Submitted:
01 September 2025
Posted:
01 September 2025
You are already at the latest version
Abstract
Backcrossing serves as an effective tool for interspecific gene introgression, facilitating germplasm improvement. To develop superior grouper aquaculture varieties, in this study, a backcrossed breed was constructed by crossing a male hybrid grouper, KGGG (Epinephelus moara ♀ × E. lanceolatus ♂), with a female parent kelp grouper (KG; E. moara). Here, we observed embryonic, larval development, and morphological changes in KG × KGGG. Additionally, the total length and body weight of the backcrossed breed and maternal parent were compared at 160 d after hatching (dAH). The results showed that fertilization and the hatching rate were 73.23 ± 4.23% and 51.74 ± 3.67%, respectively, and the egg size was 0.89 ± 0.03 mm. Hatching occurred 23:19 h after fertilization at 25 ± 1 ℃, and the newly hatched larvae were 1.94 ± 0.13 mm in length. Furthermore, at 160 dAH, the total length and body weight of the backcrossed breed were 1.5-fold and 2.4-fold greater, respectively, when compared with those of the maternal parent. Importantly, it is possible that backcrossed KG × KGGG could become a dominant strain in grouper aquaculture practices.
Keywords:
backcrossed breeding
; hybrid grouper KGGG
; embryonic development
; 27 growth performance
1. Introduction
Groupers (Epinephelinae) are commercially significant reef-associated fish that are widely distributed in tropical and temperate marine ecosystems [1]. They are highly valued in the aquaculture and seafood industry for their nutrient-rich profile and good taste, particularly in Southeast and East Asia. With the rapid expansion of grouper aquaculture, severe challenges have emerged, including the degradation of genetic resources, frequent disease outbreaks, and a critical shortage of high-quality juveniles. A potential solution to effectively address these issues is through hybridization. Interspecific hybridization of grouper species has been successfully achieved, such as the brown-marbled grouper (Epinephelus fuscoguttatus ♀ × giant grouper (E. lanceolatus) ♂ [2], humpback grouper (Cromileptes altivelis ) ♀ × giant grouper ♂ [3], red-spotted grouper (E. akaara) ♀ × giant grouper ♂ [4], and brown-marbled grouper ♀ × potato grouper (E. tukula) ♂ [5].
The KG is highly valued within groupers due to its exceptional rapid growth, remarkable environmental resilience, superior flavor profile, and considerable commercial importance [6]. The GG is the most favored species in the grouper aquaculture industry, which is driven by its high market value, fast growth, and excellent taste [7]. To combine the advantages of both fish species, our team established a novel hybrid progeny between a female kelp grouper (KG, E. moara) and a male giant grouper (GG, E. lanceolatus) [8], referred to as KGGG. However, the hybrid offspring KGGG exhibited a relatively low hatching rate of fertilized eggs (14.35 ± 8.02%). To address this issue and foster the rapid advancement of the grouper industry, in this study, we implemented a backcross breeding strategy for grouper aquaculture practices. The considerable variation in the spawning periods among the grouper species poses significant challenges for constructing backcrossed populations, which is compounded by their protogynous hermaphroditic nature. Specifically, individuals initially develop as females before undergoing a sex reversal process (requiring 5-7 years for completion) to become males [9]. Due to these challenges, there is a paucity of literature on backcross breeding in groupers. Additionally, due to the different breeding times of the hybrid parents, this study employed cryopreserved KGGG sperm for backcross breeding. Semen cryopreservation not only enables the long-term preservation of superior broodstock genetic materials but also facilitates crossbreeding between fish with asynchronous reproductive cycles or geographical isolation, such as groupers [10].
Regarding the research on backcross breeding for groupers, Sariat et al. reported the embryonic and larval development of (E. fuscoguttatus × E. lanceolatus, ♀) × (E. lanceolatus, ♂), including its morphology, sensory organ development, and behavioral changes [11]. The backcrossed offspring exhibited embryonic developmental patterns and sensory organ morphogenesis comparable to those of the purebred and hybrid groupers. Furthermore, the backcrossed progeny exhibited significantly superior growth performance and survival rates compared with the maternal grouper [12]. In addition, Aoki et al.reported on the hatchability and growth performance of (E. bruneus ♀ × E. lanceolatus ♂, ♀) × (E. lanceolatus, ♂), which suggested that the offspring had greater hatching success and growth performance than the hybrid of E. bruneus ♀ × E. lanceolatus ♂ [13].
In this study, we expanded on our previous work and, for the first time, constructed a backcrossed grouper between a female KG parent and a male hybrid KGGG grouper. The embryonic development and morphological variations in the backcrossed grouper were recorded, and a comparative analysis was conducted on the growth characteristics between the backcrossed offspring and the female KG parent. These findings will facilitate the expansion of the grouper germplasm resource pool, thereby advancing the development of the grouper aquaculture industry.
2. Materials and Methods
2.1. Experimental Materials
2.1.1. Acquisition and Hatching of the Fertilized Eggs
The processes of broodstock breeding, fertilization, and larval hatching were conducted at Laizhou Mingbo Aquaculture Co., Ltd. The KG (E. moara) and hybrid grouper KGGG (E. moara × E. lanceolatus) were used as the female and male parents, respectively. Broodstock of KG with an approximate body weight of 5.22 ± 0.89 kg and total length of 71.11 ± 4.18 cm was reared in a 40 m³ flow-through breeding tank, with a water depth of 80–150 cm. The water temperature was maintained at 20–27 ℃, with a salinity of 28–32‰, and dissolved oxygen >5 mg/L. The KGGG was also cultivated in the same conditions. They reach sexual maturity at 7 years of age, and these individuals had an average weight of 7.72 ± 1.96 kg and an average total length of 77.25 ± 6.05 cm. The sperm were collected, and the sperm freezing preservation solution EM (9 g/L NaCl, 10 g/L KHCO3, 10% calf serum, and 20% 1,2-propanediol) was used to make up a dilution at a ratio of 1:1. Then, it was transferred to 2.0 mL cryovials and frozen in liquid nitrogen for storage. The mature cloud grouper eggs were collected and placed in a 1000 mL beaker. The cryopreserved sperm was thawed in a water bath maintained at 36–39 ℃. Next, fertilization was conducted at a sperm-to-egg volume ratio of 1:1000 mL, with gentle stirring to ensure thorough mixing of the gametes. Subsequently, seawater equivalent to twice the volume of the fertilized eggs was added, and the mixture was allowed to stand for 5–6 min. Following this, seawater was introduced at 5–10 times the volume. The sunken eggs were filtered out, and rinsing and filtering of the remaining eggs were conducted 2–3 times with the same seawater. Then, the fertilized eggs were transferred into mesh cages for incubation.
2.1.2. Factory Cultivation of the Larvae
After hatching, larval feeding protocols were implemented as follows: SS-type rotifers were introduced at 3 d after hatching (dAH). L-type rotifers were added at 7 dAH. From 20 to 30 dAH, the larvae were fed Artemia nauplii, which was followed by a dietary transition to adult Artemia commencing from 31 dAH. Formulated feed was added at 37 dAH, with a complete dietary transition achieved by 55 dAH. Feeding frequency was twice daily. The rearing system comprised circular concrete tanks with a water volume of 40 m³. The environmental parameters were maintained as follows: dissolved oxygen ≥6 mg/L, salinity 30–32‰, temperature 24–27 ℃, pH 7.6–8.2, and ammonium nitrogen (NH₄⁺-N) concentration ≤0.1 mg/L.
2.2. Sampling and Observation
To investigate the developmental progression of the fertilized eggs, 10 specimens were periodically sampled and subjected to photographic observation using an optical microscope (Nikon E200). Additionally, the timing parameters and distinguishing features of each developmental phase were systematically recorded. From the newly hatched larval stage onward, 3–6 specimens displaying healthy growth status were regularly and randomly selected from the cultivation tanks for total length measurement and microscopic (Nikon E200) and stereomicroscopic (Olympus) photography combined with Image View software to document the growth performance and morphological variations. Samples were collected daily from 0 to 10 dAH. Thereafter, the larvae were further sampled at 1-day intervals from 11 to 15 dAH. Next, samples were collected at 5-day intervals. From 50 to 90 dAH, samples were collected at 10-day intervals.
2.3. Statistics
A one-way analysis of variance was conducted on the growth data using SPSS 27.0. The Least Significant Difference test and Duncan’s multiple range test were used to analyze the differences between the groups. The images generated using OriginPro 2022 software were employed for morphological analysis.
3. Results
3.1. Development of the Fertilized Eggs
The development of the fertilized eggs from the backcrossed KG and KGGG was observed under an optical microscope (Figure 1 and Figure 2). Each egg was spherical, transparent, and measured approximately 0.89 ± 0.03 mm in diameter (Figure 1a). There was a centrally located oil globule with a diameter of 0.19 ± 0.02 mm. The fertilized egg completed the entire embryonic development process under water temperature conditions of 25 ± 1 ℃ and a salinity of 29 ± 1‰.
The cleavage method of the offspring was classified as discoidal cleavage, consistent with that of most bony fish. At 00:41 h after fertilization (hAF), the blastodisc was evenly divided by a central longitudinal cleft, marking the onset of its first cleavage and producing two smooth cells (2-cell stage; Figure 1b). At 00:57 hAF, a longitudinal groove was observed perpendicular to the first cleavage furrow, dividing the blastodisc evenly into four blastomeres and marking the transition into the 4-cell stage (Figure 1c). At 01:23 hAF, irregular compression and division produced eight cells (Figure 1d). Afterward, the cells continued to divide, and the division planes became irregular. This process progressed sequentially through the 16-cell stage at 01:44 hAF (Figure 1e), 32-cell stage at 02:14 hAF (Figure 1f), 64-cell stage at 02:48 hAF (Figure 1g), 128-cell stage at 03:10 hAF (Figure 1h), and finally the multicellular stage at 03:35 hAF (Figure 1i). Subsequently, the cell volume decreased, and the embryo development progressed to the morula stage at 03:45 hAF (Figure 1j). Continuous cell accumulation at the animal pole led to the formation of a blastocoel, marking the blastocyst stage at 04:14 hAF (Figure 1k). After 4 min, the cells started to invaginate (Figure 1l). Then, early gastrula, mid gastrula, late gastrula were observed from 5:25 hAF to 7:04 hAF (Figure 1m–o). Embryo formation began at 08:20 hAF (Figure 1p), and the blastopore closed completely by 08:22 hAF (Figure 2a). Then, optic vesicles became visible at 09:34 hAF (Figure 2b), and myomeres formed at 10:20 hAF (Figure 2c). The emergence of otocyst vesicles occurred at 11:15 hAF (Figure 2d). At 13:54 hAF, 14 pairs of muscle segments were visible, their morphology became clear, and a brain vesicle appeared (Figure 2e). Kupffer’s vesicle was identified in the posterior region of the embryo, indicating progression to the tail bud stage at 16:10 hAF (Figure 2f). Furthermore, lens vesicles were observed during the late stage of egg development at 19:13 hAF (Figure 2g). Spontaneous embryonic movements increased in frequency and the heart began to beat at 20:39 hAF (Figure 2h). In the early incubation stage, the embryo’s heartbeat reached 54–57 beats per minute at 22:25 hAF (Figure 2i). Finally, hatching commenced at 23:19 hAF (Figure 2j) and concluded at 23:29 hAF (Figure 2k). The fertilization rate and hatching rate were 73.23 ± 4.23% and 51.74 ± 3.67%, respectively. The results showed that the morphological changes in the offspring’s embryonic development were generally consistent with those of KG. However, the embryonic developmental time for each stage was shorter than that of the female KG parent. Moreover, the body length of the newly hatched hybrid offspring (Figure 2k) exceeded that of the female KG parent (Figure 2m)
3.2. Characteristics of the Hatched Larvae
The morphology of the backcrossed grouper during the different growth stages is illustrated in Figure 3. The newly hatched larva was completely transparent, with undeveloped organs and no fins (Figure 2k), which hindered its ability to swim and caused it to float at the water’s surface. There were a few pigment spots on the fish’s body. Additionally, 1 dAH, the yolk sac was reduced to approximately half of the body length, the notochord had thickened, the muscle segments were clearly defined, and the digestive system had not yet developed (Figure 2l). Moreover, the yolk sac in the abdominal area of the larva had shrunk significantly, the fin membranes had widened, and the digestive tract had begun to expand, with the intestines forming a continuous tube that curved outward at the end (Figure 3a). Notably, the larva remained in the endogenous nutrition stage throughout these morphological changes. Along with the absorption of the yolk sac, eye spots, a mouth slit, and an anus became apparent. Melanophores were primarily concentrated in the eye spots and mid-section of the larva (Figure 3b). At 9 dAH, the spines of the dorsal and pelvic fins were clearly visible, indicating full development of the tripartite spines (composed of dorsal and pelvic fin spines). However, the dorsal fin spine was slightly shorter than that of the pelvic fin, with pigment deposition observed at the growing tips. Ocular development produced a clear eye structure (Figure 3c). We also noted that the growth of the tripartite spines had accelerated, which was accompanied by increased melanin accumulation at 11 dAH (Figure 3d). The entire abdominal cavity exhibited significant pigmentation at 13 dAH (Figure 3e). Protruding eyes became visible at 15 dAH (Figure 3f), and head structures were well-developed by 20 dAH (Figure 3g). At 25 dAH, the dorsal, pectoral, and caudal fins further developed with emergence of the third dorsal fin spine (Figure 3h). At 35 dAH, scale formation commenced alongside completed fin differentiation, abdominal pigmentation intensified, transparency decreased, and the first dorsal fin spine emerged, while tripartite spines exhibited progressive regression (Figure 3i). At 45 dAH, melanophore accumulation was observed along the dorsal fins and lateral line, particularly near the caudal region, and the tripartite spine length reduced by 50% (Figure 3j). At 50 dAH, the larvae reached 30 mm in total length with visible gray pigmentation and melanophore linearization along the dorsal fin base and lateral line (Figure 3k). Subsequent development resulted in body length progression to 37 mm (60 dAH) with complete tripartite spine retraction and adult-like bilateral striping (Figure 3l). At 70 dAH, the larvae reached 40 mm with well-developed muscles (Figure 3m). By 80 dAH, the fastest-growing larvae attained 80 mm in total length (Figure 3n). At 90 dAH, a significant darkening in the larval body coloration was noted (Figure 3o).
3.3. Growth Changes in the Larvae
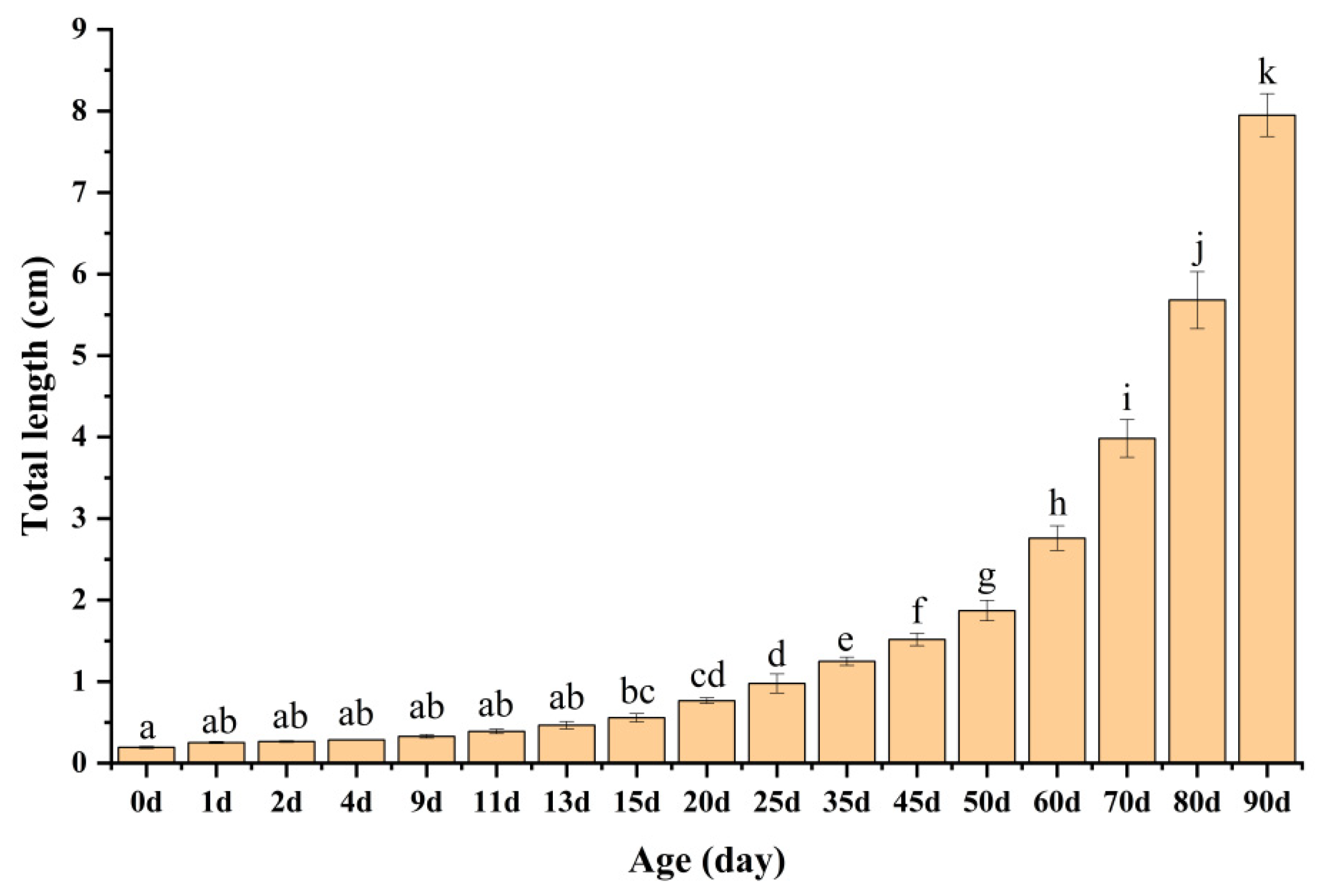
The total length of the KG × KGGG larvae progressively increased with age (Figure 4). The newly hatched larvae were 1.94 ± 0.13 mm. Their lengths increased to 2.53 ± 0.11 mm at 1 dAH, 2.65 ± 0.10 mm at 2 dAH, and 2.85 ± 0.03 mm at 4 dAH. During this period, the larvae primarily relied on endogenous nutrition from the yolk sac, and, therefore, their overall length showed minimal observable changes. The daily total length increased steadily to 3.29 ± 0.21 mm, 3.90 ± 0.28 mm, 4.66 ± 0.46 mm, and 5.57 ± 0.51 mm at 9, 11, 13, and 15 dAH, respectively. This may be related to the fry’s inability to adapt to the transition from endogenous to exogenous nutrition in the early stages. Subsequent to the commencement of the Artemia feeding protocols, rapid growth was observed at 20, 25, 35, and 45 dAH, reaching 7.67 ± 0.35 mm, 9.77 ± 1.18 mm, 12.50 ± 0.50 mm, and 15.17 ± 0.76 mm, respectively. Due to the complete transformation of their diet, the growth rate accelerated significantly, reaching values of 18.71 ± 1.25 mm, 27.60 ± 1.52 mm, 39.83 ± 2.32 mm, 56.80 ± 3.49 mm, and 79.50 ± 2.65 mm at 50, 60, 70, 80, and 90 dAH, respectively.
3.4. Comparison Between the Development of the Offspring and Maternal Groupers
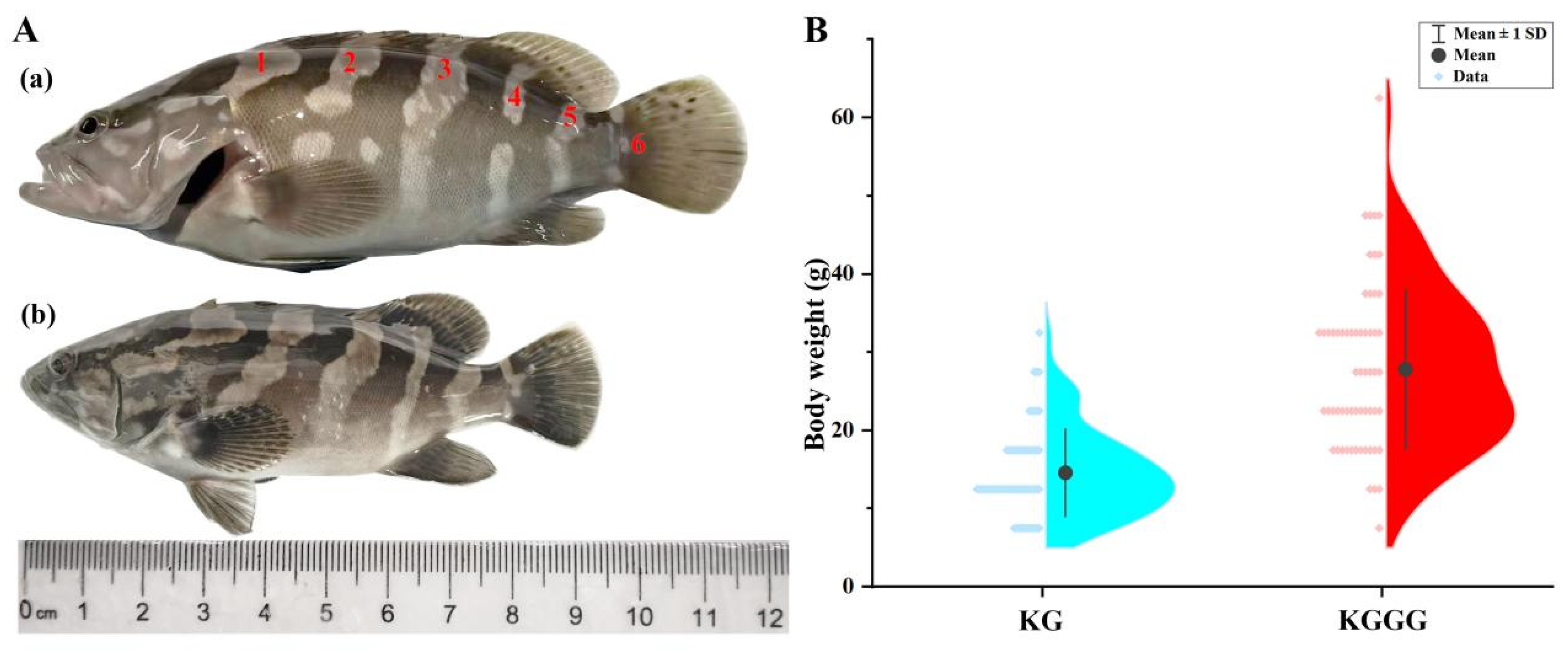
During the early hatching phase of the larvae, the hybrid offspring measured 1.94 ± 0.12 mm in total length (Figure 2k), which was significantly greater than that of the KG (1.79 ± 0.17 mm; Figure 2m). The total length and body weight of the offspring at 160 dAH were 11.63 ± 1.40 cm and 27.81 ± 10.16 g, respectively, which were approximately 1.2-fold and 1.9-fold increases compared with the KG at 160 dAH. The body coloration and patterns of the offspring were similar to those of the parent KG, with six grayish-white stripes distributed on both sides of the body. However, the offspring’s body and fins were slightly lighter in color, and the second, fourth, and sixth stripes were discontinuous.
Figure 5.
Morphological comparison between 160-day-old backcross progeny KG × KGGG and backcross maternal parent KG: (A) Body length comparison (a) KGGG (b) KG; (B) Body weight comparison.
Figure 5.
Morphological comparison between 160-day-old backcross progeny KG × KGGG and backcross maternal parent KG: (A) Body length comparison (a) KGGG (b) KG; (B) Body weight comparison.
4. Discussion
4.1. Fertilization and Hatching Rates
Fertilization and hatching rates serve as critical metrics for assessing the production of superior-quality fish offspring, indicating larval survival potential, and they can be used to optimize the cost-effectiveness of hatchery operations [14]. The fertilization and hatching rates of KG × KGGG reached 73.23 ± 4.23% and 51.74 ± 3.67%, respectively, which were significantly higher than those of OGGG (E. coioides ♀ × E. lanceolatus ♂) ♀ × GG ♂ (23.3% and 1.5%) [15], slightly higher than those of TGGG (E. fuscoguttatus ♀ × E. lanceolatus ♂) ♀ × GG ♂ (70.65% and 39.51%) [11], and markedly lower than those of TG (E. fuscoguttatus, ♀) × TGGG ♂ (94.33 ± 1.20% and 89.00 ± 1.73%) [1]. Thus, the fertilization and hatching rates were varied among the backcross combinations of groupers, being jointly influenced by the parental genetic characteristics and external cultivation conditions, including temperature. It is worth noting that the hatching rate documented in this study was significantly higher than that of the hybrid grouper KGGG (14.35 ± 8.02%) as reported by Chen et al. [8]. This was similar to LGGG (E. bruneus ♀ × E. lanceolatus ♂) ♀ × GG ♂, where the hatching rate of the backcross was higher than that of the female parent LGGG [13]. The results from the aforementioned studies demonstrated that backcrossing enhanced the hatching rate of the hybrid groupers, which contributes to the survival of the offspring. The high-quality fertilization and hatching rates in this study suggest potential genetic compatibilities between the backcrossed grouper offspring and the maternal parent, which indicates that the backcrossed offspring could be a promising candidate species for aquaculture practices.
4.2. Egg Development and Hatching Time
The egg development of the backcrossed KG and KGGG was similar to other Epinephelus species [16,17,18,19]. Their egg size was slightly larger (0.89 ± 0.03 mm) when compared with that of paternal KGGG (0.88 ± 0.03 mm) [8] and maternal KG (0.87 ± 0.01 mm) [20]. In addition, the egg size was relatively larger than other backcrossed groupers, such as TGGG × GG (0.79 ± 0.02 mm) [11] and OGGG × GG (0.76 ± 0.03 mm) [15]. Furthermore, the backcrossed egg size of KG × KGGG was also larger in comparison with those of other hybrid groupers, including: TGGG at 0.84 ± 0.03 mm [16]; OGTG at 0.83 ± 0.02 mm [19], and CGTG at 0.74 ± 0.02 mm [18]. Generally, larger eggs demonstrate enhanced hatchability, and larvae hatching from bigger eggs find it easier to consume exogenous food sources as the large yolk sac delivers more energy for development and metamorphosis [1]. Hence, the larger eggs observed in the backcrossed grouper progeny may contribute to enhanced offspring development.
The hatching time (23:19 hAF) recorded in our study was slightly later than that of the other hybrid species, such as TGGG (18:00 hAF) [16]; OGTG (17:30 hAF) [19]; and CGTG (19:00 hAF) [18]. The hatching time was also different from that of the other backcrossed grouper species, being 39 min later than that of OGGG × GG [15] and approximately 6.5 h later than that of TGGG × GG [11]. The delayed hatching time is likely attributable to variations in water temperature. The incubation water temperatures of OGGG × GG and TGGG × GG were relatively higher, around 27–29 ℃, compared with 25 ± 1 ℃ for KG × KGGG [11,15]. Temperature is a critical determinant of the duration of embryonic development [21]. As previously reported by Yoseda et al. and Song et al., the growth rates of certain fish species, including Malabar grouper (Epinephelus malabaricus), coral trout grouper (Plectropomus leopardus), and sevenband grouper (Epinephelus septemfasciatus), increased more rapidly with rising temperature [22,23,24]. Notably, compared with the maternal hatching time (38:17 hAF) [20] and paternal hatching time (32:45 hAF) [8], the hatching time of their backcrossed descendants was earlier. This finding indicates the promising potential of KG × KGGG as a newly developed hybrid for the aquaculture industry.
4.3. Growth
The total length of newly hatched KG × KGGG larvae was longer than that of the maternal species, KG, with a total length of 1.79 ± 0.17 mm (Figure 2m). The total length was also greater than those of other hybrid and backcrossed groupers, such as CGTG (1.83 mm) [18], OGGG (1.53 ± 0.01 mm) [19], OGGG × GG (1.67 ± 0.07 mm) [15], and TGGG × GG (1.336 ± 0.118 mm) [11]. However, the newly hatched larvae of KG × KGGG were slightly shorter compared with those of TGGG and KGGG, at 2.00 ± 0.30 mm and 1.95 ± 0.06 mm in length, respectively [8,16]. According to Hart and Reynolds, larger larvae may enhance survival success [25].
In this study, within the initial 48 h, growth was relatively fast. A similar conclusion was obtained by Franz et al. and Sariat et al., where rapid growth was seen upon hatching [11,26]. During the early ontogenetic stages, a smaller proportion of energy is allocated to environmental interactions, and more energy is invested in growth itself [27]. In this study, the larvae initiated feeding at 3 dAH, and a similar first feeding timing was found in the backcrossed grouper OGGG × GG [15]. At 4 dAH, the larval growth rate exhibited a gradual acceleration, which may be attributed to the adaptation to exogenous feeding. Moreover, the growth rate of the larval fish increased again after 20 dAH, which was potentially associated with the initiation of feeding regimens with Artemia nauplii. Following 50 dAH, there was a significant acceleration in the growth rate of the larvae, which was attributed to the completion of the transition to formulated feed.
4.4. Comparative Analysis with the Maternal Broodstock Grouper
At 160 dAH, the backcrossed offspring of KG and KGGG exhibited regular aligned linear stripe patterns on their lateral body sides, resembling those of their maternal parent (KG), which aligns with the findings of Wei et al. [28]. This may be attributed to the higher proportion (75%) of genetic material contributed by the kelp grouper when serving as the recurrent parent, which would have likely led to the offspring being more influenced by maternal effects [29]. However, KG × KGGG resembled the hybrid grouper KGGG in body size, demonstrating significantly larger morphometric dimensions compared with those of KG. The total lengths of KG × KGGG and KG were 11.63 ± 1.40 cm and 9.66 ± 1.25 cm, respectively, and their body weights were 27.81 ± 10.16 g and 14.59 ± 5.57 g at 160 dAH. Similar findings were documented by Lago et al., with backcrossed breeding individuals showing higher weights and longer lengths compared with the maternal Chitralada tilapia [30]. Overall, our findings indicated that the backcrossed offspring morphologically resembled the maternal parent KG, and their body size was more similar to that of the paternal hybrid grouper KGGG. The divergent traits between the backcrossed offspring and KG were mainly the body length and weight. The backcrossed groupers exhibited accelerated growth rates, which were likely attributed to paternal inheritance of superior rapid-growth traits. Liu Yang et al. found that the formation of superior growth heterosis typically requires large-sized groupers as paternal parents, with hybrids exhibiting paternal-biased growth superiority [31]. In addition, a previous study demonstrated that in the orthogonal cross (E. lanceolatus ♂ × E. moara ♀), the offspring exhibited significantly higher growth rates compared with the reciprocal cross (E. lanceolatus ♀ × E. moara ♂), revealing the dominant role of the paternal genotype in the inheritance of growth traits [32]. Hence, further studies are needed to clarify the genetic basis of growth traits in KG × KGGG.
5. Conclusions
This study showed for the first time that KG × KGGG backcrossed hybrid populations can be successfully established. Our study conducted a preliminary analysis of the embryogenesis, larval early-stage morphology, growth patterns, and heritable genetic traits of the backcrossed hybrid grouper KG × KGGG, providing a critical reference for future parental selection and germplasm authentication in grouper crossbreeding programs.
Author Contributions
Y.L. and Y.T. co-conceived this study and supervised the experiments; Y.X., S.W., T.D., L.W., Z.L. and L.L. performed the experiments, and conducted the data analysis, and created figure. Y.X. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Hainan Province Science and Technology Special Fund (ZDYF2025SXLH002); the Key Research and Development Project of Shandong Province (2022LZGC016); the National Key Research and Development Program of China (2022YFD2400502; 2022YFD2400103); China Agriculture Research System of MOF and MARA (CARS-47-G31); Qingdao Natural Science Foundation (23-2-1-53-zyyd-jch; 24-4-4-zrjj-39-jch); Qingdao Science and TechnologyBenefiting the People Demonstration Project (24-1-8-xdny-3-nsh); the Central Public-interest Scientific Institute Basal Research Fund, CAFS (2025CG02; 2020TD19); and the Yellow Sea Fisheries ResearchInstitute Research Fees (20603022025002; 20603022021010).
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| KG | Epinephelus moara |
| KGGG | Epinephelus moara ♀ × Epinephelus lanceolatus ♂ |
| dAH | days after hatching |
| hAF | hours after fertilization |
| GG | Epinepheluslanceolatus |
| OGGG | Epinephelus coioides ♀ × Epinephelus lanceolatus ♂ |
| TG | Epinephelusfuscoguttatus |
| TGGG | Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂ |
| LGGG | Epinephelus bruneus ♀ × Epinephelus lanceolatus ♂ |
| OGTG | Epinephelus coioides ♀ × Epinephelus fuscoguttatus ♂ |
| CGTG | Epinephelus corallicola ♀ × Epinephelus fuscoguttatus ♂ |
References
- Yang, Y.; Wang, T.; Chen, J.; Wu, X.; Wu, L.; Zhang, W.; Luo, J.; Xia, J.; Meng, Z.; Liu, X. First construction of interspecific backcross grouper and genome-wide identification of their genetic variants associated with early growth. Aquaculture 2021, 545, 737221. [CrossRef]
- Sun, Y.; Guo, C.-Y.; Wang, D.-D.; Li, X. F.; Xiao, L.; Zhang, X.; You, X.; Shi, Q.; Hu, G.-J.; Fang, C. Transcriptome analysis reveals the molecular mechanisms underlying growth superiority in a novel grouper hybrid (Epinephelus fuscogutatus♀× E. lanceolatus♂). BMC genetics 2016, 17 (1), 24. [CrossRef]
- Chen, J.; Ye, Z.; Yu, Z.; Wang, J.; Li, P.; Chen, X.; Liu, Y.; Zhang, Y. The complete mitochondrial genome of the hybrid grouper (Cromileptes altivelis♀× Epinephelus lanceolatus♂) with phylogenetic consideration. Mitochondrial Dna Part B 2017, 2 (1), 171-172. [CrossRef]
- Guo, Y.; Li, W.; Liu, Y.; Li, P.; Wei, R.; Liu, Y.; Zhang, Y.; Xiao, L. The complete mitochondrial genome of the hybrid grouper Epinephelus akaara ♀× Epinephelus lanceolatus ♂. Mitochondrial DNA Part B 2018, 3 (2), 599-600.
- Cheng, M.; Tian, Y.; Li, Z.; Wang, L.; Wu, Y.; Zhang, J.; Pang, Z.; Ma, W.; Zhai, J. The complete mitochondrial genome of the hybrid offspring Epinephelus fuscoguttatus♀× Epinephelus tukula♂. Mitochondrial DNA Part B 2019, 4 (2), 2717-2718.
- Lu LiJun, L. L.; Chen Chao, C. C.; Ma AiJun, M. A.; Zhai JieMing, Z. J.; Wang XinAn, W. X.; Li WeiYe, L. W. Studies on the feeding behavior and morphological developments of Epinephelus moara in early development stages. 2011.
- Chan, N. W. W.; Johnston, B. Applying the triangle taste test to wild and cultured humpback grouper (Cromileptes altivelis) in the Hong Kong market. SPC Live Reef Fish Information Bulletin 2007, 17, 31-35.
- Chen, Z. F.; Tian, Y. S.; Wang, P. F.; Tang, J.; Liu, J. C.; Ma, W. H.; Li, W. S.; Wang, X. M.; Zhai, J. M. Embryonic and larval development of a hybrid between kelp grouper Epinephelus moara♀× giant grouper E. lanceolatus♂ using cryopreserved sperm. Aquaculture Research 2018, 49 (4), 1407-1413.
- Bhandari, R. K.; Komuro, H.; Nakamura, S.; Higa, M.; Nakamura, M. Gonadal restructuring and correlative steroid hormone profiles during natural sex change in protogynous honeycomb grouper (Epinephelus merra). Zoological science 2003, 20 (11), 1399-1404. [CrossRef]
- Ding, X.Y.; Li, Z. T.; Duan, P.F.; Qiu Y.S.; Wang, X.Y.; Li, L.L.; Wang, L.N.; Liu,Y.; Li, W.S.; Wang, Q.B.; Zhao, X.; Tian, Y.S.; Li, Z.T. Effects of long-term cryopreservation on ultrastructure and enzyme activity of Epinephelus lanceolatus sperm. Journal of Fisheries of China 2023, 47(7): 079605.
- Sariat, S. A.; Ching, F. F.; Faudzi, N. M.; Senoo, S. Embryonic and larval development of backcrossed hybrid grouper between TGGG (Epinephelus fuscoguttatus × E. lanceolatus) and giant grouper (E. lanceolatus). Aquaculture 2023, 576, 739833.
- Tan, J. Backcross breeding between TGGG hybrid grouper (Epinephelus fuscoguttatus × E. lanceolatus) and giant grouper (E. lanceolatus). Journal of Survey in Fisheries Sciences 2021, 7 (2), 49-62. [CrossRef]
- Aoki, R.; Matsumasa, T.; Kumada, T.; Jin, N.; Masuma, S. Hatchability and growth performance of F1, F2, and backcross progenies of Epinephelus bruneus and Epinephelus lanceolatus. Aquaculture International 2025, 33 (4), 291. [CrossRef]
- Ching, F. F.; Othman, N.; Anuar, A.; Shapawi, R.; Senoo, S. Natural spawning, embryonic and larval development of F2 hybrid grouper, tiger grouper Epinephelus fuscoguttatus × giant grouper E. lanceolatus. International Aquatic Research 2018, 10 (4), 391-402. [CrossRef]
- Luan, G. H.; Luin, M.; Shapawi, R.; Fui Fui, C.; Senoo, S. Egg development of backcrossed hybrid grouper between OGGG (Epinephelus coioides × Epinephelus lanceolatus) and giant grouper (Epinephelus lanceolatus). Int. J. of Aquatic Science 2016, 7 (1), 13-18.
- Ch’ng, C. L.; Senoo, S. Egg and larval development of a new hybrid grouper, tiger grouper Epinephelus fuscoguttatus × giant grouper E. lanceolatus. Aquaculture Science 2008, 56 (4), 505-512.
- Zhou, L.; Weng, W.; Li, J.; Lai, Q. Studies on embryonic development, morphological development and feed changeover of Epinephelus lanceolatus larva. Chinese Agricultural Science Bulletin 2010, 26 (1), 293-302.
- Addin, A.; Senoo, S. Production of hybrid groupers: spotted grouper, Epinephelus polyphekadion × tiger grouper, E. fuscoguttatus and coral grouper, E. Corallicola × tiger grouper. In 2011 International symposium on grouper culture—technological innovation and industrial development, Taiwan, 2011.
- Koh, I. C. C.; Shaleh, S. R. M.; Senoo, S. Egg and larval development of a new hybrid orange-spotted grouper Epinephelus coioides × tiger grouper E. fuscoguttatus. Aquaculture Science 2008, 56 (3), 441-451.
- Song, Z.; Chen, C.; Zhai, J.; Li, Y.; Ma, W.; Wang, L.; Wu, L. Embryonic development and morphological characteristics of larval, juvenile and young kelp bass, Epinephelus moara. Progress in Fishery Sciences 2012, 33 (3), 26-34.
- Mandić, M.; Regner, S. Variation in fish egg size in several pelagic fish species. Studia Marina 2014, 27 (1), 31-46.
- Yoseda, K.; Dan, S.; Sugaya, T.; Yokogi, K.; Tanaka, M.; Tawada, S. Effects of temperature and delayed initial feeding on the growth of Malabar grouper (Epinephelus malabaricus) larvae. Aquaculture 2006a, 256 (1-4), 192-200. [CrossRef]
- Yoseda, K.; Teruya, K.; Yamamoto, K.; Asami, K. Effects of different temperature and delayed initial feeding on larval feeding, early survival, and the growth of coral trout grouper, Plectropomus leopardus larvae. Aquaculture Science 2006b, 54 (1), 43-50.
- Song, Y.B.; Lee, C.H.; Kang, H.C.; Kim, H.B.; Lee, Y.D. Effect of water temperature and salinity on the fertilized egg development and larval development of sevenband grouper, Epinephelus septemfasciatus. Development & reproduction 2013, 17 (4), 369. [CrossRef]
- Hart, P. J.; Reynolds, J. D.; Hart, P. J.; Reynolds, J. D. Handbook of fish biology and fisheries; Wiley Online Library, 2002.
- Franz, G. P.; Lewerentz, L.; Grunow, B. Observations of growth changes during the embryonic-larval-transition of pikeperch (Sander lucioperca) under near-natural conditions. Journal of Fish Biology 2021, 99 (2), 425-436.
- Rombough, P. The energetics of embryonic growth. Respiratory Physiology & Neurobiology 2011, 178 (1), 22-29.
- Wei, L.; Zhu, S.Q.; Liu, W.; Zhao, J.L.; Qian, Y.Z.; Wu, C.; Qian, D. Comparison on morphology and body spots characteristics between backcross progenies and their parents of mandarin fish. South China Fisheries Science 2020, 16 (2): 1-7.
- J. Marshall, D.; Uller, T. When is a maternal effect adaptive? Oikos 2007, 116 (12), 1957-1963.
- de Assis Lago, A.; Rezende, T. T.; Dias, M. A. D.; de Freitas, R. T. F.; Hilsdorf, A. W. S. The development of genetically improved red tilapia lines through the backcross breeding of two Oreochromis niloticus strains. Aquaculture 2017, 472, 17-22. [CrossRef]
- Liu, Y.; Wang, L.; Li, Z.; Li, L.; Chen, S.; Duan, P.; Wang, X.; Qiu, Y.; Ding, X.; Su, J. DNA Methylation and Subgenome Dominance Reveal the Role of Lipid Metabolism in Jinhu Grouper Heterosis. International Journal of Molecular Sciences 2024, 25 (17), 9740. [CrossRef]
- Li, Z.Q.; Cheng, M.L.; Wu, Y.P.; Zhang, J.J.; Li, Z.T.; Ma,W.H.; Pang, Z.F.; Zhai J.M.; Tian, Y.S. Early development of hybrids of Epinephelus lanceolatus(♀) × Epinephelus moara(♂) and growth characteristics of reciprocal crosses. Journal of Fisheries of China 2020, 44(3), 436-446.
Figure 1.
Embryonic development of KG × KGGG (1). a: Fertilized egg; b: 2-cell stage; c: 4-cell stage; d: 8-cell stage; e: 16-cell stage; f: 32-cell stage; g: 64-cell stage; h: 128-cell stage; i: Multi-cell stage; j: morula stage; k: High blastula stage; l: Low blastula stage; m: Early gastrula stage; n: Middle gastrula stage; o: Late gastrula stage; p: Embryo body stage.
Figure 1.
Embryonic development of KG × KGGG (1). a: Fertilized egg; b: 2-cell stage; c: 4-cell stage; d: 8-cell stage; e: 16-cell stage; f: 32-cell stage; g: 64-cell stage; h: 128-cell stage; i: Multi-cell stage; j: morula stage; k: High blastula stage; l: Low blastula stage; m: Early gastrula stage; n: Middle gastrula stage; o: Late gastrula stage; p: Embryo body stage.
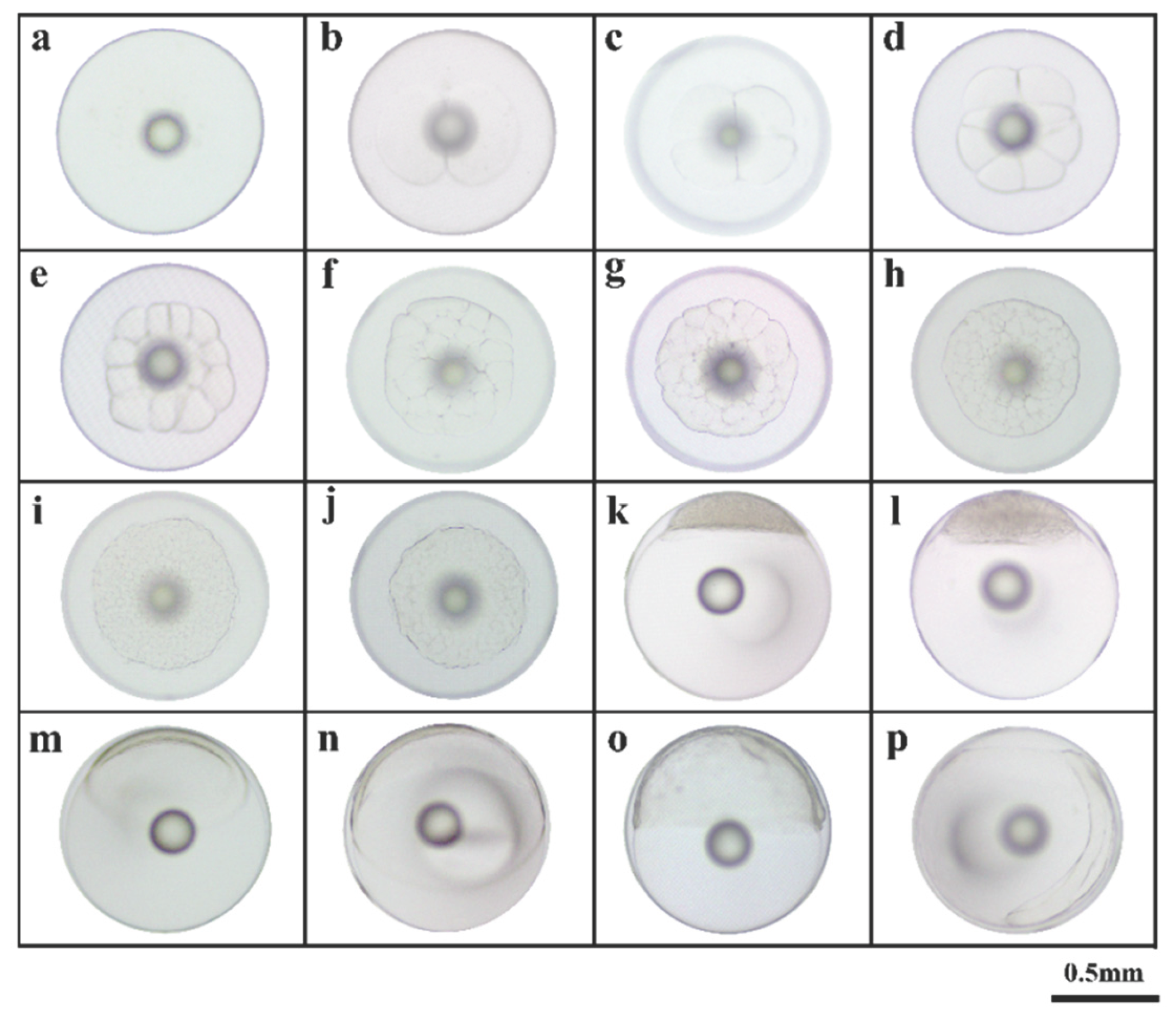
Figure 2.
Embryonic development of KG × KGGG (2). a: Closure of blastopore stage; b: Optic capsule stage; c: Muscle burl stage; d: Otocyst stage; e: Brain vesicle stage; f: Tail-bud stage; g: Crystal stage; h: Heart-beating stage; i: Pre incubation stage; j: Hatching stage; k: Newly hatched larvae; l: Larva of 1 day after hatching (dAH); m: Newly hatched larvae of KG.
Figure 2.
Embryonic development of KG × KGGG (2). a: Closure of blastopore stage; b: Optic capsule stage; c: Muscle burl stage; d: Otocyst stage; e: Brain vesicle stage; f: Tail-bud stage; g: Crystal stage; h: Heart-beating stage; i: Pre incubation stage; j: Hatching stage; k: Newly hatched larvae; l: Larva of 1 day after hatching (dAH); m: Newly hatched larvae of KG.
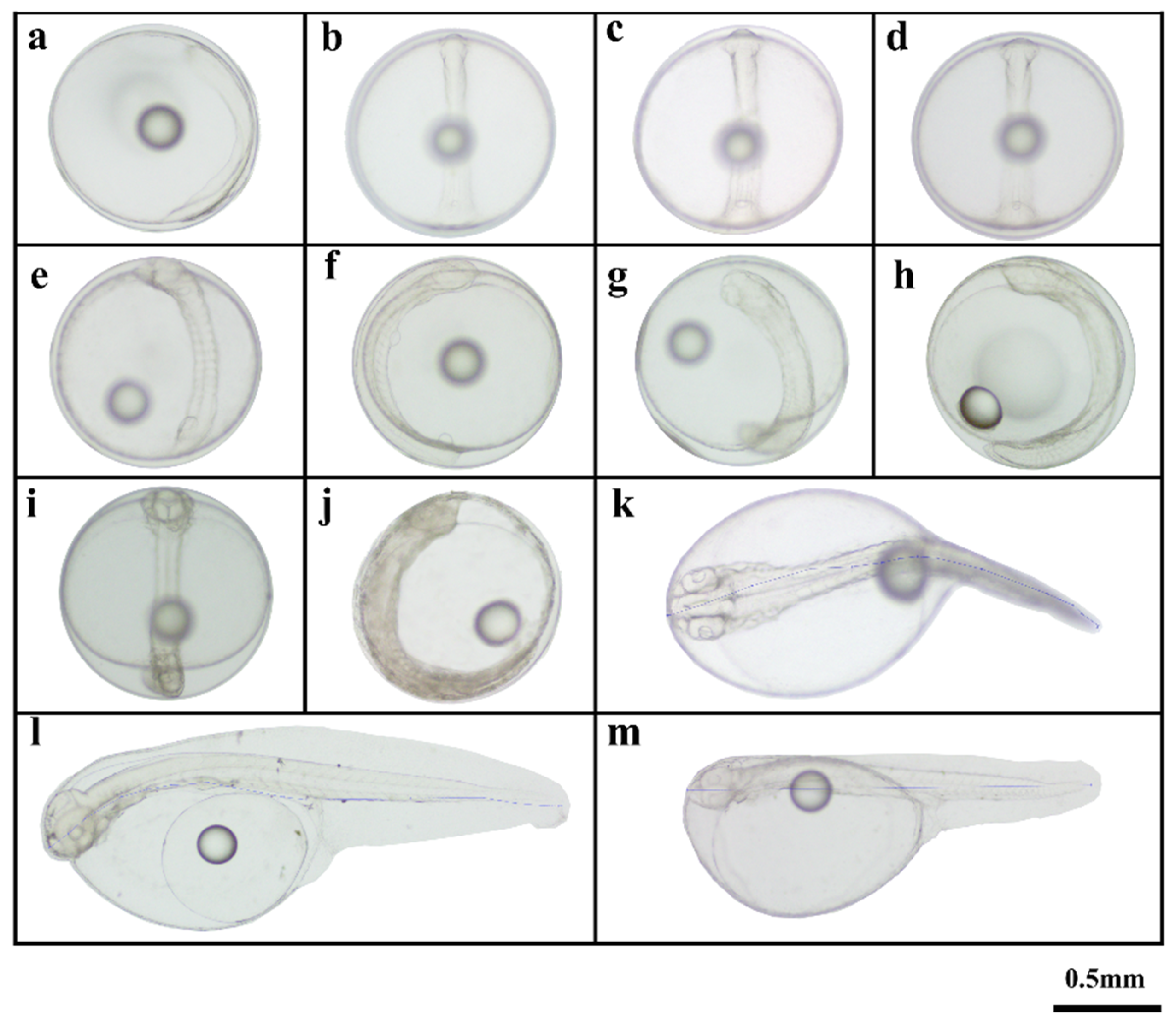
Figure 3.
Morphological development of larva juvenile and young fish of KG × KGGG. a: Larva of 2 dAH; b: Larva of 4 dAH; c: Larva of 9 dAH; d: Larva of 11 dAH; e: Larva of 13 dAH; f: Larva of 15 dAH; g: Larva of 20 dAH; h: Larva of 25 dAH; i: Juvenile of 35 dAH; j:Juvenile of 45 dAH; k: Juvenile of 50 dAH; l: Young fish of 60 dAH; m: Young fish of 70 dAH; n: Young fish of 80 dAH; o: Young fish of 90 dAH.
Figure 3.
Morphological development of larva juvenile and young fish of KG × KGGG. a: Larva of 2 dAH; b: Larva of 4 dAH; c: Larva of 9 dAH; d: Larva of 11 dAH; e: Larva of 13 dAH; f: Larva of 15 dAH; g: Larva of 20 dAH; h: Larva of 25 dAH; i: Juvenile of 35 dAH; j:Juvenile of 45 dAH; k: Juvenile of 50 dAH; l: Young fish of 60 dAH; m: Young fish of 70 dAH; n: Young fish of 80 dAH; o: Young fish of 90 dAH.

Figure 4.
Relationship between total length and age in KG × KGGG larvae.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



