Submitted:
04 September 2025
Posted:
04 September 2025
You are already at the latest version
Abstract
Plant viral diseases result in significant agricultural losses globally. Unlike fungal or bacterial plant pathogens, viruses cannot be directly managed by chemicals. Their management mostly reliant on controlling insect vectors and breeding for host resistance. However, plants possess intrinsic molecular defense mechanisms that limit viral infections. RNA silencing pathways, particularly post-transcriptional gene silencing (PTGS) driven by small RNAs, play a crucial role in antiviral defense. Other epigenetic mechanisms, including RNA-directed DNA methylation (RdDM), N6-methyladenosine (m6A) RNA modifications, histone modifications and chromatin remodeling, also contribute to gene regulation during viral infection. DNA methylation patterns can be inherited across generations in a stable way, potentially conferring viral resistance to next generations. Virus-derived 24-nt small interfering RNAs (siRNAs) can function as mobile epigenetic signals, capable of inducing systemic gene silencing. A deeper understanding of epigenetic gene silencing mechanisms in virus-infected plants could improve biotechnological approaches for virus diagnosis, pathogenesis research and disease management. This review summarizes current insights into plant epigenetic gene silencing and explores their potential applications in the development of an antiviral defense system.
Keywords:
epigenetic gene silencing
; RNA-directed DNA methylation (RdDM)
; plant viral disease
; post-transcriptional gene silencing (PTGS)
; small interfering RNAs (siRNAs)
; host–virus interaction
1. Introduction
Plant pathogenic viruses are among the most destructive agents affecting crops. Every year, viruses damage crops worth about $30 billion (Tatineni & Hein, 2023). These losses threaten the crop productivity and long-term sustainability of agriculture. The scale of economic damage caused by plant viruses is well documented worldwide, including in Bangladesh (Table 1). The effective control of viruses is therefore necessary to mitigate economic losses and increase crop productivity. However, the conventional methods such insect vector control has yielded limited success in reducing viral transmission. Moreover, since vector control is mostly chemical based, it often leaves harmful residues in the surrounding ecosystem. To cope with the evolving nature of virus and to develop more sustainable defense mechanism, molecular approaches should be prioritized. Plants adapt to different biotic and abiotic stresses associated with changing environmental conditions. These responses involve epigenetic modifications such as DNA methylation, histone modifications, chromatin remodeling and RNA modifications that regulate cellular and molecular functions in plants (Dodds & Rathjen, 2010). Some of these modifications can directly affect transcription, thereby alter stress responsive gene expression. These modifications may persist through cell divisions and in some cases be transmitted across generations (Chinnusamy & Zhu, 2009; Zhu et al.; 2016).
Plant viruses can induce heritable epigenetic changes in host plant, thereby influencing long-term defense responses (Figure 1) (Norouzitallab et al.; 2019).Viruses reprogram host cellular machinery by targeting proteins and pathways involved in defense, development, and signaling. This reprogramming alters gene expression, disrupts cell cycle, and impairs small RNA-mediated defenses. Additionally, viruses modulate host defense by interfering with DNA methylation and microRNA biogenesis pathways (Raja et al.; 2008a). Viral infection can induce de novo or altered DNA methylation patterns in host plants (Baulcombe & Dean, 2014; Castellano et al.; 2016; Raja et al.; 2008).These changes provide a relatively stable mechanism to regulate gene expression, including resistance-related genes (Dowen et al.; 2012), thereby influencing host susceptibility (Boyko et al.; 2007; Sato et al.; 2017). In addition to DNA methylation, viruses alter histone modifications, which reshape chromatin structure and reprogram host gene expression (Ramirez-Prado et al.; 2018).
Virus-derived small RNAs (vsRNAs) activate epigenetic silencing pathways in plants that suppress viral replication and gene expression (Waterhouse et al.; 2001). This sRNA-mediated silencing is a distinctive feature of plant antiviral immunity and differ from the typical responses to bacterial or fungal pathogens (Stael et al.; 2015). Beyond antiviral defense, sRNAs also regulate plant development, highlighting their dual role in gene regulation (Borges & Martienssen, 2015; Matzke et al.; 2009)
Despite significant progress in molecular studies of epigenetic mechanisms, the role of plant epigenetic gene silencing in defense against pathogenic viruses and its potential application in disease management remains poorly understand. Bridging this gap could enable innovative and translatable strategies to increase crop resistance and reduce virus-related yield losses. This review summarizes advances in understanding plant epigenetic gene silencing in response to viral infection. We examine virus–host interactions that influence epigenetic pathways and consider their potential for improving crop protection. Such insights are vital for combating the growing threat of virus-induced crop losses worldwide.
2. Epigenetic Gene Silencing
Epigenetic gene silencing is a host defense mechanism that suppresses gene expression to maintain genome integrity (Figure 1) (Rajeevkumar et al.; 2015). It protects host genomes from viral infections and the activity of transposable elements (Liu et al.; 2023). Epigenetic silencing involves two interconnected pathways: transcriptional gene silencing (TGS), which suppresses transcription through DNA methylation and histone modifications and post-transcriptional gene silencing (PTGS), which degrades target RNA molecules (Rajeevkumar et al.; 2015).
2.1. Post Transcriptional Gene Silencing
Plants use post-transcriptional gene silencing (PTGS) as a sequence-specific defense mechanism against viral infection. PTGS targets viral mRNAs in the cytosol for degradation, as a result preventing the accumulation of viral proteins (Ramirez-Prado et al.; 2018). While infecting host, plant viruses initiate PTGS through double-stranded RNA (dsRNA). For RNA viruses, these dsRNAs are produced by replication of viral genome, whereas in DNA viruses, dsRNAs often originate from bidirectional transcription of viral genome (Gong et al.; 2022). RNA-dependent RNA polymerase-6 (RDR6) uses viral single strand RNA (ssRNA) as a template for synthesizing dsRNA. The dsRNA is then processed into 21- and 22-nucleotide siRNAs by DICER-LIKE proteins DCL4 and DCL2, respectively (Borges & Martienssen, 2015). These siRNAs mostly take part in PTGS; however, they can also induce low levels of de novo DNA methylation via AGO2 mediated via non-canonical TGS pathway (Wang et al.; 2019) (Figure 2). Subsequently, siRNAs are incorporated into RNA-Induced Silencing Complex (RISC) (Agrawal et al.; 2003; Baulcombe, 2004). The double stranded siRNA unwinds upon association with RISC and only one of the strands, usually guide (antisense) strand remains, forming the ‘activated RISC`. The activated RISC binds to complementary target mRNA to guide siRNA and ARGONAUTE 1(AGO1) initiates cleavage in the binding region, executing PTGS (Lee & Carroll, 2018; Martínez De Alba et al.; 2013). Remarkably, during epigenetic PTGS, viruses that initiate the process are eliminated but silencing in the host is maintained, providing a form of epigenetic memory (Jones et al.; 1999). A well-known example of PTGS is the transgenic papaya resistance system against papaya ringspot virus (PRSV). This system consists of a non-translated or inefficiently translated coat protein (CP) transgene, providing sequence-homology dependent resistance to the homologous PRSV strains (Hamim et al.; 2018; Tripathi et al.; 2004).
2.2. Transcriptional Gene Silencing
Transcriptional Gene Silencing (TGS) occurs via RNA-directed DNA methylation (RdDM) and associated histone modifications, targeting both plant or viral genomes. This epigenetic regulation takes place in the nucleus (Deleris et al.; 2016; Matzke et al.; 2009). RdDM is the major small RNA- mediated epigenetic pathway in plants. This can occur either by a canonical or a non-canonical pathway involving small RNA biogenesis (Matzke & Mosher, 2014).
Canonical RdDM makes up the majority of RdDM activity in plant cells (Cuerda-Gil & Slotkin, 2016a). Our understanding on canonical RdDM pathway mainly originates from the studies on the model plant Arabidopsis thaliana. In this pathway, two sequential steps are involved : (a) biogenesis of 24-nucleotide siRNAs (b) de novo methylation of DNA at target loci (Matzke et al.; 2015).
RNA Polymerase IV (Pol IV) directs the siRNA biogenesis by transcribing ssRNA at the target loci. RNA-DEPENDENT RNA POLYMERASE 2 (RDR2) then converts the ssRNA into dsRNA (Figure 2). The dsRNA is then processed by DICER-LIKE 3 (DCL3) into 24-nucleotide siRNAs, which are later stabilized by HUA ENHANCER 1(HEN1) (Wambui Mbichi et al.; 2020). The Pol IV-RDR2-DCL3 pathway is sufficient for the production of 24-nucleotide siRNAs in vitro, considering contributions from other associated factors (Singh et al.; 2019).
For de-novo methylation of DNA, a single strand from each 24-nucleotide siRNA is loaded into ARGONAUTE 4 (AGO4), forming an AGO4-siRNA duplex. This duplex then guided to RNA Polymerase V (Pol V) transcription sites where the siRNA base-pairs with complementary sequences on the Pol V-generated RNA scaffold. Subsequently AGO4 recruits DOMAINS REARRANGED METHYLTRANSFERASE 2 (DRM2), which methylates the nearby DNA and suppresses the target gene expression (M. A. Matzke & Mosher, 2014a; Wierzbicki et al.; 2009; H. Zhang et al.; 2018). Histone modifications including de-acetylation, de-methylation, and de-ubiquitination, further remodel chromatin at certain RdDM targets, achieving stable transcriptional gene silencing (Wambui Mbichi et al.; 2020).
Several non-canonical RdDM pathways have been identified which differs from the canonical pathways. In plants, these are often associated with antiviral silencing (Ding & Voinnet, 2007), and arise primarily as byproducts of PTGS. The main function of non-canonical pathway is to establish silencing by sparsely methylating a targeted region, which later gets taken over by the canonical pathway to establish long term silencing (Cuerda-Gil & Slotkin, 2016). Thus, the non-canonical pathway acts as a temporary bridge between PTGS and canonical RdDM pathway (Figure 2), targeting fewer genomic region compared to the canonical pathway (Erdmann & Picard, 2020).
3. Plant Genome Modifications and Epigenetic Silencing
3.1. Plant DNA
DNA methylation primarily occurring at cytosine residues, is key epigenetic silencing system controlling gene expression and maintaining genome stability in plants (Zhang et al.; 2006). Methyl groups from S-adenosyl-L-methionine (SAM) are added to the 5th carbon of cytosine, generating 5-methylcytosine (5meC) on target sequences, including host gene promoters (Deleris et al.; 2016b).This modification is central to TGS in plants and defense against various pathogens, including viruses (Choi et al.; 2020). This modification blocks transcription of adjacent genes, establishing TGS and restricting expression of viral or host genes (Dong et al.; 2018). In the canonical RdDM pathway dsRNA is processed by DCL3 into 24-nt siRNAs (Gallego-Bartolomé, 2020). These siRNAs bind with AGO4/AGO6 guiding methyltransferases to target loci to induce de novo methylation (Gallego-Bartolomé, 2020). In non-canonical pathway, DCL4 produces 21-22nt siRNA which can bind with AGO4 and lead to DNA methylation (El-Sappah et al.; 2021). DNA methylation occurs in symmetrical CG and CHG contexts and in asymmetric CHH contexts (where H = A, T, or C) (Lucibelli et al.; 2022). Symmetric CG methylation is maintained by MET1, while CHG methylation is catalyzed by CMT3. In contrast, CHH methylation relies on CMT2 and DRM2, which establish methylation de novo (El-Sappah et al.; 2021). The RdDM pathway is integral to plant defense against viral infections. For instance, AGO4 targets viral genomes for methylation, restricting replication of geminiviruses in Arabidopsis and crop plants (Velu thambi & Sunitha, 2021).
3.2. Plant RNA
Adenosine methylation at the N6 position (m6A) is the most prevalent internal RNA modification in plants, occurring in rRNAs, mRNAs, tRNAs, miRNAs, and long noncoding RNA (Alvarado-Marchena et al.; 2022). About 80% of RNA methylation in plant cells caused by m6A modifications (Kierzek, 2003). Early analyses show that m6A was found in wheat (Triticum turgidum L.), oat (Avena sativa L.) coleoptiles, and maize (Zea mays L.) ( (Haugland & Cline, 1980; Nichols & Welder, 1981; Shinde et al.; 2023). However, the molecular mechanisms regulating m6A methylation in plants remain incompletely understood (He et al.; 2023a). m6A modification is dynamically regulated by methyltransferases (writers), demethylases (erasers), and recognized by m6A binding proteins (readers) (He et al.; 2024). This modification plays a bidirectional role, it can inhibit viral RNA replication as part of the host antiviral defense, but in some cases, reversible m6A methylation may facilitate viral infection (Yue et al.; 2022). For example, when viral RNA enters the host plant, m6A methyltransferases such as plant METTL homologs catalyze methylation as part of a stress response. Conversely, in watermelon infected with cucumber green mottle mosaic virus, ALKBH9B demethylases are activated, reducing m6A and promoting viral invasion (He et al.; 2023; Hu et al.; 2022). YTH domain proteins act as readers, recognizing m6A-modified RNAs and mediating downstream responses during the interaction of methyltransferases and demethylases (Yue et al.; 2022)
3.3. Plant Histones
In plants, genomic DNA is packed into chromatin, with the nucleosome as its functional unit. Each nucleosome contains a core histone octamer, including two copies of four core histone proteins H2A, H2B, H3, and H4 and wrapped by around 147 bp DNA. Histone N-terminal tails undergo covalent modifications, such as methylation, acetylation, ubiquitination, phosphorylation, without altering the underlying DNA sequence (Kouzarides, 2007; Xie & Duan, 2023a). Key histone modifications involved in plant epigenetic gene silencing include H3K9, H3K2and H3K4 methylation. H3K4me3 is an activation mark that can function as a molecular memory, priming plant genes for expression (Corrêa et al.; 2024). H3K9me2 and H3K27me3 act as repressive marks, silencing target genes (Kang et al.; 2022). H3K9me2 cooperates with DNA methylation to silence viral genes. H3K9 methylation recruits CMT3 to maintain CHG methylation, while DNA methylation in turn recruits SUVH enzymes to sustain H3K9me2, establishing a reinforcing loop that stabilizes epigenetic silencing (Tariq & Paszkowski, 2004). Upon viral invasion, the Polycomb Repressive Complex 2 (PRC2) deposits H3K27me3 at specific loci, compacting chromatin and repressing transcription to promote long-term gene silencing. This maintenance is largely independent of DNA methylation or RNA interference pathways, highlighting a distinct regulatory mechanism during viral stress (Tao et al.; 2023).
4. Epigenetic Gene Silencing of Plant DNA Viruses
Plant DNA viruses, particularly members of the Geminiviridae family, are major pathogens causing substantial yield losses in crops such as cassava, cotton, tomato, pepper, beans, beet, tobacco, and maize (Pooggin, 2013; Zerbini et al.; 2017). These viruses carry small circular single-stranded DNA (ssDNA) genomes (2.5–5.2 kb) with a conserved common region (~200 bp) containing the origin of replication and promoters for bidirectional transcription. After entering the host cell, viral ssDNA is transported to the nucleus in complex with coat proteins (Figure 6) (Gupta et al.; 2021). Host enzymes convert ssDNA into double-stranded DNA (dsDNA), which assembles with histones to form viral minichromosomes that serve as templates for replication and transcription while interacting with host chromatin proteins (Hanley-Bowdoin et al.; 2013). In Nicotiana benthamiana infected with Abutilon mosaic virus (AbMV), Tomato yellow leaf curl Sardinia virus (TYLCSV), African cassava mosaic virus (ACMV), and Indian cassava mosaic virus (ICMV), minichromosomes carry 11–13 nucleosomes; transcriptionally inactive forms have 13 nucleosomes, whereas active forms have 11–12 (Coursey et al.; 2018; Paprotka et al.; 2011). Infected plants display leaf curling, mosaic patterns, vein swelling, and stunted growth (Arif et al.; 2020; Hanley-Bowdoin et al.; 2013).
Plants defend themselves against geminiviruses using DNA methylation-mediated epigenetic silencing. Transcriptional gene silencing (TGS), driven by RNA-directed DNA methylation (RdDM), deposits cytosine methylation in CG, CHG, and CHH contexts, blocking viral transcription (Noris & Catoni, 2020). Host factors such as ARGONAUTE4 (AGO4), DECREASE IN DNA METHYLATION 1 (DDM1), and Ty-1 are essential for this defense (Butterbach et al.; 2014; Raja et al.; 2008a). In Arabidopsis thaliana, DNA methylation mutants are highly susceptible to Cabbage leaf curl virus (CaLCuV) and Beet curly top virus (BCTV), highlighting the critical role of methylation (Raja et al.; 2008a). Similarly, in soybean, Mungbean yellow mosaic India virus (MYMIV) DNA exhibits high methylation in intergenic regions, with 24-nt siRNAs indicating active RdDM (Sahu et al.; 2010), while in pepper, Pepper golden mosaic virus (PepGMV) induces siRNAs that silence viral transcripts and promote recovery ((Rodríguez-Negrete et al.; 2013; Rodríguez-Negrete et al.; 2009). Even low methylation levels, such as 1–2.5% of cytosines in Tomato yellow leaf curl virus (TYLCV)-infected tomato, can slow viral replication and facilitate recovery Coursey et al.; 2018; Paprotka et al.; 2011; Piedra-Aguilera et al.; 2019)
Geminiviruses also promote active DNA demethylation to bypass host defenses. This process removes 5-methylcytosine from all sequence contexts (CG, CHG, CHH) on viral DNA. For instance, the βC1 protein of Tomato yellow leaf curl China betasatellite (TYLCCNB) interacts with ROS1-like DNA glycosylase in Nicotiana benthamiana and DEMETER (DME) in Arabidopsis thaliana, reversing methylation (Gui et al.; 2022a). ROS1 also negatively regulates RdDM, further weakening antiviral defenses (Yang et al.; 2018). Similarly, the C2 protein of Beet severe curly top virus (BSCTV) reduces methylation in promoter repeats of ACCELERATED CELL DEATH 6 (ACD6), increasing gene expression and lowering host resistance (Yang et al.; 2013).
Additional viral suppressors inhibit RNA silencing and methylation pathways. AC2/AL2 from Tomato golden mosaic virus (TGMV) interferes with adenosine kinase to reduce methylation-dependent defenses (Wang et al.; 2003), while V2 from TYLCV blocks SGS3, preventing siRNA amplification (Glick et al.; 2008). Rep initiates replication and disrupts host methylation machinery (Romero-Rodríguez et al.; 2023), βC1 interacts with AGO4 and DNA glycosylases to suppress RdDM (Gui et al.; 2022a; L. Yang et al.; 2022), and C4 interferes with hormone signaling, contributing to symptoms ((Mei et al.; 2020). In Bangladesh, papaya plants infected by begomoviruses such as Tomato leaf curl Joydebpur virus (ToLCJoV), Tomato leaf curl New Delhi virus (ToLCNDV), and Tomato leaf curl Bangladesh virus (ToLCBV) may trigger RdDM defenses, but viral proteins can limit their effectiveness (Gui et al.; 2022b; Hamim et al.; 2019, 2020). Similarly, Host shift of BBTV from banana to Heliconia in Hawaii is marked by low viral titers (Hamim et al.; 2017), likely due to partial restriction by RNA-directed DNA methylation (RdDM) and siRNA-mediated silencing, suggesting that epigenetic barriers play a key role in limiting its replication and host adaptation (Guyot et al.; 2022). Together, these factors allow DNA viruses to evade host defenses and establish systemic infections (Figure 6).
Plants counter these viral strategies through RNA silencing amplification and protein phosphorylation. The Ty-1 resistance gene in tomato encodes an RNA-dependent RNA polymerase that enhances cytosine methylation on viral DNA and amplifies viral-derived siRNAs (vsiRNAs), especially 21- and 22-nt siRNAs, improving viral RNA targeting and degradation even when viral suppressors like C2/TrAP and V2 are present ((Butterbach et al.; 2014; Ma et al.; 2024) (Figure 6). Additionally, the SUCROSE-NONFERMENTING1-related kinase (SnRK1) phosphorylates viral proteins to impair their function. βC1 is phosphorylated at serine-33 and threonine-78, reducing viral DNA and symptoms (Shen et al.; 2011; Tatineni & Hein, 2023). Rep of TGMV is phosphorylated at serine-97, weakening its DNA binding and viral replication (Shen et al.; 2018) and TrAP of CaLCuV is phosphorylated at serine-109, delaying infection (Shen et al.; 2014).
Therefore, plant defense against DNA viruses is multifaceted, combining RdDM-mediated TGS, active DNA demethylation counteraction, RNA silencing amplification, and phosphorylation of viral proteins. Geminiviruses employ suppressors and demethylases to overcome these defenses, establishing a dynamic host–virus arms race that determines disease outcome.
5. Epigenetic Gene Silencing of Plant RNA Viruses
Plant RNA viruses, including Turnip mosaic virus (TuMV), Cucumber mosaic virus (CMV), Tobacco mosaic virus (TMV), and Alfalfa mosaic virus (AMV), are significant pathogens with RNA genomes that cause substantial agricultural losses (Liu et al.; 2023). Both positive-strand (+) and negative-strand (−) RNA viruses are subject to epigenetic regulation through N6-methyladenosine (m6A) modification and small interfering RNA (siRNA)-mediated silencing, reflecting a dynamic molecular arms race between viral infection strategies and host defense mechanisms (Secco et al.; 2025). m6A is a reversible RNA modification present in both cellular and viral RNAs, catalyzed by methyltransferases (“writers”), removed by demethylases (“erasers”), and recognized by m6A-binding proteins (“readers”), collectively regulating RNA stability, translation efficiency, and decay (Li et al.; 2025) (Figure 7). Depending on the host-virus interaction, m6A can stabilize or destabilize viral RNAs, influencing replication and silencing. For example, TMV infection reduces m6A levels in Nicotiana tabacum, whereas AMV infection increases m6A levels in Arabidopsis thaliana. Inhibition of the Arabidopsis RNA demethylase ALKBH9B raises m6A levels on the AMV genome, enhancing siRNA-mediated antiviral responses and limiting systemic infection, demonstrating the antiviral role of m6A modification (Martínez-Pérez et al.; 2017; Yue et al.; 2022). Similarly, Pepino Mosaic Virus (PepMV) infection in N. benthamiana and S. lycopersicum modulates m6A levels on viral RNA, altering replication and movement, highlighting the interplay between RNA methylation and viral fitness (Secco et al.; 2025).
Concurrently, plants deploy siRNA-mediated gene silencing as a post-transcriptional defense against RNA viruses (Wang et al.; 2019). Long viral double-stranded RNAs (dsRNAs) are processed by Dicer-like (DCL) enzymes into 21–25 nucleotide siRNAs, which are unwound into guide and passenger strands. The guide strand is loaded into RNA-induced silencing complexes (RISC) containing Argonaute (AGO) proteins, while the passenger strand is degraded. Activated RISC complexes target complementary viral RNAs for cleavage or translational repression, effectively reducing viral replication (Akbar et al.; 2022). Viral infections such as CMV in Arabidopsis thaliana, Tobacco Rattle Virus (TRV), and Tomato Mosaic Virus (ToMV) exemplify this mechanism, generating viral siRNAs that suppress viral gene expression.
Viruses employ multiple strategies to overcome host defenses. Viral suppressors of RNA silencing (VSRs), such as the CMV 2b and TRV 16K proteins, bind and sequester host small RNAs, thereby disrupting RISC activity (Fernández-Calvino et al.; 2016; Zhao et al.; 2018), which leading to repression of defense gene expression. Similarly, Bean common mosaic virus (BCMV) manipulates host microRNA pathways to alter development and immunity (Martin et al.; 2016), while the non-coding Hop stunt viroid (HSVd) reduces repressive histone marks such as H3K9me2, reactivating silenced genes (Marquez-Molins et al.; 2024). Importantly, the success of PRSV-resistant transgenic papaya highlights the potential of harnessing epigenetic mechanisms for crop protection, where elevated siRNA accumulation confers stable and heritable resistance (Hamim et al.; 2018).
Collectively, these studies reveal the intricate interplay between m6A modifications, siRNA-mediated silencing, and viral countermeasures, emphasizing a complex molecular arms race and suggesting that targeted manipulation of these pathways could provide innovative strategies for enhancing crop resistance against RNA viruses (Figure 7).
6. Future Perspectives and Potential Applications of Epigenetic Gene Silencing in Plant Virus Management
Epigenetic gene silencing plays a role in plant defense against viruses and is being explored for its relevance in diagnostics, functional genomics, crop improvement, and disease management. Current studies on RNA- and DNA-based epigenetic mechanisms indicate that small RNA-mediated silencing, DNA and RNA methylation, and chromatin modifications may contribute to shaping management of plant–virus interactions.
6.1. Virus Detection and Diagnostic Applications
Virus-derived small interfering RNAs (vsiRNAs), generated as part of the host defense response, serve as reliable biomarkers for viral infection. High-throughput sequencing of these small RNAs, termed virus-derived small RNA profiling (vdSAR), allows detection of both known and novel viruses even in the absence of prior genomic information (Q. Wu et al.; 2010). While 24-nucleotide siRNAs are predominantly associated with DNA viruses via RNA-directed DNA methylation (RdDM), RNA virus infections produce mainly 21–22 nucleotide vsiRNAs, which can also be profiled to infer viral presence and host response (Matzke & Mosher, 2014; Raja et al.; 2008). Integrating small RNA sequencing with DNA methylation profiling enhances diagnostic sensitivity, although distinguishing viral siRNAs from host-derived RNAs and detecting low-abundance signals remain challenges (Massart et al.; 2019; Wang et al.; 2019). Advances in sequence technologies and bioinformatics are expected to overcome these limitations in the near future (Hamim et al.; 2022).
6.2. Functional Genomics Using Virus-Induced Gene Silencing
Epigenetic gene silencing, especially via virus-induced gene silencing (VIGS), continues to serve as a useful functional genomics approach in plants (Zulfiqar et al.; 2023). Plant RNA viruses such as PVX (Potato Virus X), TRV (Tobacco Rattle Virus), and CMV (Cucumber Mosaic Virus) are commonly used as vectors in functional genomics. These viruses can induce transcriptional or post-transcriptional gene silencing of reporter transgenes, including GFP and GUS, facilitating functional studies directly in living plants (Zulfiqar et al.; 2023). Recent advancements have optimized VIGS protocols across diverse species, for instance, VIGS has been successfully applied in sunflower using TRV, and in tomato to study ripening-related genes via silencing of a specific methyltransferase (Mardini et al.; 2024; Xiong et al.; 2024). Recent methylome analyses indicate that viral infections can cause epigenetic modifications, including changes in DNA methylation, histone modifications, and chromatin structure, which are associated with the regulation of defense- and stress-related genes (Kim, 2021; Xie & Duan, 2023). Sequencing technologies, such as enzymatic methyl-seq, allow single-base resolution mapping of these changes, providing information that may help guide research toward virus-resistant cultivars (Yue et al.; 2022).
6.3. Engineering Viral Resistance Through Epigenetic Modifications
Epigenetic modifications offer a promising route for engineering durable viral resistance (Zhi & Chang, 2021). Some DNA methylation sites and histone changes help plants turn on antiviral genes( Wu & Fan, 2025), but viral proteins like AL2 and L2 from Beet Curly Top Virus (BCTV) can block these defenses (Jackel et al.; 2015). By studying these processes, scientists can use epigenetic tools to boost plant immunity and develop viral-resistant crops. Emerging tools such as long-read sequencing and CRISPR/Cas9-based epigenome editing enable precise analysis and targeted manipulation of host epigenetic marks, providing opportunities to enhance plant immunity and generate heritable viral resistance (Hamim et al.; 2022).
6.4. Exogenous Epigenetic-Based Virucides
The exogenous application of double-stranded RNA (dsRNA) or small interfering RNAs (siRNAs) offers a novel, environmentally friendly strategy to control viral infections (Vatanparast et al.; 2024). Topical dsRNA treatments targeting viral genomes reduce viral accumulation and symptoms in crops infected with Cucumber Green Mottle Mosaic Virus (CGMMV) and Potato Virus X/Y (Dubrovina & Kiselev, 2019). Recent studies demonstrate that high-pressure dsRNA spraying can induce transcriptional gene silencing via RdDM, a phenomenon termed spray-induced epigenetic modifications(Dalakouras & Ganopoulos, 2021). For example, 24-nt siRNA applications under high pressure induced methylation of the Cauliflower Mosaic Virus (CaMV) 35S promoter in Nicotiana benthamiana, leading to transcriptional silencing of viral genes (Dalakouras & Ganopoulos, 2021). These strategies provide a foundation for sequence-specific antiviral treatments and virucide development.
Collectively, epigenetic gene silencing provides versatile tools for plant-virus management. From sensitive diagnostics and functional genomics to engineering durable resistance and RNA-based virucides, these approaches offer sustainable and precise strategies to enhance plant immunity. Integrating small RNA profiling, methylome mapping, long-read sequencing, and targeted epigenome editing will advance our understanding of plant-virus interactions and facilitate the development of crops with heritable resistance to viral diseases.
Figure 3.
DNA methylation (red circles) occurring in symmetric CG and CHG contexts, as well as asymmetric CHH contexts (where H = A, T, or C). DNA methyltransferases are recruited to specific sequences: MET1 maintains CG methylation, CMT3 catalyzes CHG methylation, while CMT2 and DRM2 establish CHH methylation de novo. The red circles in the figure represent methylated cytosines at these target sites.
Figure 3.
DNA methylation (red circles) occurring in symmetric CG and CHG contexts, as well as asymmetric CHH contexts (where H = A, T, or C). DNA methyltransferases are recruited to specific sequences: MET1 maintains CG methylation, CMT3 catalyzes CHG methylation, while CMT2 and DRM2 establish CHH methylation de novo. The red circles in the figure represent methylated cytosines at these target sites.
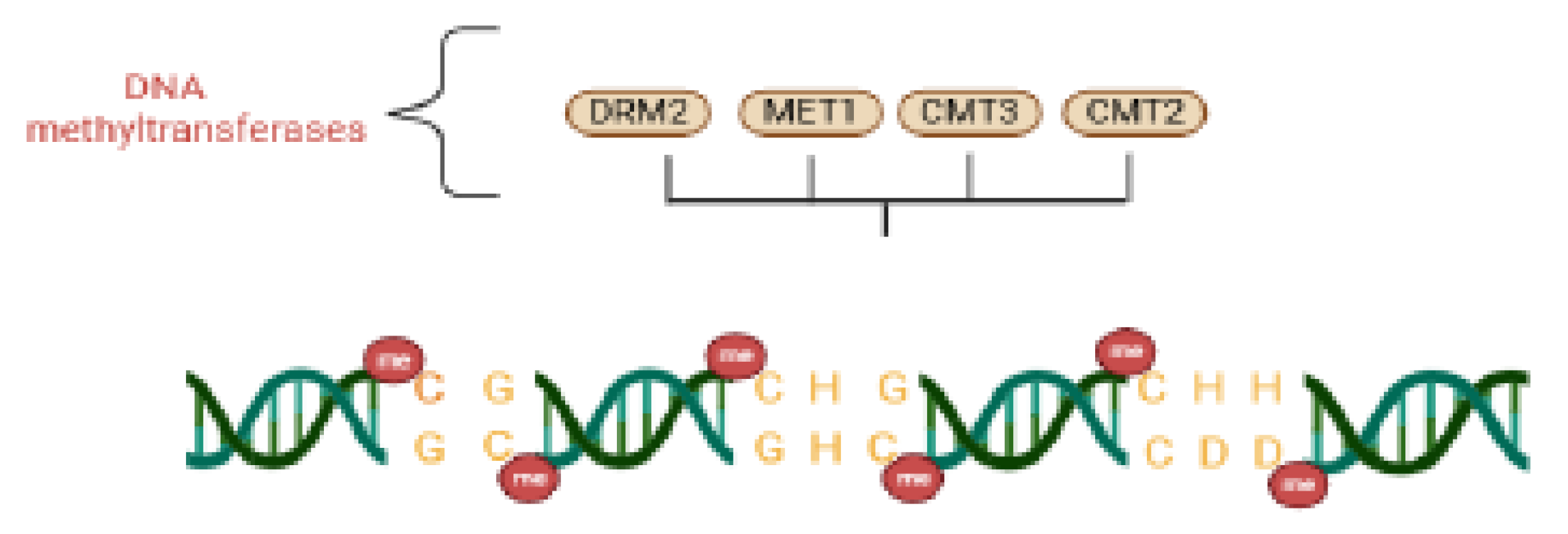
Figure 4.
The dynamic regulation of plant viral RNA by m6A modification. Viral RNA entering the host is methylated by m6A methyltransferases (‘writers’), adding m6A marks. These marks can be removed by m6A demethylases such as ALKBH9B (‘erasers’), reversing methylation. YTH domain proteins act as ‘readers,’ recognizing m6A-modified RNA and mediating downstream responses during host-virus interactions.
Figure 4.
The dynamic regulation of plant viral RNA by m6A modification. Viral RNA entering the host is methylated by m6A methyltransferases (‘writers’), adding m6A marks. These marks can be removed by m6A demethylases such as ALKBH9B (‘erasers’), reversing methylation. YTH domain proteins act as ‘readers,’ recognizing m6A-modified RNA and mediating downstream responses during host-virus interactions.
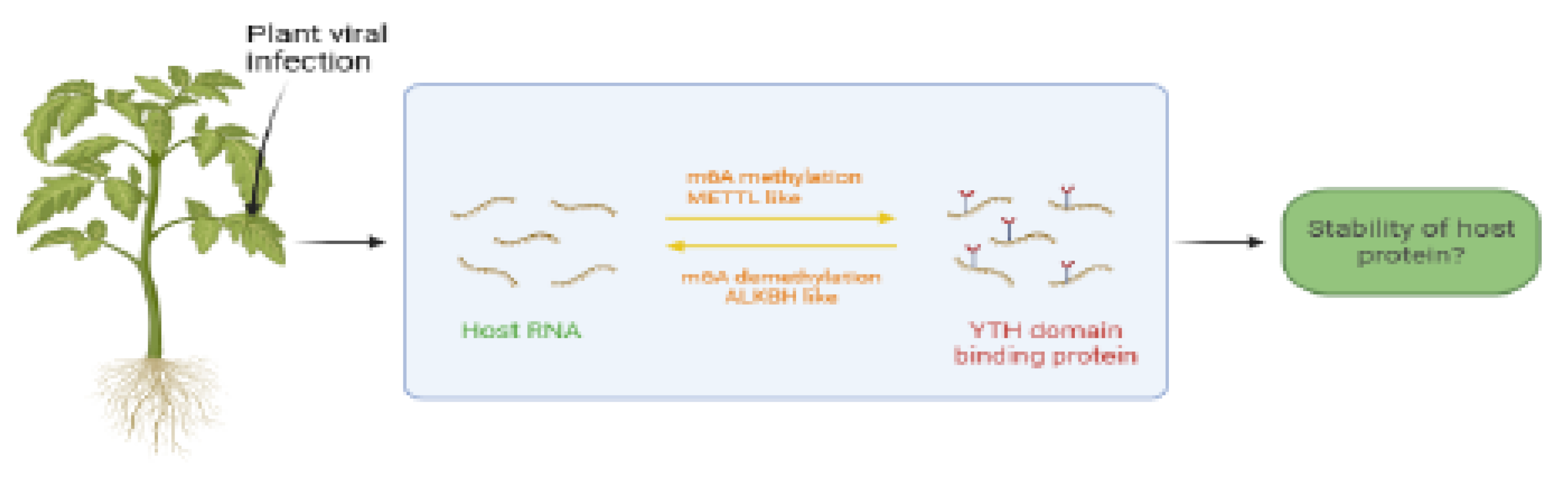
Figure 5.
Histone-mediated epigenetic mechanisms in plants. H3K4me2 marks contribute to epigenetic memory and can prime genes for transcriptional regulation. H3K9me2 collaborates with DNA methylation in a reinforcing loop: H3K9me2 recruits CMT3 to maintain CHG methylation, while DNA methylation recruits SUVH enzymes to sustain H3K9me2, collectively reinforcing gene silencing. Additionally, the Polycomb Repressive Complex 2 (PRC2) deposits H3K27me3 at specific loci, compacting chromatin and promoting long-term transcriptional repression.
Figure 5.
Histone-mediated epigenetic mechanisms in plants. H3K4me2 marks contribute to epigenetic memory and can prime genes for transcriptional regulation. H3K9me2 collaborates with DNA methylation in a reinforcing loop: H3K9me2 recruits CMT3 to maintain CHG methylation, while DNA methylation recruits SUVH enzymes to sustain H3K9me2, collectively reinforcing gene silencing. Additionally, the Polycomb Repressive Complex 2 (PRC2) deposits H3K27me3 at specific loci, compacting chromatin and promoting long-term transcriptional repression.
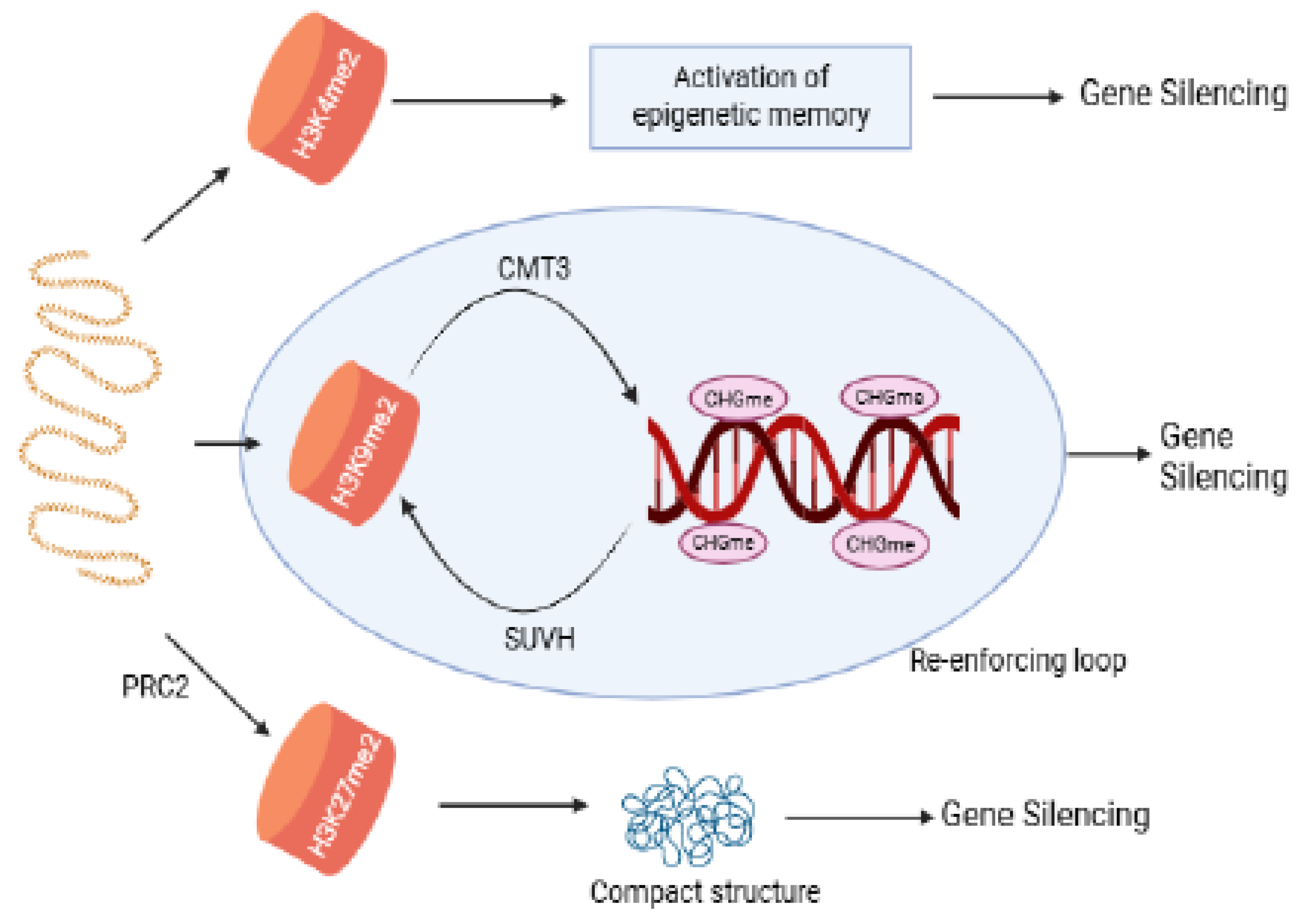
Figure 6.
Schematic of geminivirus interactions with plant epigenetic defenses and viral evasion strategies. After entering the plant cell, viral single-stranded DNA (ssDNA) is targeted by post-transcriptional gene silencing (PTGS) in the cytoplasm and transcriptional gene silencing (TGS) in the nucleus. In the nucleus, viral proteins V2 and βC1 interfere with AGO4, blocking RdDM-guided DNA methylation and promoting viral accumulation. Additional suppressors, including AC2/AL2, AC4/C4, Rep, and AC5, disrupt TGS by inhibiting key components of the methylation machinery. In the cytoplasm, βC1 degrades SGS3, impairing RDR6-dependent amplification of secondary siRNAs, while C4 also contributes to reduced siRNA accumulation. Furthermore, active DNA demethylation mediated by recruitment of host DNA glycosylases reverses methylation marks on viral genomes, enhancing replication and virulence.
Figure 6.
Schematic of geminivirus interactions with plant epigenetic defenses and viral evasion strategies. After entering the plant cell, viral single-stranded DNA (ssDNA) is targeted by post-transcriptional gene silencing (PTGS) in the cytoplasm and transcriptional gene silencing (TGS) in the nucleus. In the nucleus, viral proteins V2 and βC1 interfere with AGO4, blocking RdDM-guided DNA methylation and promoting viral accumulation. Additional suppressors, including AC2/AL2, AC4/C4, Rep, and AC5, disrupt TGS by inhibiting key components of the methylation machinery. In the cytoplasm, βC1 degrades SGS3, impairing RDR6-dependent amplification of secondary siRNAs, while C4 also contributes to reduced siRNA accumulation. Furthermore, active DNA demethylation mediated by recruitment of host DNA glycosylases reverses methylation marks on viral genomes, enhancing replication and virulence.
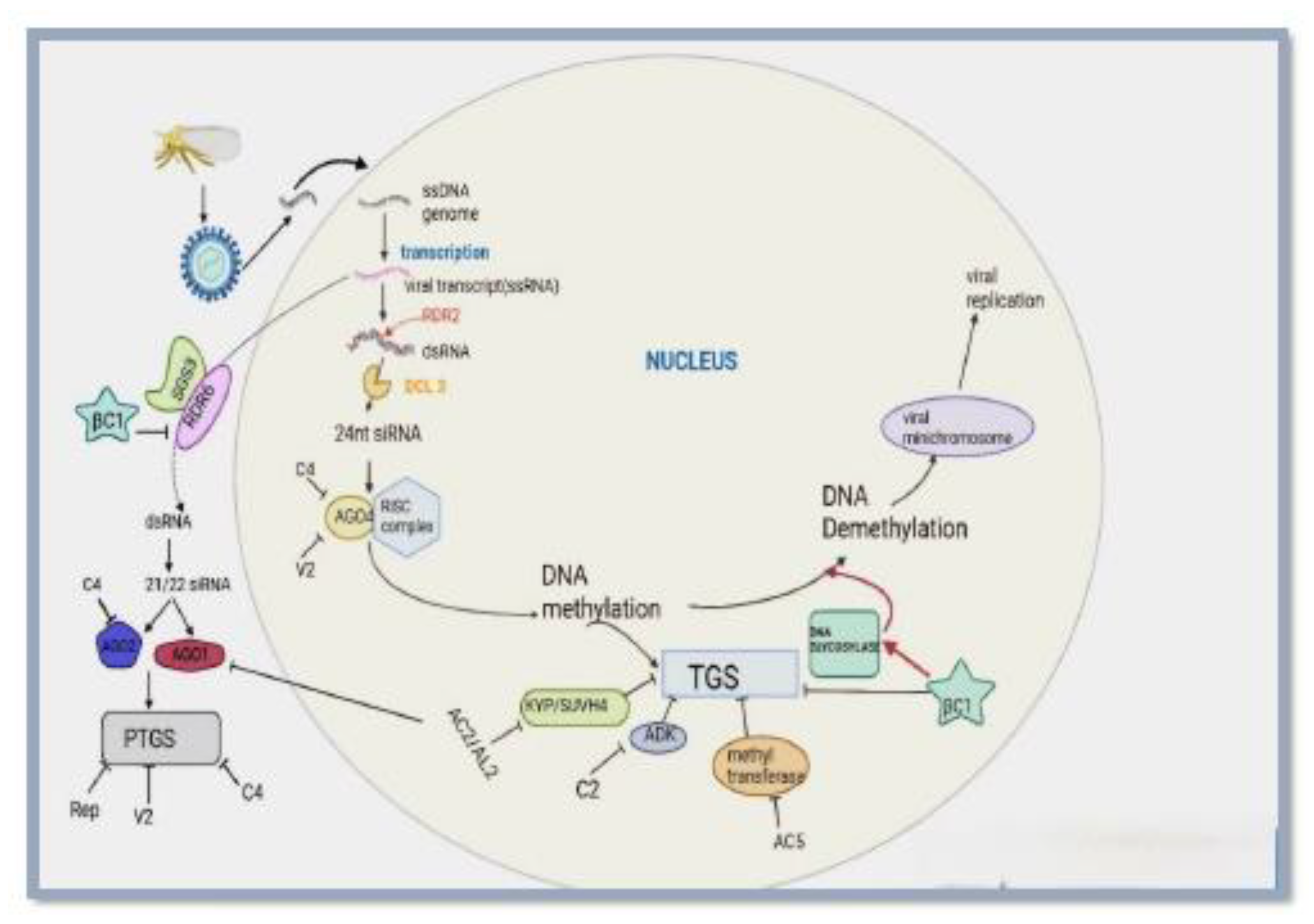
Figure 7.
Schematic representation of plant epigenetic defense against RNA viruses and viral counter-strategies. (A) m6A writers catalyze the addition of a methyl group to adenosine residues on viral or host mRNAs, creating an m6A mark. m6A readers recognize this modification, leading to targeted RNA degradation and suppression of viral replication. (B) m6A erasers remove methyl groups from mRNAs, allowing translation and replication of viral RNAs, production of viral proteins, and successful infection of plant cells. (C) RNA interference pathway: single-stranded viral RNA (ssRNA) is converted into double-stranded RNA (dsRNA), which is processed by Dicer-like (DCL) enzymes into small interfering RNAs (siRNAs). siRNAs are loaded into the RNA-induced silencing complex (RISC), where the guide strand is selected to recognize complementary viral mRNAs, resulting in cleavage, degradation, and epigenetic gene silencing.
Figure 7.
Schematic representation of plant epigenetic defense against RNA viruses and viral counter-strategies. (A) m6A writers catalyze the addition of a methyl group to adenosine residues on viral or host mRNAs, creating an m6A mark. m6A readers recognize this modification, leading to targeted RNA degradation and suppression of viral replication. (B) m6A erasers remove methyl groups from mRNAs, allowing translation and replication of viral RNAs, production of viral proteins, and successful infection of plant cells. (C) RNA interference pathway: single-stranded viral RNA (ssRNA) is converted into double-stranded RNA (dsRNA), which is processed by Dicer-like (DCL) enzymes into small interfering RNAs (siRNAs). siRNAs are loaded into the RNA-induced silencing complex (RISC), where the guide strand is selected to recognize complementary viral mRNAs, resulting in cleavage, degradation, and epigenetic gene silencing.
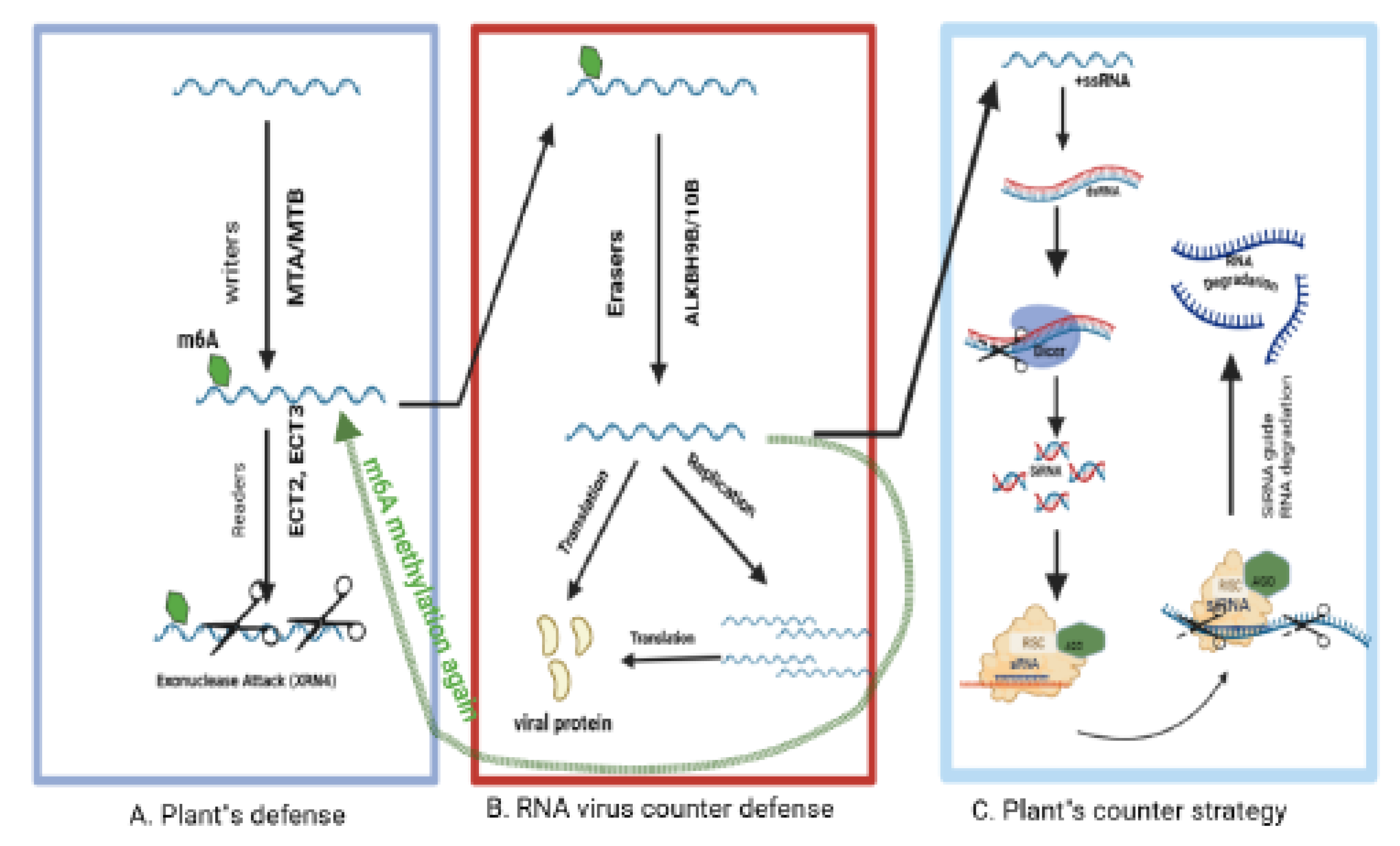
Figure 8.
dsRNA and also siRNA when applied on virus infected plant by different approaches such as infiltration, spray, and microinjection. dsRNA along with DCL then attach with RISC and they trigger transcriptional gene silencing via RdDM pathway of plant viruses and the plant have no symptoms of virus infection. While siRNA bypass the DCL then directly bind with RISC and cause the epigenetic gene silencing.
Figure 8.
dsRNA and also siRNA when applied on virus infected plant by different approaches such as infiltration, spray, and microinjection. dsRNA along with DCL then attach with RISC and they trigger transcriptional gene silencing via RdDM pathway of plant viruses and the plant have no symptoms of virus infection. While siRNA bypass the DCL then directly bind with RISC and cause the epigenetic gene silencing.
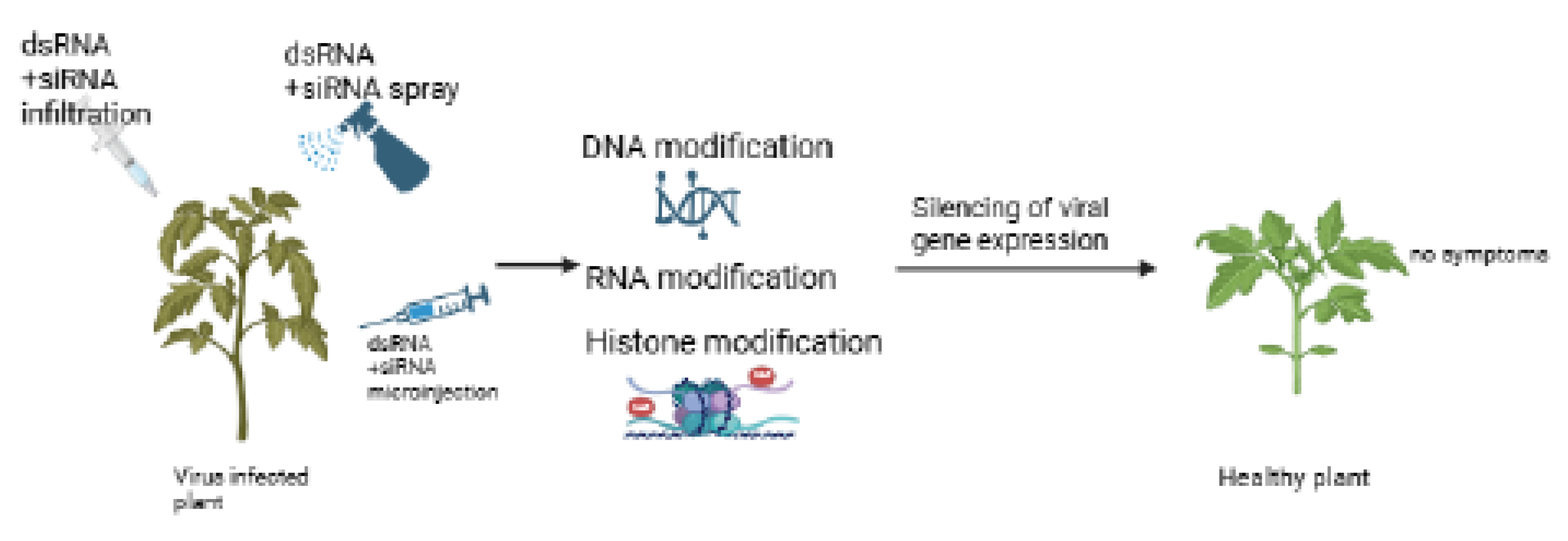

Table 2.
Key components of PTGS and RdDM pathways in plants and their roles in antiviral defense.
| Molecular Component | Category/Type | Associated Pathway (PTGS/RdDM) | Primary Function | Key References |
|---|---|---|---|---|
| DCL2, DCL4 | Dicer-like RNase III enzymes | PTGS (DCL4/DCL2); RdDM (DCL2) | DCL4 makes 21-nt siRNAs for PTGS; DCL2 makes 22-nt siRNAs for antiviral silencing and non-canonical RdDM | Erdmann & Picard, 2020; Jin et al.; 2022; Wambui Mbichi et al.; 2020 |
| DCL3 | Dicer-like RNase III enzyme | Canonical RdDM | Processes dsRNA into 24-nt siRNAs that guide DNA methylation in the RdDM pathway | Matzke & Mosher, 2014; Stroud et al.; 2013 |
| RDR6 | RNA dependent RNA polymerase | PTGS, Non-canonical RdDM | Converts single-stranded RNAs to double stranded RNAs for processing into21–22nt siRNAs by DCL1, DCL2and DCL4 | Matzke et al.; 2015 |
| RDR2 | RNA dependent RNA polymerase | Canonical RdDM | Converts Pol IV–derived ssRNAs into dsRNAs, which are processed by DCL3 into 24-nt siRNAs. | Blevins et al.; 2015; Matzke et al.; 2015 |
| AGO1 | Argonaut protein | PTGS | Forms RISC with 21–22 nt siRNAs to recognize and cleave complementary mRNAs. | Fang & Qi, 2016; Matzke et al.; 2015; Voinnet, 2008 |
| AGO2 | Argonaute protein | Non canonical RdDM | Loads 21–22 nt siRNAs to target Pol V transcripts and facilitates DRM2 recruitment for DNA methylation. | Erdmann & Picard, 2020 |
| AGO4 | Argonaute protein | Canonical RdDM | Loads 24-nt siRNAs to target Pol V transcripts and recruits DRM2 for DNA methylation. | Erdmann & Picard, 2020 |
| NERD | Plant-specific protein (PHD and zinc-finger domains) | Non-canonical RdDM | Interacts with histone H3 and AGO2–siRNA complexes to promote histone modification and transcriptional repression. | Matzke & Mosher, 2014 |
| DRM 2 | DNA methyltransferase | Canonical & non-canonical RdDM | Catalyzes de novo cytosine DNA methylation guided by AGO–siRNA complexes. | Matzke et al.; 2015 |
| HEN1 | RNA methyltransferase | PTGS, All RdDM | Adds a 2′-O-methyl group to the 3′ end of siRNAs, protecting them from degradation | Yang et al.; 2018 |
Table 3.
Gemiviral protein-mediated suppression of host defense mechanism.
| Viral Protein | Virus | Host Target | Effect on Epigenetic Gene Silencing | References |
|---|---|---|---|---|
| Rep (Replication-associated protein) | Tomato yellow leaf curl Sardinia virus (TYLCSV) | MET1, CMT3 | Reduces maintenance DNA methylation (CG context), weakening transcriptional gene silencing (TGS) | Rodríguez-Negrete et al.; 2013 |
| AC2 | Tomato golden mosaic virus (TGMV), Cabbage leaf curl virus (CaLCuV) | SUVH4/KYP (H3K9 histone methyltransferase) | Inhibits histone methylation, disrupting chromatin-based TGS | Veluthambi & Sunitha, 2021 |
| C2 | Beet severe curly top virus (BSCTV) | SAMDC1 (S-adenosyl methionine decarboxylase) | Lowers methyl donor availability, reducing DNA and histone methylation for epigenetic silencing | Zhang et al.; 2011 |
| C4 | Tomato leaf curl Yunnan virus (TLCYnV) | DRM2 (Domain Rearranged Methyltransferase 2) | Prevents de novo cytosine methylation on viral DNA, impairing RdDM-mediated TGS | Mei et al.; 2020 |
| TrAP | TGMV, BCTV | ADK (Adenosine Kinase) | Disrupts SAM biosynthesis, interfering with methylation-mediated TGS | Jackel et al.; 2015 |
| V2 | TYLCV, Cotton leaf curl Multan virus (CLCuMuV) | AGO4 | Blocks AGO4 binding to viral DNA, inhibiting RdDM and preventing transcriptional silencing | Wang et al.; 2019 |
| Pre-coat Protein | TYLCV, ToLCNDV | MET1, RDR1, HDA6 | Suppresses maintenance methylation and chromatin silencing, compromising TGS | Basu et al.; 2018; Wang et al.; 2018 |
| C4 | CLCuMuV, ToYLCGDV | SAM synthetase, BAM1 | Reduces SAM availability and inhibits TGS; disrupts epigenetic regulation of defense genes | Ismayil et al.; 2018; Li et al.; 2020; Soto-Burgos & Bassham, 2017 |
| AC5 | MYMIV | CHH cytosine methyltransferase | Suppresses RNA-induced PTGS and reverses TGS of silenced transgenes, impairing epigenetic silencing | Li et al.; 2015 |
| βC1 | Betasatellite of TYLCCNV | SAHH (S-adenosyl homocysteine hydrolase) | Disrupts methyl cycle, suppresses methylation-dependent PTGS and RdDM-mediated TGS via calmodulin-like protein (CaM) | Yang et al.; 2011; Li et al.; 2017 |
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Declaration of competing interest
All authors declare that they do not have any competing financial, professional, or non-financial interests that could be construed as influencing the interpretation of data or the presentation of information de-scribed in this manuscript.
Author Contributions
Islam Hamim and John Hu conceived and designed the review. Islam Hamim wrote the manuscript. Islam Hamim, Sadman Jawad Sakib, Md. Readoy Hossain, Jaima Noor Hia, Alvi Al Muhimine, and Maria Hasan collected and analyzed the literature and constructed the figures. Islam Hamim and John Hu edited the manuscript. All authors read and revised the manuscript.
Acknowledgements
This review was funded by the United States Agency for International Development (USAID) as part of the Feed the Future initiative under the CGIAR Fund, award number BFS-G-11-00002, and the predecessor fund of the Food Security and Crisis Mitigation II grant, award number EEM-G-0004-00013. The research was also supported by the USDA National Institute of Food and Agriculture, Hatch HAW09025-H (1001478), and the USDA Agricultural Research Service (58-5320-4-012). During the editing of the manuscript, Islam Hamim was supported through a Georg Forster fellowship from the Alexander von Humboldt Foundation. We acknowledge BioRender.com for providing the platform to create scientific figures used in this manuscript.
Conflicts of Interest
All authors declare no conflict of interest.
References
- Abo, M.E.; Sy, A.A. Rice Virus Diseases: Epidemiology and Management Strategies. Journal of Sustainable Agriculture 1997, 11, 113–134. [Google Scholar] [CrossRef]
- Agrawal, N.; Dasaradhi, P.V.N.; Mohmmed, A.; Malhotra, P.; Bhatnagar, R.K.; Mukherjee, S.K. RNA Interference: Biology, Mechanism, and Applications. Microbiology and Molecular Biology Reviews 2003, 67, 657–685. [Google Scholar] [CrossRef]
- Akbar, S.; Wei, Y.; Zhang, M.-Q. RNA Interference: Promising Approach to Combat Plant Viruses. International Journal of Molecular Sciences 2022, 23, 5312. [Google Scholar] [CrossRef]
- Alvarado-Marchena, L.; Martínez-Pérez, M.; Úbeda, J.R.; Pallas, V.; Aparicio, F. Impact of the Potential m6A Modification Sites at the 3′UTR of Alfalfa Mosaic Virus RNA3 in the Viral Infection. Viruses 2022, 14, 1718. [Google Scholar] [CrossRef]
- Arif, M.; Atta, S.; Bashir, M.A.; Hussain, A.; Khan, M.I.; Farooq, S.; Hannan, A.; Islam, S.U.; Umar, U.U.D.; Khan, M.; Lin, W.; Hashem, M.; Alamri, S.; Wu, Z. Molecular characterization and RSV Co-infection of Nicotiana benthamiana with three distinct begomoviruses. Methods 2020, 183, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Kumar Kushwaha, N.; Kumar Singh, A.; Pankaj Sahu, P.; Vinoth Kumar, R.; Chakraborty, S. Dynamics of a geminivirus-encoded pre-coat protein and host RNA-dependent RNA polymerase 1 in regulating symptom recovery in tobacco. Journal of Experimental Botany 2018, 69, 2085–2102. [Google Scholar] [CrossRef]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef]
- Baulcombe, D.C.; Dean, C. Epigenetic Regulation in Plant Responses to the Environment. Cold Spring Harbor Perspectives in Biology 2014, 6, a019471–a019471. [Google Scholar] [CrossRef]
- Blevins, T.; Podicheti, R.; Mishra, V.; Marasco, M.; Wang, J.; Rusch, D.; Tang, H.; Pikaard, C.S. Identification of Pol IV and RDR2-dependent precursors of 24 nt siRNAs guiding de novo DNA methylation in Arabidopsis. eLife 2015, 4, e09591. [Google Scholar] [CrossRef]
- Borges, F.; Martienssen, R.A. The expanding world of small RNAs in plants. Nature Reviews Molecular Cell Biology 2015, 16, 727–741. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.; Martienssen, R.A. The expanding world of small RNAs in plants. Nature Reviews Molecular Cell Biology 2015, 16, 727–741. [Google Scholar] [CrossRef]
- Boyko, A.; Kathiria, P.; Zemp, F.J.; Yao, Y.; Pogribny, I.; Kovalchuk, I. Transgenerational changes in the genome stability and methylation in pathogen-infected plants. Nucleic Acids Research 2007, 35, 1714–1725. [Google Scholar] [CrossRef]
- Butterbach, P.; Verlaan, M.G.; Dullemans, A.; Lohuis, D.; Visser, R.G.F.; Bai, Y.; Kormelink, R. Tomato yellow leaf curl virus resistance by Ty-1 involves increased cytosine methylation of viral genomes and is compromised by cucumber mosaic virus infection. Proceedings of the National Academy of Sciences 2014, 111, 12942–12947. [Google Scholar] [CrossRef]
- Castellano, M.; Martinez, G.; Marques, M.C.; Moreno-Romero, J.; Köhler, C.; Pallas, V.; Gomez, G. Changes in the DNA methylation pattern of the host male gametophyte of viroid-infected cucumber plants. Journal of Experimental Botany 2016, 67, 5857–5868. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.-K. Epigenetic regulation of stress responses in plants. Current Opinion in Plant Biology 2009, 12, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Lyons, D.B.; Kim, M.Y.; Moore, J.D.; Zilberman, D. DNA Methylation and Histone H1 Jointly Repress Transposable Elements and Aberrant Intragenic Transcripts. Molecular Cell 2020, 77, 310–323. [Google Scholar] [CrossRef]
- Corrêa, R.L.; Kutnjak, D.; Ambrós, S.; Bustos, M.; Elena, S.F. Identification of epigenetically regulated genes involved in plant-virus interaction and their role in virus-triggered induced resistance. BMC Plant Biology 2024, 24, 172. [Google Scholar] [CrossRef]
- Coursey, T.; Regedanz, E.; Bisaro, D.M. Arabidopsis RNA Polymerase V Mediates Enhanced Compaction and Silencing of Geminivirus and Transposon Chromatin during Host Recovery from Infection. Journal of Virology 2018, 92, e01320–17. [Google Scholar] [CrossRef] [PubMed]
- Cuerda-Gil, D.; Slotkin, R.K. Non-canonical RNA-directed DNA methylation. Nature Plants 2016, 2, 16163. [Google Scholar] [CrossRef]
- Cuerda-Gil, D.; Slotkin, R.K. Non-canonical RNA-directed DNA methylation. Nature Plants 2016, 2, 16163. [Google Scholar] [CrossRef] [PubMed]
- Dalakouras, A.; Ganopoulos, I. Induction of Promoter DNA Methylation Upon High-Pressure Spraying of Double-Stranded RNA in Plants. Agronomy 2021, 11, 789. [Google Scholar] [CrossRef]
- Deleris, A.; Halter, T.; Navarro, L. DNA Methylation and Demethylation in Plant Immunity. Annual Review of Phytopathology 2016, 54, 579–603. [Google Scholar] [CrossRef]
- Deleris, A.; Halter, T.; Navarro, L. DNA Methylation and Demethylation in Plant Immunity. Annual Review of Phytopathology 2016, 54, 579–603. [Google Scholar] [CrossRef]
- Ding, S.-W.; Voinnet, O. Antiviral Immunity Directed by Small RNAs. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant–pathogen interactions. Nature Reviews Genetics 2010, 11, 539–548. [Google Scholar] [CrossRef]
- Dong, Q.; Li, X.; Wang, C.-Z.; Xu, S.; Yuan, G.; Shao, W.; Liu, B.; Zheng, Y.; Wang, H.; Lei, X.; Zhang, Z.; Zhu, B. Roles of the CSE1L-mediated nuclear import pathway in epigenetic silencing. Proceedings of the National Academy of Sciences 2018, 115. [Google Scholar] [CrossRef]
- Dowen, R.H.; Pelizzola, M.; Schmitz, R.J.; Lister, R.; Dowen, J.M.; Nery, J.R.; Dixon, J.E.; Ecker, J.R. Widespread dynamic DNA methylation in response to biotic stress. Proceedings of the National Academy of Sciences 2012, 109. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Kiselev, K.V. Exogenous RNAs for Gene Regulation and Plant Resistance. International Journal of Molecular Sciences 2019, 20, 2282. [Google Scholar] [CrossRef]
- El-Sappah, A.H.; Yan, K.; Huang, Q.; Islam, Md. M.; Li, Q.; Wang, Y.; Khan, M.S.; Zhao, X.; Mir, R.R.; Li, J.; El-Tarabily, K.A.; Abbas, M. Comprehensive Mechanism of Gene Silencing and Its Role in Plant Growth and Development. Frontiers in Plant Science 2021, 12, 705249. [Google Scholar] [CrossRef]
- Erdmann, R.M.; Picard, C.L. RNA-directed DNA Methylation. PLOS Genetics 2020, 16, e1009034. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Qi, Y. RNAi in Plants: An Argonaute-Centered View. The Plant Cell 2016, 28, 272–285. [Google Scholar] [CrossRef]
- Fernández-Calvino, L.; Martínez-Priego, L.; Szabo, E.Z.; Guzmán-Benito, I.; González, I.; Canto, T.; Lakatos, L.; Llave, C. Tobacco rattle virus 16K silencing suppressor binds ARGONAUTE 4 and inhibits formation of RNA silencing complexes. Journal of General Virology 2016, 97, 246–257. [Google Scholar] [CrossRef]
- Gallego-Bartolomé, J. DNA methylation in plants: Mechanisms and tools for targeted manipulation. New Phytologist 2020, 227, 38–44. [Google Scholar] [CrossRef]
- Gallego-Bartolomé, J. DNA methylation in plants: Mechanisms and tools for targeted manipulation. New Phytologist 2020, 227, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Glick, E.; Zrachya, A.; Levy, Y.; Mett, A.; Gidoni, D.; Belausov, E.; Citovsky, V.; Gafni, Y. Interaction with host SGS3 is required for suppression of RNA silencing by tomato yellow leaf curl virus V2 protein. Proceedings of the National Academy of Sciences 2008, 105, 157–161. [Google Scholar] [CrossRef]
- Gong, Q.; Wang, Y.; Jin, Z.; Hong, Y.; Liu, Y. Transcriptional and post-transcriptional regulation of RNAi-related gene expression during plant-virus interactions. Stress Biology 2022, 2, 33. [Google Scholar] [CrossRef] [PubMed]
- Gui, X.; Liu, C.; Qi, Y.; Zhou, X. Geminiviruses employ host DNA glycosylases to subvert DNA methylation-mediated defense. Nature Communications 2022, 13, 575. [Google Scholar] [CrossRef] [PubMed]
- Gui, X.; Liu, C.; Qi, Y.; Zhou, X. Geminiviruses employ host DNA glycosylases to subvert DNA methylation-mediated defense. Nature Communications 2022, 13, 575. [Google Scholar] [CrossRef]
- Gupta, N.; Reddy, K.; Bhattacharyya, D.; Chakraborty, S. Plant responses to geminivirus infection: Guardians of the plant immunity. Virology Journal 2021, 18, 143. [Google Scholar] [CrossRef]
- Guyot, V.; Rajeswaran, R.; Chu, H.C.; Karthikeyan, C.; Laboureau, N.; Galzi, S.; Mukwa, L.F.T.; Krupovic, M.; Kumar, P.L.; Iskra-Caruana, M.-L.; Pooggin, M.M. A newly emerging alphasatellite affects banana bunchy top virus replication, transcription, siRNA production and transmission by aphids. PLOS Pathogens 2022, 18, e1010448. [Google Scholar] [CrossRef]
- Hamim, I.; Borth, W.B.; Marquez, J.; Green, J.C.; Melzer, M.J.; Hu, J.S. Transgene-mediated resistance to Papaya ringspot virus: Challenges and solutions. Phytoparasitica 2018, 46, 1–18. [Google Scholar] [CrossRef]
- Hamim, I.; Borth, W.B.; Melzer, M.J.; Suzuki, J.Y.; Wall, M.M.; Hu, J.S. Occurrence of tomato leaf curl Bangladesh virus and associated subviral DNA molecules in papaya in Bangladesh: Molecular detection and characterization. Archives of Virology 2019, 164, 1661–1665. [Google Scholar] [CrossRef]
- Hamim, I.; Borth, W.B.; Suzuki, J.Y.; Melzer, M.J.; Wall, M.M.; Hu, J.S. Molecular characterization of tomato leaf curl Joydebpur virus and tomato leaf curl New Delhi virus associated with severe leaf curl symptoms of papaya in Bangladesh. European Journal of Plant Pathology 2020, 158, 457–472. [Google Scholar] [CrossRef]
- Hamim, I.; Green, J.C.; Borth, W.B.; Melzer, M.J.; Wang, Y.N.; Hu, J.S. First Report of Banana bunchy top virus in Heliconia spp. On Hawaii. Plant Disease 2017, 101, 2153. [Google Scholar] [CrossRef]
- Hamim, I.; Sekine, K.-T.; Komatsu, K. How do emerging long-read sequencing technologies function in transforming the plant pathology research landscape? Plant Molecular Biology 2022, 110, 469–484. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Bejarano, E.R.; Robertson, D.; Mansoor, S. Geminiviruses: Masters at redirecting and reprogramming plant processes. Nature Reviews Microbiology 2013, 11, 777–788. [Google Scholar] [CrossRef]
- Haugland, R.A.; Cline, M.G. Post-transcriptional Modifications of Oat Coleoptile Ribonucleic Acids: 5′-Terminal Capping and Methylation of Internal Nucleosides in Poly(A)-Rich RNA. European Journal of Biochemistry 1980, 104, 271–277. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Ge, L.; Chen, Y.; Zhao, S.; Li, Z.; Zhou, X.; Li, F. m6A modification of plant virus enables host recognition by NMD factors in plants. Science China Life Sciences 2024, 67, 161–174. [Google Scholar] [CrossRef]
- He, H.; Jia, M.; Liu, J.; Zhou, X.; Li, F. Roles of RNA m6A modifications in plant-virus interactions. Stress Biology 2023, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Jia, M.; Liu, J.; Zhou, X.; Li, F. Roles of RNA m6A modifications in plant-virus interactions. Stress Biology 2023, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Cai, J.; Xu, T.; Kang, H. Epitranscriptomic mRNA modifications governing plant stress responses: Underlying mechanism and potential application. Plant Biotechnology Journal 2022, 20, 2245–2257. [Google Scholar] [CrossRef]
- Hull, R.; Centre, J.I. (Eds.). (2014). Plant virology (Fifth edition). Academic Press.
- Ismayil, A.; Haxim, Y.; Wang, Y.; Li, H.; Qian, L.; Han, T.; Chen, T.; Jia, Q.; Yihao Liu, A.; Zhu, S.; Deng, H.; Gorovits, R.; Hong, Y.; Hanley-Bowdoin, L.; Liu, Y. Cotton Leaf Curl Multan virus C4 protein suppresses both transcriptional and post-transcriptional gene silencing by interacting with SAM synthetase. PLOS Pathogens 2018, 14, e1007282. [Google Scholar] [CrossRef]
- Jackel, J.N.; Buchmann, R.C.; Singhal, U.; Bisaro, D.M. Analysis of Geminivirus AL2 and L2 Proteins Reveals a Novel AL2 Silencing Suppressor Activity. Journal of Virology 2015, 89, 3176–3187. [Google Scholar] [CrossRef]
- Jackel, J.N.; Buchmann, R.C.; Singhal, U.; Bisaro, D.M. Analysis of Geminivirus AL2 and L2 Proteins Reveals a Novel AL2 Silencing Suppressor Activity. Journal of Virology 2015, 89, 3176–3187. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Chen, M.; Xiang, M.; Guo, Z. RNAi-Based Antiviral Innate Immunity in Plants. Viruses 2022, 14, 432. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.; Hamilton, A.J.; Voinnet, O.; Thomas, C.L.; Maule, A.J.; Baulcombe, D.C. RNA–DNA Interactions and DNA Methylation in Post-Transcriptional Gene Silencing. The Plant Cell 1999, 11, 2291–2301. [Google Scholar] [CrossRef]
- Kang, H.; Fan, T.; Wu, J.; Zhu, Y.; Shen, W.-H. Histone modification and chromatin remodeling in plant response to pathogens. Frontiers in Plant Science 2022, 13, 986940. [Google Scholar] [CrossRef]
- Kierzek, E. The thermodynamic stability of RNA duplexes and hairpins containing N6-alkyladenosines and 2-methylthio-N6-alkyladenosines. Nucleic Acids Research 2003, 31, 4472–4480. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H. Multifaceted Chromatin Structure and Transcription Changes in Plant Stress Response. International Journal of Molecular Sciences 2021, 22, 2013. [Google Scholar] [CrossRef]
- Kouzarides, T. Chromatin Modifications and Their Function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef]
- Lee, C.H.; Carroll, B.J. Evolution and Diversification of Small RNA Pathways in Flowering Plants. Plant and Cell Physiology 2018. [CrossRef]
- Li, F.; Xu, X.; Huang, C.; Gu, Z.; Cao, L.; Hu, T.; Ding, M.; Li, Z.; Zhou, X. The AC 5 protein encoded by Mungbean yellow mosaic India virus is a pathogenicity determinant that suppresses RNA silencing-based antiviral defenses. New Phytologist 2015, 208, 555–569. [Google Scholar] [CrossRef]
- Li, F.; Zhao, N.; Li, Z.; Xu, X.; Wang, Y.; Yang, X.; Liu, S.-S.; Wang, A.; Zhou, X. A calmodulin-like protein suppresses RNA silencing and promotes geminivirus infection by degrading SGS3 via the autophagy pathway in Nicotiana benthamiana. PLOS Pathogens 2017, 13, e1006213. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.; Sun, Z. N6-methyladenosine (m6A) modification: Emerging regulators in plant-virus interactions. Virology 2025, 603, 110373. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Du, Z.; Tang, Y.; She, X.; Wang, X.; Zhu, Y.; Yu, L.; Lan, G.; He, Z. C4, the Pathogenic Determinant of Tomato Leaf Curl Guangdong Virus, May Suppress Post-transcriptional Gene Silencing by Interacting With BAM1 Protein. Frontiers in Microbiology 2020, 11, 851. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, X.; Fang, Y.; Liu, Y.; Bello, E.O.; Li, Y.; Xiong, R.; Li, Y.; Fu, Z.Q.; Wang, A.; Cheng, X. A plant RNA virus inhibits NPR1 sumoylation and subverts NPR1-mediated plant immunity. Nature Communications 2023, 14, 3580. [Google Scholar] [CrossRef] [PubMed]
- Lucibelli, F.; Valoroso, M.C.; Aceto, S. Plant DNA Methylation: An Epigenetic Mark in Development, Environmental Interactions, and Evolution. International Journal of Molecular Sciences 2022, 23, 8299. [Google Scholar] [CrossRef]
- Ma, X.; Zhou, Y.; Wu, L.; Moffett, P. Resistance gene Ty-1 restricts TYLCV infection in tomato by increasing RNA silencing. Virology Journal 2024, 21, 256. [Google Scholar] [CrossRef]
- Mardini, M.; Kazancev, M.; Ivoilova, E.; Utkina, V.; Vlasova, A.; Demurin, Y.; Soloviev, A.; Kirov, I. Advancing virus-induced gene silencing in sunflower: Key factors of VIGS spreading and a novel simple protocol. Plant Methods 2024, 20, 122. [Google Scholar] [CrossRef]
- Marquez-Molins, J.; Cheng, J.; Corell-Sierra, J.; Juarez-Gonzalez, V.T.; Villalba-Bermell, P.; Annacondia, M.L.; Gomez, G.; Martinez, G. Hop stunt viroid infection induces heterochromatin reorganization. New Phytologist 2024, 243, 2351–2367. [Google Scholar] [CrossRef]
- Martin, K.; Singh, J.; Hill, J.H.; Whitham, S.A.; Cannon, S.B. Dynamic transcriptome profiling of Bean Common Mosaic Virus (BCMV) infection in Common Bean (Phaseolus vulgaris L.). BMC Genomics 2016, 17, 613. [Google Scholar] [CrossRef] [PubMed]
- Martínez De Alba, A.E.; Elvira-Matelot, E.; Vaucheret, H. Gene silencing in plants: A diversity of pathways. Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms 2013, 1829, 1300–1308. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pérez, M.; Aparicio, F.; López-Gresa, M.P.; Bellés, J.M.; Sánchez-Navarro, J.A.; Pallás, V. Arabidopsis m6 A demethylase activity modulates viral infection of a plant virus and the m6 A abundance in its genomic RNAs. Proceedings of the National Academy of Sciences 2017, 114, 10755–10760. [Google Scholar] [CrossRef]
- Massart, S.; Chiumenti, M.; De Jonghe, K.; Glover, R.; Haegeman, A.; Koloniuk, I.; Komínek, P.; Kreuze, J.; Kutnjak, D.; Lotos, L.; Maclot, F.; Maliogka, V.; Maree, H.J.; Olivier, T.; Olmos, A.; Pooggin, M.M.; Reynard, J.-S.; Ruiz-García, A.B.; Safarova, D.; Candresse, T. Virus Detection by High-Throughput Sequencing of Small RNAs: Large-Scale Performance Testing of Sequence Analysis Strategies. Phytopathology® 2019, 109, 488–497. [Google Scholar] [CrossRef]
- Matzke, M.A.; Kanno, T.; Matzke, A.J.M. RNA-Directed DNA Methylation: The Evolution of a Complex Epigenetic Pathway in Flowering Plants. Annual Review of Plant Biology 2015, 66, 243–267. [Google Scholar] [CrossRef]
- Matzke, M.A.; Mosher, R.A. RNA-directed DNA methylation: An epigenetic pathway of increasing complexity. Nature Reviews Genetics 2014, 15, 394–408. [Google Scholar] [CrossRef]
- Matzke, M.A.; Mosher, R.A. RNA-directed DNA methylation: An epigenetic pathway of increasing complexity. Nature Reviews Genetics 2014, 15, 394–408. [Google Scholar] [CrossRef]
- Matzke, M.; Kanno, T.; Daxinger, L.; Huettel, B.; Matzke, A.J. RNA-mediated chromatin-based silencing in plants. Current Opinion in Cell Biology 2009, 21, 367–376. [Google Scholar] [CrossRef]
- Matzke, M.; Kanno, T.; Daxinger, L.; Huettel, B.; Matzke, A.J. RNA-mediated chromatin-based silencing in plants. Current Opinion in Cell Biology 2009, 21, 367–376. [Google Scholar] [CrossRef]
- Mei, Y.; Wang, Y.; Li, F.; Zhou, X. The C4 protein encoded by tomato leaf curl Yunnan virus reverses transcriptional gene silencing by interacting with NbDRM2 and impairing its DNA-binding ability. PLOS Pathogens 2020, 16, e1008829. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Wang, Y.; Li, F.; Zhou, X. The C4 protein encoded by tomato leaf curl Yunnan virus reverses transcriptional gene silencing by interacting with NbDRM2 and impairing its DNA-binding ability. PLOS Pathogens 2020, 16, e1008829. [Google Scholar] [CrossRef]
- Nichols, J.L.; Welder, L. A modified nucleotide in the poly(A) tract of maize RNA. Biochimica et Biophysica Acta (BBA) - Nucleic Acids and Protein Synthesis 1981, 652, 99–108. [Google Scholar] [CrossRef]
- Noris, E.; Catoni, M. (2020). Role of methylation during geminivirus infection. In Applied Plant Biotechnology for Improving Resistance to Biotic Stress (pp. 291–305). Elsevier. Available online: https://linkinghub.elsevier.com/retrieve/pii/B9780128160305000136.
- Norouzitallab, P.; Baruah, K.; Vanrompay, D.; Bossier, P. Can epigenetics translate environmental cues into phenotypes? Science of The Total Environment 2019, 647, 1281–1293. [Google Scholar] [CrossRef]
- Ordon, F.; Habekuss, A.; Kastirr, U.; Rabenstein, F.; Kühne, T. Virus Resistance in Cereals: Sources of Resistance, Genetics and Breeding. Journal of Phytopathology 2009, 157, 535–545. [Google Scholar] [CrossRef]
- Paprotka, T.; Deuschle, K.; Metzler, V.; Jeske, H. Conformation-Selective Methylation of Geminivirus DNA. Journal of Virology 2011, 85, 12001–12012. [Google Scholar] [CrossRef] [PubMed]
- Piedra-Aguilera, Á.; Jiao, C.; Luna, A.P.; Villanueva, F.; Dabad, M.; Esteve-Codina, A.; Díaz-Pendón, J.A.; Fei, Z.; Bejarano, E.R.; Castillo, A.G. Integrated single-base resolution maps of transcriptome, sRNAome and methylome of Tomato yellow leaf curl virus (TYLCV) in tomato. Scientific Reports 2019, 9, 2863. [Google Scholar] [CrossRef] [PubMed]
- Pooggin, M. How Can Plant DNA Viruses Evade siRNA-Directed DNA Methylation and Silencing? International Journal of Molecular Sciences 2013, 14, 15233–15259. [Google Scholar] [CrossRef] [PubMed]
- Raja, P.; Sanville, B.C.; Buchmann, R.C.; Bisaro, D.M. Viral Genome Methylation as an Epigenetic Defense against Geminiviruses. Journal of Virology 2008, 82, 8997–9007. [Google Scholar] [CrossRef]
- Raja, P.; Sanville, B.C.; Buchmann, R.C.; Bisaro, D.M. Viral Genome Methylation as an Epigenetic Defense against Geminiviruses. Journal of Virology 2008, 82, 8997–9007. [Google Scholar] [CrossRef]
- Rajeevkumar, S.; Anunanthini, P.; Sathishkumar, R. Epigenetic silencing in transgenic plants. Frontiers in Plant Science 2015, 6. [Google Scholar] [CrossRef]
- Ramirez-Prado, J.S.; Abulfaraj, A.A.; Rayapuram, N.; Benhamed, M.; Hirt, H. Plant Immunity: From Signaling to Epigenetic Control of Defense. Trends in Plant Science 2018, 23, 833–844. [Google Scholar] [CrossRef]
- Rashid, M.-O.; Wang, Y.; Han, C.-G. Molecular Detection of Potato Viruses in Bangladesh and Their Phylogenetic Analysis. Plants 2020, 9, 1413. [Google Scholar] [CrossRef]
- Rodríguez-Negrete, E.A.; Carrillo-Tripp, J.; Rivera-Bustamante, R.F. RNA Silencing against Geminivirus: Complementary Action of Posttranscriptional Gene Silencing and Transcriptional Gene Silencing in Host Recovery. Journal of Virology 2009, 83, 1332–1340. [Google Scholar] [CrossRef]
- Rodríguez-Negrete, E.; Lozano-Durán, R.; Piedra-Aguilera, A.; Cruzado, L.; Bejarano, E.R.; Castillo, A.G. Geminivirus R ep protein interferes with the plant DNA methylation machinery and suppresses transcriptional gene silencing. New Phytologist 2013, 199, 464–475. [Google Scholar] [CrossRef]
- Romero-Rodríguez, B.; Petek, M.; Jiao, C.; Križnik, M.; Zagorščak, M.; Fei, Z.; Bejarano, E.R.; Gruden, K.; Castillo, A.G. Transcriptional and epigenetic changes during tomato yellow leaf curl virus infection in tomato. BMC Plant Biology 2023, 23, 651. [Google Scholar] [CrossRef]
- Sahu, P.P.; Rai, N.K.; Chakraborty, S.; Singh, M.; Chandrappa, P.H.; Ramesh, B.; Chattopadhyay, D.; Prasad, M. Tomato cultivar tolerant to Tomato leaf curl New Delhi virus infection induces virus-specific short interfering RNA accumulation and defence-associated host gene expression. Molecular Plant Pathology 2010, 11, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Sastry, K.S.; Zitter, T.A. (2014). Management of Virus and Viroid Diseases of Crops in the Tropics. In K. S. Sastry & T. A. Zitter, Plant Virus and Viroid Diseases in the Tropics (pp. 149–480). Springer Netherlands. [CrossRef]
- Sato, Y.; Miyashita, S.; Ando, S.; Takahashi, H. Increased cytosine methylation at promoter of the NB-LRR class R gene RCY1 correlated with compromised resistance to cucumber mosaic virus in EMS-generated src mutants of Arabidopsis thaliana. Physiological and Molecular Plant Pathology 2017, 100, 151–162. [Google Scholar] [CrossRef]
- Secco, N.; Sheikh, A.H.; Hirt, H. Insights into the role of N6-methyladenosine (m6A) in plant-virus interactions. Journal of Virology 2025, 99. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Liu, Z.; Song, F.; Xie, Q.; Hanley-Bowdoin, L.; Zhou, X. Tomato SlSnRK1 Protein Interacts with and Phosphorylates βC1, a Pathogenesis Protein Encoded by a Geminivirus β-Satellite. Plant Physiology 2011, 157, 1394–1406. [Google Scholar] [CrossRef]
- Shen, W.; Bobay, B.G.; Greeley, L.A.; Reyes, M.I.; Rajabu, C.A.; Blackburn, R.K.; Dallas, M.B.; Goshe, M.B.; Ascencio-Ibáñez, J.T.; Hanley-Bowdoin, L. Sucrose Nonfermenting 1-Related Protein Kinase 1 Phosphorylates a Geminivirus Rep Protein to Impair Viral Replication and Infection. Plant Physiology 2018, 178, 372–389. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Dallas, M.B.; Goshe, M.B.; Hanley-Bowdoin, L. SnRK1 Phosphorylation of AL2 Delays Cabbage Leaf Curl Virus Infection in Arabidopsis. Journal of Virology 2014, 88, 10598–10612. [Google Scholar] [CrossRef] [PubMed]
- Shinde, H.; Dudhate, A.; Kadam, U.S.; Hong, J.C. RNA methylation in plants: An overview. Frontiers in Plant Science 2023, 14, 1132959. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Mishra, V.; Wang, F.; Huang, H.-Y.; Pikaard, C.S. Reaction Mechanisms of Pol IV, RDR2, and DCL3 Drive RNA Channeling in the siRNA-Directed DNA Methylation Pathway. Molecular Cell 2019, 75, 576–589. [Google Scholar] [CrossRef]
- Soto-Burgos, J.; Bassham, D.C. SnRK1 activates autophagy via the TOR signaling pathway in Arabidopsis thaliana. PLOS ONE 2017, 12, e0182591. [Google Scholar] [CrossRef]
- Stael, S.; Kmiecik, P.; Willems, P.; Van Der Kelen, K.; Coll, N.S.; Teige, M.; Van Breusegem, F. Plant innate immunity – sunny side up? Trends in Plant Science 2015, 20, 3–11. [Google Scholar] [CrossRef]
- Stroud, H.; Greenberg, M.V.C.; Feng, S.; Bernatavichute, Y.V.; Jacobsen, S.E. Comprehensive Analysis of Silencing Mutants Reveals Complex Regulation of the Arabidopsis Methylome. Cell 2013, 152, 352–364. [Google Scholar] [CrossRef]
- Tao, Z.; Yan, F.; Hahn, M.; Ma, Z. Regulatory roles of epigenetic modifications in plant-phytopathogen interactions. Crop Health 2023, 1, 6. [Google Scholar] [CrossRef]
- Tariq, M.; Paszkowski, J. DNA and histone methylation in plants. Trends in Genetics 2004, 20, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Tatineni, S.; Hein, G.L. Plant Viruses of Agricultural Importance: Current and Future Perspectives of Virus Disease Management Strategies. Phytopathology® 2023, 113, 117–141. [Google Scholar] [CrossRef]
- Tripathi, S.; Bau, H.; Chen, L.; Yeh, S. The ability of Papaya ringspot virusstrains overcoming the transgenic resistance of papaya conferred by the coat protein gene is not correlated with higher degrees of sequence divergence from the transgene. European Journal of Plant Pathology 2004, 110, 871–882. [Google Scholar] [CrossRef]
- Vatanparast, M.; Merkel, L.; Amari, K. Exogenous Application of dsRNA in Plant Protection: Efficiency, Safety Concerns and Risk Assessment. International Journal of Molecular Sciences 2024, 25, 6530. [Google Scholar] [CrossRef] [PubMed]
- Veluthambi, K.; Sunitha, S. Targets and Mechanisms of Geminivirus Silencing Suppressor Protein AC2. Frontiers in Microbiology 2021, 12, 645419. [Google Scholar] [CrossRef] [PubMed]
- Veluthambi, K.; Sunitha, S. Targets and Mechanisms of Geminivirus Silencing Suppressor Protein AC2. Frontiers in Microbiology 2021, 12, 645419. [Google Scholar] [CrossRef]
- Voinnet, O. Use, tolerance and avoidance of amplified RNA silencing by plants. Trends in Plant Science 2008, 13, 317–328. [Google Scholar] [CrossRef]
- Wale, S.; Platt, B.; D. Cattlin, N. Diseases, Pests and Disorders of Potatoes: A Colour Handbook (0 ed.). CRC Press. [CrossRef]
- Wambui Mbichi, R.; Wang, Q.-F.; Wan, T. (2020). RNA directed DNA methylation and seed plant genome evolution. Plant Cell Reports 2008, 39, 983–996. [Google Scholar] [CrossRef]
- Wang, B.; Yang, X.; Wang, Y.; Xie, Y.; Zhou, X. Tomato Yellow Leaf Curl Virus V2 Interacts with Host Histone Deacetylase 6 To Suppress Methylation-Mediated Transcriptional Gene Silencing in Plants. Journal of Virology 2018, 92, e00036–18. [Google Scholar] [CrossRef]
- Wang, C.; Wang, C.; Zou, J.; Yang, Y.; Li, Z.; Zhu, S. Epigenetics in the plant–virus interaction. Plant Cell Reports 2019, 38, 1031–1038. [Google Scholar] [CrossRef]
- Wang, H.; Hao, L.; Shung, C.-Y.; Sunter, G.; Bisaro, D.M. Adenosine Kinase Is Inactivated by Geminivirus AL2 and L2 Proteins. The Plant Cell 2003, 15, 3020–3032. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Y.; Gong, Q.; Ismayil, A.; Yuan, Y.; Lian, B.; Jia, Q.; Han, M.; Deng, H.; Hong, Y.; Hanley-Bowdoin, L.; Qi, Y.; Liu, Y. Geminiviral V2 Protein Suppresses Transcriptional Gene Silencing through Interaction with AGO4. Journal of Virology 2019, 93, e01675–18. [Google Scholar] [CrossRef]
- Waterhouse, P.M.; Wang, M.-B.; Lough, T. Gene silencing as an adaptive defence against viruses. Nature 2001, 411, 834–842. [Google Scholar] [CrossRef]
- Wierzbicki, A.T.; Ream, T.S.; Haag, J.R.; Pikaard, C.S. RNA polymerase V transcription guides ARGONAUTE4 to chromatin. Nature Genetics 2009, 41, 630–634. [Google Scholar] [CrossRef]
- Wu, Q.; Luo, Y.; Lu, R.; Lau, N.; Lai, E.C.; Li, W.-X.; Ding, S.-W. Virus discovery by deep sequencing and assembly of virus-derived small silencing RNAs. Proceedings of the National Academy of Sciences 2010, 107, 1606–1611. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Fan, G. The role of epigenetics in plant pathogens interactions under the changing environments: A systematic review. Plant Stress 2025, 15, 100753. [Google Scholar] [CrossRef]
- Xie, S.-S.; Duan, C.-G. Epigenetic regulation of plant immunity: From chromatin codes to plant disease resistance. aBIOTECH 2023, 4, 124–139. [Google Scholar] [CrossRef]
- Xie, S.-S.; Duan, C.-G. Epigenetic regulation of plant immunity: From chromatin codes to plant disease resistance. aBIOTECH 2023, 4, 124–139. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Liu, Y.; Wu, P.; Bian, Z.; Li, B.; Zhang, Y.; Zhu, B. Identification and virus-induced gene silencing (VIGS) analysis of methyltransferase affecting tomato (Solanum lycopersicum) fruit ripening. Planta 2024, 259, 109. [Google Scholar] [CrossRef]
- Yang, D.-L.; Zhang, G.; Wang, L.; Li, J.; Xu, D.; Di, C.; Tang, K.; Yang, L.; Zeng, L.; Miki, D.; Duan, C.-G.; Zhang, H.; Zhu, J.-K. Four putative SWI2/SNF2 chromatin remodelers have dual roles in regulating DNA methylation in Arabidopsis. Cell Discovery 2018, 4, 55. [Google Scholar] [CrossRef]
- Yang, L.; Fang, Y.; An, C.; Dong, L.; Zhang, Z.; Chen, H.; Xie, Q.; Guo, H. C 2-mediated decrease in DNA methylation, accumulation of si RNA s, and increase in expression for genes involved in defense pathways in plants infected with beet severe curly top virus. The Plant Journal 2013, 73, 910–917. [Google Scholar] [CrossRef]
- Yang, L.; Lang, C.; Wu, Y.; Meng, D.; Yang, T.; Li, D.; Jin, T.; Zhou, X. ROS1-mediated decrease in DNA methylation and increase in expression of defense genes and stress response genes in Arabidopsis thaliana due to abiotic stresses. BMC Plant Biology 2022, 22, 104. [Google Scholar] [CrossRef]
- Yang, X.; Xie, Y.; Raja, P.; Li, S.; Wolf, J.N.; Shen, Q.; Bisaro, D.M.; Zhou, X. Suppression of Methylation-Mediated Transcriptional Gene Silencing by βC1-SAHH Protein Interaction during Geminivirus-Betasatellite Infection. PLoS Pathogens 2011, 7, e1002329. [Google Scholar] [CrossRef]
- Yue, J.; Wei, Y.; Zhao, M. The Reversible Methylation of m6A Is Involved in Plant Virus Infection. Biology 2022, 11, 271. [Google Scholar] [CrossRef]
- Zerbini, F.M.; Briddon, R.W.; Idris, A.; Martin, D.P.; Moriones, E.; Navas-Castillo, J.; Rivera-Bustamante, R.; Roumagnac, P.; Varsani, A.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Geminiviridae. Journal of General Virology 2017, 98, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lang, Z.; Zhu, J.-K. Dynamics and function of DNA methylation in plants. Nature Reviews Molecular Cell Biology 2018, 19, 489–506. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yazaki, J.; Sundaresan, A.; Cokus, S.; Chan, S.W.-L.; Chen, H.; Henderson, I.R.; Shinn, P.; Pellegrini, M.; Jacobsen, S.E.; Ecker, J.R. Genome-wide High-Resolution Mapping and Functional Analysis of DNA Methylation in Arabidopsis. Cell 2006, 126, 1189–1201. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, H.; Huang, X.; Xia, R.; Zhao, Q.; Lai, J.; Teng, K.; Li, Y.; Liang, L.; Du, Q.; Zhou, X.; Guo, H.; Xie, Q. BSCTV C2 Attenuates the Degradation of SAMDC1 to Suppress DNA Methylation-Mediated Gene Silencing in Arabidopsis. The Plant Cell 2011, 23, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-H.; Liu, X.-L.; Fang, Y.-Y.; Fang, R.-X.; Guo, H.-S. CMV2b-Dependent Regulation of Host Defense Pathways in the Context of Viral Infection. Viruses 2018, 10, 618. [Google Scholar] [CrossRef]
- Zhi, P.; Chang, C. Exploiting Epigenetic Variations for Crop Disease Resistance Improvement. Frontiers in Plant Science 2021, 12, 692328. [Google Scholar] [CrossRef]
- Zhu, Q.-H.; Shan, W.-X.; Ayliffe, M.A.; Wang, M.-B. Epigenetic Mechanisms: An Emerging Player in Plant-Microbe Interactions. Molecular Plant-Microbe Interactions® 2016, 29, 187–196. [Google Scholar] [CrossRef]
- Zulfiqar, S.; Farooq, M.A.; Zhao, T.; Wang, P.; Tabusam, J.; Wang, Y.; Xuan, S.; Zhao, J.; Chen, X.; Shen, S.; Gu, A. Virus-Induced Gene Silencing (VIGS): A Powerful Tool for Crop Improvement and Its Advancement towards Epigenetics. International Journal of Molecular Sciences 2023, 24, 5608. [Google Scholar] [CrossRef]
Figure 1.
Epigenetic modifications and regulatory shifts in plants during viral infection.
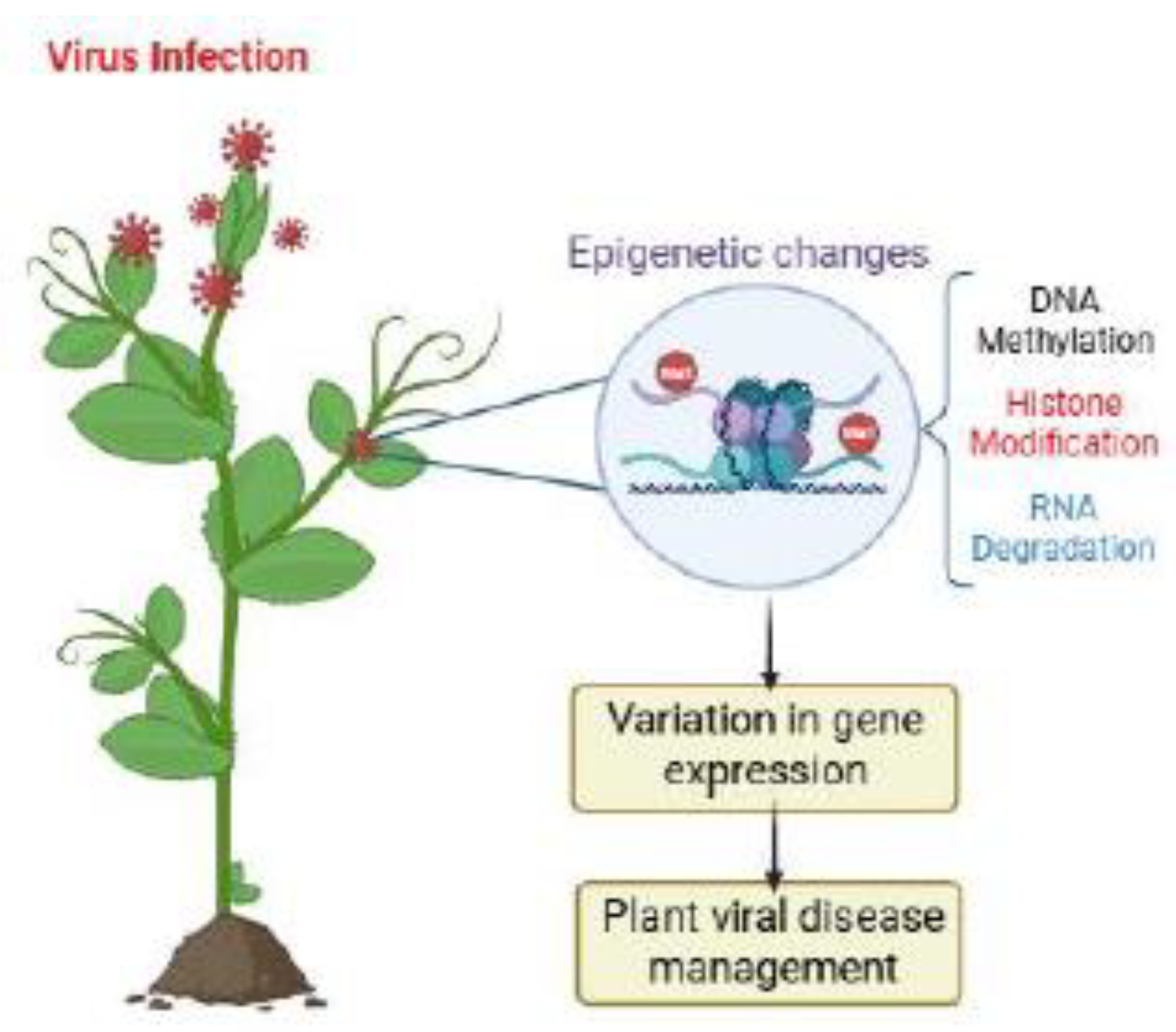
Figure 2.
A schematic diagram of post-transcriptional gene silencing (PTGS) and transcriptional gene silencing (TGS) as interconnected antiviral defense pathways in plants. In the left panel, PTGS is illustrated, where viral RNA molecules are recognized and degraded by 21–22 nt small interfering RNAs (siRNAs). These 21–22 nt siRNAs can also feed into a non-canonical RdDM pathway, resulting in sparse and locus-specific DNA methylation. This methylation recruits RNA polymerase IV (Pol IV), which initiates the canonical RdDM pathway. Pol IV transcribes single-stranded RNA (ssRNA), which is then converted into double-stranded RNA (dsRNA) by RNA-DEPENDENT RNA POLYMERASE 2 (RDR2). The dsRNA is processed into 24-nt siRNAs, which guide de novo DNA methylation through ARGONAUTE proteins and associated factors. In addition, histone modifications and chromatin remodelling occur at these loci, further reinforcing transcriptional repression. Together, these epigenetic changes establish stable TGS and effectively inhibit viral DNA transcription.
Figure 2.
A schematic diagram of post-transcriptional gene silencing (PTGS) and transcriptional gene silencing (TGS) as interconnected antiviral defense pathways in plants. In the left panel, PTGS is illustrated, where viral RNA molecules are recognized and degraded by 21–22 nt small interfering RNAs (siRNAs). These 21–22 nt siRNAs can also feed into a non-canonical RdDM pathway, resulting in sparse and locus-specific DNA methylation. This methylation recruits RNA polymerase IV (Pol IV), which initiates the canonical RdDM pathway. Pol IV transcribes single-stranded RNA (ssRNA), which is then converted into double-stranded RNA (dsRNA) by RNA-DEPENDENT RNA POLYMERASE 2 (RDR2). The dsRNA is processed into 24-nt siRNAs, which guide de novo DNA methylation through ARGONAUTE proteins and associated factors. In addition, histone modifications and chromatin remodelling occur at these loci, further reinforcing transcriptional repression. Together, these epigenetic changes establish stable TGS and effectively inhibit viral DNA transcription.
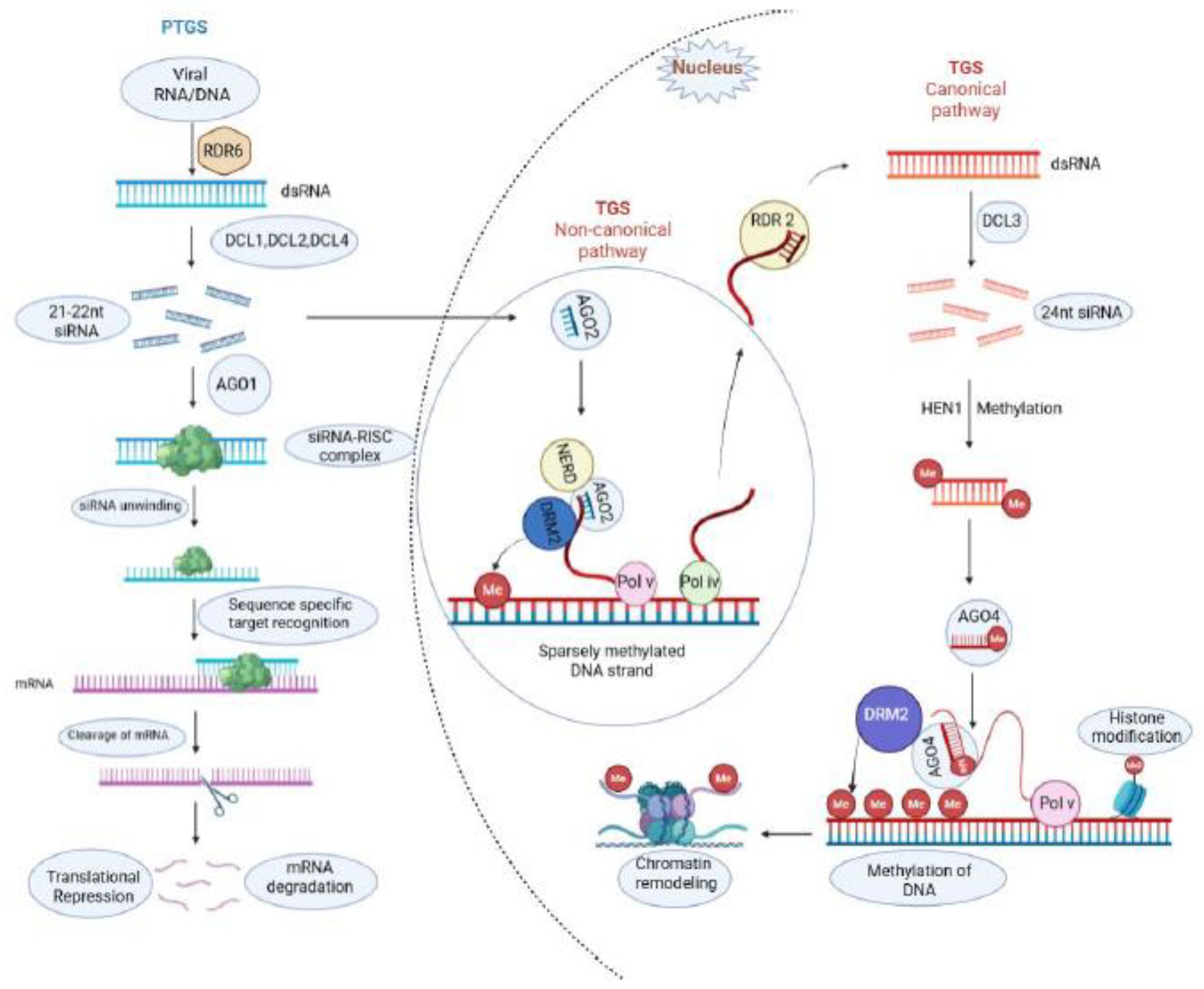
Table 1.
Estimated annual economic losses in major crops due to plant pathogenic viruses worldwide. Values are approximate and compiled from published articles.
Table 1.
Estimated annual economic losses in major crops due to plant pathogenic viruses worldwide. Values are approximate and compiled from published articles.
| Region/Country | Crop (s) | Estimated Loss (USD, approx.) | Causative Virus | Reference (s) |
|---|---|---|---|---|
| Africa/South Asia | Cassava | 1.9-2.7 billion | Cassava mosaic begomoviruses | Tatineni & Hein, 2023 |
| USA | Potato | 100-120 million | Potato leafroll Polerovirus | Sastry & Zitter, 2014; Wale et al.; 2008 |
| United Kingdom (UK) | Cereals (Barley,Oats,Rice,Wheat,Maize) | 10-15 million | Barley yellow dwarf virus | Ordon et al.; 2009 |
| South-East Asia | Rice | ~1.0 billion | Rice tungro viruses | Abo & Sy, 1997; Hull & Centre, 2014 |
| USA, Australia, Eastern Europe | Tomato, Lettuce, Eggplant, Pepper | 1.0-1.5 billion | Tomato spotted wilt virus | Tatineni & Hein, 2023 |
| Bangladesh | Potato | 0.5-1.8 billion | Potato leafroll virus, Potato virus X, Potato virus Y, Potato virus S, Potato virus H, Potato aucuba mosaic virus and Potato virus M | Rashid et al.; 2020 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



