Submitted:
24 August 2025
Posted:
25 August 2025
You are already at the latest version
Abstract
This paper addresses two aspects of molluscan shell formation. Firstly, the question of whether deposition of calcium carbonate increases atmospheric CO2 and ocean acidity as according to Ware and colleague’s hypothesis or do they form a net carbon sink? We conclude that alkaline moieties coming from a variety of sources, such as surface runoff and subsurface venting, compensate for any possible contribution of calcium carbonate formation to atmospheric CO2 and ocean acidification, but we do recognize that the current fossil fuel production of CO2 is driving both atmospheric CO2 and ocean acidification.
The second issue addresses the problems that molluscs have in getting rid of metabolically produced CO2. These include the dual function of the gills (food gathering and gas exchange), the low diffusion coefficient of CO2 in water, and the low partial pressure differential between tissue pCO2 and water pCO2. We suggest that these problems are overcome, at least in part, by sequestering tissue CO2 in shell calcium carbonate. This hypothesis can be tested by examining the effect of interferences with shell formation on molluscan pCO2 in hemolymph.
Keywords:
Carbon dioxide
; Carbon sink
; Hemolymph
; Molluscs
; Shell formation
; Respiration
1. Introduction
This paper addresses two issues with molluscan calcareous shells: i) whether shell formation contributes to net sequestration of carbon or whether it adds to atmospheric CO2 and ocean acidification, ii) whether shell formation contributes to the elimination of CO2 from molluscan tissues.
2. Does Calcareous Skeleton Formation Affect Ocean Alkalinity and Atmospheric Carbon Dioxide?
In a recent paper delineating blue carbon accumulation in the Cowichan Estuary on Vancouver Island [1], the formation of inorganic carbon accumulated in oyster shells was considered to be a source of atmospheric carbon dioxide based upon arguments presented by Ware et al. [2]. To understand the reasoning behind this, one must first consider the dynamics of the interaction of carbon dioxide with water. Carbon dioxide initially dissolves in water as a gas; some of the carbon dioxide will interact with water, forming carbonic acid that dissociates into a proton plus a bicarbonate ion. The bicarbonate ion, in turn, can dissociate into a carbonate ion and another proton. Each reaction has its own dissociation constant. The equations representing these reactions are as follows:
The formation of the inorganic calcium carbonate present in oyster shells or coral reefs is represented by the following equation:
In this second equation, we note three things: i) the formation of calcium carbonate; ii) the formation of molecular carbon dioxide from bicarbonate; and iii) the remaining presence of two protons from the first equation. Thus, there is an increase in the partial pressure of CO2 in the water and a decrease in alkalinity (increase in acidity). Theoretically, both the increased partial pressure of CO2 and increased acidity results in movement of CO2 from the water to the atmosphere. Does this actually occur? Indeed, it does appear that at a local level active calcium carbonate deposition in reefs can result in localized release of carbon dioxide from water to air, e.g. [3]. However, what we must also take into consideration is that the decrease in local alkalinity due to calcium carbonate formation is countered by the movement of alkalinity into the oceans, which will ultimately counter the decrease in local alkalinity.
Middleburg et al. discuss the difficulties in measuring how much alkalinity moves from the land into the oceans [4]. Rivers and streams carry alkaline components due to weathering of minerals, with some of these mineral-derived alkali being carried into the oceans via subsurface groundwater movement and venting. There are other sources of ocean alkalinity, such as marine phytoplankton and bacteria [5]. One can go back and forth on whether enough alkaline moieties are entering the oceans to counter the local decreases in alkalinity, since we do not know all the sources of alkalinity that enter the oceans. Perhaps the best way of determining whether the movement of alkaline components into oceans equals the decrease in alkalinity due to the formation of calcium carbonate is to look at the historical record. Zeeb and Tyrell [6] have concluded that over the past 50 million years or so, ocean pH has increased (become more alkaline) and that long-term dissolved inorganic carbon and total alkalinity was similar to pre-industrial modern values. Similar conclusions were drawn by Boudreau et al. in their studies of the past conditions [7].
It seems clear that the formation of calcareous skeletons in the ocean does not contribute to net increases in atmospheric carbon dioxide and ocean acidification. However, the increase in atmospheric carbon dioxide concentration due to human activities over the past two centuries is decreasing ocean alkalinity, which is unrelated to calcareous skeleton formation.
What is the net sequestration of carbon dioxide in molluscan shells?
The molluscan shell has two components: a mineral component comprised mainly of calcium carbonate (mostly calcite and aragonite) and an organic component comprised of insoluble chitin (a long chain polymer of N-acetylglucosamine), insoluble silk fibroin (glycine- and alanine-rich proteins), and soluble, mainly aspartic-rich acidic proteins [8]. The calcium carbonate component of the shell forms more than 95% of the mass, while the organic component forms the rest [9].
Palmer estimated that the energy costs of producing the marine gastropod shell can be divided into the cost of the calcium carbonate component (1-2 joules (J)/mg) and the cost of the organic component (29 J/mg) [10]. The major cost for the formation of calcium carbonate is the pumping of the calcium ion by Ca-ATPase. More recently, Spalding and colleagues estimate that the cost of producing the Pacific oyster shell is 0.1 J/mg for the calcium carbonate component [11]. In their calculations, Spalding and colleagues used a value of 30 J/mg for the synthesis of the organic component. Still, they suggested it could be as low as 3 J/mg organic matrix.
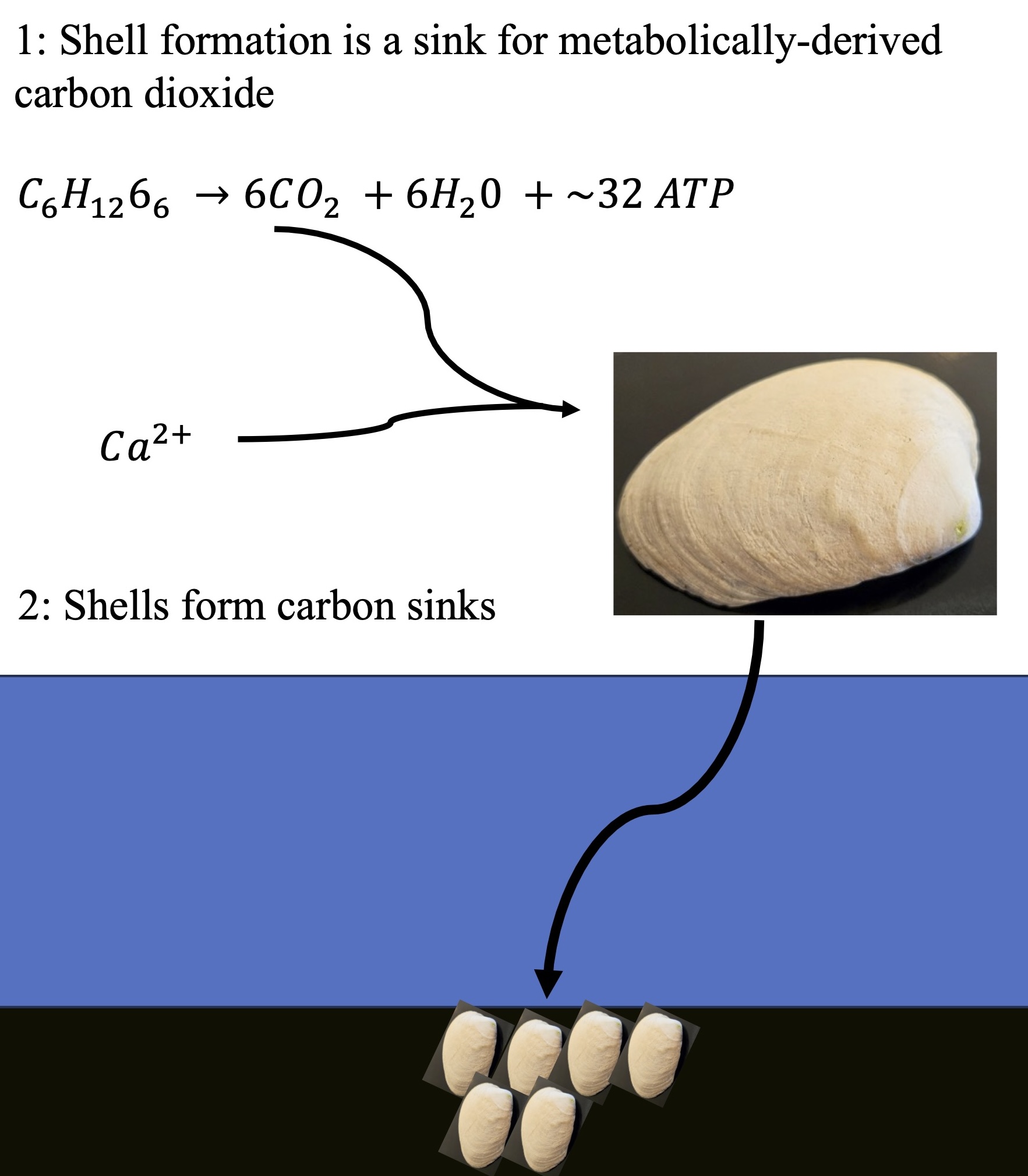
The Pacific oyster shell contains 0.9% organic matter [12]. Using the energy cost values of Spalding and colleagues, the cost for producing 1 kg of shell is 95 kJ for the calcium carbonate while the cost for the organic component is 285 kJ for a total of 380 kJ. Since the hydrolysis of one mole of ATP yields 30.5 kJ of energy, 12.46 moles of ATP were hydrolyzed to produce 1 kg of molluscan shell. The production of one mole of ATP requires the reduction of 0.167 moles of oxygen and the production of 0.167 moles of carbon dioxide; hence, the metabolism required to produce 1 kg of shell yields approximately 2.08 moles of carbon dioxide. In each kg of shell there are 991 g of calcium carbonate, or 436 g of CO2-equivalents, i.e., 9.9 moles of CO2. Thus, each kg of shell has a net sequestration of 9.9 – 2.08 = 7.82 moles of CO2, or 334 g CO2.
Thus, each tonne of shell formation results in the sequestration of about 0.334 tonnes of carbon dioxide.
3. Does Shell Production Facilitate Oyster Respiration?
The oyster shell has an obvious protective function. It may also have another function – a means to get rid of metabolically-derived CO2. The oyster respires via gills and, to some extent, the tissue surrounding the mantle cavity. The oyster gills have a dual function: respiration and capturing food particles. Because of this dual function, the gills are not as well adapted as the mammalian lung for respiration.
Gas exchange is less efficient in the oyster for additional reasons. One issue is the diffusion coefficient of CO2 in water compared to air. The diffusion coefficient of carbon dioxide at 26.85° C in water is 2.1 x 10-5 cm2/s, with slightly higher coefficients as temperature rises [13]. The diffusion coefficient in air is about four orders of magnitude greater, 0.14 cm2/sec at 26.85° C. Hence, mammals can more efficiently transfer carbon dioxide from blood to air than oysters from hemolymph to water.
Another issue is the partial pressure differential of pCO2 between hemolymph and water. In humans and other mammals, the pressure differential between venous blood pCO2 and air pCO2 is significant, with venous pCO2 ranging from 42 to 50 Torr, depending upon the level of metabolism [14], while air has a pCO2 of 0.3 Torr, i.e., a differential of 140 to 170 Torr. Exercise raises arterial pCO2 by only 1 Torr (from ~36 to 37 Torr), indicating that there is an efficient means of getting rid of carbon dioxide via the mammalian lung.
Several studies have been conducted on hemolymph collected from the adductor muscle of molluscs. The hemolymph passes through the gills and then is pumped into the tissue spaces of various organs, including the adductor muscle. Such tissue hemolymph would be comparable to a mixture of arterial and venous blood present in tissues of vertebrates. Hemolymph collected from the adductor muscle of the Densely Lamellated oyster had a pCO2 of 1.1 Torr [15]. Adductor hemolymph from the Pacific oyster had a pCO2 of 2.2 Torr [16]. pCO2 in water is not that much lower. Fresh water bodies have pCO2 levels with an average of 0.76 Torr, but with wide ranges [17]. Surface ocean water pCO2 ranges from 0.275 to 0.475 torr [18]. Estuaries have a similar range with the pCO2 in the Columbia River estuary, ranging from 0.23 to 0.44 Torr [17]. Thus, the pressure differential for pCO2 between oyster hemolymph and estuary water would range from ~2.5 to ~10. In comparison, the pressure differential in mammals ranges from 140 to 170 Torr. Thus, the oyster has several disadvantages of getting rid of metabolically-derived CO2: i) lower diffusion coefficients of CO2 in water compared to air, ii) lower partial pressures between hemolymph and water compared to between blood and air, and iii) a gas exchange organ with dual functions – respiration and food gathering.
Shell Formation
As noted above, shell formation involves the formation of calcium carbonate, which is formed by calcium cations interacting with bicarbonate anions. The bicarbonate ions are formed from the interaction of CO2 with water, catalyzed by the enzyme carbonic anhydrase in a subset of hemocytes and mantle epithelial cells [19]. We propose that this calcium carbonate formation is a significant sink for metabolically-derived CO2. This hypothesis can be tested by examining pCO2 levels in hemolymph following inhibition of shell formation.
4. Concluding Remarks
We hypothesize that molluscan shell formation acts as a carbon sink that facilitates the removal of metabolically-derived CO2. We also propose that shell formation by aquatic creatures results in net carbon sequestration. This is in agreement with the conclusions of Tamburini et al. [20].
Conflicts of Interest
There are no conflicts of interest.
References
- Douglas, T.J.; Schuerholz, G.; Juniper, S.K. Blue carbon storage in a northern temperate estuary subject to habitat loss and chronic habitat disturbance: Cowichan estuary, British Columbia, Canada. Frontiers in Marine Science 2022, 9, 857586. [Google Scholar] [CrossRef]
- Ware, J.R.; Smith, S.V.; Reaka-Kudla, M.L. Coral reefs: sources or sinks of atmospheric CO2? Coral reefs 1992, 11, 127–130. [Google Scholar] [CrossRef]
- Yan, H.; Yu, K.; Shi, Q.; Tan, Y.; Liu, G.; Zhao, M.; Li, S.; Chen, T.; Wang, Y. Seasonal variations of seawater p CO 2 and sea-air CO 2 fluxes in a fringing coral reef, northern S outh C hina S ea. Journal of Geophysical Research: Oceans 2016, 121, 998–1008. [Google Scholar] [CrossRef]
- Middelburg, J.J.; Soetaert, K.; Hagens, M. Ocean alkalinity, buffering and biogeochemical processes. Reviews of Geophysics 2020, 58, e2019RG000681. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Lee, K.; Ko, Y.H.; Lee, J.S. Contribution of marine phytoplankton and bacteria to alkalinity: An uncharacterized component. Geophysical Research Letters 2021, 48, e2021GL093738. [Google Scholar] [CrossRef]
- Zeebe, R.E.; Tyrrell, T. History of carbonate ion concentration over the last 100 million years II: Revised calculations and new data. Geochimica et Cosmochimica Acta 2019, 257, 373–392. [Google Scholar] [CrossRef]
- Boudreau, B.P.; Middelburg, J.J.; Sluijs, A.; van der Ploeg, R. Secular variations in the carbonate chemistry of the oceans over the Cenozoic. Earth and Planetary Science Letters 2019, 512, 194–206. [Google Scholar] [CrossRef]
- Furuhashi, T.; Schwarzinger, C.; Miksik, I.; Smrz, M.; Beran, A. Molluscan shell evolution with review of shell calcification hypothesis. Comparative biochemistry and physiology Part B: Biochemistry and molecular biology 2009, 154, 351–371. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Liu, Z.; Wang, L.; Song, L. Recent advances of shell matrix proteins and cellular orchestration in marine molluscan shell biomineralization. Frontiers in Marine Science 2019, 6, 41. [Google Scholar] [CrossRef]
- Palmer, A.R. Calcification in marine molluscs: how costly is it? Proceedings of the National Academy of Sciences 1992, 89, 1379–1382. [Google Scholar] [CrossRef] [PubMed]
- Spalding, C.; Finnegan, S.; Fischer, W.W. Energetic costs of calcification under ocean acidification. Global Biogeochemical Cycles 2017, 31, 866–877. [Google Scholar] [CrossRef]
- Almeida, M.; Moura, G.; Pinheiro, T.; Machado, J.; Coimbra, J. Modifications in Crassostrea gigas shell composition exposed to high concentrations of lead. Aquatic toxicology 1998, 40, 323–334. [Google Scholar] [CrossRef]
- Hou, C.; Gheorghiu, S.; Huxley, V.H.; Pfeifer, P. Reverse engineering of oxygen transport in the lung: adaptation to changing demands and resources through space-filling networks. PLoS Computational Biology 2010, 6, e1000902. [Google Scholar] [CrossRef] [PubMed]
- Hachamovitch, R.; Brown, H.V.; Rubin, S.A. Respiratory and circulatory analysis of CO2 output during exercise in chronic heart failure. Circulation 1991, 84, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Handa, T.; Araki, A.; Yamamoto, K.-i. Oxygen and acid–base status of hemolymph in the densely lamellated oyster Ostrea denselamellosa in normoxic conditions. J Nat Fish Univ 2018, 66, 203–208. [Google Scholar]
- Handa, T.; Araki, A.; Kawana, K.; Yamamoto, K.-i. Acid–base balance of hemolymph in Pacific oyster Crassostrea gigas in normoxic conditions. J Nat Fish Univ 2018, 66, 103–110. [Google Scholar]
- Hasler, C.T.; Butman, D.; Jeffrey, J.D.; Suski, C.D. Freshwater biota and rising pCO 2? Ecology Letters 2016, 19, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Rödenbeck, C.; Keeling, R.F.; Bakker, D.C.; Metzl, N.; Olsen, A.; Sabine, C.; Heimann, M. Global surface-ocean p CO 2 and sea–air CO 2 flux variability from an observation-driven ocean mixed-layer scheme. Ocean Science 2013, 9, 193–216. [Google Scholar] [CrossRef]
- Ivanina, A.V.; Falfushynska, H.I.; Beniash, E.; Piontkivska, H.; Sokolova, I.M. Biomineralization-related specialization of hemocytes and mantle tissues of the Pacific oyster Crassostrea gigas. Journal of Experimental Biology 2017, 220, 3209–3221. [Google Scholar] [CrossRef] [PubMed]
- Tamburini, E.; Turolla, E.; Lanzoni, M.; Moore, D.; Castaldelli, G. Manila clam and Mediterranean mussel aquaculture is sustainable and a net carbon sink. Science of the Total Environment 2022, 848, 157508. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



