
Submitted:
22 August 2025
Posted:
25 August 2025
You are already at the latest version
Abstract
Age-related macular degeneration (AMD), particularly its dry form, is a leading cause of irreversible vision loss due to retinal pigment epithelium (RPE) dysfunction and loss. Addressing this unmet therapeutic need requires non-invasive strategies for generating patient-specific RPE cells. This study reports the successful generation and initial characterization of RPE cells derived from urine-derived induced pluripotent stem cells (u-iPSC-RPE). Urine-derived stem cells (USCs) were isolated from healthy individuals and comprehensively characterized, confirming strong expression of renal progenitor makers and mesenchymal stem cell markers, while lacking standard hematopoietic markers. USCs were reprogrammed into iPSCs using the integration-free Sendai virus expressing the Yamanaka factors. The reprogrammed u-iPSC clones displayed characteristic pluripotency marker expression and demonstrated clearance of the Sendai virus by later passages. Subsequently, these u-iPSCs were efficiently differentiated into RPE cells, exhibiting characteristic hexagonal morphology and pigmentation which was confirmed by the expression of key RPE-specific proteins. Our findings demonstrated the feasibility of generating patient-specific RPE cells from readily accessible USCs. This reliable method for producing u-iPSC-RPE from urine-derived progenitor cells provides a promising foundation for future studies investigating their functional potential in vitro and their use in in vivo models for retinal disease modeling and therapeutic applications for AMD.
Keywords:
Age-related macular degeneration
; urine-derived stem cells
; induced pluripotent stem cells
; urine-derived iPSC
; retinal pigment epithelium
; cell therapy
; regenerative medicine
1. Introduction
Age-related macular degeneration (AMD) is a leading cause of irreversible vision loss in the USA [1,2,3] and affects close to 200 million individuals worldwide [3,4], severely diminishing quality of life and placing a significant strain on healthcare systems. Dry AMD, which accounts for approximately 80% of all AMD cases [5], can progress to geographic atrophy, which is characterized by retinal pigment epithelium (RPE) cell death, subsequent photoreceptor loss, and ultimately profound vision impairment. Currently, no effective approaches exist to prevent RPE and photoreceptor degeneration, highlighting the urgent need for innovative therapeutic interventions.
The advent of induced pluripotent stem cell (iPSC) technology has revolutionized regenerative medicine. By reprogramming somatic cells into a pluripotent state, iPSCs offer an unprecedented opportunity to generate patient-specific cell types for therapeutic transplantation, disease modeling, and drug screening. The potential of iPSC-derived RPE (iPSC-RPE) transplantation to restore retinal function in AMD patients is substantial, with initial pioneering clinical trials demonstrating their feasibility [6]. Numerous preclinical investigations involving both iPSC suspensions and monolayers continue to explore their therapeutic promise [7,8].
Despite their immense potential, several significant challenges currently impede the widespread clinical translation of iPSC therapies, and more specifically iPSC-RPE therapies. These include the risk of immune rejection associated with allogeneic (non-self) grafts, even within the immune-privileged ocular environment, and the invasive and often inefficient process of obtaining current autologous somatic cell sources, such as foreskin fibroblasts [7,9], peripheral blood mononuclear cells [8,10], bone marrow cells [11,12], and nasal mucosa [13] and conjunctival tissues [14,15]. To address these limitations, our research leverages human primary urine-derived progenitor or stem cells (USCs), a cell source we initially reported in 2008 [16,17,18], as a novel and superior alternative for iPSC generation [19,20,21,22]. USCs, a renal progenitor population, offer an easily accessible, non-invasive, and highly regenerative donor cell source [16,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55]. The ability to obtain these cells through a simple urine collection process inherently eliminates the need for invasive procedures, significantly reducing patients’ burden and simplifying the logistics of cell procurement. This non-invasive and cost-effective approach to generate autologous iPSC-RPE intrinsically eliminates the risk of immune rejection, paving the way for a personalized therapy meticulously tailored to each patient’s specific needs and disease stage.
While iPSC-RPE cells have been successfully generated from various somatic sources such as skin [7] and blood cells [8], methods for non-invasively derived iPSC-RPE are still being refined. There are clinical reliable protocols that leverage highly accessible cell sources and demonstrate efficient differentiation kinetics to enhance the clinical viability and broad accessibility of this therapy. Our previous work suggests that u-iPSCs more efficiently differentiate into neural progenitor cells [20], which are the precursor cell population for RPE cells.
In this study, we explored the feasibility of creating RPE cells from iPSCs derived from human urine samples (u-iPSC-RPE cells) and aimed to establish a reproducible protocol for generating iPSCs from USCs, rather than somatic renal tubule cells, and subsequently differentiate them into RPE-like cells. We hypothesized that the differentiated u-iPSC-RPE cells would exhibit key morphological features and express crucial protein markers comparable to native RPE cells. To achieve this, we characterized the morphology and measured the expression levels of relevant pluripotency markers in u-iPSCs, as well as RPE-specific markers in the differentiated u-iPSC-RPE cells. This work represents a crucial step toward developing an efficient and reproducible approach for autologous RPE replacement using iPSCs derived from patients’ own USCs.
2. Materials and Methods
Urine Sample Collection and USC Isolation
Urine samples were collected following a previously established protocol [16] with the approval of the Wake Forest School of Medicine Institutional Review Board. Briefly, 50-300 mL of fresh and clean-catch urine samples were obtained from consenting adult healthy male donors (n=8, aged 22-64 years). The samples were centrifuged, and the resulting cell pellet was cultured in individual wells of a 24-well tissue culture plate. Culture media and supplements were used as previously reported [16], utilizing a mixed medium of keratinocyte-serum free medium and progenitor cell medium in a 1:1 ratio. Briefly, keratinocyte serum-free medium (Thermo Fisher Scientific., Waltham, MA) was supplemented with 5 ng/ml epidermal growth factor, 50 ng/ml bovine pituitary extract, 30 ng/ml cholera toxin, 100 U/ml penicillin, and 1 mg/ml streptomycin. Progenitor cell medium consisted of 3/4 Dulbecco’s modified Eagle’s medium and 1/4 Ham’s F12 medium, supplemented with 10% fetal bovine serum (FBS, Sigma-Aldrich-Aldrich, St. Louis, MO), 0.4 µg/mL hydrocortisone (Sigma-Aldrich), 10-10 M cholera toxin (Sigma-Aldrich), 5 ng/mL insulin (Sigma-Aldrich), 1.8×10−4 M adenine (Sigma-Aldrich), 5 µg/mL transferrin (Sigma-Aldrich), 2×10−9 M 3,3’,5-triiodo-L-thyronine (Sigma-Aldrich), 10 ng/mL epidermal growth factor, and 1% penicillin-streptomycin. Unless otherwise specified, all reagents were acquired from Thermo Fisher Scientific.
Flow Cytometry Analysis for MSC and HSC Markers
For flow cytometric analysis, USCs at passage 4 (p4) were harvested by trypsinization, washed twice with ice-cold PBS, and resuspended at a concentration of 1 x 106 cells/mL Approximately 1 x 105 cells were then incubated in the dark for 30 minutes at 4°C with fluorochrome-conjugated antibodies targeting a panel of positive and negative mesenchymal stem cell (MSC) and hematopoietic stem cell (HSC) markers. The following antibodies were used: renal progenitor cell markers CD24 and CD133 [56,57,58,59]; MSC markers CD73-APC, CD90-PE, CD105-FITC; and HSC markers CD34-PerCP-Cy5.5, CD45-FITC, VEGFR2. Corresponding isotype-matched negative control antibodies (e.g., IgG1-APC, IgG1-PE, IgG1-FITC, IgG1-PerCP-Cy5.5) were used to define background fluorescence.
Following incubation, USCs were washed twice with ice-cold PBS containing 0.5% FBS and then resuspended in 300 µL of the same buffer for immediate analysis. Data acquisition was performed on a BD FACSCalibur™ flow cytometer using CellQuest™ software. A minimum of 10,000 events were acquired for each sample. Data analysis was performed using FlowJo™ software (Tree Star, Inc.). Cells were initially gated based on forward scatter (FSC) and side scatter (SSC) to exclude debris and doublets. Positive expression was determined by setting gates based on the fluorescence intensity of the isotype controls, with expression levels reported as the percentage of positive cells. All reagents were acquired from BD Biosciences (San Diego, CA).
Generation of Urine-Derived iPSCs (u-iPSCs)
To generate iPSCs, USCs were reprogrammed using the CytoTune™-iPS 2.0 Sendai Reprogramming Kit (Thermo Fisher Scientific), which utilizes Sendai virus (SeV) vectors expressing the four Yamanaka factors: OCT4, SOX2, KLF4, and c-MYC (OSKM). The methodology was based on our previously described protocol [20]. Briefly, USCs were seeded at a density of approximately 2×104 cells/well into two wells of 12-well culture plate two days prior to transduction. On the day of transduction (Day 0), a live cell count was performed on one well using a Countess™ Automated Cell Counter, and the appropriate volume of each virus added to the second well to reach the target MOI (KOS MOI = 5, hc-Myc MOI = 5, hKlf4 MOI = 3). The Sendai viral vector-containing medium was replaced with fresh medium 24 hours post-transduction.
iPSC colonies typically began to form by Day 5 post-transduction. On Day 7, cells were replated onto vitronectin-coated culture dishes at a density of 1−2×105 cells/cm2. The culture medium was then switched to Essential 8™ medium or mTeSR™ Plus medium (Thermo Fisher Scientific) for feeder-free maintenance. Colonies were manually picked and replated on Day 16-21. iPSC were routinely passaged when the colonies covered ~85% of the culture vessel surface area at a ratio of 1:3 using 0.5 mM EDTA in Calcium and Magnesium-free DPBS, Gentle Cell Dissociation Reagent or 0.02% Versene EDTA (Stemcell Technologies). For convenience, iPSCs derived from USCs are hereafter referred to as u-iPSCs.
Immunofluorescent Staining for Sendai Virus Components and Pluripotency Markers
To evaluate the clearance of Sendai viral components, iPSC colonies were analyzed by immunofluorescent staining at early passages and compared to later passages (p6). Cells were seeded onto chamber slides (Nunc™ Lab-Tek™ Chamber Slide System) and allowed to adhere. For staining, cells were fixed with 4% paraformaldehyde (PFA) for 15 minutes at room temperature, washed twice with DPBS and permeabilized with 0.1% Triton X-100 in DPBS for 10 minutes. Non-specific binding was blocked using 5% normal goat serum (NGS) and 1% BSA in DPBS for 1 hour at room temperature. Cells were incubated overnight at 4°C with primary antibody (Sendai virus HN Monoclonal Antibody 1A6, eBioscience™, Thermo Fisher Scientific) diluted in blocking solution, washed 3X with DPBS and incubated with secondary antibody (Goat anti-Rabbit IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor™ 488, Thermo Fisher Scientific) diluted in DPBS for 1 hour at room temperature. Cells were washed with DPBS 3X prior to imaging on Olympus IX-83 microscope.
Characterization of u-iPSCs Pluripotency
The following pluripotency markers were used: mouse anti-OCT4, anti-Sox2 (Santa Cruz Biotechnology), anti-SSEA4, and anti-Nanog (BD Biosciences). TRA-1-60 was used as an additional marker by live and immunofluorescent staining for early screening of iPSC clones. Following primary antibody incubation, cells were washed three times with PBS and incubated with appropriate Alexa Fluor-conjugated secondary antibodies (Thermo Fisher Scientific) diluted in blocking solution for 1 hour at room temperature in the dark. Nuclei were counterstained with DAPI (Thermo Fisher Scientific) for 5 minutes. All steps involving fluorescent antibodies were performed with minimal light exposure, with care taken to prevent sample desiccation. Appropriate controls, including negative control (omission of primary antibody), were included.
Differentiation of u-iPSCs into RPE Cells (u-iPSC-RPE cells)
USC-reprogrammed iPSCs were maintained under standard feeder-free conditions. A previously published protocol was followed to initiate differentiation [60]. In short, iPSCs were digested into small clusters using ReLeSR™. Cell clusters were transferred to Corning Ultra-Low Attachment 6-well plates and suspended as embryoid bodies (EBs) in mTeSR Plus (Stemcell Technologies) with added 10 μM ROCK inhibitor. Media was replaced after 24 hours to remove the ROCK inhibitor, and gradually replaced with differentiation induction media (DIM). After 6 days, EBs were replated on Matrigel coated 6-well plates, and the media was gradually shifted to differentiation propagation media (DPM), followed by Retinal Pigment Epithelium Maturation Media (RPEMM) at Day 22. RPEs were passaged after the onset of pigmentation at Day 40 onto fresh Matrigel-coated plates, and maintained in RPEMM. All agents and supplements were purchased from Thermo Fisher Scientific.
Characterization of u-iPSC-RPE Cells
The differentiated RPE cells were characterized using a multi-pronged approach to assess their morphology and RPE-specific protein expression. The morphology of the u-iPSC-RPE cells was assessed using phase contrast microscopy throughout the differentiation process. Immunocytochemistry was employed to confirm the expression and localization of key RPE proteins: Retinoid Isomerohydrolase (RPE65), Zona Occludens-1 (ZO-1) for tight junctions, and Melanogenesis Associated Transcription Factor (MITF). Cells were fixed with 4% paraformaldehyde, permeabilized with 0.1% Triton X-100, and blocked with 5% normal goat serum. Primary antibodies used included mouse anti-RPE65 (Abcam, Waltham, MA) and mouse anti-ZO-1 (Thermo Fisher Scientific).
3. Results
3.1. Isolation and Characterization of Urine-Derived Stem Cells
Urine-derived stem cells (USCs) were successfully isolated from the collected urine samples of healthy adult donors as described [16]. These isolated cells exhibited characteristics consistent with adult stem cells upon culture. The morphology of USCs underwent distinct changes as they were cultured and expanded in vitro (Figure 1). When initially plated, USCs started as single, isolated adherent cells, appearing by day 3 post-plating. Within a few days, the single cells have undergone several rounds of division, leading to the formation of clones and the emergence of distinct colonies by day 7. The cells continued to proliferate and displayed the expected spindle-shaped morphology often described as a “grain-of-rice” shape, common to mesenchymal stem cells (MSCs) and other fibroblast-like cells during proliferation in culture. This progression indicated successful proliferation and establishment of the cell line.
3.2. Phenotypic Characterization of USCs by Flow Cytometry
Flow cytometric analysis was performed on cultured USCs at p4 to characterize their surface marker expression, specifically evaluating the expression of common MSC and hematopoietic stem cell (HSC) markers. Representative flow cytometry plots are shown in Figure 2. The USCs consistently demonstrated positive expression of characteristic renal progenitor cell markers, markedly CD24 and CD133 [56,57,58,59], and MSC markers. Specifically, 100% of cells expressed CD24, 16.7% expressed CD133, 99.6% expressed CD29, 100% expressed CD44, 99.9% expressed CD73, 97.3% expressed CD90, and 19.8% expressed CD105 (Figure 2). These findings are consistent with the International Society for Cell & Gene Therapy (ISCT) minimal criteria for defining multipotent MSCs.
Conversely, the USCs exhibited very low or negligible expressions of hematopoietic and leukocyte surface markers. The percentage of cells positive for CD3 was <1%, CD34 was <1%, CD45 was <1%, CD14 was <1%, and VEGFR2 was <1% (Figure 2). The absence of these markers further supports the non-hematopoietic origin and mesenchymal nature of the isolated USCs, distinguishing them from hematopoietic stem cells or contaminating immune cells. Collectively, these results confirmed that the urine-derived cells isolated and expanded under the described conditions possessed a surface immunophenotype highly characteristic of multipotent MSCs.
3.3. Morphology of Urine-Derived iPSC Colonies
Reprogrammed u-iPSC colonies became visible after approximately six days post-transduction and displayed the characteristic iPSC morphology. These aggregates were presented as relatively round colonies with sharp, well-defined edges (Figure 3b). High-magnification imaging revealed tightly packed cells exhibiting a high nucleus-to-cytoplasm ratio and prominent nucleoli. Colony centers often appeared as dense areas with increased brightness under phase-contrast microscopy (Figure 3c).
3.4. Monitoring the Clearance of Sendai Viral Components in Reprogrammed u-iPSCs
A critical step for generating clinically relevant iPSCs is ensuring the complete clearance of any reprogramming vectors. This makes the use of Sendai virus for iPSC generation advantageous due to its non-integrating nature, significantly enhancing the resulting iPSCs’ safety for potential therapeutic applications. We monitored the clearance of Sendai viral components via immunofluorescence staining at various passages. Our analysis employed an anti-Sendai virus antibody, targeting hemagglutinin-neuraminidase (HN), a viral protein co-expressed with the reprogramming genes. We observed a progressive reduction in HN over time during culture of the cells. By later passages (p6), the u-iPSC lines demonstrated a complete absence of detectable signal for anti-Sendai virus (Figure 4), confirming their “virus-free” status. This crucial finding indicates that exogenous reprogramming genes were no longer expressed in the final u-iPSC lines.
3.5. Generation and Characterization of u-iPSCs
To confirm their pluripotent state, immunofluorescence staining was performed for key pluripotency markers. u-iPSCs expressed TRA-1-60, a surface marker expressed on pluripotent cells (Figure 5) and exhibited robust nuclear expression of the transcription factors SOX2, OCT3/4, and NANOG and surface expression of SSEA4 (Figure 6).
3.6. Differentiation and Initial Characterization of u-iPSC-Derived RPE Cells
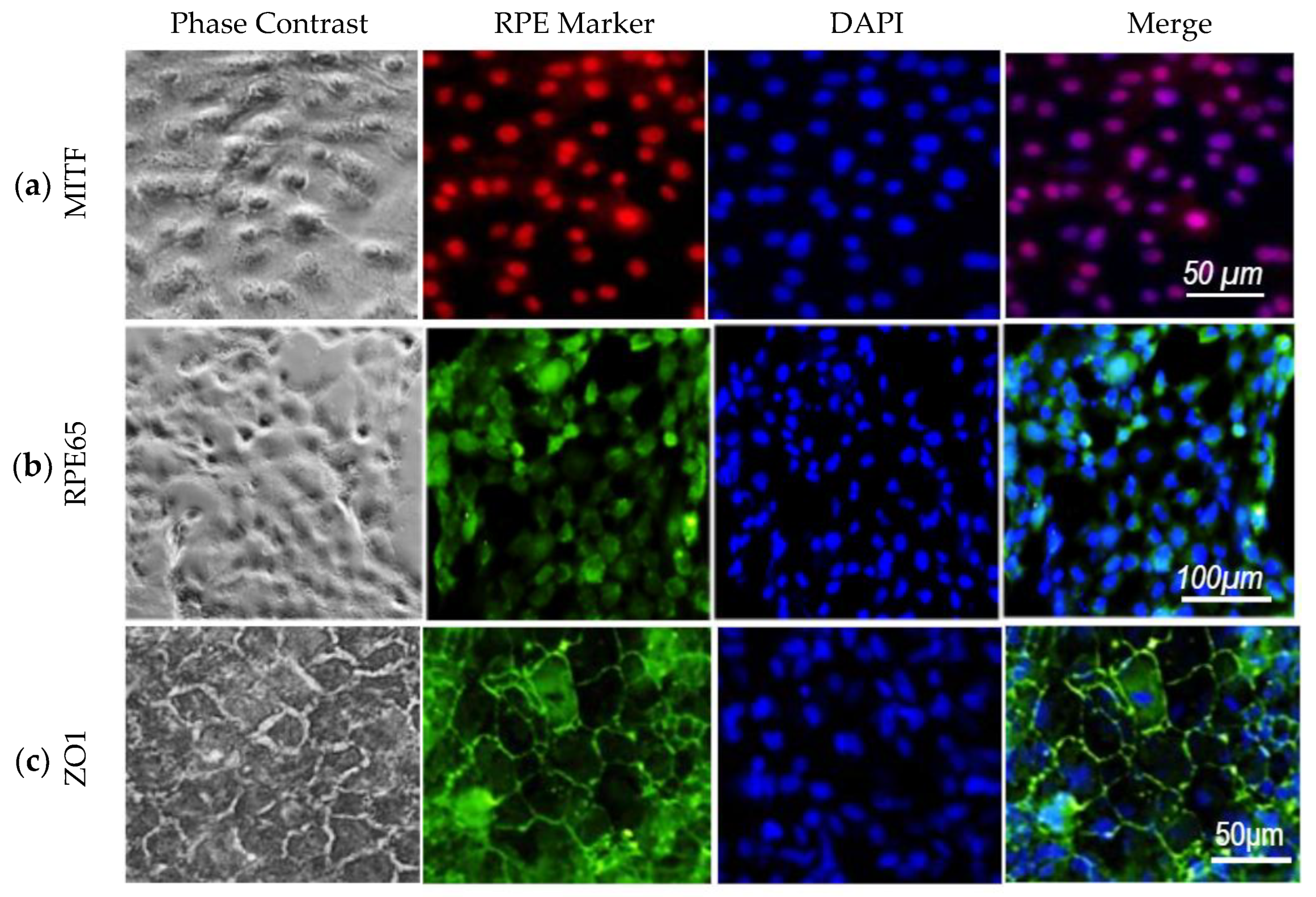
The established u-iPSC lines were subjected to a direct differentiation protocol to generate RPE cells. Throughout the differentiation process, we closely monitored the formation of key morphological hallmarks of RPE maturation. In the first 3-5 days after the start of differentiation, the cells began forming embryoid bodies (Figure 7). Several weeks post-differentiation, cells exhibited characteristic cobblestone-like arrangements and showed visible pigment (Figure 7). Immunocytochemistry was employed to confirm the expression of essential RPE-specific proteins. Immunofluorescence analysis demonstrated that the differentiated cells robustly expressed and correctly localized the key RPE markers MITF, RPE65, and ZO-1 (Figure 8).
Collectively, these findings demonstrate the successful differentiation of u-iPSCs into RPE cells that exhibit characteristic morphology and express a panel of crucial RPE-specific markers, indicating their potential for further functional maturation. While beyond the scope of this initial characterization, future studies will investigate the functional capabilities of these u-iPSC-RPE cells, including RPE65 isomerase activity and their ability to produce 11-cis retinol.
4. Discussion
In this study, we successfully established a reliable and non-invasive method for generating u-iPSC-RPE cells. Our findings demonstrate that stem cells, as opposed to other somatic cells in urine, are responsible for iPSC generation. These u-iPSCs exhibit characteristic pluripotency and can be efficiently guided towards an RPE lineage, expressing key morphological and molecular markers of RPE.
The choice of the initial cell source for iPSC generation significantly impacts the efficiency, safety, and therapeutic potential of the derived cells. While various somatic cells like skin fibroblasts, PBMCs, bone marrow cells, and nasal or conjunctival tissues have been explored, each presents unique challenges in terms of invasiveness and accessibility. Our work highlights the distinct advantages of using urine-derived stem cells (USCs) as a source for iPSC generation. Urine collection is non-invasive, cost-effective, and provides an easily accessible and virtually unlimited source of cells from healthy adult donors. This streamlined collection process significantly reduces patient burden and enhances the feasibility of generating autologous iPSCs for personalized therapeutic applications in conditions like AMD.
USCs are of epithelial lineage, most likely originating from glomerular parietal epithelial cells (PECs) [17]. PECs, which line Bowman’s capsule in the renal glomerulus, exhibit distinct differentiation patterns based on their location. Specifically, PECs located near the vascular pole are thought to act as progenitors for podocytes, while those at the tubular pole are involved in generating renal tubular epithelial cells [56,61,62,63,64,65]. Due to their embryonic origin from the intermediate mesoderm [66] and the occurrence of epithelial-mesenchymal transition (EMT), a process that promotes mesenchymal traits [67], USCs express MSC markers (Figure 2). They also express strong renal progenitor makers (CD24 and CD133 [56,57,58,59]) and are negative for hematopoietic stem cell markers, results consistent with similar serial studies from our lab [16,17,18] and others. Our previous findings also indicate that USCs possess inherent “stemness” properties, including high telomerase activity and relatively long telomeres [68], in addition to the capability of being differentiated down several cell lineages to obtain a variety of cell types, including epithelial cells [69,70,71], endothelial cells [72], skeletal myocytes [73], osteocytes [24,68,74], chondrocytes [24,71,75] and adipocytes [24]. These characteristics are highly favorable for efficient cellular reprogramming and robust proliferative capacity, offering significant advantages over other somatic cells for efficient iPSC generation.
The precise identification of the starting cell population for urine-derived iPSCs (u-iPSCs), and ultimately RPE (u-iPSC-RPE), generation is essential. Previous studies on u-iPSC often lack clarity regarding the specific cell type in urine used for reprogramming, such as exfoliated renal tubule epithelial cells [21,22] or mesenchymal stromal cells [76]. Urine contains a heterogeneous mix of cells shed from the urinary tract (kidney tubules, bladder, ureters, urethra), with a majority of differentiated epithelial cells and a smaller population of progenitor or stem cells [16]. Although the reprogramming of somatic cells into iPSC is theoretically feasible, the efficiency varies greatly depending on the initial cell type and its original differentiation state. This study, however, clearly demonstrates that the cells utilized for iPSC generation under our culture conditions are of stem cell lineage, rather than somatic cells. Somatic renal tubule epithelial cells are typically lost after subculture [16] and are not sustainable in long-term culture; only USCs consistently proliferate rapidly through multiple passages in our low-serum (2.5% FBS) culture system (Figure 1). Stromal cells are capable of extensive passaging, however they do not possess the stemness features (i.e., renal progenitor makers, MSC markers, telomerase activity, and multiple differentiation capacity) observed in our cells. This understanding of our starting material helps ensure efficient and consistent reprogramming, resulting in high-quality iPSCs.
For downstream applications requiring sufficient cell numbers, USCs at passage 3 (p3) were utilized for reprogramming. These cells exhibited robust proliferative capacity, yielding an average of over 1×107 cells and approximately 10 clones per 200 mL of urine by p3 within just 3-4 weeks. This impressive expansion highlights their potential as an abundant and accessible cell source for iPSC generation, which can be used to further induce RPE differentiation. Moreover, our study suggests that USCs may exhibit faster reprogramming into iPSCs (6-10 days for iPSC colony formation) compared to other somatic cells, such as skin fibroblasts or PBMCs, which typically require 4 weeks [19]. This accelerated timeline provides additional downstream benefits to using urine-derived iPSC for RPE generation.
Our iPSC generation protocol [20], employing Sendai viral vectors for reprogramming, consistently yielded u-iPSCs that displayed typical iPSC morphology and robust expression of pluripotency markers. A key advantage of this approach is the non-integrating nature of Sendai virus (SeV). As an RNA virus, SeV completes its life cycle exclusively in the cytoplasm, thus preventing genomic integration and eliminating the risk of insertional mutagenesis. This feature ensures the production of “transgene-free” iPSCs, where the viral genetic material is naturally cleared during cell division, thus addressing concerns about residual viral DNA and reducing potential long-term immunogenic risks, all while maintaining high reprogramming efficiency across diverse cell types. An important aspect of our methodology was the rigorous monitoring and confirmation of Sendai viral vector clearance, ensuring the generated u-iPSCs were “virus-free” and “transgene-free” by later passages (Figure 4). This non-integrating approach is paramount for the safety and clinical translation of iPSC-based therapies, addressing concerns associated with genomic integration from other reprogramming methods.
Once reprogrammed, picking u-iPSC colonies at an early stage is a critical step for successful expansion. In this study, we aimed to identify colonies matching the typical morphology of healthy, undifferentiated iPSC colonies: round, larger and sharp-edged, with large nuclei. In general, the use of multiple independent markers of iPSCs (such as SOX2/ OCT4/DAPI, and NANOG/SSEA4/DAPI used in this study) for confirmation of pluripotency is considered more precise and provides a higher level of confidence in identifying iPSCs compared to single or double staining.
The subsequent differentiation of these u-iPSCs into RPE cells resulted in cells exhibiting the characteristic hexagonal morphology and pigmentation of mature RPE (Figure 7). The expression and correct localization of key RPE-specific proteins, including RPE65, MITF, and ZO-1, confirmed by immunocytochemistry (Figure 8), unequivocally establish the capacity of u-iPSCs to differentiate into RPE. This aligns with and builds upon previous studies that have successfully differentiated iPSCs from other somatic cell sources into RPE cells [6,60,77]. The successful differentiation of u-iPSCs into RPE cells with these key characteristics of native RPE suggests their potential for therapeutic application in replacing damaged or lost RPE in dry AMD.
While our study successfully demonstrated the initial morphological and molecular characteristics of u-iPSC-RPE cells, comprehensive functional assays were beyond the scope of this investigation. We did not assess key functional properties of mature RPE, such as transepithelial electrical resistance (TEER), phagocytosis of photoreceptor outer segments, visual cycle activity (e.g., 11-cis retinal production), and growth factor secretion (e.g., PEDF). Although the expression of RPE65, a crucial visual cycle enzyme, strongly suggests functional potential, direct measurement of its isomerase activity is essential to fully verify physiological RPE function. Future studies will thoroughly characterize the functional maturity of these u-iPSC-RPE cells in both in-vitro and in-vivo models. Addressing challenges stemming from potential variability between donor lines and optimizing scalability for mass production provide additional steps pivotal to advancing this research and its translation to future clinical applications.
5. Conclusions
Our findings underscore the use of urine-derived iPSCs and the promise of autologous u-iPSC-RPE cells for future therapeutic applications in AMD. The ease of urine collection, coupled with the robust reprogramming and differentiation efficiency of multipotent USCs demonstrated in this study, positions u-iPSC-RPE as a highly attractive and non-invasive source for patient-specific cell therapy. Beyond direct clinical applications, u-iPSC-RPE also presents a versatile platform for disease modeling and drug screening, offering enhanced fidelity in recapitulating native RPE structure and function. The improved understanding of AMD pathophysiology facilitated by these cells holds promise to accelerate the identification and screening of novel therapeutic agents. Our overarching goal is to develop an efficient u-iPSC-RPE-based therapy for AMD by optimizing donor cell selection and patient targeting, including the investigation of u-iPSC-RPE derived from USCs from AMD patients with or without specific gene mutations. Following successful preclinical validation, we envision paving the way for human clinical trials, ultimately revolutionizing AMD treatment and offering hope to millions of patients worldwide through an innovative, non-invasive, and highly personalized therapeutic option.
Author Contributions
Conceptualization, Y.Z.; methodology, D.B., H.Z. and CB.; software, HZ.; validation, Y.Z., D.B., H.Z. and C.B.; formal analysis, Y.Z., D.B. and H.Z.; investigation, Y.Z., D.B., H.Z. and CB.; resources, Y.Z., J.M. and A,A.; data curation, Y.Z., D.B., H.Z. and C.B.; writing—original draft preparation, Y.Z.; writing—review and editing, Y.Z., J.M., T.C., D.B. and A.A.; visualization, Y.Z., D.B. and H.Z.; supervision, Y.Z., J.M., A.A.; project administration, Y.Z.; funding acquisition, Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Institute of Allergy and Infectious Diseases, National Institutes of Health, under Contract No. NIH/NEI R21 EY0358332024; NIH/HIAID R21 AI152832, R03 AI165170; 2025 Translational Team Science Pilot Award, Translational Eye and Vision Research Center (TrEVR, Pilot Research Grant 2024–2025), Wake Forest School of Medicine (WFSM); 2024 Pilot Research Grant, Eye Bank Association of America (EBAA); 2025 Ignition Fund Pilot award of Wake Forest School of Medicine (Y. Z: PI).
Institutional Review Board Statement
The study protocol was approved by the Institutional Review Board of the Wake Forest Institute for Regenerative Medicine (IRB00014033, 9/21/2010).
Informed Consent Statement
Informed consent was obtained from all urine-sample donors involved in the study.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AMD | Age-Related Macular Degeneration |
| USC | Urine-Derived Stem Cells |
| iPSC | Induced Pluripotent Stem Cells |
| RPE | Retinal Pigment Epithelium |
| PEC | Parietal Epithelial Cells |
| SeV | Sendai Virus |
| EB | Embryoid Body |
| DIM | Differentiation Induction Media |
| DPM | Differentiation Propagation Media |
| RPEMM | Retinal Pigment Epithelium Maturation Media |
References
- Klein, R.; Klein, B.E.K. The Prevalence of Age-Related Eye Diseases and Visual Impairment in Aging: Current Estimates. Investig. Ophthalmol. Vis. Sci. 2013, 54, ORSF5–ORSF13. [Google Scholar] [CrossRef] [PubMed]
- Group*, T.E.D.P.R. Prevalence of Age-Related Macular Degeneration in the United States. Arch. Ophthalmol. 2004, 122, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Pappas, C.M.; Zouache, M.A.; Matthews, S.; Faust, C.D.; Hageman, J.L.; Williams, B.L.; Richards, B.T.; Hageman, G.S. Protective chromosome 1q32 haplotypes mitigate risk for age-related macular degeneration associated with the CFH-CFHR5 and ARMS2/HTRA1 loci. Hum. Genom. 2021, 15, 60. [Google Scholar] [CrossRef]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.G.; Klein, R.; Cheng, C.-Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: a systematic review and meta-analysis. The Lancet Global Health 2014, 2, e106–e16. [Google Scholar] [CrossRef]
- Ebeling, M.C.; Geng, Z.; Stahl, M.R.; Kapphahn, R.J.; Roehrich, H.; Montezuma, S.R.; Ferrington, D.A.; Dutton, J.R. Testing Mitochondrial-Targeted Drugs in iPSC-RPE from Patients with Age-Related Macular Degeneration. Pharmaceuticals 2022, 15. [Google Scholar] [CrossRef]
- Mandai, M.; Watanabe, A.; Kurimoto, Y.; Hirami, Y.; Morinaga, C.; Daimon, T.; Fujihara, M.; Akimaru, H.; Sakai, N.; Shibata, Y.; Terada, M.; Nomiya, Y.; Tanishima, S.; Nakamura, M.; Kamao, H.; Sugita, S.; Onishi, A.; Ito, T.; Fujita, K.; Kawamata, S.; Go, M.J.; Shinohara, C.; Hata K-i Sawada, M.; Yamamoto, M.; Ohta, S.; Ohara, Y.; Yoshida, K.; Kuwahara, J.; Kitano, Y.; Amano, N.; Umekage, M.; Kitaoka, F.; Tanaka, A.; Okada, C.; Takasu, N.; Ogawa, S.; Yamanaka, S.; Takahashi, M. Autologous Induced Stem-Cell–Derived Retinal Cells for Macular Degeneration. New Engl. J. Med. 2017, 376, 1038–1046. [Google Scholar] [CrossRef]
- Rajendran Nair, D.S.; Zhu, D.; Sharma, R.; Martinez Camarillo, J.C.; Bharti, K.; Hinton, D.R.; Humayun, M.S.; Thomas, B.B. Long-Term Transplant Effects of iPSC-RPE Monolayer in Immunodeficient RCS Rats. Cells 2021, 10. [Google Scholar] [CrossRef]
- Sharma, R.; Khristov, V.; Rising, A.; Jha, B.S.; Dejene, R.; Hotaling, N.; Li, Y.; Stoddard, J.; Stankewicz, C.; Wan, Q.; Zhang, C.; Campos, M.M.; Miyagishima, K.J.; McGaughey, D.; Villasmil, R.; Mattapallil, M.; Stanzel, B.; Qian, H.; Wong, W.; Chase, L.; Charles, S.; McGill, T.; Miller, S.; Maminishkis, A.; Amaral, J.; Bharti, K. Clinical-grade stem cell-derived retinal pigment epithelium patch rescues retinal degeneration in rodents and pigs. Sci Transl Med 2019, 11. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Yin, J.; Huo, W.; Chaum, E. Modeling of mitochondrial bioenergetics and autophagy impairment in MELAS-mutant iPSC-derived retinal pigment epithelial cells. Stem Cell Res. Ther. 2022, 13, 260. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; David, B.T.; Trawczynski, M.; Fessler, R.G. Advances in Pluripotent Stem Cells: History, Mechanisms, Technologies, and Applications. Stem Cell Rev Rep 2020, 16, 3–32. [Google Scholar] [CrossRef] [PubMed]
- Aboutaleb Kadkhodaeian, H.; Tiraihi, T.; Ahmadieh, H.; Ziaei, H.; Daftarian, N.; Taheri, T. Generation of Retinal Pigmented Epithelium-Like Cells from Pigmented Spheres Differentiated from Bone Marrow Stromal Cell-Derived Neurospheres. Tissue Eng Regen Med 2019, 16, 253–263. [Google Scholar] [CrossRef]
- Tian, P.; Elefanty, A.; Stanley, E.G.; Durnall, J.C.; Thompson, L.H.; Elwood, N.J. Creation of GMP-Compliant iPSCs From Banked Umbilical Cord Blood. Front Cell Dev Biol 2022, 10, 835321. [Google Scholar] [CrossRef]
- Ono, M.; Hamada, Y.; Horiuchi, Y.; Matsuo-Takasaki, M.; Imoto, Y.; Satomi, K.; Arinami, T.; Hasegawa, M.; Fujioka, T.; Nakamura, Y.; Noguchi, E. Generation of induced pluripotent stem cells from human nasal epithelial cells using a Sendai virus vector. PLoS One 2012, 7, e42855. [Google Scholar] [CrossRef]
- Geng, Z.; Walsh, P.J.; Truong, V.; Hill, C.; Ebeling, M.; Kapphahn, R.J.; Montezuma, S.R.; Yuan, C.; Roehrich, H.; Ferrington, D.A.; Dutton, J.R. Generation of retinal pigmented epithelium from iPSCs derived from the conjunctiva of donors with and without age related macular degeneration. PLoS One 2017, 12, e0173575. [Google Scholar] [CrossRef] [PubMed]
- Fisher, C.R.; Ebeling, M.C.; Geng, Z.; Kapphahn, R.J.; Roehrich, H.; Montezuma, S.R.; Dutton, J.R.; Ferrington, D.A. Human iPSC- and Primary-Retinal Pigment Epithelial Cells for Modeling Age-Related Macular Degeneration. Antioxidants 2022, 11. [Google Scholar] [CrossRef]
- Zhang, Y.; McNeill, E.; Tian, H.; Soker, S.; Andersson, K.-E.; Yoo, J.J.; Atala, A. Urine Derived Cells are a Potential Source for Urological Tissue Reconstruction. J. Urol. 2008, 180, 2226–2233. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.L.; Li, Q.; Ma, J.X.; Atala, A.; Zhang, Y. Body fluid-derived stem cells - an untapped stem cell source in genitourinary regeneration. Nat Rev Urol 2023. [CrossRef]
- Yu, P.; Bosholm, C.C.; Zhu, H.; Duan, Z.; Atala, A.; Zhang, Y. Beyond waste: understanding urine’s potential in precision medicine. Trends Biotechnol 2024. [CrossRef] [PubMed]
- Guan, X.; Mack, D.L.; Moreno, C.M.; Strande, J.L.; Mathieu, J.; Shi, Y.; Markert, C.D.; Wang, Z.; Liu, G.; Lawlor, M.W.; Moorefield, E.C.; Jones, T.N.; Fugate, J.A.; Furth, M.E.; Murry, C.E.; Ruohola-Baker, H.; Zhang, Y.; Santana, L.F.; Childers, M.K. Dystrophin-deficient cardiomyocytes derived from human urine: new biologic reagents for drug discovery. Stem Cell Res 2014, 12, 467–480. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, Y.; Li, S.; Xue, H.; Schmitt, K.; Hergenroeder, G.W.; Wu, J.; Zhang, Y.; Kim, D.H.; Cao, Q. Human neural progenitors derived from integration-free iPSCs for SCI therapy. Stem Cell Res 2017, 19, 55–64. [Google Scholar] [CrossRef]
- Zhou, T.; Benda, C.; Dunzinger, S.; Huang, Y.; Ho, J.C.; Yang, J.; Wang, Y.; Zhang, Y.; Zhuang, Q.; Li, Y.; Bao, X.; Tse, H.F.; Grillari, J.; Grillari-Voglauer, R.; Pei, D.; Esteban, M.A. Generation of human induced pluripotent stem cells from urine samples. Nat Protoc 2012, 7, 2080–2089. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Benda, C.; Duzinger, S.; Huang, Y.; Li, X.; Li, Y.; Guo, X.; Cao, G.; Chen, S.; Hao, L.; Chan, Y.C.; Ng, K.M.; Ho, J.C.; Wieser, M.; Wu, J.; Redl, H.; Tse, H.F.; Grillari, J.; Grillari-Voglauer, R.; Pei, D.; Esteban, M.A. Generation of induced pluripotent stem cells from urine. J Am Soc Nephrol 2011, 22, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, S.; Liu, G.; Shi, Y.; Markert, C.; Andersson, K.E.; Atala, A.; Zhang, Y. Characterization of urine-derived stem cells obtained from upper urinary tract for use in cell-based urological tissue engineering. Tissue Eng Part A 2011, 17, 2123–2132. [Google Scholar] [CrossRef]
- Bharadwaj, S.; Liu, G.; Shi, Y.; Wu, R.; Yang, B.; He, T.; Fan, Y.; Lu, X.; Zhou, X.; Liu, H.; Atala, A.; Rohozinski, J.; Zhang, Y. Multipotential differentiation of human urine-derived stem cells: potential for therapeutic applications in urology. Stem Cells 2013, 31, 1840–1856. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wei, G.; Li, P.; Zhou, X.; Zhang, Y. Urine-derived stem cells: A novel and versatile progenitor source for cell-based therapy and regenerative medicine. Genes Dis. 2014, 1, 8–17. [Google Scholar] [CrossRef]
- Dong, X.; Zhang, T.; Liu, Q.; Zhu, J.; Zhao, J.; Li, J.; Sun, B.; Ding, G.; Hu, X.; Yang, Z.; Zhang, Y.; Li, L. Beneficial effects of urine-derived stem cells on fibrosis and apoptosis of myocardial, glomerular and bladder cells. Mol Cell Endocrinol 2016, 427, 21–32. [Google Scholar] [CrossRef]
- Guo, H.; Deng, N.; Dou, L.; Ding, H.; Criswell, T.; Atala, A.; Furdui, C.M.; Zhang, Y. 3-D Human Renal Tubular Organoids Generated from Urine-Derived Stem Cells for Nephrotoxicity Screening. ACS Biomaterials Science & Engineering 2020. [Google Scholar]
- Yang, Q.; Chen, W.; Han, D.; Zhang, C.; Xie, Y.; Sun, X.; Liu, G.; Deng, C. Intratunical injection of human urine-derived stem cells derived exosomes prevents fibrosis and improves erectile function in a rat model of Peyronie’s disease. Andrologia 2020, e13831. [Google Scholar] [CrossRef]
- Zhou, C.; Wu, X.-R.; Liu, H.-S.; Liu, X.-H.; Liu, G.-H.; Zheng, X.-B.; Hu, T.; Liang, Z.-X.; He, X.-W.; Wu, X.-J.; Smith, L.C.; Zhang, Y.; Lan, P. Immunomodulatory Effect of Urine-derived Stem Cells on Inflammatory Bowel Diseases via Downregulating Th1/Th17 Immune Responses in a PGE2-dependent Manner. J. Crohn’s Colitis 2019, 14, 654–668. [Google Scholar] [CrossRef]
- Liu, G.; Wu, R.; Yang, B.; Shi, Y.; Deng, C.; Atala, A.; Mou, S.; Criswell, T.; Zhang, Y. A cocktail of growth factors released from a heparin hyaluronic-acid hydrogel promotes the myogenic potential of human urine-derived stem cells in vivo. Acta Biomater 2020, 107, 50–64. [Google Scholar] [CrossRef]
- Yang, Q.; Chen, X.; Zheng, T.; Han, D.; Zhang, H.; Shi, Y.; Bian, J.; Sun, X.; Xia, K.; Liang, X.; Liu, G.; Zhang, Y.; Deng, C. Transplantation of Human Urine-Derived Stem Cells Transfected with Pigment Epithelium-Derived Factor to Protect Erectile Function in a Rat Model of Cavernous Nerve Injury. Cell Transpl. 2016, 25, 1987–2001. [Google Scholar] [CrossRef]
- Xiong, G.; Tao, L.; Ma, W.J.; Gong, M.J.; Zhao, L.; Shen, L.J.; Long, C.L.; Zhang, D.Y.; Zhang, Y.Y.; Wei, G.H. Urine-derived stem cells for the therapy of diabetic nephropathy mouse model. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1316–1324. [Google Scholar]
- Sun, B.; Luo, X.; Yang, C.; Liu, P.; Yang, Y.; Dong, X.; Yang, Z.; Xu, J.; Zhang, Y.; Li, L. Therapeutic Effects of Human Urine-Derived Stem Cells in a Rat Model of Cisplatin-Induced Acute Kidney Injury In Vivo and In Vitro. Stem Cells Int 2019, 2019, 8035076. [Google Scholar] [CrossRef]
- Zhu, Q.; Li, Q.; Niu, X.; Zhang, G.; Ling, X.; Zhang, J.; Wang, Y.; Deng, Z. Extracellular Vesicles Secreted by Human Urine-Derived Stem Cells Promote Ischemia Repair in a Mouse Model of Hind-Limb Ischemia. Cell Physiol Biochem 2018, 47, 1181–1192. [Google Scholar] [CrossRef]
- Zhang, Y.; Tian, H.; Bharadwaj, S.; Liu, Y.; Hodges, S.; Atala, A. Urine-derived stem cells for urological injection therapy. J. Urol. 2009, 181, 546. [Google Scholar] [CrossRef]
- Zhang, Y.; Niu, X.; Dong, X.; Wang, Y.; Li, H. Bioglass enhanced wound healing ability of urine-derived stem cells through promoting paracrine effects between stem cells and recipient cells. J Tissue Eng Regen Med 2018, 12, e1609–e22. [Google Scholar] [CrossRef]
- Zhang, C.; George, S.K.; Wu, R.; Thakker, P.U.; Abolbashari, M.; Kim, T.-H.; Ko, I.K.; Zhang, Y.; Sun, Y.; Jackson, J.; Lee, S.J.; Yoo, J.J.; Atala, A. Reno-protection of Urine-derived Stem Cells in A Chronic Kidney Disease Rat Model Induced by Renal Ischemia and Nephrotoxicity. Int. J. Biol. Sci. 2020, 16, 435–446. [Google Scholar] [CrossRef]
- Yang, H.; Chen, B.; Deng, J.; Zhuang, G.; Wu, S.; Liu, G.; Deng, C.; Yang, G.; Qiu, X.; Wei, P.; Wang, X.; Zhang, Y. Characterization of rabbit urine-derived stem cells for potential application in lower urinary tract tissue regeneration. Cell Tissue Res 2018, 374, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Liu, Y.; Bharadwaj, S.; Atala, A.; Zhang, Y. Human urine-derived stem cells seeded in a modified 3D porous small intestinal submucosa scaffold for urethral tissue engineering. Biomaterials 2011, 32, 1317–1326. [Google Scholar] [CrossRef]
- Wu, S.; Liu, G.; Bharadwaj, S.; Hoagie, S.; Atala, A.; Zhang, Y. 181 CELL THERAPY WITH AUTOLOGOUS URINE-DERIVED STEM CELLS FOR VESICOURETERAL REFLUX. J. Urol. 2011, 185, e75. [Google Scholar] [CrossRef]
- Wu, R.P.; Liu, G.; Shi, Y.A.; Bharadwaj, S.; Atala, A.; Zhang, Y. Human urine-derived stem cells originate from parietal stem cells. J Urol 2014, 191, e1–e958. [Google Scholar] [CrossRef]
- Wan, Q.; Xiong, G.; Liu, G.; Shupe, T.D.; Wei, G.; Zhang, D.; Liang, D.; Lu, X.; Atala, A.; Zhang, Y. Urothelium with barrier function differentiated from human urine-derived stem cells for potential use in urinary tract reconstruction. Stem Cell Res Ther 2018, 9, 304. [Google Scholar] [CrossRef] [PubMed]
- Tran CT, A.; Yi, H.; Balog, B.; Zhang, Y.; Damaser, M. : Human urine-derived stem cells or their secretome alone facilitate functional recovery in a rat model of stress urinary incontinence. Journal of Urology 2016, 195, No. [Google Scholar] [CrossRef]
- Tran CNT, A.; Balog, B.; Zhang, Y.; Damaser, M. Paracrine Effects of Human Urine-derived Stem Cells in Treatment of Female Stress Urinary Incontinence in a Rodent Model. Tissue Eng Part A 2015, 21, S-385. [Google Scholar]
- Qin, H.; Zhu, C.; An, Z.; Jiang, Y.; Zhao, Y.; Wang, J.; Liu, X.; Hui, B.; Zhang, X.; Wang, Y. Silver nanoparticles promote osteogenic differentiation of human urine-derived stem cells at noncytotoxic concentrations. Int J Nanomed. 2014, 9, 2469–2478. [Google Scholar] [CrossRef]
- Qin, D.; Long, T.; Deng, J.; Zhang, Y. Urine-derived stem cells for potential use in bladder repair. Stem Cell Res Ther 2014, 5, 69. [Google Scholar] [CrossRef]
- Ouyang, B.; Sun, X.; Han, D.; Chen, S.; Yao, B.; Gao, Y.; Bian, J.; Huang, Y.; Zhang, Y.; Wan, Z.; Yang, B.; Xiao, H.; Songyang, Z.; Liu, G.; Zhang, Y.; Deng, C. Human urine-derived stem cells alone or genetically-modified with FGF2 Improve type 2 diabetic erectile dysfunction in a rat model. PLoS One 2014, 9, e92825. [Google Scholar] [CrossRef]
- Liu, G.; Pareta, R.A.; Wu, R.; Shi, Y.; Zhou, X.; Liu, H.; Deng, C.; Sun, X.; Atala, A.; Opara, E.C.; Zhang, Y. Skeletal myogenic differentiation of urine-derived stem cells and angiogenesis using microbeads loaded with growth factors. Biomaterials 2013, 34, 1311–1326. [Google Scholar] [CrossRef]
- Long TJ, Y.D.; Shi, H.; Zhong, L.R.; Zhang, D.Y.; Li, W.; Zhang, Y.; Chen, D.; Jiao, Y.; Diz, D.; Zhang, Y. : Human urine-derived stem cells lessen inflammation and fibrosis within kidney tissue in a rodent model of aging-related renal insufficiency. J Urol 2017, 197, e293. [Google Scholar] [CrossRef]
- Ling, X.; Zhang, G.; Xia, Y.; Zhu, Q.; Zhang, J.; Li, Q.; Niu, X.; Hu, G.; Yang, Y.; Wang, Y.; Deng, Z. Exosomes from human urine-derived stem cells enhanced neurogenesis via miR-26a/HDAC6 axis after ischaemic stroke. J Cell Mol Med 2020, 24, 640–654. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Luo, H.; Dong, X.; Liu, Q.; Wu, C.; Zhang, T.; Hu, X.; Zhang, Y.; Song, B.; Li, L. Therapeutic effect of urine-derived stem cells for protamine/lipopolysaccharide-induced interstitial cystitis in a rat model. Stem Cell Res Ther 2017, 8, 107. [Google Scholar] [CrossRef]
- Lang, R.; Liu, G.; Shi, Y.; Bharadwaj, S.; Leng, X.; Zhou, X.; Liu, H.; Atala, A.; Zhang, Y. Self-renewal and differentiation capacity of urine-derived stem cells after urine preservation for 24 hours. PLoS One 2013, 8, e53980. [Google Scholar] [CrossRef]
- Guan, J.; Zhang, J.; Guo, S.; Zhu, H.; Zhu, Z.; Li, H.; Wang, Y.; Zhang, C.; Chang, J. Human urine-derived stem cells can be induced into osteogenic lineage by silicate bioceramics via activation of the Wnt/beta-catenin signaling pathway. Biomaterials 2015, 55, 1–11. [Google Scholar] [CrossRef]
- Fu, Y.; Guan, J.; Guo, S.; Guo, F.; Niu, X.; Liu, Q.; Zhang, C.; Nie, H.; Wang, Y. Human urine-derived stem cells in combination with polycaprolactone/gelatin nanofibrous membranes enhance wound healing by promoting angiogenesis. J Transl Med 2014, 12, 274. [Google Scholar] [CrossRef]
- Bodin, A.; Bharadwaj, S.; Wu, S.; Gatenholm, P.; Atala, A.; Zhang, Y. Tissue-engineered conduit using urine-derived stem cells seeded bacterial cellulose polymer in urinary reconstruction and diversion. Biomaterials 2010, 31, 8889–8901. [Google Scholar] [CrossRef]
- Sagrinati, C.; Netti, G.S.; Mazzinghi, B.; Lazzeri, E.; Liotta, F.; Frosali, F.; Ronconi, E.; Meini, C.; Gacci, M.; Squecco, R.; Carini, M.; Gesualdo, L.; Francini, F.; Maggi, E.; Annunziato, F.; Lasagni, L.; Serio, M.; Romagnani, S.; Romagnani, P. Isolation and characterization of multipotent progenitor cells from the Bowman’s capsule of adult human kidneys. J Am Soc Nephrol 2006, 17, 2443–2456. [Google Scholar] [CrossRef]
- Shrestha, S.; Somji, S.; Sens, D.A.; Slusser-Nore, A.; Patel, D.H.; Savage, E.; Garrett, S.H. Human renal tubular cells contain CD24/CD133 progenitor cell populations: Implications for tubular regeneration after toxicant induced damage using cadmium as a model. Toxicol. Appl. Pharmacol. 2017, 331, 116–129. [Google Scholar] [CrossRef]
- Huang, J.; Kong, Y.; Xie, C.; Zhou, L. Stem/progenitor cell in kidney: characteristics, homing, coordination, and maintenance. Stem Cell Res Ther 2021, 12, 197. [Google Scholar] [CrossRef] [PubMed]
- Al-Marsoummi, S.; Singhal, S.; Garrett, S.H.; Somji, S.; Sens, D.A.; Singhal, S.K. CD133+CD24+ Renal Tubular Progenitor Cells Drive Hypoxic Injury Recovery via Hypoxia-Inducible Factor-1A and Epidermal Growth Factor Receptor Expression. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef] [PubMed]
- Surendran, H.; Soundararajan, L.; Reddy, K.V.; Subramani, J.; Stoddard, J.; Reynaga, R.; Tschetter, W.; Ryals, R.C.; Pal, R. An improved protocol for generation and characterization of human-induced pluripotent stem cell-derived retinal pigment epithelium cells. STAR Protoc 2022, 3, 101803. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Yuan, C.; Wu, X. The recruitment mechanisms and potential therapeutic targets of podocytes from parietal epithelial cells. J. Transl. Med. 2021, 19, 441. [Google Scholar] [CrossRef]
- [62] Hong, X.; Nie, H.; Deng, J.; Liang, S.; Chen, L.; Li, J.; Gong, S.; Wang, G.; Zuo, W.; Hou, F.; Zhang, F. WT1(+) glomerular parietal epithelial progenitors promote renal proximal tubule regeneration after severe acute kidney injury. Theranostics 2023, 13, 1311–1324. [Google Scholar] [CrossRef]
- Bronstein, R.; Pace, J.; Gowthaman, Y.; Salant, D.J.; Mallipattu, S.K. Podocyte-Parietal Epithelial Cell Interdependence in Glomerular Development and Disease. J Am Soc Nephrol 2023, 34, 737–750. [Google Scholar] [CrossRef] [PubMed]
- D’Agati, V.D.; Shankland, S.J. Recognizing diversity in parietal epithelial cells. Kidney Int 2019, 96, 16–19. [Google Scholar] [CrossRef]
- Ronconi, E.; Sagrinati, C.; Angelotti, M.L.; Lazzeri, E.; Mazzinghi, B.; Ballerini, L.; Parente, E.; Becherucci, F.; Gacci, M.; Carini, M. Regeneration of glomerular podocytes by human renal progenitors. J. Am. Soc. Nephrol. : JASN 2009, 20, 322. [Google Scholar] [CrossRef] [PubMed]
- Bates, C.; Ho, J.; Sims-Lucas, S. Embryonic Development of the Kidney. Pediatric Nephrology. Edited by Avner ED, Harmon WE, Niaudet P, Yoshikawa N, Emma F, Goldstein SL. Berlin, Heidelberg: Springer Berlin Heidelberg, 2016. pp. 3–36.
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J Clin Invest 2009, 119, 1420–1428. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, G.; Wu, R.; Mack, D.L.; Sun, X.S.; Maxwell, J.; Guan, X.; Atala, A.; Zhang, Y. Differentiation Capacity of Human Urine-Derived Stem Cells to Retain Telomerase Activity. Front Cell Dev Biol 2022, 10, 890574. [Google Scholar] [CrossRef]
- Wan, Q.; Xiong, G.; Liu, G.; Shupe, T.D.; Wei, G.; Zhang, D.; Liang, D.; Lu, X.; Atala, A.; Zhang, Y. Urothelium with barrier function differentiated from human urine-derived stem cells for potential use in urinary tract reconstruction. Stem Cell Res. Ther. 2018, 9, 304. [Google Scholar] [CrossRef]
- Yin, X.; Li, Q.; McNutt, P.M.; Zhang, Y. Urine-Derived Stem Cells for Epithelial Tissues Reconstruction and Wound Healing. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Zhang, W.; Hu, J.; Huang, Y.; Wu, C.; Xie, H. Urine-derived stem cells: applications in skin, bone and articular cartilage repair. Burns Trauma 2021, 9, tkab039. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wu, R.; Yang, B.; Deng, C.; Lu, X.; Walker, S.J.; Ma, P.X.; Mou, S.; Atala, A.; Zhang, Y. Human Urine-Derived Stem Cell Differentiation to Endothelial Cells with Barrier Function and Nitric Oxide Production. Stem Cells Transl. Med. 2018, 7, 686–698. [Google Scholar] [CrossRef]
- Yi, H.; Chen, G.; Qiu, S.; Maxwell, J.T.; Lin, G.; Criswell, T.; Zhang, Y. Urine-derived stem cells genetically modified with IGF1 improve muscle regeneration. Am J Clin Exp Urol 2024, 12, 64–87. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Zhu, Y.; Kim, M.; Zhao, G.; Wang, Y.; Collins, C.P.; Mei, O.; Zhang, Y.; Duan, C.; Zhong, J.; Zhang, H.; You, W.; Shen, G.; Luo, C.; Wu, X.; Li, J.; Shu, Y.; Luu, H.H.; Haydon, R.C.; Lee, M.J.; Shi, L.L.; Huang, W.; Fan, J.; Sun, C.; Wen, L.; Ameer, G.A.; He, T.C.; Reid, R.R. Effective Bone Tissue Fabrication Using 3D-Printed Citrate-Based Nanocomposite Scaffolds Laden with BMP9-Stimulated Human Urine Stem Cells. ACS Appl Mater Interfaces 2025, 17, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Xing, F.; Zou, M.; Gong, M.; Li, L.; Xiang, Z. Comparison of chondrogenesis-related biological behaviors between human urine-derived stem cells and human bone marrow mesenchymal stem cells from the same individual. Stem Cell Res. Ther. 2021, 12, 366. [Google Scholar] [CrossRef] [PubMed]
- Baliña-Sánchez, C.; Aguilera, Y.; Adán, N.; Sierra-Párraga, J.M.; Olmedo-Moreno, L.; Panadero-Morón, C.; Cabello-Laureano, R.; Márquez-Vega, C.; Martín-Montalvo, A.; Capilla-González, V. Generation of mesenchymal stromal cells from urine-derived iPSCs of pediatric brain tumor patients. Front Immunol 2023, 14, 1022676. [Google Scholar] [CrossRef]
- Woogeng, I.N.; Kaczkowski, B.; Abugessaisa, I.; Hu, H.; Tachibana, A.; Sahara, Y.; Hon, C.-C.; Hasegawa, A.; Sakai, N.; Nishida, M.; Sanyal, H.; Sho, J.; Kajita, K.; Kasukawa, T.; Takasato, M.; Carninci, P.; Maeda, A.; Mandai, M.; Arner, E.; Takahashi, M.; Kime, C. Inducing human retinal pigment epithelium-like cells from somatic tissue. Stem Cell Rep. 2022, 17, 289–306. [Google Scholar] [CrossRef]
Figure 1.
Morphology of Urine-Derived Stem Cells. (a) Initial stage: single USC at passage 0 (p0) Day 3; (b) Clone formation: USC colony at p0 Day 7; (c) Expansion phase: USCs at p0 Day 10; (d) Subsequent passages: USCs exhibiting characteristic “grain-of-rice” appearance.
Figure 1.
Morphology of Urine-Derived Stem Cells. (a) Initial stage: single USC at passage 0 (p0) Day 3; (b) Clone formation: USC colony at p0 Day 7; (c) Expansion phase: USCs at p0 Day 10; (d) Subsequent passages: USCs exhibiting characteristic “grain-of-rice” appearance.
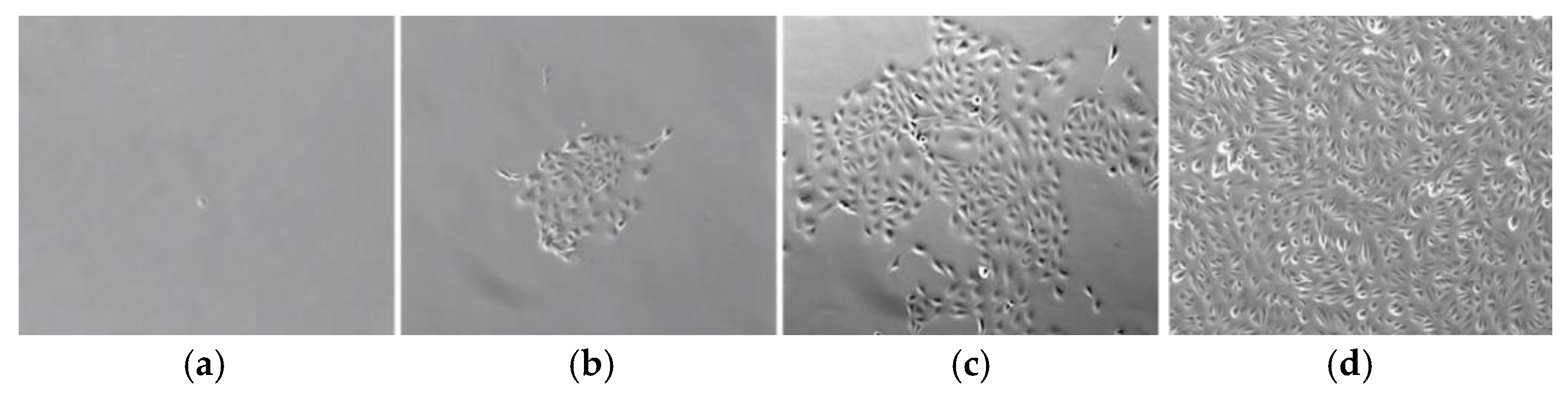
Figure 2.
Immunophenotypic Profile of USCs. Flow cytometric analysis of USCs demonstrated robust expression of characteristic renal progenitor markers CD24 and CD133, MSC surface markers, including strong positive for CD29, CD44, CD73, CD90, CD146, and SSEA-4, moderate positive for CD105, CG117, and Strol-1. Significantly. These cells were negative for hematopoietic lineage markers CD14, CD31, CD34, CD45 and VEGFR2, confirming their non-hematopoietic origin and MSC identity.
Figure 2.
Immunophenotypic Profile of USCs. Flow cytometric analysis of USCs demonstrated robust expression of characteristic renal progenitor markers CD24 and CD133, MSC surface markers, including strong positive for CD29, CD44, CD73, CD90, CD146, and SSEA-4, moderate positive for CD105, CG117, and Strol-1. Significantly. These cells were negative for hematopoietic lineage markers CD14, CD31, CD34, CD45 and VEGFR2, confirming their non-hematopoietic origin and MSC identity.
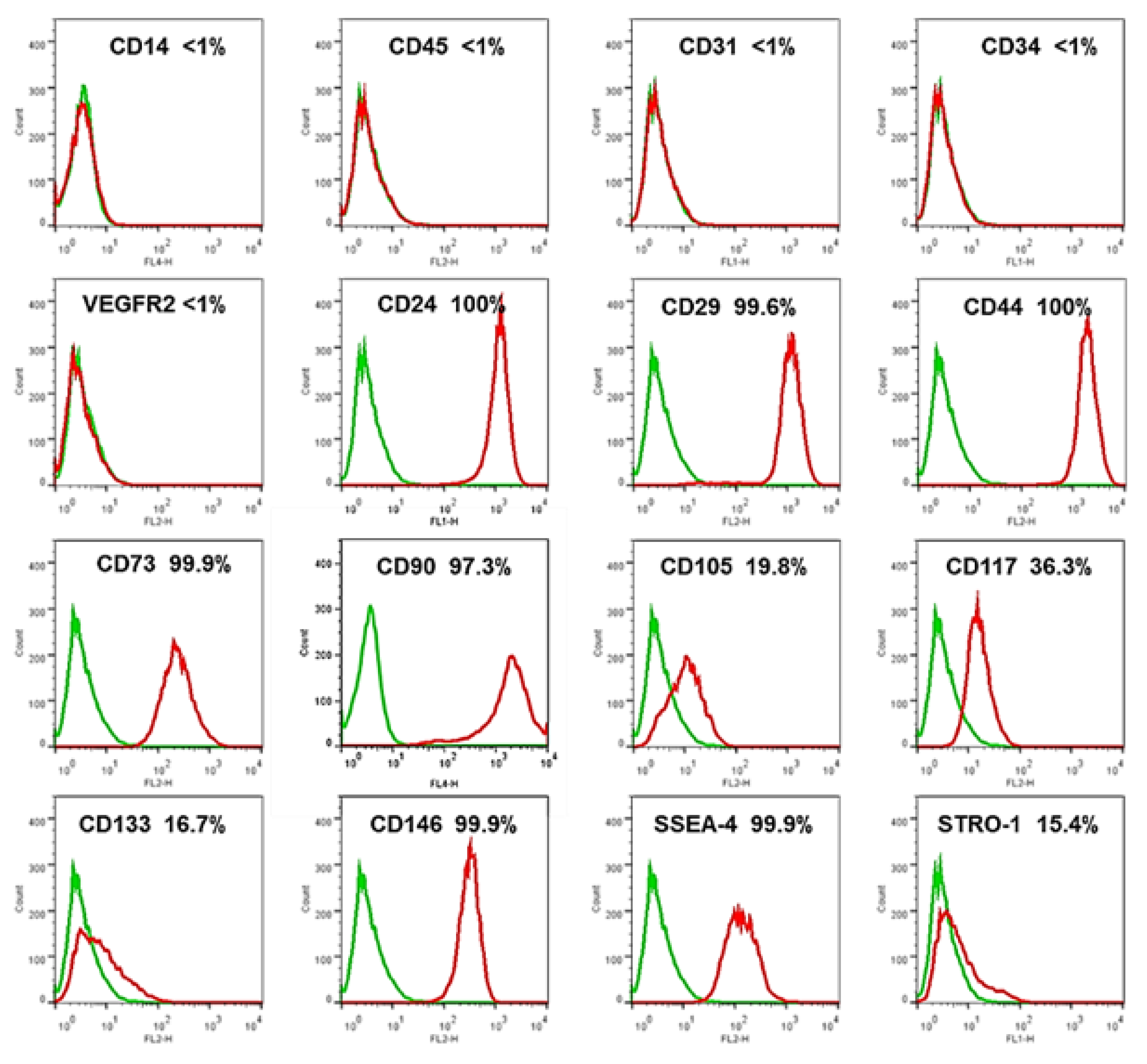
Figure 3.
Morphology of Urine-Derived iPSC Colonies. (a) Urine-derived stem cells (USCs) at passage 3 (p3) before Sendai virus (SeV) transfection, serving as a control; (b) Early u-iPSC aggregates on Day 5 after SeV transfection. (c) u-iPSC colony at passage 9 (p9) Day 3.
Figure 3.
Morphology of Urine-Derived iPSC Colonies. (a) Urine-derived stem cells (USCs) at passage 3 (p3) before Sendai virus (SeV) transfection, serving as a control; (b) Early u-iPSC aggregates on Day 5 after SeV transfection. (c) u-iPSC colony at passage 9 (p9) Day 3.
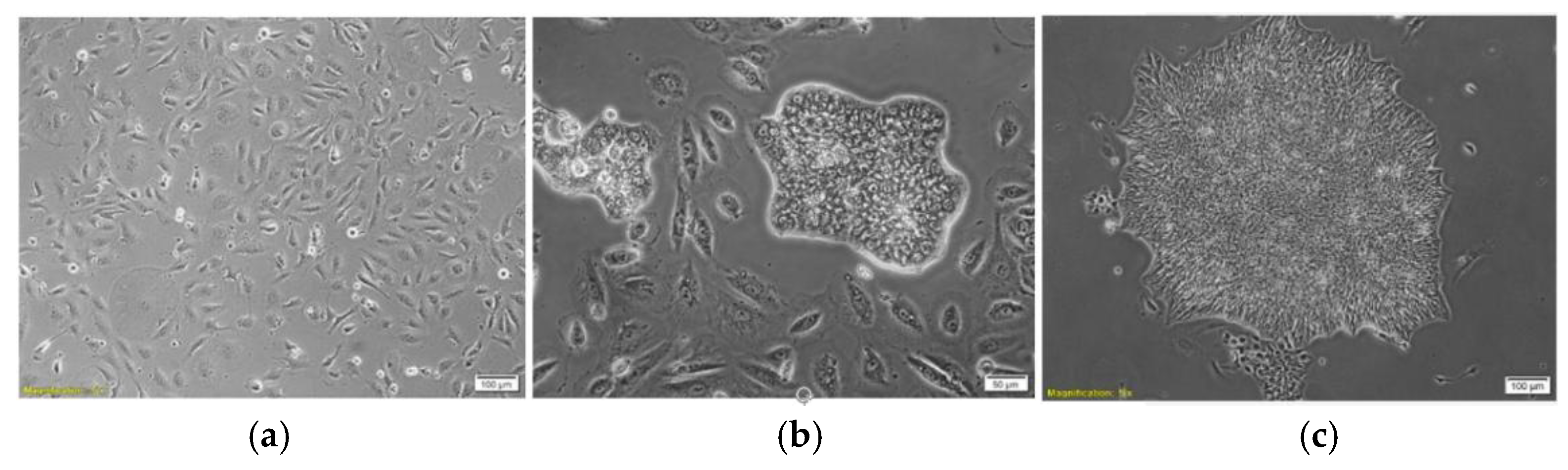
Figure 4.
Loss of Sendai Viral Protein Expression in u-iPSC Lines. (a) Phase contrast of u-iPSC at p6; (b) Immunostaining with anti-SeV antibody showing absence of viral protein expression (c) Merged image of anti-SeV antibody staining with DAPI counterstain.
Figure 4.
Loss of Sendai Viral Protein Expression in u-iPSC Lines. (a) Phase contrast of u-iPSC at p6; (b) Immunostaining with anti-SeV antibody showing absence of viral protein expression (c) Merged image of anti-SeV antibody staining with DAPI counterstain.

Figure 5.
Pluripotent State of Urine-Derived iPSC. (a) Immunofluorescence staining of u-iPSC at p12 with key pluripotency marker TRA-1-60; (b) Double staining for TRA-1-60 and DAPI (nuclear stain), positively identifying the iPSC colony.
Figure 5.
Pluripotent State of Urine-Derived iPSC. (a) Immunofluorescence staining of u-iPSC at p12 with key pluripotency marker TRA-1-60; (b) Double staining for TRA-1-60 and DAPI (nuclear stain), positively identifying the iPSC colony.
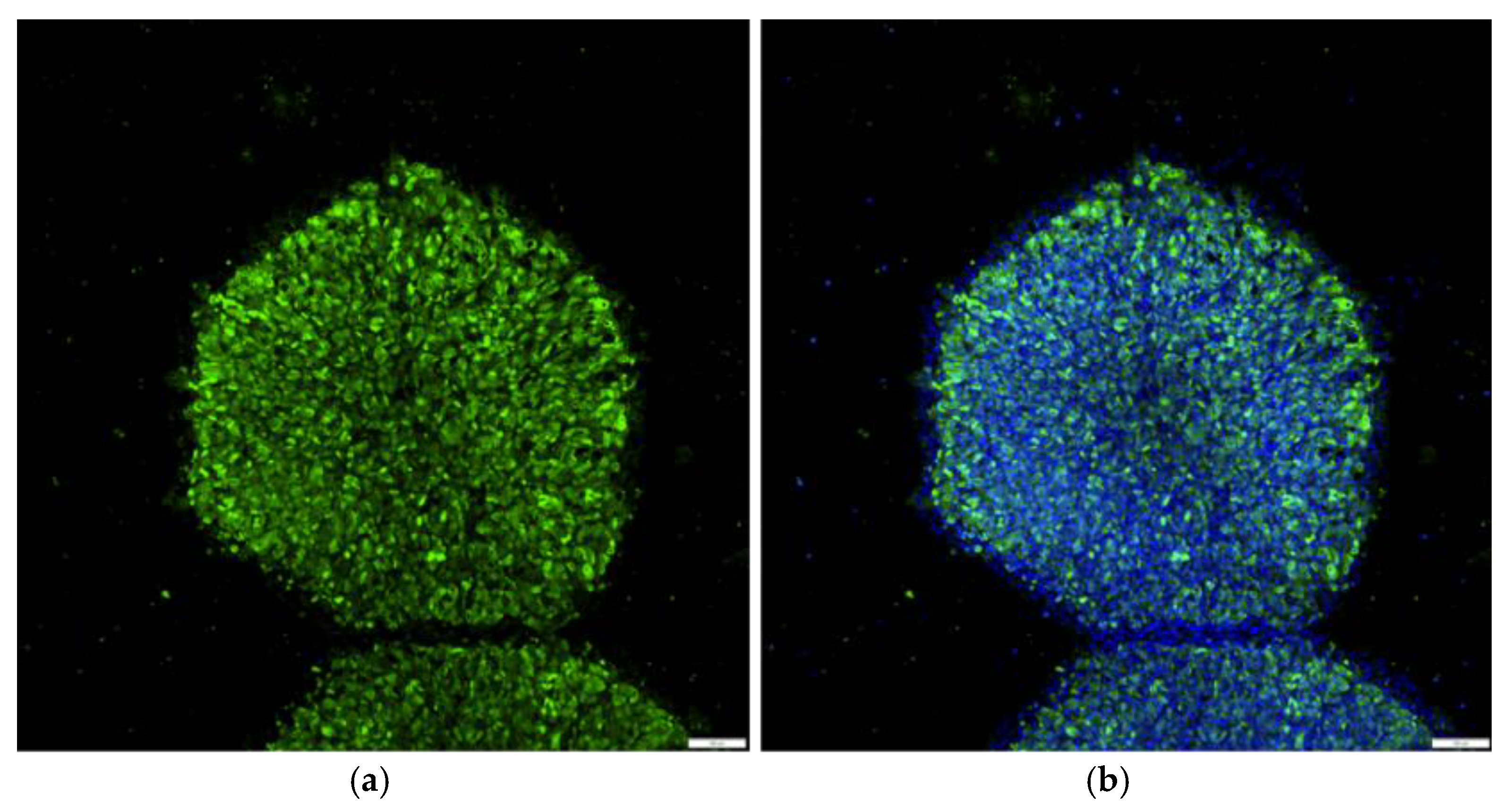
Figure 6.
Characterization of Pluripotency Markers in u-iPSC Lines. Immunofluorescence staining of u-iPSC displaying prominent nuclear expression of transcription factors SOX2, OCT4, and NANOG, along with strong surface expression of SSEA4.
Figure 6.
Characterization of Pluripotency Markers in u-iPSC Lines. Immunofluorescence staining of u-iPSC displaying prominent nuclear expression of transcription factors SOX2, OCT4, and NANOG, along with strong surface expression of SSEA4.
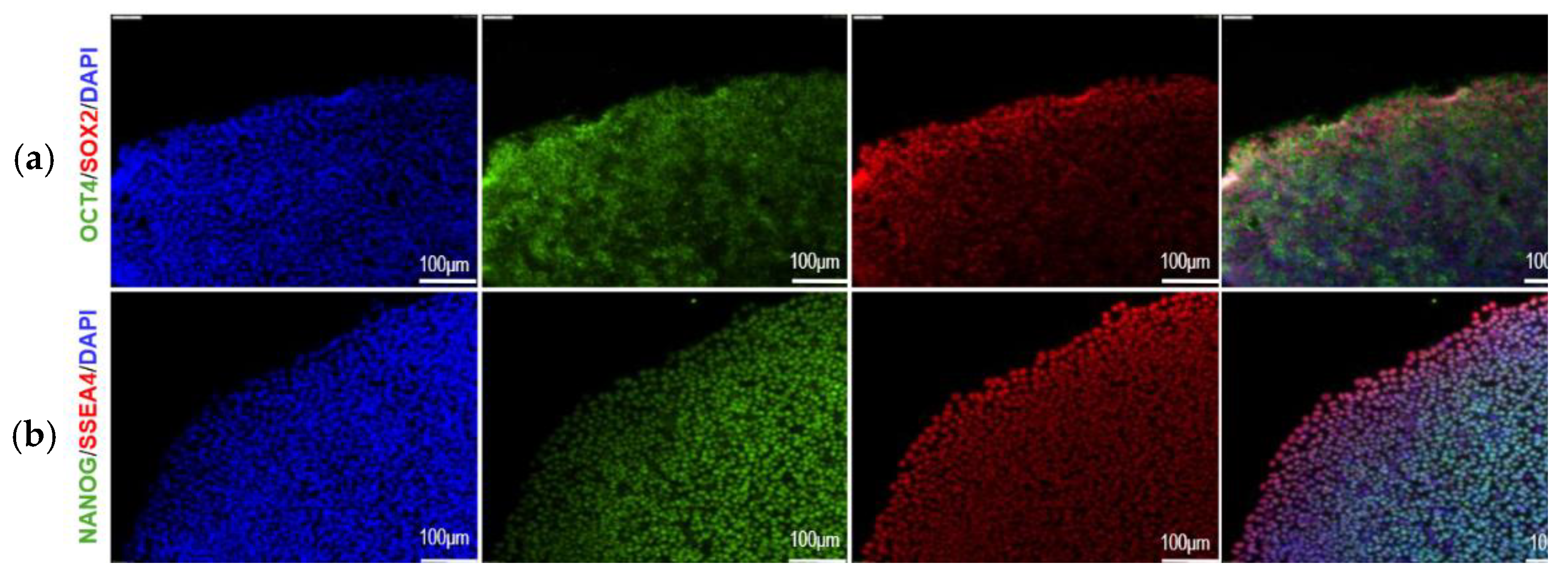
Figure 7.
Progressive Morphological Changes of u-iPSC-RPE. (a) Embryoid bodies (EBs) 3 days post-differentiation; (b) RPE with characteristic hexagonal shape in cobblestone-like arrangements 60 days post-differentiation; (c) Pigmented RPE with increased cellular density and maturation.
Figure 7.
Progressive Morphological Changes of u-iPSC-RPE. (a) Embryoid bodies (EBs) 3 days post-differentiation; (b) RPE with characteristic hexagonal shape in cobblestone-like arrangements 60 days post-differentiation; (c) Pigmented RPE with increased cellular density and maturation.

Figure 8.
Progressive Morphological Changes and RPE Marker Expression of u-iPSC-RPE. Immunofluorescence analysis of u-iPSC-RPE post-differentiation showing protein expression and localization of RPE commitment marker MITF, RPE maturation marker RPE65 and tight junction marker ZO1.
Figure 8.
Progressive Morphological Changes and RPE Marker Expression of u-iPSC-RPE. Immunofluorescence analysis of u-iPSC-RPE post-differentiation showing protein expression and localization of RPE commitment marker MITF, RPE maturation marker RPE65 and tight junction marker ZO1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



