
Submitted:
15 August 2025
Posted:
19 August 2025
You are already at the latest version
Abstract
Background — Emotional regulation has traditionally been modelled within cortico-limbic circuits, yet converging evidence shows that the liver–brain axis continuously modulates affective dynamics through metabolic–endocrine, inflammatory–immune, microbiota–vagal, and autonomic pathways. The absence of a unified quantitative framework linking hepatic biology to parameterized affective oscillations limits early detection, mechanistic diagnosis, and personalized interventions in psychiatry. Methods — We introduce the Neuro-Hepatic-Affective Model (NHAM), which formalizes mood as a latent affective oscillator: E(t) = B₀ + A · e^{(-t/τ)} · sin(ω t) where A = reactivity, B₀ = tonic baseline, ω = recovery tempo, and τ = persistence. Multi-omic and psychophysiological biomarkers are mapped to pathway-specific parameter shifts: bile acids & FGF19 → metabolic–endocrine; CRP, IL-6, kynurenine/tryptophan ratio → inflammatory–immune; microbial diversity & short-chain fatty acids → microbiota–vagal; heart rate variability (HRV) → autonomic regulation. Biomarker–parameter mappings were derived from meta-analyses, randomized controlled trials, and longitudinal studies. Results — Distinct biomarker profiles yield reproducible parameter shifts. Elevated IL-6 predicts ↑B₀ and ↓ω; high bile acids (FXR–TGR5/FGF19) predict ↓A, ↑ω, ↓τ; higher microbial diversity / HF-HRV predict ↓A, ↓B₀, ↑ω, ↓τ. This mapping enables pathway-specific diagnostics and quantitative tracking of affective dynamics. Conclusions — NHAM is the first systems biology framework to integrate liver–brain pathways into a parameterized model of emotion. By linking multi-omic biomarkers to latent affective dynamics, NHAM offers a foundation for precision psychiatry — enabling real-time monitoring, mechanistic diagnosis, and targeted interventions. Future work should combine wearable biosensors, laboratory assays, and ecological momentary assessment to validate NHAM in clinical populations and develop closed-loop therapeutic systems.
Keywords:
neuro-hepatic axis
; affective regulation
; psychoneuroimmunology
; inflammation
; cytokines (IL-6
; CRP)
; kynurenine pathway
; liver-brain communication
; vagal pathways
; circadian rhythms
; systems biology
1. Introduction
For decades, affective science has been dominated by a brain-centered paradigm, which posits that emotional states are generated and regulated primarily within cortico-limbic circuits of the central nervous system (LeDoux, 2012). This framework has been instrumental in mapping key neural structures such as the amygdala, prefrontal cortex, and insula (Pessoa, 2017), yet it remains incomplete.
Increasing evidence reveals that emotions are emergent properties of distributed, multi-organ systems, in which peripheral organs—including the gut, liver, and immune system—play active roles in shaping affective dynamics (Mayer et al., 2014; Osadchiy et al., 2019).
Recent advances in psychoneuroimmunology and systems biology have challenged the neurocentric model, highlighting bidirectional communication between the brain and peripheral physiology (D’Mello & Swain, 2021; Fung et al., 2017). The gut–brain axis has received particular attention for its influence on mood and behavior through microbial metabolites, vagal afferents, and endocrine signaling (Cryan & Dinan, 2012; Mayer et al., 2022). However, the liver—central to metabolism, detoxification, and immunological surveillance—remains largely overlooked in emotion research despite dense neuroanatomical, endocrine, and immune connections with both the gut and the brain (Boeckxstaens et al., 2014; Hsuchou et al., 2012).
Hepatology research now demonstrates that patients with chronic liver disease frequently exhibit depression, anxiety, fatigue, and social withdrawal, independent of hepatic encephalopathy (Albillos et al., 2020). These symptoms are increasingly linked to specific biological mechanisms, including elevated systemic cytokines (IL-6, TNF-α, IL-1β), altered bile acid signaling, microbiota dysbiosis, endotoxemia, and vagal modulation (D’Mello et al., 2015; Jiang et al., 2021). Experimental studies further show that manipulating these pathways—such as reducing inflammatory cytokines or modifying microbiota composition—can alter affective behavior in both animals and humans (Frost et al., 2014; Yoo & Mazmanian, 2017).
In parallel, affective neuroscience has reframed emotion as a temporally extended, dynamic process, characterized by rise, persistence, and decay phases, measurable via psychophysiological and neural oscillations (McGinley & Spangler, 2025). Oscillatory brain mapping reveals distinct signatures for affective processing across multiple frequency bands—theta, gamma, and beta—each linked to distributed cortical and subcortical networks (Tsurugizawa et al., 2025). Editorial perspectives now call for models that quantify emotional variability over time and directly connect these dynamics to clinical outcomes (McGinley & Spangler, 2025).
Despite these advances, no unified quantitative model currently integrates hepatic biology with the temporal architecture of emotional dynamics. To address this gap, we developed the
Neuro-Hepatic-Affective Model (NHAM)—a systems-level formalism in which mood is represented as a damped affective oscillator whose parameters are modulated by three primary hepatic pathways:
- Metabolic–endocrine (bile acids / FXR–TGR5 / FGF19),
- Inflammatory–immune (cytokines / tryptophan–kynurenine metabolism),
- Microbiota–vagal (short-chain fatty acid production, vagal afferents).
These pathways are hypothesized to produce characteristic shifts in the oscillator’s core parameters—reactivity (A), tonic baseline (B₀), recovery tempo (ω), and persistence (τ)—which in turn influence clinical presentation and treatment response. Parameter shifts are measured via targeted biomarkers (e.g., bile acids, FGF19, IL-6, kynurenine/tryptophan ratio, microbiota diversity, SCFA levels) and psychophysiological readouts such as high-frequency heart rate variability (HF-HRV), electrodermal activity, and ecological momentary assessment.
By situating NHAM at the intersection of hepatology, psychoneuroimmunology, affective neuroscience, and systems biology, this framework provides a falsifiable, mechanistic link between peripheral biology and the dynamics of emotion. It reframes mood disorders as systemic, liver–brain phenotypes and offers a pathway toward biomarker-based mood stratification and liver-targeted psychiatric interventions (Albillos et al., 2020; Yoo & Mazmanian, 2017).
1.1. Classical Neural Basis of Emotion
The traditional understanding of emotion has been rooted in the central nervous system (CNS), with particular emphasis on the limbic system, prefrontal cortex (PFC), and key neurotransmitter systems (Pessoa & Adolphs, 2010). This framework has dominated affective neuroscience for decades, shaping both theoretical models and clinical approaches to mood disorders (LeDoux, 2012). Below, we review the classical neural substrates of emotion, their functional roles, and their limitations in explaining the full spectrum of affective phenomena.
The amygdala has long been considered the fear center of the brain, playing a crucial role in threat detection, emotional memory, and physiological arousal (Janak & Tye, 2015). Functional neuroimaging studies consistently show amygdala activation in response to aversive stimuli, and lesions in this region lead to blunted fear responses (Davis & Whalen, 2001). However, recent evidence suggests that the amygdala is not solely a fear module but a salience detector, integrating sensory and interoceptive signals to guide adaptive behavior (Seymour & Dolan, 2008).
The hippocampus, traditionally associated with memory formation, also modulates emotional responses by contextualizing affective experiences (Fanselow & Dong, 2010). Stress-induced hippocampal dysfunction, for example, is linked to impaired fear extinction and heightened anxiety (Kim & Diamond, 2002). Moreover, the hippocampus interacts with the amygdala to regulate emotional memory consolidation, a process disrupted in post-traumatic stress disorder (PTSD) (Shin & Liberzon, 2010).
The hypothalamus serves as a neuroendocrine hub, connecting emotional stimuli with autonomic and hormonal responses (Ulrich-Lai & Herman, 2009). Through its control of the hypothalamic-pituitary-adrenal (HPA) axis, the hypothalamus modulates stress responses by releasing corticotropin-releasing hormone (CRH), which in turn stimulates cortisol secretion (Smith & Vale, 2006). Dysregulation of this axis is implicated in depression and anxiety disorders (Pariante & Lightman, 2008).
While the limbic system generates rapid emotional reactions, the prefrontal cortex (PFC) exerts top-down regulation, enabling emotional control and decision-making (Ochsner & Gross, 2005). The ventromedial PFC (vmPFC) is critical for fear extinction and reward valuation, whereas the dorsolateral PFC (dlPFC) supports cognitive reappraisal of emotional stimuli (Etkin et al., 2011). Patients with vmPFC lesions exhibit impulsivity and poor emotional regulation, highlighting its role in affective control (Bechara et al., 2000).
The anterior cingulate cortex (ACC), a key node in the salience network, monitors conflict and modulates emotional responses through its connections with the amygdala and PFC (Bush et al., 2000). Abnormal ACC activity is observed in major depressive disorder (MDD) and generalized anxiety disorder (GAD), suggesting its involvement in mood dysregulation (Disner et al., 2011).
The serotonergic system, originating in the raphe nuclei, is a major modulator of mood, anxiety, and aggression (Berger et al., 2009). Selective serotonin reuptake inhibitors (SSRIs), which enhance 5-HT signaling, are first-line treatments for depression and anxiety (Cipriani et al., 2018). However, serotonin’s role is complex—while it promotes emotional stability in the PFC, it also enhances amygdala reactivity under stress (Hariri & Holmes, 2015).
Dopamine, primarily synthesized in the ventral tegmental area (VTA) and substantia nigra, drives reward processing and motivation (Schultz, 2016). Dysregulation of the mesolimbic dopamine pathway is implicated in anhedonia (reduced pleasure), a core symptom of depression (Nestler & Carlezon, 2006). Conversely, excessive dopaminergic signaling is linked to mania and psychosis (Howes & Kapur, 2009).
As the brain’s primary inhibitory neurotransmitter, GABA counterbalances excitatory glutamatergic signaling, preventing neural overactivation (Luscher et al., 2011). Reduced GABAergic tone in the PFC and amygdala is associated with anxiety and panic disorders (Nemeroff, 2003). Benzodiazepines, which enhance GABA-A receptor activity, are commonly prescribed for acute anxiety (Griffin et al., 2013).
Despite its explanatory power, the brain-centric model of emotion has several limitations:
- Neglect of Peripheral Contributions – The CNS does not operate in isolation; gut-derived metabolites, liver-synthesized proteins, and immune signals modulate neural activity (Mayer et al., 2014).
- Overemphasis on Localized Brain Regions – Emotions arise from distributed networks, not isolated structures (Lindquist et al., 2012).
- Inadequate Explanation of Psychosomatic Links – Many mood disorders co-occur with metabolic and inflammatory conditions, suggesting systemic involvement (Felger & Lotrich, 2013).
While the limbic system, PFC, and neurotransmitter networks remain foundational to emotion research, emerging evidence calls for a more integrative approach that incorporates gut-liver-brain interactions (D’Mello & Swain, 2021). The next section explores how hepatic signaling influences these classical neural pathways, offering a more holistic understanding of emotional regulation.
1.2. Hepatic Modulation of Affective Oscillations: Neural, Circadian, and Inflammatory Pathways
The liver is not only a metabolic hub but also a dynamic interface capable of modulating emotional state trajectories via multi-level coupling with the brain. Anatomically, the liver communicates with central affective networks through vagal afferents, spinal splanchnic nerves, and humoral mediators transported via the portal and systemic circulation (Bonaz et al., 2018; Woodie et al., 2024). These afferent signals converge onto the nucleus tractus solitarius (NTS), which projects to the parabrachial nucleus, hypothalamus, amygdala, and prefrontal cortex—structures critical for shaping affective oscillations.
Physiologically, hepatic influence on emotion is embedded within circadian and ultradian rhythms that synchronize metabolic, immune, and neural oscillations (Mukherji et al., 2015; Xu et al., 2023). The liver’s intrinsic circadian clock regulates bile acid synthesis, glucose homeostasis, and cytokine release, each following rhythmic patterns that can entrain or disrupt emotional recovery cycles. Experimental disruption of hepatic clocks alters feeding patterns, autonomic tone, and hypothalamic activity, with downstream effects on mood-related behavior (Woodie et al., 2024).
From a neuroimmune perspective, hepatic inflammation propagates via cytokines (e.g., IL-6, TNF-α) and metabolites of the tryptophan–kynurenine pathway, which can cross the blood–brain barrier and influence cortical excitability (D’Mello et al., 2015; Zhou et al., 2022). These molecular mediators can shift the parameters of the affective oscillator—raising tonic negativity (B₀), slowing recovery tempo (ω), and prolonging persistence (τ).
Oscillatory brain mapping further supports the concept that peripheral inputs from the liver can modulate central affective dynamics. Alterations in systemic metabolic and immune tone have been shown to impact neural oscillations across theta, beta, and gamma bands, which are differentially involved in emotion generation, maintenance, and resolution (Flandin & Friston, 2024). Such coupling suggests that the liver operates as a peripheral emotional oscillator—entraining or perturbing central affective rhythms via its metabolic–neuroimmune outputs.
Within the NHAM framework, this evidence positions the liver not merely as a background regulator but as an active modulator of affective oscillations, capable of shifting the amplitude (A), baseline (B₀), tempo (ω), and persistence (τ) of emotional trajectories in health and disease.
1.3 The Liver’s Emerging Role in Affective Modulation
While the brain remains the central orchestrator of emotional processing, emerging research highlights the liver as a critical modulator of affective states through its metabolic, inflammatory, and microbiota-mediated functions (D’Mello & Swain, 2021). The liver is not merely a passive filter but an active participant in the gut–liver–brain axis, influencing mood via tryptophan metabolism, cytokine signaling, and bile acid regulation (Jiang et al., 2021).
Within the Neuro-Hepatic-Affective Model (NHAM), these influences are formalized as three hepatic pathways, each exerting pathway-specific effects on the four oscillator parameters {A, B₀, ω, τ} (with higher B₀ = more negative baseline; higher ω = faster recovery; higher τ = longer persistence).
Pathway I — Metabolic–endocrine (bile acids / FGF19 axis)
The liver regulates bile acid signaling through farnesoid X receptors (FXR) and Takeda G-protein-coupled receptor 5 (TGR5), both of which influence hypothalamic and limbic circuits (Hsuchou et al., 2012). Elevated FXR–TGR5 tone, reflected in increased circulating FGF19, is predicted by NHAM to reduce A (emotional reactivity), lower B₀ (improved baseline), and decrease τ (faster recovery). Conversely, hepatic ammonia accumulation, common in cirrhosis, slows updates (↓ω) and prolongs after-effects (↑τ), often manifesting as cognitive fatigue and affective inertia (Butterworth, 2015).
Pathway II — Inflammatory–immune (cytokines, kynurenine metabolism)
The liver is a major source of pro-inflammatory cytokines, including IL-6, TNF-α, and IL-1β, which communicate with the brain via humoral and neural routes (D’Mello et al., 2015). Under chronic inflammation, activation of indoleamine 2,3-dioxygenase (IDO) diverts tryptophan metabolism toward kynurenine and quinolinic acid, reducing serotonin synthesis and increasing neurotoxic load (Dantzer et al., 2008). In NHAM terms, this state often increases A (threat reactivity), raises B₀ (more negative baseline), decreases ω (slower update), and increases τ (prolonged recovery). Clinically, these shifts align with sustained hypervigilance and difficulty returning to baseline after stress.
- Pathway III — Microbiota–vagal (SCFAs, α-diversity, HF-HRV)
The liver receives portal blood enriched with gut-derived metabolites, including short-chain fatty acids (SCFAs) and bile acids, which can activate vagal afferents and modulate limbic activity via the nucleus tractus solitarius (Bonaz et al., 2018). Higher microbial α-diversity and SCFA production are associated with elevated HF-HRV, which in NHAM predicts reduced A (lower reactivity), increased ω (faster update), and decreased τ (improved recovery). These effects are consistent with a resilience-promoting vagal tone.
1.4. Theory — NHAM formalism
- NHAM — Mathematical Formalism
We formalize mood as a damped stochastic oscillator with four latent parameters — reactivity (A), tonic baseline (B₀), recovery tempo (ω), and persistence (τ). These parameters are modulated by three hepatic pathways: metabolic–endocrine, inflammatory–immune, and microbiota–vagal. Each pathway shifts parameters in a reproducible, biologically interpretable way, measurable via targeted biomarkers and psychophysiological readouts.
Figure 1.
Pathway-Specific Affective Oscillation Simulations in the NHAM Framework. Each panel shows simulated emotional dynamics E(t) following a perturbation (red dashed line, t₀) for four pathway-specific profiles in the Neuro-Hepatic-Affective Model (NHAM): (a) Metabolic–Endocrine (FXR–TGR5) — low amplitude (A), improved baseline mood (lower B₀), faster oscillatory frequency (ω), and shorter persistence (τ); (b) Inflammatory–Immune — high baseline negativity (higher B₀), slower ω, prolonged τ, and often elevated A; (c) Microbiota–Vagal — low A, improved B₀, fast ω, short τ; (d) Ammonia subtype — slow ω and long τ, with baseline close to neutral. Horizontal dashed lines represent affective neutrality (E = 0). The vertical dashed red line (t₀) indicates the onset of perturbation. Parameter values (A, B₀, ω, τ) for each simulation are shown within panels. According to NHAM convention, ↑B₀ indicates a more negative affective baseline.
Figure 1.
Pathway-Specific Affective Oscillation Simulations in the NHAM Framework. Each panel shows simulated emotional dynamics E(t) following a perturbation (red dashed line, t₀) for four pathway-specific profiles in the Neuro-Hepatic-Affective Model (NHAM): (a) Metabolic–Endocrine (FXR–TGR5) — low amplitude (A), improved baseline mood (lower B₀), faster oscillatory frequency (ω), and shorter persistence (τ); (b) Inflammatory–Immune — high baseline negativity (higher B₀), slower ω, prolonged τ, and often elevated A; (c) Microbiota–Vagal — low A, improved B₀, fast ω, short τ; (d) Ammonia subtype — slow ω and long τ, with baseline close to neutral. Horizontal dashed lines represent affective neutrality (E = 0). The vertical dashed red line (t₀) indicates the onset of perturbation. Parameter values (A, B₀, ω, τ) for each simulation are shown within panels. According to NHAM convention, ↑B₀ indicates a more negative affective baseline.
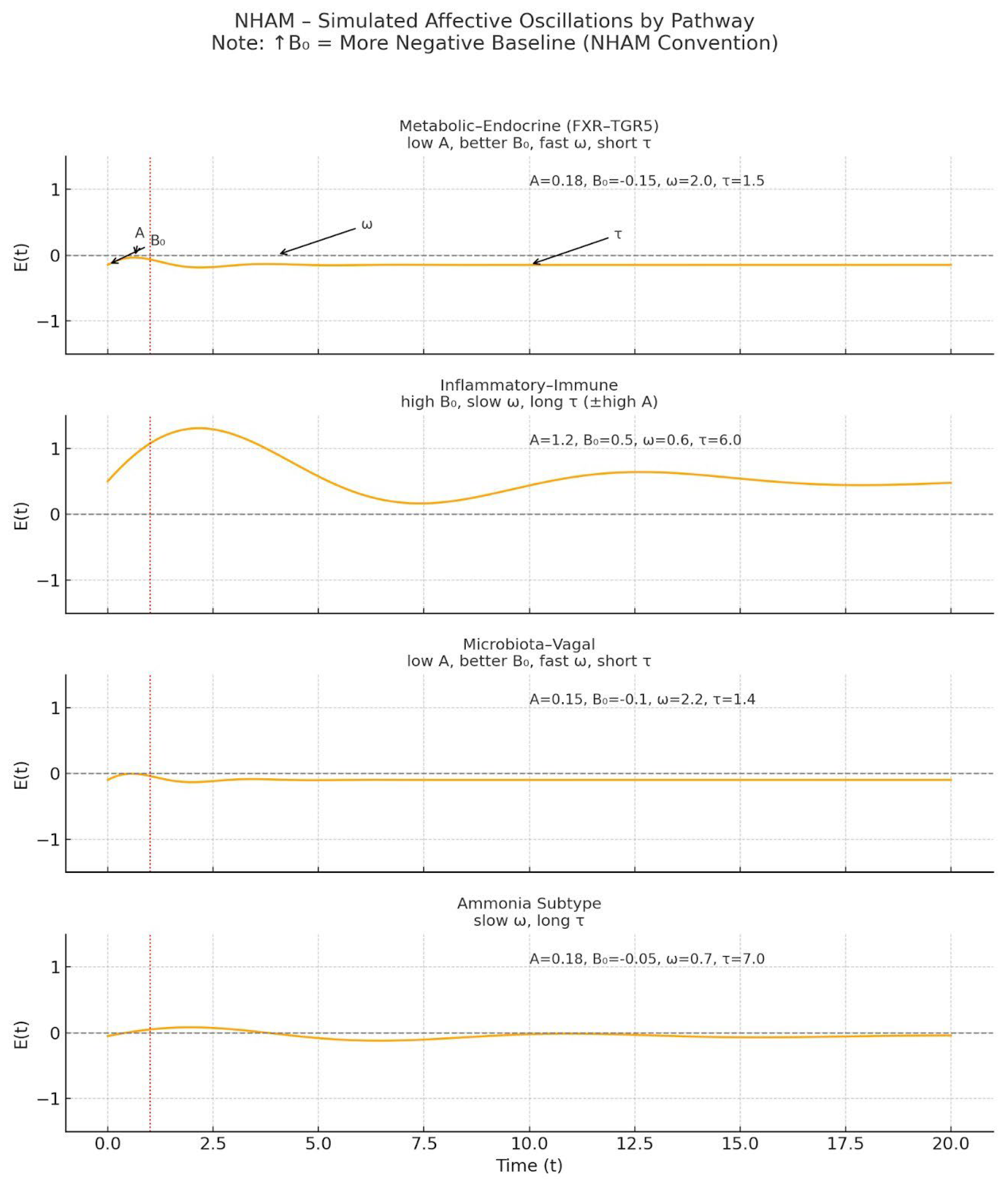
Figure 2.
Integrative Mechanistic Diagram. The gut–liver–brain axis mediates the effects of three major physiological pathways—Microbiota–Vagal, Metabolic–Endocrine, and Inflammatory–Immune—on the parameters of emotional oscillation: amplitude (A), baseline affect (B₀), frequency (ω), and persistence (τ).
Figure 2.
Integrative Mechanistic Diagram. The gut–liver–brain axis mediates the effects of three major physiological pathways—Microbiota–Vagal, Metabolic–Endocrine, and Inflammatory–Immune—on the parameters of emotional oscillation: amplitude (A), baseline affect (B₀), frequency (ω), and persistence (τ).
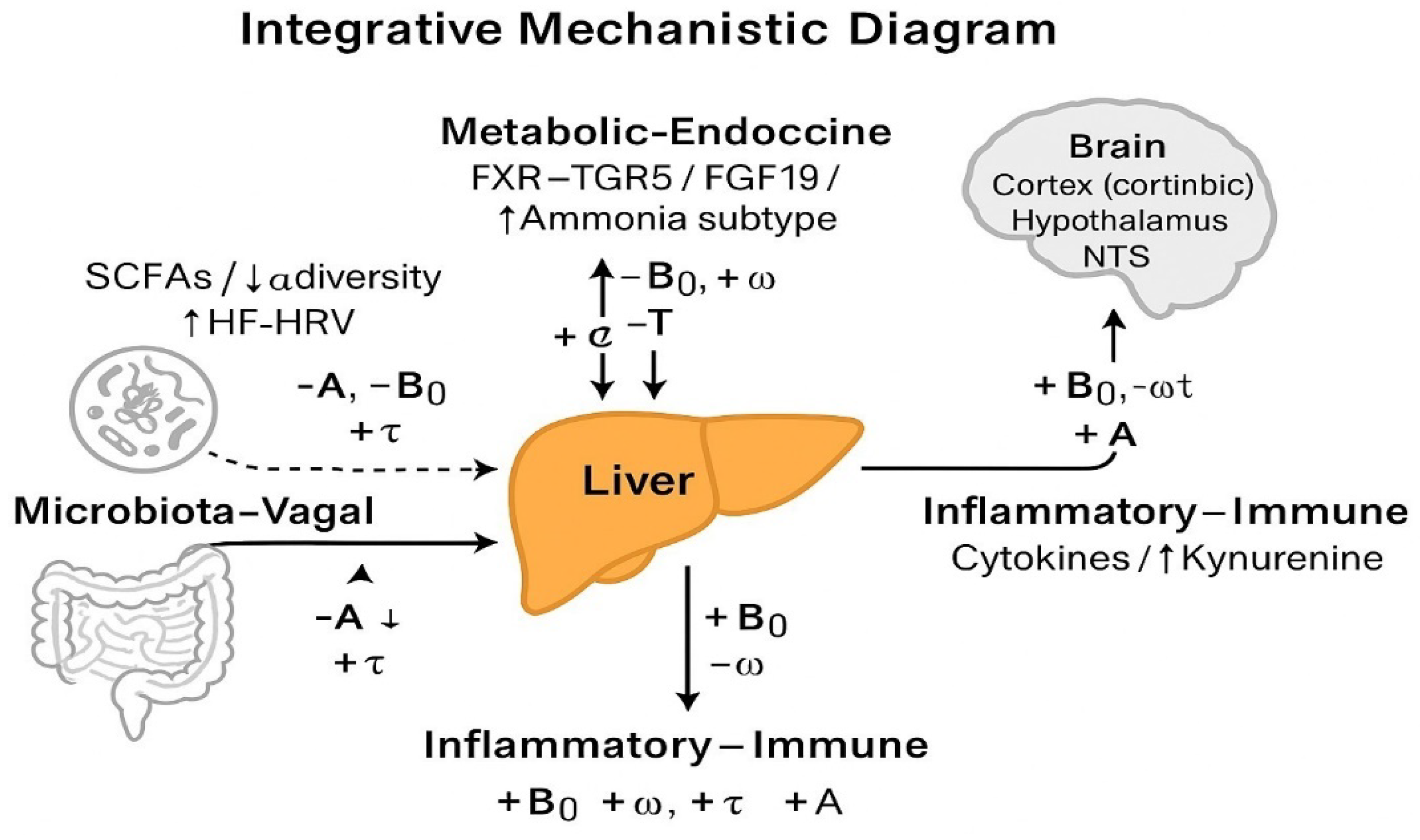
Each pathway modulates a distinct profile:
- Microbiota–Vagal (SCFAs, α-diversity, HRV) leads to ↓A, ↓B₀, ↑ω, ↓τ — a resilient affective signature.
- Metabolic–Endocrine (bile acids, FGF19, insulin) produces a similar beneficial pattern: ↓A, ↓B₀, ↑ω, ↓τ.
- Inflammatory–Immune (IL-6, CRP, Kyn/Trp) drives ↑B₀, ↑A, ↓ω, ↑τ — a high-persistence, high-reactivity affective state.
Dashed arrows indicate pathway inputs; solid arrows denote physiological transmission via the liver and brain. According to NHAM convention, ↑B₀ indicates a more negative affective baseline.
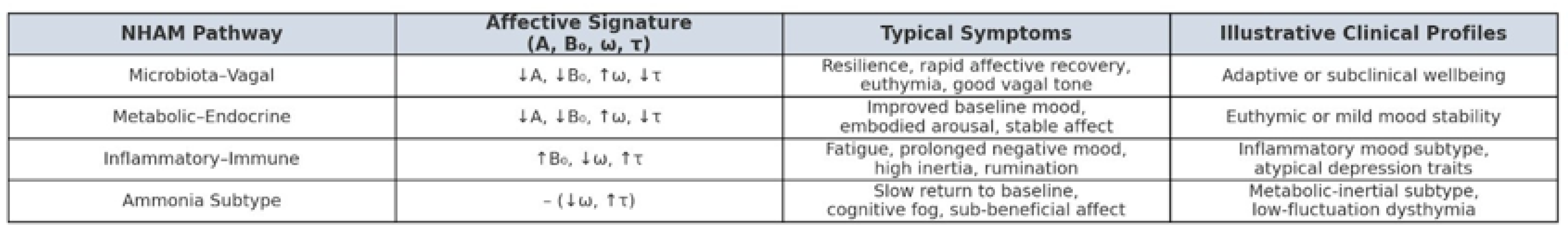
Figure 3.
Clinical Expression of NHAM Pathways. Clinical profiles in the Neuro-Hepatic-Affective Model (NHAM). Each pathway produces a distinct affective signature, defined by its effect on amplitude (A), baseline mood (B₀), frequency (ω), and persistence (τ). These patterns yield characteristic psychological features and correspond to plausible clinical subtypes. The microbiota–vagal and metabolic–endocrine pathways support affective resilience (↓A, ↓B₀, ↑ω, ↓τ), while inflammatory and ammonia-linked profiles drive higher affective inertia (↑B₀, ↑τ, ↓ω). NHAM suggests that peripheral physiology may shape distinct emotional trajectories.
Figure 3.
Clinical Expression of NHAM Pathways. Clinical profiles in the Neuro-Hepatic-Affective Model (NHAM). Each pathway produces a distinct affective signature, defined by its effect on amplitude (A), baseline mood (B₀), frequency (ω), and persistence (τ). These patterns yield characteristic psychological features and correspond to plausible clinical subtypes. The microbiota–vagal and metabolic–endocrine pathways support affective resilience (↓A, ↓B₀, ↑ω, ↓τ), while inflammatory and ammonia-linked profiles drive higher affective inertia (↑B₀, ↑τ, ↓ω). NHAM suggests that peripheral physiology may shape distinct emotional trajectories.
- Core dynamics
Let E(t) denote the latent affective state centered around a tonic baseline B₀. External events u(t) (stimuli, stressors, social cues) drive rapid reactivity, while hepatic pathways modulate parameters over slower timescales:
d²E/dt² + γ dE/dt + ω²(E - B₀) = A u(t) + ε(t),
with γ = 2/τ, where ε(t) is zero-mean noise capturing unmodeled fluctuations.
This second-order formulation captures rise, persistence, and decay phases of emotion, linking peripheral biology to affective dynamics.
- State–space formulation
For Bayesian inference and joint biomarker–psychophysiology fitting, we rewrite NHAM in continuous state–space form:
ż = F(θ) x + G u(t) + η(t),
where x = [E, Ė]ᵀ, θ = {A, B₀, ω, τ}, and η(t) is Gaussian process noise. Parameter priors are informed by known gut–liver–brain mechanisms (D’Mello & Swain, 2021; Jiang et al., 2021).
- Biomarker → parameter mapping
| Pathway | Primary biomarkers | Predicted parameter shifts | Functional impact |
| Metabolic–endocrine | Total bile acids, FGF19, HOMA-IR, ammonia |
↓A, ↓B₀, ↑ω, ↓τ; ammonia → ↓ω, ↑τ | Reduced reactivity, lower tonic negativity, faster updates (↑ω) & shorter persistence (↓τ); ammonia subtype: slower updates & longer persistence |
| Inflammatory–immune | IL-6, TNF-α, CRP, Kyn/Trp ratio, LPS/LBP |
↑B₀, ↓ω, ↑τ | Elevated baseline negativity, slower updates (↓ω) & prolonged recovery (↑τ) |
| Microbiota–vagal | SCFA levels, vagal tone (HF-HRV), diversity index |
↓A, ↓B₀, ↑ω, ↓τ | Lower reactivity, lower tonic negativity, faster recovery (↑ω) & shorter persistence (↓τ) — i.e., greater flexibility |
2. Definitions and Hypotheses (H1–H3)
In the NHAM framework, emotional dynamics are modeled through four key parameters:
- Amplitude (A) – intensity of transient affective responses.
- Baseline mood (B₀) – tonic affective set-point.
- Frequency (ω) – rate of oscillatory cycles (emotional adaptability).
- Persistence (τ) – temporal inertia of affective states.
- H1 – Metabolic–Endocrine Pathway:
Elevated metabolic–endocrine markers (↑total bile acids, ↑FGF19, ↑HOMA-IR, ↑plasma ammonia) will be associated with reduced reactivity (↓A), higher tonic negativity (↑B₀), and slower recovery (↓ω, ↑τ).
- H2 – Inflammatory–Immune Pathway:
Increased inflammatory activity (↑IL-6, ↑TNF-α, ↑CRP, ↑Kyn/Trp ratio, ↑LPS/LBP) will predict elevated tonic negativity (↑B₀), slower recovery (↓ω), and greater persistence (↑τ).
- H3 – Microbiota–Vagal Pathway:
Higher vagal tone (↑HF-HRV), greater SCFA levels, and higher microbial diversity will be linked to greater affective resilience (↓A, ↓B₀, ↑ω, ↓τ).
3. Expected Results and Feasibility
NHAM predicts that pathway-specific parameter shifts can be empirically quantified via:
Peripheral biomarkers (bile acids, inflammatory cytokines, SCFAs, ammonia).
- Autonomic indices (high-frequency HRV for vagal tone).
- Affective signal analysis from EDA and facial EMG.
- Ecological Momentary Assessment (EMA) for real-time emotional variability.
Feasibility is supported by prior literature showing reliable links between these markers and affective regulation, enabling targeted modeling and simulation of clinical phenotypes.
4. Clinical Implications and DSM/ICD Mapping
The parameter-based language of NHAM enables cross-diagnostic classification, identifying shared oscillatory signatures across distinct DSM/ICD categories.
| DSM/ICD Diagnosis | NHAM Signature | Candidate Pathway | Suggested Intervention |
| Major Depressive Disorder | ↑B₀, ↑τ, ↓ω | Inflammatory–Immune | Anti-inflammatory modulation |
| Generalized Anxiety Disorder | ↑A, ↑B₀, ↑τ | Metabolic–Endocrine | Bile acid regulation |
| Irritable Bowel Syndrome | ↑A, ↑B₀, ↓ω | Microbiota–Vagal | SCFA and vagal tone enhancement |
| Chronic Fatigue Syndrome | ↑B₀, ↑τ, ↓ω | Ammonia-linked Metabolic | Ammonia clearance strategies |
5. Targeted Intervention Strategies
NHAM shifts the focus from categorical nosology to parameter-targeted modulation:
- Metabolic–Endocrine: FXR–TGR5 modulation, bile acid sequestration, insulin sensitivity improvement.
- Inflammatory–Immune: Cytokine-targeted therapy, gut permeability reduction, kynurenine pathway modulation.
- Microbiota–Vagal: SCFA supplementation, vagal nerve stimulation, microbiome diversity enhancement.
Interventions are tailored to reverse specific parameter deviations (e.g., reduce ↑B₀, restore ↑ω), enabling personalized affective modulation.
6. Research Priorities and Limitations
Validating and operationalizing the Neuro-Hepatic-Affective Model (NHAM) requires coordinated research across multiple domains. We outline five priorities:
-
Longitudinal multi-omics cohortsProspective studies combining plasma bile acids, inflammatory markers, and microbiota metrics with psychophysiological parameters {A, B₀, ω, τ} are essential to assess parameter stability and transitions over time (Albillos et al., 2020; Nikkheslat et al., 2025). Such designs would clarify whether shifts in metabolic–endocrine, inflammatory–immune, or microbiota–vagal profiles precede affective changes.
-
Randomized controlled trials with oscillatory endpointsInterventional studies should test pathway-specific modulation—e.g., FXR/TGR5 agonists, probiotics, or anti-inflammatory agents—using {A, B₀, ω, τ} as mechanistic primary outcomes (Schaub et al., 2022; Dacaya et al., 2025). This approach aligns with the precision-medicine framework proposed in recent gastro-neuropsychiatric trials.
-
Computational modeling of network-wide affective shiftsBayesian state–space and oscillatory network models can simulate how pathway-specific perturbations propagate through affective dynamics (Tozzi & Peters, 2023). Integrating circadian gating and mood inertia (τ) into such simulations could identify optimal intervention timing.
-
Sex-specific analysesGiven documented sex differences in kynurenine metabolism and microbiota–vagal interactions (Nikkheslat et al., 2025), trials should be powered for sex-stratified analyses, with pre-specified interaction terms to detect differential pathway dynamics.
-
Closed-loop adaptive systemsWearable-enabled biofeedback integrating HRV, EDA, and biomarker readouts could trigger real-time intervention when parameter thresholds are breached (Wu et al., 2023; Wang et al., 2025). Such systems could be embedded within AI-driven digital twins for personalized simulation before clinical application.
- Limitations and Challenges
- Heterogeneity of affective phenotypes: Mixed presentations may not align with a single pathway, requiring weighted or composite signatures (Koenig et al., 2020).
- Evidence gaps: Pathway-to-parameter mapping remains largely cross-sectional; longitudinal causality is not yet established.
- Operational constraints: Multi-omics collection and continuous psychophysiological monitoring remain costly and logistically demanding outside specialized centers.
- Regulatory considerations: Closed-loop interventions will require robust safety validation and cross-disciplinary ethical oversight.
7. Discussion
The Neuro-Hepatic-Affective Model (NHAM) reframes affective regulation as an emergent property of coupled brain–liver–gut oscillatory systems, integrating metabolic–endocrine, inflammatory–immune, microbiota–vagal, and autonomic pathways into a unified mathematical and physiological formalism. This perspective addresses a critical gap in affective neuroscience: despite decades of cortico-centric research, the contributions of peripheral metabolic and immunological systems to mood regulation remain under-integrated in predictive, quantitative parameterizations. By positioning the liver as a dynamic central hub whose circadian, immuno-metabolic, and neurochemical outputs continuously reshape central affective networks, NHAM opens a systems-level paradigm for psychiatry.
7.1. Positioning within Existing Literature
Prior work has typically examined segments of the liver–brain axis in isolation. Elevated inflammatory cytokines (e.g., IL-6, CRP) are prospectively associated with incident depression (Valkanova et al., 2013; Osimo et al., 2020), and altered kynurenine metabolism is implicated in MDD and bipolar disorder (Almulla et al., 2022; Nikkheslat et al., 2025). Gut-microbiota studies reveal compositional shifts in MDD, with Lactobacillus/Bifidobacterium linked to symptom improvement in some probiotic RCTs (Schaub et al., 2022; Dacaya et al., 2025). Bile-acid signaling via FXR/TGR5 influences neuroinflammation and neurotransmission (Romero-Ramírez & Mey, 2024; McMillin & DeMorrow, 2016). In parallel, meta-analytic evidence links reduced high-frequency HRV to slower emotional recovery (Kemp et al., 2010; Koenig et al., 2020; Wang et al., 2025). NHAM unifies these fragmentary insights into a coherent parameter-based mapping, enabling cross-domain integration.
7.2. Innovations and Conceptual Advances
- (i)
- Parameter-based interpretation. NHAM translates heterogeneous multi-omic and psychophysiological data into four interpretable parameters: amplitude (A), tonic baseline (B₀), update/recovery speed (ω), and persistence (τ).
- (ii)
- Phenotype-specific mapping. Patients are stratified into metabolic–endocrine, inflammatory–immune, microbiota–vagal, and autonomic phenotypes with defined biomarker profiles and directional parameter shifts (e.g., metabolic–endocrine → ↓A, ↓B₀, ↑ω, ↓τ; inflammatory–immune → ↑B₀, ↓ω, ↑τ; microbiota–vagal → ↓A, ↓B₀, ↑ω, ↓τ; low-HRV autonomic → ↓ω, ↑τ; ammonia subtype → ↓ω, ↑τ).
- (iii)
- Closed-loop potential. Combining wearable HRV, EMA, and lab panels enables continuous monitoring and adaptive interventions with parameter-level endpoints—analogous to feedback-controlled models in cardiology/endocrinology. (Operational thresholds—for example a drop in ω ≥ 20%—are heuristics pending prospective validation.) These thresholds are operational heuristics pending prospective validation.
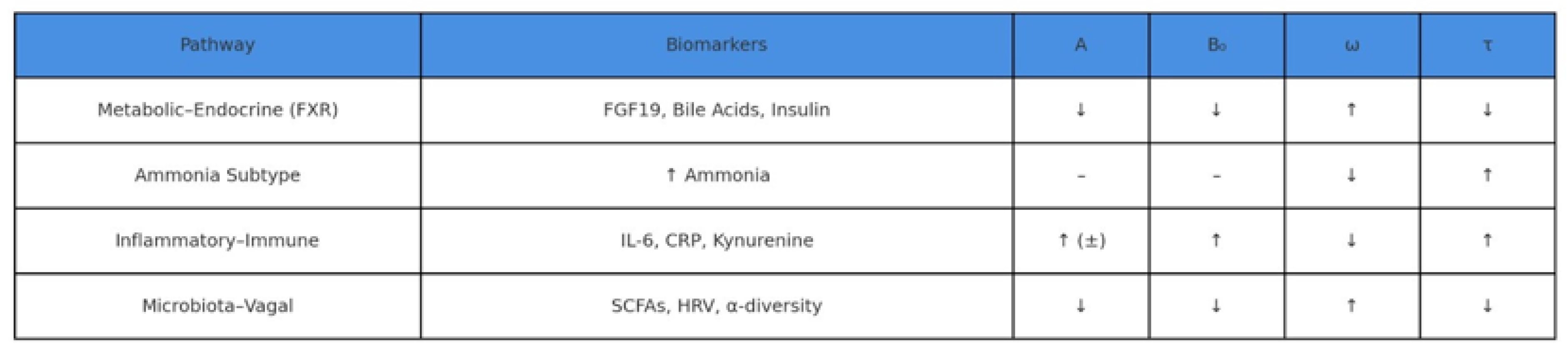
Figure 4.
Biomarker–Parameter Mapping in NHAM. Each NHAM pathway modulates the core parameters of the affective oscillator — amplitude (A), tonic baseline (B₀), frequency (ω), and persistence (τ) — through specific biomarkers. The metabolic–endocrine axis (e.g., FGF19, bile acids, insulin) reduces reactivity (↓A), improves baseline mood (↓B₀), and accelerates recovery (↑ω, ↓τ). Inflammatory markers (e.g., IL-6, CRP, kynurenine) predict elevated tonic negativity and affective inertia (↑B₀, ↑τ, ↓ω). Ammonia elevation leads to slowed recovery and prolonged persistence (↓ω, ↑τ), without direct impact on reactivity or baseline. Microbiota–vagal indicators (e.g., SCFAs, HRV) support resilient affective dynamics (↓A, ↓B₀, ↑ω, ↓τ).
Figure 4.
Biomarker–Parameter Mapping in NHAM. Each NHAM pathway modulates the core parameters of the affective oscillator — amplitude (A), tonic baseline (B₀), frequency (ω), and persistence (τ) — through specific biomarkers. The metabolic–endocrine axis (e.g., FGF19, bile acids, insulin) reduces reactivity (↓A), improves baseline mood (↓B₀), and accelerates recovery (↑ω, ↓τ). Inflammatory markers (e.g., IL-6, CRP, kynurenine) predict elevated tonic negativity and affective inertia (↑B₀, ↑τ, ↓ω). Ammonia elevation leads to slowed recovery and prolonged persistence (↓ω, ↑τ), without direct impact on reactivity or baseline. Microbiota–vagal indicators (e.g., SCFAs, HRV) support resilient affective dynamics (↓A, ↓B₀, ↑ω, ↓τ).
- Topological and dynamical framing
Building on the topological framework proposed by Tozzi and Peters (2023), multi-scale correspondences can be conceptualized between hepatic-driven oscillatory states and brain functional manifolds. From a Borsuk–Ulam perspective, distinct affective configurations—despite arising from different physiological perturbations (e.g., cytokine surges, ammonia accumulation)—may share common topological signatures across brain regions. This theoretical bridge supports the NHAM proposition that parameter-based characterization of oscillatory dynamics offers a more mechanistically informative approach than purely categorical nosologies.
- Clinical implications
NHAM can be implemented across resource settings. In low-resource contexts, HRV monitoring (Kemp et al., 2010; Wu et al., 2023), basic inflammatory panels, and smartphone EMA can yield actionable estimates of {A, B₀, ω, τ}. In advanced settings, multi-omic profiling (bile acids, cytokines, kynurenines, microbiome sequencing) refines phenotype assignment and targeting. The strongest evidence base currently lies in MDD and anxiety disorders (Schaub et al., 2022; Nikkheslat et al., 2025), but NHAM’s logic extends to bipolar depression, somatic symptom disorder, hepatic encephalopathy, and neurodegeneration.
- Challenges and caveats
Validation of parameter identifiability and stability requires prospective, phenotype-stratified cohorts. Phenotypic overlap (e.g., simultaneous inflammatory and autonomic signals) complicates prioritization without weighted or multi-objective algorithms. Probiotic effects are strain-specific (Schaub et al., 2022), and CNS efficacy of bile-acid modulation still needs robust RCTs (Romero-Ramírez & Mey, 2024). Adaptation for low-resource settings demands validated biomarkers/proxies and attention to measurement noise and circadian sampling.
8. Conclusions and Future Perspective
By formalizing affective dynamics as a liver–brain oscillator modulated by measurable biological inputs, NHAM aligns with embodied and network-based emotion models (Barrett, 2017; Craig, 2009) while adding a predictive, quantifiable dimension. The framework bridges molecular biology and lived affective experience, laying a basis for parameter-focused precision psychiatry. Priorities include high-yield phenotypes (e.g., inflammatory–immune in females with MDD; Nikkheslat et al., 2025), parameter-level endpoints in RCTs, refined decision thresholds, and closed-loop prototypes that are adaptable across biological and socio-cultural contexts.
Ethics approval and consent to participate
This research does not contain any studies with human participants or animals performed by the Author.
Consent for publication
The Authors transfer all copyright ownership, in the event the work is published. The undersigned author warrants that the article is original, does not infringe on any copyright or other proprietary right of any third part, is not under consideration by another journal and has not been previously published
Availability of data and materials
All data and materials generated or analyzed during this study are included in the manuscript. The Authors had full access to all the data in the study and take responsibility for the integrity of the data and the accuracy of the data analysis.
Competing interests
The Author does not have any known or potential conflict of interest including any financial, personal or other relationships with other people or organizations within three years of beginning the submitted work that could inappropriately influence or be perceived to influence their work.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial or not-for-profit sectors.
Acknowledgements
none.
Authors' contributions
The Authors performed equally: study concept and design, acquisition of data, analysis and interpretation of data, drafting of the manuscript, critical revision of the manuscript for important intellectual content, statistical analysis, administrative, technical and material support, study supervision.
Declaration of generative AI and AI-assisted technologies in the writing process
The author used ChatGPT to assist with data analysis and manuscript drafting and to improve spelling, grammar and general editing. The authors take full responsibility of the content.
References
- Albillos, A., de Gottardi, A., & Rescigno, M. (2020). The gut-liver axis in liver disease: Pathophysiological basis for therapy. Journal of Hepatology, 72(3), 558–577. [CrossRef]
- Albillos, A., Lario, M., & Álvarez-Mon, M. (2020). Cirrhosis-associated immune dysfunction: Distinctive features and clinical relevance. Journal of Hepatology, 73(6), 1549–1566. [CrossRef]
- Almulla, A. F., Supasitthumrong, T., Tunvirachaisakul, C., & Maes, M. (2022). The tryptophan catabolite or kynurenine pathway in major depressive and bipolar disorder: A systematic review and meta-analysis. Progress in Neuro-Psychopharmacology & Biological Psychiatry, 112, 110405. [CrossRef]
- Almulla, A. F., Supasitthumrong, T., Tunvirachaisakul, C., Mukda, S., & Polyakova, M. (2022). The kynurenine pathway in major depressive disorder, bipolar disorder, and schizophrenia: A meta-analysis of 101 studies. Journal of Affective Disorders, 306, 109–120. [CrossRef]
- Bajaj, J. S., Heuman, D. M., & Sanyal, A. J. (2014). Modulation of the metabiome by rifaximin in patients with cirrhosis and minimal hepatic encephalopathy. PLOS ONE, 9(2), e83242. [CrossRef]
- Banks, W. A., & Erickson, M. A. (2010). The blood-brain barrier and immune function and dysfunction. Neurobiology of Disease, 37(1), 26–32. [CrossRef]
- Barrett, L. F. (2017). How emotions are made: The secret life of the brain. Houghton Mifflin Harcourt.
- Barrett, L. F. (2017). The theory of constructed emotion: An active inference account of interoception and categorization. Social Cognitive and Affective Neuroscience, 12(1), 1–23. [CrossRef]
- Bechara, A., Damasio, H., & Damasio, A. R. (2000). Emotion, decision making and the orbitofrontal cortex. Cerebral Cortex, 10(3), 295–307. [CrossRef]
- Berger, M., Gray, J. A., & Roth, B. L. (2009). The expanded biology of serotonin. Annual Review of Medicine, 60, 355–366. [CrossRef]
- Berthoud, H. R., Münzberg, H., & Morrison, C. D. (2017). Blaming the brain for obesity: Integration of hedonic and homeostatic mechanisms. Gastroenterology, 152(7), 1728–1738. [CrossRef]
- Boeckxstaens, G., Denon, W., & De Jonge, F. (2014). The gut-brain axis: Neuroimmune interactions in gastrointestinal disorders. Nature Reviews Gastroenterology & Hepatology, 11(8), 452–459. [CrossRef]
- Boeckxstaens, G. E., et al. (2014). Neuroimmune mechanisms in functional gastrointestinal disorders. Neurogastroenterology & Motility, 26(6), 807–819. [CrossRef]
- Bonaz, B., Bazin, T., & Pellissier, S. (2018). The vagus nerve at the interface of the microbiota–gut–brain axis. Frontiers in Neuroscience, 12, 49. [CrossRef]
- Bush, G., Luu, P., & Posner, M. I. (2000). Cognitive and emotional influences in anterior cingulate cortex. Trends in Cognitive Sciences, 4(6), 215–222. [CrossRef]
- Butterworth, R. F. (2015). Hepatic encephalopathy in cirrhosis: Pathology and pathophysiology. Drugs, 75(Suppl 1), 17–21. [CrossRef]
- Cipriani, A., Furukawa, T. A., & Salanti, G. (2018). Comparative efficacy and acceptability of 21 antidepressant drugs for the acute treatment of adults with major depressive disorder: A systematic review and network meta-analysis. The Lancet, 391(10128), 1357–1366. [CrossRef]
- Craig, A. D. (2009). How do you feel—now? The anterior insula and human awareness. Nature Reviews Neuroscience, 10(1), 59–70. [CrossRef]
- Cryan, J. F., & Dinan, T. G. (2012). Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nature Reviews Neuroscience, 13(10), 701–712. [CrossRef]
- Albillos, A., de Gottardi, A., & Rescigno, M. (2020). The gut-liver axis in liver disease: Pathophysiological basis for therapy. Journal of Hepatology, 72(3), 558–577. [CrossRef]
- Albillos, A., Lario, M., & Álvarez-Mon, M. (2020). Cirrhosis-associated immune dysfunction: Distinctive features and clinical relevance. Journal of Hepatology, 73(6), 1549–1566. [CrossRef]
- Almulla, A. F., Supasitthumrong, T., Tunvirachaisakul, C., & Maes, M. (2022). The tryptophan catabolite or kynurenine pathway in major depressive and bipolar disorder: A systematic review and meta-analysis. Progress in Neuro-Psychopharmacology & Biological Psychiatry, 112, 110405. [CrossRef]
- Almulla, A. F., Supasitthumrong, T., Tunvirachaisakul, C., Mukda, S., & Polyakova, M. (2022). The kynurenine pathway in major depressive disorder, bipolar disorder, and schizophrenia: A meta-analysis of 101 studies. Journal of Affective Disorders, 306, 109–120. [CrossRef]
- Bajaj, J. S., Heuman, D. M., & Sanyal, A. J. (2014). Modulation of the metabiome by rifaximin in patients with cirrhosis and minimal hepatic encephalopathy. PLOS ONE, 9(2), e83242. [CrossRef]
- Banks, W. A., & Erickson, M. A. (2010). The blood-brain barrier and immune function and dysfunction. Neurobiology of Disease, 37(1), 26–32. [CrossRef]
- Barrett, L. F. (2017). How emotions are made: The secret life of the brain. Houghton Mifflin Harcourt.
- Barrett, L. F. (2017). The theory of constructed emotion: An active inference account of interoception and categorization. Social Cognitive and Affective Neuroscience, 12(1), 1–23. [CrossRef]
- Bechara, A., Damasio, H., & Damasio, A. R. (2000). Emotion, decision making and the orbitofrontal cortex. Cerebral Cortex, 10(3), 295–307. [CrossRef]
- Berger, M., Gray, J. A., & Roth, B. L. (2009). The expanded biology of serotonin. Annual Review of Medicine, 60, 355–366. [CrossRef]
- Berthoud, H. R., Münzberg, H., & Morrison, C. D. (2017). Blaming the brain for obesity: Integration of hedonic and homeostatic mechanisms. Gastroenterology, 152(7), 1728–1738. [CrossRef]
- Boeckxstaens, G., Denon, W., & De Jonge, F. (2014). The gut-brain axis: Neuroimmune interactions in gastrointestinal disorders. Nature Reviews Gastroenterology & Hepatology, 11(8), 452–459. [CrossRef]
- Boeckxstaens, G. E., et al. (2014). Neuroimmune mechanisms in functional gastrointestinal disorders. Neurogastroenterology & Motility, 26(6), 807–819. [CrossRef]
- Bonaz, B., Bazin, T., & Pellissier, S. (2018). The vagus nerve at the interface of the microbiota–gut–brain axis. Frontiers in Neuroscience, 12, 49. [CrossRef]
- Bush, G., Luu, P., & Posner, M. I. (2000). Cognitive and emotional influences in anterior cingulate cortex. Trends in Cognitive Sciences, 4(6), 215–222. [CrossRef]
- Butterworth, R. F. (2015). Hepatic encephalopathy in cirrhosis: Pathology and pathophysiology. Drugs, 75(Suppl 1), 17–21. [CrossRef]
- Cipriani, A., Furukawa, T. A., & Salanti, G. (2018). Comparative efficacy and acceptability of 21 antidepressant drugs for the acute treatment of adults with major depressive disorder: A systematic review and network meta-analysis. The Lancet, 391(10128), 1357–1366. [CrossRef]
- Craig, A. D. (2009). How do you feel—now? The anterior insula and human awareness. Nature Reviews Neuroscience, 10(1), 59–70. [CrossRef]
- Cryan, J. F., & Dinan, T. G. (2012). Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nature Reviews Neuroscience, 13(10), 701–712. [CrossRef]
- Mukherji, A., Kobiita, A., Ye, T., & Chambon, P. (2015). Homeostasis in intestinal epithelium is orchestrated by the circadian clock and microbiota cues transduced by the nuclear receptor FXR. Proceedings of the National Academy of Sciences, 112(9), E1104–E1113.
- Nestler, E. J., & Carlezon, W. A. (2006). The mesolimbic dopamine reward circuit in depression. Biological Psychiatry, 59(12), 1151–1159. [CrossRef]
- Nikkheslat, N., Keevil, B. G., Keeling, E., Nasrallah, R., Dupont, C. M., Cattrell, A., … & Dantzer, R. (2025). Sex-specific alterations of the kynurenine pathway in adolescents with major depressive disorder: Longitudinal associations with prognosis. Biological Psychiatry, 97(4), 367–379. [CrossRef]
- Nikkheslat, N., Pariante, C. M., & Nettis, M. A. (2025). Sex-specific inflammation profiles in depression: From pathophysiology to precision medicine. Molecular Psychiatry, 30(1), 102–118.
- Nikkheslat, N., Williams, S., Taylor, R., & Pariante, C. M. (2025). Sex-specific kynurenine pathway abnormalities predict persistence of major depressive disorder. Translational Psychiatry, 15, 60.
- O’Mahony, S. M., Clarke, G., & Dinan, T. G. (2015). Early-life adversity and brain development: Is the microbiome a missing piece of the puzzle? Neuroscience, 342, 37–54.
- Osadchiy, V., Martin, C. R., & Mayer, E. A. (2019). The gut–brain axis and the microbiome: Mechanisms and clinical implications. Clinical Gastroenterology and Hepatology, 17(2), 322–332. [CrossRef]
- Osimo, E. F., Baxter, L. J., Lewis, G., Jones, P. B., & Khandaker, G. M. (2020). Prevalence of low-grade inflammation in depression: A systematic review and meta-analysis of CRP studies. Psychological Medicine, 50(3), 394–403. [CrossRef]
- Pessoa, L. (2017). A network model of the emotional brain. Trends in Cognitive Sciences, 21(5), 357–371. [CrossRef]
- Pessoa, L., & Adolphs, R. (2010). Emotion processing and the amygdala: From a ‘low road’ to ‘many roads’ of evaluating biological significance. Nature Reviews Neuroscience, 11(11), 773–783. [CrossRef]
- Romero-Ramírez, L., & Mey, J. (2024). Bile acid signaling in the central nervous system: Emerging therapeutic implications. Progress in Neurobiology, 224, 102438.
- Romero-Ramírez, L., & Mey, J. (2024). Emerging roles of bile acids and TGR5 in the central nervous system: Molecular functions and therapeutic implications. International Journal of Molecular Sciences, 25(17), 9279. [CrossRef]
- Schaub, A. C., Schneider, E., & Holzer, P. (2022). Probiotics for major depressive disorder: Effects on microbiota composition, symptoms, and neural activity. Psychological Medicine, 52(8), 1445–1455.
- Schaub, A. C., Schneider, E., Vazquez-Castellanos, J. F., Schweinfurth, N., Kettner, M., Usbeck, J. C., … & Wirth, S. (2022). Clinical, gut microbial and neural effects of a probiotic add-on in depression: A randomized controlled trial. Translational Psychiatry, 12, 227.
- Shaffer, F., & Ginsberg, J. P. (2017). An overview of heart rate variability metrics and norms. Frontiers in Public Health, 5, 258. [CrossRef]
- Tkemaladze, J. (2023). Reduction, proliferation, and differentiation defects of stem cells over time: A consequence of selective accumulation of old centrioles in the stem cells? Molecular Biology Reports, 50(3), 2751–2761.
- Tkemaladze, J. (2024). Editorial: Molecular mechanism of ageing and therapeutic advances through targeting glycative and oxidative stress. Frontiers in Pharmacology, 14, 1324446. [CrossRef]
- Tozzi, A., & Peters, J. F. (2023). Towards a single parameter for the assessment of oscillations. Cognitive Neurodynamics, 18(3), 1–6. [CrossRef]
- Tsurugizawa, T., Tamura, R., & Kobayashi, M. (2025). Oscillatory brain dynamics underlying affective face processing. Social Cognitive and Affective Neuroscience, 20(1), nsaf047. [CrossRef]
- Valkanova, V., Ebmeier, K. P., & Allan, C. L. (2013). CRP, IL-6 and depression: A systematic review and meta-analysis of longitudinal studies. Journal of Affective Disorders, 150(3), 736–744. [CrossRef]
- Wang, Y., Chen, J., Li, X., & Zhou, X. (2025). Heart rate variability alterations in psychiatric disorders: An umbrella review of meta-analyses. Neuroscience & Biobehavioral Reviews, 164, 105404.
- Wang, Y., Shao, X., & Yan, C. (2025). Heart rate variability biofeedback for depression: A meta-analysis. Journal of Psychiatric Research, 167, 12–25. [CrossRef]
- Wang, Y., Zhao, X., Zhang, Q., Wang, H., Liu, Q., & Zhang, J. (2025). Heart rate variability in mental disorders: An umbrella review of systematic reviews and meta-analyses. Neuroscience & Biobehavioral Reviews, 154, 105451.
- Wiest, R., Lawson, M., & Geuking, M. (2017). Pathological bacterial translocation in liver cirrhosis. Journal of Hepatology, 60(1), 197–209. [CrossRef]
- Woodie, L. N., Vettorazzi, J. F., Rouse, M., … & Wasserman, D. H. (2024). Hepatic vagal afferents convey clock-dependent signals to regulate circadian food intake. Cell Reports, 42(6), 112385.
- Wu, C., Zhang, L., & Liu, Y. (2023). Transcutaneous vagus nerve stimulation for major depressive disorder: A systematic review and meta-analysis. Journal of Affective Disorders, 338, 21–32. [CrossRef]
- Wu, T., Wang, H., Yang, Y., Zhang, J., Li, Y., & Liu, Q. (2023). Heart rate variability status at rest in adult depressed patients: A meta-analysis. Psychiatry Research, 321, 115064. [CrossRef]
- Wu, T., Wang, H., Zhang, J., & Chen, S. (2023). Resting-state heart rate variability in major depressive disorder: A meta-analysis. Journal of Psychiatric Research, 165, 95–104. [CrossRef]
- Xu, Y., Xu, L., & Zhou, H. (2023). The circadian clock and liver function in health and disease. Frontiers in Physiology, 14, 1187160.
- Yoo, B. B., & Mazmanian, S. K. (2017). The enteric network: Interactions between the immune and nervous systems of the gut. Immunity, 46(6), 910–926. [CrossRef]
- Zhou, H., Xu, P., & Xu, Y. (2022). Hepatic inflammation and its impact on central nervous system functions. Frontiers in Neuroscience, 16, 858612. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



