Submitted:
27 August 2025
Posted:
01 September 2025
You are already at the latest version
Abstract
Norway spruce (Picea abies [L.] Karst.) is a primary forest-forming species in the European part of Russia, both in terms of its distribution and economic importance. A number of studies indicate that one of the reasons for the disturbance of spruce forests is linked to rising temperatures, particularly the detrimental effects of extreme droughts. In our study, we propose a hypothesis regarding the critical condition of mature spruce forests within the coniferous-broadleaved vegetation zone at the center of the East European Plain, the decline of which is escalating each year due to global climate change. The studies were conducted in intact spruce forests using resurveyed vegetation relevés within the Smolensk-Moscow Upland by repeating relevés after 40 years. We estimated beta diversity of species composition using the Mann-Whitney U test to study succession processes. Interspecific relationships were estimated using the Spearman criterion. The functional significance was assessed using the species activity index. Interpretation of the species composition of communities of different time periods based on indirect ordination methods (NMDS). Results showed that significant disturbances of the tree layer have led to changes in the vegetation of subordinate layers. An analysis of the complete species composition of spruce forests based on scoring assessments using Ellenberg's scales revealed changes in the ecological conditions of habitats over 40 years. A noticeable trend was observed towards an increase in the proportion of thermophilic species and those favoring less soil reaction, indicating a shift towards a nemoral vegetation spectrum. It is expected that without implementing additional forestry measures such as silviculture and thinning, the next 40 to 60 years will see a decline in the proportion of spruce within mixed stands, potentially culminating in the complete collapse of monospecific spruce forests in the center of the East European Plain.
Keywords:
1. Introduction
2. Materials and Methods
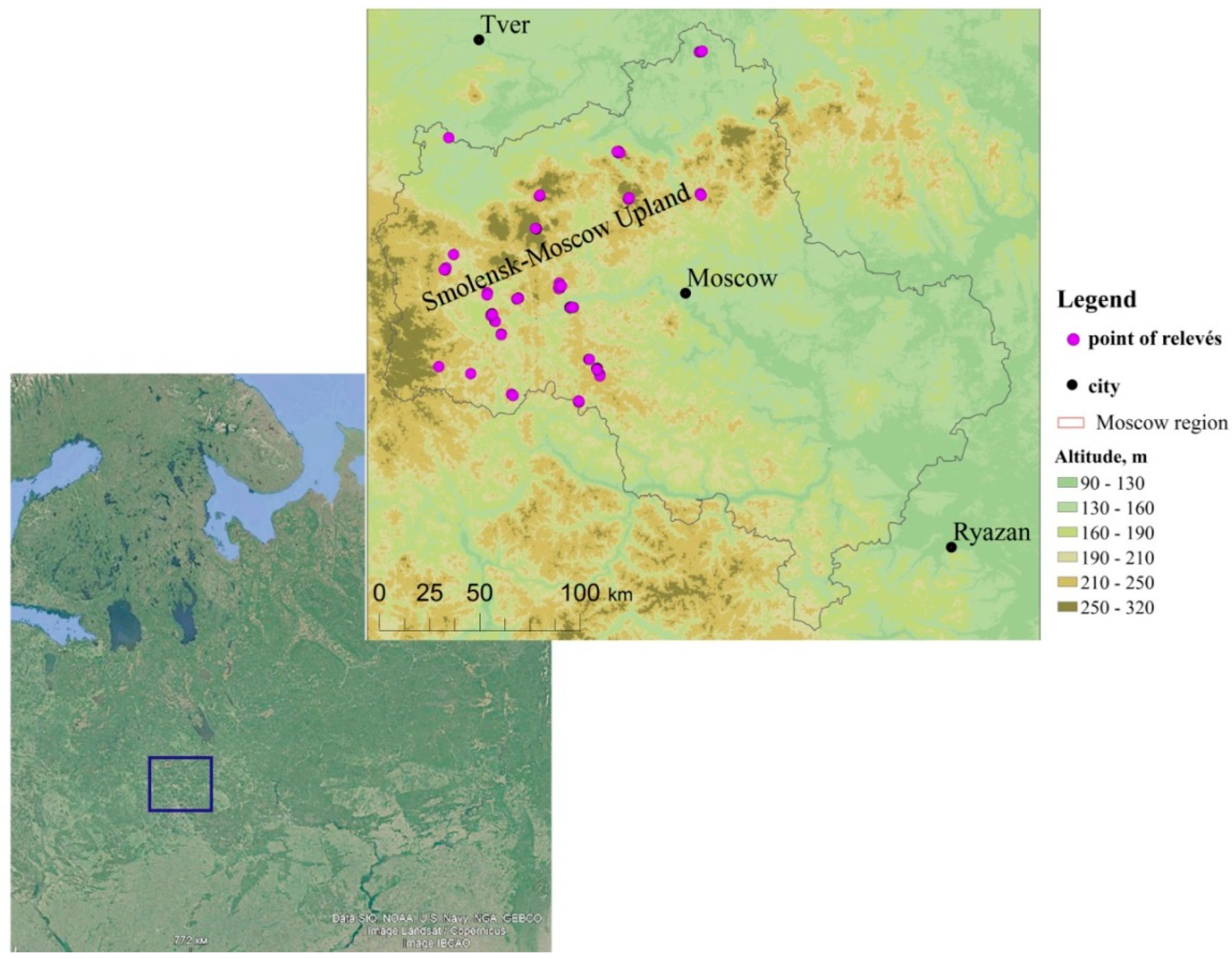
2.1. Study Area
2.2. Data Acquisition
2.3. Data Analysis
3. Results
3.1. Changes in Structural and Functional Properties in Spruce Forests
3.2. Changes in Common Patterns of Tree and Shrub Layers
3.3. Changes in Common Patterns in Field and Ground Layers
3.4. Relationship with Environmental Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ECG | Ecological and cenotic group |
| Br | Boreal |
| PC | Projective cover |
| Nm | Nemoral |
| NW | Nitrophilic-wet |
| Md | Meadow |
| Eg | Edge-herb |
| Ad | Adventive |
| NMDS | Non-metric multidimensional scaling |
| L | Light |
| N | Nitrogen richness |
| R | Soil reaction |
| M | Soil moisture |
| d.s. | data set |
References
- Jandt, U.; Bruelheide, H.; Jansen, F.; Bonn, A.; Grescho, V.; Klenke, R.A.; Sabatini, F.M.; Bernhardt-Römermann, M.; Blüml, V.; Dengler, J.; et al. More Losses than Gains during One Century of Plant Biodiversity Change in Germany. Nature 2022, 611, 512–518. [CrossRef]
- Bolam, F. Over Half of Threatened Species Require Targeted Recovery Actions to Avert Human-induced Extinction. Frontiers in Ecology and the Environment 2023, 22, 64–70. [CrossRef]
- Zhang, L.; Ameca, E.I.; Otero-Jimenez, B.; Montaño, S.K.; Shea, A.; Kelly, T.; Andrianoely, D.; Wright, P.C. Human-Induced Deforestation Increases Extinction Risk Faster than Climate Pressures: Evidence from Long-Term Monitoring of the Globally Endangered Milne-Edward’s Sifaka. Biological Conservation 2022, 274, 109716.
- Dimitrova, A.; Csilléry, K.; Klisz, M.; Lévesque, M.; Heinrichs, S.; Cailleret, M.; Andivia, E.; Madsen, P.; Böhenius, H.; Cvjetkovic, B.; et al. Risks, Benefits, and Knowledge Gaps of Non-Native Tree Species in Europe. Front. Ecol. Evol. 2022, 10, 908464. [CrossRef]
- Ellis, E.C.; Antill, E.C.; Kreft, H. All Is Not Loss: Plant Biodiversity in the Anthropocene. PLOS ONE 2012, 7, e30535. [CrossRef]
- Klinkovská, K.; Sperandii, M.G.; Knollová, I.; Danihelka, J.; Hájek, M.; Hájková, P.; Hroudová, Z.; Jiroušek, M.; Lepš, J.; Navrátilová, J.; et al. Half a Century of Temperate Non-Forest Vegetation Changes: No Net Loss in Species Richness, but Considerable Shifts in Taxonomic and Functional Composition. Glob Chang Biol 2025, 31, e70030. [CrossRef]
- Pakeman, R.J.; Lepš, J.; Kleyer, M.; Lavorel, S.; Garnier, E.; the VISTA consortium Relative Climatic, Edaphic and Management Controls of Plant Functional Trait Signatures. J Vegetation Science 2009, 20, 148–159. [CrossRef]
- Martinez del Castillo, E.; Zang, C.; Buras, A.; Hacket-Pain, A.; Esper, J.; Serrano-Notivoli, R.; Hartl, C.; Weigel, R.; Klesse, S.; Resco de Dios, V.; et al. Climate-Change-Driven Growth Decline of European Beech Forests. Communications Biology 2022, 5. [CrossRef]
- Viljur, M.-L.; Abella, S.; Adámek, M.; Alencar, J.; Barber, N.; Beudert, B.; Burkle, L.; Cagnolo, L.; Campos, B.; Chao, A.; et al. The Effect of Natural Disturbances on Forest Biodiversity: An Ecological Synthesis. Biological reviews of the Cambridge Philosophical Society 2022, 97, 1930–1947. [CrossRef]
- Rosenfeld, J. Functional Redundancy in Ecology and Conservation. Oikos 2002, 98.
- Functional Plant Ecology; 2nd Edition; edited by Francisco Pugnaire and Fernando Valladares.; Taylor & Francis Group, 2007;
- Nock, C.; Vogt, R.; Beisner, B. Functional Traits. In; 2016 ISBN 978-0-470-01617-6.
- Vasilevich, V. Functional diversity in plant communities. Botanical Journal 2016, 101, 776–795. [CrossRef]
- Bakker, J.P.; Olff, H.; Willems, J.H.; Zobel, M. Why Do We Need Permanent Plots in the Study of Long-Term Vegetation Dynamics? Journal of Vegetation Science 1996, 7, 147–155. [CrossRef]
- Chytry, M.; Tichý, L.; Hennekens, S.; Schaminée, J. Assessing Vegetation Change Using Vegetation-Plot Databases: A Risky Business. Applied Vegetation Science 2014, 17. [CrossRef]
- Kapfer, J.; Hédl, R.; Jurasinski, G.; Kopecký, M.; Schei, F.H.; Grytnes, J.-A. Resurveying Historical Vegetation Data - Opportunities and Challenges. Appl Veg Sci 2016, 20, 164–171. [CrossRef]
- Kneeshaw, D.; Gauthier, S. Old Growth in the Boreal Forest: A Dynamic Perspective at the Stand and Landscape Level | Request PDF. Environmental Reviews 2003, 11, 99–114. [CrossRef]
- Mosseler, A.; Thompson, I.; Pendrel, B. Overview of Old-Growth Forests in Canada from a Science Perspective. Environmental Reviews 2011, 11. [CrossRef]
- Rees, M.; Condit, R.; Crawley, M.; Pacala, S.; Tilman, D. Long-Term Studies of Vegetation Dynamics. Science 2001, 293, 650–655. [CrossRef]
- Litvinenko, L.; Kalinina, A. Territorial Distribution of Precipitation in the Moscow Region in the Presence and Absence of the Large Anthropogenic Formation. Ecology of Urban Areas 2018.
- Hytteborn, H.; Maslov, A.A.; Nazimova, D.I.; Rysin, L.P. Boreal Forests of Eurasia. In Coniferous forests; etc.: Elsevier: Amsterdam, 2005; pp. 23–99.
- Maslov, A. Monitoring biodiversity and natural dynamics processes in protected forest areas: program and results of work over 25 years. In Structure and functions of forests in European Russia; Scientific Publications Association KMK: Moscow, 2009; pp. 172–190 ISBN 978-5-87317-585-7.
- Plans of the Russian Empire Land Survey 1777.
- Three-Verst Military Topographic Map of the Russian Empire 1850.
- Topographic military maps of Red Army (RKKA) 1936.
- Mirkin, B. Theoretical basis of modern phytocenology; Nauka.; USSR Academy of Sciences, Bashkir branch, Institute of Biology: Moscow, 1985;
- Rodwell, J.S.; Mucina, L.; Pignatti, S.; Schaminée, J.H.J.; Chytry, M. European Vegetation Survey; the Context of the Case Studies. Folia geobotanica et phytotaxonomica 1997, 32, 113–115.
- Moore, D.M. Flora Europaea Check-List and Chromosome Index; Cambridge Univ. Press: Cambridge., 1982;
- Ignatov, M.; Ignatova, E. Moss Flora of the Middle Eropean Russia; KMK: Moscow, 2003; Vol. 1; ISBN 5-87317-149-1.
- Maslov, A. Biodiversity of the Native Forest Types in Strict Scientific Forest Reserves of the Moscow Region. Russian journal of forest science 2022, 631–342. [CrossRef]
- Smirnov, V.; Khanina, L.; Bobrovsky, M. Validation of the ecological-coenotical groups of vascular plant species for European Russian forests on the basis of ecological indicator values, vegetation releves and statistical analysis. Bulletin of Moscow Society of Naturalists 2006, 111, 36–47.
- Mucina, L.; Grabherr, G.; Wallnöfer, S. Die Pflanzengesellschaften Österrreichs. Teil III; Wälder und Gebüsche: Jena, 1993;
- Malyshev, L. Floristic Zoning Based on Quantitative Characteristics. Botanical Journal 1973, 1581–1588.
- Chernenkova, T.V.; Kotlov, I.P.; Belyaeva, N.G.; Suslova, E.G.; Morozova, O.V. Assessment and mapping of the cenotic diversity of the Moscow region’s forest. Russian Journal of Forest Science (Lesovedenie) 2022. [CrossRef]
- Tichý, L. JUICE, Software for Vegetation Classification. Journal of Vegetation Science 2002, 13, 451–453. [CrossRef]
- R Core Team 2020.
- Remmert, H. The Mosaic-Cycle Concept of Ecosystems — An Overview. In The Mosaic-Cycle Concept of Ecosystems; Springer, Berlin, Heidelberg, 1991; pp. 1–21 ISBN 978-3-642-75650-4.
- Král, K.; Vrška, T.; Hort, L.; Adam, D.; Šamonil, P. Developmental Phases in a Temperate Natural Spruce-Fir-Beech Forest: Determination by a Supervised Classification Method. Eur J Forest Res 2010, 129, 339–351. [CrossRef]
- Bayer, D.; Pretzsch, H. Reactions to Gap Emergence: Norway Spruce Increases Growth While European Beech Features Horizontal Space Occupation–Evidence by Repeated 3D TLS Measurements. Silva Fennica 2017, 51.
- Neal, J.; Hawker, L. FABDEM V1-2 2023.
- Šamonil, P.; Schaetzl, R.J.; Valtera, M.; Goliáš, V.; Baldrian, P.; Vašíčková, I.; Adam, D.; Janík, D.; Hort, L. Crossdating of Disturbances by Tree Uprooting: Can Treethrow Microtopography Persist for 6000 Years? Forest Ecology and Management 2013, 307, 123–135. [CrossRef]
- Schelhaas, M.; Nabuurs, G.; Schuck, A. Natural Disturbances in the European Forests in the 19th and 20th Centuries. Global Change Biology 2003, 9, 1620–1633. [CrossRef]
- Maslov, A. BARK BEETLE AND DRYING OUT OF SPRUCE FORESTS; FBU VNIILM: Moscow, 2010; ISBN 978-5-94219-170-2.
- Korotkov, S.A. Change in the Composition of Tree Stands and the Stability of Protective Forests in the Central Part of the Russian Plain; Moscow, 2023;
- Malakhova, E.; Lyamtsev, N. Extent and Structure of Moscow Region Spruce Forest Dieback in 2010-2012. Izvestia Sankt-Peterburgskoj lesotehniceskoj akademii 2014, 193–201.
- Maslov, A.; Komarova, I.; Kotov, A. The Dynamics of Reproduction of Bark Beetles in Central Russia in 2010-2013. And the Forecast for 2014. Forestry information 2014, 38–46.
- Pugachevsky, A. Spruce coenopopulations. Structure, dynamics, regulatory factors; Nauka i Tekhika.; Minsk, 1992; ISBN 5-343-00783-X.
- Purves, D.W.; Lichstein, J.W.; Pacala, S.W. Crown Plasticity and Competition for Canopy Space: A New Spatially Implicit Model Parameterized for 250 North American Tree Species. PLoS ONE 2007, 2, e870. [CrossRef]
- Karpov, V. Experimental Phytocenology of Taiga; Nauka.; Leningrad Department: Leningrad, 1969;
- Alekseev, V. Light Regime of the Forest; Nauka.; Leningrad, 1975;
- Dyrenkov, S. Structure and dynamics of taiga spruce forests; Nauka: Leningrad, 1984;
- Rysin, L.; Savelyeva, L. Cadastres of forest types and forest biogeocenoses; Scientific publications partnership KMK: Moscow, 2007; ISBN 978-5-87317-397-6.
- Third Assessment Report on Climate Change and Its Consequences in the Russian Federation; Наукoемкие технoлoгии.; Saint Petersburg, 2022;
- Kobyakov, K.; Titova, S.; Shmatkov, N.; Korotkov, V.; Kazakov, P. Assessment of Possibilities for Increasing Greenhouse Gas Absorption by Forests in the Central European Russia. Sustainable forest management 2019, 4–20.
- Maslov, A.A. Dynamics of Ecological Species Groups and Forest Types during Natural Successions in Central Russia Preserved Forests. Bulletin of Moscow Society of Naturalists. Biological series 1998, 103, 34–43.
- Enyedi, Z.M.; Ruprecht, E.; Deák, M. Long-term Effects of the Abandonment of Grazing on Steppe-like Grasslands. Applied Vegetation Science 2008, 11, 55–62. [CrossRef]
- Ruprecht, E.; Enyedi, M.Z.; Eckstein, R.L.; Donath, T.W. Restorative Removal of Plant Litter and Vegetation 40 Years after Abandonment Enhances Re-Emergence of Steppe Grassland Vegetation. Biological Conservation 2010, 143, 449–456. [CrossRef]
- Klinkovská, Sperandii et Al. Available online: https://scholar.google.com/scholar?lookup=0&q=Klinkovsk%C3%A1,+Sperandii+et+al.+2024&hl=en&as_sdt=0,9&as_vis=1 (accessed on 28 March 2025).
- Chételat, J.; Kalbermatten, M.; Lannas, K.S.M.; Spiegelberger, T.; Wettstein, J.-B.; Gillet, F.; Peringer, A.; Buttler, A. A Contextual Analysis of Land-Use and Vegetation Changes in Two Wooded Pastures in the Swiss Jura Mountains. E&S 2013, 18, art39. [CrossRef]
- Peringer, A.; Siehoff, S.; Chételat, J.; Spiegelberger, T.; Buttler, A.; Gillet, F. Past and Future Landscape Dynamics in Pasture-Woodlands of the Swiss Jura Mountains under Climate Change. Ecology and Society 2013, 18.
- Konatowska, M.; Młynarczyk, A.; Rutkowski, P.; Kujawa, K. Impact of Site Conditions on Quercus Robur and Quercus Petraea Growth and Distribution under Global Climate Change. Remote Sensing 2024, 16, 4094.
- Pretzsch, H. Forest Dynamics, Growth and Yield: From Measurement to Model; Springer Berlin Heidelberg: Berlin, Heidelberg, 2009; ISBN 978-3-540-88306-7.
- Hari, P. Theoretical Aspects of Eco-Physiological Research. In Crop physiology of forest trees.; Helsinki, 1985; pp. 21–30.
- Rabotnov, T. Phytocenology: Textbook for universities in the field of “Biology” and specialty “Botany”; Moscow State University Publishing House: Moscow, 1992; ISBN 5-211-02401-X.
- Korotkov, V. Basic Concepts and Methods of Restoration of Natural Forests in Eastern Europe. Russian Journal of Ecosystem Ecology 2017, 2, 1–18. [CrossRef]
- Chernenkova, T.V.; Kotlov, I.P.; Belyaeva, N.G.; Suslova, E.G. Spatiotemporal Modeling of Coniferous Forests Dynamics along the Southern Edge of Their Range in the Central Russian Plain. Remote Sensing 2021, 13. [CrossRef]
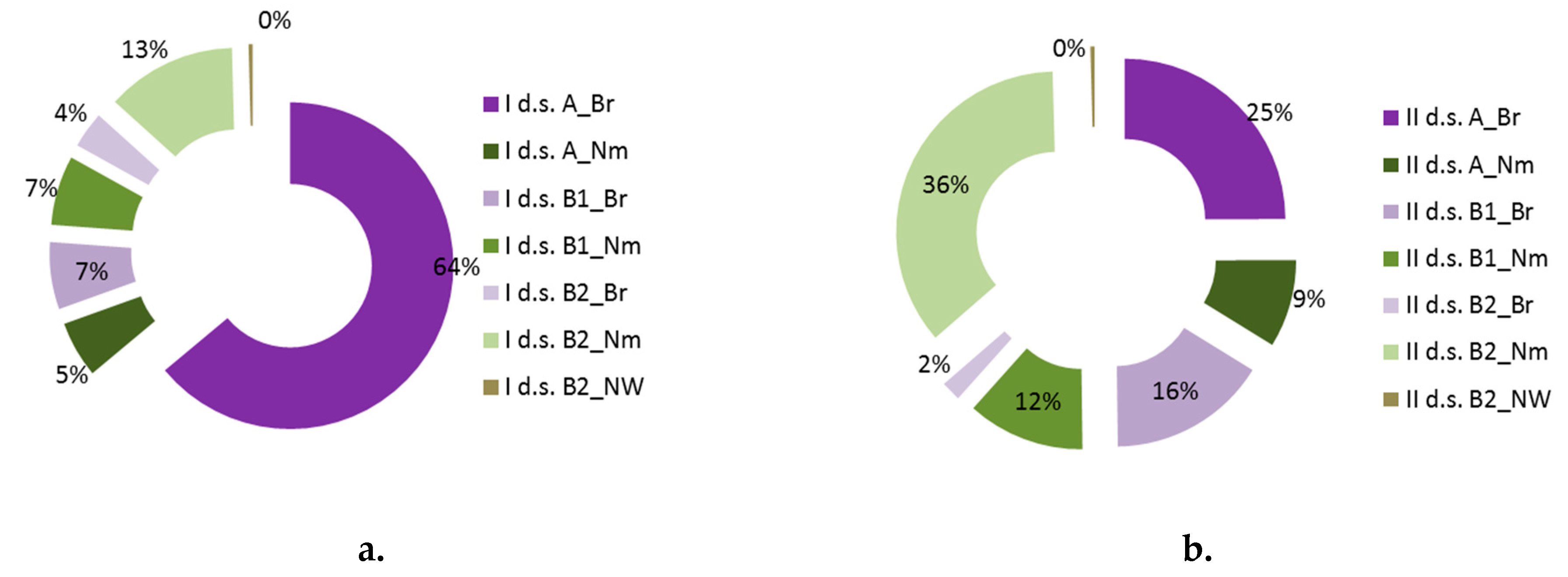
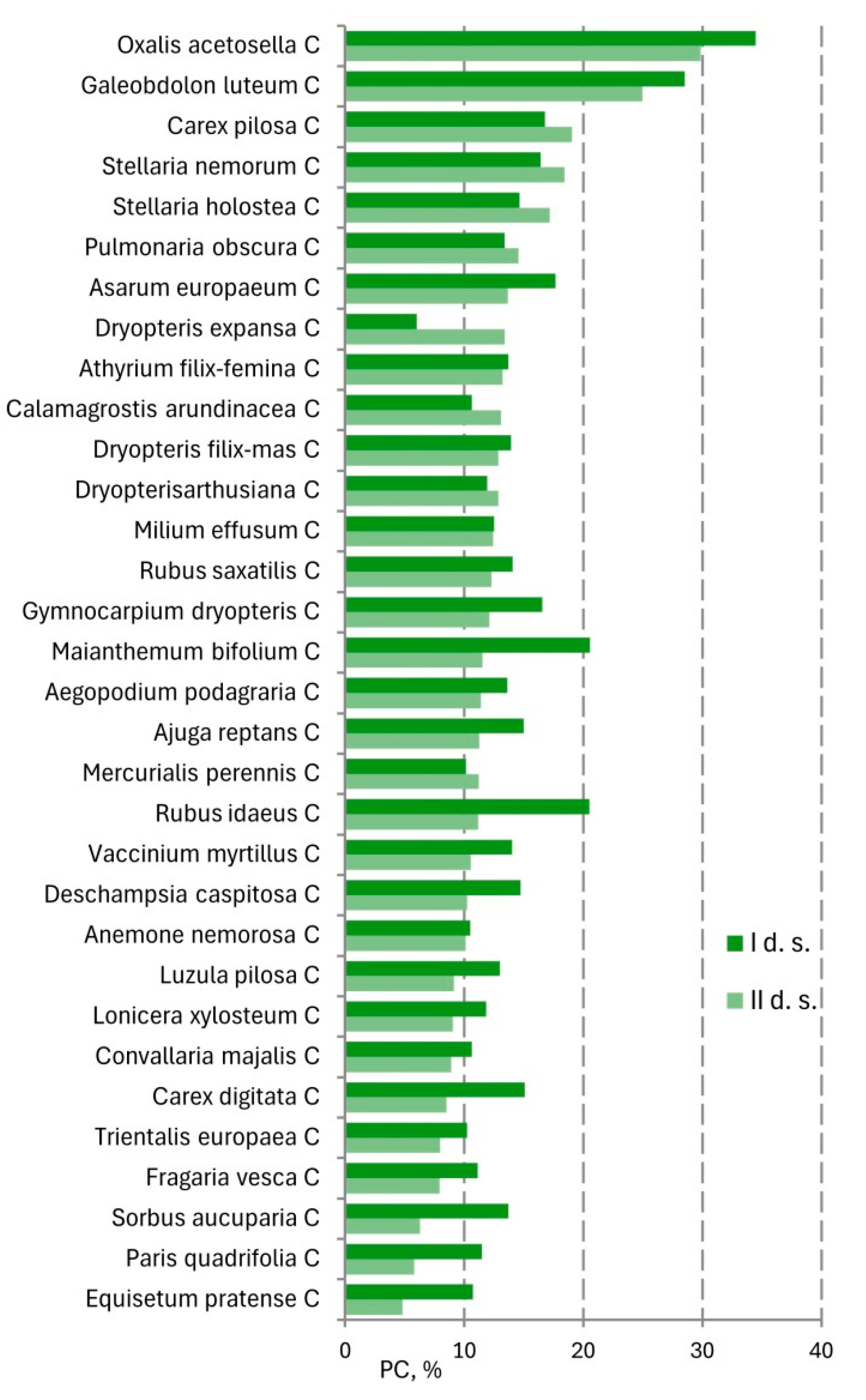
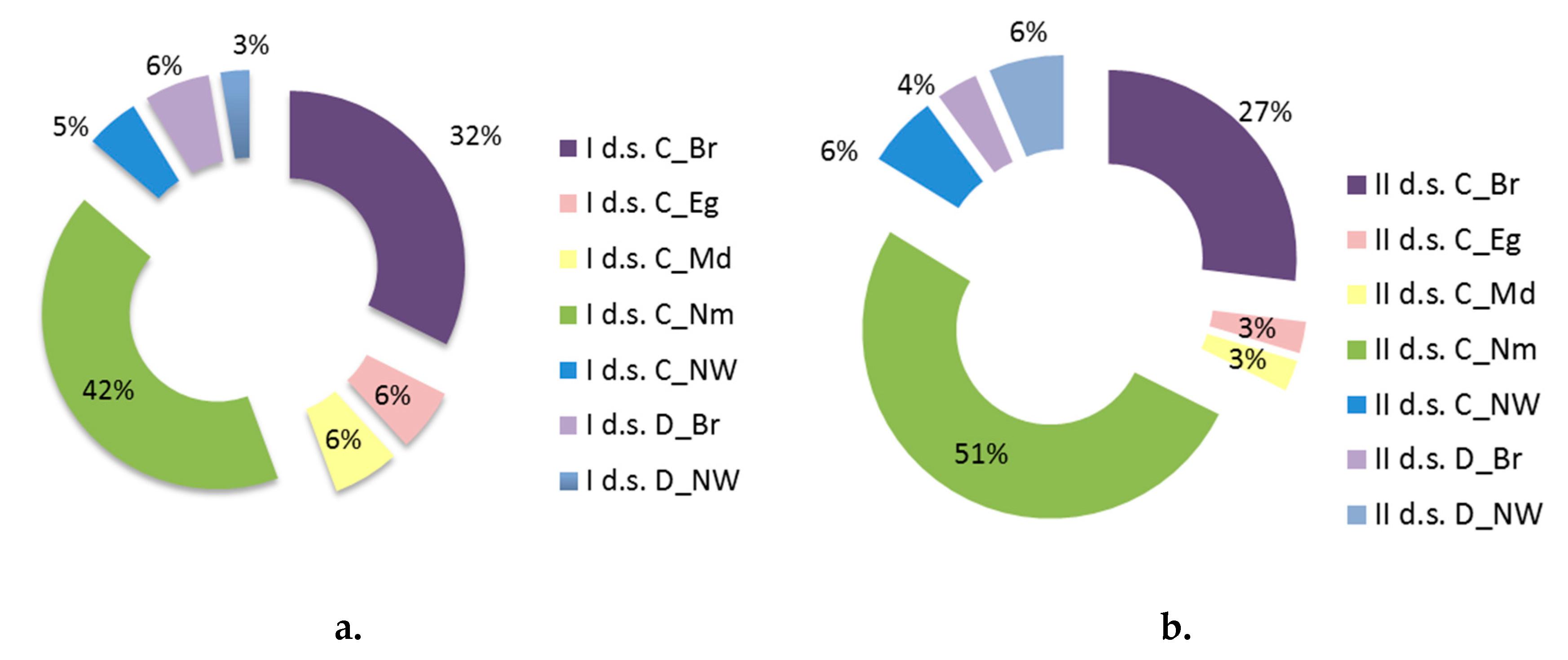
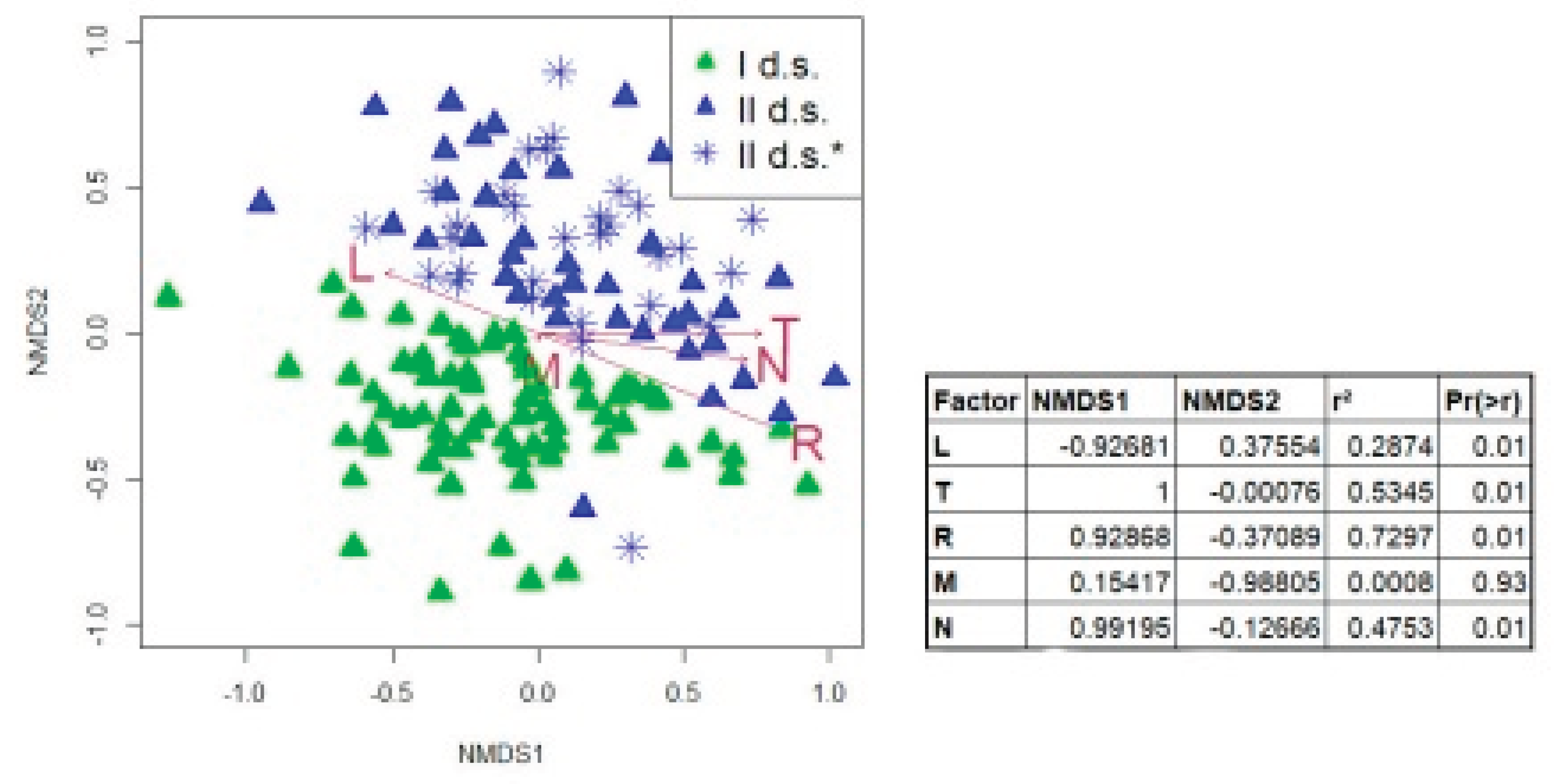

| Layers | I d.s. | II d.s. | p | ||
|---|---|---|---|---|---|
| M, % | S.D. | M, % | S.D. | ||
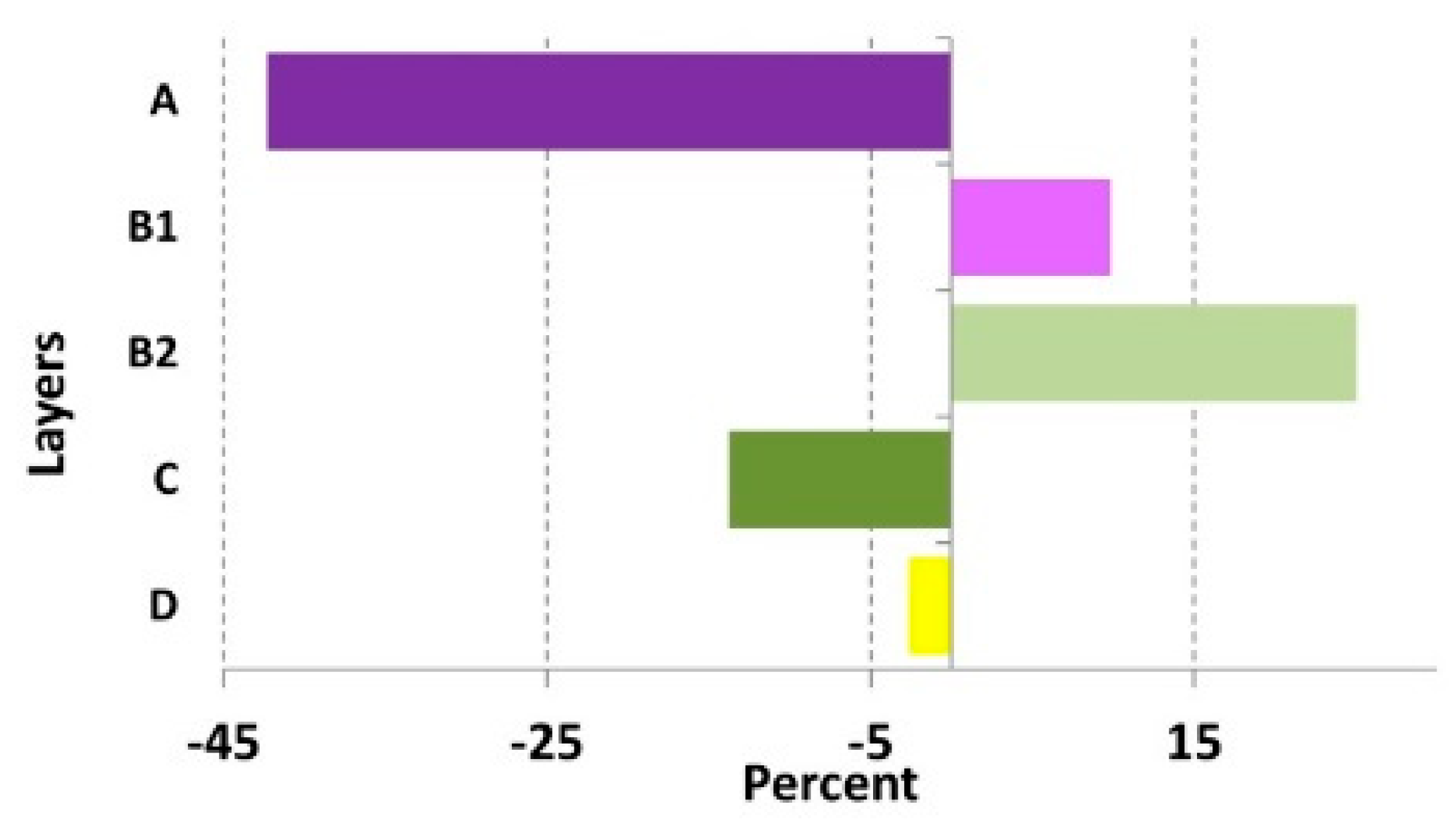
| A | 71.4 | 9.2 | 29.1 | 22.4 | 0.000 |
| B1 | 11.0 | 13.7 | 20.8 | 16.8 | 0.000 |
| B2 | 12.2 | 16.6 | 37.2 | 24.2 | 0.000 |
| C | 76.7 | 17.6 | 62.9 | 18.2 | 0.012 |
| D | 16.6 | 25.1 | 14.0 | 21.7 | 0,000 |
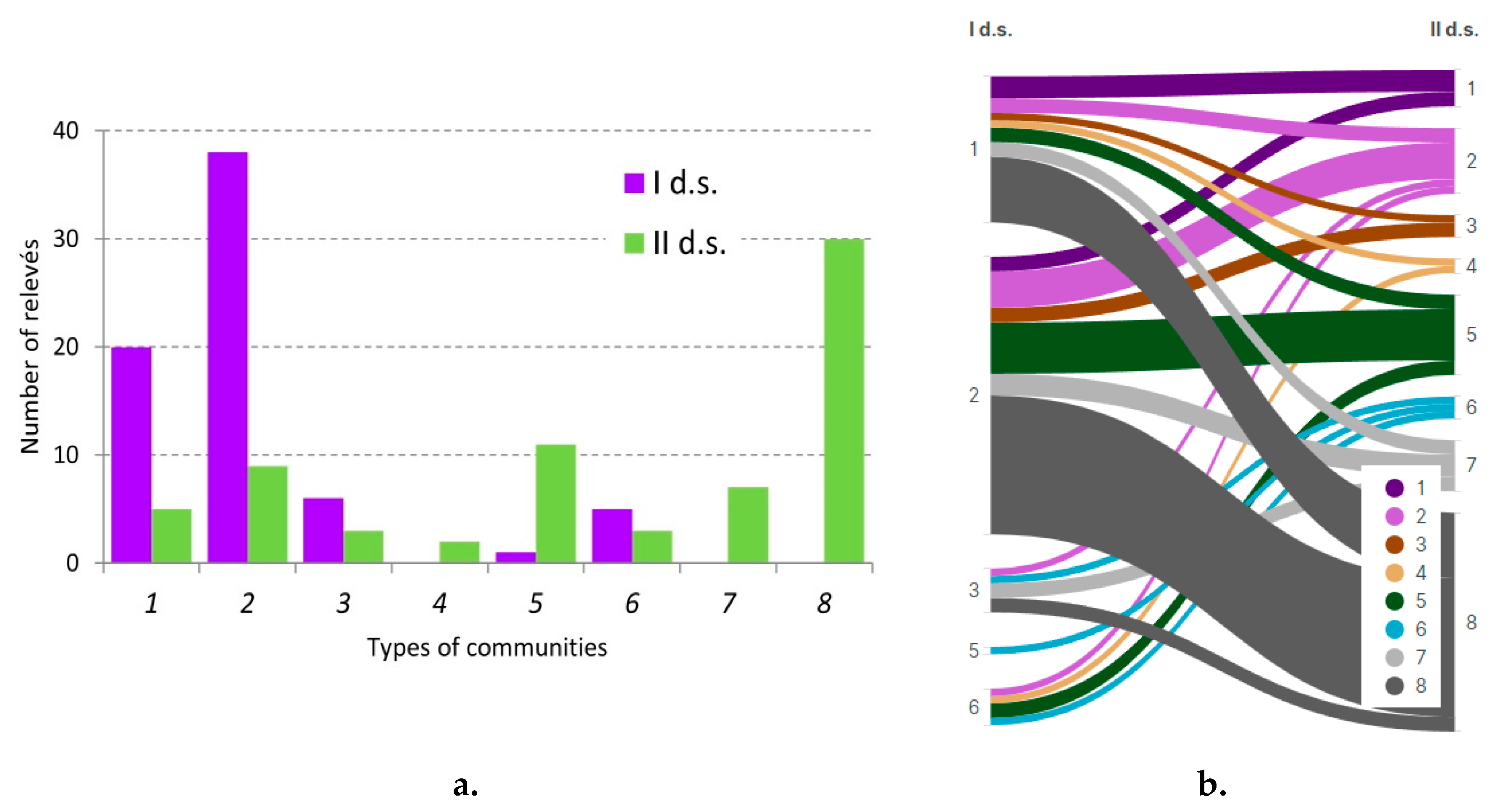
| №№ | Community types |
|---|---|
| 1 | Spruce with birch, aspen forests dwarf shrubs–small herb–green moss and small herb (Vaccinium myrtillus, V. vitis idaea, Oxalis acetosella, Dryopteris carthusiana, Calamagrostis arundinacea, Luzula pilosa, Carex digitata, Orthilia secunda, Pleurozium schreberi, Hylocomium splendens, Rhytidiadelphus triquetrus) |
| 2 | Spruce with birch, aspen, oak and linden forests small herb–broad herb and broad herb (Stellaria_holostea, Aegopodium podagraria, Carex pilosa, Anemonoides nemorosa, Oxalis acetosella, Veronica chamaedrys, Carex pilosa, Ajuga reptans, Lamiastrum galeobdolon, Atrichum undulatum) |
| 3 | Spruce-pine with birch forests dwarf shrubs–small herb–green moss and small herb (Vaccinium myrtillus, Vaccinium vitis-idaea, Oxalis acetosella, Dryopteris carthusiana, Calamagrostis arundinacea, Convallaria majalis, Pleurozium schreberi, Hylocomium splendens) |
| 4 | Spruce-pine with birch forests small herb–broad herb and broad herb (Corylus avellana, Oxalis acetosella, Carex pilosa, Lamiastrum galeobdolon, Athyrium filix-femina, Dryopteris carthusiana). |
| 5 | Oak-linden forests broad herbs (Aegopodium podagraria, Carex pilosa, Anemonoides ranunculoides, Galeobdolon luteum, Mercurialis perennis, Lamiastrum galeobdolon, Dryopteris filix-mas, Pulmonaria obscura, Asarum europaeum, Ranunculus cassubicus, Stellaria nemorum, Aconitum septentrionale) |
| 6 | Birch-aspen forests with broad herb (Aegopodium podagraria, Ranunculus_cassubicus, Carex pilosa, Glechoma_hirsuta, Equisetum_pratense, Lamiastrum galeobdolon, Pulmonaria obscura, Stellaria nemorum, Calamagrostis arundinacea) |
| 7 | Disintegrated spruce forests with spruce undergrowth and dwarf shrubs–small herb–green moss and small herb (Equisetum sylvaticum, E. pratense, Lysimachia vulgaris, Circaea alpina, Dryopteris expansa, Filipendula ulmaria, Trientalis europaea, Luzula pilosa, Orthilia secunda, Climacium dendroides) |
| 8 | Disintegrated spruce forests with hazel and small herb–broad herb and broad herb (Corylus avellana, Stellaria nemorum, Carex sylvatica, Athyrium filix-femina, Rubus idaeus, Dryopteris carthusiana) |
| Species | I d.s. | II d.s. | F II/I | p | ||
|---|---|---|---|---|---|---|
| M | S.D. | M | S.D. | |||
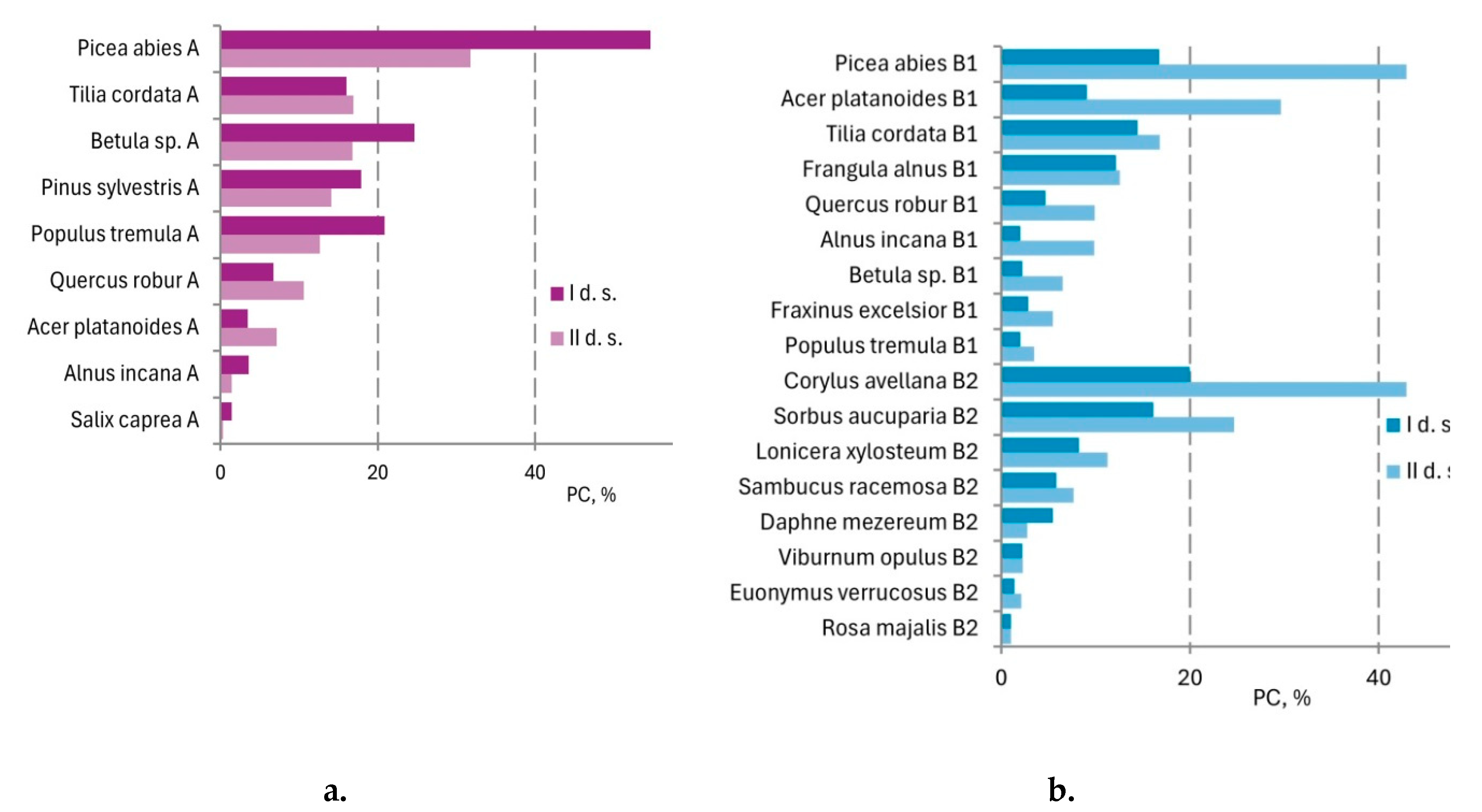
| Acer platanoides | 0.17 | 0.66 | 0.73 | 3.2 | 0.56 | 0.000* |
| Betula species | 8.70 | 19.8 | 4.03 | 7.9 | -4.67 | 0.006* |
| Picea abies | 42.76 | 11.6 | 16.16 | 16.8 | -26.60 | 0.000* |
| Pinus sylvestris | 4.56 | 11.0 | 2.84 | 6.9 | -1.71 | 0.415 |
| Populus tremula | 6.23 | 1.5 | 1.60 | 7.6 | -4.63 | 0.001* |
| Quercus robur | 0.64 | 0.17 | 2.29 | 7.3 | 1.64 | 0.753 |
| Tilia cordata | 3.67 | 0.66 | 4.11 | 11.4 | 0.44 | 0.780 |
| Species | I d.s. | II d.s. | F II/I | p | ||
|---|---|---|---|---|---|---|
| M | S.D. | M | S.D. | |||
| Acer platanoides | 1.17 | 4.15 | 4.03 | 7.4 | 2.86 | 0.010* |
| Betula sp. | 0.07 | 0.31 | 0.43 | 1.6 | 0.36 | 0.532 |
| Picea abies | 4.00 | 6.5 | 13.7 | 14.8 | 9.7 | 0.000* |
| Populus tremula | 0.1 | 0.23 | 0.1 | 0.46 | 0 | 0.879 |
| Quercus robur | 0.31 | 1.9 | 1.6 | 4.0 | 1.29 | 0.000* |
| Tilia cordata | 2.97 | 9.9 | 3.0 | 7.8 | 0.03 | 0.553 |
| Species | I d.s. | II d.s. | F II/I | p | ||
|---|---|---|---|---|---|---|
| M | S.D. | M | S.D. | |||
| Corylus avellana | 5.70 | 11.7 | 26.36 | 23.7 | 20.66 | 0.000 * |
| Daphne mezereum | 0.43 | 0.91 | 0.11 | 0.62 | -0.32 | 0.000 * |
| Euonymus verrucosa | 0.03 | 0.17 | 0.06 | 0.29 | 0.03 | 0.475 |
| Frangula alnus | 2.10 | 5.5 | 1.40 | 2.6 | -0.70 | 0.218 |
| Lonicera xylosteum | 0.97 | 3.2 | 1.82 | 2.9 | 0.85 | 0.000 * |
| Sambucus racemosa | 0.49 | 1.3 | 0.83 | 2.6 | 0.35 | 0.572 |
| Sorbus aucuparia | 3.70 | 7.3 | 8.68 | 12.5 | 4.98 | 0.000 * |
| Viburnum opulus | 0.07 | 0.26 | 0.08 | 0.31 | 0.00 | 0.788 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



