Submitted:
22 August 2025
Posted:
27 August 2025
You are already at the latest version
Abstract
Despite the oligotrophic conditions of the southeastern Levantine Sea and northern Red Sea, six lobster species—five slipper lobsters (Scyllaridae) and one spiny lobster (Palinuridae) maintain permanent, reproducing populations in the study area. Additionally, there are isolated records of four other sporadic lobster species. In the southeastern Mediterranean, permanent species include the Mediterranean Slipper Lobster, Scyllarides latus, Small European Locust Lobster, Scyllarus arctus, and Pygmy Locust Lobster, Scyllarus pygmaeus. In the northern Red Sea, they include the Clamkiller Slipper Lobster, Scyllarides tridacnophaga, Lewinsohn Locust Slipper Lobster, Eduarctus lewinsohni, and Pronghorn Spiny Lobster, Panulirus penicillatus. This review synthesizes current knowledge of their biology and ecology, including distribution, habitat, reproduction and development, feeding, predators and anti-predatory adaptations, behavior, sensory modalities, environmental impacts, threats, and conservation. Recent advances focus mainly on larger, commercially valuable species (S. latus, S. tridacnophaga, P. penicillatus), while major gaps remain for oceanic post-embryonic stages and the nektonic niso postlarva, as well as for smaller, often cryptic species (S. arctus, S. pygmaeus, E. lewinsohni). Addressing these gaps will require targeted research, using modern methodologies, in coastal, deep, and open waters, coupled with citizen-science surveys. While many Indo-Pacific decapods have been established in the Mediterranean, no immigrant lobster species have successfully colonized Levant waters, despite rare records of three non-indigenous species (NIS). However potential NIS predators and shifts in mollusk compositions, the main prey of some native lobsters, may affect the latter. Large lobsters remain targeted by fisheries despite protective regulations, which are not always effective or obeyed. No-take marine protected areas (MPAs) or nature reserves can be effective if sufficiently large and well-managed. Habitat loss from marine construction can be partly compensated by stable, environmentally safe artificial reefs tailored to lobster behavioral ecology. The categories of the studied lobsters’ species in the International Union for Conservation of Nature (IUCN) Red List of Threatened Species, last updated over fifteen years ago, should be re-evaluated.
Keywords:
slipper lobsters
; spiny lobsters
; phyllosoms
; biology
; ecology
; anthropogenic effects
; overfishing
; marine protected areas
; management
; future research
1. Introduction
Lobsters are crustaceans that, according to the classical taxonomy, belong to the suborder Macrura Reptantia, order Decapoda (all of which have 5 pairs of limbs), class Malacostraca, suborder Crustacea, order Arthropoda. Members of the latter group are characterized by a segmented body, two pairs of tentacles (antennae and antennulae) and biramous limbs that have undergone various specializations for feeding, grasping, walking, swimming, reproduction and sensory functions. Their body is covered with a hard exoskeleton composed mainly of chitin and calcium. Malacostracans include the largest crabs, including some species of lobsters, which are found exclusively in seawater.
Lobsters are generally divided into three infraorders. The first, Astacidea, includes the clawed lobsters’ family, which comprise species of significant commercial importance, most notably the American lobster, Homarus americanus. The second, Palinuridea, contains the spiny lobster family, which includes commercially valued species, and the slipper lobster family, some of which have economic value. Few members of the third group, Thalassinidea, have economic importance [1]. In more innovative classification approaches, lobsters are grouped into two main categories: those with claws (Homarida) and those without claws (Achelata), the latter encompassing the families Palinuridae, Scyllaridae and Synaxidae [1,2]. The present review focuses on clawed, spiny and slipper lobsters.
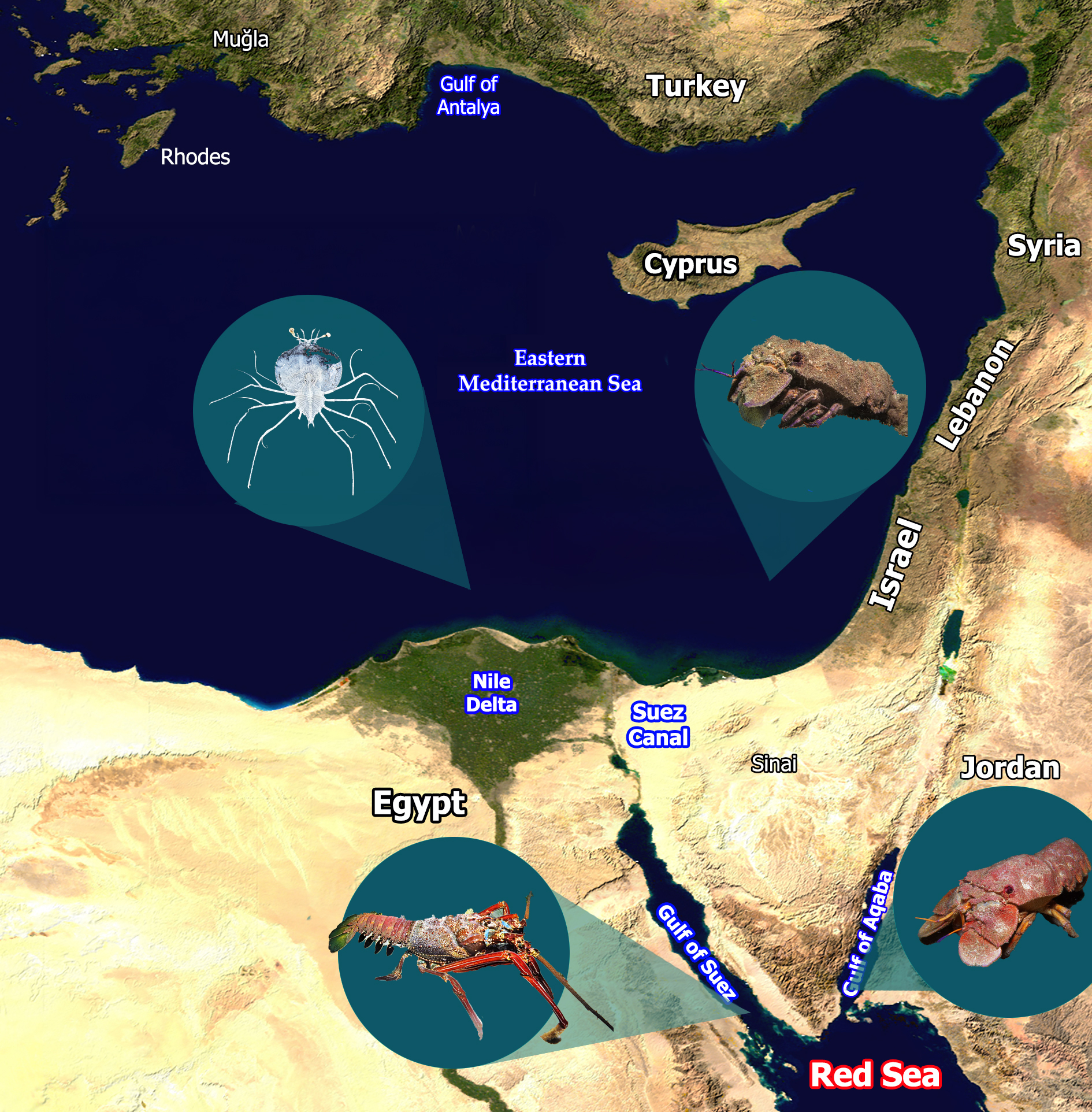
The Levantine Sea is the easternmost part of the Mediterranean Sea (Figure 1), located far from the Mediterranean’s connection to the Atlantic Ocean at the Gibraltar Straits. As a result, Levantine water has distinct physical and chemical, and therefore ecological, characteristics that are more extreme than the rest of the Mediterranean.
Environmental conditions in the Mediterranean change gradually from west to east: evaporation increases, precipitation decreases, salinity rises, water temperatures climb, nutrient levels in the photic zone drop, and primary productivity declines. These trends affect the entire food web, including fisheries, and result in the ultra-oligotrophic nature of the Levant Basin. Therefore, the southeastern Levantine coast is marked by low biological productivity and dynamic conditions, including continuous abiotic changes (e.g., rising water temperatures) and biotic shifts (e.g., the invasion of alien species). In recent decades, rapid population growth, particularly along coastal areas, has intensified human impacts, creating environmental threats, harming marine life, and generating conflicts between anthropogenic activities and the marine ecosystem [3].
Most indigenous marine biota in the Levantine Sea are of Atlantic origin, but since the opening of the Suez Canal in 1869, there has been a substantial influx of alien species from the Red Sea and Indian Ocean, a process known as Lessepsian (or Erythrean) migration (e.g., [4,5,6]). Levantine waters are warming at nearly twice the global average [7] and faster than other parts of the Mediterranean [8]. Climate change plays a key role in facilitating the spread of Lessepsian species [9]. Despite its oligotrophic nature and relatively low biomass, the Levant may host locally high biodiversity, driven by the prevalence of numerous species, including many non-indigenous species (NIS), and the presence of some unique habitats, such as hard substrates.
The coast of the Southeastern Levantine Sea and its adjacent inner shelf is divided into two main sedimentological provinces. The southern province consists of the margin of northern Sinai and of southern and central Israel, extending northward to northern Haifa Bay (Figure 1). This margin is relatively wide (ranging from about 50 km off Sinai to 10 km off Haifa) and the sea floor is generally smooth [3,10]. It is composed mainly of Nile-derived fine quartz sand and represents the northern flank of the Nile littoral cell, which extends roughly 650 km along the southeastern Mediterranean from Abu Quir Bay near Alexandria, Egypt, to Haifa Bay [3]. Since the construction of the Aswan dams, most sand supplied to this coastal system originates from erosion of the Nile Delta. The sediments are transported by longshore and offshore currents along the coasts of northern Sinai and Israel [11]. The northern province comprises the margins of northern Israel and Lebanon. It is narrow (3 to 12 km) and cut by numerous canyons. The latter are primarily submarine extensions of rivers of the Northern Israel and Lebanon [11]. The seabed here is dominated by hard substrates, with some areas covered by local coarse carbonate sand [12,13,14].
Several types of hard substrates occur along the Levantine coast, including kurkar, a local name for an aeolian, carbonate-cemented quartz sandstone, forming ridges parallel to the coastline. These ridges create rocky substrates rich in crevices and caves. Such complex submerged natural, as well as artificial, hard-bottom habitats, are important for many marine species, including certain lobsters’ species (e.g., [15,16]). Hard substrates are more common along the northern coasts. Kurkar ridges occur underwater mainly from the shoreline to depths of about 30 m. Some of the kurkar substrates appear, in the intertidal zone, in the form of abrasion platforms, especially prevalent in the northern part of the region [3]. Another rocky feature is beachrock, which contains abundant marine-derived particles such as shells and coarse sediment, rapidly cemented by calcium carbonate within the intertidal zone. Beachrock outcrops may extend hundreds of meters in length, exceed 40 m in width, and reach about 1 m in thickness. There are also conglomerate and lime rocks along the Northern Province of the region.
The Gulf of Suez (GoS) and the Gulf of Aqaba (Eilat) (GoA) form the northern arms of the Red Sea, separated by the triangular Sinai Peninsula (Figure 1). The GoS, the northwestern arm of the Red Sea, is relatively shallow, with depths measured in tens of meters. Only in its southern part, off Ras Mohamed, does a deeper trench extend into the gulf from the Red Sea. The southern boundary of the gulf is defined by a line running from Ras Mohamed to Shadwan Island and Hurghada. This line roughly follows the 200 m depth contour, marking the point where the Red Sea proper begins [17].
The northeastern arm of the Red Sea, the GoA, is deep, with a maximum depth of 1,829 m. Its coasts are either very narrow or absent altogether. On the western side, large alluvial fans extend below the sea surface and are cut by numerous marine canyons. The eastern side drops steeply into deep basins [18].
Coral reefs are the most important habitats in the northern Red Sea. Although the open water is extremely oligotrophic, these reefs support high biological diversity and feature efficient nutrient recirculation. The spatial complexity of coral reefs includes structures such as caves, crevices, tunnels, holes, and cavities, which provide habitat for a wide range of marine life, including some lobster species. Bays in the northern Red Sea contain coral patches, seagrass beds, muddy substrates, and coral reefs.
Globally, the future stability and persistence of coral reefs is uncertain due to global warming and recurrent bleaching events, which reduce the resilience of these ecologically and socioeconomically important ecosystems [19]. The Sea Surface Temperature (SST) of the Northern Red Sea is warming at a rate of a few hundredths ºC per year, comparable to the warming observed in the global oceans and the Mediterranean Sea [20]. Despite this, coral bleaching linked to rising sea temperatures has not yet been reported in the region. It has been suggested that the GoA, and possibly the GoS, may serve as reef refugia due to a unique combination of environmental conditions. This hypothesis is supported by experimental evidence showing an exceptionally high bleaching threshold for corals in the Northern Red Sea, as well as by ocean modeling that indicates potential dispersal of coral planula larvae through a selective thermal barrier. It is proposed that millennia of natural selection at the southernmost end of the Red Sea have favored coral genotypes less susceptible to thermal stress, potentially delaying bleaching events in the GoA by at least a century [19]. Nevertheless, even in the northern Red Sea, increased anthropogenic pressures from tourism, shipping, ports, and industrial activities have negatively affected these sensitive and unique ecosystems.
Other important marine habitats in the northern Red Sea include seagrass meadows, such as those formed by the fern seagrass, Halophila stipulacea [21]. These meadows provide coastal protection and support high biodiversity. They also efficiently fixate carbon, making a significant contribution to mitigating climate change.
Salinity in the northern Red Sea ranges from ~ 39‰ in the south to ~ 41‰ in the north, around the GoS [22]. This is similar to the average salinity in the upper mixed layer (0–10 m depth) of the open Levant Sea, of 39.75‰ [3]. Despite the similarities in salinity and temperature regimes between the northern Red Sea and the southeastern Levantine Sea, far fewer species have migrated from north to south (anti-Lessepsian migration) compared with the south-to-north Lessepsian migration. Lessepsian migrants may outnumber anti-Lessepsian migrants by three or more orders of magnitude [9]. Fewer than 10 species are recognized as anti-Lessepsian, compared with nearly 500 Lessepsian species [23]. The Red Sea is open to southern seas, and non-indigenous species (NIS) of lobsters may also be introduced via ballast water from ships travelling to commercial ports such as Aqaba in Jordan, Eilat in Israel (both at the northern end of the GoA), and Duba (NEOM) on the northern Red Sea coast of Saudi Arabia. Similar introductions may occur via ships traveling to the main ports in GoS: Suez Port, Ain Sokhna Port, and Adabiya Port, as well as vessels transiting the Suez Canal.
Lobsters are currently important resources across the world’s oceans, providing food security, employment, and valuable trade commodities. Humans have exploited lobsters since prehistoric times. They were a valuable food and economic resource for early coastal communities. Ancient Greek and Roman Mediterranean civilizations accumulated considerable knowledge about lobster biology and fisheries [24]. Despite their oligotrophic nature, the southeastern Mediterranean and northern Red Sea support a variety of spiny and slipper lobster species, some of which have been, and continue to be, harvested for food. Lobster use in ancient times varied widely, from complete prohibition in Jewish dietary laws to being regarded as a delicacy in the Roman world [25]. One of the earliest known depictions of a lobster is a wall carving in Egypt, illustrating Queen Hatshepsut’s five-ship trade expedition to the Red Sea in the 15th century BC (e.g., [26]). This bas-relief mural on a temple wall in Deir el-Bahari, Egypt, depicts a spiny lobster alongside other mostly edible tropical marine animals. Some scholars suggest that this lobster represents the Pronghorn spiny lobster, Panulirus penicellatus [27].
The present article reviews the latest available information on the biology, ecology, distribution, reproduction, development, feeding, predation, environmental impacts, threats, and conservation of the known lobster species in the studied area.
2. Lobsters from the Southeastern Mediterranean Coast
Only slipper lobsters of the family Scyllaridae are known to be permanent residents along the Mediterranean coast of the Levant. These include the Mediterranean slipper lobster, Scyllarides latus, the Small European locust lobster, Scyllarus arctus, and the pygmy locust lobster, Scyllarus pygmaeus. They are also called “shovel-nosed’ lobsters, a name that refers to their flattened, shovel-like antennae characteristic of slipper lobsters.
2.1. The Mediterranean Slipper Lobster
The Mediterranean slipper lobster, Scyllarides latus (Latreille, 1802) (Figure 2), is a relatively common species along the southeastern Mediterranean coast. The etymology of its scientific name derives from skylarlos (Greek) meaning a kind of crab, and latus (Latin) meaning broad [28,29]. According to the FAO, its common names include Grande Cigale or Grosse Cigale in French, and Cigarra or Cigarra de Mar in Spanish [1,29]. Other vernacular names for this species are: Flat Lobster (UK); Magnosa, Cicala Grande (Italy); Cavaco, Lagosta de pedra (Portugal); Astakoudaki Megalo (Greek); Büyük Ayı Istakozu (Turkey); Cigale Noire (meaning "black cicada") and Farzit (Tunisia) [29]; additional common/local names include Cigale courte (Algeria); Feritah (Morocco); Caravida (Greece); Sapa (Spain); Homard plat, Macieta (France); Ccala hamra, Ccala seula (Malta) [30]; and Kapan Gushmani (referring to a spoon-shaped creature with connected, lumpy parts) in Israel (e.g., [31]).
Due to its considerable size and its economic and ecological importance, knowledge about S. latus is relatively well-established compared to other scyllarids species reported along the Levant coast. This is largely thanks to studies conducted mainly in Israel, Italy, Spain, and the Azores Islands. Nevertheless, several aspects of its biology and ecology remain unknown.
The Mediterranean slipper lobster belongs to the genus Scyllarides, which includes 14 species found in temperate and tropical waters. These lobsters are members of the subfamily Arctidinae, which also includes the genus Arctides, comprising three species [32]. The family Scyllaridae is the most diverse among lobster groups in terms of both species number and variety, encompassing four subfamilies and at least 89 species. Slipper lobsters are taxonomically closely related to spiny lobsters (family Palinuridae) and furry lobsters (family Synaxidae); together, these families form the Achelata group of lobsters [1,2]. They differ from other decapod crustaceans in several ways, primarily due to their unique larval stage known as the phyllosoma (Figure 3 and Figure 7). Additionally, their cephalothorax is flattened and features lateral lips, while the second pair of antennae are short and broadened into large plates—often mistakenly referred to as "shovels" or "fins."
Like other lobster species, S. latus is primarily nocturnal. During the day, it typically hides in dens, burrows, caves, and dark crevices within the rocky substrate [33]. However, initial data from a recent citizen science survey along the Mediterranean coast of Israel surprisingly revealed several instances of S. latus being observed during daylight hours. This species is found close to the coast on the shallow continental shelf (up to about 30 m depth) only part of the year, from around February to July, when seawater temperatures are relatively cool. During most of the summer and autumn, it is rarely seen in these shallow areas [15,34], likely migrating to deeper waters and returning to the shallows the following winter.
The maximum length of Scyllarides latus can reach is up to 45 cm, although it typically does not exceed 30 cm, with an average weight of about 1.5 kg. In recent decades, smaller sizes have been reported, likely due to fishing pressure and fishermen’s preference for larger individuals [33]. Notably, Atlantic specimens tend to be heavier than those from the southeastern Mediterranean [15]. Nonetheless, S. latus remains the largest lobster species in the southeastern Mediterranean Sea and has historically been, and continues to be, a target for fisheries. It is commonly found in rocky habitats at depths ranging from approximately 4 to 100 meters [1], though it has occasionally been reported at depths of up to 400 meters [35]. Its distribution spans the Mediterranean Sea as well as the central eastern Atlantic Ocean, from the coast of Portugal to Senegal, including Madeira, the Azores, Canary, and Cape Verde Islands [1]. The lobster is yellowish-brown in color, with the edges of its flattened antennae (the "shovels"), antennulae, and legs often displaying a bright purple hue (Figure 2).
2.1.1. Reproduction and Development
The breeding season in the study area lasts from April to July. The sexes are separate, with the female-to-male ratio in the population being roughly equal [35]. In adults, distinguishing females from males is straightforward by examining the ventral surface of the body. Females have larger, leafier, and broader pleopods (abdominal appendages) used to hold the eggs. Their fifth walking legs (pereiopods), which handle the eggs, are chelated (bearing pincers), whereas males’ fifth legs lack pincers. The genital openings are located at the base of the third walking legs in females and at the base of the fifth in males. In the Levant, males with white gelatinous spermatophores at the base of the fourth and fifth walking legs have been observed both at sea and in the laboratory during early spring, when water temperatures reach at least 17ºC [15]. Unlike some other lobster species, mating in this species is not associated with molting. In laboratory observations in the Levant, females were found carrying spermatophores 6–10 days before eggs appeared. Egg-bearing females have been reported both at sea and in the laboratory at water temperatures between 17º and 27ºC. The eggs have a diameter of approximately 0.7 mm, and their number is directly proportional to the female’s size, ranging from about 100,000 to 356,000 [32,34]. Such high fecundity is assumed as an adaptation to compensate for larval losses in the open sea and the variable recruitment of juveniles due to cyclical changes in the marine environment [36]. The eggs initially appear yellow orange (Figure 3) and darken brown as they develop. Along the Levantine coast, females carry their eggs on the abdomen for 4–6 weeks [15], while in the Atlantic Ocean, this period extends to 6–8 weeks [37] before the eggs are released into the water. It has been reported that females may lose some or all of their eggs if disturbed by predators or humans, either at sea or in laboratory settings [24,35].
The egg hatches into a “prelarval” stage called the naupliosoma, which measures about 1.3 mm in length. This stage does not feed and survives for no more than five hours before molting into the first stage of the phyllosoma larva [38]. The first stage of the phyllosoma larva lives for approximately six days and reaches about 1.8 mm long. It is a flat, transparent, leaf-like larva (Figure 3) that drifts in the pelagic zone, far from the shore. Feeding primarily on soft, gelatinous animals, it grows for nearly a year, undergoing 11 molts during this period. The final phyllosoma stage, which can reach up to 48 mm in length [38], then metamorphoses into the nisto, or post-larva. The nisto stage is neither fully planktonic nor entirely benthic; it represents the transition from life in the water column to life on the substrate. For many years, this stage was not observed in nature, but a lengthy search of museum collections uncovered a specimen in the Zoological Museum in Turin [39]. This tiny and rare specimen, measuring 38.7 mm (Figure 3), was collected in southern Italy in the early 20th century, likely from waters deeper than 800 m. It is assumed that individuals in this stage settle in deeper waters where they are better protected from predators. They then develop into small juveniles and gradually migrate along the bottom toward shallow adult habitats, molting and growing until they join the juvenile and adult population in coastal waters.
Although Mediterranean slipper lobsters mate and lay eggs that hatch readily in the laboratory, scientists have not yet been able to complete their full life cycle or successfully raise them from eggs in captivity.
2.1.2. Food and Predators
The Mediterranean slipper lobsters feed mainly on clams [40], which they forage in the open area after sunset. They sometimes bring clams into their dens, probably those they have collected before dawn [15]. They open the clams with their strong pereiopods after investigating the clam using their antennulae and cutting the clam's closing muscles [41].
S. latus appears, at first glance, defenseless. Clawed lobsters, such as H. americanus (Figure 10), are equipped with large claws that may be used for defense (but also for gripping and cracking food), and the bodies of spiny lobsters, such as the Pronghorn spiny lobster from the northern Red Sea (Figure 15), are covered with spiny armor and have long, spiny antennae that may be used to ward off predators. Apparently, scyllarids have no morphological adaptations against predators. However, it turned out that they have both morphological and behavioral adaptations against predation. First, they usually hide during the day in horizontal dens, whose shade protects them from detection and physical harm, usually by diurnal predators. They prefer natural or artificial shelters (artificial reefs (AR), ships’ wrecks) that have at least one extra escape opening in addition to the entrance [42]. They also tend to choose dens with small openings, which add to the physical protection of the dens as well as increase their shading. They often hide in dens with other individuals of their own species [41]. When they emerge from the dens in the evening to forage or for social interactions, their color blends with the rocky environment in the twilight light. If they are attacked, their thick armor may protect them from the initial assault of a typical diurnal predator such as the Mediterranean triggerfish, Batistes carotinensis, with its strong teeth. Their coarse carapace (exoskeleton) has been found to be at least twice as thick as that of clawed and spiny lobsters [43]. The special internal structure of the carapace [44] also makes it more resistant to mechanical damage than the carapace of lobsters from other families. During an attack, the slipper lobsters protect their mouthparts and delicate antennulae with their flat, stiff antennae ("shovels"), and their eyes converge into eye sockets protected by strong spines. Their short, strong legs, equipped with strong cloves, are used to cling to the rocky bottom or the walls of their den, to prevent the predator from turning them over and exposing their ventral side, where their carapace is thinner and more vulnerable. Measurements, using spring scales, have indicated that the clinging force of slipper lobsters to a rough, hard substrate (or the force required to detach the lobsters from such a substrate) ranges from 3 to 15 kg and is directly proportional to the size (weight) of the lobster. This clinging force is equivalent to 8 to 29 times the lobster's body weight [15]. Experiments at sea have shown that the survival of S. latus that hid during the day in dens in an AR was significantly higher than that of their conspecifics that were at the same time in the open, and most of them were prey upon by Mediterranean trigger fish [45].
If the “fortress defense” strategy fails, or if a predator manages to enter the den, the lobsters escape through alternative openings and switch to a rapid getaway swim. Slipper lobsters, which normally walk slowly on the substrate, use powerful tail flips in emergencies, rapidly propelling themselves backward by repeatedly striking their muscular and flexible abdomens. This burst-and-coast swimming style, characterized by large tail movements propelling the lobster quickly backward, followed by a powerless glide, is also seen in some fast-swimming, negatively buoyant fish and other crustaceans. During acceleration, swimming speeds can reach up to three body lengths per second (about one meter per second), while during the glide phase speeds drop below one body length per second [46]. Escape swimming is short-lived and used only in emergencies to reach safety, such as an alternative den, due to its high energy demand. Lobsters can also abruptly change swimming direction, likely to confuse predators [32]. The flattened second antennae of S. latus, often mistaken for “shovels” or “flippers”, have movable joints that act as stabilizers or rudders to control swimming movements [46,47] (Figure 4). The tail fan (telson) may also contribute to steering and stabilization during swimming.
Another potential predator of S. latus is the common octopus, Octopus vulgaris. The lobsters have frequently been observed sharing dens with Mediterranean moray eels, Muraena helena, without any evident predator-prey interactions between them [42]. It is speculated that this shared shelter may be mutually beneficial: since the octopus is prey for the moray, the lobsters might gain protection from the octopus by co-sheltering with the moray. Conversely, the moray may benefit from the presence of lobsters attracting octopuses to the den. However, these possible mutualistic relationships require further investigation.
2.1.3. Sensory Modalities
The sensory world of adult S. latus is less understood compared to other lobster groups. In other lobsters, chemical sensation is typically divided into near-field “taste” and far-field “smell.” However, observations of S. latus feeding behavior suggest that this distinction may not be as clear [41]. Lobsters possess tufts of sensory hairs (setae) across many parts of their bodies, but notably not at the tips of their legs. Despite this, S. latus can locate and retrieve clams buried several centimeters deep by using the claws of their walking legs, while simultaneously increasing the fluttering rate of their antennulae [15]. This behavior has also been observed in laboratory aquariums when clams were put into it.
The antennulae are covered with various types of sensory hair setae that likely serve as chemical and mechanical receptors [50]. The mechanical receptors are also used for hydrodynamic sensing during swimming, as are the numerous sensory hair setae located on the lobsters' "shovels," which aid in steering during “take-off,” “landing,” and changes in direction. These sensory structures provide crucial feedback during escape swimming.
The ventral, and especially the dorsal, side of the S. latus exoskeleton, like those of other decapod crustaceans, is covered with numerous stiff sensory hair setae, which probably function as mechanical receptors.
The eyes, situated on stalks, also allow for backward vision. In response to a direct threat, they converge into foveae, each protected by a stiff spine. Anatomical studies of S. latus eyes [51] indicate that they resemble those of nocturnal decapod crustaceans and are adapted for vision in dim light. Additionally, these lobsters appear capable of perceiving polarized light—a crucial adaptation against predation, considering that several species of triggerfish, which are among the lobsters’ predators, also possess this ability.
Statocysts are known in other lobster families and are used to maintain balance, but they have not yet been reported in slipper lobsters. The fact that S. latus responds immediately with a powerful “tail flip” of its muscular abdomen when turned over on its back [45] suggests that its statocysts are well developed.
Since S. latus appears capable of returning to the same preferred dens in hard substrate after a year [52] or even several years [34], how do they navigate such great distances? They may use magnetic cues or a geomagnetic “map,” as reported in spiny lobsters [53]. However, this aspect has not yet been studied in slipper lobsters.
Despite their French common name, Cigale de Mer [29], meaning “sea cicada,” which refers to the sound the lobster makes when rubbing its lobes together upon being taken out of the water, it remains unclear whether S. latus can hear or produce sounds like some other lobster species. Additionally, the sensory mechanisms of juvenile slipper lobsters are currently unknown.
2.1.4. Environmental Impacts, Threats and Conservation
The main threats to S. latus populations are overfishing, habitat loss, and climate change [31,32]. Due to its large size and high market value, S. latus is heavily fished throughout its range [54], including in the southeastern Levant. Fishing methods primarily include diving and the use of trammel and gill nets. The removal of large individuals with high reproductive potential poses a serious threat to the species’ populations. In some areas of the western Mediterranean, such as Italy, it is estimated that the population may be unable to recover at all [55]. However, genetic studies comparing S. latus individuals from different sites in the eastern, central, and western Mediterranean, as well as the eastern Atlantic Ocean [56,57], have revealed little genetic variation across this broad distribution. This phenomenon, known as panmixia, refers to a population’s ability to reproduce freely across a very wide geographic range spanning hundreds to thousands of kilometers. The researchers attribute this to the species’ high fecundity and prolonged larval stage in the open sea. Given these findings, which indicate strong connectivity between geographically distant S. latus populations, researchers suggest that future conservation efforts should treat all populations as a single stock [57]. For example, larvae from healthy populations around the Balearic Islands in Spain could potentially help restore depleted habitats in Italy, where Mediterranean slipper lobster populations have been nearly wiped out.
The main threats to the populations of S. latus are overfishing, habitat loss and climate change [31,32]. Due to its size and its high market value, S. latus is intensively fished throughout its range [54], including the southeastern Levant. Fishing is mainly done by diving and using trammel and gill nets. Removing large individuals with high reproductive potential from the sea is a serious threat to populations of this species. It is estimated that in some areas of the western Mediterranean, such as Italy, the population will not be able to recover at all [55]. However, genetic studies comparing individuals of S. latus from different sites in the eastern, central and western Mediterranean, and the eastern Atlantic Ocean [56,57] have indicated a lack of genetic variation between specimens of S. latus across the wide distribution range examined. This is a phenomenon of panmixia - the ability of individuals in a population to reproduce without restrictions over a very wide geographical range of hundreds to thousands of kilometers. The researchers explained this by the high fecundity of the S. latus and the prolonged larval stage in the open sea. Considering these findings, which indicated a high connectivity between geographically distant populations of S. latus, the researchers suggested [57] that future conservation strategies should manage all populations of this species together (as a single stock). For example, larvae from healthy populations of S. latus from the Balearic Islands in Spain may, therefore, restore suitable habitats in Italy where the populations of the Mediterranean slipper lobsters have been almost completely depleted.
In Israel, S. latus has been a protected species since 2005; however, illegal fishing continues. Due to the extensive marine area and insufficient number of inspectors/ rangers, the most effective method for protecting this species is through marine nature reserves. A study conducted in the Rosh Hanikra–Achziv Marine Reserve, a fully protected Marine Protected Area (MPA), and a comparable control area on Israel’s northern coast [52] found that lobster abundance, density, and size were significantly higher within the protected reserve. Additionally, marked lobsters in the reserve tended to return to their dens during winter, unlike those in the control area. It is assumed that reserves may also benefit areas outside them by allowing adults, juveniles, and larvae to migrate into unprotected areas. Similar MPAs exist along the Mediterranean coast of Lebanon, such as the Palm Islands Natural Reserve in northern Lebanon (e.g., [58]), the Tyre Coast Nature Reserve in southern Lebanon [59], and the Sallum MPA, the first MPA on Egypt’s Mediterranean coast [60].
The destruction of preferred habitats for S. latus, such as Submerged Kurkar Ridges (SKR) rich in caves, ravines, and natural dens [3], is another factor impacting their populations. Preserving these habitats is essential for the conservation and recovery of Mediterranean slipper lobsters and other threatened species. Environmentally safe and stable ARs, designed based on the lobsters’ behavioral ecology, may provide effective hiding places and potentially serve as substitutes for destroyed natural habitats [61].
Rapid coastal urbanization, demographic growth, and industrial development can lead to chemical pollution in the shallow shelf. Elevated concentrations of trace metals, copper, zinc, and iron, were detected in S. latus specimens collected near Beirut compared to those from other coastal sites in Lebanon [62].
The sharp increase in seawater temperatures off the southeastern Levant coasts in recent years—due to climate change, reaching values above 31°C [3] may be harmful to S. latus. In laboratory conditions, lobsters’ molting can be partial and complicated [63], sometimes resulting in death when kept for two months in seawater at 26°C or higher [41]. This may explain why lobsters migrate during the warm seasons to deeper, colder waters, which allow them to molt normally from a physiological standpoint. Molting involves shedding the old shell (exuvia) and growing a new one underneath, enabling the lobster to grow. However, during this time, the lobster’s new outer shell remains soft, making it more vulnerable to predation. The migration to deeper waters may also be linked to reduced predation pressure in the darker depths. Information on the timing of molting is very limited and mainly based on a few reports of lobsters’ exuviae found on the Israeli coast, mostly between November and February [15].
A laboratory study [64] indicated that slipper lobsters increase their activity as water temperature rises. These findings suggest that rising temperatures in the southeastern Mediterranean could affect the lobster’s movements and seasonal distribution, potentially causing specimens to migrate to deeper waters earlier in the season. Climate change is also increasing the frequency and intensity of extreme weather events such as storms [3]. Notably, stranding of S. latus during two severe storms was recently reported off the northern coast of Israel [65].
The Mediterranean slipper lobster is classified as “Data Deficient” on the International Union for Conservation of Nature (IUCN) Red List of Threatened Species [29]. Protecting a threatened species requires comprehensive information about its biology and ecology. However, because S. latus is primarily nocturnal and its populations are scattered, it is challenging for a small group of scientists to gain a complete understanding of its ecology. Consequently, the public, especially sea enthusiasts along the Mediterranean coast of Israel, was invited to assist in gathering data, such as the locations and timing of lobster exuviae and live sightings, through a “citizen science” approach [66]. Although the Israeli project has been brief, it has already yielded a few important reports that have contributed to expanding the biological knowledge of this species.
2.2. Locust Lobsters
Locust lobsters belong to a single subfamily, Scyllarinae, which included only one genus, Scyllarus, and at least 40 species. Most of these species are small and only a few have marginal commercial value, typically caught as by-catch [1]. Since 2002, some of the species have been assigned to other genera [67], but the two species known from the southeastern Mediterranean coasts remain in the genus Scyllarus.
2.2.1. The Small European Locust Lobster
The Small European locust lobster, Scyllarus arctus (Linnaeus, 1758) (Figure 5). Etymology of the scientific name: skylarlos (Greek) – a kind of crab or perhaps derived from skullow (Greek) – torn or ragged (alluding to the antennal margins); arctus (Latin) –narrow (i.e., compared with other species), bear [28,29]. The FAO names for this species are - Petite Cigale (French) and Santiaguiño (Spanish)[1]. Other common or vernacular names include: Broad lobster (UK); Chambre, Petit scyllare (France); Cicala di mare, Magnosella (Italy); Lameiro, Lagosta da pedra (Portugal); Bujias, Cigarra, Toribo, Xuius (Spain); Chkal, Ziz il bahr, Petit cigal de mer, Cigal blnche (Tunisia); Astakouáki (Greece); Kapanit dubit (Israel)(refers to a small spoon-shaped bear-like lobster); Lesser slipper lobster (highlights its smaller size compared to other slipper lobsters) [30].
S. arctus is widespread throughout the Mediterranean Sea and the eastern Atlantic Ocean from the southern coast of the British Isles to the Azores, Madeira and the Canary Islands. It inhabits rocky substrates, often with caves, crevices and other shelters, as well as mudflats and seagrass beds, at depths of 4 - 50 m and occasionally beyond 200 m [35]. The species also occurs on ARs such as shipwrecks. Reports indicate seasonal migrations, with individuals moving to deeper waters in autumn and returning to shallower areas for breeding in spring. Total body length typically ranges from 5 - 10 cm, with a maximum body length of ~ 16 cm. S. arctus is of secondary importance, taken mainly as by-catch in coastal fisheries, for example along Spain’s Atlantic coast [68]. @@@
The Small European locust lobster is a strikingly colorful species. Its rectangular cephalothorax bears three prominent central teeth. The body is reddish-brown with whitish scale tips and a dark brown spot in the center of each abdominal somite, though this marking is not sharply defined. The antennae ("shovels") are serrated. The abdominal joints are light blue with an orange stripe, forming a complex, fern-like pattern (Figure 5). The legs are yellow with dark blue stripes and red tips, while the bases of the eyes are orange red. Males are smaller than females. Like other slipper lobsters, this species is nocturnal, spending the day hidden in dens and caves where it is sheltered from predators, particularly benthic fish such as groupers
2.2.1.1. Reproduction and Development
In the Mediterranean Sea, egg-bearing females have been observed year-round, with peak occurrences in March–April and July–September. Histological examinations confirm these as the main breeding periods. Each female carries 30,000–70,000 eggs, measuring 0.4–0.5 mm in diameter and yellow orange in color [35]. In laboratory conditions, larvae pass through 12–16 instars before molting to the benthic stage (nisto) after 192 days [35]. An oceanographic survey found that larvae are most abundant in eddies located about 250 km off the Israeli coast, which likely provide a suitable habitat for their early developmental stages [69].
2.2.1.2. Threats, Environmental Impacts and Conservation
S. arctus is listed as “Least Concern” on the IUCN Red List of Threatened Species [70] due to its wide distribution and the fact that it is only fished in part of its range. While local overexploitation does occur in some areas, and population declines have been recorded there, the species’ ecological characteristics have made it relatively resistant to extinction. This resilience is largely attributed to its high fertility and well-connected populations, supported by long-lived larvae that can survive for more than six months.
A 2009 assessment recommended continued monitoring of exploitation levels to detect potential increases in fishing pressure, alongside stricter enforcement and adjustments to current fishery management regimes [70]. For example, in northwestern Spain, the legal minimum body length for harvesting S. arctus (9 cm) ensures that sexually mature individuals have the opportunity to reproduce at least once a year. However, the area’s long reproductive cycle (about 11 months), combined with the near year-round presence of egg-bearing females and the prohibition on catching them, results in fishing being heavily biased toward males. This imbalance could reduce the reproductive potential of the stock due to a shortage of males [68].
The species is considered one of the most common crustaceans along the Lebanese coast [71], but it is likely less abundant in the southern waters of Israel and Egypt. It has not been recorded off the North African coast east of Morocco. Protection of Posidonia seagrass beds, a preferred habitat for S. arctus in areas where they occur, further enhances conservation of the species [70].
2.2.2. The Pygmy Locust Lobster
The pygmy locust lobster, Scyllarus pygmaeus (Bate, 1888) (Figure 6), is the smallest species of slipper lobster, with males reaching a maximum length of 54 mm and females up to 65 mm.Etymology of the scientific name: skylarlos (Greek) – a kind of crab, or perhaps derived from skullow (Greek) – torn or ragged (alluding to the antennal margins); pygmaeus (Latin) – dwarf or pygmy [28].FAO names: in French – Cigale naine; in Spanish – Cigarra enana [1]. Other common or vernacular names include: nain cigale (France); Cicala minore (Italy) [30]; Kapanit Zutit (Israel) [72], meaning “small spoon-shaped miniature lobster.”
The pygmy locust lobster occurs at depths of 5–100 m [1], inhabiting sandy substrates, deep coral areas, and seagrass beds dominated by Caulerpa prolifera and Neptune grass, Posidonia oceanica, as well as caves in the Mediterranean Sea and around the Canary, Cape Verde, and Madeira Islands in the Atlantic Ocean. It has not been recorded off the coast of North Africa east of Morocco [1]. This species sometimes burrows into soft substrates and has also been collected at depths of up to 1,200 m [35]. In the Mediterranean Sea, it is considered less common, likely because individuals are sometimes misidentified as juveniles of the sympatric S. arctus, due to their similar serrated, flattened antennae (“shovels”). They can be distinguished mainly by size, S. arctus being larger, but also by specific morphological traits, such as the tubercle on the last thoracic sternite, which is flattened in S. arctus and conical in S. pygmaeus [73].
The coloration of S. pygmaeus is pale brownish or pinkish, with patches of darker setae (Figure 6) and bluish or orange stripes along the abdominal joints. The carapace is compact and bears small ridges. Their diet consists primarily of small live bivalves and occasionally dead fish.
2.2.2.1. Reproduction and Development
In the western Mediterranean Sea, females carrying eggs have been observed year-round, with peak occurrence between April and July. In the eastern Mediterranean, the breeding season appears to be longer, and females with eggs have even been recorded in December at depths of 27–82 m [74]. Little is known about the larvae and early benthic stages. Recently, a stage VII phyllosoma larva of this species was reported from the Levant (Figure 7). It was collected far from shore, in the central part of the Israeli exclusive economic zone, at a depth of 0.5–1 m in waters with a bottom depth of 1,100 m [75]. This constitutes the first record of the larva of S. pygmaeus in the eastern Mediterranean Sea.
2.2.2.2. Threats, Environmental Impacts and Conservation
S. pygmaeus is classified as "Least Concern" on the IUCN Red List of Threatened Species. This is due to its wide distribution in the Mediterranean and eastern Atlantic, its small size and lack of economic interest. There is a local decline in the population in certain areas [76].
2.3. Occasional Species on the Mediterranean Coast
Only three lobster species have been recorded on the southeastern Mediterranean coasts, each reported just once. These include an adult male ornate spiny lobster, Panulirus ornatus (Fabricius, 1798) (Figure 8), caught in 1988 near the Haifa Port breakwater [77]; and a complete exuvia of an adult female long-legged spiny lobster, Panulirus longipes longipes (A. Milne Edwards, 1868) (Figure 9), found in the Haifa coastal waters in 2018 [78]. Both specimens were likely Lessepsian migrants [3]. In late 2022, two American clawed lobsters, Homarus americanus (H. Milne Edwards, 1837) (Figure 10), were reported on Arches Beach in Caesarea [79], on Israel’s central coast, probably originating from lobsters imported from North America for restaurants in Israel.
3. Lobsters from the Northern Red Sea
On the coasts of the GoA and GoS there are representatives of both the families of slipper lobsters, Scyllaridae, and spiny lobsters, Palinuridae. Lobsters of the later family are characterized by a very long thick, spiny antennae that are longer than the body length and the four front pectoral legs without chelae.
3.1. Clamkiller Slipper Lobster
The Clamkiller Slipper Lobster, Scyllarides tridacnophaga Holthuis, 1967, closely resembles its Mediterranean congener, S. latus. Likewise, it is primarily nocturnal, hiding in dens during the day [80]. The etymology of its scientific name derives from Greek: skylarlos meaning a kind of crab, and tridacnophaga combining tridacna (referring to giant clams) and the suffix -phaga (meaning "eater" or "devourer"), reflecting its habit of preying on giant clams [29]. FAO common names include “Clamkiller slipper lobster” (English), “Cigale de mer” or “Cigale Marie-carogne” (French) and in Saudi Arabia and Egypt (Arabic) “sartan al-baḥr 'abu shḥāṭa” (the sea crab abu shkhta) and “Saltu'un malikat al-rukhawiyat” (literally, Crab of the Queen of Mollusks). In Hebrew, it is called “Kapan Hatsdafot” (literally, The Clams’ Slipper Lobster) [72].
This species is the largest slipper lobster in the Northern Red Sea, with a maximum length up to 40 cm, though the average size is about 30 cm. It typically inhabits coral reef areas or rocky substrates in shallow waters, ranging from 5 to 112 meters deep, and has also been found in seagrass meadows, such as those dominated by Halophila stipulacea, and sandy bottoms. Its geographical distribution spans the Indo-Pacific region, from the western Pacific Ocean along the western coasts of Thailand, India, and Pakistan, extending to East Africa (Somalia and Kenya), the Gulf of Aden, and the Red Sea [1].
The color of S. tridacnophaga is generally reddish to yellowish-brown, featuring a mottled pattern on the dorsal side and pale orange on the ventral side (Figure 11). Most of the bumps on the shell, as well as the edges of the flattened antennae, are reddish. The flagella at the tip of the orange antennulae are deep purple. The legs are orange, with purple joints. The upper part of the first abdominal segment is mostly smooth and bears three large, sharply defined red spots—one central and two lateral—all round in shape (Figure 12). The spaces between the spots are approximately as wide as the spots themselves. In contrast, the Mediterranean S. latus displays a central red spot surrounded by a yellow ring, flanked on each side by two prominent, non-round red spots separated by very narrow spaces [80].
3.1.1. Food
S. tridacnophaga is able to open giant clams of the genus Tridacna. These clams anchor themselves to hard substrates using byssal threads. The lobster manipulates the clam to expose its dorsal surface, where the byssal thread attachments protrude, and then plunges its dactyls (the terminal segments of the legs) into this vulnerable area, causing the clam to gape. It likely destroys the adductor muscle or damages the nervous system that controls shell closure. At this stage, the lobster overturns the shell, inserts its legs to further wedge the valves open, and feeds on the living tissues [81]. In addition to giant clams, it also preys on other clams and scavenges dead fish [1]. Individuals sometimes forage in groups of three or more and exhibit gregarious sheltering during the day.
3.1.2. Reproduction
The breeding season occurs during the spring and summer months. During courtship, S. tridacnophaga rely on their senses of smell and touch. Fertilization is usually external, with the female carrying the eggs until they hatch, after which the larvae are released. The Marine Science Station (MSS) in Aqaba, Jordan, has successfully maintained several clamkiller slipper lobsters in captivity. At the field laboratory of the Central Marine Fisheries Research Institute (CMFRI) in Madras, India, four individuals, both males and females, were kept. Reproduction occurred in late June, producing round eggs approximately 0.5 mm in diameter. The eggs were initially light yellow, gradually turning orange and then rusty brown. Adult growth is slow, particularly in males [82].
Recently, a single-file queue of three S. tridacnophaga was observed in the evening on a sandy bottom north of a shipwreck in Eilat, Israel, at the northern edge of the GoA (Figure 12). This behavior resembles the queuing reported for Caribbean spiny lobsters, Panulirus argus, during their autumnal mass migrations (e.g., [83]). It remains unclear whether this behavior is related to foraging, reproduction, defense, migration, or other activities.
The breeding season occurs in the spring and summer months. During courtship, S. tridacnophaga use their sense of smell and touch. Fertilization is usually external, and the female carries the eggs until they hatch, and the larvae are released. The Marine Science Station (MSS) in Aqaba, Jordan, has successfully kept several clamkiller slipper lobsters. In the field laboratory of the Marine Fisheries Research Institute (CMFl) in Madras, India, four individuals, males and females, of were kept. Reproduction took place in late June, and the eggs were round, about 0.5 mm in diameter, light yellow in color, which gradually turned orange and rusty brown. Adult growth is slow, especially in males [82].
3.1.3. Threats, Environmental Impacts and Conservation
S. tridacnophaga is listed as “Least Concern” on the IUCN Red List of Threatened Species. However, due to its large size, it is caught in Kenya, either by diving or as bycatch in trawl fisheries, and probably also in other parts of its range. A continuing decline in the number of adult individuals has been reported [84]. It is also occasionally entangled in ghost nets (discarded or abandoned fishing gear).
3.2. The Lewinsohn Locust Slipper Lobster
The Lewinsohn Locust Slipper Lobster, Scyllarus lewinsohni (Holthuis, 1967) [85], has since been reclassified as Eduarctus lewinsohni (Holthuis, 1967) [85,86].Etymology – The genus name Eduarctus combines part of the name Eduard, honoring Eduard von Martens, for whom the type species was named, with the generic name Arctus De Haan, 1849. The word arctus (Latin) means “narrow” (i.e., compared with other species) or “bear” [28]. The species name lewinsohni commemorates the late Israeli crustacean expert Chanan Lewinsohn [85]. Its Hebrew name is “Kapanit Lewinsohn” (Lewinsohn locust lobster) [72].
Very little is known about this small species. On the IUCN Red List of Threatened Species, it is listed as “Data Deficient,” with a noted decline in the number of mature individuals in its fragmented population. However, this assessment dates back to 2009 [85].
Due to its small size, it has no commercial value and is not threatened by fishing. It has been recorded at depths of 20–82 meters in the Red Sea north of Bab-el-Mandeb (the southern entrance to the Red Sea), in the southern GoS, and in Eilat, Israel, the GoA. The cephalothorax measures 6–12 mm in length, with a maximum overall size estimated at 3–4 cm. Like other locust lobsters, its carapace is highly compressed and features small ribs on the surface. The coloration of E. lewinsohni is reddish-brown on the sides of the cephalothorax, marked with several large white spots and a few smaller dots of similar color. The abdominal segments display a complex red and white fern-like pattern in the center and along the sides [85]. The legs are yellow with dark lines (Figure 13).
3.3. Evidence of a New Species of Pygmy Slipper Lobster in Eilat the Gulf of Aqaba—Biarctus sordidus
A phyllosoma larva of a pygmy slipper lobster species, previously unknown in the Red Sea [87], was recently sampled in the northern GoA (Figure 14). The distribution of this species, the pygmy slipper lobster, Biarctus sordidus (Stimpson, 1860), whose English name is similar to the Mediterranean Scyllarus pygmaeus, was previously known only from the Far East to the Persian Gulf [85]. Etymology: Bi- means “life” (Greek), arctus means “narrow” or “bear” (Latin), referring to its comparatively narrow shape [28,29]. Sordidus (Latin) means “dirty” or “foul.” The taxonomy of B. sordidus has changed over time. Initially, it was classified as Scyllarus sordidus and Arctus sordidus, but subsequent studies reclassified it as Biarctus sordidus [85,88]. This species is closely related to other slipper lobsters, with its classification based on both morphological and molecular data. The larva was collected using a plankton net towed horizontally at a depth of 0.5–1 m, over a bottom depth of 700 m. The specimen was identified as a phyllosoma stage III/V of B. sordidus based on morphology and DNA barcoding. Researchers suggest that previous Red Sea biodiversity surveys may have confused B. sordidus with its congener B. pumilus, which is known from the Red Sea [85]. An additional/alternative explanation for the late detection of B. sordidus is that the species was recently introduced via ballast water of a merchant ship that arrived at the port of Eilat, Israel at the north edge of the GoA, or the close-by port of Aqaba, Jordan from the Far East. This is despite the fact that there is still no report of the transfer of phyllosoms via ballast water.
3.4. The Pronghorn Spiny Lobster
The Pronghorn Spiny Lobster, Panulirus penicillatus (Olivier, 1791) is the largest spiny lobster in the Northern Red Sea. Its maximum body length is about 40 cm or more, the average length of an adult lobster is about 30 cm, and males are larger than females [1]. Etymology: The genus name Panulirus is likely derived from the Greek "pan" (all, whole) and "oura" (tail), possibly describing the lobster's large, fan-like tail. It may also be an anagram (a word formed by rearranging the letters of a different word) of the lobster genus Palinurus [28]. The species name, penicillatus, is derived from the Latin "penicillus," meaning "paintbrush" or "pencil [28]", likely referring to the brush-like tufts of setae (bristles) found on its legs and body (Figure 15]. FAO names: Pronghorn spiny lobster (English), Langouste fourchette (French), Langosta horquilla (Spanish) [1]. Other common /vernacular names are: Common spiny lobster, Red lobster, Golden lobster, Rock lobster, Double spined lobster, Fourspine rock lobster, Coral cray, Socorro spiny lobster, and “Jirad albahr hasir” (Arabic) in Saudi Arabia [89] and “Mekhoshtan Kotsani” in Hebrew [90] (literally: has spiny tentacles).
The body of this lobster is elongated, with the second pair of antennae extending beyond the length of the body. It is very spiny and lacks banding [1]. The legs (pereiopods) do not have pincers, and the front of the cephalothorax features large spines that protrude above the eyes. P. penicillatus displays striking coloration (Figure 15). Its body color varies widely, ranging from yellowish-green and greenish brown to dark rusty-brown or bluish black. The head, thorax, and abdomen are mottled with tiny whitish spots, with two fairly large whitish spots located on the sides of the first abdominal segment. The abdominal appendages (pleopods) are dark with light margins, while the legs display longitudinal yellowish-white lines or stripes set against a darker greenish or reddish background. The tail fan at the end of the abdomen is very wide, predominantly bright blue or greenish-gray, and edged with vivid orange.
This species has an extremely wide circumtropical distribution, ranging from the Red Sea and the western Indian Ocean to the eastern Pacific Ocean [1]. Despite its broad range, biological knowledge about this species remains limited compared to other spiny lobsters.
During the day, it hides in rock crevices and beneath the outer edges of coral reefs, typically in fixed locations, becoming primarily active at night. It inhabits shallow waters between 1 and 16 meters deep, most commonly at depths less than 4 meters, though it has occasionally been observed in deeper waters (e.g., [91]). The pronghorn spiny lobster favors rocky substrates in clear waters, away from river mouths, often in areas with wave breaks, as well as on arid beaches and small islands [92].
It usually shelters alone, but gregarious behavior has been observed, with groups often exceeding 20 individuals [93], a tendency also noted in some other spiny lobster species (e.g., [94]).
3.4.1. Reproduction and Development
The sex ratio did not differ significantly from 1:1 [92,93]. Females reach sexual maturity in the wild at a cephalothorax length (CL) of 50 mm. The breeding season in the GoA spans from February to October, during which females lay 2–4 clutches. The number of eggs is directly proportional to cephalothorax length. The incubation period for the eggs is approximately 35 days at temperatures between 24 and 27 ºC [92]. Phyllosoma larvae of this species have been successfully reared in aquaria from hatching until they metamorphosed into active pueruli, which can swim freely and settle on substrates when near suitable shallow habitats. In the laboratory, completing this life cycle required 8–10 months and involved 20 molting stages [95]. In terms of size distribution and size-specific fecundity, the Saudi Red Sea population closely resembles those in the GoA and the Philippines but differs markedly from populations in Palau and Enewetak [96].
3.4.2. Food and Anti-Predatory Adaptations
The pronghorn spiny lobsters are generalist feeders, primarily consuming benthic invertebrates such as mollusks, crustaceans, and sea urchins, whose hard parts they break with their strong upper jaws. Their diet may also include polychaetes, brittle stars, and fish remains. In some regions, they also feed on algae. Lobsters inhabiting carbonate reefs tend to consume more mollusks than those from granite reefs, and larger lobsters likely feed closer to the coast or rely more on detritus feeders than smaller individuals [97].
There are no specific reports on their predators. However, based on known predators of other lobster species, large fish such as the orange-lined triggerfish, Balistapus undulatus, and groupers may prey on adult P. penicillatus in the northern Red Sea. The lobsters’ diurnal sheltering and nocturnal activity serve as anti-predator adaptations against daytime predators. P. penicillatus has a notably large head equipped with considerable defensive mechanisms in the wild [98]. It is protected by numerous spines on its shell and by long, spiny antennae that may be used to fend off predators. The seaward reef margin preferred by this species may also present a lower risk of predation. Additionally, the robust pereiopods characteristic of P. penicillatus enable it to forage effectively in strong surge conditions, likely providing a selective advantage that allowed this species to colonize and thrive in this habitat, which may serve as a refuge from predators [97].
3.4.3. Threats, Environmental Impacts and Conservation
P. penicillatus is fished throughout its wide range, primarily caught by hand or spear gun during daytime skin and SCUBA diving, or using torchlight near the surface at night. Traps have proven ineffective for this species, whereas trammel nets tend to yield better results [1]. Overexploitation has caused local declines in some areas, including the Northern Red Sea. Despite this, the species is classified as "Least Concern" on the IUCN Red List of Threatened Species due to its broad distribution. A 2009 assessment concluded that fishing did not significantly impact on the global population [99]. However, since this assessment is now almost 16 years old, current fishing levels should be re-monitored to detect any increases in exploitation and to provide an updated measure of the species’ abundance. P. penicillatus is protected in Israeli waters of the GoA, with particularly effective protection within nature reserves MPAs) such as the Eilat Coral Nature Reserve (e.g., [100]), the Aqaba MPA in Jordan [101], and Egyptian marine reserves in the GoA, including Abu Galum Managed Resources Protected Area, Nabq Managed Resources Protected Area, and Ras Mohammed National Park [60]. Several studies have documented positive effects of MPAs on lobsters, including increased abundance, density, biomass, catch-per-unit-effort (CPUE), and size [102]. However, this trend was not observed for P. penicillatus within the Galapagos Marine Reserve, where populations declined, likely due to weak enforcement and illegal fishing [103]. No significant changes in lobster size or size-frequency distribution were detected after more than 11 years of protection. Scientists suggest that poaching inside MPAs is a major factor limiting the recovery of lobster populations in this region.
4. Discussion and Conclusions
Despite the oligotrophic nature of the Southeastern Levantine Sea and the Northern Red Sea, six species (five slipper lobsters and one spiny lobster) maintain permanent reproductive populations in the study area, along with single reports of four additional sporadic lobster species. Biological knowledge of the permanent lobster species in this region has expanded in recent years, particularly for the larger, commercially valuable species (S. latus, S. tridacnophaga, and P. penicillatus). However, even for the adult stages of these species, there remain significant gaps in understanding their genetics, life history, reproduction, behavior, sensory biology, diet and feeding habits, predators and anti-predator adaptations, movement, physiology, growth, distribution, fisheries, and conservation status. This lack of information is even more pronounced for the post-embryonic oceanic stages, the naupliosoma in some, perhaps all species, and the free-living phyllosoma larvae, whose larval period can range from a few weeks to over nine months depending on the species. Data on the nektonic niso postlarvae, which settle and initiate the benthic juvenile phase, are rare. Biological knowledge of the smaller and often cryptic species (S. arctus, S. pygmaeus, and E. lewinsohni) is even more limited.
To bridge this knowledge gap, focused studies employing modern technologies, such as underwater video cameras, Remotely Operated Vehicles (ROVs), acoustic tagging, and DNA barcoding, are needed both in coastal waters and in the deep, open sea. However, because lobsters are widely dispersed across time and space, it is challenging for a limited number of scientists to systematically collect data across such vast marine and coastal areas. Therefore, the scientific community must engage the general public active along coasts and at sea—professional and sport fishermen, skin and SCUBA divers, and beachgoers—through Citizen Science initiatives (e.g., [66,104]). Since lobster distributions often cross international borders, international Citizen Science surveys are necessary whenever possible. The findings from the few existing Citizen Science studies on lobsters emphasize the importance of integrated approaches to monitoring cryptic, sometimes low-density species like lobsters.
Notwithstanding four remote records of different lobster species from other biogeographic regions, mainly the Indo-Pacific, there is currently no evidence of established reproductive populations of these species in the new areas. Many Lessepsian crustaceans have formed viable, permanent reproductive populations in the Mediterranean, where crustacean NIS are the second-most abundant invertebrate group, comprising more than a hundred species belonging to the order Decapoda [105,106]. Some of these species contribute positively to commercial fisheries, while others outcompete native commercial crustaceans. However, no lobster NIS has established a permanent reproductive population in the study area. It has been suggested [107] that the passage of lobster sensitive propagules via ship ballast water is unlikely due to the long and complex life cycle of these crustaceans. Additionally, it is doubtful whether the delicate planktonic early stages of lobsters can survive and complete their development under the environmental conditions of the destination regions.
However, Lessepsian migration may impact Mediterranean lobsters indirectly through changes in their food sources and predators. A recent study reported a significant collapse in marine mollusk populations in the eastern Mediterranean over the past decades [108]. The authors attributed this decline primarily to seawater warming, which has rendered conditions unsuitable for native mollusk populations. They described this event as the largest climate-driven, regional-scale biodiversity loss documented in the oceans to date. Furthermore, they predict that as climate warming continues, this native biodiversity collapse will worsen and spread geographically, with only Lessepsian species entering through the Suez Canal potentially mitigating the decline. Restoration to historical baselines, they argue, is unlikely. How will such a dramatic shift in mollusk populations affect lobsters, which heavily rely on mollusks, especially bivalves, as a food source? Lessepsian predators may also influence Mediterranean lobsters. For example, the lionfish, Pterois miles, was first recorded in the eastern Mediterranean in 2012. Since then, its population density has increased dramatically, making it extremely common in the coastal waters of the Levant, from very shallow areas to depths greater than 100 m. P. miles has a high reproductive potential: individuals reach maturity within a year, and adults are capable of spawning year-round, with peak spawning during summer. Lionfish in the Levant grow faster and larger than in their native range, and females outnumber males. As generalist predators, lionfish consume a wide range of teleost and crustacean prey, including species of high economic value [109]. Given their effectiveness in preying on benthic fish and invertebrates, they are assumed to prey on juvenile S. latus and probably also on adults of smaller scyllarid species. For instance, a 17 mm juvenile Spanish slipper lobster (Scyllarides aequinoctialis) was found in the gut contents of a 310 mm invasive red lionfish (Pterois volitans) in Belize [110]. Additionally, the highest attack rates and shortest handling times by adult P. volitans were observed for the Norway lobster (Nephrops norvegicus) [111]. However, this assumption regarding P. miles in the Levant still needs to be confirmed by stomach content analyses.
The most effective “predator” of lobsters, although evolutionarily recent, is Homo sapiens.Lobsters have been targeted by fisheries from prehistoric times through the 21st century [24]. However, modern fishing pressure has sometimes been too intense, leading in some cases to the depletion of local lobster populations to levels from which they cannot recover (e.g., [55]).
One approach to protecting lobster populations primarily involves regulating fishing seasons, limiting lobster size, controlling fishing gear (such as traps and nets), and setting harvesting quotas to promote sustainable practices. In some cases, lobsters have even been declared protected species, making their removal from the sea illegal. However, these regulations are not always effective or enforced, due to the vastness of marine inspection areas relative to the limited number of inspectors/rangers [102], as well as insufficient public education among those active at sea.
Another approach is to establish MPAs or nature reserves. The protection offered by no-take MPAs can lead to significant improvements in four key criteria within their boundaries compared to fished control areas outside the reserves: biomass, density, individual size, and species richness [112]. Several studies have demonstrated positive effects on lobsters within MPAs, including increased abundance, density, biomass, catch-per-unit-effort (CPUE), and individual size [102]. The single quantitative study on the effect of an MPA on S. latus in the southeastern Mediterranean highlighted some of these positive outcomes [52]. These benefits may also extend to lobster fisheries, as MPAs can supply propagules, juveniles, and adults to unprotected areas through the “spillover” process. However, this aspect was not examined in the recent study on the effect of a no-take reserve on S. latus in the southeastern Mediterranean [52] and warrants further investigation. Some reserves fail to show positive effects, potentially due to factors such as MPAs being too small relative to the home ranges of lobster species, their location, size, shape, protection of lobster predators, ineffective enforcement against illegal fishing, and “edge effects” (e.g., [103]). Therefore, there is a clear need for more and larger MPAs to protect lobsters (and other important or threatened species) in the study area, alongside stronger enforcement in existing marine reserves. In cases where lobster fishing is allowed, establishing a minimum legal size for the fishery and protecting egg-bearing females should be strictly enforced.
Additional important aspect of lobster conservation in the study area is the destruction of their preferred natural habitats, such as the morphologically complex SKR, which provides natural dens for lobsters [3]. Marine construction activities, such as building ports, can cause significant damage to SKR. For example, during the construction of the new Bay Port in Haifa Bay, a 2.2 km long and up to 19 m deep entrance channel was excavated through SKR to accommodate the newest giant ships [3], resulting in the loss of a considerable portion of natural lobster habitat. Extensive underwater sand dredging associated with port construction may also cause silting. With the influence of wind and currents, this silting can be carried to nearby SKR areas, disrupting the vital filtration processes of bivalves [3], which, as previously mentioned, are an important food source for lobsters. To mitigate the loss of essential natural habitats, environmentally safe and stable ARs can be deployed. These should be designed based on the behavioral ecology of the specific lobster species, considering factors such as the size, number, and configuration of the ARs, as has been successfully implemented in the southeastern Mediterranean [113] and the GoA [114]. The best practice would be, rather than crushing natural rock (kurkar or coral rocks) and pumping out the fragments, to cut large pieces of natural rock and use this durable material to build the ARs.
The lobster species discussed in this review were previously classified by the IUCN Red List as either “Data Deficient” or “Least Concern” [55,70,76,84,86,99]. However, these assessments were made at least 15–16 years ago and are due for updating. Given the environmental changes and new data accumulated since then, it is timely to re-evaluate these IUCN statuses.
Finally, unfortunately, in recent years, parts of the study area have been affected by war activities. Missiles, detonated drones, and other explosive devices have landed in coastal waters and exploded. What impact have these events had on lobsters, their habitats, and fisheries? No direct studies have specifically examined this subject in lobsters. However, research on other marine animals (e.g., [115]) indicates that underwater blasts can cause significant harm, ranging from direct physical injury to behavioral disturbances and ecosystem damage. Explosions generate powerful shock waves that may damage internal organs, rupture cell membranes, and even cause death. Additionally, these blasts can destroy habitats and disrupt animals’ abilities to communicate, find food, and navigate their environment. The severity of the impact depends on factors such as species, size, and proximity to the explosion. Recent studies (e.g., [116]) on the effects of seismic surveys using sub-surface air guns, which create intense sound waves penetrating the substrate, have shown that these sound waves can affect lobster physiology and short-term behavior. For example, Western rock lobsters, Panulirus cygnus, exposed to air guns exhibited short-term behavioral changes related to predator avoidance, and fewer tagged lobsters were recaptured by commercial fishers.
On the other hand, civilian anthropogenic pressure from commercial and sport fishermen, divers, and others, on lobsters is considerably reduced during wartime due to restrictions on non-military activities at sea (e.g., [117]). However, this reduction in civilian pressure is not expected to offset the significant damage caused by underwater warfare explosions on lobster populations.
Author Contributions
Conceptualization.; writing, review and editing, E.S.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author gratefully acknowledges the photographic contributions of Amit Zeevi, Stephen Breitstein, Jonathan Gottlieb, Dov Zingerman, Luis Sánchez Tocino, Prof. Tamar Guy-Haim, Prof. Clive Jones, Erik Friedmann, Michael Tuval, Ziv Regev, Dr. Gil Koplovitz, Bader Levi, and Dr. Dani Barchana, as well as the illustrations by Rachel Pollak. Special thanks are extended to Prof. Dov Zviely for assistance in designing the map of the study area, and to Eran Shushu Spanier for his support in designing the images and preparing the graphical abstract. The author also thanks Gideon Brand and Lior Ben Ari for their help in translating the Arabic names of lobsters into English.
Conflicts of Interest
The author declares no conflict of interest.
References
- Holthuis, L. B. Marine lobsters of the world. FAO fisheries synopsis, /: Italy, 1991. https, 1991. [Google Scholar]
- Lavalli, K. L.; Spanier, E. Infraorder Palinura Latreille, 1802. Chapter 68. In The Crustacea, Traite de Zoologie 9A – Decapoda; Forest, J., von Vaupel Klein, J.C., Eds, *!!! REPLACE !!!*, Eds.; Koninklijke Brill: Leiden, Holland, 2010; pp. 426–532. [Google Scholar] [CrossRef]
- Spanier, E. , Zviely, D. Key environmental impacts along the Mediterranean coast of Israel in the last 100 years. J. Mar. Sci. Eng. 2022, 11. [Google Scholar] [CrossRef]
- Spanier, E; Galil, B. S. Lessepsian migrationִ a continuous biogeographical process. Endeavour 1991, 15, 102–106. [Google Scholar] [CrossRef]
- Galil, B.S.; Mienis, H.K.; Hoffman, R.; Goren, M. Non-indigenous species along the Israeli Mediterranean coast: tally, policy, outlook. Hydrobiologia 2021, 848, 2011–2029. [Google Scholar] [CrossRef]
- Galil, B.S. A Sea, a Canal, a Disaster: The Suez Canal and the Transformation of the Mediterranean Biota. In The Suez Canal: Past Lessons and Future Challenges. Palgrave Studies in Maritime Politics and Security. Lutmar, C.; Rubinovitz, Z., Eds, *!!! REPLACE !!!*, Eds.; Palgrave Macmilla: Cham, Switzerland, 2023; pp. 199–215. [Google Scholar] [CrossRef]
- Zittis, G.; Almazroui, M.; Alpert, P.; Ciais, P.; Cramer, W.; Dahdal, Y. . Lelieveld, J. Climate change and weather extremes in the Eastern Mediterranean and Middle East. Rev. Geophys. 2022, 60, e2021RG000762–1. [Google Scholar] [CrossRef]
- Pastor, F.; Valiente, J. A.; Khodayar, S. A warming Mediterranean: 38 years of increasing sea surface temperature. Remote Sen. 2020, 12, 2687. [Google Scholar] [CrossRef]
- Khalil, M.T.; Mostafa, A.B.; El-Naggar, M.M. Climate Change and Lessepsian Migration to the Mediterranean Sea. In Khalil, M.T., Emam, W.W.M., Negm, A., Eds; Climate Changes Impacts on Aquatic Environment. Springer Nature: London, UK, 2025; pp. 85–118. [Google Scholar] [CrossRef]
- Eytam, Y.; Ben-Avraham, Z. Morphology and sediments of the inner shelf off northern Israel. Isr. J. Earth Sci. 1992, 41, 27–44. [Google Scholar]
- Almagor, G.; Gill. D.; Perath, I. Marine sand resources offshore Israel. Mar. Georesources Geotech. 2000, 18, 1–42. [Google Scholar] [CrossRef]
- Goldsmith, V.; Golik, A. Sediment transport model of the southeastern Mediterranean coast. Mar. Geol. 1980, 37, 135–147. [Google Scholar] [CrossRef]
- Nir, Y. Offshore artificial structures and their influence on the Israel and Sinai Mediterranean beaches. Coast. Eng. 1982, 1982, 1837–1856. [Google Scholar]
- Emery, K.O.; Neev, D. Mediterranean beaches of Israel. Israel Geol. Surv. Bull. 1960, 26, 1–24. [Google Scholar]
- Spanier, E.; Lavalli, K.L. Natural history of Scyllarides latus (Crustacea Decapoda): a review of the contemporary biological knowledge of the Mediterranean slipper lobster. J. Nat. His. 1998, 32, 1769–1786. [Google Scholar] [CrossRef]
- Spanier, E. 14. In Lobsters: Biology Management, Aquaculture and Fisheries Phillips, B. F. Ed; Part 2: Lobsters of Commercial Importance, Blackwell: Oxford, UK, 2006; pp. 462–496. [Google Scholar] [CrossRef]
- Schütz, K. I. Structure and stratigraphy of the Gulf of Suez, Egypt. In Interior rift basins; Landon, S. M. Ed; AAPG Mem. 1994, 59, 57–96. [Google Scholar] [CrossRef]
- Ben-Avraham, Z. Structural framework of the gulf of Elat (Aqaba), northern Red Sea. J. Geophys. Res. Solid Earth 1985, 90, 703–726. [Google Scholar] [CrossRef]
- Fine, M. ; Gildor, H;, Genin, A. A coral reef refuge in the Red Sea. Glob. Chang. Biol. 2013, 19, 3640–3647. [Google Scholar] [CrossRef]
- Sengupta, S.; Gildor, H.; Ashkenazy, Y. Depth-dependent warming of the Gulf of Eilat (Aqaba). Clim. Change 2024, 177, 107. [Google Scholar] [CrossRef]
- Essa, B.; Geneid, Y. Temporal change of seagrass meadows: estimating species composition, coverage and carbon stock trends over 16 years in NIOF studied area, Hurghada, Egypt. Egypt. J. Aquatic Biol. Fish. 2025, 29, 1361–1381. [Google Scholar] [CrossRef]
- BadrElDin, A. M.; Hallock, P. Benthic foraminifers in coastal habitats of Ras Mohamed Nature Reserve, southern Sinai, Red Sea, Egypt. J. f. Micropaleontology. 2024, 43, 239–267. [Google Scholar] [CrossRef]
- Belmonte, G. Elements of Pelagos Biology: With Focus on the Mediterranean Sea., Springer Nature: Cham., Switzerland, 2025, 1-118 pp. [CrossRef]
- Spanier, E.; Lavalli, K.L.; Goldstein, J. S.; Groeneveld, J.C.; Jordaan G., L.; Jones, C.M.; Phillips, B. F.; Bianchini, M. L.; Kibler, R. D.; Díaz, D.; Mallol, S.; Goñi, R.; van der Meeren, G. I. ; Agnalt, A-L.; Behringer, D. C.; Keegan, W. F.; Jeffs A. A concise review of lobster utilization by worldwide human populations from pre-history to the modern era. ICES J. Mar. Sci. [CrossRef]
- Spanier, E. The utilization of lobsters by humans in the Mediterranean basin from the Prehistoric Era to the Modern Era – an interdisciplinary short review. AJMS 2015, 1, 223–234. [Google Scholar] [CrossRef]
- Glenister, C. L. Profiling Punt: using trade relations to locate “God’s Land”. Doctoral dissertation, University of Stellenbosch, South Africa, 2008.
- Danelius, E.; Steinitz, H. The fishes and other aquatic animals on the Punt-reliefs at Deir el-Bahri. JEA 1967, 53, 15–24. [Google Scholar]
- Jaeger, E. C. A source-book of biological names and terms; ar">CABI Charles C. Thomas: Springfield, Illinois, US,, Third Edition, Fourth Printing, 1966. [Google Scholar]
- Ingle, R. Crayfishes, lobsters and crabs of Europe: an illustrated guide to common and traded species. Springer Science & Business Media, BV, 2012. [CrossRef]
- Common names of Scyllarides latus. https://www.sealifebase.se/comnames/CommonNamesList.php?ID=26141&GenusName=Scyllarides&SpeciesName=latus&StockCode=386. (accessed on 24 June 2025).
- Spanier, E. The Mediterranean Slipper Lobster - the known and the concealed in the ecology of a threatened species. Ecol. Environ. 2023, 14, 1–15. [Google Scholar]
- Lavalli, K. L.; Spanier, E.; Goldstein, J. S. Scyllarid Lobster Biology and Ecology, Chapter 3. In Crustacea Diarte-Plata, G.; Escamilla-Montes, R., Eds, *!!! REPLACE !!!*, Eds.; IntechOpen: London, UK, 2019; pp. 25–51. [Google Scholar] [CrossRef]
- Spanier, E.; Lavalli, K. L. Commercial Scyllarids. Chapter 13 In Lobsters: biology, management, aquaculture and fisheries. Phillips, B. F., Ed; Blackwell: Oxford, UK, 2013; pp. 414–467. [Google Scholar] [CrossRef]
- Spanier, E.; Tom, M.; Pisanty, S.; Almog, G. Seasonality and shelter selection by the slipper lobster Scyllarides latus in the southeastern Mediterranean. Mar. Ecol. Prog. Ser. 1988, 42, 247–255. [Google Scholar] [CrossRef]
- Pessani, D.; Mura, M. The Biology of the Mediterranean Scyllarids. In The Biology and Fisheries of the Slipper Lobsters Lavalli, K. L.; Spanier, E. Eds, Ed.; CRC Press, Taylor & Francis Group: London, UK, 2007. [Google Scholar]
- Sekiguchi, H.; Booth, D.; Webber, W. R. Early life histories of slipper lobsters. In The Biology and Fisheries of the Slipper Lobsters Lavalli, K. L., Spanier, E. Eds; CRC Press, Taylor & Francis Group: London, UK, 2007; pp. 9–90. [Google Scholar]
- Martins, H. R. Biological studies on the exploited stock of the Mediterranean locust lobster, Scyllarides latus, (Latrielle, 1803) (Decapoda, Scyllaridae) in the Azores. J. Crustac. Biol. 1985, 5, 294–305. [Google Scholar] [CrossRef]
- Martins, H. R. Some observations on the naupliosoma and phyllosoma larvae of the Mediterranean locust lobster, Scyllarides latus, (Latrielle, 1803) from the Azores. 1985b Intl. Council for the Exploration of the Sea C.M.K. 52 (13) Shellfish Committee.
- Spanier, E.; Lavalli, K. L. First record of an early benthic juvenile likely to be that of the Mediterranean slipper lobster, Scyllarides latus (Latreille, 1802) Crustaceana 2013, 86, 259-267. 86. [CrossRef]
- Spanier, E. Mollusca as food for the slipper lobster Scyllarides latus in the coastal waters of Israel. Levantina 1987, 68, 713–716. [Google Scholar]
- Lavalli, K.L.; Spanier, E.; Grasso, F. Behavior and sensory biology of slipper lobsters. In The Biology and Fisheries of the Slipper Lobsters Lavalli, K. L. Spanier, E. Eds; CRC Press, Taylor & Francis Group: London, UK, 2007. [Google Scholar]
- Spanier, E.; Almog-Shtayer, G. Shelter preferences in the Mediterranean slipper lobster: effects of physical properties. J. Experiment. Mar. Biol. Ecol. 1992, 164, 103–116. [Google Scholar] [CrossRef]
- Barshaw, D. E.; Lavalli, K. L.; Spanier, E. Offense versus defense: responses of three different morphological types of lobsters to predation. Mar. Ecol. Prog. Ser. 2003, 256, 171–182. [Google Scholar] [CrossRef]
- Tarsitano, S. F.; Lavalli, K. L.; Horne, F.; Spanier, E. The constructional properties of the exoskeleton of Homarid, Palinurid, and Scyllarid Lobsters. Hydrobiologia 2006, 557, 9–20. [Google Scholar] [CrossRef]
- Barshaw, D.E.; Spanier, E. Anti-predator behaviors of the Mediterranean slipper lobster Scyllarides latus. Bull. Mar. Sci. 1994, 55, 375–382. [Google Scholar]
- Spanier, E.; Weihs, D.; Almog-Shtayer, G. Swimming of the Mediterranean Slipper lobster J. Experiment. Mar. Biol. Ecol. 1991, 145, 15–31. [Google Scholar] [CrossRef]
- Spanier, E.; Weihs, D. Why do shovel-nosed (slipper) lobsters have shovels? The Lobster Newsletter 1992, 5, 8–9. [Google Scholar]
- Lavalli, K. L.; Spanier, E. Predator adaptations of decapods, Chapter 8, In The Natural History of the Crustacea, Vol. 2: Lifestyles and Feeding Biology Thiel, M.; Watling, L, Eds, Ed.; Oxford University Press: NY, USA, 2015; pp. 190–228. [Google Scholar]
- Lavalli, K. L.; Spanier, E. Does gregariousness function as an antipredator mechanism in the Mediterranean slipper lobster, Scyllarides latus? Mar. Freshw. Res. 2001, 52, 1133–1143. [Google Scholar] [CrossRef]
- Weisbaum, D.; Lavalli, K. L. Morphology and distribution of antennular setae of scyllarid lobsters (Scyllarides aequinoctialis, S. latus, and S. nodifer) with comments on their possible function. Invert. Biol. 2004, 123, 324–342. [Google Scholar] [CrossRef]
- Lau, S. T. F.; Spanier, E.; Meyer-Rochow, V. B. Anatomy and ultrastructural organisation of the eye of the Mediterranean slipper lobster, Scyllarides latus: preliminary results. N. Z. J. Mar. Freshwater Res. 2009, 43, 233–245. [Google Scholar] [CrossRef]
- Miller, E.; Spanier, E.; Diamant, R.; Yahel, R.; Spanier, E. ; Diamant, R.; Yahel R. Nature reserves facilitate the conservation of the Mediterranean slipper lobster, Scyllarides latus. Mar. Ecol. Prog. Ser. 2023, 714, 57–69. [Google Scholar] [CrossRef]
- Boles, L.; Lohmann, K. True navigation and magnetic maps in spiny lobsters. Nature 2003, 421, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Spanier, E.; Lavalli, K. L. Slipper lobster fisheries—Present status and future perspectives In The Biology and Fisheries of the Slipper Lobsters Lavalli, K. L.; Spanier, E. Eds, Ed.; CRC Press, Taylor & Francis Group: London, UK, 2007; pp. 377–391. [Google Scholar] [CrossRef]
- Butler, M. , MacDiarmid, A. & Cockcroft A. Scyllarides latus, /: List of Threatened Species, 2011. . https; (accessed on 2 July 2025)2011. [Google Scholar]
- Bianchini, M. L.; Spanier, E.; Ragonese, S. Enzymatic variability of Mediterranean slipper lobsters, Scyllarides latus, from Sicilian waters. Ann. Ser. Hist. Nat. 2003, 13, 43–50. [Google Scholar]
- Faria, J.; Froufe, E.; Tuya, F.; Alexandrino, P.; Pérez-Losada, M. Panmixia in the endangered slipper lobster Scyllarides latus from the Northeastern Atlantic and Western Mediterranean. J. Crustac. Biol. 2013, 33, 557–566. [Google Scholar] [CrossRef]
- Nehmeh, B.; Haydous, F.; Ali, H.; Hdaifi, A.; Abdlwahab, B.; Orm, M. B.; Abrahamian, Z.; Akoury, E. Emerging contaminants in the Mediterranean Sea endangering Lebanon's Palm Islands Natural Reserve. RSC adv., 2025, 15, 2034–2044. [Google Scholar] [CrossRef]
- Saad, R.; Gérard, J. A.; Gérard, P. Impacts of local Community perception and participation on environmental issues in integrated coastal zone Management: a case study of Tyre, southern Lebanon. IJHSS, 2024, 11, 1–15. [Google Scholar] [CrossRef]
- Samy-Kamal, M.; Sánchez-Lizaso, J. L.; Forcada, A. (2011). Status of marine protected areas in Egypt. Anim. Biodivers. Conser. 2011, 34, 165–177. [Google Scholar] [CrossRef]
- Spanier, E.; Lavalli, K.L.; Edelist, D. Artificial reefs for lobsters – an overview of their application for fisheries enhancement, management, and conservation In Chapter 6 of Artificial Reefs in Fisheries Management Bortone, S.; Brandini, F. P., Fabi, G., Otake, S., Eds, *!!! REPLACE !!!*, Eds.; CRC Press, Taylor & Francis Group: NY, USA, 2011; pp. 77–109. [Google Scholar]
- Shiber, J. G. Trace metals in edible marine molluscs and crustaceans from Lebanon. Hydrobiologia 1981, 83, 465–476. [Google Scholar] [CrossRef]
- Spanò, S.; Ragonese, S.; Bianchini, M. An anomalous specimen of Scyllarides latus (Decapoda, Scyllaridae). Crustaceana 2003, 76, 885–889. [Google Scholar] [CrossRef]
- Goldstein, J. S.; Spanier, E.; Spanier, E. Increasing seawater temperatures in the Levantine basin, eastern Mediterranean may elicit increased activity in slipper lobsters, Scyllarides latus (Latreille, 1803) (Decapoda: Achelata: Scyllaridae). J. Crustac. Biol. 2023, 42, 1–8. [Google Scholar] [CrossRef]
- Spanier, E.; Miller, E. ; Zviely. D. Winter stranding of Mediterranean slipper lobsters, Scyllarides latus. Reg. Stud. Mar. Sci. 2017, 14, 126-131. 14,. [CrossRef]
- Gennaro, G. Ampliamento delle conoscenze sulla distribuzione e sullo stato di conservazione della specie protetta Scyllarides latus nell'Area Marina Protetta" Isola di Bergeggi" e nella ZSC" Fondali Noli-Bergeggi".. Master Thesis, Universitá Delgi Studi Di Genova, Italy, 2025. [Google Scholar]
- Holthuis L., B. The Indo-Pacific scyllarine lobsters (Crustacea, Decapoda, Scyllaridae). Zoosystema 2002, 24(3), 499–683. [Google Scholar]
- Alborés, I.; García-Soler, C.; Fernández, L. Reproductive biology of the slipper lobster Scyllarus arctus in Galicia (NW Spain): Implications for fisheries management. Fish. Res. 2019, 212, 1–11. [Google Scholar] [CrossRef]
- Fiedler, U.; Spanier, E. Occurrence of larvae of Scyllarus arctus (Crustacea, Decapoda, Scyllaridae) in the Eastern Mediterranean — preliminary results. Ann. Ser. Hist. Nat. 1999, 17, 153–158. [Google Scholar]
- Butler, M.; MacDiarmid, A.; Wahle, R.; Cockcroft, A.; Chan, T.Y. (2011). Scyllarus arctus, /: List of Threatened Species. 2009 . https; (accessed on 5 July 2025)2009. [Google Scholar]
- Majdalani, S. The present status of fishery and information system in Lebanon. Enabling participation in the fishery statistics and information system in the Mediterranean (TCP/INT/2904).
- Fishelson, L. Lexicon of the Flora and Fauna of the Land of Israel, Ministry of Defense Publishing House: Raanana, Israel, 1996; Volume 3 - Aquatic Life, (in Hebrew), pp. 183-164.
- Lewinsohn, C. The occurrence of Scyllarus pygmaeus (Bate) in the eastern Mediterranean (Deacpoda, Scyllaridae) Crustaceana 1974, 27, 43–46. 27. [CrossRef]
- Lewinsohn, C.; Holthuis, L.B. The Crustacea Decapoda of Cyprus. Zool. Verh. 1986, 230, 1–64. [Google Scholar]
- Guy-Haim, T.; Iakoleva, A.; Ermak, K.; Spanier, E.; Morov, A. R. First record of the phyllosoma larva of the pygmy locust lobster Scyllarus pygmaeus in the Eastern Mediterranean Sea J. Mar. Biol. Assoc. U.K., 2024, 104, e49, 1-4. , 104. [CrossRef]
- Wahle, R.; MacDiarmid, A.; Cockcroft, A.; Chan, T.Y.; Butler, M. Scyllarus pygmaeus, The IUCN Red List of Threatened Species. 2009. . https://www.iucnredlist.org/search? (accessed on 5 July 2025).
- Galil, B.; Pisanty, S.; Spanier, E.; Tom, M. The Indo-Pacific lobster Panulirus ornatus (Fabricius, 1798) (Crustacea: Decapoda) a new Lessepsian migrant to the Eastern Mediterranean Isr. J. Zool. 1989, 35, 241–243. [Google Scholar]
- Spanier, E.; Friedmann, E. The collection of an exuvia identified as Panulirus longipes longipes (A. Milne-Edwards, 1868) from off Haifa, Israel. Mediterr. Mar. Sci., 2019, 20. [Google Scholar] [CrossRef]
- Spanier, E. First record of the American clawed lobster, Homarus americanus (H. Milne Edwards, 1837), in the southeastern Mediterranean. Bioinvations Rec. 2023, 12, 775–786. [Google Scholar] [CrossRef]
- Holthuis, L.B. Some new species of Scyllaridae. Proc. K. Ned. Akad. Wet. 1967, C 70, 305–308. [Google Scholar]
- Holthuis, L. The second Israel South Red Sea expedition, 1965: Report no. 7. The Palinuridae and Scyllaridae of the Red Sea. Zool. Meded. 1968, 42, 281–301. [Google Scholar]
- Kizhakudan, J. K.; Sundar, R.; Mohamed, A.A.; Anassery, A.A.; Thiagu, R. , Midhun, M. Growth, maturation and breeding of the clamkiller slipper lobster Scyllarides tridacnophaga Holthuis, 1967 in captivity. Lobster Newsletter, 2018, 31, 8–11. [Google Scholar]
- Herrnkind, W. Queuing behavior of spiny lobsters. Science 1969, 164, 1425-1427.–content. [Google Scholar] [CrossRef]
- MacDiarmid, A.; Cockcroft, A.; Butler, M. Scyllarides tridacnophaga, The IUCN Red List of Threatened Species, 2011. https://www.iucnredlist.org/search?query=Scyllarides%20tridacnophaga&searchType=species. (accessed on 8 July 2025).
- Holthuis, L. B. The Indo-Pacific scyllarine lobsters (Crustacea, Decapoda, Scyllaridae). Zoosystema 2002, 24, 499–683. [Google Scholar]
- Chan, T.Y. Eduarctus lewinsohni, The IUCN Red List of Threatened Species, 2009. https://www.iucnredlist.org/search?
- Guy-Haim, V.; Farstey, T.; Iakovleva, A.; Spanier, E.; Morov, A.R. First report of the pygmy slipper lobster Biarctus sordidus (Crustacea, Decapoda) in the Red Sea following he finding of its phyllosoma larva. J. Mar. Biol. Assoc. U. K. 2024, 104, e110–1. [Google Scholar] [CrossRef]
- Yang, C. H.; Lin, C. W.; Chan, T. Y. Additional slipper lobsters of the subfamily Scyllarinae Latreille, 1825 (Crustacea, Achelata, Scyllaridae) from Taiwan. Zootaxa, /: 336-346. https, 2636. [Google Scholar]
- Sealife Base, version o4/2025. https://www.sealifebase.se/comnames/CommonNamesList.php?ID=9316&GenusName=Panulirus&SpeciesName=penicillatus&StockCode=346. (accessed on 12 July 2025).
- Fishelson, L. Lexicon of the Flora and Fauna of the Land of Israel, Ministry of Defense Publishing House: Raanana, Israel, 1996; Volume 3 - Aquatic Life, (in Hebrew), p. 185.
- Vaitheeswaran, T. On rare occurrence of pronghorn spiny lobster, Panulirus penicillatus (Olivier, 1791) off Tuticorin coast, India (08 35.912’N and 78 25.327’E) (25 M). J. Agric. Sci. Bot. 2017, 1, 7–8. [Google Scholar] [CrossRef]
- Plaut, I. Sexual maturity, reproductive season and fecundity of the spiny lobster Panulirus penicillatus from the Gulf of Eilat (Aqaba), Red Sea. Mar. Freshwater Res. 1993, 44, 527–535. [Google Scholar] [CrossRef]
- Hearn, A.; Murillo, J. C. Life History of the Red Spiny Lobster, Panulirus penicillatus (Decapoda: Palinuridae), in the Galápagos Marine Reserve, Ecuador1. Pac. Sci. 2008, 62, 191–204. [Google Scholar] [CrossRef]
- Zimmer-Faust, R.K.; Spanier, E. Gregariousness and sociality in spiny lobsters: implication for den habitation. J. Exp. Mar. Biol. Ecol. 1987, 105, 51–71. [Google Scholar] [CrossRef]
- Matsuda, H.; Takenouchi, T.; Goldstein, J. S. The complete larval development of the pronghorn spiny lobster Panulirus penicillatus (Decapoda: Palinuridae) in culture. J. Crustac. Biol. 2006, 26, 579–600. [Google Scholar] [CrossRef]
- Hogarth, P. J.; Barratt, L. A. Size distribution, maturity and fecundity of the spiny lobster Panulirus penicillatus (Olivier 1791) in the Red Sea. Trop. Zoo. 1996, 9, 399–408. [Google Scholar] [CrossRef]
- Sardenne, F.; Bodin, N.; Barret, L.; Blamey, L.; Govinden, R.; Gabriel, K. . Rowat, D. Diet of spiny lobsters from Mahé Island reefs, Seychelles inferred by trophic tracers. Reg. Stud. Mar. Sci. 2021, 42, 101640. [Google Scholar] [CrossRef]
- MacDonald, C. D. Fecundity and reproductive rates in Indo-West Pacific spiny lobsters. Micronesica 1988, 21, 103–114. [Google Scholar]
- Cockcroft, A.; MacDiarmid, A.; Butler, M. Panulirus penicillatus. IUCN Red List of Threatened Species, 2009. https://www.iucnredlist.org/search?query=Panulirus%20penicillatus.&searchType=species. (accessed on 14 July 2025).
- Loya, Y. How to influence environmental decision makers? The case of Eilat (Red Sea) coral reefs. J. Exp. Mar. Biol. Ecol. 2007, 344, 35–53. [Google Scholar] [CrossRef]
- Al-Zibdah, M. K. The Aqaba Marine Protected Area--Integration of Marine Science and Resource Management in the Gulf of Aqaba-Red Sea. Int. J. Mar. Sci. 2013, 3, 361–367. [Google Scholar] [CrossRef]
- Spanier, E. Marine protected areas – a review of their potential effects on lobster population size and structure and fisheries management Fish. Res. 2024, 275, 107025. [Google Scholar] [CrossRef]
- Buglass, S.; Reyes, H.; Ramirez-González, J.; Eddy, T. D.; Salinas-de-León, P. Jarrin, J. M. Evaluating the effectiveness of coastal no-take zones of the Galapagos Marine Reserve for the red spiny lobster, Panulirus penicillatus. Mar. Policy 2018, 88, 204–212. [Google Scholar] [CrossRef]
- Kampouris, T. E.; Koutsoubas, D.; kanelopoulou, K.; Zannaki, C. , Batjakas, I. E. Informing the general public on the threat status of the European spiny lobster, Palinurus elephas (Fabricius, 1787) through Citizen-Science and social media platforms. A case study from the Aegean Sea. Mediterr. Mar. Sci. [CrossRef]
- Galil, B.S.; Boero, F.; Campbell, M.L.; Carlton, J.T.; Cook, E.; Fraschetti, S.; Gollasch, S.; Hewitt, C.L.; Jelmert, A.; MacPherson, E.; et al. ‘Double trouble’: The expansion of the Suez Canal and marine bioinvasions in the Mediterranean Sea. Biol. Invasions 2015, 17, 973–976. [Google Scholar] [CrossRef]
- Galil, B. S; Zirler, R.; Feldstein-Farkash, T.; Bronstein, O. A call to ARMS: new records of invasive alien decapods in the Mediterranean Sea. Manag. Biol. Invas. 2024, 15(4), 581–600. [Google Scholar] [CrossRef]
- Spanier, E. Why there are almost no reports on non-Indigenous lobsters? AOMB. [CrossRef]
- Albano, P.G.; Steger, J.; Bošnjak, M.; Dunne, B.; Guifarro, Z.; Turapova, E.; Hua, Q.; Kaufman, D.S.; Rilov, G.; Zuschin, M. Native biodiversity collapse in the eastern Mediterranean. Proc. R. Soc. B Boil. Sci. 2021, 288, 20202469. [Google Scholar] [CrossRef]
- Savva, I.; Chartosia, N.; Antoniou, C.; Kleitou, P.; Georgiou, A.; Stern, N.; Hadjioannou, L.; Jimenez, C. ; Andreou,V.; Hall-Spencer, J. M.; Kletou, D. They are here to stay: the biology and ecology of lionfish (Pterois miles) in the Mediterranean Sea. J. Fish Biol. [CrossRef]
- Mizrahi, M. I.; Chapman, J. K.; Gough, C. L.; Humber, F.; Anderson, L. G. Management implications of the influence of biological variability of invasive lionfish diet in Belize. Management of Biol. Invasions 2017, 8(1), 61-70.–70. [Google Scholar] [CrossRef]
- McCard, M.; McCard, N.; Coughlan, N. E.; South, J.; Kregting, L.; Dick, J. T. Functional response metrics explain and predict high but differing ecological impacts of juvenile and adult lionfish. R. Soc. Open Sci. 2024, 11. [Google Scholar] [CrossRef] [PubMed]
- Lester, S.E.; Halpern, B.S.; Grorud-Colvert, K.; Lubchenco, J.; Ruttenberg, B.I.; Gaines, S. D.; Airam´e, S.; Warner, R.R. Biological effects within no-take marine reserves: a global synthesis. Mar. Ecol. Prog. Ser. 2009, 384, 33–46. [Google Scholar] [CrossRef]
- Spanier, E.; Tom, M.; Pisanty, S.; Almog-Shtayer, G. Artificial reefs in the low productive marine environment of the Southeastern Mediterranean. P.S.Z.N.I: Mar. Ecol. [CrossRef]
- Spanier, E.; Angel, D.L. Benthic artificial reefs and mid-water artificial substrates in the northern Gulf of Aqaba: Biodiversity and potential environmental applications. In Aqaba-Eilat, the Improbable Gulf: Environment, Biodiversity and Preservation; Por, F.D., Ed.; Hebrew University Magnes Press: Jerusalem, Israel, 2008; pp. 281–298. [Google Scholar]
- Jenkins, A. K.; Kotecki, S. E.; Dahl, P. H.; Bowman, V. F.; Casper, B. M.; Boerger, C.; Popper, A. N. Physical effects from underwater explosions on two fish species. In The Effects of Noise on Aquatic Life: Principles and Practical Considerations; Popper, A.N., Sisneros, J.A., Hawkins, A.D., Thomsen, F., Eds.; Springer International Publishing: Cham, Switzerland, 2024; pp. 979–987. [Google Scholar] [CrossRef]
- de Lestang, S.; How, J.; Erbe, C.; Rushworth, K. Boom, shake the room: Seismic surveys affect behaviour and survival of western rock lobster. Fish. Res. 2024, 277, 107072. [Google Scholar] [CrossRef]
- Siluvaithasan, A. S.; Stokke, K. Fisheries under fire: Impacts of war and challenges of reconstruction and development in Jaffna fisheries, Sri Lanka. Nor. Geogr. Tidsskr. 2006, 60(3), 240–248. [Google Scholar] [CrossRef]
Figure 1.
The study area. Background: part of Blue Marble: Land Surface, Shallow Water, and Shaded Topography. NASA Goddard Space Flight Center Image by Reto Stöckli, Robert Simmon and MODIS Groups; https://visibleearth.nasa.gov/images/57752/blue-marble-land-surface-shallow-water-and-shaded-topography(accessed on 11 February 2002). Courtesy of Professor Dov Zviely, Faculty of Marine Sciences, Ruppin Academic Center, Israel.
Figure 1.
The study area. Background: part of Blue Marble: Land Surface, Shallow Water, and Shaded Topography. NASA Goddard Space Flight Center Image by Reto Stöckli, Robert Simmon and MODIS Groups; https://visibleearth.nasa.gov/images/57752/blue-marble-land-surface-shallow-water-and-shaded-topography(accessed on 11 February 2002). Courtesy of Professor Dov Zviely, Faculty of Marine Sciences, Ruppin Academic Center, Israel.
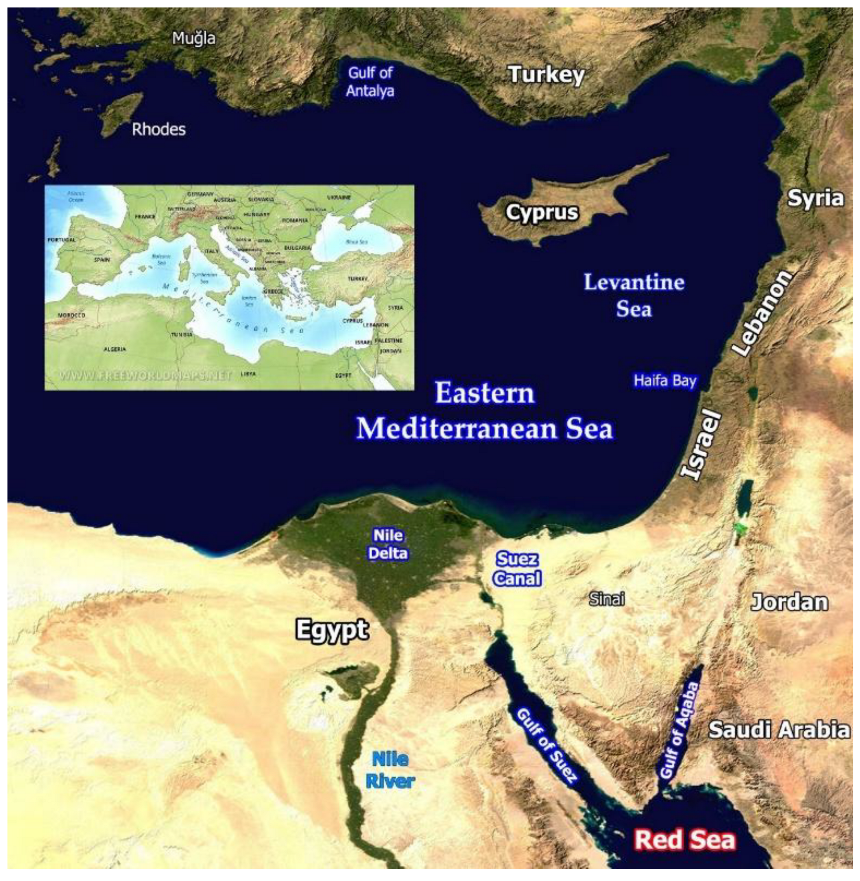
Figure 2.
The Mediterranean slipper lobster, Scyllarides latus (photographed by Amit Zeevi on November 5, 2021, at 11:00 AM at a depth of 5 meters in the Akhziv-Rosh HaNikra Marine Nature Reserve on the northern coast of Israel).
Figure 2.
The Mediterranean slipper lobster, Scyllarides latus (photographed by Amit Zeevi on November 5, 2021, at 11:00 AM at a depth of 5 meters in the Akhziv-Rosh HaNikra Marine Nature Reserve on the northern coast of Israel).

Figure 3.
Left: Female Scyllarides latus with eggs (photographed by S. Breitstein, University of Haifa), Center: Phyllosoma larva of Scyllarides sp. (from the Martin Wiggo Johnson Phyllosoma slide collection of the Scripps Institute of Oceanography, Pelagic Invertebrate Collection website (https://www.flickr.com/photos/siopiczp/collections/72157641176701533/) Right: nisto – young benthic stage of S. latus (photographed by J.J. Gottlieb, University of Haifa).
Figure 3.
Left: Female Scyllarides latus with eggs (photographed by S. Breitstein, University of Haifa), Center: Phyllosoma larva of Scyllarides sp. (from the Martin Wiggo Johnson Phyllosoma slide collection of the Scripps Institute of Oceanography, Pelagic Invertebrate Collection website (https://www.flickr.com/photos/siopiczp/collections/72157641176701533/) Right: nisto – young benthic stage of S. latus (photographed by J.J. Gottlieb, University of Haifa).
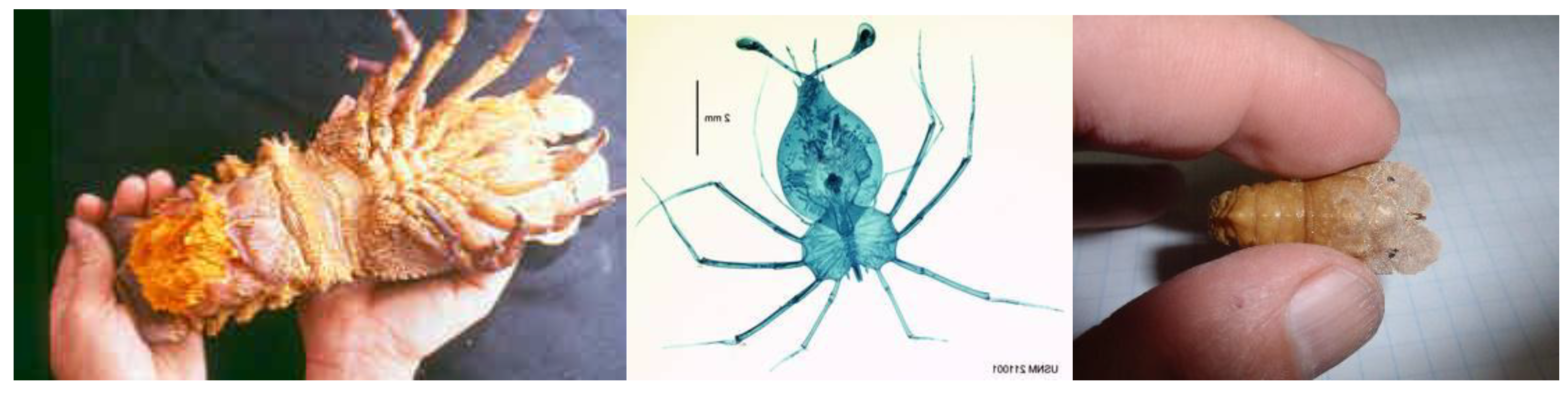
Figure 4.
Demonstration of the Mediterranean slipper lobster's "burst and coast" swimming shown from above (top image) and from the side (middle image). T marks the start of the muscular abdomen movement (from [46] with permission). Bottom right: Illustration of the use of the flattened second antennae for steering while swimming (drawn, from video footage, by Rachel Pollak, University of Haifa).
Figure 4.
Demonstration of the Mediterranean slipper lobster's "burst and coast" swimming shown from above (top image) and from the side (middle image). T marks the start of the muscular abdomen movement (from [46] with permission). Bottom right: Illustration of the use of the flattened second antennae for steering while swimming (drawn, from video footage, by Rachel Pollak, University of Haifa).
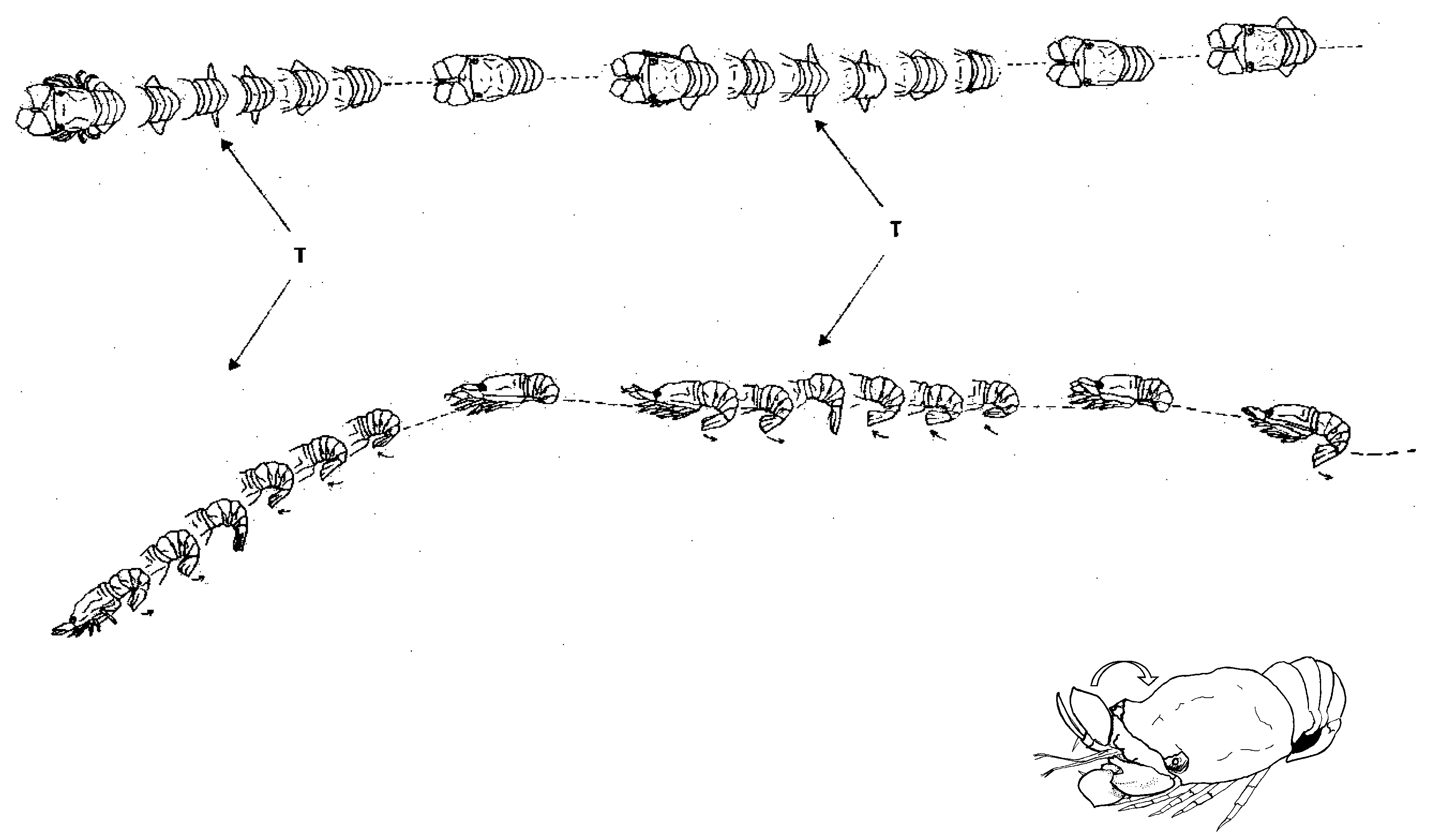
Figure 5.
The Small European locust lobster, Scyllarus arctus, photographed by Dov Zingerman on 27 July 2019 at a depth of 18 m near Vela Luka, Croatia, in the Adriatic Sea.
Figure 5.
The Small European locust lobster, Scyllarus arctus, photographed by Dov Zingerman on 27 July 2019 at a depth of 18 m near Vela Luka, Croatia, in the Adriatic Sea.

Figure 6.
The pygmy locust lobster, Scyllarus pygmaeus. Credit: Luis Sánchez Tocino (https://litoraldegranada.ugr.es), Granada University, Spain.
Figure 6.
The pygmy locust lobster, Scyllarus pygmaeus. Credit: Luis Sánchez Tocino (https://litoraldegranada.ugr.es), Granada University, Spain.

Figure 7.
Dorsal view of a Scyllarus pygmaeus phyllosoma larva (stage VII), collected on 12 December 2022 using a horizontal tow with a Bongo plankton net (mesh size 65–200 μm) at a depth of 0.5–1 m, offshore in the central Israeli Exclusive Economic Zone (bottom depth 1100 m), southeastern Mediterranean Sea. Scale bar: 1 cm. (Photograph by Prof. T. Guy-Haim).
Figure 7.
Dorsal view of a Scyllarus pygmaeus phyllosoma larva (stage VII), collected on 12 December 2022 using a horizontal tow with a Bongo plankton net (mesh size 65–200 μm) at a depth of 0.5–1 m, offshore in the central Israeli Exclusive Economic Zone (bottom depth 1100 m), southeastern Mediterranean Sea. Scale bar: 1 cm. (Photograph by Prof. T. Guy-Haim).

Figure 8.
Ornate spiny lobster, Panulirus ornatus (Credit: Prof. Clive Jones, James Cook University, Australia).
Figure 8.
Ornate spiny lobster, Panulirus ornatus (Credit: Prof. Clive Jones, James Cook University, Australia).

Figure 9.
Exuvia of an adult female long-legged spiny lobster, Panulirus longipes longipes (Photographed by E. Friedmann, Israel).
Figure 9.
Exuvia of an adult female long-legged spiny lobster, Panulirus longipes longipes (Photographed by E. Friedmann, Israel).

Figure 10.
Agonistic interaction between two American clawed lobsters, Homarus americanus (photographed by Michael Tuval in very shallow water, 20–30 cm deep, off the coast of Caesarea, Israel, on 30 December 2022).
Figure 10.
Agonistic interaction between two American clawed lobsters, Homarus americanus (photographed by Michael Tuval in very shallow water, 20–30 cm deep, off the coast of Caesarea, Israel, on 30 December 2022).

Figure 11.
The Clamkiller Slipper Lobster (Scyllarides tridacnophaga) on fern seagrass (Halophila stipulacea). Photographed by Amit Zeevi in Eilat, Gulf of Aqaba, Israel, at 10 m depth on May 14, 2022.
Figure 11.
The Clamkiller Slipper Lobster (Scyllarides tridacnophaga) on fern seagrass (Halophila stipulacea). Photographed by Amit Zeevi in Eilat, Gulf of Aqaba, Israel, at 10 m depth on May 14, 2022.

Figure 12.
A single-file queue of three Clamkiller Slipper Lobsters, Scyllarides tridacnophaga, on a sandy bottom north of the Sun Boat diving site, Eilat, northern Gulf of Aqaba, Israel, at a depth of 8 m on 8 June 2025 (video by Ziv Regev; still image prepared by Eran Shushu Spanier).
Figure 12.
A single-file queue of three Clamkiller Slipper Lobsters, Scyllarides tridacnophaga, on a sandy bottom north of the Sun Boat diving site, Eilat, northern Gulf of Aqaba, Israel, at a depth of 8 m on 8 June 2025 (video by Ziv Regev; still image prepared by Eran Shushu Spanier).

Figure 13.
Lewinsohn locust lobster (Eduarctus lewinsohni), Eilat, northern Gulf of Aqaba, Israel, at approximately 5 m depth on the shallow reef opposite the Interuniversity Institute for Marine Sciences, 9 August 2019. Estimated total length: 3–4 cm. (Photographed by Dr. Gil Koplovitz).
Figure 13.
Lewinsohn locust lobster (Eduarctus lewinsohni), Eilat, northern Gulf of Aqaba, Israel, at approximately 5 m depth on the shallow reef opposite the Interuniversity Institute for Marine Sciences, 9 August 2019. Estimated total length: 3–4 cm. (Photographed by Dr. Gil Koplovitz).

Figure 14.
Ventral view of a phyllosoma stage III/V larva of the pygmy slipper lobster, Biarctus sordidus, sampled on 17 April 2024 off Eilat, Israel, in the northern Gulf of Aqaba (29°28′N, 34°55′E). The specimen was collected at a bottom depth of approximately 700 m using a plankton net (200-μm mesh size, 50 cm opening diameter) towed horizontally at 0.5–1 m depth. Scale bar: 500 μm (Photographed by Prof. T. Guy-Haim).
Figure 14.
Ventral view of a phyllosoma stage III/V larva of the pygmy slipper lobster, Biarctus sordidus, sampled on 17 April 2024 off Eilat, Israel, in the northern Gulf of Aqaba (29°28′N, 34°55′E). The specimen was collected at a bottom depth of approximately 700 m using a plankton net (200-μm mesh size, 50 cm opening diameter) towed horizontally at 0.5–1 m depth. Scale bar: 500 μm (Photographed by Prof. T. Guy-Haim).

Figure 15.
The pronghorn spiny lobster, Panulirus penicillatus, Coral Coast Nature Reserve, Eilat, northern Gulf of Aqaba, Israel, 22 July 2015, at a depth of 2.5 m. Estimated length (excluding antennae) ~ 40 cm (Photographed by Bader Levi).
Figure 15.
The pronghorn spiny lobster, Panulirus penicillatus, Coral Coast Nature Reserve, Eilat, northern Gulf of Aqaba, Israel, 22 July 2015, at a depth of 2.5 m. Estimated length (excluding antennae) ~ 40 cm (Photographed by Bader Levi).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



