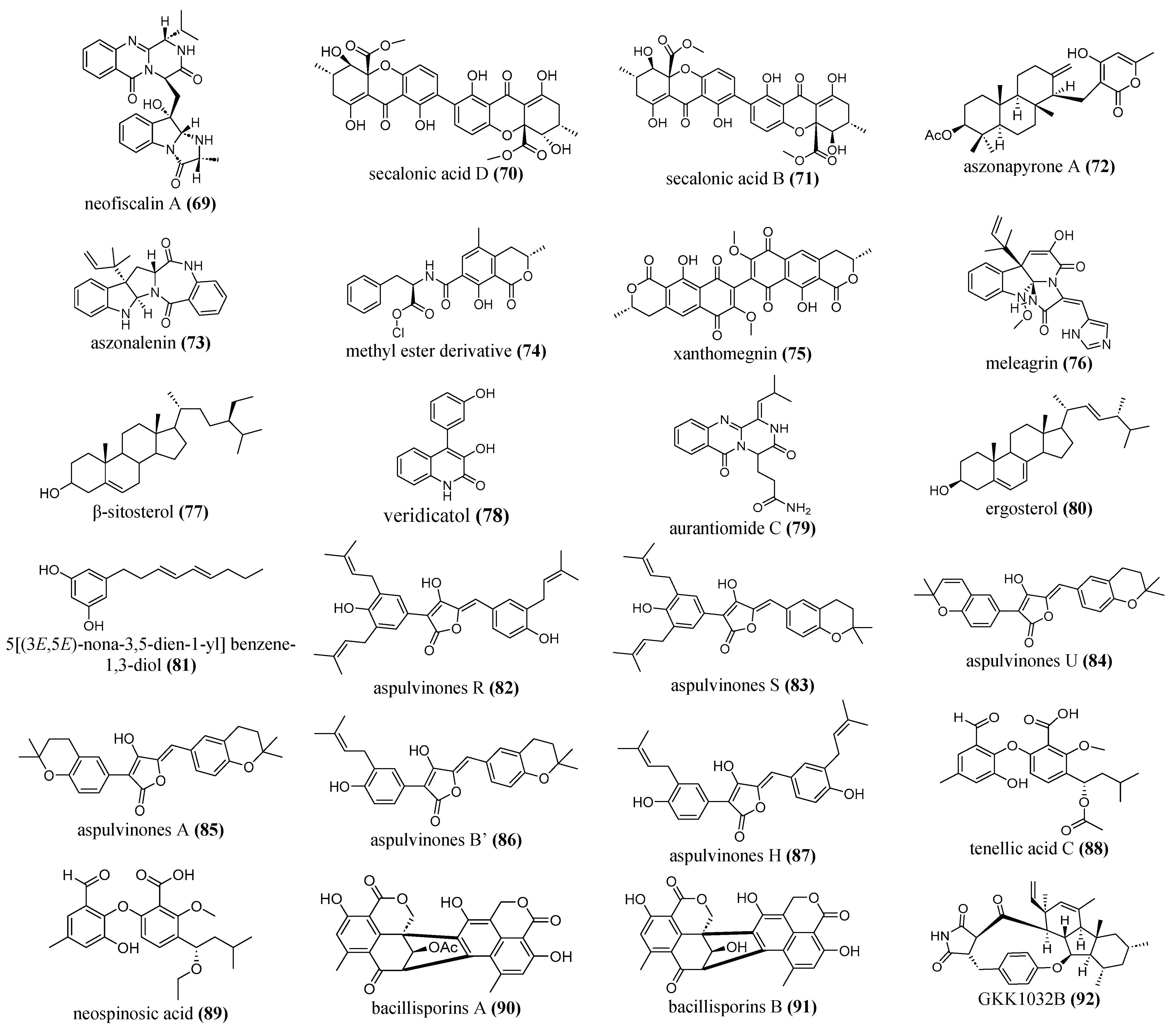
Table 1.
Number of papers obtained from Google Scholar (2015–2024) using specified search phrases before and after application of exclusion criteria.
Table 1.
Number of papers obtained from Google Scholar (2015–2024) using specified search phrases before and after application of exclusion criteria.
| Year | Marine endophytic fungi antibiofilm compounds | Seaweed endophytic fungi antibiofilm compounds | ||
| Before Exclusion | After Exclusion | Before Exclusion | After Exclusion | |
| 2015 | 46 | 0 | 7 | 0 |
| 2016 | 53 | 0 | 12 | 0 |
| 2017 | 83 | 2 | 30 | 0 |
| 2018 | 138 | 1 | 22 | 0 |
| 2019 | 182 | 0 | 56 | 1 |
| 2020 | 249 | 1 | 73 | 0 |
| 2021 | 447 | 4 | 140 | 1 |
| 2022 | 544 | 3 | 181 | 1 |
| 2023 | 846 | 1 | 246 | 0 |
| 2024 | 764 | 0 | 233 | 0 |
Exclusion criteria: conference abstracts, review papers, non-English language articles, papers that did not mention the search keyword or mentioned it only in the conclusion/discussion sections, and studies that did not use the keyword "endophytic" and did not describe a methodology compatible with isolating endophytic fungi or not pertaining to a host-associated fungus or fungal symbiont.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



