
Submitted:
27 August 2025
Posted:
28 August 2025
You are already at the latest version
Abstract
The increase in incidence and antibiotic-resistant strains show a need for a broadly protective vaccine to prevent gonorrhea. OMVax has develop a combination vaccine based on native outer membrane vesicles (NOMV) from two Neisseria meningitidis (Nm) and two Neisseria gonorrhoeae (Ng) strains. The strains had the acyl transferase LpxL1 knocked out to increase safety and reduction modifiable protein was also knocked out in the Ng strains. Factor H binding protein (FHbp) mutants with reduced Factor H (FH) binding from subfamilies A and B, respectively, were overexpressed in the Nm strains. The Ng strains individually expressed porin PorB.1a or PorB.1b. Antibodies elicited by the Nm-Ng NOMV vaccine had SBA with human complement against diverse Nm and Ng strains grown in the presence of CMP-NANA, had no significant reduction in SBA compared to the respective individual vaccines, inhibited adhesion to human cervical and vaginal cells by five out of six Ng strains tested, and inhibited Nm and Ng colonization in a transgenic mouse model. In conclusion, the Nm-Ng NOMV vaccine has the potential to protect against disease and inhibit colonization by diverse Nm and Ng strains, which may be an advantage for controlling disease through vaccination, particularly in the adolescent/young adult age group.
Keywords:
factor H binding protein
; PorB
; CEACAM1
; colonization
; transgenic mouse model
1. Introduction
Gonorrhea caused by the bacterial pathogen Neisseria gonorrhoeae (Ng), is currently the second most common sexually transmitted infection (STI) world-wide with an incidence of >600,000 cases per year in the US and ~82 million globally. Ng colonizes the mucosa of men and women but disproportionately affects the health of women resulting in reduced fertility and other complications of the upper and lower genital tract. The general increase in the incidence of disease and emergence of multi-drug-resistant strains has increased concern for the potential of untreatable gonorrhea. While there is no vaccine specifically designed to prevent gonorrhea, epidemiological studies conducted by Petousis-Harris and co-workers in New Zealand revealed that a detergent-extracted outer membrane vesicle vaccine (dOMV) produced from closely related Neisseria meningitidis bacteria (MeNZB) had 31% efficacy in preventing gonorrhea [1]. The MeNZB vaccine was developed by Chiron, Corp. as a tailor-made vaccine to control a meningococcal disease epidemic caused by Neisseria meningitidis serogroup B (MenB) [2]. The effect of MeNZB on reducing gonorrhea in New Zealand was subsequently confirmed for other MenB OMV-containing vaccines [3,4,5] including the current market leading MenB vaccine, 4CMenB (Bexsero®) [6], and led to the development of several new vaccines for Ng based on Ng OMV [7,8,9]. However, a recent Phase 2 clinical trial of an Ng OMV vaccine produced by Galaxo Smith Kline (GSK) failed to meet pre-defined efficacy criteria and it is unclear whether the vaccine will continue to be developed by the company [10].
Currently there are two MenB vaccines licensed in the U.S, 4CMenB and MenB-FHbp (Trumenba®). Both MenB vaccines, however, provide incomplete meningococcal strain coverage, and elicit protection of relatively short duration [11,12]. In particular, 4CMenB was predicted to not provide protection against strains causing outbreaks in countries where it is used for routine vaccination of infants such as the UK, France, Spain, and Canada. Recently, MenB vaccine strain coverage in these countries was reported to be as little as 5.3% against emerging clonal complexes based on prediction by the genetic Meningococcal Antigen Typing System [13].
Recognizing the limitations of current MenB vaccines, OMVax has developed a next generation MenB vaccine based on native outer membrane vesicles (NOMV) with over-expressed mutant Factor H binding proteins (FHbp) from subfamilies A and B with reduced Factor H binding [14,15]. In addition, the NOMV were made safer by eliminating sialic acid antigens that mimic host antigens and by deleting the acyl transferase LpxL1 resulting in penta-acyl lipooligosaccharide (LOS), which has reduced endotoxin activity [7]. This latter modification eliminates the need for detergent extraction, which can remove important lipoprotein antigens such as FHbp from OMV and alter the structure of membrane protein epitopes that elicit protective antibodies [7].
Recent studies have shown that antibodies elicited by 4CMenB that are reactive with diverse Ng strains are directed to the MenB porin protein, PorB, and lipooligosaccharide (LOS) [16]. Since MenB OMV have been shown to have 30%-40% efficacy against Ng in fully vaccinated adults, OMVax considered whether protection against Ng by MenB OMV could be increased by also including an NOMV component produced from Ng strains that express porin proteins that are representative of each of the two sub-families of gonococcal porins, PorB.1a and PorB.1b (Ng NOMV). We show here that a combination MenB NOMV vaccine component (Nm NOMV) with overexpressed mutant FHbp from subfamilies A and B provide broader coverage and higher serum bactericidal activity (SBA) against MenB than existing MenB vaccines and, when combined with Ng NOMV components (Nm-Ng NOMV) elicit antibodies having broad SBA against MenB and Ng strains, inhibit colonization of Ng to human cervical and vaginal cell lines and in a transgenic mouse model of gonorrhea. Further, a combined MenB-Ng vaccine that protects against both pathogens may have greater acceptance than a stand-alone Ng vaccine since MenB vaccines are routinely administered to control meningococcal disease in schools, colleges, universities and the military in adolescents and young adults who have a higher risk of acquiring new Ng infections compared to other age groups [17].
2. Materials and Methods
The MenB vaccine component was comprised of NOMV naturally blebbed from two mutant strains of N. meningitidis. Each strain was derived from an H44/76 parent strain. Each strain has deletions of the wild-type fhbp, lpxL1, and siaD-galE genes by insertion of DNA containing mutant fhbp genes (Subfamily A containing three amino acid substitutions, or Subfamily B containing a single amino acid substitution), along with an engineered promoter, and drug resistance markers (erythromycin, kanamycin and spectinomycin, respectively). The Subfamily A and Subfamily B mutants of Factor H binding protein (FHbp) have reduced human Factor H (FH) binding [14,15]. Deletion of lpxL1 results in penta-acylated lipooligosaccharide (LOS), which has attenuated endotoxin activity, compared with the wildtype hexa-acylated LOS [18]. Interruption of the siaD and galE genes eliminates expression of the group B capsular polysaccharide and LOS derivatives, which may elicit antibodies cross-reactive with human tissues [19,20]. To enhance FHbp expression further, each strain also carries a modified version of a multi-copy plasmid containing a chloramphenicol resistance gene [21] and coding for the respective mutant FHbp driven by the engineered promoter. Both N. gonorrhoeae strains had LpxL1 and reduction modifiable protein knocked out.
NOMV were purified from culture supernatant of bacteria grown in chemically defined media, for N. meningitidis, or GCLB [22], for N. gonorrhoeae with supplements that enable the bacteria to grow to high density. After reaching stationary phase, bacteria were harvested by centrifugation. The culture supernatant was filter sterilized using 0.2 µm filters and concentrated using 100 KDa filters (ThermoFisher Scientific). NOMV were collected by ultracentrifugation and further purified by size exclusion chromatography (ToyoPearl HW-55F; Sigma-Aldrich). Total protein concentration was measured by BCA (Pierce). The combination vaccine contained a 1:1 mixture of NOMV from N. meningitidis and N. gonorrhoeae. The combination vaccine referred to as Nm-Ng NOMV was combined with aluminum hydroxide adjuvant (Alhydrogel® 2%, from Croda).
Five to six week old CD-1 mice were immunized I.P. on days 0, and 21 with Nm NOMV at 5 µg/dose, Ng NOMV at 5 µg/Dose, and Nm-Ng NOMV at 10 µg/dose, all adjuvanted with 50 µg/dose of Al(OH)3. Adjuvant alone was given at 50 µg/dose as negative control. Blood was collected by terminal cardiac puncture on day 35. Human CEACAM-1 and FH transgenic FVB-BALB/c hybrid mice were immunized I.P. on days 0, 21 and 42 with Ng NOMV at 5 µ/dose, and Nm-Ng NOMV at 10 µg/dose, both adjuvanted with 50 µg/Dose of Al(OH)3. Bexsero® was administered at 1/5 of the human dose. Blood was collected by terminal cardiac puncture on day 56.
Binding of the post immunization mouse sera to wild-type rFHbps and purified NOMV was measured by enzyme-linked immunosorbent assay (ELISA). The wells of a microtiter plate (Immulon 2B; ThermoFisher Scientific) were coated with 2 µg/ml of rFHbp or NOMV in PBS and incubated overnight at 4°C. The plates were blocked with PBS containing 0.1% Tween-20 (Sigma) (PBST) and 1% (weight/volume) bovine serum albumin (BSA) (Sigma). Concentration-dependent binding of the anti-FHbp and anti-NOMV Abs was measured at 1:100 to 1:200,000 dilution in PBST-BSA. After incubation (2 h at room temperature), the plates were washed and rabbit anti-mouse immunoglobulin G (IgG)-alkaline phosphatase (Zymed; 1:5,000 diluted in PBST/BSA) was added. After 1 h at room temperature, alkaline phosphatase substrate (Sigma) was added, and the absorbance at 405 nm was measured after 30 min. ELISA titers were defined as the serum dilution giving an OD405nm of 1.
For serum bactericidal activity (SBA) assay, bacteria were grown to early log phase in liquid Frantz medium supplemented with 4 mM d,l-lactate (Sigma-Aldrich) and 2 mM cytidine 5′-monophospho-N-acetylneuraminic acid (CMP-NANA; Carbosynth) to enhance the sialylation of lipooligosaccharide [23]. Test sera were heated for 30 min at 56°C to inactivate endogenous complement. The 40-μl reaction mixture used to measure bactericidal activity contained 2-fold serial dilutions of the test serum sample, 300 to 400 CFU of bacteria, and 30% IgG-IgM depleted human complement (Pel-Freez Biologicals). The SBA titer was the interpolated dilution resulting in 50% survival of the bacteria after 60 minutes incubation at 37 °C and 5% CO2 compared to the number of CFU per milliliter in negative-control sera and complement.
For Ng SBA assay, bacterial cultures were prepared the day prior to testing by streaking frozen stocks onto Chocolate II Agar (GC II Agar with Hemoglobin and IsoVitaleX™, Fisher Scientific) and re-passaged onto fresh Chocolate II Agar supplemented with CMP-NANA (50 µg/ml) on the day of assay. After 6 h at 37 °C in an atmosphere containing 5% CO2, bacteria were then suspended in HBSS containing 1 mM MgCl2 and 0.15 mM CaCl2 and 2% BSA. Test sera were heated for 30 min at 56°C to inactivate endogenous complement. The 75-μl reaction mixture used to measure bactericidal activity contained 2-fold serial dilutions of the test serum sample, 800 to 1000 CFU of bacteria, and 20% (volume/volume) IgG-IgM depleted human complement (Pel-Freez Biologicals). The SBA titer was the interpolated dilution resulting in 50% survival of the bacteria after 60 minutes incubation at 37 °C and 5% CO2 compared to the number of CFU per milliliter in negative-control sera and complement.
IgG from postimmunization serum pools from mice immunized with each vaccine and adjuvant control were purified using Protein G HP SpinTrap columns (Cytiva Lifesciences). The concentrations of the purified mouse serum IgG were determined by a capture ELISA in which a goat anti-mouse κ chain specific antibody (Biosource) was adsorbed to the wells of a microtiter plate and used to capture the Abs. The bound Abs were detected with goat anti-mouse κ chain specific antibody conjugated with alkaline phosphatase (Biosource). The concentrations of the mouse IgG Abs were assigned by comparison with binding of a purified mouse IgG standard (BioRad).
For colonization assays, bacterial cultures were prepared the day prior to testing by streaking frozen stocks onto Chocolate II Agar (GC II Agar with Hemoglobin and IsoVitaleX™, Fisher Scientific). Bacteria were then suspended in PBS to an OD620nm of 0.3 and 10 µM CellTrace™ CFSE Cell Proliferation dye (Invitrogen) was added to the solution. The labeling reaction was stopped after 30 minutes with fetal bovie serum (FBS), and free label removed with two washes. Labeled bacteria were spun down and respuspended in PBS supplemented with CMP-NANA (50 µg/ml) to an OD620nm of 0.3. Bacteria were further diluted in cell culture media to reach ∼1 × 105 CFU/mL. For the adherence assays, monolayers of human cervical (ME-180, ATCC #HTP-33) and vaginal epithelial (VK2/E6E7, ATCC #CRL-2616) cells were incubated at 37 °C, 5 % CO2 for four hours with ∼1 × 105 CFU sialylated N. gonorrhoeae strains that had been pre-incubated with 25 µg/ml purified mouse IgG for 15 min in cell culture media. Following infection, cells were washed three times with PBS to remove non-adherent bacteria, the cells were detached using Gibco™ StemPro™ Accutase™, and transferred to microcentrifuge tubes. Cells were washed twice with PBS and fixed with 4% Paraformaldehyde. Labeled adherent bacteria were quantitated using ImageStreamX (Cytek Biosciences). Data are presented as percentage of inhibition of adherent bacteria relative to adjuvant only antibody control.
Transgenic mice were screened for the human CEACAM1 gene and human FH protein expression as described elsewhere [24]. MenB strain 8047 and Ng strain WHO F were grown as described as for the SBA assay protocol above. After reaching an OD620nm of 0.6 for Nm and after being resuspended to an OD of 0.3 for Ng, bacteria were diluted to get to the targeted inoculum of 103 CFU/animal for 8047, and 107 CFU/animal for WHO F. Immediately after the challenge, a sample of the bacteria preparation was plated on an agar plate to confirm the inoculum. Mice were challenged intranasally and euthanized after 48 hours for Nm challenge and 72 hours for Ng challenge. Tracheal wash and tissue samples were collected for each animal and plated on GC agar plates (REF) supplemented with IsoVitalex and VCNT inhibitor. Colonies were visible after 16-24 hours incubation at 37˚C with 5% CO2. The tracheal wash was performed using an insulin syringe filled with 500µL of PBS, 1 mM Mg buffer inserted into the trachea, and the wash was collected from the nares into a 1.5mL microtube. To obtain bacteria adhering to the nasal pharynx mucosal surfaces, the lower jaw of the mouse was removed. The soft palette was collected into a tube filled with 500µL of 0.05% trypsin-EDTA or Accutase™ (ThermoFisher Scientific). The hard palette and nasal passages were swabbed with an ultrafine aluminum shaft applicator with a calcium alginate fiber tip (Puritan), which was resuspended into the tube with soft palette tissue sample. The soft palette and fiber tip were incubated at 37°C for 10 minutes before plating the entire solution on a chocolate agar plate. The tracheal wash was plated on a separate plate. CFU were counted the following day after overnight incubation at 37°C in an atmosphere of 5% CO2. Statistical comparison between the results for mice given adjuvant only with mice immunized with the Nm-Ng NOMV vaccine was performed using a Welch’s two tailed t test with Prism (GraphPad).
3. Results
The IgG anti-FHbp titers of all CD1 WT mice before vaccination were below the lowest serum dilution tested (<1:100). At two weeks after dose 2, the IgG titers remained below 1:100 in the adjuvant negative-control group and Ng NOMV vaccine group. IgG titers increased in both vaccine groups (Figure 1a) where the Nm NOMV component was present (alone and in combination with the Ng NOMV vaccine). The anti-FHbp IgG titers elicited by the Nm-Ng NOMV combination were not significantly reduced compared to Nm NOMV alone. To test the breadth of SBA activity, MenB strains were chosen to represent both close (FHbp IDs 13 and 24) and distant (ID15 and ID28) homology to the FHbp antigens present in the vaccines (Figure 1b) for FHbp subfamilies B and A, respectively. The SBA titers using IgG/IgM-depleted human complement are shown in Figure 1c. There was no significant difference between titers elicited by the combination Nm-Ng NOMV vaccine or Nm NOMV alone. The Ng NOMV component and adjuvant alone antisera had no SBA against the four MenB test strains.
The IgG titers against NOMV from two Ng strains, WHO M (PorB.1a) and WHO N (PorB.1b) that are heterologous to the vaccine NOMV are shown in Figure 2a. The titers of mice given Ng NOMV alone or in combination are approximately 10-fold higher than for mice given the Nm NOMV vaccine only and there was no significant difference between the titers of mice immunized with Ng NOMV or the Nm-Ng NOMV combination vaccine.
The Nm NOMV antisera had little or no SBA against the Ng test strains. Ng NOMV and Nm-Ng NOMV antisera had high SBA titers against strains expressing PorB.1a and PorB.1b. The difference in Ng NOMV SBA titers were not significantly different from those of the Nm-Ng NOMV vaccine except for Ng strain FA1090, which is homologous to the strains used to prepare the Ng NOMV component. PorB.1b and PorB.1a in the strains used to prepare the NOMV vaccine were heterologous to PorB in WHO M, WHO G and WHO N.
Currently, there is no in vitro correlate of protection against Ng infection. However, for N. gonorrhoeae to cause an infection, it must adhere to mucosal epithelium. Therefore, we investigated whether purified IgG antibodies from mice immunized with the NOMV vaccines was able to inhibit adherence to VK2/E6E7 vaginal and ME-180 cervical epithelial cells using sialylated, fluorescently labeled bacteria from Ng strains expressing PorB.1a (WHO F, WHO G, WHO M) and PorB.1b (FA1090, MS11, and WHO M). The data were obtained and analyzed using an ImageStreamX Mark II flow cytometer that combines flow cytometry with fluorescence microscopy to identify and count individual cells with adherent bacteria. Purified IgG from mice immunized with Nm NOMV had significant inhibition activity against WHO G to VK2/E6E7 vaginal cells compared to the adjuvant only control but not any other Ng strain or any strain to ME-180 cervical cells (Figure 3a,b). In contrast, IgG from mice immunized with Ng NOMV and the combination Nm-Ng NOMV vaccine inhibited adherence of all Ng strains to both cell lines except for WHO M on VK2/E6E7 cells and to FA1090 and WHO M on ME-180 cells (Figure 3a,b). The differences in inhibitory activity between purified IgG from Ng NOMV and Nm-Ng NOMV-immunized mice was not significant.
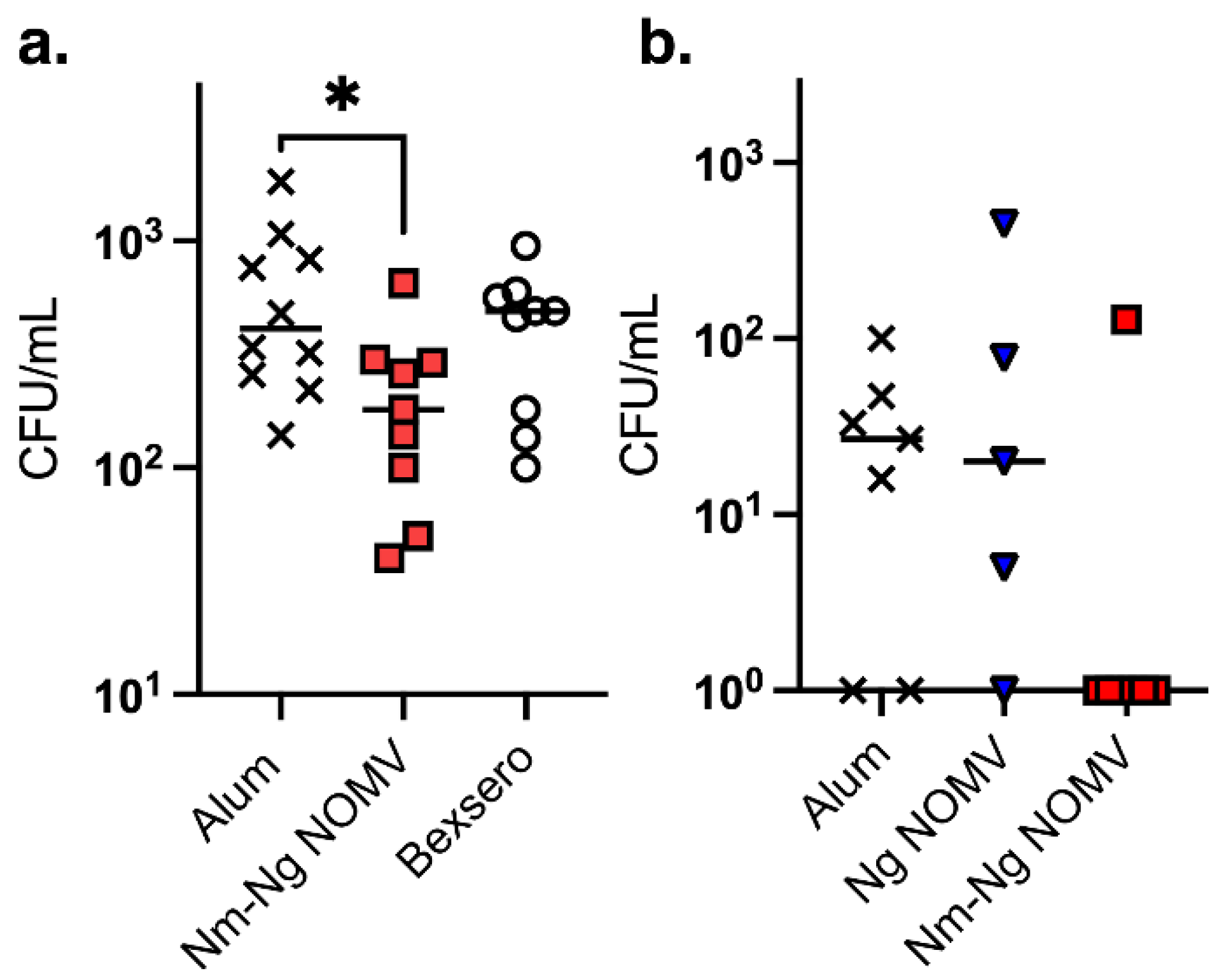
Ng is a human restricted pathogen. Although a wild-type mouse Ng challenge model is commonly used to test whether vaccine elicited antibodies can inhibit vaginal colonization by Ng strains, the wild-type mice used lack human-specific receptors and immune shielding mechanisms and, therefore, do not replicate Ng colonization of human mucosal tissues. To determine whether mice immunized with NOMV vaccines can inhibit colonization in an animal model, we constructed a transgenic (Tg) mouse line that expresses human CEACAM1 for adhesion through Opa proteins and Factor H to provide immune shielding. The human CEACAM1/FH Tg mice were challenged intranasally with MenB strain 8047 or Ng strain WHO F after immunization with two doses of each NOMV vaccine or adjuvant only. The Nm 8047 strain was heterologous to the strains used to prepare the NOMV used in the vaccines. For MenB colonization, an additional control included 4CMenB, which provides protection against MenB disease but does not appear to affect colonization in humans [25]. As shown in Figure 4, mice given NOMV-NmNg vaccine inhibited colonization of MenB strain 8047 that was significant compared to the adjuvant only control but not to 4CMenB. Also, Tg mice immunized with NOMV-NmNg inhibited colonization of WHO F compared to mice immunized with adjuvant or NOMV-Ng (p>0.0001).
4. Discussion
To date, the only vaccines shown to have efficacy in preventing gonorrhea are vaccines containing vesicles isolated from the closely related bacterial species Neisseria meningitidis. Building upon this data, we constructed a novel vaccine, Nm-Ng NOMV, that combines a next generation NOMV MenB vaccine with NOMV produced from Ng strains that express PorB.1a and PorB.1b. We focused on PorB for both Nm and Ng strains since PorB is a major component of OMV from both Nm and Ng. Further support for this approach comes from a recent study of human monoclonal antibodies produced from subjects vaccinated with 4CMenB that were reactive with Nm and Ng PorB epitopes as well as LOS [16]. The anti-PorB antibodies had SBA against Ng strains and provided cross-protection in a wild-type mouse vaginal challenge model of gonococcal infection. The PorB amino acid sequence from the production strain for Nm NOMV is identical to PorB in the OMV component of 4CMenB. The two Ng strains used for production of NOMV were chosen based on PorB.1a and PorB.1b sequences where the surface exposed variable loops were among the most common for Ng isolates of each PorB subfamily circulating world-wide [26].
OMVax’s Nm-Ng NOMV vaccine elicited antibodies that had SBA against diverse Ng strains. In our SBA assay, the bacteria were grown in the presence of CMP-NANA, which is scavenged from the human host and used by the bacteria to sialylate LOS. Sialylation of LOS results in bacteria that are considerably more resistant to SBA than strains grown without CMP-NANA [27]. SBA assays were performed using IgG/IgM-depleted human complement, which more closely replicates SBA activity in humans compared to SBA done with non-human complement [28]. SBA activity confirms that Nm-Ng NOMV vaccine elicits antibodies that recognize antigens on the bacterial surface of the strains tested that are present in amounts sufficient for robust SBA activity. SBA has not been established as a surrogate of protection against disease but is likely to be important in preventing gonorrhea.
Nm and Ng are obligate human pathogens that use mechanisms for attachment (CEACAMs, CD46), invasion [29], and immune shielding (FH [30], TspB [31], OpaCEA [32]) that specifically interact with human receptors. Antibodies elicited by vaccines (e.g., IgA and IgG) are present in secretions enveloping epithelial cells that are in direct contact with Nm and Ng during the earliest stages of infection and can prevent colonization and invasion [33]. Antibodies that interfere with the mechanisms of colonization protect the host during the initial stages of infection from more advanced stages of disease and potentially limit transmission between vaccinated and unvaccinated individuals (i.e., community immunity). The most cost effective and widely used vaccines provide both individual and community protection.
Purified IgG from mice immunized with Ng NOMV or Nm-Ng NOMV were able to inhibit adherence of 5 out of 6 Ng strains to human vaginal and/or cervical cell lines. Interestingly, antibodies elicited by both vaccines inhibited adherence of Ng strain FA1090 to vaginal cells but not cervical cells and did not inhibit adherence of Ng strain WHO M to either cell line even though both vaccines produced high titers of bactericidal antibodies against WHO M (Figure 3a,b). Inhibiting adherence is also likely important for protection, but adherence mediated by bacterial Opa protein binding to host CEACAM receptors is complicated by expression of different CEACAMs (e.g., CEACAM1 in cervical tissues and CEACAM5 in vaginal tissues) and expression of different Opa proteins by the bacteria, which also vary depending on environmental factors [34]. Although we did not quantitate antibodies to Opa proteins elicited by Ng NOMV or Nm-Ng NOMV, it is possible that presence of multiple Opa proteins in the mixture of NOMVs may be important for eliciting antibodies that can inhibit adherence by diverse strains in the vaginal and cervical cell culture models of Ng adherence.
To evaluate ability to inhibit Ng colonization in a live animal, we used a transgenic mouse model of Ng colonization that replicates some aspects of pathogenesis in humans in expressing human CEACAM1, a receptor specific for Opa proteins expressed by Nm and Ng that mediates adhesion, and human FH to provide immune protection. The mice were challenged intranasally, which enabled testing both male and female mice. Although the results were not statistically significant in this under powered preliminary experiment, 6 of 7 mice vaccinated with Nm-Ng NOMV and challenged intranasally with Ng strain WHO F had no colonizing bacteria compared to only 2 of 7 mice given adjuvant alone or 1 of 6 given Ng NOMV alone (Figure 4). The trend suggests the combination vaccine that included the MenB NOMV component provided superior protection compared to the Ng NOMV component alone and are consistent with the results of the epidemiological studies showing vaccines with a MenB OMV provide some protection against gonorrhea in humans [1,3,4,5,6].
Although there are two MenB vaccines approved for use in humans in multiple countries, neither 4CMenB or MenB-FHbp is broadly protective or able to protect against disease caused by an increasing number of strains currently causing outbreaks in several European countries and Canada [11,12,13]. The situation calls for development of a next generation MenB vaccine that can provide broader coverage. The Nm NOMV component of Nm-Ng NOMV vaccine, contains mutant FHbps from subfamilies A and B with reduced FH binding overexpressed in NOMV such that epitope structure and native environment is preserved. As a result, the FHbp antigens elicit high titers of antibodies that bind to FHbp and mediate complement-dependent bactericidal activity but can also block FH binding, which further enhances SBA [35]. SBA is a surrogate of protection in humans against meningococcal disease [36]. As shown in Figure 1c, the ability of Nm NOMV to elicit antibodies with broad SBA activity was not diminished by combining with Ng NOMV in the Nm-Ng NOMV vaccine. Similarly, Nm-Ng NOMV elicited antibodies with high SBA titers against diverse Ng strains compared to Ng NOMV alone (Figure 2) and inhibited colonization of a heterologous MenB strain in a mouse model of meningococcal disease (Figure 4a).
Finally, a combination MenB-Ng vaccine may be advantageous with respect to acceptance and potential to control the spread of gonorrhea. Adolescents and young adults are commonly vaccinated for protection against MenB and this age group is also of greatest risk of acquiring new gonorrhea infections [17]. The Nm-Ng NOMV vaccine has the potential to address both problems since a vaccine with potential to prevent life-threatening meningococcal disease and gonorrhea in a population of sexually active subjects at highest risk of Ng infections may be more acceptable to the public than a vaccine specifically for prevention of a sexually transmitted infection.
Author Contributions
The study was conceived and designed by S.G., and G.R.M.; data were acquired by S.G., S.W.T., A.B., A.R.R., and G.R.M.; data were analyzed and interpreted by S.G., S.W.T., A.B., A.R.R., and G.R.M.; drafting of the article was performed by and critical intellectual content was provided by S.G. and G.R.M.; final approval of the version to be submitted was provided by S.G., S.W.T., A.B., A.R.R., and G.R.M.
Funding
This research was funded in part by NIH NIAID grant numbers R01AI046464 and R44AI124759 to G.R.M.
Institutional Review Board Statement
All experiments involving the use of animals in this study were conducted using protocols approved by the Institutional Animal Use and Care Committee of the University of California, San Francisco in facilities certified by the American Association for Laboratory Animal Science (AALAS).
Data Availability Statement
All data described is included in this manuscript.
Conflicts of Interest
S.G., S.W.T, A.B., A.R.R. and G.R.M. are or were employees of OMVax, Inc. S.G. and G.R.M. hold shares in OMVax, Inc. and declare no other financial and nonfinancial relationships and activities related to this study.
Abbreviations
The following abbreviations are used in this manuscript:
| dOMV | detergent-treated outer membrane vesicles |
| ELISA | enzyme-linked immunoabsorbant assay |
| FH | Factor H |
| FHbp | Factor H binding protein |
| GalE KO | inactivated UDP-glucose-4-epimerase gene, resulting in synthesis of LOS that cannot by sialylated |
| GMT | geometric mean titer |
| IP | intraperitoneal |
| KO | knockout |
| LpxL1 KO | Genetically inactivated LpxL1 (attenuates endotoxin) |
| MenB | Neisseria meningitidis capsular group B or serogroup B |
| 4CMenB | Bexsero®, consisting of 3 recombinant proteins and 25 µg of dOMV |
| MenB-FHbp | Trumenba® , consisting of recombinant FHbp from subfamilies A and B |
| NOMV | native, non-detergent-treated outer membrane vesicles |
| Nm | Neisseria meningitidis |
| Ng | Neisseria gonorrheae |
| OMV | outer membrane vesicles |
| LOS | lipooligosaccharide (endotoxin) (because of the absence of the O-antigen, N. meningitidis LPS is referred to as LOS |
| SBA | serum bactericidal activity |
| WT | wild-type |
| siaD KO | inactivated sialyl transferase gene (deletes group B capsular synthesis) |
| STI | Sexually transmitted infection |
References
- Petousis-Harris, H.; Paynter, J.; Morgan, J.; Saxton, P.; McArdle, B.; Goodyear-Smith, F.; Black, S. Effectiveness of a group B outer membrane vesicle meningococcal vaccine against gonorrhoea in New Zealand: a retrospective case-control study. Lancet 2017, 390, 1603-1610. [CrossRef]
- Oster, P.; Lennon, D.; O’Hallahan, J.; Mulholland, K.; Reid, S.; Martin, D. MeNZB: a safe and highly immunogenic tailor-made vaccine against the New Zealand Neisseria meningitidis serogroup B disease epidemic strain. Vaccine 2005, 23, 2191-2196.
- Azze, R.F.O. A meningococcal B vaccine induces cross-protection against gonorrhea. Clin Exp Vaccine Res 2019, 8, 110-115. [CrossRef]
- Reyes Diaz, L.M.; Lastre Gonzalez, M.; Cuello, M.; Sierra-Gonzalez, V.G.; Ramos Pupo, R.; Lantero, M.I.; Harandi, A.M.; Black, S.; Perez, O. VA-MENGOC-BC Vaccination Induces Serum and Mucosal Anti Neisseria gonorrhoeae Immune Responses and Reduces the Incidence of Gonorrhea. Pediatr Infect Dis J 2021, 40, 375-381. [CrossRef]
- Whelan, J.; Klovstad, H.; Haugen, I.L.; Holle, M.R.; Storsaeter, J. Ecologic Study of Meningococcal B Vaccine and Neisseria gonorrhoeae Infection, Norway. Emerg Infect Dis 2016, 22, 1137-1139. [CrossRef]
- Longtin, J.; Dion, R.; Simard, M.; Betala Belinga, J.F.; Longtin, Y.; Lefebvre, B.; Labbé, A.C.; Deceuninck, G.; De Wals, P. Possible Impact of Wide-scale Vaccination Against Serogroup B Neisseria Meningitidis on Gonorrhea Incidence Rates in One Region of Quebec,. Open Forum Infect Dis. 2017, 4, S734-S735.
- Beernink, P.T.; Ispasanie, E.; Lewis, L.A.; Ram, S.; Moe, G.R.; Granoff, D.M. A Meningococcal Native Outer Membrane Vesicle Vaccine With Attenuated Endotoxin and Overexpressed Factor H Binding Protein Elicits Gonococcal Bactericidal Antibodies. J Infect Dis 2019, 219, 1130-1137. [CrossRef]
- Liu, Y.; Hammer, L.A.; Daamen, J.; Stork, M.; Egilmez, N.K.; Russell, M.W. Microencapsulated IL-12 Drives Genital Tract Immune Responses to Intranasal Gonococcal Outer Membrane Vesicle Vaccine and Induces Resistance to Vaginal Infection with Diverse Strains of Neisseria gonorrhoeae. mSphere 2023, 8, e0038822. [CrossRef]
- Matthias, K.A.; Connolly, K.L.; Begum, A.A.; Jerse, A.E.; Macintyre, A.N.; Sempowski, G.D.; Bash, M.C. Meningococcal Detoxified Outer Membrane Vesicle Vaccines Enhance Gonococcal Clearance in a Murine Infection Model. J Infect Dis 2022, 225, 650-660. [CrossRef]
- Ruiz Garcia, Y.; Marrazzo, J.; Martinon-Torres, F.; Workowski, K.; Giordano, G.; Pizza, M.; Sohn, W.Y. Urgent Need to Understand and Prevent Gonococcal Infection: From the Laboratory to Real-World Context. J Infect Dis 2024, 230, e758-e767. [CrossRef]
- Lujan, E.; Partridge, E.; Giuntini, S.; Ram, S.; Granoff, D.M. Breadth and Duration of Meningococcal Serum Bactericidal Activity in Health Care Workers and Microbiologists Immunized with the MenB-FHbp Vaccine. Clin Vaccine Immunol 2017, 24. [CrossRef]
- Lujan, E.; Winter, K.; Rovaris, J.; Liu, Q.; Granoff, D.M. Serum Bactericidal Antibody Responses of Students Immunized With a Meningococcal Serogroup B Vaccine in Response to an Outbreak on a University Campus. Clin Infect Dis 2017, 65, 1112-1119. [CrossRef]
- Roca-Grande, J.; Mir-Cros, A.; Munoz-Almagro, C.; Lung, M.; Belles-Belles, A.; Camara, J.; Cercenado, E.; Galan-Ladero, M.A.; Martin-Nalda, A.; Moreno-Mingorance, A.; et al. Genomic characterization of invasive Neisseria meningitidis in Spain (2011/12-2022/23): expansion of clonal complex 213 and the potential threat to 4CMenB vaccine strain coverage. Emerg Microbes Infect 2025, 14, 2482696. [CrossRef]
- Konar, M.; Rossi, R.; Walter, H.; Pajon, R.; Beernink, P.T. A Mutant Library Approach to Identify Improved Meningococcal Factor H Binding Protein Vaccine Antigens. PLoS One 2015, 10, e0128185. [CrossRef]
- Rossi, R.; Konar, M.; Beernink, P.T. Meningococcal Factor H Binding Protein Vaccine Antigens with Increased Thermal Stability and Decreased Binding of Human Factor H. Infect Immun 2016, 84, 1735-1742. [CrossRef]
- Vezzani, G.; Viviani, V.; Audagnotto, M.; Rossi, A.; Cinelli, P.; Pacchiani, N.; Limongi, C.; Santini, L.; Giusti, F.; Tomei, S.; et al. Isolation of human monoclonal antibodies from 4CMenB vaccinees reveals PorB and LOS as the main OMV components inducing cross-strain protection. Front Immunol 2025, 16, 1565862. [CrossRef]
- National Center for HIV, V.H., STD, and Tuberculosis Prevention. National Overview of STIs in 2023. Available online: https://www.cdc.gov/sti-statistics/annual/summary.html (accessed on August 6).
- Steeghs, L.; Tommassen, J.; Leusen, J.H.; van de Winkel, J.G.; van der Ley, P. Teasing apart structural determinants of ‘toxicity’ and ‘adjuvanticity’: implications for meningococcal vaccine development. J Endotoxin Res 2004, 10, 113-119. [CrossRef]
- Finne, J.; Leinonen, M.; Makela, P.H. Antigenic similarities between brain components and bacteria causing meningitis. Implications for vaccine development and pathogenesis. Lancet 1983, 2, 355-357.
- Harvey, H.A.; Swords, W.E.; Apicella, M.A. The mimicry of human glycolipids and glycosphingolipids by the lipooligosaccharides of pathogenic neisseria and haemophilus. J Autoimmun 2001, 16, 257-262.
- Pagotto, F.J.; Salimnia, H.; Totten, P.A.; Dillon, J.R. Stable shuttle vectors for Neisseria gonorrhoeae, Haemophilus spp. and other bacteria based on a single origin of replication. Gene 2000, 244, 13-19. [CrossRef]
- Ragland, S.A.; Criss, A.K. Protocols to Interrogate the Interactions Between Neisseria gonorrhoeae and Primary Human Neutrophils. Methods Mol Biol 2019, 1997, 319-345. [CrossRef]
- Mandrell, R.E.; Kim, J.J.; John, C.M.; Gibson, B.W.; Sugai, J.V.; Apicella, M.A.; Griffiss, J.M.; Yamasaki, R. Endogenous sialylation of the lipooligosaccharides of Neisseria meningitidis. J Bacteriol 1991, 173, 2823-2832.
- Beernink, P.T.; Shaughnessy, J.; Pajon, R.; Braga, E.M.; Ram, S.; Granoff, D.M. The effect of human factor H on immunogenicity of meningococcal native outer membrane vesicle vaccines with over-expressed factor H binding protein. PLoS Pathog 2012, 8, e1002688. [CrossRef]
- Marshall, H.S.; McMillan, M.; Koehler, A.P.; Lawrence, A.; Sullivan, T.R.; MacLennan, J.M.; Maiden, M.C.J.; Ladhani, S.N.; Ramsay, M.E.; Trotter, C.; et al. Meningococcal B Vaccine and Meningococcal Carriage in Adolescents in Australia. N Engl J Med 2020, 382, 318-327. [CrossRef]
- McKnew, D.L.; Lynn, F.; Zenilman, J.M.; Bash, M.C. Porin variation among clinical isolates of Neisseria gonorrhoeae over a 10-year period, as determined by Por variable region typing. J Infect Dis 2003, 187, 1213-1222. [CrossRef]
- Wetzler, L.M.; Barry, K.; Blake, M.S.; Gotschlich, E.C. Gonococcal lipooligosaccharide sialylation prevents complement-dependent killing by immune sera. Infect Immun 1992, 60, 39-43. [CrossRef]
- Matthias, K.A.; Reveille, A.; Dhara, K.; Lyle, C.S.; Natuk, R.J.; Bonk, B.; Bash, M.C. Development and validation of a standardized human complement serum bactericidal activity assay to measure functional antibody responses to Neisseria gonorrhoeae. Vaccine 2025, 43, 126508. [CrossRef]
- Criss, A.K.; Seifert, H.S. A bacterial siren song: intimate interactions between Neisseria and neutrophils. Nature reviews. Microbiology 2012, 10, 178-190. [CrossRef]
- Madico, G.; Welsch, J.A.; Lewis, L.A.; McNaughton, A.; Perlman, D.H.; Costello, C.E.; Ngampasutadol, J.; Vogel, U.; Granoff, D.M.; Ram, S. The meningococcal vaccine candidate GNA1870 binds the complement regulatory protein factor H and enhances serum resistance. J Immunol 2006, 177, 501-510.
- Muller, M.G.; Moe, N.E.; Richards, P.Q.; Moe, G.R. Resistance of Neisseria meningitidis to human serum depends on T and B cell stimulating protein B. Infect Immun 2015, 83, 1257-1264. [CrossRef]
- Boulton, I.C.; Gray-Owen, S.D. Neisserial binding to CEACAM1 arrests the activation and proliferation of CD4+ T lymphocytes. Nat Immunol 2002, 3, 229-236. [CrossRef]
- Khalil, M.; Al-Mazrou, Y.; Findlow, H.; Chadha, H.; Bosch Castells, V.; Oster, P.; Borrow, R. Meningococcal serogroup C serum and salivary antibody responses to meningococcal quadrivalent conjugate vaccine in Saudi Arabian adolescents previously vaccinated with bivalent and quadrivalent meningococcal polysaccharide vaccine. Vaccine 2014, 32, 5715-5721. [CrossRef]
- Islam, E.A.; Anipindi, V.C.; Francis, I.; Shaik-Dasthagirisaheb, Y.; Xu, S.; Leung, N.; Sintsova, A.; Amin, M.; Kaushic, C.; Wetzler, L.M.; et al. Specific Binding to Differentially Expressed Human Carcinoembryonic Antigen-Related Cell Adhesion Molecules Determines the Outcome of Neisseria gonorrhoeae Infections along the Female Reproductive Tract. Infect Immun 2018, 86. [CrossRef]
- Beernink, P.T.; Vianzon, V.; Lewis, L.A.; Moe, G.R.; Granoff, D.M. A Meningococcal Outer Membrane Vesicle Vaccine with Overexpressed Mutant FHbp Elicits Higher Protective Antibody Responses in Infant Rhesus Macaques than a Licensed Serogroup B Vaccine. MBio 2019, 10. [CrossRef]
- Frasch, C.E.; Borrow, R.; Donnelly, J. Bactericidal antibody is the immunologic surrogate of protection against meningococcal disease. Vaccine 2009, 27 Suppl 2, B112-116. [CrossRef]
Figure 1.
(a). Serum IgG anti-FHbp titers by ELISA (recombinant FHbp on the plate). Nm-Ng NOMV vaccine showed comparable IgG titers to the Nm NOMV only vaccine. (b). FHbp ID splits-Tree dendrogram. FHbp sequence IDs for a set of currently circulating invasive MenB strains from the CDC are compared to those in Nm-Ng NOMV (shown in green), 4CMenB and MenB-FHbp vaccines (shown in red). FHbp IDs for strains used for SBA are highlighted in blue. (c). SBA titers against MenB strains. SBA assays performed using IgG-IgM depleted human serum as source of complement and bacteria grown in the presence of CMP-NANA. Nm-Ng NOMV vaccine showed similar SBA compared to Nm NOMV against MenB strains irrespective of matching or mismatching FHbp sequences. Serum from 4CMenB immunized mice had no SBA, except for Quebec and UK240476 strains (GMT 1:210 and 1:25 respectively).
Figure 1.
(a). Serum IgG anti-FHbp titers by ELISA (recombinant FHbp on the plate). Nm-Ng NOMV vaccine showed comparable IgG titers to the Nm NOMV only vaccine. (b). FHbp ID splits-Tree dendrogram. FHbp sequence IDs for a set of currently circulating invasive MenB strains from the CDC are compared to those in Nm-Ng NOMV (shown in green), 4CMenB and MenB-FHbp vaccines (shown in red). FHbp IDs for strains used for SBA are highlighted in blue. (c). SBA titers against MenB strains. SBA assays performed using IgG-IgM depleted human serum as source of complement and bacteria grown in the presence of CMP-NANA. Nm-Ng NOMV vaccine showed similar SBA compared to Nm NOMV against MenB strains irrespective of matching or mismatching FHbp sequences. Serum from 4CMenB immunized mice had no SBA, except for Quebec and UK240476 strains (GMT 1:210 and 1:25 respectively).
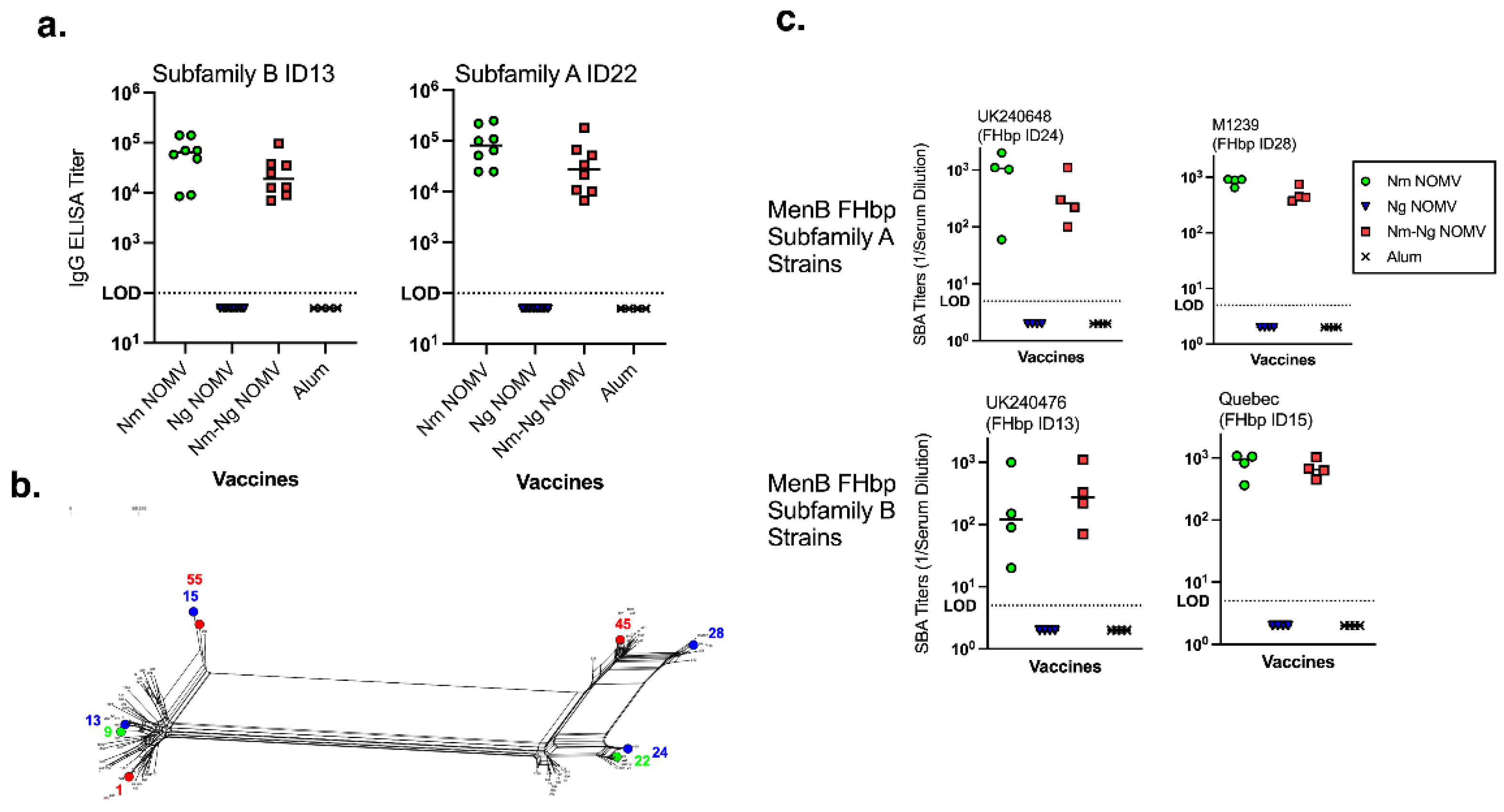
Figure 2.
(a). Serum IgG anti-Ng titers by ELISA (NOMV from two heterologous Ng strains on the plate). Nm-Ng NOMV vaccine showed comparable IgG titers to the Ng NOMV only vaccine. (b) SBA Titers against Ng strains. SBA assays performed using IgG-IgM depleted human serum as source of complement and bacteria grown in the presence of CMP-NANA. Nm-Ng NOMV vaccine elicited high SBA titers, similar to the Ng NOMV vaccine, against a broad panel of Ng strains. Serum from 4CMenB-immunized mice had no SBA against FA1090 and it was not tested against other Ng strains.
Figure 2.
(a). Serum IgG anti-Ng titers by ELISA (NOMV from two heterologous Ng strains on the plate). Nm-Ng NOMV vaccine showed comparable IgG titers to the Ng NOMV only vaccine. (b) SBA Titers against Ng strains. SBA assays performed using IgG-IgM depleted human serum as source of complement and bacteria grown in the presence of CMP-NANA. Nm-Ng NOMV vaccine elicited high SBA titers, similar to the Ng NOMV vaccine, against a broad panel of Ng strains. Serum from 4CMenB-immunized mice had no SBA against FA1090 and it was not tested against other Ng strains.
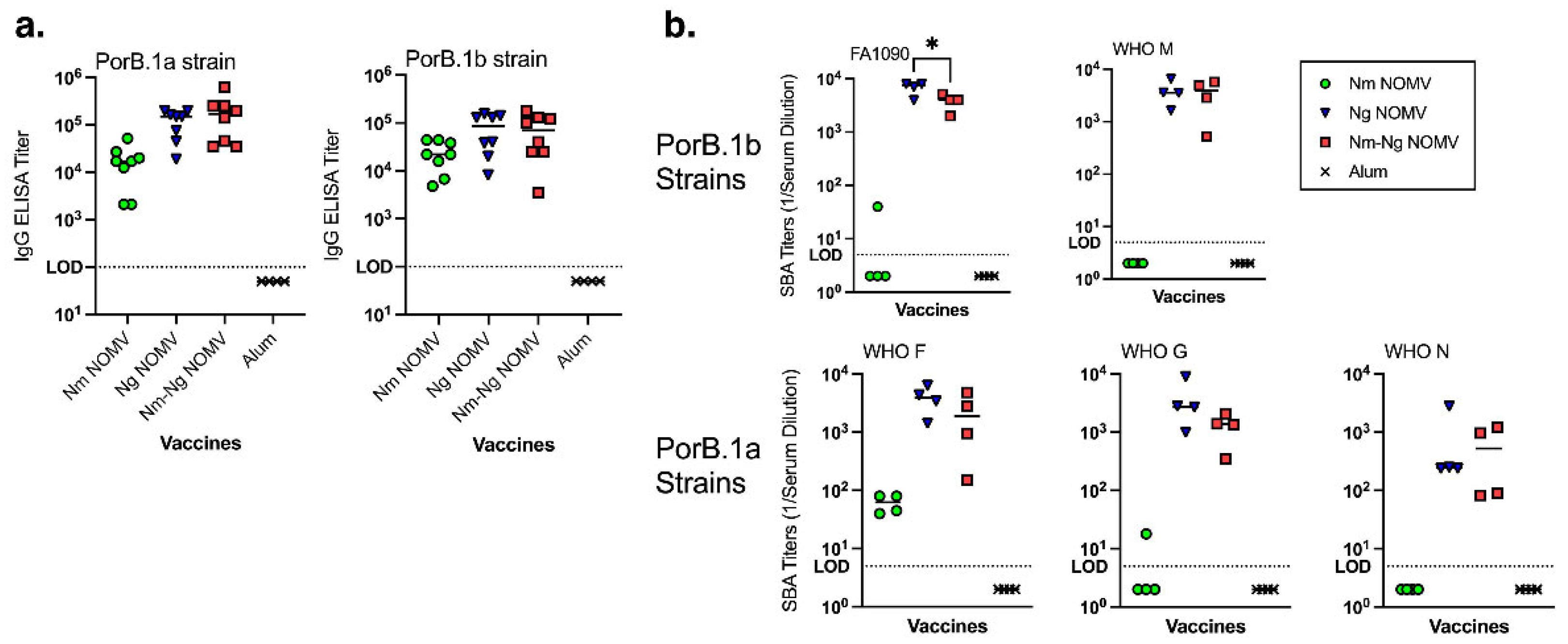
Figure 3.
Inhibition of adhesion by Ng strains to cervical and vaginal human cells by IgG purified from serum pools. Sialylated bacteria and 25 µg/mL of purified IgG were added to VK2 or ME-180 cells to test antibody ability to prevent Ng adhesion to human cells. Adhering labeled bacteria were quantified using ImageStreamX. Test strains included both PorB.1a (WHO F, WHO G, and WHO N) and PorB.1b strains (FA1090, MS11, WHO M). (a) Inhibition of adherence to VK2 vaginal cells and (b) ME-180 cervical cells.
Figure 3.
Inhibition of adhesion by Ng strains to cervical and vaginal human cells by IgG purified from serum pools. Sialylated bacteria and 25 µg/mL of purified IgG were added to VK2 or ME-180 cells to test antibody ability to prevent Ng adhesion to human cells. Adhering labeled bacteria were quantified using ImageStreamX. Test strains included both PorB.1a (WHO F, WHO G, and WHO N) and PorB.1b strains (FA1090, MS11, WHO M). (a) Inhibition of adherence to VK2 vaginal cells and (b) ME-180 cervical cells.
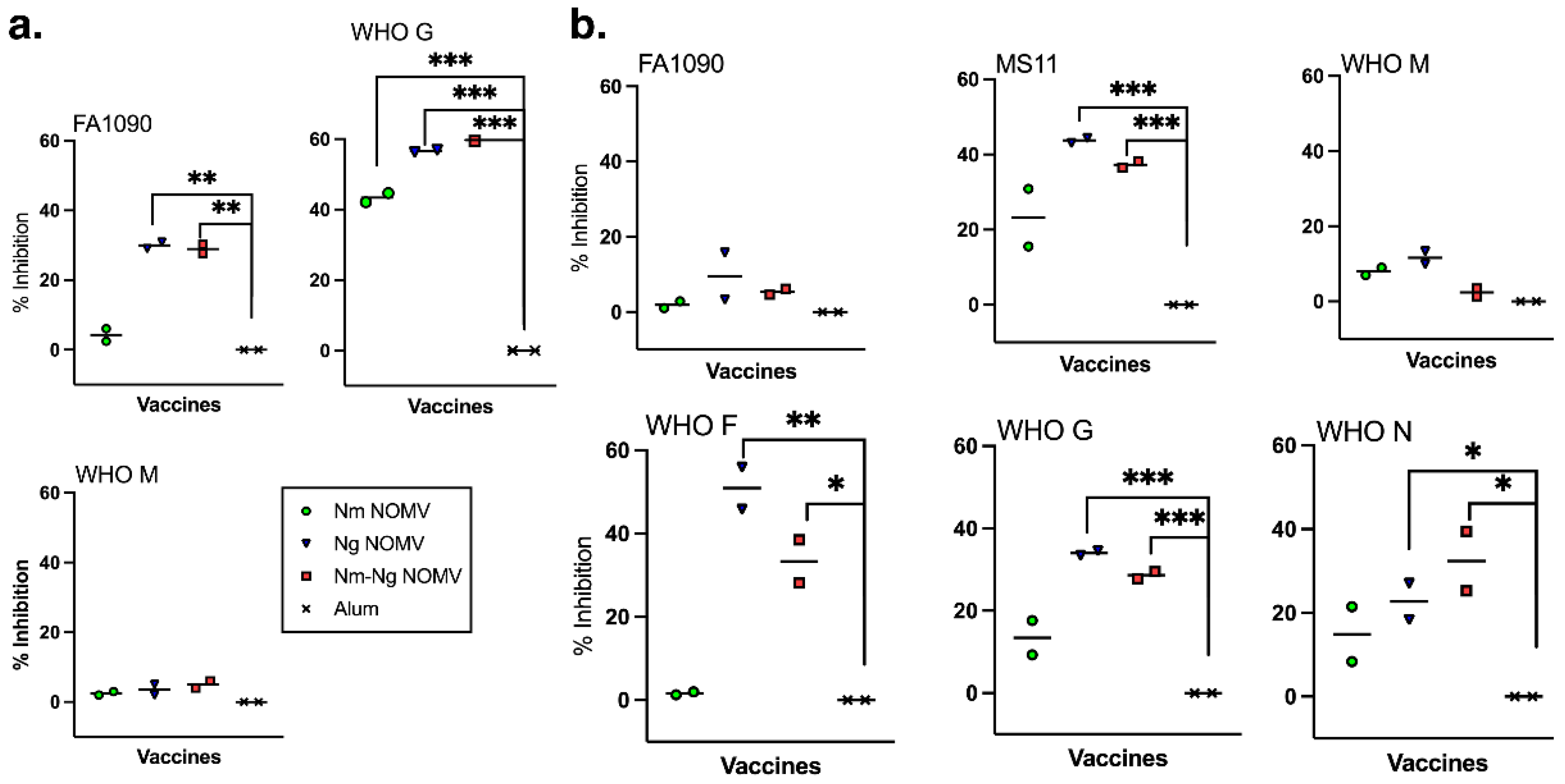
Figure 4.
In vivo inhibition of colonization. (a) MenB colonization. Tg mice immunized with either Nm-Ng NOMV, Bexsero or alum were challenged with 1x103 CFU of heterologous MenB strain 8047. Bacteria were recovered after 48 hours from the challenge. Nm-Ng NOMV decreased CFU in the nasopharynx compared to the alum control (P=0.04). (b) Ng colonization. Tg mice immunized with either Ng NOMV, Nm-Ng NOMV, or alum were challenged intranasally with 1x107 CFU of Ng strain WHO F. Bacteria were recovered after 72 hours from the challenge. The difference between the alum control and the Tg mice receiving the NOMV vaccines was not statistically significant. Variability in each group was high. However, a trend was observed as 6 of 7 mice vaccinated with Nm-Ng NOMV had no CFU, suggesting that the combination vaccine was more effective. A larger number of animals per group will be required to detect differences.
Figure 4.
In vivo inhibition of colonization. (a) MenB colonization. Tg mice immunized with either Nm-Ng NOMV, Bexsero or alum were challenged with 1x103 CFU of heterologous MenB strain 8047. Bacteria were recovered after 48 hours from the challenge. Nm-Ng NOMV decreased CFU in the nasopharynx compared to the alum control (P=0.04). (b) Ng colonization. Tg mice immunized with either Ng NOMV, Nm-Ng NOMV, or alum were challenged intranasally with 1x107 CFU of Ng strain WHO F. Bacteria were recovered after 72 hours from the challenge. The difference between the alum control and the Tg mice receiving the NOMV vaccines was not statistically significant. Variability in each group was high. However, a trend was observed as 6 of 7 mice vaccinated with Nm-Ng NOMV had no CFU, suggesting that the combination vaccine was more effective. A larger number of animals per group will be required to detect differences.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



