Submitted:
24 August 2025
Posted:
25 August 2025
You are already at the latest version
Abstract
Psoriasis is a chronic inflammatory autoimmune skin disease characterized by erythematous plaques covered with silvery-white scales, often accompanied by systemic complications such as psoriatic arthritis and cardiovascular diseases. The occurrence and progression of this disease are related to the IL-23/IL-17 axis and involve aberrant activation and interactions of multiple immune cells, including keratinocytes, dendritic cells, Th17 cells, and neutrophils, with genetic predispositions and environmental triggers. Currently, psoriasis remains incurable. Therapeutic strategies that primarily focus on immunosuppression, anti-inflammation, and palliative care can only control and delay disease progression. These strategies face challenges relating to cost, safety, efficacy and precision of targeting. Future research should prioritize individualized treatment, immune tolerance induction, and microbiome modulation to achieve long-term disease control and potential cure. This review is primarily focused on the progress and prospects of the pathogenesis and therapies for psoriasis, aiming to provide new insights for individualized treatment in the future.
Keywords:
psoriasis
; IL-23/IL-17 axis
; biologicals
; targeted therapy
; CAR-T cell therapy
1. Introduction
The erroneous attack and destruction of self-tissue or -organ by the immune system can result in autoimmune diseases. To date, over 150 distinct autoimmune disorders have been documented, including psoriasis, type 1 diabetes mellitus, multiple sclerosis, systemic lupus erythematosus, rheumatoid arthritis (RA), and so on [1]. Psoriasis is a persistent, recurrent, immune-mediated inflammatory dermatosis that can affect individuals across diverse geographic regions, ethnicities, and age groups globally [2]. Psoriasis typically presents clinically as well-demarcated erythematous plaques or papules covered with characteristic silvery-white scales, frequently accompanied by marked pruritus, pain, and other distressing symptoms. As a systemic inflammatory disorder, psoriasis manifests not only as cutaneous lesions but is also associated with multiple comorbidities, including psoriatic arthritis [3], metabolic syndrome [4,5] and depression [6]. These multisystem complications severely affect patients’ quality of life and their social activities.
The pathogenesis of psoriasis involves multiple cell types and cytokines as well as both genetic predisposition and environmental triggers. Current evidence indicates interleukin (IL)-23/IL-17 axis as the central driver of psoriatic inflammation [7,8,9]. Cytokines such as IL-17A and IL-23 induce keratinocyte proliferation and inflammatory responses, leading to the chronic and relapsing features of psoriasis [10,11]. In addition, genetic factors are al
o important in the development of psoriasis. Studies have identified 109 distinct psoriasis susceptibility loci [12]. Environmental factors such as infections, stress, smoking, and climate changes are also considered important triggers of psoriasis [10,13]. Microbial infections, such as those caused by streptococci and human immunodeficiency virus (HIV), can trigger or exacerbate psoriasis through mechanisms involving superantigens or epitope spreading [14,15]. Furthermore, lifestyle factors such as psychological stress and sleep deprivation can exacerbate psoriasis symptoms by affecting the immune system and skin barrier function.
Currently, the treatment strategies for psoriasis primarily include topical therapy, systemic therapy, and biologic therapy. Conventional topical treatments commonly involve corticosteroids and vitamin D3 analogs. Corticosteroids can rapidly control symptoms, but prolonged use may lead to adverse effects [16], whereas vitamin D3 analogs are relatively safer but exhibit a slower onset of action. Conventional systemic therapies encompass phototherapy, methotrexate, and cyclosporine. Phototherapy is widely accessible and cost-effective, yet prolonged use of this therapy poses risks of photoaging. Methotrexate and cyclosporine demonstrate significant efficacy in moderate-to-severe psoriasis, but their use requires careful monitoring due to potential hepatotoxicity, nephrotoxicity, and immunosuppressive effects [17,18]. Recent advances in biologic agents, particularly monoclonal antibodies against IL-17 and IL-23, have demonstrated significant clinical efficacy and a favorable safety profile in psoriasis treatment. However, their long-term use may be associated with increased risks of drug resistance and infections [19,20]. Moreover, the advent of innovative cellular therapies, such as CAR-T therapy, hold much promise for the treatment and control of psoriasis. This review mainly covers pathological features, advances in the pathogenesis and therapeutics of psoriasis, and future trends in autoimmune disease therapy. We also discuss on novel individualized precision treatment and eradication strategies in the future.
2. Epidemiology and Disease Burden
Psoriasis is a globally distributed disease [21]. The worldwide prevalence of psoriasis exhibits significant geographical variation, ranging from 11.4% [22] to 0.09% [23]. It was reported that the global prevalence of psoriasis was increasing [24,25], which might be associated with the misuse of antimicrobial drugs and environmental pollution. An UK-based study revealed that individuals with elevated genetic susceptibility who were exposed to nitrogen dioxide (NO2), nitrogen oxides (NOₓ), and particular materials under 10 µm (PM10) showed significantly increased risk of psoriasis development [26]. There is no significant difference between genders in the prevalence of psoriasis [27].
Psoriasis induces multidimensional health impairments, including dermatological-articular comorbidities [3], Crohn’s disease [28], nonalcoholic fatty liver disease [29], cardiovascular disease [5,30], metabolic disorders [31], and so on. The loss rate of disability-adjusted life years (DALYs) has been recorded at up to 1.2%, causing economic burden, mental stress, and even endangering the lives of patients and their families [32,33]. Furthermore, psoriasis can involve the face, hands, and reproductive organs, thereby impairing psychosocial well-being and elevating risks of depression, social withdrawal, and suicidality [34,35]. It has been reported that 43.2% of individuals living with moderate-to-severe plaque psoriasis exhibit depressive symptoms [36].
3. Clinical Presentation
Based on clinical and pathophysiological characteristics, psoriasis is mainly categorized into eight subtypes, such as chronic plaque, guttate psoriasis, erythrodermic psoriasis, psoriatic arthritis (PsA), pustular psoriasis, inverse psoriasis, seborrheic psoriasis, and nail psoriasis.
Chronic plaque psoriasis, accounting for approximately 90% of all psoriasis cases [37,38], initially presents as erythematous papules or plaques that progressively develop into well-demarcated, polymorphic erythematous plaques with thick, silvery-white scales. Upon removal of the outermost scales, stratified scaling (referred to as the “wax-drop phenomenon”) becomes apparent. Further scraping reveals a reddish, glossy, translucent film (also known as the thin-film phenomenon), which can be peeled off to reveal punctate haemorrhages (i.e., Auspitz’s sign, which is due to scraping of dilated, tortuous capillaries at the top of the papillae of the dermis) [39]. The lesions can occur symmetrically on any part of the body, with morphological variations depending on the affected region. Pruritus is a frequently associated symptom.
Guttate psoriasis accounts for about 2% of psoriasis and is common in adolescents, usually following a streptococcal infection of the throat, and presents with numerous scaly papules 0.3 to 0.5 cm in size [40]. The onset of the disease is rapid, with flushing of the skin and varying degrees of itchiness. With appropriate treatment, the symptoms may ameliorate within a few weeks, and in a few patients the disease may become chronic [41].
Erythrodermic psoriasis, a severe and potentially life-threatening variant of psoriasis, accounts for approximately 1–2% of all psoriasis cases. It is characterized by generalized erythema, edema, and prominent scaling, typically affecting >75% of the body surface areas [42]. The disease often follows a prolonged and relapsing course, requiring intensive therapeutic intervention.
PsA, an inflammatory joint disorder associated with psoriasis, may develop concurrently with or subsequent to cutaneous manifestations. Clinical features include joint swelling, pain, and progressive functional impairment, with severe cases leading to joint deformities.
Pustular psoriasis is characterized by the eruption of sterile, superficial micropustules ranging from pinhead to millet-sized (1–3 mm), appearing as yellowish-white to pale yellow lesions. The pustules generally desiccate and crust within 1–2 weeks, followed by spontaneous resolution. However, the disease often exhibits a relapsing-remitting course, with periodic recurrence of pustular.
Inverse psoriasis affects skin folds, such as the buttocks, groin and armpits. The skin lesions are red, shiny and without characteristic scales.
Seborrheic psoriasis occurs in the sebaceous areas of the scalp and face, as well as in the area behind ears or in front of the sternum.
Nail psoriasis manifests itself in a variety of ways, including small pits on the nail plate, nail separation, oil droplets (orange discolouration of the nail bed), or fragmentation of the nail plate. It affects about 40.9% of patients with plaque psoriasis, and patients with nail involvement have a longer duration of disease, higher severity of disease, and a higher frequency of PsA [43].
The diagnosis of psoriasis is primarily determined by the location, type and history of skin injury. Histopathological features have been shown to possess a certain degree of diagnostic value, while skin imaging has been identified as having auxiliary diagnostic value.
Certain diseases exhibit symptoms similar to those of psoriasis and require differential diagnosis. For instance, atopic dermatitis typically occurs in the flexural regions of the body, such as the antecubital and popliteal fossae. The lesions lack a clear demarcation from unaffected skin. Scalp psoriasis should be differentiated from seborrheic dermatitis and tinea capitis. Psoriatic lesions on the trunk and extremities must be distinguished from secondary syphilitic eruptions, lichen planus, and chronic eczema. Psoriatic arthritis requires differentiation from RA in diagnosis. Inverse psoriasis should be differentiated from candidiasis and other fungal infections. Pityriasis rosea can be distinguished from guttate psoriasis by its fine scaling and the presence of a herald patch, which appears 1–2 weeks before the generalized eruption. Additionally, early lesions of cutaneous T-cell lymphoma lack well-defined borders and thick scales, unlike those of psoriasis.
4. Pathophysiology
4.1. Histological features
The histopathological characteristics of psoriasis predominantly consist of epidermal hyperplasia, hyperkeratosis with parakeratosis, presence of Munro microabscesses (focal collections of neutrophils within the epidermis), and marked diminution or complete absence of the granular layer [27]. Psoriasis shows twisted and widened capillaries in the dermal papillae with swollen lining cells and thinning of the suprapapillary epidermal plates. A predominantly T-lymphocytic inflammatory infiltrate with neutrophils is observed in both the dermis and perivascular areas. Erythrodermic psoriasis is characterized by marked dilation and congestion of superficial dermal vasculature, while pustular psoriasis exhibits the pathognomonic Kogoj micropustules—multifocal spongiform pustules containing neutrophil aggregates in the upper spinous layer [44].
4.2. Genetic contributions
Psoriasis demonstrates strong genetic predisposition. Genome-wide association studies (GWAS) have identified 109 distinct susceptibility loci for psoriasis [45,46,47]. Approximately 30% of psoriasis patients report a family history, and the disease concordance rate in monozygotic twins reaches 68% [48], highlighting the significant genetic contribution to psoriasis pathogenesis. Environmental triggers may modulate disease phenotype through immunoregulatory and epigenetic mechanisms [49,50]. HLA-C*06, a human leukocyte antigen (HLA) allele, triggers psoriasis by inducing an autoimmune response against melanocytes through presentation of the “a disintegrin and metalloproteinase with thrombospondin motifs-like 5” (ADAMTSL5) autoantigen [51]. The erap1 gene encodes an interferon (IFN)-γ-induced aminopeptidase that in the endoplasmic reticulum trims precursor peptides of melanocyte autoantigens to the right length for HLA-C*06:02 presentation, activating CD8+ T-cell-mediated autoimmune disease [52]. In addition to the genes previously mentioned, HLA-DRB1*07 [53], il23r [54], ddx58 [55], traf3ip2 [56] and other genes have also been found to be associated with psoriasis [57].
4.3. Triggers
In addition to genetic predisposition, environmental factors, hormonal levels, and lifestyles also contribute to the onset and progression of psoriasis. Microbial infections have been strongly implicated in both disease initiation and exacerbation. For example, infections with Streptococcus pyogenes may trigger or exacerbate psoriasis through superantigen-mediated mechanisms or epitope spreading [58,59]. CD8+ T cells in psoriasis patients cross-recognize streptococcal M proteins and keratin 17 (K17) via molecular mimicry [60,61]. Psoriasis incidence increased in HIV-infected individuals compared to the general population [15]. HIV may directly trigger psoriasis as a source of superantigens or as a co-stimulatory factor for antigen delivery [62], and activated CD8+ T cells produce more IFN-γ during HIV infection [63], or as HIV infection destroys CD4+ T cells, thereby affecting the immunomodulatory role of regulatory T cells (Tregs). In addition to the aforementioned pathogens, intestinal dysbiosis has been consistently associated with increased psoriasis incidence [64,65,66]. 31–88% of psoriasis patients experienced stressful events prior to disease onset, and individuals experiencing significant stress within 12 months demonstrate higher psoriasis incidence [67]. It was proposed that stress may mediate the pathophysiology of psoriasis through the hypothalamic-pituitary-adrenal axis (HPA), the immune pathway and the peripheral nervous system [68].
Fluctuations in estrogen levels can potentially impact the progression of psoriasis. Estrogen may exert anti-psoriatic effects via estrogen receptors α and β (ERα/ERβ)-mediated downregulation of proinflammatory cytokines in immune cells. Paradoxically, under specific conditions, estrogen can promote IL-23 secretion by dendritic cells, potentially exacerbating disease [69,70,71], suggesting that estrogen may play dual regulatory roles on psoriasis in an environmentally dependent manner [70,72]. Lifestyle also influences the course of psoriasis, and emerging evidence indicates that elevated cortisol levels in sleep disorders such as insomnia stimulate cutaneous mast cells, compromise skin barrier function, and upregulate proinflammatory cytokines expression, thereby exacerbating psoriasis [73,74]. Furthermore, research has indicated that individuals with a history of cigarette smoking exhibit a higher incidence of psoriasis compared to the general population [47]. At present, there are still no sufficient evidence to determine whether alcohol consumption is associated with the development and recurrence of psoriasis [75,76,77,78]. Thus, the efforts will be required to investigate the effect of alcohol on psoriasis.
4.4. Pathogenesis
The pathogenesis of psoriasis involves intricate interactions of genetic, immunological, and environmental factors, characterized by the synergistic dysregulation of immune system activation and epidermal cell dysfunction [79,80]. This process is driven by aberrant immune responses, which fuels a self-amplifying cascade of IL-17 and IL-23 secretion, resulting in the hallmark pathological features of chronic inflammation (erythema) and epidermal hyperproliferation (scaling). Upon cutaneous injury or microbial infection, the damage-associated molecular patterns (DAMPs) of host cell and pathogen-associated molecular patterns (PAMPs) of pathogens collectively activate Toll-like receptor (TLR) signaling pathways in various cutaneous cell populations, thereby triggering the cascade release of proinflammatory cytokines that ultimately perpetuate the chronic inflammatory response characteristic of psoriatic pathogenesis [81].
Keratinocytes (KCs), constituting approximately 90% of epidermal cells, exhibit abnormal proliferation and differentiation that directly contribute to psoriatic acanthosis [82,83]. Beyond DAMPs activation, KCs respond to both exogenous antigens and endogenous autoantigens by producing antimicrobial peptides, S100 proteins, and other effector molecules that initiate or amplify innate and adaptive immune responses, thereby participating in psoriatic inflammation [55,84]. Research demonstrates that KC-derived S100A9 induces dendritic cells to produce IL-23, driving the IL-23/IL-17 axis, while IL-17A further stimulates KCs to express IL-25 (also termed IL-17E), which mediates proinflammatory phenotypes and hyperproliferation through STAT3 activation [85]. Additionally, KC-secreted chemokine C-C motif chemokine ligand 20 (CCL20) recruits CD4+ T helper (Th)-17 cells and γδ T cells, exacerbating inflammatory responses [86]. KCs also respond to cytokines such as IFN-γ, IL-17, and IL-36, establishing positive feedback loops [87,88,89]. Notably, loss of protein phosphatase 6 in keratinocytes activates the C/EBP-β-ARG1 pathway, increasing polyamine (e.g., spermidine) production. These polyamines bind self-RNA, enhancing dendritic cell recognition through TLR7 signaling and worsening psoriasis inflammation [90].
4.5. Innate Immunity
Aberrant activation of the innate immune system may contribute to psoriatic inflammation, wherein immune cells including neutrophils, dendritic cells, and macrophages become hyperactivated through pattern recognition receptor (PRR)-mediated detection of DAMPs or PAMPs, subsequently releasing excessive proinflammatory cytokines that further stimulate adaptive immune responses (as shown in Figure 1).
In psoriatic skin lesions, neutrophils accumulate within the epidermal compartment [91,92]. Alarmins, selectins, and cytokines mediate neutrophil recruitment to inflammatory sites [93,94]. The recruited neutrophils release neutrophil extracellular traps (NETs) [95,96]. These NETs induce the release of IL-36γ and lipocalin2 (LCN2) from KCs by activating the TLR4/IL-36R signaling pathway, thereby promoting both neutrophil recruitment and the activation of additional immune cells, and creating a neutrophil-driven, self-propagating inflammatory cycle [97,98]. Furthermore, the neutrophils promote the initiation of inflammatory responses and early lesion development through interactions with macrophages, DCs, and other immune cells [99]. Neutrophil-derived IL-17A potently induces KCs to express proinflammatory cytokines and chemokines, enhancing their migratory phenotype and infiltration [100,101]. Concurrently, migrating neutrophils overexpress matrix metalloproteinase-9 (MMP-9), promoting psoriasis progression by inducing vascular remodeling, enhancing endothelial activation, and driving the accumulation of CD4+ T cells [102]. Interestingly, it was reported that Ly6-targeted immunonanocarriers loaded with roflumilast, which specifically bound to neutrophils, significantly alleviated psoriasiform dermatitis by suppressing neutrophil activation and inflammatory responses [103].
In healthy skin tissue, dendritic cells (DCs) are predominantly confined to the dermal compartment, whereas approximately 50% of DCs migrate to the epidermal layer in psoriatic lesions [104,105,106]. Notably, the recruited DCs in psoriatic lesions stimulate a distinct subset of dual-function T cells that secrete IFN-γ and IL-17, a phenomenon not observed in non-lesional areas or healthy skin [107]. The IFN-γ exacerbates skin barrier dysfunction by inducing apoptosis in KCs [108], while IL-17 promotes hyperproliferation of KCs [88], collectively resulting in characteristic psoriatic acanthosis and chronic inflammation. Plasmacytoid DCs (pDCs) infiltrating psoriatic lesions induce IFN-α secretion, driving T-cell expansion. Xenotransplantation experiments further demonstrated that blocking IFN-α signaling completely prevents psoriasis development [109].
Cutaneous macrophages are broadly categorized into epidermal Langerhans cells (LCs) and dermal macrophages. As tissue-resident immune sentinels, LCs are critical in the early pathogenesis of psoriasis [110], where activated keratinocytes in psoriatic lesions overproduce bone morphogenetic protein 7 (BMP7) to promote progenitor differentiation into pro-inflammatory LCs [111]. These LCs secrete pro-inflammatory cytokines, driving the accumulation of CD4+ T cells and γδ T cells in psoriatic skin [112,113]. Furthermore, psoriatic LCs demonstrate upregulated expression of S100A8/S100A9 proteins that stimulate IL-15 production, while concurrently generating IL-1β and IL-23 [114]. Notably, LCs in psoriasis patients exhibit upregulation of C-X-C motif chemokine ligand 9 (CXCL9), which mediate the targeted recruitment of T lymphocytes to inflammatory foci, thereby amplifying the psoriatic immune cascade [115].
Psoriatic lesions demonstrate increased activated macrophages, particularly during late-stage inflammation [105,116]. Patients with psoriasis show elevated circulating monocyte-derived macrophages in plasma that predominantly exhibit an M1 phenotype; these monocytes infiltrate the dermo-epidermal junction of skin lesions, serving as critical sources of inflammation [117,118]. In psoriasis, IL-17 directly promotes inflammation and activates proinflammatory polarization of monocyte-macrophages, while also enhancing their sensitivity to pathogen-derived TLR4 ligands, thereby amplifying inflammatory signaling [119]. It was found that in the CD18 hypomorphic psoriasiform mouse model, activated macrophages played a pivotal role in chronic inflammation through TNF-α release, with either macrophage depletion or TNF-α neutralization significantly ameliorated skin pathology [117]. Furthermore, macrophages contribute to psoriatic pathogenesis not only through secretion of proinflammatory cytokines but also via intricate interactions with T lymphocytes, dendritic cells, and other immune cells, collectively driving disease progression [120].
Mast cells are critical in acute allergic and chronic inflammatory pathologies [121,122]. Upon IFN-α stimulation, mast cells release exosomes that translocate cytosolic PLA2 enzymatic activity to juxtaposed CD1a-expressing antigen-presenting cells, thereby instigating a cytokine milieu dominated by IL-22 and IL-17A secretion [123]. It was reported that the suprabasin (SBSN)-derived polypeptide, SBSN (50–63), activates mast cells via TLR4 signaling, exacerbating inflammatory responses in psoriasis [124]. The Koebner phenomenon refers to the formation of new psoriatic lesions at locations of cutaneous trauma or other physical stimuli in psoriasis patients. Mechanistically, this process is likely mediated by injury-induced mast cell activation and subsequent release of inflammatory mediators [125].
γδ T cells significantly contribute to psoriatic inflammation, with Vγ5+γδ T andVγ4+γδ T cell subsets exhibiting specific cutaneous tropism [126]. It has been demonstrated that IL-1β, acting through the IL-1R axis, drives γδ T-cell activation and expansion [127]. Pro-inflammatory cytokines secreted by γδ T cells activate keratinocytes, which in turn produce inflammatory mediators that collectively promote inflammatory responses in psoriasis [128].
Natural killer cells (NKs) play a significant immunomodulatory role in psoriasis through cytokine-mediated regulation of Th1/Th2 and Th17/Treg cell network, thereby contributing to the disease pathogenesis [129,130,131]. Research has revealed that psoriasis patients exhibit an increased proportion of the IL-17-secreting NKT cell subset, and targeted therapy against IL-17 can effectively correct this imbalance [132].
Myeloid-derived suppressor cells (MDSCs) typically secrete immunosuppressive cytokines like IL-10 and TGF-β to downregulate the expression of pro-inflammatory cytokines and inhibit immune responses. However, emerging evidence points to a paradoxical role for these cells in psoriasis. It was found that MDSCs from psoriasis patients paradoxically promoted Th17 responses by enhancing Th17 cell differentiation, increasing production of IL-23, IL-1β, and CCL4, while showing reduced expression of PD-1 and PD-L1 and failing to generate Tregs, thereby disrupting T cell homeostasis [133]. Furthermore, in psoriasis, myeloid-derived adjuster cells (MDACs) drive pathological inflammation by promoting differentiation into pro-inflammatory M1 macrophages and DCs. Notably, recent literature indicates that suppression of the RORγt/NFAT1 axis can restore the ability of MDACs to suppress T cell activity in psoriasis [134].
4.6. Adaptive Immunity
It was found that psoriatic skin lesions are enriched with CD4+ T cells [120]. Upon skin injury or infection, DAMPs are released and activate DCs via TLR7/8, triggering the release of proinflammatory cytokines, notably TNF-α and IL-17, which drive the differentiation of CD4+ T cells into Th17 and Th1 subsets. Th17 cells produce cytokines including IL-17A, IL-21 and IL-22, with IL-17A directly activating the NF-κB and MAPK cascades in KCs, inducing the expression of antimicrobial peptides (AMPs) and proinflammatory factors, thereby promoting epidermal hyperplasia and amplifying inflammation [135,136,137,138]. IL-21 and IL-22 disrupt the epithelial barrier, increasing epithelial exposure to pathogens or DAMPs, which perpetuates DC activation [139,140]. Furthermore, under the influence of IL-17 and IL-6, DCs secrete elevated levels of IL-23, a cytokine essential for the development and functional maturation of Th17 cells, thereby further expanding the Th17 population [141].
Th1 cells produce IFN-γ and TNF-α, where IFN-γ enhances the antigen-presenting capacity of DCs and promotes M1 macrophage polarization (secreting IL-12 and NO), thereby reinforcing Th1 differentiation. TNF-α directly contributes to tissue damage, releasing additional DAMPs that sustain TLR7/8 pathway activation, establishing a proinflammatory positive feedback loop [120,142].
Th22 cell is characterized by producing IL-22, which promotes abnormal KCs proliferation and parakeratosis while establishing a positive feedback loop through CCL20 induction [143,144]. Th9 cells display skin tropism and tissue-resident properties in cutaneous environments. Notably, psoriatic lesions exhibit significantly elevated levels of both Th9 cells and their secreted IL-9 compared to healthy skin controls [145]. Mechanistically, IL-9 contributes to psoriasis pathogenesis by potentiating Th17-mediated inflammatory responses and promoting pathological angiogenesis [146]. The frequency of T follicular helper (Tfh) cells increased in both peripheral blood and lesional skin of psoriasis patients, and this increase correlated positively with IL-21 levels and disease severity [147].
However, in psoriasis patients, Th2 cells and their secreted cytokines, IL-4 and IL-10, are reduced [148]. Studies have shown that IL-4 suppresses IL-23 secretion by DCs, thereby limiting Th17 activation [149]. Additionally, activation of acid-sensing ion channel 3 (ASIC3) triggers the production of calcitonin gene-related peptide (CGRP) from sensory neurons, subsequently enhancing IL-23 production by DCs and thereby amplifying Th17 cell activation [150]. These findings not only delineate the systemic inflammatory characteristics in psoriasis but also highlight the pivotal role of the IL-23/IL-17 signaling axis in driving cutaneous inflammation.
It was reported that specific K17 truncated variants (e.g., aa118-aa132) exhibited HLA-DRB1-restricted immunogenicity, inducing T cell proliferation and IFN-γ production in psoriatic patients in vitro [53]. K17 may also trigger CD8+ T cell-specific immune responses through an HLA-C*06:02-restricted mechanism, with this differential MHC restriction likely attributable to its multiple antigenic epitopes. It was found that the presence of peptidoglycan (PG)-specific Th1 cells in psoriatic skin lesions that recognize streptococcal or staphylococcal PG in an HLA-DR-restricted manner and secrete IFN-γ, with enhanced PG recognition exacerbating streptococcus-induced psoriasis [151,152]. Furthermore, the presence of autoantigens (e.g., ADAMTSL5, PLA2G4D) and pathogenic immune complexes (e.g., polyamine-RNA-peptide, LL37-self DNA/RNA), and the consequent activation of immune responses, underscore the pivotal role of T cell-mediated immunity in disease pathogenesis. Psoriatic lesions exhibit predominant expression of T cell receptor (TCR) Vβ2 and Vβ6 gene families, with CDR3 sequence analysis revealing oligoclonal expansion, supporting an antigen-driven T-cell response [153]. In HLA-C*06:02-positive individuals, specific CD8+ T cells expressing Vα3S1/Vβ13S1 TCRs demonstrate cytotoxic activity against HLA-C*06:02-expressing melanocytes, confirming antigen-specific T cell activation through HLA-mediated presentation of autoantigens such as ADAMTSL5 [51]. However, subsequent studies analyzing the TCR repertoire in five psoriasis patients revealed heterogeneous Vβ gene usage patterns without consistent expansion of Vβ6.1–3 or Vβ13.1 clones, suggesting potential interpatient variability in T cell responses [154].
CD8+ T cells, exhibit significant infiltration and activation in psoriatic skin lesions, particularly within the epidermal layer. These cells are activated through antigen presentation by DCs and the cytokine microenvironment, with IL-23 promoting their differentiation into Tc17 cells. Tc17 cells secrete proinflammatory cytokines, directly driving epidermal hyperplasia and inflammatory responses [155]. It was found that IFN-γ produced by Tc17 cells activated the JAK1/STAT1 signaling pathway, upregulating KCs expression of K17 and exacerbating aberrant epidermal proliferation. Meanwhile, Tc22-derived IL-22 directly stimulated KCs proliferation while inhibiting differentiation, leading to the characteristic psoriatic hyperkeratosis [156]. Furthermore, CD8+ T cells interacted with DCs to promote CD4+ T cell differentiation into Th1, Th17, and Th22 subsets, influencing KCs apoptosis, and with fibroblasts to induce CCL20 secretion, enhancing inflammatory cell tissue residency [157]. Additionally, CD8+ T cells recognized multiple psoriasis-associated autoantigens via MHC class I molecules. For instance, ADAMTSL5, produced by melanocytes, was presented by HLA-C*06:02, activating CD8+ T cells and inducing IL-17 and IFN-γ secretion [51]. Therapeutic strategies depleting CD8+ T cells have shown significant efficacy [158], further underscoring their pivotal role in psoriasis pathogenesis.
Compared to healthy skin, psoriatic lesions exhibit a 100-fold increase in T cell numbers, and a 50-fold increase compared to non-lesional skin, indicating significant tissue-resident memory T cells (TRM) expansion in psoriatic plaques [159]. The recurrence of psoriasis at previously affected sites can be attributed to the accumulation and reactivation of TRM cells at these locations [160,161]. Skin TRM cells exhibit remarkable longevity, persisting for more than a year even without local antigen presentation in murine models [159,162]. The development of therapeutic agents targeting aberrantly activated T cells in psoriatic skin, particularly through modulation of their immunological memory functions to prevent disease recurrence, may hold great promise to alleviation or complete cure of psoriasis.
The functional and phenotypic alterations of Tregs in psoriasis remain incompletely understood. It was found that although the number of Treg increased systemically in psoriasis patients, their migration and survival within psoriatic lesions were impaired, resulting in insufficient immunosuppressive capacity and uncontrolled inflammation [163]. Tregs from psoriatic lesions and peripheral blood exhibit diminished capacity to suppress effector T cell proliferation [164,165]. With compromised immunosuppressive function, psoriatic Tregs paradoxically acquire the ability to drive inflammation through IL-17 secretion [165]. Furthermore, obesity-related factors, particularly long-chain fatty acids, have been shown to reduce cutaneous Treg populations, thereby aggravating psoriatic pathogenesis [166].
Psoriasis patients exhibit increased levels of B cell activation in peripheral blood, correlating with the severity of clinical symptoms. It was reported that differential proportions of CD19+ B cell subsets in peripheral blood among various psoriasis subtypes were observed, with these alterations closely being associated with clinical disease activity, particularly during flare-ups [167]. Notably, autoantibodies against K13 [168], heterogeneous nuclear ribonucleoprotein A1 (hnRNPA1) [169], K17 [60,170], and Rab coupling protein isoform 3 (FLJ00294) [168] have been identified in psoriasis patients. Additionally, IgG autoantibodies targeting LL-37 [171] and ADAMTS-L5 [51,172] were observed in a subset of psoriasis patients. Surprisingly, a 2024 case report documented complete and sustained remission of psoriasis following CD19 CAR-T cell therapy targeting B cells, with no relapse observed over 3.5 years of follow-up [173]. Although the presence and role of B cells in psoriatic skin lesions remain elusive, current findings suggest their involvement in disease pathogenesis through multiple mechanisms, including autoantibody production and potential antigen presentation.
Regulatory B cells (Bregs) suppress psoriasiform inflammation through IL-10-dependent mechanisms, which concurrently enhance Tregs expansion and restrain Th17 differentiation [174]. However, it was found that a reduction in Bregs in both peripheral blood and psoriatic lesions showed an inverse correlation with increased populations of IL-17- and IFN-γ- producing T cells [175,176]. Future investigations into therapeutic strategies aimed at enhancing Bregs functionality for optimized psoriasis management should be highly encouraged.
5. Management and Treatment
5.1. Conventional Topical Therapy
Current treatment strategies for psoriasis primarily involve immunosuppression, anti-inflammatory therapy, and palliative care (Conventional treatment approaches are shown in Table 1). In addition to cutaneous and articular manifestations, psoriasis is associated with various comorbidities that require prompt therapeutic intervention.
Corticosteroids serve as first-line topical therapeutic agents, demonstrating favorable safety and tolerability profiles. Clinically, formulations of varying potencies and durations of action can be selected based on disease severity, enabling rapid symptom control [177,178]. Glucocorticoids exert multiple functions through the leucine zipper protein (GILZ), which suppresses Th17-mediated inflammation by regulating Th17-inducing cytokine expression in dendritic cells and inhibiting Th17 cell proliferation and phenotypic expression in T cells [179]. However, prolonged use of topical corticosteroids may induce localized adverse cutaneous reactions, including tachyphylaxis and suppression of the HPA axis [16].
Salicylic acid-based drugs are utilized in the treatment of psoriasis, as they promote desquamation of hyperkeratotic scales and enhance the penetration of concomitant topical therapies, such as corticosteroids [180]. However, their efficacy as monotherapy is limited. Caution is warranted regarding potential systemic absorption, particularly when applied to extensive body surface areas or used concurrently with other topical medications.
Calcipotriol, a vitamin D3 analog, primarily acts by binding to vitamin D receptors on keratinocytes to modulate keratinocyte proliferation and differentiation. It is commonly used for topical treatment of plaque psoriasis [181]. Clinical trials have demonstrated that calcipotriol exhibits comparable efficacy to most topical corticosteroids in the treatment of mild plaque psoriasis, along with a favorable safety profile. Combination therapy with vitamin D3 analogs and topical corticosteroids significantly reduces the incidence of adverse events [181,182].
Tapinarof cream, an aryl hydrocarbon receptor modulator, regulates IL-17 expression and skin barrier proteins (such as filaggrin), demonstrating significant efficacy in mild-to-severe plaque psoriasis while potentially causing local adverse reactions including folliculitis [183].
5.2. Conventional Systemic Therapy
Phototherapy is considered a first-line intervention for moderate-to-severe psoriasis following the failure of topical therapies. Evidence-based medicine has demonstrated that narrowband UVB (NB-UVB) phototherapy exhibits well-established efficacy and an excellent safety profile, making it suitable for nearly all patient populations, including children and pregnant women [184]. Despite its efficacy, clinical implementation is limited by the scarcity of phototherapy centers and need for frequent hospital visits, creating practical burdens for patients.
Acitretin, a synthetic retinoid used for moderate-to-severe psoriasis, has been shown to enhance therapeutic efficacy when combined with other systemic treatments while allowing dose reduction and minimizing adverse effects [185,186,187]. However, robust clinical trial data supporting its efficacy and safety as monotherapy remain insufficient. Of particular importance is its well-documented teratogenicity, which mandates absolute contraindication in women of childbearing potential and continuation of effective contraception for at least three years following treatment [188,189].
Methotrexate (MTX), a folate antagonist, is recommended for the systemic treatment of moderate-to-severe psoriasis and psoriatic arthritis because it interferes with purine and pyrimidine synthesis by inhibiting dihydrofolate reductase (DHFR), thereby exerting antiproliferative and immunomodulatory effects. Serious adverse effects include hepatotoxicity and immunosuppression [190].
Cyclosporine, a calcineurin inhibitor, primarily exerts immunosuppressive effects through selective inhibition of T-lymphocyte activity and is indicated for moderate-to-severe psoriasis and psoriatic arthritis [191,192]. The drug demonstrates rapid therapeutic onset, with relatively low risks of myelosuppression and hepatotoxicity. Major adverse effects include nephrotoxicity, immunosuppression, and potential malignancy [193].

Table 1.
Conventional Drug Therapies for Psoriasis.
| Category | Drug Name | Target/ Mechanism |
Efficacy Profile | Adverse Effects /Potential Risks |
Notes |
|---|---|---|---|---|---|
| Topical Therapies |
Corticosteroids [178] | Suppression of Th17-mediated inflammation; modulation of dendritic and T-cell activity | Rapid symptom control; suitable for mild-to-moderate plaque psoriasis | hypothalamic-pituitary-adrenal axis suppression with prolonged use | Avoid continuous long-term application |
| Calcipotriol (Vitamin D3 analog) [194] | Regulation of keratinocyte proliferation/differentiation; immunomodulation | Improves hyperproliferation and differentiation; safe and effective | Local irritation (erythema, pruritus) | First-line for plaque psoriasis | |
| Tacrolimus ointment [195] | Calcineurin inhibition | High local tolerability | Burning sensation, infection risk | Suitable for mild cases or combination therapy | |
| Calcipotriol/betamethasone combo [196] | Synergistic anti-inflammatory effects of Vitamin D3 analog + corticosteroid | Superior efficacy to monotherapy; reduced relapse | Reduced skin irritation vs. monotherapy | Indicated for moderate-to-thick plaques | |
| Tapinarof cream [183] | Aryl hydrocarbon receptor, modulation; IL-17 suppression; skin barrier enhancement | Significant improvement in mild-to-severe plaque psoriasis | Folliculitis, local irritation | Favorable safety profile | |
| Systemic Therapies | NB-UVB [184] | local immunosuppression | First-line therapy with high safety | Frequent clinic visits; limited phototherapy center access | Suitable for pregnant patients and children |
| MTX [178] | Folate metabolism inhibition; anti-inflammatory | Effective for moderate-to-severe psoriasis and psoriatic arthritis | Hepatotoxicity myelosuppression, gastrointestinal disturbances | Requires regular liver function monitoring | |
| Cyclosporine [191] | Selective T-cell inhibition; IL-2 blockade | Rapid onset; used for acute flares | Nephrotoxicity, immunosuppression | renal monitoring | |
| Acitretin [189,197] | Keratinocyte differentiation modulation; anti-inflammatory | Controls hyperkeratosis; adjunct for severe psoriasis | Teratogenicity, xerosis, photosensitivity | Prohibited for pregnant women |
5** Due to space limitation, only representative drugs per category are listed.
5.3. Biologicals
The ideal therapeutic approach should precisely target pathological immune responses while preserving protective immunity, though this goal requires further investigation to achieve. In cases where conventional systemic therapies demonstrate inadequate efficacy, significant adverse effects, or contraindications due to comorbidities, biologicals have emerged as the treatment of choice.
Antibodies: Monoclonal antibody demonstrates significant efficacy in psoriasis treatment, and bispecific antibodies targeting dual or multiple signaling pathways exhibit synergistic therapeutic effects. Ustekinumab (anti-IL-12/23p40 antibody) [198] and Tildrakizumab (anti-IL-23p19 antibody) [199] have been successfully employed in psoriasis management. The anti-IL-17A antibodies Ixekizumab and Secukinumab demonstrate both safety and clinical efficacy in psoriasis treatment [200,201]. Clinical trials indicate the bispecific antibody Bimekizumab (targeting IL-17A and IL-17F) shows superior efficacy in alleviating psoriasis symptoms compared to inhibitors targeting either IL-17 or TNF-α alone, though with higher incidence of adverse effects including oral candidiasis and diarrhea [202,203]. Spesolimab, an IL-36 receptor monoclonal antibody, effectively controls acute flares and reduces recurrence risk in generalized pustular psoriasis [204]. Cyclophilin A (CypA), a proinflammatory factor interacting with ACE2 and CD147 to promote psoriatic inflammation, shows promising therapeutic potential as anti-CypA monoclonal antibody demonstrates superior efficacy compared to combined anti-IL-17a antibody and methotrexate therapy [205].
Small RNA-mediated gene silencing strategy: Small interfering RNAs (siRNAs) have demonstrated therapeutic potential for psoriasis in recent years. MiR-340 specifically binds to the 3′UTR in mice, targeting and suppressing IL-17A expression, thereby alleviating disease severity in imiquimod-induced psoriatic mouse models [206]. Additionally, intradermal injection of HMGCS1 siRNA modulates immune responses and keratinocyte function, reducing PASI scores, epidermal hyperplasia, and IL-23 expression while inhibiting STAT3 phosphorylation levels in psoriatic mouse models [207]. The rapid advancement of siRNA applications is closely associated with progress in lipid nanoparticle (LNP) technology and nucleic acid modification methods [208,209]. Utilizing a lipid-based nanocarrier (CYnLIP) to co-deliver IL-36α siRNA and erlotinib (a classical tyrosine kinase inhibitor) significantly decreased psoriasis-like plaque severity (PASI score reduction from 4 to 1) and improved pathological features by suppressing inflammatory pathways in murine models [210]. These findings indicate that siRNA and miRNA-based technologies, through targeted gene suppression, offers novel strategies and approaches for psoriasis treatment.
Chimeric antigen receptor T-cell (CAR-T) immunotherapy: CAR-T is an innovative therapeutic approach that combines molecular biology, virology, and immunology to genetically engineer T-cells with synthetic CARs, enabling MHC-independent target cell recognition and elimination for precision medicine. While achieving groundbreaking success in treatment of cancers, its precise targeting capability has expanded applications to autoimmune diseases [211]. In 2024, it was reported that complete remission of 45-year chronic plaque psoriasis coincided with CD19 CAR-T treatment for relapsed large B-cell lymphoma [173]. Subsequently in 2025, CD19 CAR-T therapy induced profound remission in precursor B-cell acute lymphoblastic leukemia while simultaneously resolving refractory severe plaque psoriasis [212]. CD19 CAR-T therapy unexpectedly ameliorated psoriatic symptoms through B-cell depletion. These findings suggest that CD19-targeted CAR-T cells may restore immune homeostasis and suppress autoimmune responses in psoriasis by eliminating aberrantly activated B cells or modulating T-cell subsets. Furthermore, it was found that CRISPR-engineered allogeneic CD19 CAR-T cells safely induced durable remission in refractory autoimmune diseases, including severe myositis and systemic sclerosis, by achieving profound B cell depletion and reversing tissue damage [213]. This groundbreaking study laid a crucial foundation for the clinical advancement of universal CAR-T cell therapy.
Treg-targeted strategy: Tregs are crucial for maintaining immune tolerance by suppressing effector T cell response, and restoring Treg number and function has become a novel therapeutic target in psoriasis treatment. Genetically engineered CAR-FoxP3 Tregs, administered intranasally, effectively suppressed inflammation, ameliorated disease symptoms, and conferred sustained protection against secondary experimental autoimmune encephalomyelitis (EAE) induction in a murine model of EAE [214]. CAR-Tregs represent a novel therapeutic strategy for autoimmune diseases by “restoring immune tolerance”, offering promising applications in the treatment of psoriasis. It was reported that a perforated microneedle (PMN) system loaded with Tregs significantly improved inflammatory symptoms in a psoriasis mouse model by locally delivering cells and releasing propionic acid to enhance Treg function [215]. Furthermore, a clinical trial demonstrated that low-dose IL-2 can safely and selectively expand Tregs, showing potential clinical efficacy in various autoimmune diseases, including psoriasis [216]. However, the broad application of Treg-targeted strategies remains constrained by key challenges, including the complexity of bioengineering techniques, optimization of cell delivery efficiency and tissue-specific targeting, long-term stability of therapeutic effects, and mitigation of risks linked to non-specific immunosuppression. Therefore, future research must focus on addressing these bottlenecks to achieve safe, durable, and personalized immune tolerance induction.
Hematopoietic stem cell transplantation (HSCT): HSCT employs myeloablative conditioning to eliminate pathogenic immune cells (including T and B lymphocytes) and reconstitute normal hematopoietic and immune systems. This therapy is primarily indicated for patients with treatment-refractory severe psoriasis, particularly those with comorbid autoimmune disorders or hematologic abnormalities. Case reports have documented complete remission of both immunoglobulin light-chain amyloidosis and psoriasis following autologous HSCT [217,218]. Due to significant risks including infections, graft-versus-host disease (GVHD), secondary malignancies, and other autoimmune conditions, HSCT is strictly limited to carefully selected severe cases after thorough risk-benefit evaluation. Future research should focus on optimizing regimens, refining patient selection criteria, and developing strategies to reduce complications through clinical trials.
Mesenchymal stromal cells (MSCs) are adult stromal cells possessing self-renewal capacity, multilineage differentiation potential, and immunomodulatory properties. Their low immunogenicity enables allogeneic transplantation. Clinical trials demonstrate that psoriatic patients receiving allogeneic umbilical cord-derived MSCs (UMSCs) transplantation exhibited favorable safety profiles with significant therapeutic effects observed in some cases [219,220]. It was reported that intravenous administration of adipose-derived MSCs (AD-MSCs) for moderate-to-severe psoriasis showed good safety and tolerability [221], with some patients achieving long-term remission [222], thereby providing a foundation for larger-scale investigations. However, current evidence primarily derived from small-scale trials or short-term follow-up, requires further validation of long-term risks and treatment protocol standardization. Future directions should include multicenter randomized double-blind controlled trials to provide high-level evidence, reduce production costs, and improve treatment accessibility. The paracrine mechanism represents the primary therapeutic mode of MSCs, involving both direct cell-cell contact and secretion of soluble factors to modulate immune responses. In psoriasis treatment, significant clinical effects can be achieved using only MSC-conditioned medium (MSC-CM) without direct MSC administration [223]. This strategy avoids potential risks associated with live cell transplantation while offering better standardization potential and improving clinical translation prospects.
Exosomes are 30–150 nm vesicular particles capable of transporting proteins, nucleic acids (e.g., miRNA), and lipids, participating in immunomodulation and inflammatory responses. In psoriasis, MSC-derived exosomes restore immune balance by delivering molecules such as miR-146a to inhibit Th17 differentiation and promote Treg proliferation [224]. Clinical studies demonstrate that adipose-derived mesenchymal stem cell exosomes exhibit optimal therapeutic efficacy, significantly improving erythema, induration, and scaling while modulating immune homeostasis [225]. Furthermore, dimethyl fumarate (DMF)-loaded Treg exosomes (rExo@DMF MNs) delivered via microneedles reduce proinflammatory cytokine release, inhibit keratinocyte proliferation and migration, and ameliorate psoriasiform inflammation in murine models [226]. Future directions may involve exosome-based drug delivery systems (e.g., microneedles, ointments) as novel therapeutic approaches for psoriasis [226,227]. However, exosome therapy faces challenges including heterogeneity and difficulties in clinical-scale production.
Bispecific T-cell engagers (BiTEs) represent a therapeutic approach that redirects T-cell cytotoxicity against specific target cells by simultaneously engaging CD3 on T cells and disease-associated antigens. The CD19xCD3 BiTE blinatumomab mediates T-B cell conjugation to induce B-cell apoptosis, demonstrating clinical improvement in rheumatoid arthritis patients even at low doses [228], while no clinical data yet exists for psoriasis.
Gene therapy: Targeting genetically altered loci or their transcriptional products in psoriatic patients represents a promising therapeutic strategy. As discussed earlier, the modulation of specific gene transcripts using siRNA approaches will not be further elaborated. It was demonstrated that CRISPR/Cas9-mediated PD-L1 overexpression ameliorated psoriatic manifestations in murine models [229]. Furthermore, the application of CRISPR/CAS9 to target NLRP3 has been shown to ameliorate the inflammatory response in murine models [230]. Current research on CRISPR/Cas9-based therapies for psoriasis remains exploratory due to limitations in delivery systems and off-target effects. It was found that the elevated level of H4K16 acetylation (H4K16ac) in macrophages from psoriatic lesions showed positive correlation with self-RNA accumulation and disease severity, the lysine acetyltransferase8 (KAT8) was recruited by AP-1 transcription complexes to CXCL2/CCL3 chemokine promoter regions, promoting neutrophil infiltration and NETs formation, and that genetic ablation or pharmacological inhibition of KAT8 substantially attenuated TLR7-mediated cutaneous inflammation and arthritic pathology in murine models, establishing this epigenetic modulator as a novel therapeutic target [231].
Immunometabolism has emerged as a research focus in psoriasis, with metabolic reprogramming of immune cells, offering novel therapeutic opportunities. Inhibition of Vγ4+γδ T cell mitochondrial translation has been demonstrated to effectively reduce erythema, desquamation, and skin thickening [232]. In psoriatic keratinocytes, protein phosphatase 6 (PP6) deficiency promotes inflammation through enhanced oxidative phosphorylation (OXPHOS), while OXPHOS inhibition significantly ameliorates disease manifestations in murine models [90]. Glutaminase 1 (GLS1)-mediated glutamine metabolism drives Th17 and γδT17 cell differentiation via the mucosa-associated lymphoid tissue lymphoma translocation protein 1 (MALT1)/c-Jun axis, and pharmacological inhibition of this pathway markedly decreases epidermal hyperplasia and inflammation in psoriatic mouse models [233]. These findings collectively suggest that targeting immunometabolic pathways represents a promising therapeutic strategy for psoriasis.
Emerging therapeutic strategies have demonstrated that engineered pyrrole-imidazole polyamides specifically bind to the binding site of c-Rel transcription factor within the IL-23p19 subunit promoter, selectively suppressing IL-23 expression and showing therapeutic efficacy in both imiquimod-induced psoriatic and experimental autoimmune uveitis murine models [234]. Cytotoxic T-lymphocyte antigen-4 (CTLA-4) serves as a negative regulator of T-cell responses through competitive binding with CD80/CD86 to inhibit CD28 co-stimulatory signaling, thereby exerting protective effects against autoimmune pathogenesis [235,236,237]. CTLA4-targeting therapeutics have advanced into clinical trials for the treatment of psoriasis and juvenile idiopathic arthritis [237,238]. The synthetic peptide dNP2-ctCTLA-4, comprising a cell-penetrating peptide (dNP2) fused to CTLA-4′s cytoplasmic signaling domain, facilitates intracellular delivery of therapeutic cargo. By expanding Tregs populations, this construct significantly ameliorates psoriasiform skin inflammation [239].
Bacterial Extracts: The novel exopolysaccharide Ebosin, isolated from Streptomyces sp. 139, suppresses Th17 cell differentiation while enhancing regulatory Tregs proportions, exerting anti-inflammatory effects via modulation of the miR-155- tnfaip3-IL-17 axis and ameliorating imiquimod induced psoriatic inflammation [240]. Similarly, leukotoxin (LtxA), a naturally occurring leukocyte-targeting bacterial protein, binds lymphocyte function-associated antigen-1 (LFA-1) to inhibit hyperproliferation of activated leukocytes in psoriasis patients, demonstrating potent therapeutic efficacy in psoriatic xenograft models [241].
5.4. Small-Molecule Inhibitors
Significant progress in the treatment of psoriasis using small-molecule inhibitors has been made, which provides patients with more therapeutic options. RORγt is a key regulator that promotes Th17 cell differentiation and IL-17 production. Currently, RORγt is considered a novel target for psoriasis drug development. VTP-43742 showed efficacy in alleviating psoriasis symptoms in a Phase IIa clinical trial, though potential safety concerns may exist due to insufficient participant numbers [242]. Apremilast, a phosphodiesterase-4 (PDE4) inhibitor, has been approved for treating psoriasis [243]. H4 receptor antagonists exhibited both antipruritic and anti-inflammatory effects not only in psoriasis mouse models but also for the first time in human clinical trials [244]. In K14-VEGF transgenic mice, flonoltinib maleate (FM), a dual JAK2/FLT3 inhibitor, ameliorated psoriasis-like pathology by concurrently suppressing splenic Th1/Th17 cell differentiation and limiting dendritic cell infiltration [245]. Deucravacitinib, a selective tyrosine kinase 2 inhibitor, was reported to outperform both placebo and apremilast while maintaining favorable tolerability in moderate-to-severe plaque psoriasis [246].
5.5. Alternative Therapy
In traditional Chinese medicine (TCM), psoriasis is termed “Bai Bi” and is treated through a holistic syndrome differentiation approach, forming a multi-target, multi-pathway integrated intervention system with the therapeutic principle of combined internal and external treatments. Clinical patterns mainly include blood-heat syndrome, blood-stasis syndrome, blood-dryness syndrome, and dampness-heat syndrome, treated following principles like “clearing heat-cooling blood (qing re liang xue)”, “activating blood-resolving stasis (huo xue hua yu)”, “nourishing blood-moistening dryness (yang xue run zao)”, and “clearing heat-resolving dampness (qing re hua shi)” [247,248]. Internal treatments primarily employ classical formulas: Rhinoceros Horn & Rehmannia Decoction (Xijiao Dihuang Decoction) or Compound Qingdai Capsules/Pills for blood-heat syndrome; Peach Kernel & Carthamus Four Substances Decoction (Taohong Siwu Decoction) or Yinxie Capsules for blood-stasis syndrome; Xiaoyin Granules or Angelica Drink (Danggui Yinzi) for blood-dryness syndrome; Gentiana Drain the Liver Decoction (Longdan Xiegan Decoction) for dampness-heat syndrome. These medications work by regulating immune-inflammatory response, improving microcirculation, and suppressing keratinocyte hyperproliferation [249,250,251,252]. TCM external therapies demonstrate unique advantages, including topical agents like Indigo Naturalis Ointment (Qingdai Ointment) and Qinbai Ointment, along with non-pharmacological therapies such as filiform needling, fire needling, cupping, and moving cupping [253,254,255]. Herbal bath therapy (e.g., Liangxue Zhiyang Formula) can reduce PASI scores and improve hemorheology [256]. Fire needling and moving cupping modulate Th17/Treg balance and reduce STAT3 pathway activity to alleviate inflammation [257]. While herbal fumigation combined with NB-UVB significantly improves efficacy rates [258,259]. For psoriasis patients with comorbid depression, TCM emphasizes the “liver stagnation-blood heat” (gan yu xue re) pathogenesis, employing liver-soothing formulas or five-element music therapy to regulate HPA axis function and monoamine neurotransmitters, improving psychological status [260,261]. However, the current limitations of TCM research on psoriasis treatment include small number of samples in randomized clinical trials, relatively weak basic research, and the need for further optimization in topical drug formulations and mechanistic studies.
Future treatment strategies for psoriasis should prioritize multidisciplinary cooperation. For patients with psoriasis comorbid with depression, particular emphasis should be placed on psychological counseling and treatment. Comprehensive management involving enhanced patient education, lifestyle modification, and integration of artificial intelligence (AI) technologies spanning diagnostic assessment, therapeutic decision-making, treatment response prediction, risk surveillance, and patient communication/education can significantly improve clinical outcomes. This approach holds promise for advancing future psoriasis management toward timely, intelligent, and efficient paradigms. By managing psoriasis and its complications in an integrated manner, the overall health and quality of life of patients can be much improved.
6. Outlook
The pathogenesis and treatment of psoriasis still have many questions to be addressed, which require a step forward to analyze the pathogenesis of psoriasis, the interaction between genetic and environmental factors and mechanisms, and the excavation and translation of novel immunotherapeutic targets. For example, the use of multi-omics technology to elucidate how susceptibility genes (e.g., HLA-C*06:02, il23r) affect the disease phenotype through epigenetic regulation-mediated immune response. Neuroimmune interaction is also an important research direction. Further studies will be needed to elucidate the molecular mechanisms by which stress regulates the activation and function of immune cells via HPA axis or peripheral nervous system, thereby providing a theoretical foundation for psychological interventions or neuromodulation therapies. To elucidate the mechanism of disease recurrence, efforts will be required to investigate the long-term survival mechanism of TRM cells, and develop targeted CD103-CAR-T cells or small-molecule inhibitors to remove TRM cells, so as to prolong the remission period.
Therapeutic strategies for psoriasis have achieved significant advancement over the past few decades (currently available biologicals and drug targets are listed in Table 2). Although existing treatments demonstrate excellent efficacy in controlling psoriasis symptoms and delaying disease progression, the condition remains incurable, and some patients exhibit suboptimal responses or develop resistance to current therapies, leading to disease recurrence or exacerbation [262]. Consequently, the development of more precise and individualized treatment strategies represents a critical future direction. By employing genetic testing, immunophenotyping, and other advanced methodologies to identify specific immune abnormalities and genetic profiles in individual patients, tailored therapeutic regimens can be designed to maximize efficacy. Despite the demonstrated success of existing biologicals in psoriasis treatment, their limitations, including high treatment costs, potential infection risks and long-term safety concerns, remain significant barriers to broader clinical application [243].
In the field of immunotherapy, it is imperative to overcome the limitations of existing drug targets, particularly the key molecular mediators in the pathogenesis of psoriasis, though these targets still require further in-depth investigation. Future research should be encouraged to explore novel intervention strategies targeting molecular pathways such as the RORγt/NFAT1 axis [134] to address resistance to IL-17/IL-23 inhibitors in certain patient populations. Of note, small-molecule inhibitors of RORγt (e.g., VTP-43742) [242,263] and JAK inhibitors (e.g., Tofacitinib) [264] have demonstrated promising potential in clinical trials and may emerge as viable therapeutic alternatives for psoriasis in the future.
In recent years, cell-based therapies have achieved groundbreaking advances in oncology, and their potential applications in autoimmune diseases are increasingly gaining attention. Notably, an unexpected case of psoriasis remission following CD19 CAR-T cell therapy mediated through B-cell depletion has unveiled a novel therapeutic avenue for this disease [173,265]. CAR-T cell therapy may disrupt the IL-23/Th17 axis-driven pathological paradigm and restore immune homeostasis by depleting aberrantly activated B cells or modulating T-cell subsets. Moving forward, CAR-T cell therapy and other cell-based strategies (e.g., CAR-Tregs, MSCs) hold great promise as transformative treatment modalities for psoriasis, though their long-term safety and efficacy require further validation through large-scale clinical trials. MSCs and their exosomes demonstrate therapeutic potential through immunomodulation and paracrine effects; however, challenges remain in standardized production, heterogeneity control, and long-term safety. While gene-editing technologies (e.g., CRISPR/Cas9) have shown promise in murine models, the efficiency of delivery systems and off-target risks represent major barriers to clinical translation.
Current therapeutic strategies for psoriasis predominantly rely on non-specific immunosuppression or immunomodulation, which may lead to broad immune suppression and potential adverse effects. Future research should focus on exploring immune tolerance induction as a novel treatment approach, particularly through antigen-specific immunotherapy targeting psoriasis-associated autoantigens such as K17 [60,170] and ADAMTSL5 [51,172]. For instance, the targeting of antigen-specific T cells or B cells by blocking antigen-specific responses or inducing tolerance to self-antigens may effectively control the inflammatory response of psoriasis without compromising systemic immunity. Additionally, bacterial extracts (e.g., Ebosin) have demonstrated anti-inflammatory effects in psoriatic animal models by modulating immune responses and inflammatory pathways [240], suggesting their potential as future therapeutic options. With rapid advancements in nanotechnology and RNA interference (RNAi), biomaterial-based delivery systems and siRNA therapeutics hold considerable promise for psoriasis treatment. For instance, HMGCS1 siRNA [207] and IL-36α siRNA [210] have exhibited significant anti-inflammatory efficacy in murine models, highlighting their potential for clinical translation.
Immunometabolic studies have revealed the pivotal role of metabolic reprogramming (e.g., OXPHOS, glutaminolysis) in psoriasis pathogenesis. Targeting these pathways (e.g., via GLS1 inhibitors) may represent a promising therapeutic strategy. Epigenetic regulation (e.g., KAT8 inhibition) offers precise intervention targets by modulating inflammatory gene expression, though its tissue specificity and safety profile require further investigation.
TCM demonstrates unique advantages in psoriasis treatment through multi-target regulation (e.g., IL-17 inhibition, Th17/Treg balance modulation), particularly for mild-to-moderate cases or in synergistic combination with Western medicine. However, the lack of high-quality evidence-based medical data and standardized protocols remains a bottleneck for its global adoption. Future development requires integration with modern technologies to advance precision medicine in integrative TCM-Western approaches.
In summary, the therapeutic landscape for psoriasis holds immense promise, with future research continuing to explore precise, safe, and effective treatment strategies. Interdisciplinary collaboration and technological innovation will facilitate the translation of the research results from bench to clinical applications, ultimately advancing psoriasis therapy toward individualized, precision-based and integrated approaches. While significant challenges remain, the continuous scientific and technological advancements will ultimately render the cure for psoriasis an achievable reality.
Table 2.
Novel and Investigational Therapies for Psoriasis.
| Category | Drug Name | Target/Mechanism | Approval Status | Efficacy Profile | Adverse Effects/Potential Risks | Notes |
|---|---|---|---|---|---|---|
| IL-17 monoclonal antibody | Secukinumab [201] | IL-17A neutralization; blocks IL-17A signaling | Approved (US, EU, CN, etc.) | PASI 90 response: 80–90%; rapid lesion clearance | Injection-site reactions, candidiasis | Requires infection monitoring |
| Vunakizumab (China-developed) [266] | Blocks IL-17A signaling | Approved (China) | Annual dosing: 14 injections; PASI 100 response: >70% | Low infection risk | Cost-effective domestic innovator | |
| Bimekizumab [202] | Dual IL-17A/F inhibition | Approved (EU, UK; US pending) | Superior to IL-17A monotherapy (higher PASI 100 rates) | Oral candidiasis, diarrhea | Synergistic dual-target action | |
| IL-23 monoclonal antibody | Guselkumab [267] | IL-23p19 blockade | Approved (US, EU, CN, etc.) | Q8W dosing; PASI 90 response >80% | Mild injection-site reactions, low Tuberculosis risk | Sustained long-term remission |
| Ustekinumab [198] | Dual IL-12/23p40 blockade; inhibits Th1/Th17 pathways | Approved (Global) | Long-term disease control in moderate-to-severe plaque psoriasis | Respiratory infections | First dual-target biologic for IL-12 and IL-23 | |
| TNF-α monoclonal antibody | Adalimumab [268] | TNF-α blockade | Approved (Global) | PASI 75 response: 70–80% in moderate-to-severe cases | Tuberculosis reactivation, potential malignancy risk | Preferred for psoriatic arthritis comorbidity |
| IL-36 monoclonal antibody | Spesolimab [204] | IL-36 blockade | Approved (US, EU, CN) | Rapid control of generalized pustular psoriasis (GPP) | Infections, infusion reactions | First-in-class IL-36 pathway inhibitor |
| Small-Molecule Inhibitors | Apremilast [243] | DE4 inhibitor; reduces pro-inflammatory cytokines | Approved in multiple countries | Suitable for mild-to-moderate cases; oral administration | Diarrhea, nausea, weight loss | Superior safety profile compared to traditional immunosuppressants |
| Tofacitinib [264] | JAK1/3 inhibition; blocks JAK-STAT signaling | Approved for PsA in multiple countries | Oral administration; rapid relief of articular symptoms | Infection risk, thromboembolic events | Requires long-term safety monitoring | |
| VTP-43742/PF-06763809 [242,269] | RORγt inhibition; reduces IL-17 production | Clinical trials | Novel Th17 pathway suppression with promising efficacy | Good tolerability/safety pending | First-in-class RORγt inhibitors | |
| Cell Therapies | CD19 CAR-T [173] | CD19-targeted B-cell depletion | Case reports only | Complete psoriasis remission sustained | CRS, B-cell aplasia-related infections | Mechanism unclear; target optimization needed |
| CAR-Tregs [214] | Engineered Tregs for enhanced immune suppression | Preclinical studies | Effective in experimental autoimmune models (exploratory for psoriasis) | Technical complexity, graft rejection risks | Potential tolerance-restoring approach | |
| Umbilical/Adipose MSCs [220,221,222] | Immunomodulation (paracrine effects) | Clinical trials | Good safety profile, preliminary evidence of sustained improvement in some patients | Transient fever, infusion-related reactions | Requires stringent quality control | |
| TCM Therapy | Compound Indigo Capsule [270] | Multi-target modulation | Approved (China) | Significant improvement in erythema and infiltration | Diarrhea, abdominal pain | Contraindicated in pregnancy |
| miRNA | miR-340 siRNA [206] | Downregulates IL-17A expression via RNA interference | Preclinical studies | Attenuates inflammation in murine models | Low delivery efficiency, poor stability | Requires nanocarrier optimization |
| Immunometabolic Modulator | GLS1 Inhibitor [233] | Glutamine metabolism blockade (Th17 differentiation) | Preclinical studies | Markedly improves IMQ-induced psoriasiform dermatitis in mice | Unknown | Targets metabolic reprogramming, avoids direct immunosuppression |
| Epigenetic Modulator | KAT8 Inhibitor [231] | Reduces H4K16ac (suppresses CXCL2/CCL3) | Preclinical studies | Ameliorates IMQ-induced murine model symptoms | Unknown | N/A |
| Other Investigative Drugs | CYnLIP(Nanocarrier) [210] | Co-delivers IL-36α siRNA + erlotinib | Preclinical studies | Significantly reduces murine PASI scores | Human safety unverified | Combines gene therapy + chemical drug |
| Ebosin(Streptomyces exopolysaccharide) [240] | Inhibits Th17 differentiation; modulates miR-155-TNFAIP3-IL-17 axis | Preclinical studies | Attenuates inflammation in murine models | Unclear toxicity profile | Natural product with multi-pathway modulation | |
| Antibody-Nanoparticle Conjugate [103] | Neutrophil-specific delivery of anti-inflammatory payload | Preclinical studies | Reduces systemic toxicity; site-specific action | Preclinical safety pending | Precision delivery technology prototype |
* Due to space limitation, not all investigational psoriasis therapies are shown.
Acknowledgments
This work was supported by Start-Up fundings for faculty recruitment from Henan University (CX3050A1000178, CX3050A1000179).
Conflicts of Interest
The authors declare no conflict of interest. The funding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Song Y, Li J, Wu Y: Evolving understanding of autoimmune mechanisms and new therapeutic strategies of autoimmune disorders. Signal Transduction and Targeted Therapy 2024, 9.
- Theo Vos SSL, Cristiana Abbafati, Kaja M Abbas, Mohammad Abbasi, Mitra Abbasifard, Mohsen Abbasi-Kangevari, Hedayat Abbastabar. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef]
- Ritchlin CT, Colbert RA, Gladman DD: Psoriatic Arthritis. N Engl J Med 2017, 376, 957–970. [CrossRef]
- Gelfand JM, Yeung H: Metabolic syndrome in patients with psoriatic disease. J Rheumatol Suppl 2012, 89, 24–28. [CrossRef] [PubMed]
- Robati RM, Partovi-Kia M, Haghighatkhah HR, Younespour S, Abdollahimajd F: Increased serum leptin and resistin levels and increased carotid intima-media wall thickness in patients with psoriasis: is psoriasis associated with atherosclerosis? J Am Acad Dermatol 2014, 71, 642–648. [CrossRef] [PubMed]
- Sampogna F, Tabolli S, Abeni D: Living with psoriasis: prevalence of shame, anger, worry, and problems in daily activities and social life. Acta Derm Venereol 2012, 92, 299–303. [CrossRef] [PubMed]
- Di Cesare A, Di Meglio P, Nestle FO: The IL-23/Th17 axis in the immunopathogenesis of psoriasis. J Invest Dermatol 2009, 129, 1339–1350. [CrossRef]
- Ghoreschi K, Balato A, Enerbäck C, Sabat R: Therapeutics targeting the IL-23 and IL-17 pathway in psoriasis. Lancet 2021, 397, 754–766. [CrossRef]
- van der Fits L, Mourits S, Voerman JS, Kant M, Boon L, Laman JD, Cornelissen F, Mus AM, Florencia E, Prens EP, Lubberts E: Imiquimod-induced psoriasis-like skin inflammation in mice is mediated via the IL-23/IL-17 axis. J Immunol 2009, 182, 5836–5845. [CrossRef]
- Griffiths CEM, Armstrong AW, Gudjonsson JE, Barker J: Psoriasis. Lancet 2021, 397, 1301–1315. [CrossRef]
- Li H, Yao Q, Mariscal AG, Wu X, Hülse J, Pedersen E, Helin K, Waisman A, Vinkel C, Thomsen SF, et al: Epigenetic control of IL-23 expression in keratinocytes is important for chronic skin inflammation. Nat Commun 2018, 9, 1420. [CrossRef]
- Dand N, Stuart PE, Bowes J, Ellinghaus D, Nititham J, Saklatvala JR, Teder-Laving M, Thomas LF, Traks T, Uebe S, et al.: GWAS meta-analysis of psoriasis identifies new susceptibility alleles impacting disease mechanisms and therapeutic targets. Nat Commun 2025, 16, 2051. [CrossRef]
- Mallon E, Bunce M, Savoie H, Rowe A, Newson R, Gotch F, Bunker CB: HLA-C and guttate psoriasis. Br J Dermatol 2000, 143, 1177–1182.
- Chen YL, Ng JSW, Ottakandathil Babu R, Woo J, Nahler J, Hardman CS, Kurupati P, Nussbaum L, Gao F, Dong T, et al.: Group A Streptococcus induces CD1a-autoreactive T cells and promotes psoriatic inflammation. Sci Immunol, 2023; 8, eadd9232.
- Yen YF, Chuang PH, Jen IA, Chen M, Lan YC, Liu YL, Lee Y, Chen YH, Chen YA: Incidence of autoimmune diseases in a nationwide HIV/AIDS patient cohort in Taiwan, 2000–2012. Ann Rheum Dis 2017, 76, 661–665. [CrossRef]
- Kim Alexander Papp WG, Charles W Lynde, Yves Poulin, David Adam, Benjamin Barankin, Kirk Barber, Marc Bourcier, Melinda Gooderham, Lyn C Guenther, Vincent C Ho, Andrei Metelitsa, Neil H Shear, Ronald B Vender, Norman Wasel, Marni C Wiseman: 2016 Addendum to the Canadian Guidelines for the Management of Plaque Psoriasis 2009. J Cutan Med Surg 2016, 20, 375–431.
- Khoroshun K, Bantel C, Hoffmann F, Jobski K: Methotrexate-related drug reactions on kidneys and liver in rheumatoid arthritis: an analysis of spontaneous reports in EudraVigilance. Arthritis Res Ther 2025, 27, 80. [CrossRef]
- Krupp P, Monka C: Side-effect profile of cyclosporin A in patients treated for psoriasis. Br J Dermatol 1990, 122 Suppl 36, 47–56.
- Langley RG, Elewski BE, Lebwohl M, Reich K, Griffiths CE, Papp K, Puig L, Nakagawa H, Spelman L, Sigurgeirsson B, et al: Secukinumab in plaque psoriasis--results of two phase 3 trials. N Engl J Med 2014, 371, 326–338. [CrossRef] [PubMed]
- Reich K, Armstrong AW, Langley RG, Flavin S, Randazzo B, Li S, Hsu MC, Branigan P, Blauvelt A: Guselkumab versus secukinumab for the treatment of moderate-to-severe psoriasis (ECLIPSE): results from a phase 3, randomised controlled trial. Lancet 2019, 394, 831–839. [CrossRef] [PubMed]
- Murray CJ, Vos T, Lozano R, Naghavi M, Flaxman AD, Michaud C, Ezzati M, Shibuya K, Salomon JA, Abdalla S, et al.: Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990–2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2197–2223. [CrossRef]
- Danielsen K, Olsen AO, Wilsgaard T, Furberg AS: Is the prevalence of psoriasis increasing? A 30-year follow-up of a population-based cohort. Br J Dermatol 2013, 168, 1303–1310. [CrossRef]
- Gibbs S: Skin disease and socioeconomic conditions in rural Africa: Tanzania. Int J Dermatol 1996, 35, 633–639. [CrossRef]
- Luo H: Global burden and cross-country inequalities in six major immune-mediated inflammatory diseases from 1990 to 2021: A systemic analysis of the Global Burden of Disease Study 2021. Autoimmun Rev 2024, 23, 103639. [CrossRef]
- Wang K, Zhao Y, Cao X: Global burden and future trends in psoriasis epidemiology: insights from the global burden of disease study 2019 and predictions to 2030. Arch Dermatol Res 2024, 316, 114. [CrossRef]
- Wu J, Ma Y, Yang J, Tian Y: Exposure to Air Pollution, Genetic Susceptibility, and Psoriasis Risk in the UK. JAMA Netw Open 2024, 7, e2421665. [CrossRef] [PubMed]
- Boehncke WH, Schön MP: Psoriasis. Lancet 2015, 386, 983–994. [CrossRef] [PubMed]
- Fu Y, Lee CH, Chi CC: Association of Psoriasis With Inflammatory Bowel Disease: A Systematic Review and Meta-analysis. JAMA Dermatol 2018, 154, 1417–1423. [CrossRef] [PubMed]
- Ruan Z, Lu T, Chen Y, Yuan M, Yu H, Liu R, Xie X: Association Between Psoriasis and Nonalcoholic Fatty Liver Disease Among Outpatient US Adults. JAMA Dermatol 2022, 158, 745–753. [CrossRef]
- Ramessur R, Saklatvala J, Budu-Aggrey A, Ostaszewski M, Möbus L, Greco D, Ndlovu M, Mahil SK, Barker JN, Brown S, et al.: Exploring the Link Between Genetic Predictors of Cardiovascular Disease and Psoriasis. JAMA Cardiol 2024, 9, 1009–1017. [CrossRef]
- Takeshita J, Grewal S, Langan SM, Mehta NN, Ogdie A, Van Voorhees AS, Gelfand JM: Psoriasis and comorbid diseases: Epidemiology. J Am Acad Dermatol 2017, 76, 377–390.
- Branisteanu DE, Pirvulescu RA, Spinu AE, Porumb EA, Cojocaru M, Nicolescu AC, Branisteanu DC, Branisteanu CI, Dimitriu A, Alexa AI, Toader MP: Metabolic comorbidities of psoriasis (Review). Exp Ther Med 2022, 23, 179.
- Pouplard C, Brenaut E, Horreau C, Barnetche T, Misery L, Richard MA, Aractingi S, Aubin F, Cribier B, Joly P, et al.: Risk of cancer in psoriasis: a systematic review and meta-analysis of epidemiological studies. J Eur Acad Dermatol Venereol, 2013; 27, (Suppl 3), 36–46.
- Ryan C, Sadlier M, De Vol E, Patel M, Lloyd AA, Day A, Lally A, Kirby B, Menter A: Genital psoriasis is associated with significant impairment in quality of life and sexual functioning. J Am Acad Dermatol 2015, 72, 978–983. [CrossRef] [PubMed]
- Singh S, Taylor C, Kornmehl H, Armstrong AW: Psoriasis and suicidality: A systematic review and meta-analysis. J Am Acad Dermatol 2017, 77, 425–440.e422. [CrossRef]
- Rompoti N, Tsiori S, Kontoangelos K, Kouzoupis A, Papageorgiou C, Gregoriou S, Stratigos A, Rigopoulos D: Psychopathological Profile of Patients with Moderate-to-Severe Plaque Psoriasis and Its Correlation to DLQI: Results from a Prospective, Monocentric Clinical Study. J Clin Med 2024, 13.
- Rendon A, Schäkel K: Psoriasis Pathogenesis and Treatment. International Journal of Molecular Sciences 2019, 20, 1475. [CrossRef]
- Griffiths CE, Barker JN: Pathogenesis and clinical features of psoriasis. Lancet 2007, 370, 263–271. [CrossRef]
- Mehta S, Singal A, Singh N, Bhattacharya SN: A study of clinicohistopathological correlation in patients of psoriasis and psoriasiform dermatitis. Indian J Dermatol Venereol Leprol 2009, 75, 100. [CrossRef]
- Dupire G, Droitcourt C, Hughes C, Le Cleach L: Antistreptococcal interventions for guttate and chronic plaque psoriasis. Cochrane Database Syst Rev 2019, 3, Cd011571.
- Martin BA, Chalmers RJ, Telfer NR: How great is the risk of further psoriasis following a single episode of acute guttate psoriasis? Arch Dermatol 1996, 132, 717–718. [CrossRef]
- Singh RK, Lee KM, Ucmak D, Brodsky M, Atanelov Z, Farahnik B, Abrouk M, Nakamura M, Zhu TH, Liao W: Erythrodermic psoriasis: pathophysiology and current treatment perspectives. Psoriasis (Auckl) 2016, 6, 93–104.
- Augustin M, Reich K, Blome C, Schäfer I, Laass A, Radtke MA: Nail psoriasis in Germany: epidemiology and burden of disease. Br J Dermatol 2010, 163, 580–585.
- Park JH, Park YJ, Kim SK, Kwon JE, Kang HY, Lee ES, Choi JH, Kim YC: Histopathological Differential Diagnosis of Psoriasis and Seborrheic Dermatitis of the Scalp. Ann Dermatol 2016, 28, 427–432. [CrossRef] [PubMed]
- Tsoi LC, Stuart PE, Tian C, Gudjonsson JE, Das S, Zawistowski M, Ellinghaus E, Barker JN, Chandran V, Dand N, et al.: Large scale meta-analysis characterizes genetic architecture for common psoriasis associated variants. Nat Commun 2017, 8, 15382. [CrossRef] [PubMed]
- Tang H, Jin X, Li Y, Jiang H, Tang X, Yang X, Cheng H, Qiu Y, Chen G, Mei J, et al.: A large-scale screen for coding variants predisposing to psoriasis. Nat Genet 2014, 46, 45–50. [CrossRef] [PubMed]
- Sun LD, Cheng H, Wang ZX, Zhang AP, Wang PG, Xu JH, Zhu QX, Zhou HS, Ellinghaus E, Zhang FR, et al.: Association analyses identify six new psoriasis susceptibility loci in the Chinese population. Nat Genet 2010, 42, 1005–1009. [CrossRef]
- Lønnberg AS, Skov L, Skytthe A, Kyvik KO, Pedersen OB, Thomsen SF: Heritability of psoriasis in a large twin sample. Br J Dermatol 2013, 169, 412–416. [CrossRef]
- Antonatos C, Grafanaki K, Asmenoudi P, Xiropotamos P, Nani P, Georgakilas GK, Georgiou S, Vasilopoulos Y: Contribution of the Environment, Epigenetic Mechanisms and Non-Coding RNAs in Psoriasis. Biomedicines 2022, 10.
- Della Bella C, Corrà A, Mantengoli E, Galano A, Benagiano M, Bonciani D, Mariotti EB, Pratesi S, Quintarelli L, Aimo C, et al. : Skin IL-17A and IFN-γ Production Correlate with Disease Severity in Patients with Psoriasis and Streptococcal Infection. J Invest Dermatol 2023, 143, 925–932. [Google Scholar] [CrossRef]
- Arakawa A, Siewert K, Stöhr J, Besgen P, Kim SM, Rühl G, Nickel J, Vollmer S, Thomas P, Krebs S, et al.: Melanocyte antigen triggers autoimmunity in human psoriasis. J Exp Med 2015, 212, 2203–2212. [CrossRef]
- Arakawa A, Reeves E, Vollmer S, Arakawa Y, He M, Galinski A, Stöhr J, Dornmair K, James E, Prinz JC: ERAP1 Controls the Autoimmune Response against Melanocytes in Psoriasis by Generating the Melanocyte Autoantigen and Regulating Its Amount for HLA-C*06:02 Presentation. J Immunol 2021, 207, 2235–2244.
- Shen Z, Wang G, Fan JY, Li W, Liu YF: HLA DR B1*04, *07-restricted epitopes on Keratin 17 for autoreactive T cells in psoriasis. J Dermatol Sci 2005, 38, 25–39. [CrossRef]
- Cargill M, Schrodi SJ, Chang M, Garcia VE, Brandon R, Callis KP, Matsunami N, Ardlie KG, Civello D, Catanese JJ, et al.: A large-scale genetic association study confirms IL12B and leads to the identification of IL23R as psoriasis-risk genes. Am J Hum Genet 2007, 80, 273–290. [CrossRef] [PubMed]
- Zhu H, Lou F, Yin Q, Gao Y, Sun Y, Bai J, Xu Z, Liu Z, Cai W, Ke F, et al.: RIG-I antiviral signaling drives interleukin-23 production and psoriasis-like skin disease. EMBO Mol Med 2017, 9, 589–604. [CrossRef]
- Hayashi M, Hirota T, Saeki H, Nakagawa H, Ishiuji Y, Matsuzaki H, Tsunemi Y, Kato T, Shibata S, Sugaya M, et al.: Genetic polymorphism in the TRAF3IP2 gene is associated with psoriasis vulgaris in a Japanese population. J Dermatol Sci 2014, 73, 264–265. [CrossRef] [PubMed]
- Harden JL, Krueger JG, Bowcock AM: The immunogenetics of Psoriasis: A comprehensive review. J Autoimmun 2015, 64, 66–73. [CrossRef] [PubMed]
- Teng Y, Xie W, Tao X, Liu N, Yu Y, Huang Y, Xu D, Fan Y: Infection-provoked psoriasis: Induced or aggravated (Review). Exp Ther Med 2021, 21, 567. [CrossRef]
- Assarsson M, Söderman J, Seifert O: Significant Correlation Between Cutaneous Abundance of Streptococcus and Psoriasis Severity in Patients with FBXL19 Gene Variants. Acta Derm Venereol, 2024; 104, adv34892.
- Gudmundsdottir AS, Sigmundsdottir H, Sigurgeirsson B, Good MF, Valdimarsson H, Jonsdottir I: Is an epitope on keratin 17 a major target for autoreactive T lymphocytes in psoriasis? Clin Exp Immunol 1999, 117, 580–586.
- Johnston A, Gudjonsson JE, Sigmundsdottir H, Love TJ, Valdimarsson H: Peripheral blood T cell responses to keratin peptides that share sequences with streptococcal M proteins are largely restricted to skin-homing CD8(+) T cells. Clin Exp Immunol 2004, 138, 83–93. [CrossRef]
- Morar N, Willis-Owen SA, Maurer T, Bunker CB: HIV-associated psoriasis: pathogenesis, clinical features, and management. Lancet Infect Dis 2010, 10, 470–478. [CrossRef]
- Fuchs D, Hausen A, Reibnegger G, Werner ER, Dierich MP, Wachter H: Psoriasis, gamma-interferon, and the acquired immunodeficiency syndrome. Ann Intern Med 1987, 106, 165. [CrossRef]
- Liu S, He M, Jiang J, Duan X, Chai B, Zhang J, Tao Q, Chen H: Triggers for the onset and recurrence of psoriasis: a review and update. Cell Communication and Signaling 2024, 22.
- Aram K, Patil A, Goldust M, Rajabi F: COVID-19 and exacerbation of dermatological diseases: A review of the available literature. Dermatol Ther 2021, 34, e15113. [CrossRef]
- Chen ML, Kao WM, Huang JY, Hung YM, Wei JC: Human papillomavirus infection associated with increased risk of new-onset psoriasis: a nationwide population-based cohort study. Int J Epidemiol 2020, 49, 786–797. [CrossRef]
- Rousset L, Halioua B: Stress and psoriasis. Int J Dermatol 2018, 57, 1165–1172. [CrossRef]
- Woźniak E, Owczarczyk-Saczonek A, Placek W: Psychological Stress, Mast Cells, and Psoriasis-Is There Any Relationship? Int J Mol Sci 2021, 22.
- Kanda N, Watanabe S: Regulatory roles of sex hormones in cutaneous biology and immunology. J Dermatol Sci 2005, 38, 1–7. [CrossRef]
- Adachi A, Honda T: Regulatory Roles of Estrogens in Psoriasis. J Clin Med 2022, 11.
- Cemil BC, Cengiz FP, Atas H, Ozturk G, Canpolat F: Sex hormones in male psoriasis patients and their correlation with the Psoriasis Area and Severity Index. J Dermatol 2015, 42, 500–503. [CrossRef]
- Kobayashi K, Chikazawa S, Chen Y, Suzuki S, Ichimasu N, Katagiri K: Oestrogen inhibits psoriasis-like dermatitis induced by imiquimod in mice in relation to increased IL-10 producing cells despite elevated expression of IL-22, IL-23, IL-17 mRNA. Exp Dermatol 2023, 32, 203–209. [CrossRef] [PubMed]
- Terán-Pérez G, Arana-Lechuga Y, Esqueda-León E, Santana-Miranda R, Rojas-Zamorano J, Velázquez Moctezuma J: Steroid hormones and sleep regulation. Mini Rev Med Chem 2012, 12, 1040–1048.
- Paus R, Theoharides TC, Arck PC: Neuroimmunoendocrine circuitry of the ‘brain-skin connection’. Trends Immunol 2006, 27, 32–39. [CrossRef] [PubMed]
- Wei J, Zhu J, Xu H, Zhou D, Elder JT, Tsoi LC, Patrick MT, Li Y: Alcohol consumption and smoking in relation to psoriasis: a Mendelian randomization study. Br J Dermatol 2022, 187, 684–691. [CrossRef]
- Brenaut E, Horreau C, Pouplard C, Barnetche T, Paul C, Richard MA, Joly P, Le Maître M, Aractingi S, Aubin F, et al.: Alcohol consumption and psoriasis: a systematic literature review. J Eur Acad Dermatol Venereol, 2013; 3, (Suppl 3), 30–35.
- Murzaku EC, Bronsnick T, Rao BK: Diet in dermatology: Part II. Melanoma, chronic urticaria, and psoriasis. J Am Acad Dermatol 2014, 71, 1053.e1051–1053.e1016.
- Dai YX, Wang SC, Chou YJ, Chang YT, Chen TJ, Li CP, Wu CY: Smoking, but not alcohol, is associated with risk of psoriasis in a Taiwanese population-based cohort study. J Am Acad Dermatol 2019, 80, 727–734. [CrossRef]
- Nestle FO, Di Meglio P, Qin JZ, Nickoloff BJ: Skin immune sentinels in health and disease. Nat Rev Immunol 2009, 9, 679–691. [CrossRef]
- Perera GK, Di Meglio P, Nestle FO: Psoriasis. Annu Rev Pathol 2012, 7, 385–422. [CrossRef]
- Zhang LJ, Sen GL, Ward NL, Johnston A, Chun K, Chen Y, Adase C, Sanford JA, Gao N, Chensee M, et al.: Antimicrobial Peptide LL37 and MAVS Signaling Drive Interferon-β Production by Epidermal Keratinocytes during Skin Injury. Immunity 2016, 45, 119–130. [CrossRef]
- Zhou X, Chen Y, Cui L, Shi Y, Guo C: Advances in the pathogenesis of psoriasis: from keratinocyte perspective. Cell Death Dis 2022, 13, 81. [CrossRef] [PubMed]
- Albanesi C, Madonna S, Gisondi P, Girolomoni G: The Interplay Between Keratinocytes and Immune Cells in the Pathogenesis of Psoriasis. Front Immunol 2018, 9, 1549. [CrossRef] [PubMed]
- Yan S, Xu Z, Lou F, Zhang L, Ke F, Bai J, Liu Z, Liu J, Wang H, Zhu H, et al: NF-κB-induced microRNA-31 promotes epidermal hyperplasia by repressing protein phosphatase 6 in psoriasis. Nat Commun 2015, 6, 7652. [CrossRef]
- Silva de Melo BM, Veras FP, Zwicky P, Lima D, Ingelfinger F, Martins TV, da Silva Prado D, Schärli S, Publio G, Hiroki CH, et al: S100A9 Drives the Chronification of Psoriasiform Inflammation by Inducing IL-23/Type 3 Immunity. J Invest Dermatol 2023, 143, 1678–1688.e1678. [CrossRef] [PubMed]
- Homey B, Dieu-Nosjean MC, Wiesenborn A, Massacrier C, Pin JJ, Oldham E, Catron D, Buchanan ME, Müller A, deWaal Malefyt R, et al: Up-regulation of macrophage inflammatory protein-3 alpha/CCL20 and CC chemokine receptor 6 in psoriasis. J Immunol 2000, 164, 6621–6632. [CrossRef]
- Müller A, Hennig A, Lorscheid S, Grondona P, Schulze-Osthoff K, Hailfinger S, Kramer D: IκBζ is a key transcriptional regulator of IL-36-driven psoriasis-related gene expression in keratinocytes. Proc Natl Acad Sci U S A 2018, 115, 10088–10093. [CrossRef]
- Ekman AK, Bivik Eding C, Rundquist I, Enerbäck C: IL-17 and IL-22 Promote Keratinocyte Stemness in the Germinative Compartment in Psoriasis. J Invest Dermatol 2019, 139, 1564–1573.e1568. [CrossRef] [PubMed]
- Lowes MA, Suárez-Fariñas M, Krueger JG: Immunology of psoriasis. Annu Rev Immunol 2014, 32, 227–255. [CrossRef]
- Lou F, Sun Y, Xu Z, Niu L, Wang Z, Deng S, Liu Z, Zhou H, Bai J, Yin Q, et al: Excessive Polyamine Generation in Keratinocytes Promotes Self-RNA Sensing by Dendritic Cells in Psoriasis. Immunity 2020, 53, 204–216.e210. [CrossRef] [PubMed]
- Rodriguez-Rosales YA, Langereis JD, Gorris MAJ, van den Reek J, Fasse E, Netea MG, de Vries IJM, Gomez-Muñoz L, van Cranenbroek B, Körber A, et al.: Immunomodulatory aged neutrophils are augmented in blood and skin of psoriasis patients. J Allergy Clin Immunol 2021, 148, 1030–1040. [CrossRef]
- Katayama H: Development of psoriasis by continuous neutrophil infiltration into the epidermis. Exp Dermatol 2018, 27, 1084–1091. [CrossRef]
- Schön MP, Krahn T, Schön M, Rodriguez ML, Antonicek H, Schultz JE, Ludwig RJ, Zollner TM, Bischoff E, Bremm KD, et al.: Efomycine M, a new specific inhibitor of selectin, impairs leukocyte adhesion and alleviates cutaneous inflammation. Nat Med 2002, 8, 366–372. [CrossRef]
- Senra L, Mylonas A, Kavanagh RD, Fallon PG, Conrad C, Borowczyk-Michalowska J, Wrobel LJ, Kaya G, Yawalkar N, Boehncke WH, Brembilla NC: IL-17E (IL-25) Enhances Innate Immune Responses during Skin Inflammation. J Invest Dermatol 2019, 139, 1732–1742.e1717. [CrossRef]
- Lin AM, Rubin CJ, Khandpur R, Wang JY, Riblett M, Yalavarthi S, Villanueva EC, Shah P, Kaplan MJ, Bruce AT: Mast cells and neutrophils release IL-17 through extracellular trap formation in psoriasis. J Immunol 2011, 187, 490–500. [CrossRef]
- Herster F, Bittner Z, Archer NK, Dickhöfer S, Eisel D, Eigenbrod T, Knorpp T, Schneiderhan-Marra N, Löffler MW, Kalbacher H, et al.: Neutrophil extracellular trap-associated RNA and LL37 enable self-amplifying inflammation in psoriasis. Nat Commun 2020, 11, 105. [CrossRef]
- Shao S, Fang H, Dang E, Xue K, Zhang J, Li B, Qiao H, Cao T, Zhuang Y, Shen S, et al.: Neutrophil Extracellular Traps Promote Inflammatory Responses in Psoriasis via Activating Epidermal TLR4/IL-36R Crosstalk. Front Immunol 2019, 10, 746. [CrossRef]
- Ma F, Plazyo O, Billi AC, Tsoi LC, Xing X, Wasikowski R, Gharaee-Kermani M, Hile G, Jiang Y, Harms PW, et al: Single cell and spatial sequencing define processes by which keratinocytes and fibroblasts amplify inflammatory responses in psoriasis. Nat Commun 2023, 14, 3455. [CrossRef]
- Hu SC, Yu HS, Yen FL, Lin CL, Chen GS, Lan CC: Neutrophil extracellular trap formation is increased in psoriasis and induces human β-defensin-2 production in epidermal keratinocytes. Sci Rep 2016, 6, 31119. [CrossRef]
- Shao S, Fang H, Zhang J, Jiang M, Xue K, Ma J, Zhang J, Lei J, Zhang Y, Li B, et al.: Neutrophil exosomes enhance the skin autoinflammation in generalized pustular psoriasis via activating keratinocytes. Faseb j 2019, 33, 6813–6828. [CrossRef]
- Liu XT, Shi ZR, Lu SY, Hong D, Qiu XN, Tan GZ, Xiong H, Guo Q, Wang L: Enhanced Migratory Ability of Neutrophils Toward Epidermis Contributes to the Development of Psoriasis via Crosstalk With Keratinocytes by Releasing IL-17A. Front Immunol 2022, 13, 817040. [CrossRef] [PubMed]
- Chen J, Zhu Z, Li Q, Lin Y, Dang E, Meng H, Sha N, Bai H, Wang G, An S, Shao S: Neutrophils Enhance Cutaneous Vascular Dilation and Permeability to Aggravate Psoriasis by Releasing Matrix Metallopeptidase 9. J Invest Dermatol 2021, 141, 787–799. [CrossRef] [PubMed]
- Lin CY, Yu HP, Chang YT, Lin ZC, Alalaiwe A, Hwang TL, Fang JY: Targeting anti-inflammatory immunonanocarriers to human and murine neutrophils via the Ly6 antigen for psoriasiform dermatitis alleviation. Biomater Sci 2023, 11, 873–893. [CrossRef] [PubMed]
- Lowes MA, Chamian F, Abello MV, Fuentes-Duculan J, Lin SL, Nussbaum R, Novitskaya I, Carbonaro H, Cardinale I, Kikuchi T, et al.: Increase in TNF-alpha and inducible nitric oxide synthase-expressing dendritic cells in psoriasis and reduction with efalizumab (anti-CD11a). Proc Natl Acad Sci U S A 2005, 102, 19057–19062. [CrossRef]
- Terhorst D, Chelbi R, Wohn C, Malosse C, Tamoutounour S, Jorquera A, Bajenoff M, Dalod M, Malissen B, Henri S: Dynamics and Transcriptomics of Skin Dendritic Cells and Macrophages in an Imiquimod-Induced, Biphasic Mouse Model of Psoriasis. J Immunol 2015, 195, 4953–4961. [CrossRef]
- Cheng JB, Sedgewick AJ, Finnegan AI, Harirchian P, Lee J, Kwon S, Fassett MS, Golovato J, Gray M, Ghadially R, et al: Transcriptional Programming of Normal and Inflamed Human Epidermis at Single-Cell Resolution. Cell Rep 2018, 25, 871–883. [CrossRef]
- Zaba LC, Fuentes-Duculan J, Eungdamrong NJ, Abello MV, Novitskaya I, Pierson KC, Gonzalez J, Krueger JG, Lowes MA: Psoriasis is characterized by accumulation of immunostimulatory and Th1/Th17 cell-polarizing myeloid dendritic cells. J Invest Dermatol 2009, 129, 79–88. [CrossRef]
- Rebane A, Zimmermann M, Aab A, Baurecht H, Koreck A, Karelson M, Abram K, Metsalu T, Pihlap M, Meyer N, et al.: Mechanisms of IFN-γ-induced apoptosis of human skin keratinocytes in patients with atopic dermatitis. J Allergy Clin Immunol 2012, 129, 1297–1306. [CrossRef]
- Nestle FO, Conrad C, Tun-Kyi A, Homey B, Gombert M, Boyman O, Burg G, Liu YJ, Gilliet M: Plasmacytoid predendritic cells initiate psoriasis through interferon-alpha production. J Exp Med 2005, 202, 135–143. [CrossRef] [PubMed]
- Zhou Y, Xu F, Chen XY, Yan BX, Wang ZY, Chen SQ, Zheng M, Man XY: The epidermal immune microenvironment plays a dominant role in psoriasis development, as revealed by mass cytometry. Cell Mol Immunol 2022, 19, 1400–1413. [CrossRef] [PubMed]
- Borek I, Köffel R, Feichtinger J, Spies M, Glitzner-Zeis E, Hochgerner M, Sconocchia T, Krump C, Tam-Amersdorfer C, Passegger C, et al.: BMP7 aberrantly induced in the psoriatic epidermis instructs inflammation-associated Langerhans cells. J Allergy Clin Immunol 2020, 145, 1194–1207.e1111. [CrossRef] [PubMed]
- Yoshiki R, Kabashima K, Honda T, Nakamizo S, Sawada Y, Sugita K, Yoshioka H, Ohmori S, Malissen B, Tokura Y, Nakamura M: IL-23 from Langerhans cells is required for the development of imiquimod-induced psoriasis-like dermatitis by induction of IL-17A-producing γδ T cells. J Invest Dermatol 2014, 134, 1912–1921.
- Zheng T, Zhao W, Li H, Xiao S, Hu R, Han M, Liu H, Liu Y, Otsu K, Liu X, Huang G: p38α signaling in Langerhans cells promotes the development of IL-17-producing T cells and psoriasiform skin inflammation. Sci Signal 2018, 11.
- Martini E, Wikén M, Cheuk S, Gallais Sérézal I, Baharom F, Ståhle M, Smed-Sörensen A, Eidsmo L: Dynamic Changes in Resident and Infiltrating Epidermal Dendritic Cells in Active and Resolved Psoriasis. J Invest Dermatol 2017, 137, 865–873. [CrossRef]
- Fujita H, Shemer A, Suárez-Fariñas M, Johnson-Huang LM, Tintle S, Cardinale I, Fuentes-Duculan J, Novitskaya I, Carucci JA, Krueger JG, Guttman-Yassky E: Lesional dendritic cells in patients with chronic atopic dermatitis and psoriasis exhibit parallel ability to activate T-cell subsets. J Allergy Clin Immunol 2011, 128, e571–e512.
- Wang Y, Edelmayer R, Wetter J, Salte K, Gauvin D, Leys L, Paulsboe S, Su Z, Weinberg I, Namovic M, et al.: Monocytes/Macrophages play a pathogenic role in IL-23 mediated psoriasis-like skin inflammation. Sci Rep 2019, 9, 5310. [CrossRef] [PubMed]
- Wang H, Peters T, Kess D, Sindrilaru A, Oreshkova T, Van Rooijen N, Stratis A, Renkl AC, Sunderkötter C, Wlaschek M, et al.: Activated macrophages are essential in a murine model for T cell-mediated chronic psoriasiform skin inflammation. J Clin Invest 2006, 116, 2105–2114. [CrossRef] [PubMed]
- Stratis A, Pasparakis M, Rupec RA, Markur D, Hartmann K, Scharffetter-Kochanek K, Peters T, van Rooijen N, Krieg T, Haase I: Pathogenic role for skin macrophages in a mouse model of keratinocyte-induced psoriasis-like skin inflammation. J Clin Invest 2006, 116, 2094–2104. [CrossRef]
- Erbel C, Akhavanpoor M, Okuyucu D, Wangler S, Dietz A, Zhao L, Stellos K, Little KM, Lasitschka F, Doesch A, et al.: IL-17A influences essential functions of the monocyte/macrophage lineage and is involved in advanced murine and human atherosclerosis. J Immunol 2014, 193, 4344–4355. [CrossRef]
- Sieminska I, Pieniawska M, Grzywa TM: The Immunology of Psoriasis-Current Concepts in Pathogenesis. Clin Rev Allergy Immunol 2024, 66, 164–191. [CrossRef]
- Kolkhir P, Elieh-Ali-Komi D, Metz M, Siebenhaar F, Maurer M: Understanding human mast cells: lesson from therapies for allergic and non-allergic diseases. Nat Rev Immunol 2022, 22, 294–308. [CrossRef] [PubMed]
- Galli SJ, Gaudenzio N, Tsai M: Mast Cells in Inflammation and Disease: Recent Progress and Ongoing Concerns. Annu Rev Immunol 2020, 38, 49–77. [CrossRef]
- Cheung KL, Jarrett R, Subramaniam S, Salimi M, Gutowska-Owsiak D, Chen YL, Hardman C, Xue L, Cerundolo V, Ogg G: Psoriatic T cells recognize neolipid antigens generated by mast cell phospholipase delivered by exosomes and presented by CD1a. J Exp Med 2016, 213, 2399–2412. [CrossRef] [PubMed]
- Zhou T, Du X, Zhang L, Zheng Y, Jia T, Song X, Che D, Geng S: Suprabasin-derived polypeptides: SBSN(50–63) induces inflammatory response via TLR4-mediated mast cell activation. Inflammopharmacology 2023, 31, 1329–1339. [CrossRef]
- Ji YZ, Liu SR: Koebner phenomenon leading to the formation of new psoriatic lesions: evidences and mechanisms. Biosci Rep 2019, 39.
- Ribot JC, Lopes N, Silva-Santos B: γδ T cells in tissue physiology and surveillance. Nat Rev Immunol 2021, 21, 221–232. [CrossRef]
- Cai Y, Xue F, Quan C, Qu M, Liu N, Zhang Y, Fleming C, Hu X, Zhang HG, Weichselbaum R, et al.: A Critical Role of the IL-1β-IL-1R Signaling Pathway in Skin Inflammation and Psoriasis Pathogenesis. J Invest Dermatol 2019, 139, 146–156. [CrossRef]
- Laggner U, Di Meglio P, Perera GK, Hundhausen C, Lacy KE, Ali N, Smith CH, Hayday AC, Nickoloff BJ, Nestle FO: Identification of a novel proinflammatory human skin-homing Vγ9Vδ2 T cell subset with a potential role in psoriasis. J Immunol 2011, 187, 2783–2793. [CrossRef]
- Peng P, Lou Y, Wang S, Wang J, Zhang Z, Du P, Zheng J, Liu P, Xu LX: Activated NK cells reprogram MDSCs via NKG2D-NKG2DL and IFN-γ to modulate antitumor T-cell response after cryo-thermal therapy. J Immunother Cancer 2022, 10.
- Choi Y, Saron WA, O’Neill A, Senanayake M, Wilder-Smith A, Rathore AP, St John AL: NKT cells promote Th1 immune bias to dengue virus that governs long-term protective antibody dynamics. J Clin Invest 2024, 134.
- Venken K, Jacques P, Mortier C, Labadia ME, Decruy T, Coudenys J, Hoyt K, Wayne AL, Hughes R, Turner M, et al.: RORγt inhibition selectively targets IL-17 producing iNKT and γδ-T cells enriched in Spondyloarthritis patients. Nat Commun 2019, 10, 9. [CrossRef] [PubMed]
- Hu Y, Chen Y, Chen Z, Zhang X, Guo C, Yu Z, Xu P, Sun L, Zhou X, Gong Y, et al: Dysregulated Peripheral Invariant Natural Killer T Cells in Plaque Psoriasis Patients. Front Cell Dev Biol 2021, 9, 799560.
- Liu P, Peng C, Chen X, Wu L, Yin M, Li J, Qin Q, Kuang Y, Zhu W: Acitretin Promotes the Differentiation of Myeloid-Derived Suppressor Cells in the Treatment of Psoriasis. Front Med (Lausanne) 2021, 8, 625130.
- Sarkar D, Pramanik A, Das D, Bhattacharyya S: Shifting phenotype and differentiation of CD11b(+)Gr1(+) immature heterogeneous myeloid derived adjuster cells support inflammation and induce regulators of IL17A in imiquimod induced psoriasis. Inflamm Res 2024, 73, 1581–1599. [CrossRef] [PubMed]
- Jovanovic DV, Di Battista JA, Martel-Pelletier J, Jolicoeur FC, He Y, Zhang M, Mineau F, Pelletier JP: IL-17 stimulates the production and expression of proinflammatory cytokines, IL-beta and TNF-alpha, by human macrophages. J Immunol 1998, 160, 3513–3521. [CrossRef]
- Albanesi C, Cavani A, Girolomoni G: IL-17 is produced by nickel-specific T lymphocytes and regulates ICAM-1 expression and chemokine production in human keratinocytes: synergistic or antagonist effects with IFN-gamma and TNF-alpha. J Immunol 1999, 162, 494–502. [CrossRef]
- Kawaguchi M, Kokubu F, Odaka M, Watanabe S, Suzuki S, Ieki K, Matsukura S, Kurokawa M, Adachi M, Huang SK: Induction of granulocyte-macrophage colony-stimulating factor by a new cytokine, ML-1 (IL-17F), via Raf I-MEK-ERK pathway. J Allergy Clin Immunol 2004, 114, 444–450. [CrossRef]
- Johnston A, Fritz Y, Dawes SM, Diaconu D, Al-Attar PM, Guzman AM, Chen CS, Fu W, Gudjonsson JE, McCormick TS, Ward NL: Keratinocyte overexpression of IL-17C promotes psoriasiform skin inflammation. J Immunol 2013, 190, 2252–2262. [CrossRef]
- Yeste A, Mascanfroni ID, Nadeau M, Burns EJ, Tukpah AM, Santiago A, Wu C, Patel B, Kumar D, Quintana FJ: IL-21 induces IL-22 production in CD4+ T cells. Nat Commun 2014, 5, 3753. [CrossRef]
- Hammer AM, Morris NL, Cannon AR, Khan OM, Gagnon RC, Movtchan NV, van Langeveld I, Li X, Gao B, Choudhry MA: Interleukin-22 Prevents Microbial Dysbiosis and Promotes Intestinal Barrier Regeneration Following Acute Injury. Shock 2017, 48, 657–665. [CrossRef]
- Bunte K, Beikler T: Th17 Cells and the IL-23/IL-17 Axis in the Pathogenesis of Periodontitis and Immune-Mediated Inflammatory Diseases. Int J Mol Sci 2019, 20.
- Chang HW, Yan D, Singh R, Liu J, Lu X, Ucmak D, Lee K, Afifi L, Fadrosh D, Leech J, et al: Alteration of the cutaneous microbiome in psoriasis and potential role in Th17 polarization. Microbiome 2018, 6, 154.
- Zheng Y, Danilenko DM, Valdez P, Kasman I, Eastham-Anderson J, Wu J, Ouyang W: Interleukin-22, a T(H)17 cytokine, mediates IL-23-induced dermal inflammation and acanthosis. Nature 2007, 445, 648–651. [CrossRef] [PubMed]
- Trifari S, Kaplan CD, Tran EH, Crellin NK, Spits H: Identification of a human helper T cell population that has abundant production of interleukin 22 and is distinct from T(H)-17, T(H)1 and T(H)2 cells. Nat Immunol 2009, 10, 864–871. [CrossRef]
- Schlapbach C, Gehad A, Yang C, Watanabe R, Guenova E, Teague JE, Campbell L, Yawalkar N, Kupper TS, Clark RA: Human TH9 cells are skin-tropic and have autocrine and paracrine proinflammatory capacity. Sci Transl Med 2014, 6, 219ra218.
- Singh TP, Schön MP, Wallbrecht K, Gruber-Wackernagel A, Wang XJ, Wolf P: Involvement of IL-9 in Th17-associated inflammation and angiogenesis of psoriasis. PLoS One, 2013; 8, e51752.
- Niu J, Song Z, Yang X, Zhai Z, Zhong H, Hao F: Increased circulating follicular helper T cells and activated B cells correlate with disease severity in patients with psoriasis. J Eur Acad Dermatol Venereol 2015, 29, 1791–1796. [CrossRef]
- Austin LM, Ozawa M, Kikuchi T, Walters IB, Krueger JG: The majority of epidermal T cells in Psoriasis vulgaris lesions can produce type 1 cytokines, interferon-gamma, interleukin-2, and tumor necrosis factor-alpha, defining TC1 (cytotoxic T lymphocyte) and TH1 effector populations: a type 1 differentiation bias is also measured in circulating blood T cells in psoriatic patients. J Invest Dermatol 1999, 113, 752–759.
- Guenova E, Skabytska Y, Hoetzenecker W, Weindl G, Sauer K, Tham M, Kim KW, Park JH, Seo JH, Ignatova D, et al: IL-4 abrogates T(H)17 cell-mediated inflammation by selective silencing of IL-23 in antigen-presenting cells. Proc Natl Acad Sci U S A 2015, 112, 2163–2168. [CrossRef]
- Huang C, Sun P-Y, Jiang Y, Liu Y, Liu Z, Han S-L, Wang B-S, Huang Y-X, Ren A-R, Lu, J.-F, et al.: Sensory ASIC3 channel exacerbates psoriatic inflammation via a neurogenic pathway in female mice. Nature Communications, 2024; 15.
- Baker BS, Laman JD, Powles A, van der Fits L, Voerman JS, Melief MJ, Fry L: Peptidoglycan and peptidoglycan-specific Th1 cells in psoriatic skin lesions. J Pathol 2006, 209, 174–181. [CrossRef]
- Baker BS, Powles A, Fry L: Peptidoglycan: a major aetiological factor for psoriasis? Trends Immunol 2006, 27, 545–551. [CrossRef]
- Menssen A, Trommler P, Vollmer S, Schendel D, Albert E, Gürtler L, Riethmüller G, Prinz JC: Evidence for an antigen-specific cellular immune response in skin lesions of patients with psoriasis vulgaris. J Immunol 1995, 155, 4078–4083. [CrossRef]
- Lin WJ, Norris DA, Achziger M, Kotzin BL, Tomkinson B: Oligoclonal expansion of intraepidermal T cells in psoriasis skin lesions. J Invest Dermatol 2001, 117, 1546–1553. [CrossRef] [PubMed]
- Pantelyushin S, Haak S, Ingold B, Kulig P, Heppner FL, Navarini AA, Becher B: Rorγt+ innate lymphocytes and γδ T cells initiate psoriasiform plaque formation in mice. J Clin Invest 2012, 122, 2252–2256. [CrossRef]
- Zhang YY, Lin YT, Wang L, Sun XW, Dang EL, Xue K, Zhang WG, Zhang KM, Wang G, Li B: CD8αα(+)T cells exert a pro-inflammatory role in patients with psoriasis. Skin Health Dis, 2021; 1, e64.
- Diani M, Casciano F, Marongiu L, Longhi M, Altomare A, Pigatto PD, Secchiero P, Gambari R, Banfi G, Manfredi AA, et al: Increased frequency of activated CD8(+) T cell effectors in patients with psoriatic arthritis. Sci Rep 2019, 9, 10870. [CrossRef] [PubMed]
- Di Meglio P, Villanova F, Navarini AA, Mylonas A, Tosi I, Nestle FO, Conrad C: Targeting CD8(+) T cells prevents psoriasis development. J Allergy Clin Immunol 2016, 138, 274–276.e276.
- Cheuk S, Wikén M, Blomqvist L, Nylén S, Talme T, Ståhle M, Eidsmo L: Epidermal Th22 and Tc17 cells form a localized disease memory in clinically healed psoriasis. J Immunol 2014, 192, 3111–3120. [CrossRef]
- Migayron L, Merhi R, Seneschal J, Boniface K: Resident memory T cells in nonlesional skin and healed lesions of patients with chronic inflammatory diseases: Appearances can be deceptive. J Allergy Clin Immunol 2024, 153, 606–614. [CrossRef] [PubMed]
- Deng G, Zhang Y, Song J, Zhang Y, Zheng Q, Luo Y, Fei X, Yang Y, Kuai L, Li B, Luo Y: The role and therapeutic strategies for tissue-resident memory T cells, central memory T cells, and effector memory T cells in psoriasis. Immunology 2024, 173, 470–480. [CrossRef]
- Matos TR, O’Malley JT, Lowry EL, Hamm D, Kirsch IR, Robins HS, Kupper TS, Krueger JG, Clark RA: Clinically resolved psoriatic lesions contain psoriasis-specific IL-17-producing αβ T cell clones. J Clin Invest 2017, 127, 4031–4041. [CrossRef] [PubMed]
- Lee BH, Bang YJ, Lim SH, Kang SJ, Kim SH, Kim-Schulze S, Park CG, Kim HJ, Kim TG: High-dimensional profiling of regulatory T cells in psoriasis reveals an impaired skin-trafficking property. EBioMedicine 2024, 100, 104985.
- Sugiyama H, Gyulai R, Toichi E, Garaczi E, Shimada S, Stevens SR, McCormick TS, Cooper KD: Dysfunctional blood and target tissue CD4+CD25high regulatory T cells in psoriasis: mechanism underlying unrestrained pathogenic effector T cell proliferation. J Immunol 2005, 174, 164–173. [CrossRef]
- Yang L, Li B, Dang E, Jin L, Fan X, Wang G: Impaired function of regulatory T cells in patients with psoriasis is mediated by phosphorylation of STAT3. J Dermatol Sci 2016, 81, 85–92. [CrossRef]
- Sivasami P, Elkins C, Diaz-Saldana PP, Goss K, Peng A, Hamersky Mt, Bae J, Xu M, Pollack BP, Horwitz EM, et al: Obesity-induced dysregulation of skin-resident PPARγ(+) Treg cells promotes IL-17A-mediated psoriatic inflammation. Immunity 2023, 56, 1844–1861.e1846. [CrossRef]
- Lu J, Ding Y, Yi X, Zheng J: CD19+ B cell subsets in the peripheral blood and skin lesions of psoriasis patients and their correlations with disease severity. Braz J Med Biol Res 2016, 49, e5374.
- Jones DA, Yawalkar N, Suh KY, Sadat S, Rich B, Kupper TS: Identification of autoantigens in psoriatic plaques using expression cloning. J Invest Dermatol 2004, 123, 93–100. [CrossRef]
- Guarneri C, Aguennouz M, Guarneri F, Polito F, Benvenga S, Cannavò SP: Autoimmunity to heterogeneous nuclear ribonucleoprotein A1 in psoriatic patients and correlation with disease severity. J Dtsch Dermatol Ges 2018, 16, 1103–1107. [CrossRef]
- Yunusbaeva M, Valiev R, Bilalov F, Sultanova Z, Sharipova L, Yunusbayev B: Psoriasis patients demonstrate HLA-Cw*06:02 allele dosage-dependent T cell proliferation when treated with hair follicle-derived keratin 17 protein. Sci Rep 2018, 8, 6098. [CrossRef]
- Lande R, Botti E, Jandus C, Dojcinovic D, Fanelli G, Conrad C, Chamilos G, Feldmeyer L, Marinari B, Chon S, et al.: The antimicrobial peptide LL37 is a T-cell autoantigen in psoriasis. Nat Commun 2014, 5, 5621. [CrossRef] [PubMed]
- Orbai AM, Fiorentino D, Perin J, Darrah E, Yang Q, Gutierrez-Alamillo L, Bingham CO, Petri M, Rosen A, Casciola-Rosen L: SOX-5 Transcription Factor: a Novel Psoriatic Autoantigen Preferentially Found in Women. ACR Open Rheumatol 2024, 6, 807–819. [CrossRef] [PubMed]
- Wang SY, An WH, Wang ZS, Wang WL, Zhang B, Xu KL, Guo SL, Gao M, Li B, Huang L, et al: Incidentally cured psoriasis in a patient with refractory/relapsed diffuse large B-cell lymphoma receiving CD19 CAR-T cell therapy: a case report. Front Immunol 2024, 15, 1418768. [CrossRef]
- Mizumaki K, Horii M, Kano M, Komuro A, Matsushita T: Suppression of IL-23-mediated psoriasis-like inflammation by regulatory B cells. Sci Rep 2021, 11, 2106. [CrossRef]
- Mavropoulos A, Zafiriou E, Simopoulou T, Brotis AG, Liaskos C, Roussaki-Schulze A, Katsiari CG, Bogdanos DP, Sakkas LI: Apremilast increases IL-10-producing regulatory B cells and decreases proinflammatory T cells and innate cells in psoriatic arthritis and psoriasis. Rheumatology (Oxford) 2019, 58, 2240–2250.
- Mavropoulos A, Varna A, Zafiriou E, Liaskos C, Alexiou I, Roussaki-Schulze A, Vlychou M, Katsiari C, Bogdanos DP, Sakkas LI: IL-10 producing Bregs are impaired in psoriatic arthritis and psoriasis and inversely correlate with IL-17- and IFNγ-producing T cells. Clin Immunol 2017, 184, 33–41.
- Menter A, Korman NJ, Elmets CA, Feldman SR, Gelfand JM, Gordon KB, Gottlieb A, Koo JY, Lebwohl M, Leonardi CL, et al: Guidelines of care for the management of psoriasis and psoriatic arthritis: Section 6. Guidelines of care for the treatment of psoriasis and psoriatic arthritis: case-based presentations and evidence-based conclusions. J Am Acad Dermatol 2011, 65, 137–174.
- Nast A, Boehncke WH, Mrowietz U, Ockenfels HM, Philipp S, Reich K, Rosenbach T, Sammain A, Schlaeger M, Sebastian M, et al.: S3—Guidelines on the treatment of psoriasis vulgaris (English version). Update. J Dtsch Dermatol Ges, 2012; 10, (Suppl 2), S1–95.
- Jones SA, Perera DN, Fan H, Russ BE, Harris J, Morand EF: GILZ regulates Th17 responses and restrains IL-17-mediated skin inflammation. J Autoimmun 2015, 61, 73–80. [CrossRef]
- Koo J, Cuffie CA, Tanner DJ, Bressinck R, Cornell RC, DeVillez RL, Edwards L, Breneman DL, Piacquadio DJ, Guzzo CA, Monroe EW: Mometasone furoate 0.1%-salicylic acid 5% ointment versus mometasone furoate 0.1% ointment in the treatment of moderate-to-severe psoriasis: a multicenter study. Clin Ther 1998, 20, 283–291. [CrossRef] [PubMed]
- Mason A, Mason J, Cork M, Hancock H, Dooley G: Topical treatments for chronic plaque psoriasis: an abridged Cochrane systematic review. J Am Acad Dermatol 2013, 69, 799–807. [CrossRef] [PubMed]
- Segaert S, Duvold LB: Calcipotriol cream: a review of its use in the management of psoriasis. J Dermatolog Treat 2006, 17, 327–337. [CrossRef]
- Lebwohl MG, Stein Gold L, Strober B, Papp KA, Armstrong AW, Bagel J, Kircik L, Ehst B, Hong HC, Soung J, et al: Phase 3 Trials of Tapinarof Cream for Plaque Psoriasis. N Engl J Med 2021, 385, 2219–2229. [CrossRef] [PubMed]
- Menter A, Korman NJ, Elmets CA, Feldman SR, Gelfand JM, Gordon KB, Gottlieb A, Koo JYM, Lebwohl M, Lim HW, et al: Guidelines of care for the management of psoriasis and psoriatic arthritis: Section 5. Guidelines of care for the treatment of psoriasis with phototherapy and photochemotherapy. J Am Acad Dermatol 2010, 62, 114–135.
- Rim JH, Park JY, Choe YB, Youn JI: The efficacy of calcipotriol + acitretin combination therapy for psoriasis: comparison with acitretin monotherapy. Am J Clin Dermatol 2003, 4, 507–510. [CrossRef]
- Menter A, Gottlieb A, Feldman SR, Van Voorhees AS, Leonardi CL, Gordon KB, Lebwohl M, Koo JY, Elmets CA, Korman NJ, et al: Guidelines of care for the management of psoriasis and psoriatic arthritis: Section 1. Overview of psoriasis and guidelines of care for the treatment of psoriasis with biologics. J Am Acad Dermatol 2008, 58, 826–850. [CrossRef] [PubMed]
- Gisondi P, Del Giglio M, Cotena C, Girolomoni G: Combining etanercept and acitretin in the therapy of chronic plaque psoriasis: a 24-week, randomized, controlled, investigator-blinded pilot trial. Br J Dermatol 2008, 158, 1345–1349. [CrossRef]
- Katz HI, Waalen J, Leach EE: Acitretin in psoriasis: an overview of adverse effects. J Am Acad Dermatol, 1999; 41, S7–s12.
- Goldfarb MT, Ellis CN, Gupta AK, Tincoff T, Hamilton TA, Voorhees JJ: Acitretin improves psoriasis in a dose-dependent fashion. J Am Acad Dermatol 1988, 18, 655–662. [CrossRef] [PubMed]
- de Eusebio E, Armario-Hita JC, de Miquel VA: Treatment of psoriasis: focus on clinic-based management with infliximab. Am J Clin Dermatol, 2014; 15, (Suppl 1), S5–16.
- Ho VC, Griffiths CE, Albrecht G, Vanaclocha F, León-Dorantes G, Atakan N, Reitamo S, Ohannesson A, Mørk NJ, Clarke P, et al: Intermittent short courses of cyclosporin (Neoral(R)) for psoriasis unresponsive to topical therapy: a 1-year multicentre, randomized study. The PISCES Study Group. Br J Dermatol 1999, 141, 283–291. [CrossRef]
- Ho VC, Griffiths CE, Berth-Jones J, Papp KA, Vanaclocha F, Dauden E, Beard A, Puvanarajan L, Paul C: Intermittent short courses of cyclosporine microemulsion for the long-term management of psoriasis: a 2-year cohort study. J Am Acad Dermatol 2001, 44, 643–651. [CrossRef]
- Lin VW, Ringold S, Devine EB: Comparison of Ustekinumab With Other Biological Agents for the Treatment of Moderate to Severe Plaque Psoriasis: A Bayesian Network Meta-analysis. Arch Dermatol 2012, 148, 1403–1410. [CrossRef]
- Ortonne JP, van de Kerkhof PC, Prinz JC, Bieber T, Lahfa M, Rubins A, Wozel G, Lorette G: 0.3% Tacrolimus gel and 0.5% Tacrolimus cream show efficacy in mild to moderate plaque psoriasis: Results of a randomized, open-label, observer-blinded study. Acta Derm Venereol 2006, 86, 29–33. [CrossRef] [PubMed]
- Remitz A, Reitamo S, Erkko P, Granlund H, Lauerma AI: Tacrolimus ointment improves psoriasis in a microplaque assay. Br J Dermatol 1999, 141, 103–107. [CrossRef] [PubMed]
- Douglas WS, Poulin Y, Decroix J, Ortonne JP, Mrowietz U, Gulliver W, Krogstad AL, Larsen FG, Iglesias L, Buckley C, Bibby AJ: A new calcipotriol/betamethasone formulation with rapid onset of action was superior to monotherapy with betamethasone dipropionate or calcipotriol in psoriasis vulgaris. Acta Derm Venereol 2002, 82, 131–135.
- Elias KM, Laurence A, Davidson TS, Stephens G, Kanno Y, Shevach EM, O’Shea JJ: Retinoic acid inhibits Th17 polarization and enhances FoxP3 expression through a Stat-3/Stat-5 independent signaling pathway. Blood 2008, 111, 1013–1020. [CrossRef]
- Jairath V, Acosta Felquer ML, Cho RJ: IL-23 inhibition for chronic inflammatory disease. Lancet 2024, 404, 1679–1692. [CrossRef]
- Mease PJ, Chohan S, Fructuoso FJG, Luggen ME, Rahman P, Raychaudhuri SP, Chou RC, Mendelsohn AM, Rozzo SJ, Gottlieb A: Efficacy and safety of tildrakizumab in patients with active psoriatic arthritis: results of a randomised, double-blind, placebo-controlled, multiple-dose, 52-week phase IIb study. Ann Rheum Dis 2021, 80, 1147–1157.
- Gordon KB, Blauvelt A, Papp KA, Langley RG, Luger T, Ohtsuki M, Reich K, Amato D, Ball SG, Braun DK, et al: Phase 3 Trials of Ixekizumab in Moderate-to-Severe Plaque Psoriasis. N Engl J Med 2016, 375, 345–356. [CrossRef]
- Reich K, Sullivan J, Arenberger P, Jazayeri S, Mrowietz U, Augustin M, Elewski B, You R, Regnault P, Frueh JA: Secukinumab shows high and sustained efficacy in nail psoriasis: 2.5-year results from the randomized placebo-controlled TRANSFIGURE study. Br J Dermatol 2021, 184, 425–436. [CrossRef] [PubMed]
- Warren RB, Blauvelt A, Bagel J, Papp KA, Yamauchi P, Armstrong A, Langley RG, Vanvoorden V, De Cuyper D, Cioffi C, et al.: Bimekizumab versus Adalimumab in Plaque Psoriasis. N Engl J Med 2021, 385, 130–141. [CrossRef] [PubMed]
- Reich K, Warren RB, Lebwohl M, Gooderham M, Strober B, Langley RG, Paul C, De Cuyper D, Vanvoorden V, Madden C, et al.: Bimekizumab versus Secukinumab in Plaque Psoriasis. N Engl J Med 2021, 385, 142–152. [CrossRef]
- Morita A, Strober B, Burden AD, Choon SE, Anadkat MJ, Marrakchi S, Tsai TF, Gordon KB, Thaçi D, Zheng M, et al.: Efficacy and safety of subcutaneous spesolimab for the prevention of generalised pustular psoriasis flares (Effisayil 2): an international, multicentre, randomised, placebo-controlled trial. Lancet 2023, 402, 1541–1551. [CrossRef]
- Yang W, Bai X, Jia X, Li H, Min J, Li H, Zhang H, Zhou J, Zhao Y, Liu W, et al: The binding of extracellular cyclophilin A to ACE2 and CD147 triggers psoriasis-like inflammation. J Autoimmun 2024, 148, 103293. [CrossRef] [PubMed]
- Bian J, Liu R, Fan T, Liao L, Wang S, Geng W, Wang T, Shi W, Ruan Q: miR-340 Alleviates Psoriasis in Mice through Direct Targeting of IL-17A. The Journal of Immunology 2018, 201, 1412–1420. [CrossRef] [PubMed]
- Chen L, Huang D, Huang Z, Liu X, He M, Luo M, Tang Z, Tan G, Guo Q, Xiong H: Decreased HMGCS1 inhibits proliferation and inflammatory response of keratinocytes and ameliorates imiquimod-induced psoriasis via the STAT3/IL-23 axis. Int Immunopharmacol 2024, 133, 112033. [CrossRef]
- Whitehead KA, Langer R, Anderson DG: Knocking down barriers: advances in siRNA delivery. Nat Rev Drug Discov 2009, 8, 129–138. [CrossRef]
- Tang Q, Khvorova A: RNAi-based drug design: considerations and future directions. Nat Rev Drug Discov 2024, 23, 341–364. [CrossRef]
- Boakye CHA, Patel K, Doddapaneni R, Bagde A, Marepally S, Singh M: Novel amphiphilic lipid augments the co-delivery of erlotinib and IL36 siRNA into the skin for psoriasis treatment. J Control Release 2017, 246, 120–132. [CrossRef]
- Song Y, Li J, Wu Y: Evolving understanding of autoimmune mechanisms and new therapeutic strategies of autoimmune disorders. Signal Transduct Target Ther 2024, 9, 263. [CrossRef]
- Zhang A, Zhang G, Yang H, Gong B, Li S, Wei N, Xue H, Wei H, Wang J, Qiu S: Treatment of pro-B acute lymphoblastic leukemia and severe plaque psoriasis with anti-CD19 CAR T cells: a case report. Frontiers in Immunology 2025, 16.
- Wang X, Wu X, Tan B, Zhu L, Zhang Y, Lin L, Xiao Y, Sun A, Wan X, Liu S, et al: Allogeneic CD19-targeted CAR-T therapy in patients with severe myositis and systemic sclerosis. Cell 2024, 187, 4890–4904.e4899. [CrossRef] [PubMed]
- Fransson M, Piras E, Burman J, Nilsson B, Essand M, Lu B, Harris RA, Magnusson PU, Brittebo E, Loskog AS: CAR/FoxP3-engineered T regulatory cells target the CNS and suppress EAE upon intranasal delivery. J Neuroinflammation 2012, 9, 112.
- Zhang W, Chen Y, Zhao Z, Zheng H, Wang S, Liao Z, Sheng T, Zhao S, Hou W, Yu X, et al.: Adoptive T(reg) therapy with metabolic intervention via perforated microneedles ameliorates psoriasis syndrome. Sci Adv, 2023; 9, eadg6007.
- Rosenzwajg M, Lorenzon R, Cacoub P, Pham HP, Pitoiset F, El Soufi K, C RI, Bernard C, Aractingi S, Banneville B, et al: Immunological and clinical effects of low-dose interleukin-2 across 11 autoimmune diseases in a single, open clinical trial. Ann Rheum Dis 2019, 78, 209–217. [CrossRef]
- Chen W, Ren G, Zuo K, Huang X: Complete remission of both immunoglobulin light chain amyloidosis and psoriasis after autologous hematopoietic stem cell transplantation: A case report. Medicine (Baltimore), 2018; 97, e13589.
- Kaffenberger BH, Wong HK, Jarjour W, Andritsos LA: Remission of psoriasis after allogeneic, but not autologous, hematopoietic stem-cell transplantation. J Am Acad Dermatol 2013, 68, 489–492. [CrossRef] [PubMed]
- Ahn H, Lee SY, Jung WJ, Pi J, Lee KH: Psoriasis treatment using minimally manipulated umbilical cord-derived mesenchymal stem cells: A case report. World J Clin Cases 2021, 9, 6798–6803. [CrossRef]
- Cheng L, Wang S, Peng C, Zou X, Yang C, Mei H, Li C, Su X, Xiao N, Ouyang Q, et al: Human umbilical cord mesenchymal stem cells for psoriasis: a phase 1/2a, single-arm study. Signal Transduct Target Ther 2022, 7, 263. [CrossRef]
- Yao D, Ye S, He Z, Huang Y, Deng J, Wen Z, Chen X, Li H, Han Q, Deng H, et al.: Adipose-derived mesenchymal stem cells (AD-MSCs) in the treatment for psoriasis: results of a single-arm pilot trial. Ann Transl Med 2021, 9, 1653. [CrossRef]
- Tang L, Yao D, He Z, Ye S, Chen X, Huang Y, Han Q, Zeng X, Zheng X, Liu T, et al: Distinct adaptive immune receptor feature of adipose-derived mesenchymal stem cells (AD-MSCs) treatment of psoriasis. Arch Dermatol Res 2024, 316, 542. [CrossRef]
- Seetharaman R, Mahmood A, Kshatriya P, Patel D, Srivastava A: Mesenchymal Stem Cell Conditioned Media Ameliorate Psoriasis Vulgaris: A Case Study. Case Rep Dermatol Med 2019, 2019, 8309103.
- Shao H, Chen J: Extracellular Vesicles from miR-146a Overexpressing Mesenchymal Stem Cells Attenuate Imiquimod-Induced Psoriasis by Regulating Cytokine Expression. Iran J Immunol 2025, 22, 119–130.
- Mohseni Meybodi MA, Nilforoushzadeh MA, KhandanDezfully N, Mansouri P: The safety and efficacy of adipose tissue-derived exosomes in treating mild to moderate plaque psoriasis: A clinical study. Life Sci 2024, 353, 122915. [CrossRef] [PubMed]
- Ou M, Cao J, Luo R, Zhu B, Miao R, Yu L, Wang X, Li W, Fu Y, Zhang J, et al.: Drug-loaded microneedle patches containing regulatory T cell-derived exosomes for psoriasis treatment. Acta Biomater 2025, 198, 452–466. [CrossRef] [PubMed]
- Chandran NS, Bhupendrabhai MN, Tan TT, Zhang B, Lim SK, Choo ABH, Lai RC: A phase 1, open-label study to determine safety and tolerability of the topical application of mesenchymal stem/stromal cell (MSC) exosome ointment to treat psoriasis in healthy volunteers. Cytotherapy 2025, 27, 633–641.
- Bucci L, Hagen M, Rothe T, Raimondo MG, Fagni F, Tur C, Wirsching A, Wacker J, Wilhelm A, Auger JP, et al: Bispecific T cell engager therapy for refractory rheumatoid arthritis. Nat Med 2024, 30, 1593–1601. [CrossRef]
- Gao C, Cai Y, Wu X, Song J, Zheng Q, Wang M, Luo Y, Luo Y, Fei X, Zhang Y, et al.: CRISPR/Cas9-Mediated Knockout and Overexpression Studies Unveil the Role of PD-L1 in Immune Modulation in a Psoriasis-like Mouse Model. Inflammation, 2025.
- Tan E, Wan T, Pan Q, Duan J, Zhang S, Wang R, Gao P, Lv J, Wang H, Li D, et al: Dual-responsive nanocarriers for efficient cytosolic protein delivery and CRISPR-Cas9 gene therapy of inflammatory skin disorders. Sci Adv 2024, 10, eadl4336. [CrossRef] [PubMed]
- Xiang Y, Jiang Y, Wang Z, Wang X, Ma L, Ding Y, Rui B, Zhao C, Li X, Xu M, et al.: Targeting KAT8 alleviates self-RNA-driven skin inflammation by modulating histone H4 lysine 16 acetylation in psoriasis. Cell Death Differ, 2025.
- Dhillon-LaBrooy A, Braband KL, Tantawy E, Rampoldi F, Kao YS, Boukhallouk F, Velasquez LN, Mamareli P, Silva L, Damasceno LEA, et al: Inhibition of Mitochondrial Translation Ameliorates Imiquimod-Induced Psoriasis-Like Skin Inflammation by Targeting Vγ4+ γδ T Cells. J Invest Dermatol 2024, 144, 844–854.e842. [CrossRef]
- Xia X, Cao G, Sun G, Zhu L, Tian Y, Song Y, Guo C, Wang X, Zhong J, Zhou W, et al: GLS1-mediated glutaminolysis unbridled by MALT1 protease promotes psoriasis pathogenesis. J Clin Invest 2020, 130, 5180–5196. [CrossRef]
- He X, Liu R, Fan T, Huang X, Wu C, Su W, Wang T, Ruan Q: Treating Autoimmune Diseases by Targeting IL-23 with Gene-Silencing Pyrrole-Imidazole Polyamide. J Immunol 2020, 204, 2053–2063. [CrossRef]
- Hossen MM, Ma Y, Yin Z, Xia Y, Du J, Huang JY, Huang JJ, Zou L, Ye Z, Huang Z: Current understanding of CTLA-4: from mechanism to autoimmune diseases. Front Immunol 2023, 14, 1198365. [CrossRef]
- Yu C, Sonnen AF, George R, Dessailly BH, Stagg LJ, Evans EJ, Orengo CA, Stuart DI, Ladbury JE, Ikemizu S, et al.: Rigid-body ligand recognition drives cytotoxic T-lymphocyte antigen 4 (CTLA-4) receptor triggering. J Biol Chem 2011, 286, 6685–6696. [CrossRef] [PubMed]
- Hosseini A, Gharibi T, Marofi F, Babaloo Z, Baradaran B: CTLA-4: From mechanism to autoimmune therapy. Int Immunopharmacol 2020, 80, 106221. [CrossRef] [PubMed]
- Brunner HI, Tzaribachev N, Vega-Cornejo G, Louw I, Berman A, Calvo Penadés I, Antón J, Ávila-Zapata F, Cuttica R, Horneff G, et al.: Subcutaneous Abatacept in Patients With Polyarticular-Course Juvenile Idiopathic Arthritis: Results From a Phase III Open-Label Study. Arthritis Rheumatol 2018, 70, 1144–1154. [CrossRef] [PubMed]
- Lee WS, Nam KH, Kim JH, Kim WJ, Kim JE, Shin EC, Kim GR, Choi JM: Alleviating psoriatic skin inflammation through augmentation of Treg cells via CTLA-4 signaling peptide. Front Immunol 2023, 14, 1233514. [CrossRef]
- Guo W, Xu F, Zhuang Z, Liu Z, Xie J, Bai L: Ebosin Ameliorates Psoriasis-Like Inflammation of Mice via miR-155 Targeting tnfaip3 on IL-17 Pathway. Front Immunol 2021, 12, 662362. [CrossRef]
- Stenderup K, Rosada C, Dam TN, Salerno E, Belinka BA, Kachlany SC: Resolution of psoriasis by a leukocyte-targeting bacterial protein in a humanized mouse model. J Invest Dermatol 2011, 131, 2033–2039. [CrossRef]
- Gege C: RORγt inhibitors as potential back-ups for the phase II candidate VTP-43742 from Vitae Pharmaceuticals: patent evaluation of WO2016061160 and US20160122345. Expert Opin Ther Pat 2017, 27, 1–8. [CrossRef]
- Armstrong AW, Read C: Pathophysiology, Clinical Presentation, and Treatment of Psoriasis: A Review. Jama 2020, 323, 1945–1960. [CrossRef]
- Schaper-Gerhardt K, Rossbach K, Nikolouli E, Werfel T, Gutzmer R, Mommert S: The role of the histamine H(4) receptor in atopic dermatitis and psoriasis. Br J Pharmacol 2020, 177, 490–502.
- Zhu J, Yang T, Tang M, Yang Z, Pei H, Ye H, Tang Y, Cheng Z, Lin P, Chen L: Studies on the anti-psoriasis effects and its mechanism of a dual JAK2/FLT3 inhibitor flonoltinib maleate. Biomed Pharmacother 2021, 137, 111373.
- Armstrong AW, Gooderham M, Warren RB, Papp KA, Strober B, Thaçi D, Morita A, Szepietowski JC, Imafuku S, Colston E, et al.: Deucravacitinib versus placebo and apremilast in moderate to severe plaque psoriasis: Efficacy and safety results from the 52-week, randomized, double-blinded, placebo-controlled phase 3 POETYK PSO-1 trial. J Am Acad Dermatol 2023, 88, 29–39. [CrossRef]
- Deng BX, Jiang, C.Y, Wang P, Liu, W.L, Qu X, Zhao, Y.M, Chen K, Cai, N.N, & Tao, Y: Distribution and evolution patterns of TCM syndromes in psoriasis. Journal of Traditional Chinese Medicine, 2006; 770–772.
- Liu AM, Zhang, BX, Zhao W, & Xu, S. D: Discussion on etiology and pathogenesis of psoriasis vulgaris. Journal of Beijing University of Traditional Chinese Medicine 2021, 44, 266–271.
- Huang G, Yuan X, Hu, C.X, Fu, J.J, Shi, H.Y, Xin Q, Yang M, & Gong, L. P: Study on the effect of Xijiao Dihuang Decoction on intestinal Th17/Treg imbalance in psoriasis mice based on the gut-immune-skin axis. Clinical Journal of Chinese Medicine 2023, 35, 1569–1572.
- Chen H, & Wang, S. P: Therapeutic effect of Compound Indigo Naturalis Capsule on psoriasis vulgaris and its influence on serum IL-2 and IL-8 levels. Journal of Chinese Medicinal Materials 2004, 885–886.
- Feng J, Xu, H.Q, & Su, B. S: Effect of Compound Qingdai Capsule on c-myc expression in epidermal keratinocytes of psoriasis patients. Chinese Journal of Integrated Traditional and Western Medicine 1996, 146–148.
- Lei M, Liu R, Yao B, Hu, Y.J, Wang, W.J, & Zhang, X. J: Expression of p38MAPK/Th17 signaling pathway-related cytokines in psoriasis vulgaris patients and the regulatory effect of Modified Gentian Liver-Draining Decoction. Lishizhen Medicine and Materia Medica Research 2020, 31, 1342–1347.
- Chen LH: Clinical observation on Compound Indigo Naturalis Ointment for treating psoriasis vulgaris. Journal of Gansu College of Traditional Chinese Medicine 2010, 27, 45–47.
- Zou JH, Gong LP, Huang G, Hu, C.X, Yan J, & Zhang, F. Y: Meta-analysis of acupuncture efficacy for psoriasis vulgaris. Hunan Journal of Traditional Chinese Medicine 2020, 36, 127–130.
- Li B, Li Z, Chen Q, Fang, W.H, & Zhang, C. H: Effects of moving cupping therapy on peripheral blood CD4+ T lymphocyte subsets in plaque psoriasis patients with blood stasis syndrome. China Journal of Traditional Chinese Medicine and Pharmacy 2021, 36, 1769–1772.
- Zhou Y, Yang M, Bai, X.P, Gao Y, Han H, & Guan, H. W: Efficacy of NB-UVB phototherapy combined with self-prepared Cool-Blood Anti-Itching decoction herbal bath for psoriasis vulgaris and its effects on hemorheology. Modern Journal of Integrated Traditional Chinese and Western Medicine 2017, 26, 3023–3026.
- Feng F, Wang Y, Zhao, J.X, Di, T.T, Meng, Y.J, Chen, Z.X, Qi C, Hu, X.Q, Wang, Y.Z, & Li, P: Effects of fire needle therapy on psoriasiform skin lesions and STAT3 pathway in imiquimod-induced mice. Chinese Acupuncture & Moxibustion 2022, 42, 541–548.
- Song X, Tang, S.W, Jiang, W.C, Yang Y, Wang, Q.L, & Xie, S. Q: Chinese herbal bath combined with NB-UVB phototherapy for stable plaque psoriasis and its impact on patients’ quality of life. Chinese Journal of Dermatovenereology 2017, 31, 757–759.
- Guang YJ: Clinical observation of NB-UVB combined with Chinese herbal fumigation for psoriasis vulgaris. Medical Diet and Health 2020, 18, 28–30.
- Chen CF, Zheng, YZ, Jia, L.Y, & Yu, T. G: Clinical study of Shugan Jieyu Capsule in treating psoriasis vulgaris. Chinese Journal of Clinical Pharmacology 2017, 33, 222–225.
- Dai XR, Peng, FF, Miao, Y.X, Dong M, & Jiang, G. F: Nursing observation on the interventional effects of Five-Element Jue-tone music therapy for psoriasis patients with Qi-stagnation constitution. Journal of Hunan University of Chinese Medicine 2018, 38, 715–718.
- Langley RG, Krueger GG, Griffiths CE: Psoriasis: epidemiology, clinical features, and quality of life. Ann Rheum Dis, 2005; 64, (Suppl. 2), ii18–23; discussion ii24–15.
- Chen L, Su M, Jin Q, Wang CG, Assani I, Wang MX, Zhao SF, Lv SM, Wang JW, Sun B, et al.: Discovery of N-(2-benzyl-4-oxochroman-7-yl)-2-(5-(ethylsulfonyl) pyridin-2-yl) acetamide (b12) as a potent, selective, and orally available novel retinoic acid receptor-related orphan receptor γt inverse agonist. Bioorg Chem 2022, 119, 105483.
- Mease P, Hall S, FitzGerald O, van der Heijde D, Merola JF, Avila-Zapata F, Cieślak D, Graham D, Wang C, Menon S, et al.: Tofacitinib or Adalimumab versus Placebo for Psoriatic Arthritis. N Engl J Med 2017, 377, 1537–1550. [CrossRef] [PubMed]
- Zhang A, Zhang G, Yang H, Gong B, Li S, Wei N, Xue H, Wei H, Wang J, Qiu S: Treatment of pro-B acute lymphoblastic leukemia and severe plaque psoriasis with anti-CD19 CAR T cells: a case report. Front Immunol 2025, 16, 1529745. [CrossRef]
- Yan K, Li F, Bi X, Han L, Zhang Z, Chen R, Li Y, Zhang L, Wang X, Li L, et al: Efficacy and safety of vunakizumab in moderate-to-severe chronic plaque psoriasis: A randomized, double-blind, placebo-controlled phase 3 trial. J Am Acad Dermatol 2025, 92, 92–99. [CrossRef]
- Blauvelt A, Papp KA, Griffiths CE, Randazzo B, Wasfi Y, Shen YK, Li S, Kimball AB: Efficacy and safety of guselkumab, an anti-interleukin-23 monoclonal antibody, compared with adalimumab for the continuous treatment of patients with moderate to severe psoriasis: Results from the phase III, double-blinded, placebo- and active comparator-controlled VOYAGE 1 trial. J Am Acad Dermatol 2017, 76, 405–417.
- Gordon KB, Langley RG, Leonardi C, Toth D, Menter MA, Kang S, Heffernan M, Miller B, Hamlin R, Lim L, et al: Clinical response to adalimumab treatment in patients with moderate to severe psoriasis: double-blind, randomized controlled trial and open-label extension study. J Am Acad Dermatol 2006, 55, 598–606. [CrossRef]
- Berstein G, Zhang Y, Berger Z, Kieras E, Li G, Samuel A, Yeoh T, Dowty H, Beaumont K, Wigger-Alberti W, et al.: A phase I, randomized, double-blind study to assess the safety, tolerability and efficacy of the topical RORC2 inverse agonist PF-06763809 in participants with mild-to-moderate plaque psoriasis. Clin Exp Dermatol 2021, 46, 122–129. [CrossRef] [PubMed]
- Chen L, Zhao W: Compound Indigo Naturalis Ointment in the Treatment of 60 Cases of Plaque Psoriasis. Guangming Journal of Chinese Medicine 2014, 29, 1413–1414.
Figure 1.
Schematic diagram of dysregulated immune responses in psoriasis. In genetically predisposed individuals, skin injury or infectious triggers induce neutrophil infiltration, releasing neutrophil extracellular traps (NETs) that activate the TLRs-IFN-α pathway in dendritic cells (DCs), while exosomes and matrix metalloproteinase-9 (MMP9) further enhance the secretion of cytokines by keratinocytes (KCs). KCs release antimicrobial peptides (AMPs), cytokines, and alarmins, recruiting other immune cells to the inflammatory site. Additionally, DCs and Langerhans cells (LCs) drive T-cell differentiation via IL-23. CD4+ Th17 cells, CD8+ Tc17 cells, and γδ T cells collectively produce cytokines such as IL-17, further stimulating KCs proliferation and pro-inflammatory cytokine release, establishing a self-amplifying immune feedback loop. Moreover, tumor necrosis factor (TNF)-α and IL-1β secreted by macrophages and KCs synergistically amplify inflammation. These dysregulated immune responses lead to profound immune cell infiltration, excessive production of cytokines and chemokines, and KCs pathological hyperproliferation, resulting in characteristic psoriatic lesions featuring hyperkeratosis and parakeratosis.
Figure 1.
Schematic diagram of dysregulated immune responses in psoriasis. In genetically predisposed individuals, skin injury or infectious triggers induce neutrophil infiltration, releasing neutrophil extracellular traps (NETs) that activate the TLRs-IFN-α pathway in dendritic cells (DCs), while exosomes and matrix metalloproteinase-9 (MMP9) further enhance the secretion of cytokines by keratinocytes (KCs). KCs release antimicrobial peptides (AMPs), cytokines, and alarmins, recruiting other immune cells to the inflammatory site. Additionally, DCs and Langerhans cells (LCs) drive T-cell differentiation via IL-23. CD4+ Th17 cells, CD8+ Tc17 cells, and γδ T cells collectively produce cytokines such as IL-17, further stimulating KCs proliferation and pro-inflammatory cytokine release, establishing a self-amplifying immune feedback loop. Moreover, tumor necrosis factor (TNF)-α and IL-1β secreted by macrophages and KCs synergistically amplify inflammation. These dysregulated immune responses lead to profound immune cell infiltration, excessive production of cytokines and chemokines, and KCs pathological hyperproliferation, resulting in characteristic psoriatic lesions featuring hyperkeratosis and parakeratosis.
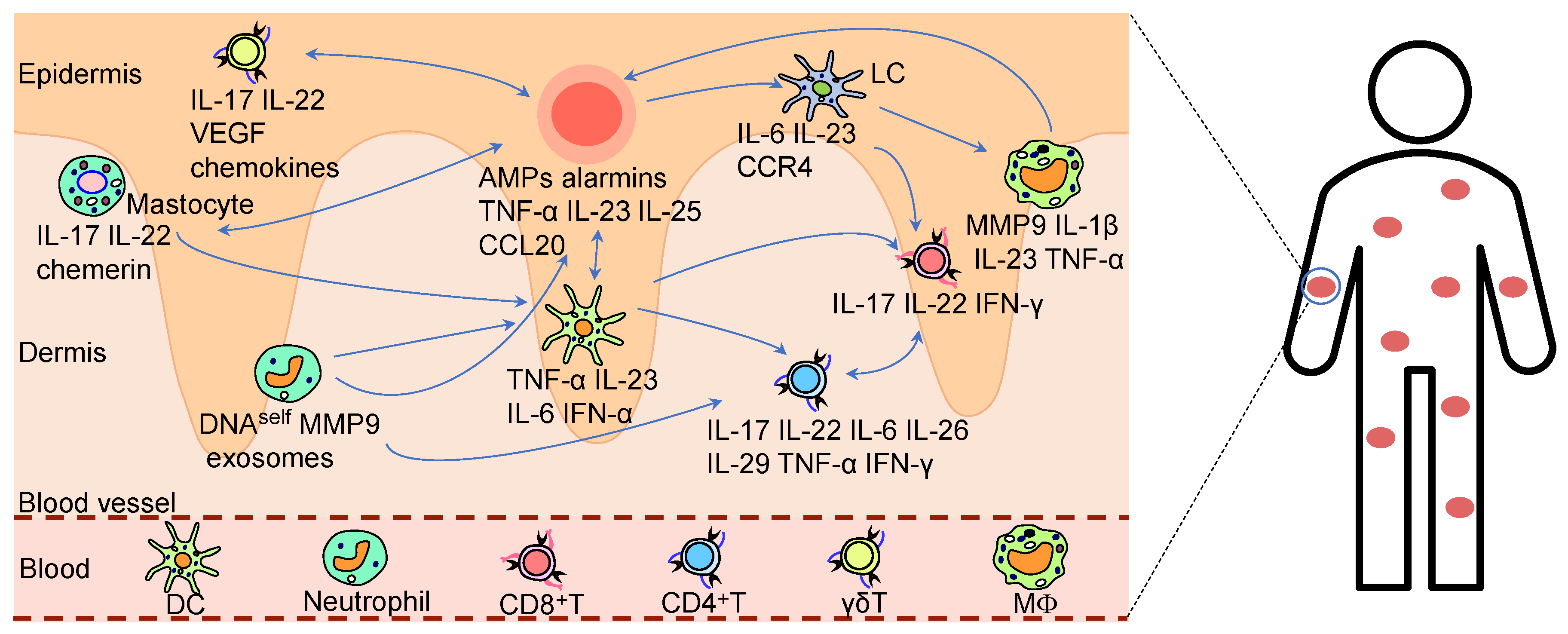
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



