
Submitted:
25 August 2025
Posted:
26 August 2025
You are already at the latest version
Abstract
Maternal effects have long been recognized as powerful non-genetic influences on offspring development, yet the molecular mechanisms underpinning these effects remain only partially understood. In parallel, epitranscriptomics, an emerging field centered on chemical modifications to RNA, has revealed new layers of gene regulation with implications for cell fate, plasticity, and response to environmental cues. In this perspective article, a conceptual link is proposed between maternal effects and epitranscriptomic mechanisms, focusing on how maternal environments may shape offspring phenotypes through RNA modifications. Evidence is examined from diverse systems, including maternal deposition of modified RNAs, environmental modulation of RNA-modifying enzymes, and early developmental windows sensitive to maternal inputs. Particular attention is given to how such mechanisms might operate in utero or during early embryogenesis to mediate lasting phenotypic outcomes. While direct experimental evidence remains limited, the potential for maternal cues to influence the offspring epitranscriptome is highlighted as a promising area for future research. To articulate this emerging connection, the concept of “maternal RNA imprinting” is proposed, the idea that offspring development is shaped by maternal cues via targeted RNA modifications. This article aims not only to synthesize emerging insights across fields but also to stimulate interdisciplinary discussion and encourage investigation into the unexplored intersections of maternal biology and RNA regulation.
Keywords:
Maternal effects
; Epitranscriptomics
; RNA modifications
; Developmental programming
; Non-genetic inheritance
1. Maternal Effects: Beyond Genetic Inheritance
Maternal effects have been increasingly recognized as important regulators of offspring phenotype, operating independently of the inherited DNA sequence (Mousseau et al., 2009; Wolf and Wade, 2009). Traditionally viewed through the lens of genetics, developmental outcomes were once largely attributed to allelic combinations passed from parents to progeny (Mitchell, 2022; Wolf, 2000). However, it has become evident that the maternal environment exerts a substantial influence on offspring development through a variety of non-genetic mechanisms (Galloway, 2005; Potticary and Duckworth, 2020; Räsänen and Kruuk, 2007; Wells, 2007)(Potticary and Duckworth, 2020). These maternal effects encompass physiological, biochemical, and behavioral cues, transmitted during oogenesis, gestation, or early postnatal life (Melnik et al., 2024; Mitchell, 2022; Ruebel and Latham, 2020).
In animal systems, maternal effects have been well-documented in contexts such as nutrition, stress exposure, metabolic state, and immune status (Besson et al., 2016; Burgueño et al., 2020; Edwards et al., 2021; Grueber et al., 2018; Kaiser and Sachser, 2005; Lecorguillé et al., 2021; Ma and Cain, 2025; Pollak and Weber-Stadlbauer, 2020). These factors have been shown to influence offspring traits ranging from growth rate and metabolism to neurodevelopment and immune function. In many cases, such effects have persisted into adulthood and even across generations, suggesting involvement of mechanisms capable of inducing stable, long-term changes in gene expression or cellular state (Bjorklund, 2006; Cummings et al., 2010; Ho, 2014; Melnik et al., 2024; Pollak and Weber-Stadlbauer, 2020; Shama et al., 2016; Zhang et al., 2006). Importantly, these influences are often mediated through the maternal provisioning of molecular substrates during early development, including hormones, metabolites, proteins, and RNAs (Dworkin and Dworkin-Rastl, 1990; Groothuis et al., 2019; Harry and Zakas, 2023; Macartney et al., 2022; Meylan et al., 2012; Moore et al., 2019). Recent research has expanded the understanding of maternal effects to include the regulation of gene expression programs during embryogenesis (Adrian-Kalchhauser et al., 2018; Almeida et al., 2019; Chille et al., 2021; Harry and Zakas, 2023; Hu et al., 2024; Videvall et al., 2016). Notably, maternal contributions have been implicated in modulating chromatin architecture and DNA methylation landscapes in the developing embryo (Agrelius and Dudycha, 2025; Branco et al., 2016; Champagne and Curley, 2009; Das et al., 2022; Meaney and Szyf, 2005). These epigenetic alterations offer a mechanistic explanation for how transient environmental exposures in the mother can produce sustained phenotypic outcomes in the offspring (Agrelius and Dudycha, 2025; Champagne, 2011; Sharp et al., 2018, 2017). However, while considerable attention has been given to DNA-level epigenetic marks, a growing body of evidence now points to RNA as another critical layer of regulatory control.
Maternal RNAs, transcripts deposited into the oocyte during oogenesis, serve as essential regulators of early embryonic development, particularly in organisms where zygotic genome activation is delayed (Schier, 2007; Sha et al., 2019; Winata and Korzh, 2018; Yang et al., 2024). These RNAs direct early cell divisions, patterning, and axis formation before the embryo begins transcribing its own genome (Bettegowda and Smith, 2007; Farley and Ryder, 2008; Kojima et al., 2024). The stability, translation, and function of these transcripts can be influenced by post-transcriptional modifications, some of which are now understood to be dynamic and responsive to environmental conditions (Despic and Neugebauer, 2018; Lorenzo-Orts and Pauli, 2024; Vejnar et al., 2019; Winata and Korzh, 2018). As such, maternal effects may extend into the realm of post-transcriptional regulation, potentially involving mechanisms such as RNA methylation or editing (Ding et al., 2022; J. Liu et al., 2022; Wu et al., 2022; Zhu et al., 2023). These findings raise the possibility that maternal effects may be exerted, in part, through modulation of the embryonic epitranscriptome (Frapin et al., 2020; Wang et al., 2022; Suqi Wu et al., 2023). Though this concept remains underexplored, it introduces a compelling hypothesis: that maternal effects and epitranscriptomic regulation may intersect to form a unified system of non-genetic developmental programming. By considering maternal effects as more than mere nutrient or hormonal signals, and by integrating recent discoveries from RNA biology, this article sets the stage for examining how RNA modifications may act as a mechanistic bridge linking maternal cues to offspring phenotype. This emerging perspective opens new avenues for research and invites a reevaluation of how maternal environments shape development at the molecular level.
2. Epitranscriptomics: Adding Regulatory Dimension to RNA Biology
Epitranscriptomics, the study of post-transcriptional chemical modifications on RNA molecules, has added an important new layer of regulatory complexity to gene expression. Unlike canonical epigenetics, which primarily targets DNA and histone proteins, epitranscriptomic modifications act directly on RNA, influencing its splicing, stability, localization, translation, and degradation (Arzumanian et al., 2022; Moshitch-Moshkovitz et al., 2022; Motorin and Helm, 2022; Zhao et al., 2016). More than 170 types of RNA modifications have been identified to date, with N6-methyladenosine (m6A) emerging as the most prevalent and well-characterized mark on messenger RNA (mRNA) in eukaryotes (Meyer, 2019; Meyer and Jaffrey, 2014). These modifications are catalyzed by specific enzymes, classified as “writers” (e.g., METTL3, METTL14), “erasers” (e.g., FTO, ALKBH5), and “readers” (e.g., YTH domain proteins), that together orchestrate the dynamic and reversible regulation of RNA fate (Wiener and Schwartz, 2020).
Epitranscriptomic control is now known to be tightly integrated with developmental processes. For example, m6A modifications have been shown to guide cell fate transitions during early embryogenesis, neural differentiation, and skeletogenesis (Ahi, 2025; Che et al., 2022; Haran and Lenka, 2019; Livneh et al., 2020; Yao et al., 2024; M. Zhang et al., 2020). By selectively destabilizing certain transcripts while stabilizing others, RNA modifications serve as molecular timers that fine-tune gene expression in a context- and stage-specific manner. These regulatory features are especially relevant in systems undergoing rapid and tightly coordinated transcriptional changes, such as the early embryo. Importantly, the deposition and removal of RNA modifications are influenced by cellular and environmental signals. Stress conditions, nutrient availability, hypoxia, and inflammatory cues have all been shown to alter the activity or expression of RNA-modifying enzymes (Ahi and Singh, 2024; Cayir et al., 2020). This raises the possibility that external factors, such as those originating from the maternal environment, could influence the transcriptomic output of developing tissues through epitranscriptomic means (Frapin et al., 2020; Wang et al., 2022; Suqi Wu et al., 2023). In this way, epitranscriptomics not only contributes to intrinsic gene regulatory logic but also offers a potential interface between environmental exposure and cellular response. Although the field is still in its early stages, advances in techniques such as MeRIP-seq, miCLIP, single-cell m6A profiling, direct RNA nanopore sequencing and live-cell RNA biosensors can accelerate the identification of functional RNA modifications across developmental contexts (Ahi and Khorshid, 2025; Sağlam and Akgül, 2024; Zhong et al., 2023). Future applications of these tools in maternal–fetal models may reveal novel regulatory pathways whereby maternal signals modulate the fetal transcriptome via direct chemical modification of RNA (P. Li et al., 2025; H. Liu et al., 2022). In this framework, epitranscriptomics emerges not just as a standalone regulatory system but as a candidate mediator of environmentally responsive, non-genetic inheritance.
3. Early Developmental Windows and Maternal Regulation of the Fetal Epitranscriptome
Early embryonic development is marked by a series of tightly regulated transitions, during which the embryo is highly sensitive to environmental cues, including those derived from the maternal organism (Fawcett and Frankenhuis, 2015; Wells, 2019). These windows of developmental plasticity offer unique opportunities for maternal signals to influence gene expression and cell fate decisions (Bedzhov et al., 2014). It is being proposed that such influence may extend to the epitranscriptomic layer, wherein RNA modifications could mediate adaptive responses to maternal inputs during critical periods of embryogenesis (Frapin et al., 2020; H. Liu et al., 2022; Quarto et al., 2025; Yao et al., 2024).
Several lines of evidence support the idea that maternal environments can shape the molecular landscape of the early embryo in a lasting manner (need a broad reference here). Maternal diet, metabolic state, microbiome and exposure to environmental stressors and chemicals have been shown to affect not only embryonic development but also the expression of genes involved in RNA metabolism and modification (Du et al., 2024; Dvoran et al., 2022; Frapin et al., 2020; Kunovac et al., 2023, 2021; J. Li et al., 2023; X. Li et al., 2025; Shen et al., 2022; Song et al., 2023; Xiao et al., 2022; Xiong et al., 2024; S. Zhang et al., 2020; Zheng et al., 2024). For instance, maternal protein restriction in rat has been associated with altered m6A methylation in the developing fetal hypothalamus, coinciding with changes in metabolic gene expression (Frapin et al., 2020). On the other hand, maternal obesity has been shown to dramatically affect m6A mRNA profile in placenta with adverse effects on fetal growth (Shen et al., 2022). Another study in human showed that maternal stress and psychological disturbances can lead to substantial changes in fetal m6A RNA modification patterns in association with developmental growth (J. Li et al., 2023). These findings suggest that maternal nutritional status may modulate the fetal epitranscriptome, potentially leading to persistent physiological consequences.
Timing appears to be a key factor in this interaction. During pre-implantation development, the zygote undergoes maternal-to-zygotic transition (MZT), during which maternal RNAs are degraded and zygotic transcription begins (Bai et al., 2019). It is during this phase that maternal signals, including metabolites and hormones, could influence the expression or activity of RNA-modifying enzymes in the zygote or early embryo (Despic and Neugebauer, 2018; Jiang and Fan, 2022; Sui et al., 2020; Winata et al., 2018; Xiang et al., 2025; Zhu et al., 2023). In later stages, such as gastrulation and organogenesis (Xiao et al., 2023; Yang et al., 2019; Yao et al., 2024), tissue-specific RNA modification patterns may also be shaped by maternal conditions, especially in metabolically active organs like the brain, liver, and adipose (Fang et al., 2023; Frasch et al., 2017; Heng et al., 2019; Li et al., 2016; Wang et al., 2023; Shengyu Wu et al., 2023; Xiao et al., 2019). Furthermore, the placenta itself represents a dynamic interface between maternal and fetal environments (O’Brien and Wang, 2023). Epitranscriptomic regulation in placental tissues has been increasingly recognized, with studies identifying m6A-dependent pathways that affect nutrient transport, vascular development, and immune modulation (Bian et al., 2022; Kumar et al., 2024; H. Liu et al., 2022; Qiu et al., 2021; Taniguchi et al., 2020; Wang et al., 2020; Shengyu Wu et al., 2023; Zhou et al., 2022) Given the placenta’s role in interpreting and relaying maternal cues, it is plausible that epitranscriptomic changes in placental RNA may also reflect or mediate maternal effects. Collectively, these findings support a model in which maternal influences on the fetal epitranscriptome are not only possible but likely occur during defined developmental windows when cells are transcriptionally and metabolically primed for such modulation. Investigating these windows further could yield insights into how early environmental exposures are translated into molecular phenotypes, with consequences for long-term health and development.
4. Potential Mediators: Maternal RNAs and RNA-Modifying Enzymes
A central question in linking maternal effects to epitranscriptomic regulation is identifying the molecular mediators responsible for transmitting information from the mother to the embryonic RNA landscape. Two highly plausible routes have been proposed: maternal RNAs deposited during oogenesis and the regulation of RNA-modifying enzymes in the embryo or placenta. Maternal RNAs are known to play an indispensable role in early development (Schier, 2007; Winata and Korzh, 2018; Yang et al., 2024), particularly in species where transcription from the zygotic genome is delayed (Lee et al., 2014). These transcripts, deposited in the oocyte during its maturation, are responsible for directing key developmental events until zygotic genome activation occurs. Recent studies have suggested that these maternal RNAs are not merely inert templates but may carry post-transcriptional modifications such as m6A, which influence their translation efficiency, subcellular localization, and degradation kinetics (Wu et al., 2022; Zhang et al., 2022; Zhao et al., 2017; Zhu et al., 2023). Thus, the maternal RNA pool may encode regulatory information both in sequence and in epitranscriptomic form.
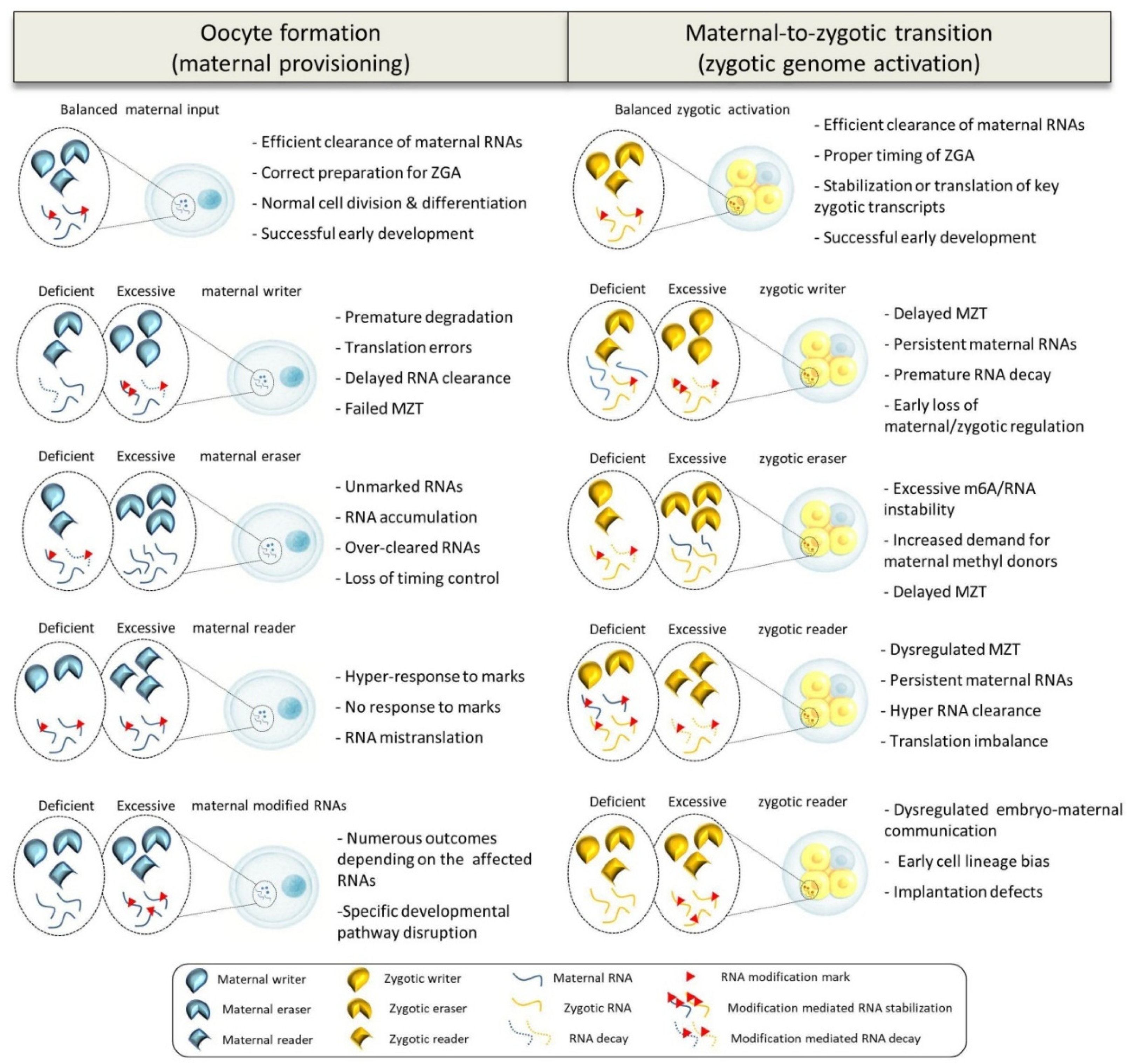
In parallel, maternal influences on the enzymatic machinery responsible for RNA modifications have also been noted. For example, the expression of METTL3, a major m6A writer enzyme, has been shown to be responsive to maternal metabolic and hormonal conditions in certain models (Sui et al., 2020; Xia et al., 2018; Zhang et al., 2022; Zheng et al., 2024, 2023). Similarly, maternal stress has been linked to altered expression of FTO, an m6A demethylase, in fetal tissues (P. Li et al., 2025; Sun et al., 2023). These enzymes may be maternally regulated either through direct maternal, fetal signaling or indirectly via placental pathways. The possibility that maternal hormones, cytokines, or metabolites affect the abundance or activity of these RNA-modifying enzymes presents a promising avenue for mechanistic studies (see Figure 2 for different potential scenarios). Moreover, recent work has shown that extracellular vesicles (EVs) released from maternal tissues can carry RNAs and RNA-binding proteins into fetal circulation (Aguilera et al., 2024; Fabbiano et al., 2020; Segura-Benítez et al., 2025). Some of these RNAs have been found to be chemically modified, raising the hypothesis that maternal EVs could serve as delivery vehicles for epitranscriptomically active cargo. Though largely speculative at present, this mechanism would provide a direct route for maternal epitranscriptomic signals to reach developing fetal tissues. Taken together, these mechanisms underscore the plausibility that maternal effects could influence offspring development via modulation of RNA modifications, either through the deposition of modified RNAs or the regulation of RNA-modifying enzymes. As tools for detecting and manipulating RNA modifications become more precise, these hypotheses are increasingly testable and could reveal a new class of maternally derived regulatory cues in development.
Figure 1.
A general overview of intersections between maternal effects and RNA modifications at developmental level.
Figure 1.
A general overview of intersections between maternal effects and RNA modifications at developmental level.
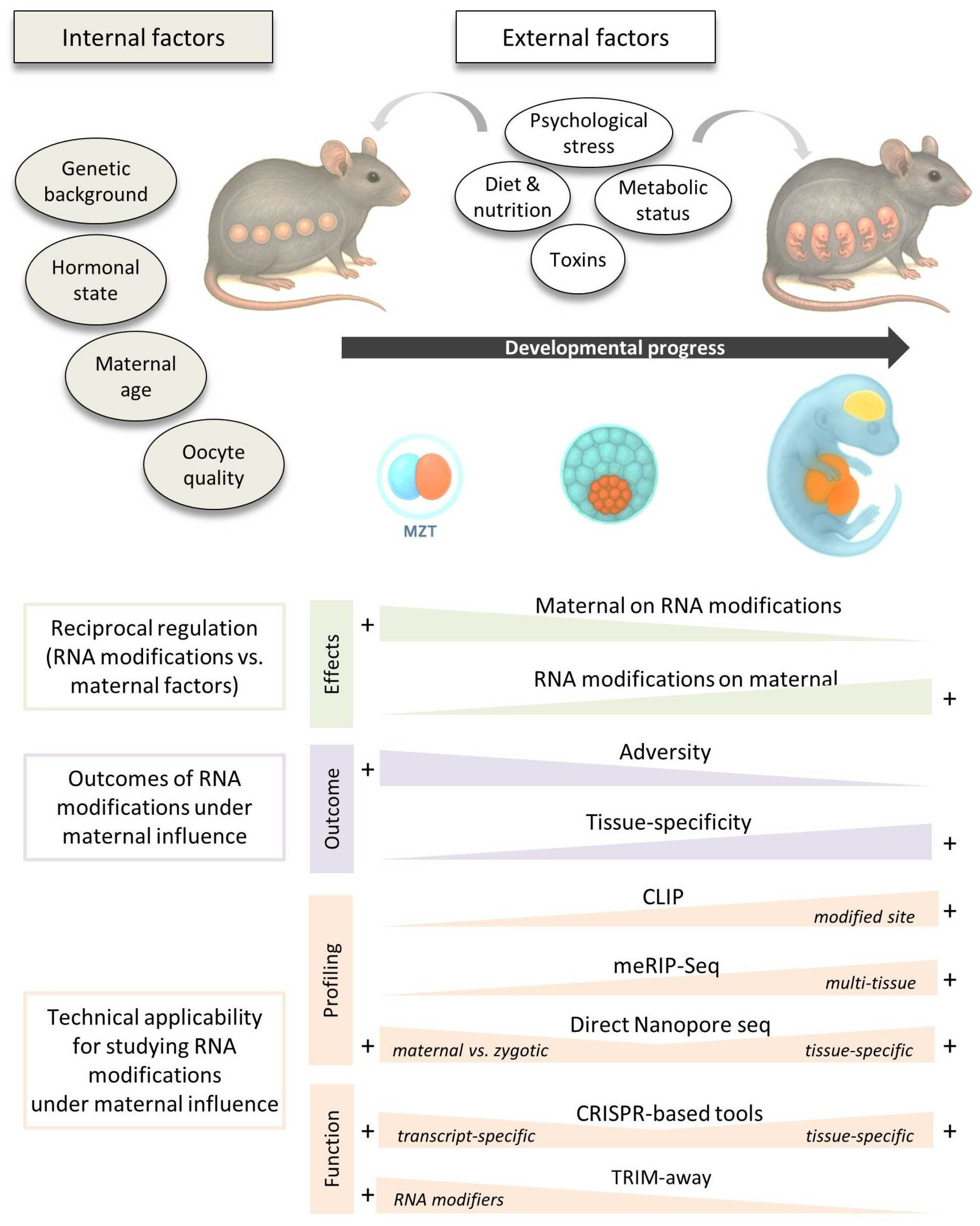
Figure 2.
Potential outcomes for regulatory influence of maternal versus zygotic RNA modification machinery in early developmental stages.
Figure 2.
Potential outcomes for regulatory influence of maternal versus zygotic RNA modification machinery in early developmental stages.
5. Evidence for Maternal-Epitranscriptomic Crosstalk: What Do We Know So Far?
While the direct regulatory connections between maternal effects and epitranscriptomics are still emerging, several independent lines of research support the plausibility of their interaction. Early clues come from models in which maternal conditions alter RNA methylation landscapes in embryonic and fetal tissues. In one notable example, the maternal microbiome was shown to affect m6A patterns in the developing brain and intestine of mouse fetuses, suggesting that maternal-derived microbial metabolites may modulate RNA methylation through metabolic or signaling pathways (Xiao et al., 2022). This finding provided one of the first functional demonstrations of maternal influence on the fetal epitranscriptome. Further supporting evidence arises from maternal nutritional studies (Frapin et al., 2020; Kaspi et al., 2018; Melnik et al., 2024; Xiao et al., 2022; Yan et al., 2025). For instance, maternal methionine supplementation, known to affect methyl group availability, has been associated with shifts in fetal brain m6A levels; this implies that methyl donors in the maternal diet could influence the activity of methyltransferase complexes (Yan et al., 2025). Similarly, maternal exposure to high-fat or low-protein diets has been linked to altered expression of m6A writers and erasers in both placental and fetal tissues, with downstream consequences for metabolic gene expression and differentiation outcomes (Frapin et al., 2020; Izquierdo et al., 2021; Kaspi et al., 2018; Melnik et al., 2024; Xiao et al., 2022).
Stress and inflammation in the maternal organism have also been associated with epitranscriptomic changes in offspring (Du et al., 2022; H. Liu et al., 2022; Ma et al., 2025; Wang et al., 2022; Yu et al., 2024; S. Zhang et al., 2020). Inflammatory cytokines, glucocorticoids, and oxidative stress, factors commonly elevated during gestational stress, can influence the abundance of RNA-modifying enzymes and alter the translation of key regulatory transcripts (Chen et al., 2024; Schroeder et al., 2024). These stress-responsive changes have been observed in brain, liver, and hematopoietic systems during fetal development (Schroeder et al., 2024; S. Zhang et al., 2020); they point toward an environmentally sensitive epitranscriptomic layer that may reflect maternal physiological states. Although most of these studies are correlational, they set the stage for more mechanistic investigations. A major limitation so far has been the difficulty in temporally and spatially resolving RNA modifications in early development. However, innovations in RNA immunoprecipitation and sequencing, combined with targeted gene editing tools, are beginning to overcome these barriers (Binan et al., 2025; Ren et al., 2023). Single-cell m6A mapping, for example, has allowed researchers to track RNA methylation states across cell types and developmental stages (Y. Li et al., 2023; Ren et al., 2025); while still technically challenging, it holds promise for tracing maternal signal-dependent changes in RNA modification patterns. The accumulating evidence underscores a growing consensus that maternal conditions do influence the epitranscriptomic state of offspring; however, the mechanistic links remain to be fully clarified. The current data justify targeted experiments to determine whether these RNA modifications are merely correlative signatures of maternal states or functionally involved in transmitting phenotypic effects across developmental time.
6. Evolutionary and Ecological Implications of Maternal Epitranscriptomic Influence
If maternal regulation of offspring epitranscriptomics proves to be a functional mechanism, it could represent a powerful form of developmental plasticity (Livneh et al., 2020; Park et al., 2020), enabling rapid phenotypic tuning in response to environmental variability without requiring genetic change (Ahi and Singh, 2024; Cayir et al., 2020; Wei et al., 2025). From an evolutionary standpoint, such a system would provide a mechanism for the transgenerational transmission of adaptive traits in response to maternal environmental cues (Legoff et al., 2019; Xiang et al., 2025), particularly under conditions where environmental conditions fluctuate more rapidly than genetic adaptation can occur (Ahi and Singh, 2024). In ecological systems, maternal effects have often been interpreted as anticipatory mechanisms, preparing offspring for expected environmental conditions based on maternal experience (Galloway, 2005; Potticary and Duckworth, 2020). Epitranscriptomic modifications, being reversible yet stable across short developmental windows, may offer an ideal molecular substrate for such transient, tunable responses (Park et al., 2020; Yoon et al., 2018). By modulating RNA stability and translation in specific tissues, maternal environments might prime offspring physiology or behavior in context-specific ways; for example, enhancing stress resilience, altering metabolic capacity, or modifying developmental timing (Kisliouk et al., 2020; H. Liu et al., 2022; Wagner and Schosserer, 2022; Yang et al., 2025). This potential utility is further amplified in species with external development, where maternal investment through the oocyte becomes the primary avenue for environmental signaling. In these systems, maternal provisioning of modified RNAs or regulators of RNA modification may serve as early warning systems, tuning gene expression programs in advance of environmental exposure.
Despite these possibilities, the evolutionary dynamics of RNA modification systems remain poorly understood. Few studies have assessed the heritability, variability, or fitness consequences of altered RNA modification profiles across generations (Bresnahan et al., 2023; Xiong et al., 2024; Zheng et al., 2024). If maternal epitranscriptomic regulation is indeed a widespread mechanism, it may challenge conventional models of non-genetic inheritance by providing a rapid, semi-stable, and reversible method for transferring information from mother to offspring. Furthermore, different species may rely on distinct epitranscriptomic strategies based on their reproductive biology, embryonic timing, or environmental unpredictability. Comparative studies across taxa, especially those with contrasting maternal provisioning modes (Navarro-Martín et al., 2020), will be essential for understanding how and why such mechanisms may have evolved. These questions open up not only new empirical challenges but also opportunities to refine theoretical models of inheritance, adaptation, and plasticity.
7. Experimental Challenges and Emerging Tools to Uncover Mechanistic Links
Investigating the potential mechanistic connections between maternal effects and epitranscriptomic regulation presents significant experimental hurdles, primarily due to the dynamic and context-specific nature of both systems. First, RNA modifications are often transient and highly cell-type-specific; detecting these marks in early embryos, which are composed of rapidly dividing and differentiating cells, requires both spatial and temporal resolution that few current methods fully provide. Another challenge lies in distinguishing whether observed RNA modifications are a direct consequence of maternal signals or arise as secondary responses during embryonic development. This distinction necessitates experimental systems that allow precise manipulation of maternal environments while tracking downstream molecular consequences in fetal tissues at defined stages.
To address these challenges, a growing set of molecular tools has been applied. Antibody-based techniques such as MeRIP-seq and miCLIP have enabled transcriptome-wide mapping of RNA modifications, particularly m6A (Sağlam and Akgül, 2024). These methods, though powerful, are limited by antibody specificity and often require high input material, making them less suitable for early developmental samples. More recently, the development of single-cell epitranscriptomic methods has begun to fill this gap, allowing for resolution of RNA modifications at the level of individual cells or lineages (Crespo-García et al., 2024). In addition, the advent of direct RNA nanopore sequencing has enabled transcriptome-wide profiling of m6A modifications without the need for chemical conversion or enrichment, offering a powerful advantage for studying RNA methylation at the zygotic or fetal stage with single-molecule precision (Zhong et al., 2023). In parallel, CRISPR-based tools have been adapted to target RNA methylation enzymes to specific transcripts; although still in early development, these systems allow direct testing of the functional role of epitranscriptomic marks in gene regulation and development (Breunig et al., 2021; Fang et al., 2020). Of particular relevance to maternal–fetal studies is the application of TRIM-away, a proteolysis-based technique that enables acute depletion of specific endogenous proteins, including RNA-modifying enzymes (Clift et al., 2018). TRIM-away has proven effective in depleting RNA methyltransferases or demethylases in early embryos without genetic manipulation, offering a rapid and reversible method for testing the requirement of these enzymes during sensitive developmental windows (Chen et al., 2019; Clift et al., 2018; Weir et al., 2021). This tool may be especially valuable for dissecting the temporal dynamics of maternal influences on the fetal epitranscriptome. Despite these advancements, technical and interpretive challenges remain. Cross-reactivity of antibodies, the functional redundancy of RNA modification enzymes, and the incomplete understanding of modification “readers” complicate the attribution of phenotypic effects to specific molecular events. Nevertheless, with the continuing refinement of tools and model systems, the field is well-positioned to explore how maternal cues may operate through RNA modifications to shape developmental trajectories.
8. Concluding Thoughts: Conceptual Bridges and Open Questions
The possibility that maternal effects and epitranscriptomic regulation intersect to shape offspring development presents a compelling framework for rethinking non-genetic inheritance. While both domains have been extensively studied in isolation, their potential integration opens a new dimension in developmental biology, one where environmental cues from the maternal organism could be translated into post-transcriptional regulatory changes in the offspring, with implications for phenotype, health, and even evolutionary fitness. This perspective article has outlined several lines of evidence that support such a connection; however, it must be emphasized that definitive mechanistic links remain largely speculative at present. Observations that maternal nutritional, microbial, or stress-related environments alter RNA modification patterns in fetal tissues are intriguing, yet causality has not been consistently demonstrated. Whether these RNA modifications are central mediators or merely responsive markers remains unresolved in many contexts.
Key questions remain to be addressed. For instance, how specific and selective are maternal influences on RNA modification machinery? Are these changes preserved through cell divisions, and can they be transmitted beyond one generation? Which transcripts are most susceptible to maternal modulation, and how do these modifications intersect with other regulatory layers such as microRNAs, alternative splicing, or chromatin state? Furthermore, to what extent do these interactions vary across tissues, developmental windows, or species?
Addressing these questions will require multidisciplinary collaboration; developmental biologists, molecular geneticists, and RNA biochemists must work in concert with systems biologists and evolutionary theorists to establish a common language and set of priorities. The development of temporally controlled, tissue-specific tools, such as TRIM-away, inducible CRISPR interference systems, and advanced imaging of modified RNAs in vivo, will be essential to disentangle maternal inputs from autonomous embryonic processes. Moreover, more comparative studies across model organisms and ecological systems are needed to test the generality and adaptability of this regulatory axis.
The purpose of this article has not been to assert a definitive model but rather to highlight an underexplored conceptual space that may prove fruitful for future research. By framing maternal effects and epitranscriptomics as potentially interconnected systems, the goal is to stimulate discussion, generate testable hypotheses, and encourage researchers in both fields to consider overlapping mechanisms. Whether or not maternal regulation of the offspring epitranscriptome proves to be widespread or only context-specific, the exploration of this intersection promises to deepen our understanding of how environment, regulation, and inheritance are woven together in shaping organismal development. In conclusion, the maternal–epitranscriptomic axis, while still in its theoretical infancy, offers a promising conceptual bridge in developmental biology—one that may ultimately transform how we understand phenotypic plasticity, intergenerational signaling, and the molecular logic of early life.
| Box 1 | Key concepts referenced in this article |
|
Maternal effects: Non-genetic influences from the mother that shape offspring phenotype. Epitranscriptomics: Study of chemical modifications on RNA that regulate its function post-transcription. m6A (N6-methyladenosine): The most abundant mRNA modification; affects RNA stability, splicing, and translation. RNA-modifying enzymes: Proteins that install (writers), remove (erasers), or recognize (readers) RNA modifications. Maternal RNAs: Transcripts deposited into the oocyte, guiding early embryonic development. Maternal-to-zygotic transition (MZT): Developmental phase where embryonic control shifts from maternal RNAs to the zygotic genome. Placental interface: The maternal–fetal exchange site; transmits nutrients, signals, and possibly RNA regulators. Developmental plasticity: The capacity of an organism to alter its developmental trajectory in response to environmental cues. Non-genetic inheritance: Transmission of traits across generations independent of DNA sequence changes. Single-cell epitranscriptomics: Techniques that profile RNA modifications at the single-cell level for spatial and temporal precision. TRIM-away: A proteolysis method used to rapidly degrade endogenous proteins, including RNA-modifying enzymes. Phenotypic programming: The process by which environmental factors during development shape long-term traits. |
Ethics approval statement
Not applicable
Consent for publication
Not applicable
Data availability statement
Data sharing is not applicable to this article as no datasets were generated or analysed during the current study.
Conflict of interest disclosure
The author declares that he has no competing interests.
Funding statement
The author received no specific funding for this work
Authors’ contributions
E.P.A. conceived the study, drafted the manuscript and designed the figures.
References
- Adrian-Kalchhauser, I., Walser, J.C., Schwaiger, M., Burkhardt-Holm, P., 2018. RNA sequencing of early round goby embryos reveals that maternal experiences can shape the maternal RNA contribution in a wild vertebrate. BMC Evol. Biol. 18, 1–14. [CrossRef]
- Agrelius, T.C., Dudycha, J.L., 2025. Maternal effects in the model system Daphnia: the ecological past meets the epigenetic future. Hered. 2025 1342 134, 142–154. [CrossRef]
- Aguilera, C., Wong, Y.S., Gutierrez-Reinoso, M.A., Velásquez, A.E., Melo-Báez, B., Cabezas, J., Caamaño, D., Navarrete, F., Castro, F.O., Rodriguez-Alvarez, L.L., 2024. Embryo-maternal communication mediated by extracellular vesicles in the early stages of embryonic development is modified by in vitro conditions. Theriogenology 214, 43–56. [CrossRef]
- Ahi, E.P., 2025. Regulation of Skeletogenic Pathways by m6A RNA Modification: A Comprehensive Review. Calcif. Tissue Int. 2025 1161 116, 1–23. [CrossRef]
- Ahi, E.P., Khorshid, M., 2025. Potentials of RNA biosensors in developmental biology. Dev. Biol. 526, 173–188. [CrossRef]
- Ahi, E.P., Singh, P., 2024. An emerging orchestrator of ecological adaptation: m6A regulation of post-transcriptional mechanisms. Mol. Ecol. 17545. [CrossRef]
- Almeida, M.V., de Jesus Domingues, A.M., Ketting, R.F., 2019. Maternal and zygotic gene regulatory effects of endogenous RNAi pathways. PLOS Genet. 15, e1007784. [CrossRef]
- Arzumanian, V.A., Dolgalev, G. V., Kurbatov, I.Y., Kiseleva, O.I., Poverennaya, E. V., 2022. Epitranscriptome: Review of Top 25 Most-Studied RNA Modifications. Int. J. Mol. Sci. 23, 13851. [CrossRef]
- Bai, S., Fu, K., Yin, H., Cui, Y., Yue, Q., Li, W., Cheng, L., Tan, H., Liu, X., Guo, Y., Zhang, Y., Xie, J., He, W., Wang, Y., Feng, H., Xin, C., Zhang, J., Lin, M., Shen, B., Sun, Z., Guo, X., Zheng, K., Ye, L., 2019. The maternal-to-zygotic transition revisited. Development 146. [CrossRef]
- Bedzhov, I., Graham, S.J.L., Leung, C.Y., Zernicka-Goetz, M., 2014. Developmental plasticity, cell fate specification and morphogenesis in the early mouse embryo. Philos. Trans. R. Soc. B Biol. Sci. 369. [CrossRef]
- Besson, A.A., Lagisz, M., Senior, A.M., Hector, K.L., Nakagawa, S., 2016. Effect of maternal diet on offspring coping styles in rodents: a systematic review and meta-analysis. Biol. Rev. 91, 1065–1080. [CrossRef]
- Bettegowda, A., Smith, G.W., 2007. Mechanisms of maternal mRNA regulation: implications for mammalian early embryonic development. Front. Biosci. 12, 3713–3726. [CrossRef]
- Bian, Y., Li, J., Shen, H., Li, Y., Hou, Y., Huang, L., Song, G., Qiao, C., 2022. WTAP dysregulation-mediated HMGN3-m6A modification inhibited trophoblast invasion in early-onset preeclampsia. FASEB J. 36, e22617. [CrossRef]
- Binan, L., Jiang, A., Danquah, S.A., Valakh, V., Simonton, B., Bezney, J., Manguso, R.T., Yates, K.B., Nehme, R., Cleary, B., Farhi, S.L., 2025. Simultaneous CRISPR screening and spatial transcriptomics reveal intracellular, intercellular, and functional transcriptional circuits. Cell 188, 2141-2158.e18. [CrossRef]
- Bjorklund, D.F., 2006. Mother knows best: Epigenetic inheritance, maternal effects, and the evolution of human intelligence. Dev. Rev. 26, 213–242. [CrossRef]
- Branco, M.R., King, M., Perez-Garcia, V., Bogutz, A.B., Caley, M., Fineberg, E., Lefebvre, L., Cook, S.J., Dean, W., Hemberger, M., Reik, W., 2016. Maternal DNA Methylation Regulates Early Trophoblast Development. Dev. Cell 36, 152–163. [CrossRef]
- Bresnahan, S.T., Lee, E., Clark, L., Ma, R., Rangel, J., Grozinger, C.M., Li-Byarlay, H., 2023. Examining parent-of-origin effects on transcription and RNA methylation in mediating aggressive behavior in honey bees (Apis mellifera). BMC Genomics 24, 1–13. [CrossRef]
- Breunig, C.T., Köferle, A., Neuner, A.M., Wiesbeck, M.F., Baumann, V., Stricker, S.H., 2021. Crispr tools for physiology and cell state changes: Potential of transcriptional engineering and epigenome editing. Physiol. Rev. 101, 177–211. [CrossRef]
- Burgueño, A.L., Juárez, Y.R., Genaro, A.M., Tellechea, M.L., 2020. Prenatal stress and later metabolic consequences: Systematic review and meta-analysis in rodents. Psychoneuroendocrinology 113, 104560. [CrossRef]
- Cayir, A., Byun, H.M., Barrow, T.M., 2020. Environmental epitranscriptomics. Environ. Res. 189, 109885. [CrossRef]
- Champagne, F.A., 2011. Maternal imprints and the origins of variation. Horm. Behav. 60, 4–11. [CrossRef]
- Champagne, F.A., Curley, J.P., 2009. Epigenetic mechanisms mediating the long-term effects of maternal care on development. Neurosci. Biobehav. Rev. 33, 593–600. [CrossRef]
- Che, Y.H., Lee, H., Kim, Y.J., 2022. New insights into the epitranscriptomic control of pluripotent stem cell fate. Exp. Mol. Med. 2022 5410 54, 1643–1651. [CrossRef]
- Chen, R., Wang, T., Tong, H., Zhang, X., Ruan, J., Qi, H., Liu, X., He, G., 2024. METTL3 and IGF2BP2 coordinately regulate FOSL1 mRNA via m6A modification, suppressing trophoblast invasion and contributing to fetal growth restriction. FASEB J. 38, e70154. [CrossRef]
- Chen, X., Liu, M., Lou, H., Lu, Y., Zhou, M.T., Ou, R., Xu, Y., Tang, K.F., 2019. Degradation of endogenous proteins and generation of a null-like phenotype in zebrafish using Trim-Away technology. Genome Biol. 20, 1–6. [CrossRef]
- Chille, E., Strand, E., Neder, M., Schmidt, V., Sherman, M., Mass, T., Putnam, H., 2021. Developmental series of gene expression clarifies maternal mRNA provisioning and maternal-to-zygotic transition in a reef-building coral. BMC Genomics 22, 1–17. [CrossRef]
- Clift, D., So, C., McEwan, W.A., James, L.C., Schuh, M., 2018. Acute and rapid degradation of endogenous proteins by Trim-Away. Nat. Protoc. 2018 1310 13, 2149–2175. [CrossRef]
- Crespo-García, E., Bueno-Costa, A., Esteller, M., 2024. Single-cell analysis of the epitranscriptome: RNA modifications under the microscope. RNA Biol. 21, 1–8. [CrossRef]
- Cummings, J.A., Clemens, L.G., Nunez, A.A., 2010. Mother counts: How effects of environmental contaminants on maternal care could affect the offspring and future generations. Front. Neuroendocrinol. 31, 440–451. [CrossRef]
- Das, A., Iwata-Otsubo, A., Destouni, A., Dawicki-McKenna, J.M., Boese, K.G., Black, B.E., Lampson, M.A., 2022. Epigenetic, genetic and maternal effects enable stable centromere inheritance. Nat. Cell Biol. 2022 245 24, 748–756. [CrossRef]
- Despic, V., Neugebauer, K.M., 2018. RNA tales - how embryos read and discard messages from mom. J. Cell Sci. 131. [CrossRef]
- Ding, Y., Zheng, Y., Wang, J., Li, H., Zhao, C., Tao, H., Li, Y., Xu, K., Huang, X., Gao, G., Chen, H., Bo, X., 2022. Recurrent RNA edits in human preimplantation potentially enhance maternal mRNA clearance. Commun. Biol. 5, 1400. [CrossRef]
- Du, R., Li, L., Wang, Y., 2022. N6-Methyladenosine-Related Gene Signature Associated With Monocyte Infiltration Is Clinically Significant in Gestational Diabetes Mellitus. Front. Endocrinol. (Lausanne). 13, 853857. [CrossRef]
- Du, Z.Y., Zhu, H.L., Chang, W., Zhang, Y.F., Ling, Q., Wang, K.W., Zhang, J., Zhang, Q.B., Kan, X.L., Wang, Q.N., Wang, H., Zhou, Y., 2024. Maternal prednisone exposure during pregnancy elevates susceptibility to osteoporosis in female offspring: The role of mitophagy/FNDC5 alteration in skeletal muscle. J. Hazard. Mater. 469, 133997. [CrossRef]
- Dvoran, M., Nemcova, L., Kalous, J., 2022. An Interplay between Epigenetics and Translation in Oocyte Maturation and Embryo Development: Assisted Reproduction Perspective. Biomed. 2022, Vol. 10, Page 1689 10, 1689. [CrossRef]
- Dworkin, M.B., Dworkin-Rastl, E., 1990. Functions of maternal mRNA in early development. Mol. Reprod. Dev. 26, 261–297. [CrossRef]
- Edwards, P.D., Lavergne, S.G., McCaw, L.K., Wijenayake, S., Boonstra, R., McGowan, P.O., Holmes, M.M., 2021. Maternal effects in mammals: Broadening our understanding of offspring programming. Front. Neuroendocrinol. 62, 100924. [CrossRef]
- Fabbiano, F., Corsi, J., Gurrieri, E., Trevisan, C., Notarangelo, M., D’Agostino, V.G., Giuseppe, V., Agostino, D.’, Risparmio, D., Trento, D., Rovereto, E., 2020. RNA packaging into extracellular vesicles: An orchestra of RNA-binding proteins? J. Extracell. Vesicles 10, e12043. [CrossRef]
- Fang, J., Wu, X., He, J., Zhang, Hanwen, Chen, X., Zhang, Hua, Novakovic, B., Qi, H., Yu, X., 2023. RBM15 suppresses hepatic insulin sensitivity of offspring of gestational diabetes mellitus mice via m6A-mediated regulation of CLDN4. Mol. Med. 29, 1–16. [CrossRef]
- Fang, L., Wang, W., Li, G., Zhang, L., Li, J., Gan, D., Yang, J., Tang, Y., Ding, Z., Zhang, M., Zhang, W., Deng, D., Song, Z., Zhu, Q., Cui, H., Hu, Y., Chen, W., 2020. CIGAR-seq, a CRISPR/Cas-based method for unbiased screening of novel mRNA modification regulators. Mol. Syst. Biol. 16, 10025. [CrossRef]
- Farley, B.M., Ryder, S.P., 2008. Regulation of Maternal mRNAs in Early Development. Crit. Rev. Biochem. Mol. Biol. 43, 135–162. [CrossRef]
- Fawcett, T.W., Frankenhuis, W.E., 2015. Adaptive explanations for sensitive windows in development. Front. Zool. 12, 1–14. [CrossRef]
- Frapin, M., Guignard, S., Meistermann, D., Grit, I., Moullé, V.S., Paillé, V., Parnet, P., Amarger, V., 2020. Maternal Protein Restriction in Rats Alters the Expression of Genes Involved in Mitochondrial Metabolism and Epitranscriptomics in Fetal Hypothalamus. Nutr. 2020, Vol. 12, Page 1464 12, 1464. [CrossRef]
- Frasch, M.G., Schulkin, J., Metz, G.A.S., Antonelli, M., 2017. Animal Models of Fetal Programming: Focus on Chronic Maternal Stress During Pregnancy and Neurodevelopment. Anim. Model. Study Hum. Dis. Second Ed. 839–849. [CrossRef]
- Galloway, L.F., 2005. Maternal effects provide phenotypic adaptation to local environmental conditions. New Phytol. 166, 93–100. [CrossRef]
- Groothuis, T.G.G., Hsu, B.Y., Kumar, N., Tschirren, B., 2019. Revisiting mechanisms and functions of prenatal hormone-mediated maternal effects using avian species as a model. Philos. Trans. R. Soc. B 374. [CrossRef]
- Grueber, C.E., Gray, L.J., Morris, K.M., Simpson, S.J., Senior, A.M., 2018. Intergenerational effects of nutrition on immunity: a systematic review and meta-analysis. Biol. Rev. 93, 1108–1124. [CrossRef]
- Haran, V., Lenka, N., 2019. Deciphering the Epitranscriptomic Signatures in Cell Fate Determination and Development. Stem Cell Rev. Reports 15, 474–496. [CrossRef]
- Harry, N.D., Zakas, C., 2023. Maternal patterns of inheritance alter transcript expression in eggs. BMC Genomics 24, 1–13. [CrossRef]
- Heng, J., Tian, M., Zhang, W., Chen, F., Guan, W., Zhang, S., 2019. Maternal heat stress regulates the early fat deposition partly through modification of m6A RNA methylation in neonatal piglets. Cell Stress Chaperones 24, 635–645. [CrossRef]
- Ho, D.H., 2014. Transgenerational Epigenetics: The Role of Maternal Effects in Cardiovascular Development. Integr. Comp. Biol. 54, 43–51. [CrossRef]
- Hu, Y., Feng, B., Wang, F., 2024. Analysis of maternal effect genes from maternal mRNA in eggs of Sogatella furcifera. Heliyon 10, e34014. [CrossRef]
- Izquierdo, V., Palomera-ávalos, V., Pallàs, M., Griñán-Ferré, C., 2021. Resveratrol Supplementation Attenuates Cognitive and Molecular Alterations under Maternal High-Fat Diet Intake: Epigenetic Inheritance over Generations. Int. J. Mol. Sci. 22, 1–18. [CrossRef]
- Jiang, Z.Y., Fan, H.Y., 2022. Five questions toward mRNA degradation in oocytes and preimplantation embryos: when, who, to whom, how, and why? Biol. Reprod. 107, 62–75. [CrossRef]
- Kaiser, S., Sachser, N., 2005. The effects of prenatal social stress on behaviour: mechanisms and function. Neurosci. Biobehav. Rev. 29, 283–294. [CrossRef]
- Kaspi, A., Khurana, I., Ziemann, M., Connor, T., Spolding, B., Zimmet, P., Walder, K., El-Osta, A., 2018. Diet during Pregnancy is Implicated in the Regulation of Hypothalamic RNA Methylation and Risk of Obesity in Offspring. Mol. Nutr. Food Res. 62, 1800134. [CrossRef]
- Kisliouk, T., Rosenberg, T., Ben-Nun, O., Ruzal, M., Meiri, N., 2020. Early-Life m6A RNA Demethylation by Fat Mass and Obesity-Associated Protein (FTO) Influences Resilience or Vulnerability to Heat Stress Later in Life. eNeuro 7, 1–14. [CrossRef]
- Kojima, M.L., Hoppe, C., Giraldez, A.J., 2024. The maternal-to-zygotic transition: reprogramming of the cytoplasm and nucleus. Nat. Rev. Genet. 2024 264 26, 245–267. [CrossRef]
- Kumar, R.P., Kumar, R., Ganguly, A., Ghosh, A., Ray, S., Islam, M.R., Saha, A., Roy, N., Dasgupta, P., Knowles, T., Niloy, A.J., Marsh, C., Paul, S., 2024. METTL3 shapes m6A epitranscriptomic landscape for successful human placentation. bioRxiv 2024.07.12.603294. [CrossRef]
- Kunovac, A., Hathaway, Q.A., Pinti, M. V., Durr, A.J., Taylor, A.D., Goldsmith, W.T., Garner, K.L., Nurkiewicz, T.R., Hollander, J.M., 2021. Enhanced antioxidant capacity prevents epitranscriptomic and cardiac alterations in adult offspring gestationally-exposed to ENM. Nanotoxicology 15, 812–831. [CrossRef]
- Kunovac, A., Hathaway, Q.A., Thapa, D., Durr, A.J., Taylor, A.D., Rizwan, S., Sharif, D., Valentine, S.J., Hollander, J.M., 2023. N6-methyladenosine (M6A) in fetal offspring modifies mitochondrial gene expression following gestational nano-TiO2 inhalation exposure. Nanotoxicology 17, 651–668. [CrossRef]
- Lecorguillé, M., Teo, S., Phillips, C.M., 2021. Maternal Dietary Quality and Dietary Inflammation Associations with Offspring Growth, Placental Development, and DNA Methylation. Nutr. 2021, Vol. 13, Page 3130 13, 3130. [CrossRef]
- Lee, M.T., Bonneau, A.R., Giraldez, A.J., 2014. Zygotic genome activation during the maternal-to-zygotic transition. Annu. Rev. Cell Dev. Biol. 30, 581–613. [CrossRef]
- Legoff, L., D’Cruz, S.C., Tevosian, S., Primig, M., Smagulova, F., 2019. Transgenerational Inheritance of Environmentally Induced Epigenetic Alterations during Mammalian Development. Cells 2019, Vol. 8, Page 1559 8, 1559. [CrossRef]
- Li, J., Gao, X., Wang, S., Yao, D., Shao, S., Wu, H., Xu, M., Yi, Q., Xie, L., Zhu, Z., Song, D., Li, H., 2023. The role of m6A methylation in prenatal maternal psychological distress and birth outcome. J. Affect. Disord. 338, 52–59. [CrossRef]
- Li, P., Lin, Y., Ma, H., Zhang, J., Zhang, Q., Yan, R., Fan, Y., 2025. Epigenetic regulation in female reproduction: the impact of m6A on maternal-fetal health. Cell Death Discov. 2025 111 11, 1–30. [CrossRef]
- Li, X., Yang, J., Zhu, Y., Liu, Y., Shi, X., Yang, G., 2016. Mouse Maternal High-Fat Intake Dynamically Programmed mRNA m6A Modifications in Adipose and Skeletal Muscle Tissues in Offspring. Int. J. Mol. Sci. 2016, Vol. 17, Page 1336 17, 1336. [CrossRef]
- Li, X., Zhao, S., Zhai, M., Ma, Y., Jiang, B., Jiang, Y., Chen, T., 2025. Extractable organic matter from PM2.5 inhibits cardiomyocyte differentiation via AHR-mediated m6A RNA methylation. J. Hazard. Mater. 486, 137110. [CrossRef]
- Li, Y., Wang, Y., Vera-Rodriguez, M., Lindeman, L.C., Skuggen, L.E., Rasmussen, E.M.K., Jermstad, I., Khan, S., Fosslie, M., Skuland, T., Indahl, M., Khodeer, S., Klemsdal, E.K., Jin, K.X., Dalen, K.T., Fedorcsak, P., Greggains, G.D., Lerdrup, M., Klungland, A., Au, K.F., Dahl, J.A., 2023. Single-cell m6A mapping in vivo using picoMeRIP–seq. Nat. Biotechnol. 2023 424 42, 591–596. [CrossRef]
- Liu, H., Zheng, J., Liao, A., 2022. The regulation and potential roles of m6A modifications in early embryonic development and immune tolerance at the maternal-fetal interface. Front. Immunol. 13, 988130. [CrossRef]
- Liu, J., Huang, T., Chen, W., Ding, C., Zhao, T., Zhao, X., Cai, B., Zhang, Y., Li, S., Zhang, L., Xue, M., He, X., Ge, W., Zhou, C., Xu, Y., Zhang, R., 2022. Developmental mRNA m5C landscape and regulatory innovations of massive m5C modification of maternal mRNAs in animals. Nat. Commun. 2022 131 13, 1–13. [CrossRef]
- Livneh, I., Moshitch-Moshkovitz, S., Amariglio, N., Rechavi, G., Dominissini, D., 2020. The m6A epitranscriptome: Transcriptome plasticity in brain development and function. Nat. Rev. Neurosci. 21, 36-51. [CrossRef]
- Lorenzo-Orts, L., Pauli, A., 2024. The molecular mechanisms underpinning maternal mRNA dormancy. Biochem. Soc. Trans. 52, 861. [CrossRef]
- Ma, J., Cain, K.D., 2025. Maternal effects on offspring immunity in fish. Fish Shellfish Immunol. 161, 110261. [CrossRef]
- Ma, X., Chen, X., Mu, X., Cao, M., Zhang, Y., 2025. Epigenetics of maternal-fetal interface immune microenvironment and placental related pregnancy complications. Front. Immunol. 16, 1549839. [CrossRef]
- Macartney, E.L., Crean, A.J., Bonduriansky, R., 2022. Parental dietary protein effects on offspring viability in insects and other oviparous invertebrates: a meta-analysis. Curr. Res. Insect Sci. 2, 100045. [CrossRef]
- Meaney, M.J., Szyf, M., 2005. Maternal care as a model for experience-dependent chromatin plasticity? Trends Neurosci. 28, 456–463. [CrossRef]
- Melnik, B.C., Weiskirchen, R., Stremmel, W., John, S.M., Schmitz, G., 2024. Risk of Fat Mass- and Obesity-Associated Gene-Dependent Obesogenic Programming by Formula Feeding Compared to Breastfeeding. Nutr. 2024, Vol. 16, Page 2451 16, 2451. [CrossRef]
- Meyer, K.D., 2019. m6A-mediated translation regulation. Biochim. Biophys. Acta - Gene Regul. Mech. 1862, 301–309. [CrossRef]
- Meyer, K.D., Jaffrey, S.R., 2014. The dynamic epitranscriptome: N6-methyladenosine and gene expression control. Nat. Rev. Mol. Cell Biol. 2014 155 15, 313–326. [CrossRef]
- Meylan, S., Miles, D.B., Clobert, J., 2012. Hormonally mediated maternal effects, individual strategy and global change. Philos. Trans. R. Soc. B Biol. Sci. 367, 1647–1664. [CrossRef]
- Mitchell, L.E., 2022. Maternal effect genes: Update and review of evidence for a link with birth defects. Hum. Genet. Genomics Adv. 3, 100067. [CrossRef]
- Moore, M.P., Whiteman, H.H., Martin, R.A., 2019. A mother’s legacy: the strength of maternal effects in animal populations. Ecol. Lett. 22, 1620–1628. [CrossRef]
- Moshitch-Moshkovitz, S., Dominissini, D., Rechavi, G., 2022. The epitranscriptome toolbox. Cell 185, 764–776. [CrossRef]
- Motorin, Y., Helm, M., 2022. RNA nucleotide methylation: 2021 update. Wiley Interdiscip. Rev. RNA 13, e1691. [CrossRef]
- Mousseau, T.A., Uller, T., Wapstra, E., Badyaev, A. V., 2009. Evolution of maternal effects: past and present. Philos. Trans. R. Soc. B Biol. Sci. 364, 1035–1038. [CrossRef]
- Navarro-Martín, L., Martyniuk, C.J., Mennigen, J.A., 2020. Comparative epigenetics in animal physiology: An emerging frontier. Comp. Biochem. Physiol. Part D Genomics Proteomics 36, 100745. [CrossRef]
- O’Brien, K., Wang, Y., 2023. The Placenta: A Maternofetal Interface. Annu. Rev. Nutr. 43, 301–325. [CrossRef]
- Park, C.W., Lee, S.M., Yoon, K.J., 2020. Epitranscriptomic regulation of transcriptome plasticity in development and diseases of the brain. BMB Rep. 53, 551. [CrossRef]
- Pollak, D.D., Weber-Stadlbauer, U., 2020. Transgenerational consequences of maternal immune activation. Semin. Cell Dev. Biol. 97, 181–188. [CrossRef]
- Potticary, A.L., Duckworth, R.A., 2020. Multiple environmental stressors induce an adaptive maternal effect. Am. Nat. 196, 487–500. [CrossRef]
- Qiu, W., Zhou, Y., Wu, H., Lv, X., Yang, L., Ren, Z., Tian, H., Yu, Q., Li, J., Lin, W., Zhao, L., Luo, S., Gao, J., 2021. RNA Demethylase FTO Mediated RNA m6A Modification Is Involved in Maintaining Maternal-Fetal Interface in Spontaneous Abortion. Front. Cell Dev. Biol. 9, 617172. [CrossRef]
- Quarto, G., Li Greci, A., Bizet, M., Penning, A., Primac, I., Murisier, F., Garcia-Martinez, L., Borges, R.L., Gao, Q., Cingaram, P.K.R., Calonne, E., Hassabi, B., Hubert, C., Herpoel, A., Putmans, P., Mies, F., Martin, J., Van der Linden, L., Dube, G., Kumar, P., Soin, R., Kumar, A., Misra, A., Lan, J., Paque, M., Gupta, Y.K., Blomme, A., Close, P., Estève, P.O., Caine, E.A., Riching, K.M., Gueydan, C., Daniels, D.L., Pradhan, S., Shiekhattar, R., David, Y., Morey, L., Jeschke, J., Deplus, R., Collignon, E., Fuks, F., 2025. Fine-tuning of gene expression through the Mettl3-Mettl14-Dnmt1 axis controls ESC differentiation. Cell 188, 998-1018.e26. [CrossRef]
- Räsänen, K., Kruuk, L.E.B., 2007. Maternal effects and evolution at ecological time-scales. Funct. Ecol. 21, 408–421. [CrossRef]
- Ren, J., Zhou, H., Zeng, H., Wang, C.K., Huang, J., Qiu, X., Sui, X., Li, Q., Wu, X., Lin, Z., Lo, J.A., Maher, K., He, Y., Tang, X., Lam, J., Chen, H., Li, B., Fisher, D.E., Liu, J., Wang, X., 2023. Spatiotemporally resolved transcriptomics reveals the subcellular RNA kinetic landscape. Nat. Methods 20, 695–705. [CrossRef]
- Ren, Z., He, J., Huang, X., Gao, Y., Wei, C., Wu, Z., Guo, W., Wang, F., Zhao, Q., Sun, X., Zhang, J., Cao, N., Lin, L., Wang, J., Cun, Y., 2025. Isoform characterization of m6A in single cells identifies its role in RNA surveillance. Nat. Commun. 2025 161 16, 1–19. [CrossRef]
- Ruebel, M.L., Latham, K.E., 2020. Listening to mother: Long-term maternal effects in mammalian development. Mol. Reprod. Dev. 87, 399–408. [CrossRef]
- Sağlam, B., Akgül, B., 2024. An Overview of Current Detection Methods for RNA Methylation. Int. J. Mol. Sci. 2024, Vol. 25, Page 3098 25, 3098. [CrossRef]
- Schier, A.F., 2007. The maternal-zygotic transition: death and birth of RNAs. Science 316, 406–407. [CrossRef]
- Schroeder, M., Fuenzalida, B., Yi, N., Shahnawaz, S., Gertsch, J., Pellegata, D., Ontsouka, E., Leiva, A., Gutiérrez, J., Müller, M., Brocco, M.A., Albrecht, C., 2024. LAT1-dependent placental methionine uptake is a key player in fetal programming of metabolic disease. Metabolism 153, 155793. [CrossRef]
- Segura-Benítez, M., Carbajo-García, M.C., Quiñonero, A., De Los Santos, M.J., Pellicer, A., Cervelló, I., Ferrero, H., 2025. Endometrial extracellular vesicles regulate processes related to embryo development and implantation in human blastocysts. Hum. Reprod. 40, 56–68. [CrossRef]
- Sha, Q.Q., Zhang, J., Fan, H.Y., 2019. A story of birth and death: mRNA translation and clearance at the onset of maternal-to-zygotic transition in mammals. Biol. Reprod. 101, 579–590. [CrossRef]
- Shama, L.N.S., Mark, F.C., Strobel, A., Lokmer, A., John, U., Mathias Wegner, K., 2016. Transgenerational effects persist down the maternal line in marine sticklebacks: gene expression matches physiology in a warming ocean. Evol. Appl. 9, 1096–1111. [CrossRef]
- Sharp, G.C., Lawlor, D.A., Richardson, S.S., 2018. It’s the mother!: How assumptions about the causal primacy of maternal effects influence research on the developmental origins of health and disease. Soc. Sci. Med. 213, 20–27. [CrossRef]
- Sharp, G.C., Salas, L.A., Monnereau, C., Allard, C., Yousefi, P., Everson, T.M., Bohlin, J., Xu, Z., Huang, R.C., Reese, S.E., Xu, C.J., Baïz, N., Hoyo, C., Agha, G., Roy, R., Holloway, J.W., Ghantous, A., Merid, S.K., Bakulski, K.M., Küpers, L.K., Zhang, H., Richmond, R.C., Page, C.M., Duijts, L., Lie, R.T., Melton, P.E., Vonk, J.M., Nohr, E.A., Williams-DeVane, C.L., Huen, K., Rifas-Shiman, S.L., Ruiz-Arenas, C., Gonseth, S., Rezwan, F.I., Herceg, Z., Ekström, S., Croen, L., Falahi, F., Perron, P., Karagas, M.R., Quraishi, B.M., Suderman, M., Magnus, M.C., Jaddoe, V.W.V., Taylor, J.A., Anderson, D., Zhao, S., Smit, H.A., Josey, M.J., Bradman, A., Baccarelli, A.A., Bustamante, M., Håberg, S.E., Pershagen, G., Hertz-Picciotto, I., Newschaffer, C., Corpeleijn, E., Bouchard, L., Lawlor, D.A., Maguire, R.L., Barcellos, L.F., Smith, G.D., Eskenazi, B., Karmaus, W., Marsit, C.J., Hivert, M.F., Snieder, H., Fallin, M.D., Melén, E., Munthe-Kaas, M.C., Arshad, H., Wiemels, J.L., Annesi-Maesano, I., Vrijheid, M., Oken, E., Holland, N., Murphy, S.K., Sørensen, T.I.A., Koppelman, G.H., Newnham, J.P., Wilcox, A.J., Nystad, W., London, S.J., Felix, J.F., Relton, C.L., 2017. Maternal BMI at the start of pregnancy and offspring epigenome-wide DNA methylation: findings from the pregnancy and childhood epigenetics (PACE) consortium. Hum. Mol. Genet. 26, 4067–4085. [CrossRef]
- Shen, W. Bin, Ni, J., Yao, R., Goetzinger, K.R., Harman, C., Reece, E.A., Wang, B., Yang, P., 2022. Maternal obesity increases DNA methylation and decreases RNA methylation in the human placenta. Reprod. Toxicol. 107, 90–96. [CrossRef]
- Song, Y.P., Lv, J.W., Zhang, Z.C., Qian, Q.H., Fan, Y.J., Chen, D.Z., Zhang, H., Xu, F.X., Zhang, C., Huang, Y., Wang, H., Wei, W., Xu, D.X., 2023. Effects of Gestational Arsenic Exposures on Placental and Fetal Development in Mice: The Role of Cyr61 m6A. Environ. Health Perspect. 131, 097004. [CrossRef]
- Sui, X., Hu, Y., Ren, C., Cao, Q., Zhou, S., Cao, Y., Li, M., Shu, W., Huo, R., 2020. METTL3-mediated m6A is required for murine oocyte maturation and maternal-to-zygotic transition. Cell Cycle 19, 391–404. [CrossRef]
- Sun, M.H., Jiang, W.J., Li, X.H., Lee, S.H., Heo, G., Zhou, D., Guo, J., Cui, X.S., 2023. High Temperature–Induced m6A Epigenetic Changes Affect Early Porcine Embryonic Developmental Competence in Pigs. Microsc. Microanal. 29, 2174–2183. [CrossRef]
- Taniguchi, K., Kawai, T., Kitawaki, J., Tomikawa, J., Nakabayashi, K., Okamura, K., Sago, H., Hata, K., 2020. Epitranscriptomic profiling in human placenta: N6-methyladenosine modification at the 5′-untranslated region is related to fetal growth and preeclampsia. FASEB J. 34, 494–512. [CrossRef]
- Vejnar, C.E., Messih, M.A., Takacs, C.M., Yartseva, V., Oikonomou, P., Christiano, R., Stoeckius, M., Lau, S., Lee, M.T., Beaudoin, J.D., Musaev, D., Darwich-Codore, H., Walther, T.C., Tavazoie, S., Cifuentes, D., Giraldez, A.J., 2019. Genome wide analysis of 3′ UTR sequence elements and proteins regulating mRNA stability during maternal-to-zygotic transition in zebrafish. Genome Res. 29, 1100–1114. [CrossRef]
- Videvall, E., Sletvold, N., Hagenblad, J., Agren, J., Hansson, B., 2016. Strong Maternal Effects on Gene Expression in Arabidopsis lyrata Hybrids. Mol. Biol. Evol. 33, 984–994. [CrossRef]
- Wagner, A., Schosserer, M., 2022. The epitranscriptome in ageing and stress resistance: A systematic review. Ageing Res. Rev. 81, 101700. [CrossRef]
- Wang, J., Gao, F., Zhao, X., Cai, Y., Jin, H., 2020. Integrated analysis of the transcriptome- wide m6A methylome in preeclampsia and healthy control placentas. PeerJ 8, e9880. [CrossRef]
- Wang, Q., Pan, M., Zhang, T., Jiang, Y., Zhao, P., Liu, X., Gao, A., Yang, L., Hou, J., 2022. Fear Stress During Pregnancy Affects Placental m6A-Modifying Enzyme Expression and Epigenetic Modification Levels. Front. Genet. 13, 927615. [CrossRef]
- Wang, S., Chen, S., Sun, J., Han, P., Xu, B., Li, X., Zhong, Y., Xu, Z., Zhang, P., Mi, P., Zhang, C., Li, L., Zhang, H., Xia, Y., Li, S., Heikenwalder, M., Yuan, D., 2023. m6A modification-tuned sphingolipid metabolism regulates postnatal liver development in male mice. Nat. Metab. 2023 55 5, 842–860. [CrossRef]
- Wei, S., Tao, H.Y., Duan, Z., Wang, Y., 2025. Environmental Exposure, Epitranscriptomic Perturbations, and Human Diseases. Environ. Sci. Technol. [CrossRef]
- Weir, E., McLinden, G., Alfandari, D., Cousin, H., 2021. Trim-Away mediated knock down uncovers a new function for Lbh during gastrulation of Xenopus laevis. Dev. Biol. 470, 74–83. [CrossRef]
- Wells, J.C.K., 2019. Developmental plasticity as adaptation: adjusting to the external environment under the imprint of maternal capital. Philos. Trans. R. Soc. B 374. [CrossRef]
- Wells, J.C.K., 2007. The thrifty phenotype as an adaptive maternal effect. Biol. Rev. 82, 143–172. [CrossRef]
- Wiener, D., Schwartz, S., 2020. The epitranscriptome beyond m6A. Nat. Rev. Genet. 2020 222 22, 119–131. [CrossRef]
- Winata, C.L., Korzh, V., 2018. The translational regulation of maternal mRNAs in time and space. FEBS Lett. 592, 3007–3023. [CrossRef]
- Winata, C.L., Łapinśki, M., Pryszcz, L., Vaz, C., Ismail, M.H. Bin, Nama, S., Hajan, H.S., Lee, S.G.P., Korzh, V., Sampath, P., Tanavde, V., Mathavan, S., 2018. Cytoplasmic polyadenylation-mediated translational control of maternal mRNAs directs maternal-to-zygotic transition. Dev. 145. [CrossRef]
- Wolf, J.B., 2000. Gene interactions from maternal effects. Evolution 54, 1882–1898. [CrossRef]
- Wolf, J.B., Wade, M.J., 2009. What are maternal effects (and what are they not)? Philos. Trans. R. Soc. B Biol. Sci. 364, 1107–1115. [CrossRef]
- Wu, Shengyu, Xie, H., Su, Y., Jia, X., Mi, Y., Jia, Y., Ying, H., 2023. The landscape of implantation and placentation: deciphering the function of dynamic RNA methylation at the maternal-fetal interface. Front. Endocrinol. (Lausanne). 14, 1205408. [CrossRef]
- Wu, Suqi, Liu, K., Zhou, B., Wu, Suwen, 2023. N6-methyladenosine modifications in maternal-fetal crosstalk and gestational diseases. Front. Cell Dev. Biol. 11, 1164706. [CrossRef]
- Wu, Y., Xu, X., Qi, M., Chen, C., Li, M., Yan, R., Kou, X., Zhao, Y., Liu, W., Li, Y., Liu, X., Zhang, M., Yi, C., Liu, H., Xiang, J., Wang, H., Shen, B., Gao, Y., Gao, S., 2022. N6-methyladenosine regulates maternal RNA maintenance in oocytes and timely RNA decay during mouse maternal-to-zygotic transition. Nat. Cell Biol. 2022 246 24, 917–927. [CrossRef]
- Xia, H., Zhong, C., Wu, X., Chen, J., Tao, B., Xia, X., Shi, M., Zhu, Z., Trudeau, V.L., Hu, W., 2018. Mettl3 mutation disrupts gamete maturation and reduces fertility in zebrafish. Genetics 208, 729–743. [CrossRef]
- Xiang, Y., Chang, H.-M., Leung, P.C.K., Bai, L., Zhu, Y., 2025. RNA modifications in female reproductive physiology and disease: emerging roles and clinical implications. Hum. Reprod. Update. [CrossRef]
- Xiao, S., Cao, S., Huang, Q., Xia, Linjian, Deng, M., Yang, M., Jia, G., Liu, X., Shi, J., Wang, W., Li, Y., Liu, S., Zhu, H., Tan, K., Luo, Q., Zhong, M., He, C., Xia, Laixin, 2019. The RNA N6-methyladenosine modification landscape of human fetal tissues. Nat. Cell Biol. 2019 215 21, 651–661. [CrossRef]
- Xiao, Y., Chen, J., Yang, S., Sun, H., Xie, L., Li, J., Jing, N., Zhu, X., 2023. Maternal mRNA deadenylation and allocation via Rbm14 condensates facilitate vertebrate blastula development. EMBO J. 42. [CrossRef]
- Xiao, Z., Liu, S., Li, Z., Cui, J., Wang, H., Wang, Zihan, Ren, Q., Xia, L., Wang, Zhijian, Li, Y., 2022. The Maternal Microbiome Programs the m6A Epitranscriptome of the Mouse Fetal Brain and Intestine. Front. Cell Dev. Biol. 10, 882994. [CrossRef]
- Xiong, Y.W., Zhu, H.L., Zhang, J., Geng, H., Tan, L.L., Zheng, X.M., Li, H., Fan, L.L., Wang, X.R., Zhang, X.D., Wang, K.W., Chang, W., Zhang, Y.F., Yuan, Z., Duan, Z.L., Cao, Y.X., He, X.J., Xu, D.X., Wang, H., 2024. Multigenerational paternal obesity enhances the susceptibility to male subfertility in offspring via Wt1 N6-methyladenosine modification. Nat. Commun. 2024 151 15, 1–20. [CrossRef]
- Yan, C., He, B., Wang, C., Li, W., Tao, S., Chen, J., Wang, Y., Yang, L., Wu, Y., Wu, Z., Liu, N., Qin, Y., 2025. Methionine in embryonic development: metabolism, redox homeostasis, epigenetic modification and signaling pathway. Crit. Rev. Food Sci. Nutr. [CrossRef]
- Yang, G., Xin, Q., Dean, J., 2024. Degradation and translation of maternal mRNA for embryogenesis. Trends Genet. 40, 238–249. [CrossRef]
- Yang, L., Ma, M., Gao, Y., Liu, J., 2025. Decoding N6-methyladenosine’s dynamic role in stem cell fate and early embryo development: insights into RNA–chromatin interactions. Curr. Opin. Genet. Dev. 91, 102311. [CrossRef]
- Yang, Y., Wang, L., Han, X., Yang, W.L., Zhang, M., Ma, H.L., Sun, B.F., Li, A., Xia, J., Chen, J., Heng, J., Wu, B., Chen, Y.S., Xu, J.W., Yang, X., Yao, H., Sun, J., Lyu, C., Wang, H.L., Huang, Y., Sun, Y.P., Zhao, Y.L., Meng, A., Ma, J., Liu, F., Yang, Y.G., 2019. RNA 5-Methylcytosine Facilitates the Maternal-to-Zygotic Transition by Preventing Maternal mRNA Decay. Mol. Cell 75, 1188-1202.e11. [CrossRef]
- Yao, Y., Liu, P., Li, Y., Wang, W., Jia, H., Bai, Y., Yuan, Z., Yang, Z., 2024. Regulatory role of m6A epitranscriptomic modifications in normal development and congenital malformations during embryogenesis. Biomed. Pharmacother. 173. [CrossRef]
- Yoon, K.J., Vissers, C., Ming, G. li, Song, H., 2018. Epigenetics and epitranscriptomics in temporal patterning of cortical neural progenitor competence. J. Cell Biol. 217, 1901–1914. [CrossRef]
- Yu, X., Xu, J., Song, B., Zhu, R., Liu, J., Liu, Y.F., Ma, Y.J., 2024. The role of epigenetics in women’s reproductive health: the impact of environmental factors. Front. Endocrinol. (Lausanne). 15, 1399757. [CrossRef]
- Zhang, G., Xu, Y., Wang, X., Zhu, Y., Wang, L., Zhang, W., Wang, Y., Gao, Y., Wu, X., Cheng, Y., Sun, Q., Chen, D., 2022. Dynamic FMR1 granule phase switch instructed by m6A modification contributes to maternal RNA decay. Nat. Commun. 2022 131 13, 1–16. [CrossRef]
- Zhang, M., Zhai, Y., Zhang, S., Dai, X., Li, Z., 2020. Roles of N6-Methyladenosine (m6A) in Stem Cell Fate Decisions and Early Embryonic Development in Mammals. Front. Cell Dev. Biol. 8, 566543. [CrossRef]
- Zhang, S., Meng, P., Cheng, S., Jiang, X., Zhang, J., Qin, X., Tang, Q., Bai, L., Zou, Z., Chen, C., 2020. Pregnancy exposure to carbon black nanoparticles induced neurobehavioral deficits that are associated with altered m6A modification in offspring. Neurotoxicology 81, 40–50. [CrossRef]
- Zhang, T.Y., Bagot, R., Parent, C., Nesbitt, C., Bredy, T.W., Caldji, C., Fish, E., Anisman, H., Szyf, M., Meaney, M.J., 2006. Maternal programming of defensive responses through sustained effects on gene expression. Biol. Psychol. 73, 72–89. [CrossRef]
- Zhao, B.S., Roundtree, I.A., He, C., 2016. Post-transcriptional gene regulation by mRNA modifications. Nat. Rev. Mol. Cell Biol. 2016 181 18, 31–42. [CrossRef]
- Zhao, B.S., Wang, X., Beadell, A. V., Lu, Z., Shi, H., Kuuspalu, A., Ho, R.K., He, C., 2017. M6 A-dependent maternal mRNA clearance facilitates zebrafish maternal-to-zygotic transition. Nature 542, 475–478. [CrossRef]
- Zheng, D., Jiang, J., Shen, A., Zhong, Y., Zhang, Y., Xiu, J., 2024. Maternal Hypertension Aggravates Vascular Dysfunction After Injury in Male Adult Offspring Through Transgenerational Transmission of N6-Methyladenosine. Hypertension. [CrossRef]
- Zheng, Z.H., Zhang, G. Le, Jiang, R.F., Hong, Y.Q., Zhang, Q.Y., He, J.P., Liu, X.R., Yang, Z.S., Yang, L., Jiang, X., Qu, L.J., Ding, C.H., Xu, Y.W., Yang, S.H., Liu, J.L., 2023. METTL3 is essential for normal progesterone signaling during embryo implantation via m6A-mediated translation control of progesterone receptor. Proc. Natl. Acad. Sci. U. S. A. 120, e2214684120. [CrossRef]
- Zhong, Z.D., Xie, Y.Y., Chen, H.X., Lan, Y.L., Liu, X.H., Ji, J.Y., Wu, F., Jin, L., Chen, J., Mak, D.W., Zhang, Z., Luo, G.Z., 2023. Systematic comparison of tools used for m6A mapping from nanopore direct RNA sequencing. Nat. Commun. 2023 141 14, 1–14. [CrossRef]
- Zhou, W., Xue, P., Yang, Y., Xia, L., Yu, B., 2022. Research progress on N6-methyladenosine in the human placenta. J. Perinat. Med. 50, 1115–1123. [CrossRef]
- Zhu, W., Ding, Y., Meng, J., Gu, L., Liu, W., Li, L., Chen, H., Wang, Y., Li, Z., Li, C., Sun, Y., Liu, Z., 2023. Reading and writing of mRNA m6A modification orchestrate maternal-to-zygotic transition in mice. Genome Biol. 24, 1–16. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



