Submitted:
15 August 2025
Posted:
18 August 2025
You are already at the latest version
Abstract
The emergence of purinergic signaling has given ATP and extracellular nucleotides a whole new perspective. This concept refers to the fact that extracellular nucleotides released by most cells act as signaling molecules via specific membrane receptors called purinergic receptors, thus regulating several cellular functions. Although purinergic signaling has been well studied in several physiological systems, recent work has shown that this signaling pathway is also essential in the immune system. In this minireview, we will discuss the complex role of purinergic signaling in the regulation of effector T cell functions, including migration, differentiation, and activation. We will also cover the role of P2 receptors in the development of autoimmune diseases. Un-derstanding how P2 receptors regulate effector T cells will likely further our under-standing of protective immunity and immune pathogenesis and may lead to new therapeutic approaches and agents in immune diseases
Keywords:
P2 receptors
; effector T cells
; human Th17
; autoimmunity
1. Introduction
Nucleotides are widely dispersed molecules that are essential for regulating various biological processes. They are the basic building blocks of nucleic acids and play crucial functions as cofactors, energy intermediates, and allosteric modulators. Furthermore, they also act as ubiquitous triggers for the release of intracellular messengers [1,2,3]. Burnstock introduced the term "purinergic signalling" in the 1970s to emphasize the role of the nucleoside adenosine and the purine nucleotides ATP and ADP in the intricate process of intercellular communication [4]. Initially, there was considerable skepticism due to the unusual possibility that cells, which are thought to be in a state of normal physiological functioning, might release ATP into the extracellular environment [1,2]. Purinergic signaling has undergone significant expansion since then, and it is now clear that extracellular nucleotides, particularly ATP, are widely dispersed signaling molecules released during neurotransmission and in response to various cellular stressors, including osmotic swelling, mechanical deformation, hypoxia, and cytotoxic substances [1,2]. Following their initial classification as neurotransmitters, ATP and its dephosphorylated derivatives have been found to serve as signaling molecules in various physiological processes [2].
In a multicellular system, purinergic signaling enables individual cells to adjust their reactions, ultimately benefiting the organism as a whole [5]. Three fundamental components make up purinergic signaling: (i) mechanisms that generate and release ATP into the pericellular space; (ii) Purinergic receptors that detect released ATP and its metabolites and trigger intracellular signals that control cell functions; (iii) mechanisms that stop purinergic signaling through cellular re-uptake, enzymatic breakdown of ATP, or simple diffusion of ATP and its metabolites away from cells [5].
The release of nucleotides occurs under both physiological and pathological conditions, either through cell lysis or via other mechanisms, including the exocytosis of ATP-containing vesicles and nucleotide-permeable membrane channels, such as connexin and pannexin semichannels [1,6,7]. Once in the extracellular environment, ATP acts as a signaling molecule via so-called purinergic type 2 (P2) receptors to regulate cellular functions. Purinergic signaling can act in an autocrine manner, i.e., on the cells that released it, or in a paracrine manner on neighboring cells. P2 receptors comprise two subfamilies: the P2Y and P2X receptors [8,9]. P2Y receptors are G-protein-coupled metabotropic receptors that, in addition to ATP, can bind other nucleotides such as ADP or UTP. There are eight functional P2Y receptors, namely P2Y1, P2Y2, P2Y4, P2Y6, P2Y11, P2Y12, P2Y13, and P2Y14 [10,11]. P2X receptors, on the other hand, are ionotropic receptors that bind exclusively to ATP and act as ion channels, inducing ionic movements, including Na+, K+, and Ca2+ ions [11]. The P2X receptor subfamily comprises seven subtypes (P2X1–P2X7) with varying sensitivities to ATP [12,13].
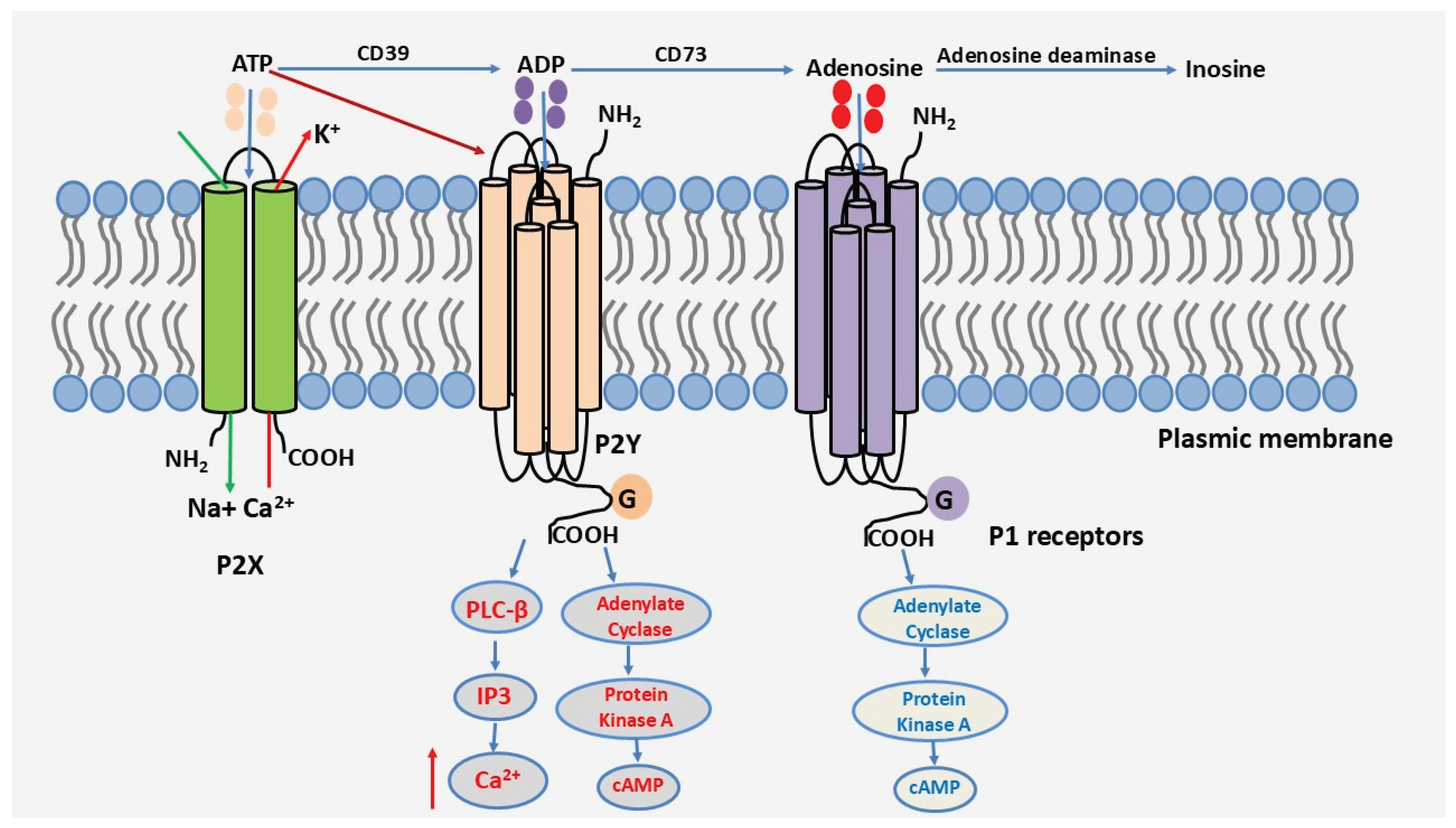
After exerting its effect on P2 receptors, ATP comes into contact with ectonucleotidases expressed on the surface of most cell types, with distinct distribution profiles [14]. ATP will be sequentially degraded by CD39 and CD73 nucleotidases, leading to the subsequent generation of adenosine, a biologically active metabolite that activates P1 receptors [15,16,17]. P1 receptors belong to the family of G protein-coupled transmembrane receptors. There are four subtypes of adenosine receptors, each with distinct affinity and signaling properties: A1, A2A, A2B, and A3 [18,19]. These receptors participate in terminating the action of ATP and P2 receptors, often exhibiting anti-inflammatory effects. Of note, adenosine can also be degraded by adenosine deaminase to inosine. A schematic representation of the purinergic signaling cascade is shown in Figure 1.
This review will mainly discuss P2 receptors, which correspond to the purinergic receptors most studied and involved in T cells, and in the pro-inflammatory response.
Figure 1.
The purinergic signaling cascade.
2. P2 Receptor Expression in the Immune System
P2 receptors are expressed, either constitutively or following cellular activation, on a range of cells, including immune cells [20]. Several studies have demonstrated the importance of extracellular ATP in the activation processes and regulation of various immune system cell functions [20,21]. Furthermore, ATP released by damaged cells acts as a danger signal. Indeed, when cellular damage occurs, for example, following tissue injury or when cells become apoptotic or necrotic, the ATP released into the extracellular space induces several cellular responses, including the recruitment of innate immune cells to the injured site, to ensure clearance of the danger [22,23,24]. Released ATP can have autocrine and paracrine effects not only on innate immune cells, such as neutrophils, macrophages, dendritic cells, and natural killer (NK) cells, but also on adaptive immune cells, including T cells [14,21]. This review will focus on the role of purinergic signaling in T cells, with a particular emphasis on effector T cells.
3. Role of P2 Receptors in T Cell Activation and Chemotaxis
Purinergic signaling has been much less studied in T cells compared to other leukocytes. However, T cells can also release ATP, which facilitates autocrine T cell activation and intercellular communication [25,26,27]. Several stimuli induce ATP release from T cells, including during T cell activation upon antigen recognition of naive T cells by the T cell receptor (TCR), mechanical stimulation, membrane deformation, and osmotic stress, among others. ATP released by semichannels, particularly pannexin-1, plays a critical role in the activation of naive T cells [28]. Several studies have demonstrated that T cells can express multiple P2 receptors, including P2X1, P2X4, P2X5, P2X7, and P2Y11 [17,20,25,27,29,30,31,32,33,34]. Notably, the P2X5 receptor is non-functional in humans [35].
Initial studies in the context of T cell activation showed that TCR activation induces ATP release and autocrine stimulation of P2X1, P2X4, and P2X7 receptors. Activation of these receptors contributes to increased intracellular calcium, the activation of the transcription factor NFAT, and IL-2 expression, all of which are associated with CD4+ T cell proliferation [22,23,25,36]. From a functional point of view, activation of the TCR/CD28 complex of CD4+ T cells during immunological synapse formation induces ATP production from mitochondria. It causes the translocation of pannexin-1 semichannels, as well as P2X1, P2X4, and P2X7 receptors to the immunological synapse to orchestrate autocrine purinergic signaling, leading to T cell activation, cytokine production and proliferation [33].
However, more recent studies have shown that CD4+ T cell activation and proliferation are primarily and strongly dependent on P2X4, but not on the P2X1 and P2X7 receptors [25,34]. Rather, the P2X1 receptor has been implicated in maintaining the metabolism of quiescent T cells, which prepares them for antigen recognition, but is not required for T cell activation [37]. P2X7 receptors are found at the immunological synapse and are also uniformly distributed on the cell surface during lymphocyte activation. This distribution would allow T cells to remain sensitive to ATP generated by tissues and cells that are not directly involved in antigen presentation [14]. P2X7 plays a role in the apoptosis of naive CD4+ T cells through a mechanism involving ERK1/2 signaling [38,39]. In addition to P2X receptors, the P2Y11 receptor supports T lymphocyte activation by directing mitochondrial trafficking to the immunological synapse [40]. On the other hand, studies have shown that the P2Y11 receptor has a protective effect against the apoptotic effects of the P2X7 receptor in naive CD4+ T cells, via a mechanism that involves inhibiting the P2X7 receptor's ability to form nonspecific pores [41,42]. In addition to the P2Y11 receptor, although the P2Y1 receptor is weakly expressed in T cells, a study has shown that it can also participate in the activation of naive T cells [27].
Taken together, these studies suggest that different P2 receptors play distinct roles in T cell physiology, depending on the steps involved in T cell activation, with a significant role assigned to the P2X4 receptor [43,44]. These studies also suggest the existence of two distinct autocrine purinergic signaling systems in T cells: one system that facilitates T cell vigilance and involves the P2X1 receptor, and a second system that regulates T cell functional responses following antigen recognition and is predominantly dependent on the P2X4 receptor [37,43].
T cells must migrate to encounter antigen-presenting cells (APCs) and to execute their varied functions in immune defense and inflammation [25]. In addition to T cell activation, purinergic signaling is also essential for chemotaxis and migration of T cells towards antigen-presenting cells in response to the chemokine CXCL12 or SDF-1α [25]. To this end, P2X4 and P2Y11 receptors have been identified as critical. These receptors act synergistically to orchestrate the metabolic program that regulates T cell polarization and migration [25,27,45]. Indeed, P2X4 will localize in front of the cell in the direction of migration by amplifying the chemokine signal through autocrine stimulation. At the same time, the P2Y11 receptor participates in the retraction of the uropods of migrating T cells [45].
Most of the above studies have been conducted with peripheral blood CD4+ T cells and in the context of naïve CD4+ T cell activation. Since the primary focus of this review is on effector T cells and autoimmunity, we will not elaborate further on the associated mechanisms. These have been elegantly addressed by the research group of W.G. Junger, and the readers are invited to check these papers for more details [14,26,27,37,40].
4. Role of P2 Receptors in Effector T Cells
While the role of P2 receptors in T cell activation is established, few studies, especially human studies, have investigated how purinergic receptors regulate effector T cell functions. Several studies have addressed this issue by utilizing animal models of autoimmunity. Th17 cells represent a crucial effector T cell subset in the development of autoimmune diseases [46,47], and growing evidence suggests that P2 receptors are crucial for the functions of Th17 cells and in the regulation of autoimmune diseases. In the following, we will discuss these findings as well as address other effector T cell subsets, including Th1, Th2, and Tregs
4.1. Role of P2 Receptors in Th17 Differentiation and Activation
Naïve CD4+ T cells are polarized towards the Th17 lineage during antigen recognition and in the presence of specific cytokines including IL-6, TGFβ, IL-23 and IL-1β, which lead to the expression of the transcription factor RORc, the master regulator of the Th17 lineage, which in turn, lead to the production of IL-17 cytokine. Under certain circumstances like in response to IL-6, Tregs can also differentiate into inflammatory Th17 cells.
Little work has been conducted on how P2 receptors regulate Th17 cells. Nevertheless, early work reported that P2 receptors could influence Th17 differentiation indirectly by acting on dendritic cells during antigen presentation. ATP, via the P2X7 receptor, acts through dendritic cells to promote Th17 generation in a mouse model of ovalbumin-induced asthma [48]. Similarly, P2X7 receptor was shown to enhance Th17 cell differentiation and collagen-induced arthritis in mice by enhancing the production of Th17 cell-polarizing cytokines by dendritic cells [22]. In a mouse model of experimental autoimmune encephalomyelitis (EAE), it was found that P2Y6 receptor signaling inhibits the production by dendritic cells of the Th1- and Th17-polarizing cytokines IL-12 and IL-23, respectively, and leads to the inhibition of the differentiation of Th1 and Th17 subpopulations and to the reduction of EAE severity [49]. Furthermore, during the acute inflammation of inflammatory bowel disease, IL-6 increases ATP production and activates the P2X7 receptor on Tregs, which induces their conversion into Th17 cells.[50]. Along these lines, in an allergy mouse model, mast cells can counteract Tregs partially through IL-6-mediated Tregs differentiation towards Th17 cells [51].
Although the above studies performed in mouse models of inflammation provided evidence of the role of P2 receptors in the regulation of Th17 cells, it is not clear if P2 receptors can directly regulate effector T cell differentiation and whether this can occur in human effector T cells. In this context, we recently reported that P2X4 is a key player in the differentiation of human Th17 cells. We have shown that human Th17 cells exhibit a P2 receptor expression profile similar to that of naive CD4+ T cells, although their expression was higher on Th17 cells. Thus, Th17 cells express mostly P2X4, P2X5, P2X7, and P2Y11 receptors along with weak levels of P2X1, and we have identified the P2X4 receptor as a critical pathway for the differentiation and activation of human Th17 cells [34]. Indeed, inhibition of these receptors showed that only P2X4 receptor inhibition interfered with the differentiation and generation of human Th17 cells from naive CD4+ T cells isolated from peripheral blood. In contrast, P2X4 inhibition did not affect the differentiation of human Th1 cells, a T cell subpopulation that also plays a vital role in the development of autoimmune diseases [52]. We also examined whether the different receptors regulated cellular expansion and proliferation during the differentiation process. 75% fewer differentiated Th17 cells in comparison with control cells were obtained when the P2X4 receptor was blocked. However, the P2X7 and P2Y11 inhibitors did not affect the cellular expansion of Th17 cells but decreased Th1 cell growth. These findings suggest that the purinergic receptor P2X4 is necessary for the proliferation of both human Th1 and Th17 cells but only affects Th17 differentiation. The requirement of P2X4 for both Th1 and Th17 proliferation is thought to be due to the fact that naive CD4+ T cells require P2X4 activation for proliferation [25].
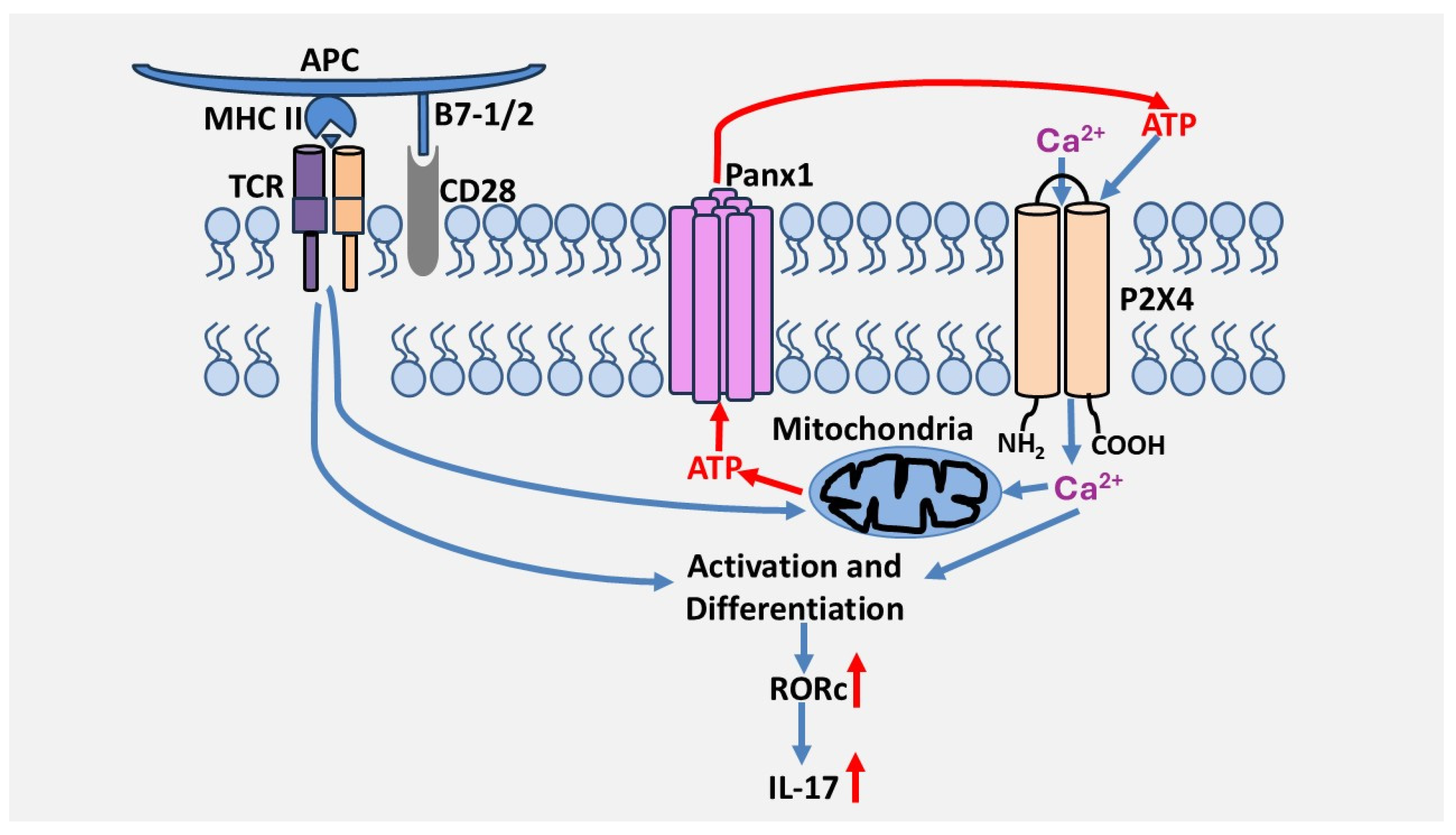
We also investigated the involvement of P2 receptors in the reactivation of Th17 cells, which refers to the capacity of polarized/differentiated Th17 cells to produce IL-17. To this end, receptor antagonists were not added during differentiation but only during the reactivation of polarized Th17 cells with anti-CD3/CD28 antibodies. Following this activation, we found that inhibition of the P2X4 receptor with 5-BDBD decreased IL-17 production but had no effect on IFN-γ production. On the other hand, inhibition of P2X7 and P2Y11 receptors did not affect the output of either cytokine. Furthermore, combined inhibition of P2X4, P2X7, and P2Y11 receptors did not produce a more potent inhibition on IL-17 production than inhibition of the P2X4 receptor alone. These results suggest that P2X4 is the most critical P2 receptor in IL-17 production. This is supported by the fact that the P2X4 receptor is involved in the expression of RORc but not T-bet. These two transcription factors are the primary regulators of IL-17 and INF-γ gene expression, respectively [34]. These studies suggest that purinergic signaling differentially regulates helper T cell programs and identify for the first time the P2X4 receptor as a primary receptor in the activation and differentiation of human Th17 lymphocytes. However, it remains to be determined by which mechanisms the P2X4 receptor contributes to the differentiation of human Th17 cells. Interestingly, P2X4 also regulates human Th2 activation, suggesting it could also be important in Th2-dependent immunity [34]. A model by which the P2X4 receptor promotes Th17 cell differentiation and activation is depicted in Figure 2.
Figure 2.
Proposed model of the role of the P2X4 receptor in the regulation of Th17 cells. T cell activation through the TCR/CD28 complex in the presence of Th17-polarizing cytokines induces ATP release via pannexin-1 channels, which leads to P2X4 activation, which is necessary for TCR/CD28-induced RORc expression levels, Th17 cell differentiation, and IL-17 production.
Figure 2.
Proposed model of the role of the P2X4 receptor in the regulation of Th17 cells. T cell activation through the TCR/CD28 complex in the presence of Th17-polarizing cytokines induces ATP release via pannexin-1 channels, which leads to P2X4 activation, which is necessary for TCR/CD28-induced RORc expression levels, Th17 cell differentiation, and IL-17 production.
4.2. Role of P2 Receptors in Th17 Cell Migration
After their differentiation in lymphoid organs, Th17 cells migrate through extracellular matrix of target tissues to reach inflammatory sites where their reactivation leads, to the production of IL-17, which orchestrates the inflammatory response and tissue damage [53,54]. Although the role of P2 receptors has been demonstrated in the chemotaxis of immune cells including T cells [55], their involvement in Th17 adhesion and migration through extracellular matrix has not been studied. T cell migration through extracellular matrix requires the intervention of integrins, a large family of α/β transmembrane receptors that play a key role in cell-cell interactions and cell adhesion [55,56].
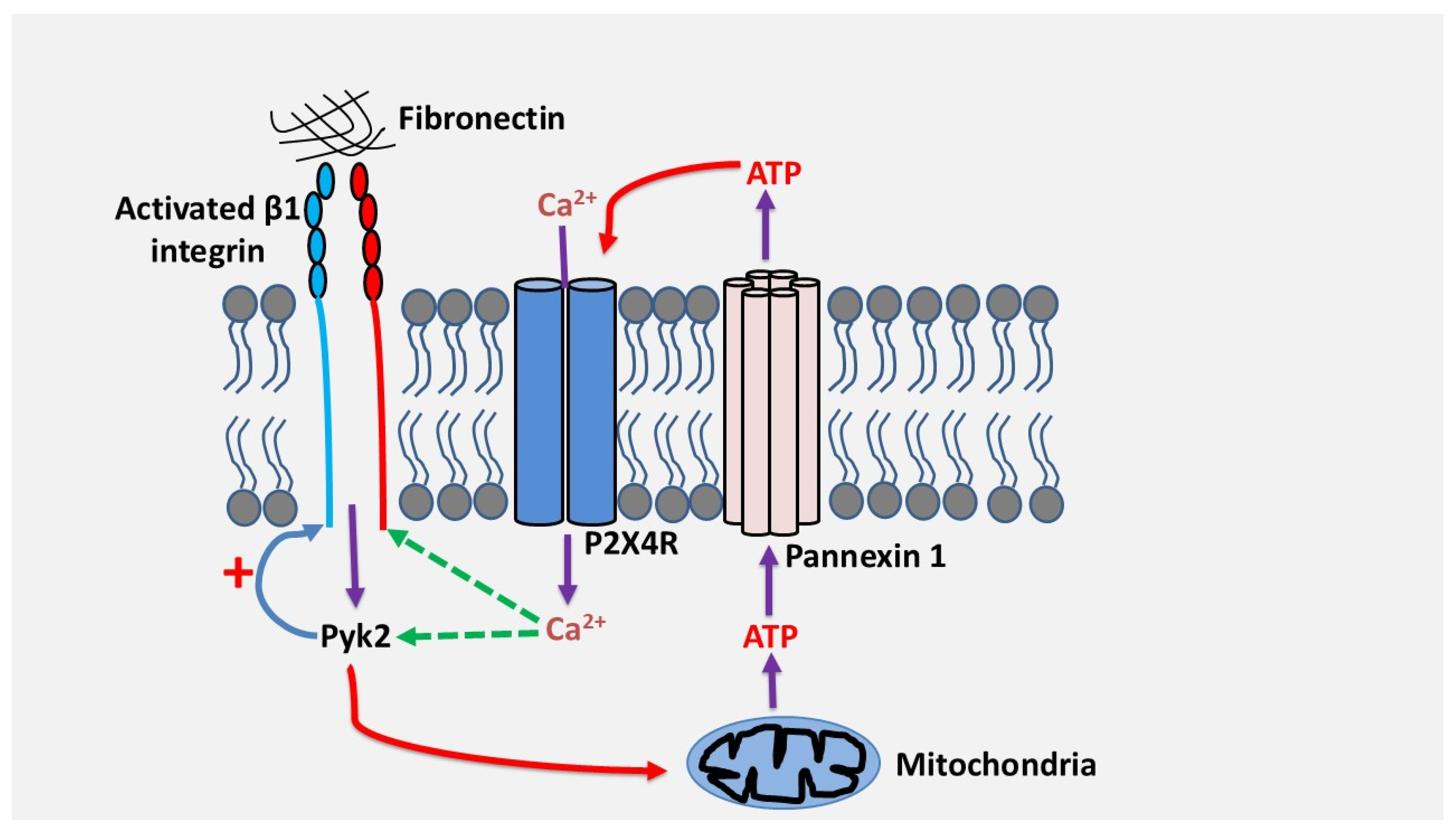
We have demonstrated in our laboratory that the adhesion and migration of human Th17 cells through fibronectin, a major extracellular matrix protein whose expression is strongly increased in inflammatory tissues, depends on the signaling of the purinergic receptor P2X4 [57]. Adhesion to fibronectin via β1 integrins, α4β1 and α5β1, induces a sustained release of ATP in quantities sufficient to activate the P2X4 receptor [57]. This occurred via mitochondrial ATP and pannexin-1 channels. Inhibition of the P2X4 receptor but not of the P2X7 and P2Y11 receptors inhibited the adhesion and migration of Th17 cells through fibronectin, suggesting the existence of an autocrine activation pathway between β1 integrins and the P2X4 receptor [57]. Mechanistically, the P2X4 receptor, but not other receptors, strengthens the activation of β1 integrins , which is important to enhance cell adhesion and migration [57]. This likely occurred through increased intracellular calcium via P2X4 and activation of the focal adhesion kinase PYK2, which can be activated by calcium, and which plays a critical role in cell adhesion and migration. On the other hand, we found that PYK2, was also essential for fibronectin-induced ATP release suggesting a positive feedback loop of activation regulation between integrin β1, P2X4 receptor and PYK2 in human Th17 cells [57]. Based on these results, the model below (Figure 3) illustrates how P2X4 receptor regulates Th17 cell migration in fibronectin. It remains to be determined if Th17 cells and other leukocytes also depend on purinergic signaling and P2X4 to migrate in different contexts and invade other matrices like collagens or laminins. However, human Th17 cells migrate through laminin by using P2X4 receptor (unpublished observations) suggesting that the implication of purinergic receptors in leukocyte migration and invasion could be a general mechanism. This is reinforced by studies that reported that activation of P2Y1 and P2Y12 receptors induced calcium-dependent activation of platelet integrin αIIbβ3 in megakaryocytes [58] and that Thy-1-induced astrocyte migration depend on activation of the P2X7 receptor by integrin αvβ3 [59].
Figure 3.
Proposed model by which the P2X4 receptor promotes adhesion and migration of human Th17 lymphocytes via β1 integrins. Cell adhesion to fibronectin via β1 integrin activates PYK2, leading to ATP release through pannexin-1. ATP/P2X4 receptor signaling triggers calcium influx, which in turn activates PYK2, resulting in enhanced β1 integrin activation, further promoting adhesion and inducing migration of Th17 cells.
Figure 3.
Proposed model by which the P2X4 receptor promotes adhesion and migration of human Th17 lymphocytes via β1 integrins. Cell adhesion to fibronectin via β1 integrin activates PYK2, leading to ATP release through pannexin-1. ATP/P2X4 receptor signaling triggers calcium influx, which in turn activates PYK2, resulting in enhanced β1 integrin activation, further promoting adhesion and inducing migration of Th17 cells.
5. P2 Receptors in Effector T Cells and Autoimmune Diseases
Autoimmune diseases are characterized by loss of immune tolerance, the presence of autoreactive lymphocytes, and the generation of autoantibodies against self-components and persistent inflammation leading to tissue damage [60].
It is now widely accepted that P2 receptors play a crucial role in regulating the inflammatory response by activating immune cells [17,20,61]. The propagation cascade of the inflammatory response and tissue damage can lead to the release of a significant amount of ATP and the stimulation of P2 receptors on immune cells, resulting in a positive feedback mechanism that extends the inflammation response [62]. Furthermore, given the role of purinergic signaling in effector T cells strongly suggest that P2 receptors are important players in the development of autoimmune diseases.
5.1. Rheumatoid Arthritis
The role of purinergic signaling in the development of rheumatoid arthritis was suggested in the 1990s, following the proposal of adenosine as a therapeutic option [63]. Most studies have shown that ATP in rheumatoid arthritis acts via the P2X7 receptor. Indeed, the expression and function of the P2X7 receptor are increased on immune cells isolated from patients with arthritis [64,65,66].
Additionally, the P2X7 receptor enhances the production of cathepsins by macrophages, which may promote bone resorption associated with rheumatoid arthritis [67]. Finally, the promoting effect of P2X7 receptor on Th17 cell differentiation was investigated in CD4+ T cells co-cultured with dendritic cells in the collagen-induced arthritis mouse model [22]. The results of this study revealed that blocking P2X7 with antagonists decreases the development of arthritis in mice by inhibiting Th17 differentiation, which occurs after blocking the expression of Th17-polarizing cytokines (IL-1β, TGF-β1, IL-23, and IL-6) by dendritic cells [22]. However, one study showed that monocytes and lymphocytes from arthritic patients exhibit reduced expression of the P2X7R gene [65]. In addition, clinical trials targeting the P2X7 receptor have had no effect in the treatment of rheumatoid arthritis, suggesting that the P2X7 receptor is insufficient for the development of rheumatoid arthritis and that other receptors are also important [68]. Indeed, several studies have shown that the P2X7 receptor requires the presence of the P2X4 receptor for optimal functioning. [44,69,70,71,72].
The P2X4 receptor has been linked to joint pain [73] and the upregulation of human Th17 cell differentiation and migration [34,57]. Along these lines, we also showed that the blockade of P2X4 receptor decreased the production of IL-17 by 30-40% in activated effector/memory CD4+ T cells isolated from the peripheral blood of patients with rheumatoid arthritis [34]. In contrast, inhibition of the P2X7 receptor had no effect on IL-17 production, which may explain why P2X7 inhibitors failed in clinical trials. The blockade of P2X4 and P2X7 also did not affect the production of IFN-γ by rheumatoid effector/memory CD4+ T cells, demonstrating no influence on Th1 function [34]. These results suggest that the P2X4 receptor is a pathogenic pathway in rheumatoid arthritis, primarily through the impairment of Th17 function [34]. Moreover, the P2X4 receptor antagonist, 5-BDBD, also reduced the severity of collagen-induced arthritis in mice by inhibiting Th17 activation [34]. Along these lines, the P2X4 antisense oligonucleotides reduced synovial inflammation in the collagen-induced mouse model of arthritis by modulating the serum levels of IL-1β, TNF-α, IL-6, and IL-17 [74]. Together, these results suggest that the P2X4 receptor is a critical pathway in the development of rheumatoid arthritis, arguing in favor of assessing P2X4 inhibitors in clinical trials for the treatment of rheumatoid arthritis.
Regarding P2Y receptors, the P2Y11 receptor has been implicated in the cytokine-induced inflammatory response of patients' primary fibroblast-like synoviocytes [75]. Indeed, blockade of this receptor with the specific antagonist NF340 inhibited IL-1β-induced expression of TNF-α and IL-6 [75]. The P2Y6 and P2Y12 receptors participate in bone resorption during osteoporosis via osteoclast activation [76,77], while the P2Y14 receptor has the opposite effect by stimulating osteogenesis via osteoblasts [78,79]. However, the role of these P2Y receptors in animal models of arthritis has not been investigated.
5.2. Systemic Lupus Erythematosus
The P2X7 receptor is involved in systemic lupus erythematosus through the activation of the NLRP3 inflammasome and increased production of IL-1β and IL-18 by lupus patient-derived macrophages [80,81], thereby contributing to the cardiovascular, cutaneous, and renal manifestations of lupus [82,83,84]. Th1 and Th17 cells exhibit higher expression of the P2X7 receptor in lupus patients compared to controls, and P2X7 levels on Th17 cells correlate with diseases activities of both lupus and rheumatoid arthritis, suggesting an important role for P2X7 in those two diseases. [85]. However, in experimental murine lupus, P2X7 receptor plays a protective role by limiting the expansion of pathological T follicular helper cells through the induction of pyroptosis and decreasing the generation of autoreactive antibodies [86]. These contrasting studies could be due to the presence of different P2X7 receptor variants in humans, each with distinct functional characteristics [87]. In this context, a recent study identified a P2Y8 receptor variant that has a role in B cell self-tolerance to be dysregulated in lupus patients, indicating a role for P2Y8 in immunological tolerance and lupus pathogenesis [88].
As B cells and plasma cells are critical effectors in lupus, it has recently been shown that bone marrow plasma cells (PCs) sense extracellular ATP via P2X4 receptor [89]. In this study, the authors demonstrated that bone marrow PCs utilize the ligand-gated purinergic ion channel P2X4 to sense extracellular ATP released by bone marrow osteoblasts through the gap junction protein Pannexin 3 (Panx3) [89]. Mutating P2X4 in developing B-lineage cells using Mb1-Cre resulted in lower serum antibody titers and significantly reduced numbers of bone marrow PCs, demonstrating that the P2X4 receptor is necessary to establish normal bone marrow PC populations [89]. The P2X4-specific inhibitor 5-BDBD abrogated the impact of extracellular ATP on PCs in vitro and depleted bone marrow PCs in vivo [89]. P2X4 blockade also reduced autoantibody titers and kidney disease in two mouse models of systemic lupus erythematosus characterized by serum antibodies against double-stranded DNA (dsDNA) and progressive proteinuria [89]. While the role of P2X4 on Th17 cells in lupus has not been examined, it is likely from the studies on rheumatoid arthritis that inhibition of Th17 cells also contribute along with the inhibition of plasma cells to the protective effect of P2X4 antagonist in lupus.
5.3. Multiple Sclerosis
Genetic mutations in the P2X4 and P2X7 receptors, which result in a functional loss of these receptors, have been associated with familial multiple sclerosis [90]. Along these lines, activating P2X4 receptor promoted the remyelination response and improved clinical signs of experimental EAE in mice [91].
Conflicting results have been reported regarding the function of P2Y12 in the development of EAE. One study reported that P2Y12 knockout mice were protected from EAE [92]. Indeed, deficiency of P2Y12 led to a sharp decrease in the percentage of Th17 cells, accompanied by decreased IL-17A production and a low mRNA level of Th17-related genes [92]. In contrast, a second study found that the absence of P2Y12 exacerbated the disease [93]. In this study, the deletion of P2Y12 boosted the expression of IL-17A in the serum and the proportion of Th17 cells in the spleen and central nervous system. Loss of P2Y12 significantly increased the production of the Th17-polarizing cytokine IL-23 in contrast to the wild-type (WT) BMDCs [93]. FACS analysis also indicated that the culture supernatant from P2Y12-deficient DCs promoted the differentiation of more naïve CD4+ T cells into Th17 cells [93]. Furthermore, a pro-inflammatory environment, which damages neurons, has been associated with loss of P2Y12 receptor expression in microglia from multiple sclerosis patients [94].
Loss of P2Y6 receptor expression has also been associated with exacerbating EAE in mice [49]. Increased expression of IL-12 and IL-23 was detected in P2Y6-deficient bone marrow-derived dendritic cells compared to controls [49]. This increased expression was correlated with enhanced Th1/Th17 polarization by mature dendritic cells [49]. These data demonstrate that P2Y6 functions as a crucial regulator of DC maturation, and its deletion results in worsened EAE [49].
Together, these studies tend to suggest that P2 receptors may play a protective role in the development of multiple sclerosis.
5.4. Inflammatory Bowel Diseases (IBD)
P2X7 receptor plays a vital role in the development of inflammatory bowel diseases. It upregulates the production of pro-inflammatory cytokines (Myd88, NF-κB, IL-6, IL-1β, and TNF-α) and facilitates the infiltration of immune cells in animal models of ulcerative colitis [95,96,97]. In another study, it has been demonstrated that P2X7 receptor activation promotes intestinal inflammation in TNBS (2,4,6-trinitrobenzenesulfonic acid) and oxazolone-induced colitis, two models of chemically induced colitis. The study revealed that P2X7 knockout mice have a higher number of FoxP3-positiveT regulatory cells (Tregs) in the colonic lamina propria [98]. Furthermore, flow cytometry analysis of lymph nodes revealed that P2X7R activation by ATP triggered death and retention of Tregs, thereby impairing gut immune tolerance [98]. This study provides substantial evidence in support of a significant role for the P2X7 receptor in the establishment of the enhanced inflammatory response during IBD, by promoting Treg cell death and compromising immune system tolerance in the gut [98].
Regarding P2Y receptors, increased expression of the P2Y6 receptor on epithelial cells has been associated with inflammation in DSS (dextran sodium sulfate)-induced mouse colitis [99]. However, another study highlighted a protective role for this receptor in regulating the quality of mucus in a DSS-induced colitis model. Indeed, it has been reported that P2Y6-deficient mice exhibited poor mucus quality and were more sensitive to DSS, resulting in increased disease activity index [100]. A study also demonstrated that deletion of P2Y6 receptor in mice exacerbated DSS-induced intestinal colitis by increasing the infiltration of Th17/Th1 cells and neutrophils in their colons, which correlated with increased levels of IFN-γ and IL-17A in the sera as well as increased mRNA levels of IFN-γ, IL-17A, IL-6, IL-23, and IL-1β in P2ry6-/- colons [101]. However, it is not clear if P2Y6 is expressed on Th1/Th17 cells and if its effect is direct or mediated through the dysregulation of immune tolerance mechanisms.
The importance of purinergic signaling in the regulation of IBD comes also from studies on ectonucleotidases. Indeed, the use of knockout mice showed in preclinical models that both CD39 and CD73 can protect from IBD likely through the generation of Tregs, thus limiting the expansion of pathogenic Th17 cells [102,103,104]. These pathways however seem to be dysregulated in patients and therefore therapeutic agents that can enhance the development of CD39+Tregs could be considered as promising therapeutic strategy. This is the case with two agents, namely indole-3′-carbonyl-thiazole-4-carboxylic acid methyl ester (ITE) and unconjugated bilirubin [102,105,106]. Of note, treatment with GM-CSF, which was reported to reduce clinical symptoms of Crohn’ disease, led to the expression of CD39 and CD73 on activated monocytes which reduce inflammation in the context of DSS-induced colitis model, either directly by hydrolyzing pro-inflammatory extracellular ATP into adenosine or indirectly by promoting the formation of Tregs from naïve T cells [107].
The majority of studies on P2 receptors in IBD have been conducted in models of chemical-induced colitis in mice. It will be interesting to study these receptors in T cell-dependent models like IL-10 knock-out mice and the T cell transfer model.
5.5. Liver Autoimmunity
Autoimmune hepatitis (AIH) is an organ-specific autoimmune illness characterized by hypergammaglobulinemia, autoantibody positivity, and histological presence of interface hepatitis [108]. It is a severe liver disease caused by the abnormal activation of CD8+ and CD4+ effector T cells, including Th17 cells [109].
P2X4-deficient receptor mice have been utilized to investigate the role of this receptor in three distinct models of acute liver injury induced by concanavalin A (ConA), carbon tetrachloride, and acetaminophen [110]. The findings of the study revealed that ablation of the P2X4 receptor significantly reduced the severity of hepatitis in mice caused by ConA by restraining inflammation, oxidation, and cell death programs (apoptosis, autophagy, and NLRP3 inflammasome-activated pyroptosis [110]. Accordingly, ELISA analysis revealed increased levels of serum inflammatory mediators IL-1β, IL-6, IL-17A, IFN-γ, and TNF-α in ConA-treated WT mice when compared to P2X4R-/- mice [110]. Furthermore, treatment with the P2X4R antagonist (5-BDBD) alleviated ConA-induced autoimmune hepatitis [110]. This study is the first to demonstrate that the absence of the P2X4 receptor may reduce immune-mediated liver damage, potentially by inhibiting inflammatory, oxidative, and programmed cell death processes [110]. The study also emphasizes that ConA-induced acute hepatitis requires the P2X4 receptor activation.
Research conducted with AIH patients has indicated functional impairment in CD39+ Treg cells. The authors demonstrated that Tregs expressing the ectonucleotidase CD39 are present at low levels and are also dysfunctional. Hence, these Tregs fail to hydrolyze proinflammatory ATP adequately, are unable to synthesize adenosine, and subsequently to inhibit Th17 cell immunity [111].
Table 1 below summarizes the established roles of P2 receptors and their function in autoimmune diseases.
6. Conclusions
This article provides a review of the role of P2 receptors in effector T cells and their implications in autoimmune disease. Thus, P2 receptors can promote effector T cells' function either directly or by acting on the production of polarizing cytokines by antigen-presenting cells. On the other hand, P2 responses can also inhibit effector T cells and limit inflammatory response, mostly via dendritic cells or by enhancing Treg cells through CD39. Additional work is needed to further understand how P2 receptors affect effector T cells, especially with human models, and decipher how P2 receptors regulate the transcriptional programs of effector T cells. There are also some conflicting studies regarding the implication of P2 receptors in autoimmune diseases. This may be due to the use of different or less specific antagonists, or to variations in study models. Further studies are necessary to elucidate the role of purinergic signaling in effector T cells and the progression of autoimmune diseases by using more specific antagonists and uniformizing animal models.
Conversely, studies on the P2X4 receptor seem to be conclusive. Indeed, P2X4 has been implicated in autoimmune arthritis, lupus, and autoimmune hepatitis. This receptor also seems to be protective in the EAE model. Similarly, P2Y6 is protective in IBD and EAE. Clinical trials are necessary to determine whether these receptors constitute viable therapeutic targets in those autoimmune diseases.
Author Contributions
All authors contributed to the manuscript’s preparation and approved the final submitted version. F.A supervised this work.
Funding
This research was funded by a grant (RGPIN-2017-06116) from the Natural Sciences and Engineering Research Council of Canada to F.A and by The Fonds de recherche du Québec through the research center funding grant (#30641). C.H was supported by a doctoral scholarship from Fonds Pierre Borgeat sur les Maladies Rhumatismales of Université Laval and is a recipient of a doctoral scholarship from Fonds de Recherche du Québec. The funding sources played no role in the conception of the study or the manuscript content.
References
- Giuliani AL, Sarti AC, Di Virgilio F. Extracellular nucleotides and nucleosides as signalling molecules. Immunol Lett. 2019. 205: p. 16-24. [CrossRef]
- Vultaggio-Poma V, Falzoni S, Salvi G, Giuliani AL, Di Virgilio F. Signalling by extracellular nucleotides in health and disease. Biochim Biophys Acta Mol Cell Res. 2022. 1869: p. 119237. [CrossRef]
- Zimmermann H. Extracellular ATP and other nucleotides-ubiquitous triggers of intercellular messenger release. Purinergic Signal. 2016. 12: p. 25-57. [CrossRef]
- Burnstock G, Campbell G, Satchell D, Smythe A. Evidence that adenosine triphosphate or a related nucleotide is the transmitter substance released by non-adrenergic inhibitory nerves in the gut. Br J Pharmacol. 1970. 40: p. 668-88. [CrossRef]
- Ledderose C, Junger WG. Mitochondria Synergize With P2 Receptors to Regulate Human T Cell Function. Front Immunol. 2020. 11: p. 549889. [CrossRef]
- Evans RJ, Derkach V, Surprenant A. ATP mediates fast synaptic transmission in mammalian neurons. Nature. 1992. 357: p. 503-5. [CrossRef]
- Zimmermann H. ATP and acetylcholine, equal brethren. Neurochem Int. 2008. 52: p. 634-48. [CrossRef]
- Burnstock G. Purinergic Signalling: Therapeutic Developments. Front Pharmacol. 2017. 8: p. 661. [CrossRef]
- Kennedy C. The P2Y/P2X divide: How it began. Biochem Pharmacol. 2021. 187: p. 114408. [CrossRef]
- Jacobson KA, Delicado EG, Gachet C, Kennedy C, Von Kugelgen I, Li B, Miras-Portugal MT, Novak I, Schoneberg T, Perez-Sen R, Thor D, Wu B, Yang Z, Muller CE. Update of P2Y receptor pharmacology: IUPHAR Review 27. Br J Pharmacol. 2020. 177: p. 2413-2433. [CrossRef]
- Von Kugelgen I. Molecular pharmacology of P2Y receptor subtypes. Biochem Pharmacol. 2021. 187: p. 114361.
- Stokes L, Bidula S, Bibic L, Allum E. To Inhibit or Enhance? Is There a Benefit to Positive Allosteric Modulation of P2X Receptors? Front Pharmacol. 2020. 11: p. 627. [CrossRef]
- Sheng D, Hattori M. Recent progress in the structural biology of P2X receptors. Proteins. 2022. 90: p. 1779-1785. [CrossRef]
- Junger WG. Immune cell regulation by autocrine purinergic signalling. Nat Rev Immunol. 2011. 11: p. 201-12. [CrossRef]
- Antonioli L, Pacher P, Vizi ES, Hasko G. CD39 and CD73 in immunity and inflammation. Trends Mol Med. 2013. 19: p. 355-67.
- Allard B, Longhi MS, Robson SC, Stagg J. The ectonucleotidases CD39 and CD73: Novel checkpoint inhibitor targets. Immunol Rev. 2017. 276: p. 121-144. [CrossRef]
- Antonioli L, Blandizzi C, Pacher P, Hasko G. The Purinergic System as a Pharmacological Target for the Treatment of Immune-Mediated Inflammatory Diseases. Pharmacol Rev. 2019. 71: p. 345-382. [CrossRef]
- Alencar AKN, Montes GC, Barreiro EJ, Sudo RT, Zapata-Sudo G. Adenosine Receptors As Drug Targets for Treatment of Pulmonary Arterial Hypertension. Front Pharmacol. 2017. 8: p. 858. [CrossRef]
- Antonioli L, Fornai M, Blandizzi C, Pacher P, Hasko G. Adenosine signaling and the immune system: When a lot could be too much. Immunol Lett. 2019. 205: p. 9-15. [CrossRef]
- Jacob F, Perez Novo C, Bachert C, Van Crombruggen K. Purinergic signaling in inflammatory cells: P2 receptor expression, functional effects, and modulation of inflammatory responses. Purinergic Signal. 2013. 9: p. 285-306. [CrossRef]
- Burnstock G, Boeynaems JM. Purinergic signalling and immune cells. Purinergic Signal. 2014. 10: p. 529-64.
- Fan ZD, Zhang YY, Guo YH, Huang N, Ma HH, Huang H, Yu HG. Involvement of P2X7 receptor signaling on regulating the differentiation of Th17 cells and type II collagen-induced arthritis in mice. Sci Rep. 2016. 6: p. 35804. [CrossRef]
- Cekic C, Linden J. Purinergic regulation of the immune system. Nat Rev Immunol. 2016. 16: p. 177-92. [CrossRef]
- Chen J, Zhao Y, Liu Y. The role of nucleotides and purinergic signaling in apoptotic cell clearance - implications for chronic inflammatory diseases. Front Immunol. 2014. 5: p. 656. [CrossRef]
- Ledderose C, Liu K, Kondo Y, Slubowski CJ, Dertnig T, Denicolo S, Arbab M, Hubner J, Konrad K, Fakhari M, Lederer JA, Robson SC, Visner GA, Junger WG. Purinergic P2X4 receptors and mitochondrial ATP production regulate T cell migration. J Clin Invest. 2018. 128: p. 3583-3594. [CrossRef]
- Ledderose C, Bao Y, Lidicky M, Zipperle J, Li L, Strasser K, Shapiro NI, Junger WG. Mitochondria are gate-keepers of T cell function by producing the ATP that drives purinergic signaling. J Biol Chem. 2014. 289: p. 25936-45. [CrossRef]
- Woehrle T, Ledderose C, Rink J, Slubowski C, Junger WG. Autocrine stimulation of P2Y1 receptors is part of the purinergic signaling mechanism that regulates T cell activation. Purinergic Signal. 2019. 15: p. 127-137. [CrossRef]
- Schenk U, Westendorf AM, Radaelli E, Casati A, Ferro M, Fumagalli M, Verderio C, Buer J, Scanziani E, Grassi F. Purinergic control of T cell activation by ATP released through pannexin-1 hemichannels. Sci Signal. 2008. 1: p. ra6. [CrossRef]
- Burnstock G, Knight GE. Cellular distribution and functions of P2 receptor subtypes in different systems. Int Rev Cytol. 2004. 240: p. 31-304.
- Manohar M, Hirsh MI, Chen Y, Woehrle T, Karande AA, Junger WG. ATP release and autocrine signaling through P2X4 receptors regulate gammadelta T cell activation. J Leukoc Biol. 2012. 92: p. 787-94.
- Kennedy C. P2Y11 Receptors: Properties, Distribution and Functions. Adv Exp Med Biol. 2017. 1051: p. 107-122.
- Di Virgilio F, Vuerich M. Purinergic signaling in the immune system. Auton Neurosci. 2015. 191: p. 117-23. [CrossRef]
- Woehrle T, Yip L, Elkhal A, Sumi Y, Chen Y, Yao Y, Insel PA, Junger WG. Pannexin-1 hemichannel-mediated ATP release together with P2X1 and P2X4 receptors regulate T-cell activation at the immune synapse. Blood. 2010. 116: p. 3475-84. [CrossRef]
- Hamoudi C, Zhao C, Abderrazak A, Salem M, Fortin PR, Sévigny J, Aoudjit F. The Purinergic Receptor P2X4 Promotes Th17 Activation and the Development of Arthritis. J Immunol. 2022. 208: p. 1115-1127. [CrossRef]
- Kotnis S, Bingham B, Vasilyev DV, Miller SW, Bai Y, Yeola S, Chanda PK, Bowlby MR, Kaftan EJ, Samad TA, Whiteside GT. Genetic and functional analysis of human P2X5 reveals a distinct pattern of exon 10 polymorphism with predominant expression of the nonfunctional receptor isoform. Mol Pharmacol. 2010. 77: p. 953-60. [CrossRef]
- Savio LEB, De Andrade Mello P, Da Silva CG, Coutinho-Silva R. The P2X7 Receptor in Inflammatory Diseases: Angel or Demon? Front Pharmacol. 2018. 9: p. 52.
- Ledderose C, Bao Y, Ledderose S, Woehrle T, Heinisch M, Yip L, Zhang J, Robson SC, Shapiro NI, Junger WG. Mitochondrial Dysfunction, Depleted Purinergic Signaling, and Defective T Cell Vigilance and Immune Defense. J Infect Dis. 2016. 213: p. 456-64. [CrossRef]
- Aswad F, Dennert G. P2X7 receptor expression levels determine lethal effects of a purine based danger signal in T lymphocytes. Cell Immunol. 2006. 243: p. 58-65. [CrossRef]
- Tsukimoto M, Maehata M, Harada H, Ikari A, Takagi K, Degawa M. P2X7 receptor-dependent cell death is modulated during murine T cell maturation and mediated by dual signaling pathways. J Immunol. 2006. 177: p. 2842-50. [CrossRef]
- Ledderose C, Bromberger S, Slubowski CJ, Sueyoshi K, Junger WG. Frontline Science: P2Y11 receptors support T cell activation by directing mitochondrial trafficking to the immune synapse. J Leukoc Biol. 2021. 109: p. 497-508. [CrossRef]
- Dreisig K, Kornum BR. A critical look at the function of the P2Y11 receptor. Purinergic Signal. 2016. 12: p. 427-37. [CrossRef]
- Dreisig K, Sund L, Dommer MW, Kristensen NP, Boddum K, Viste R, Fredholm S, Odum N, Jaattela M, Skov S, Kornum BR. Human P2Y11 Expression Level Affects Human P2X7 Receptor-Mediated Cell Death. Front Immunol. 2018. 9: p. 1159. [CrossRef]
- Brock VJ, Wolf IMA, Er-Lukowiak M, Lory N, Stahler T, Woelk LM, Mittrucker HW, Muller CE, Koch-Nolte F, Rissiek B, Werner R, Guse AH, Diercks BP. P2X4 and P2X7 are essential players in basal T cell activity and Ca2+ signaling milliseconds after T cell activation. Sci Adv. 2022. 8: p. eabl9770. [CrossRef]
- Suurvali J, Boudinot P, Kanellopoulos J, Ruutel Boudinot S. P2X4: A fast and sensitive purinergic receptor. Biomed J. 2017. 40: p. 245-256. [CrossRef]
- Ledderose C, Bromberger S, Slubowski CJ, Sueyoshi K, Aytan D, Shen Y, Junger WG. The purinergic receptor P2Y11 choreographs the polarization, mitochondrial metabolism, and migration of T lymphocytes. Sci Signal. 2020. 13. [CrossRef]
- Han L, Yang J, Wang X, Li D, Lv L, Li B. Th17 cells in autoimmune diseases. Front Med. 2015. 9: p. 10-9.
- Yasuda K, Takeuchi Y, Hirota K. The pathogenicity of Th17 cells in autoimmune diseases. Semin Immunopathol. 2019. 41: p. 283-297.
- Zhang F, Su X, Huang G, Xin XF, Cao EH, Shi Y, Song Y. Adenosine Triphosphate Promotes Allergen-Induced Airway Inflammation and Th17 Cell Polarization in Neutrophilic Asthma. J Immunol Res. 2017. 2017: p. 5358647. [CrossRef]
- Li Z, He C, Zhang J, Zhang H, Wei H, Wu S, Jiang W. P2Y6 Deficiency Enhances Dendritic Cell-Mediated Th1/Th17 Differentiation and Aggravates Experimental Autoimmune Encephalomyelitis. J Immunol. 2020. 205: p. 387-397. [CrossRef]
- Schenk U, Frascoli M, Proietti M, Geffers R, Traggiai E, Buer J, Ricordi C, Westendorf AM, Grassi F. ATP inhibits the generation and function of regulatory T cells through the activation of purinergic P2X receptors. Sci Signal. 2011. 4: p. ra12. [CrossRef]
- Piconese S, Gri G, Tripodo C, Musio S, Gorzanelli A, Frossi B, Pedotti R, Pucillo CE, Colombo MP. Mast cells counteract regulatory T-cell suppression through interleukin-6 and OX40/OX40L axis toward Th17-cell differentiation. Blood. 2009. 114: p. 2639-48. [CrossRef]
- Kamali AN, Noorbakhsh SM, Hamedifar H, Jadidi-Niaragh F, Yazdani R, Bautista JM, Azizi G. A role for Th1-like Th17 cells in the pathogenesis of inflammatory and autoimmune disorders. Mol Immunol. 2019. 105: p. 107-115. [CrossRef]
- Blauvelt A, Chiricozzi A. The Immunologic Role of IL-17 in Psoriasis and Psoriatic Arthritis Pathogenesis. Clin Rev Allergy Immunol. 2018. 55: p. 379-390. [CrossRef]
- Lubberts E. The IL-23-IL-17 axis in inflammatory arthritis. Nat Rev Rheumatol. 2015. 11: p. 562. [CrossRef]
- El Azreq MA, Arseneault C, Boisvert M, Page N, Allaeys I, Poubelle PE, Tessier PA, Aoudjit F. Cooperation between IL-7 Receptor and Integrin alpha2beta1 (CD49b) Drives Th17-Mediated Bone Loss. J Immunol. 2015. 195: p. 4198-209.
- El Azreq MA, Boisvert M, Cesaro A, Page N, Loubaki L, Allaeys I, Chakir J, Poubelle PE, Tessier PA, Aoudjit F. alpha2beta1 integrin regulates Th17 cell activity and its neutralization decreases the severity of collagen-induced arthritis. J Immunol. 2013. 191: p. 5941-50. [CrossRef]
- Hamoudi C, Muheidli A, Aoudjit F. beta1 Integrin induces adhesion and migration of human Th17 cells via Pyk2-dependent activation of P2X4 receptor. Immunology. 2023. 168: p. 83-95. [CrossRef]
- Bye AP, Gibbins JM, Mahaut-Smith MP. Ca2+ waves coordinate purinergic receptor-evoked integrin activation and polarization. Sci Signal. 2020. 13.
- Alvarez A, Lagos-Cabre R, Kong M, Cardenas A, Burgos-Bravo F, Schneider P, Quest AF, Leyton L. Integrin-mediated transactivation of P2X7R via hemichannel-dependent ATP release stimulates astrocyte migration. Biochim Biophys Acta. 2016. 1863: p. 2175-88. [CrossRef]
- Rai R, Cross Talk of Purinergic and Immune Signaling: Implication in Inflammatory and Pathogenic Diseases, in Purinergic System, IntechOpen, Editor. 2022: Brazil.
- Liu JP, Liu SC, Hu SQ, Lu JF, Wu CL, Hu DX, Zhang WJ. ATP ion channel P2X purinergic receptors in inflammation response. Biomed Pharmacother. 2023. 158: p. 114205. [CrossRef]
- Di Virgilio F, Sarti AC, Coutinho-Silva R. Purinergic signaling, DAMPs, and inflammation. Am J Physiol Cell Physiol. 2020. 318: p. C832-C835.
- Da Silva JLG, Passos DF, Bernardes VM, Leal DBR. ATP and adenosine: Role in the immunopathogenesis of rheumatoid arthritis. Immunol Lett. 2019. 214: p. 55-64. [CrossRef]
- Di Virgilio F, Dal Ben D, Sarti AC, Giuliani AL, Falzoni S. The P2X7 Receptor in Infection and Inflammation. Immunity. 2017. 47: p. 15-31. [CrossRef]
- Portales-Cervantes L, Nino-Moreno P, Doniz-Padilla L, Baranda-Candido L, Garcia-Hernandez M, Salgado-Bustamante M, Gonzalez-Amaro R, Portales-Perez D. Expression and function of the P2X7 purinergic receptor in patients with systemic lupus erythematosus and rheumatoid arthritis. Hum Immunol. 2010. 71: p. 818-25. [CrossRef]
- Portales-Cervantes L, Nino-Moreno P, Salgado-Bustamante M, Garcia-Hernandez MH, Baranda-Candido L, Reynaga-Hernandez E, Barajas-Lopez C, Gonzalez-Amaro R, Portales-Perez DP. The His155Tyr (489C>T) single nucleotide polymorphism of P2RX7 gene confers an enhanced function of P2X7 receptor in immune cells from patients with rheumatoid arthritis. Cell Immunol. 2012. 276: p. 168-75. [CrossRef]
- Lopez-Castejon G, Theaker J, Pelegrin P, Clifton AD, Braddock M, Surprenant A. P2X7 receptor-mediated release of cathepsins from macrophages is a cytokine-independent mechanism potentially involved in joint diseases. J Immunol. 2010. 185: p. 2611-9. [CrossRef]
- Keystone EC, Wang MM, Layton M, Hollis S, Mcinnes IB, Team DCS. Clinical evaluation of the efficacy of the P2X7 purinergic receptor antagonist AZD9056 on the signs and symptoms of rheumatoid arthritis in patients with active disease despite treatment with methotrexate or sulphasalazine. Ann Rheum Dis. 2012. 71: p. 1630-5. [CrossRef]
- Sakaki H, Fujiwaki T, Tsukimoto M, Kawano A, Harada H, Kojima S. P2X4 receptor regulates P2X7 receptor-dependent IL-1beta and IL-18 release in mouse bone marrow-derived dendritic cells. Biochem Biophys Res Commun. 2013. 432: p. 406-11. [CrossRef]
- Casas-Pruneda G, Reyes JP, Perez-Flores G, Perez-Cornejo P, Arreola J. Functional interactions between P2X4 and P2X7 receptors from mouse salivary epithelia. J Physiol. 2009. 587: p. 2887-901.
- Schneider M, Prudic K, Pippel A, Klapperstuck M, Braam U, Muller CE, Schmalzing G, Markwardt F. Interaction of Purinergic P2X4 and P2X7 Receptor Subunits. Front Pharmacol. 2017. 8: p. 860. [CrossRef]
- Antonioli L, Blandizzi C, Fornai M, Pacher P, Lee HT, Hasko G. P2X4 receptors, immunity, and sepsis. Curr Opin Pharmacol. 2019. 47: p. 65-74. [CrossRef]
- Zhang WJ, Luo HL, Zhu ZM. The role of P2X4 receptors in chronic pain: A potential pharmacological target. Biomed Pharmacother. 2020. 129: p. 110447.
- Li F, Guo N, Ma Y, Ning B, Wang Y, Kou L. Inhibition of P2X4 suppresses joint inflammation and damage in collagen-induced arthritis. Inflammation. 2014. 37: p. 146-53. [CrossRef]
- Gao F, Li X. P2Y11 receptor antagonist NF340 ameliorates inflammation in human fibroblast-like synoviocytes: An implication in rheumatoid arthritis. IUBMB Life. 2019. 71: p. 1552-1560. [CrossRef]
- Orriss IR, Wang N, Burnstock G, Arnett TR, Gartland A, Robaye B, Boeynaems JM. The P2Y6 receptor stimulates bone resorption by osteoclasts. Endocrinology. 2011. 152: p. 3706-16. [CrossRef]
- Su X, Floyd DH, Hughes A, Xiang J, Schneider JG, Uluckan O, Heller E, Deng H, Zou W, Craft CS, Wu K, Hirbe AC, Grabowska D, Eagleton MC, Townsley S, Collins L, Piwnica-Worms D, Steinberg TH, Novack DV, Conley PB, Hurchla MA, Rogers M, Weilbaecher KN. The ADP receptor P2RY12 regulates osteoclast function and pathologic bone remodeling. J Clin Invest. 2012. 122: p. 3579-92. [CrossRef]
- Mikolajewicz N, Komarova SV. Role of UDP-Sugar Receptor P2Y14 in Murine Osteoblasts. Int J Mol Sci. 2020. 21. [CrossRef]
- Lazarowski ER, Harden TK. UDP-Sugars as Extracellular Signaling Molecules: Cellular and Physiologic Consequences of P2Y14 Receptor Activation. Mol Pharmacol. 2015. 88: p. 151-60. [CrossRef]
- Kahlenberg JM, Carmona-Rivera C, Smith CK, Kaplan MJ. Neutrophil extracellular trap-associated protein activation of the NLRP3 inflammasome is enhanced in lupus macrophages. J Immunol. 2013. 190: p. 1217-26. [CrossRef]
- Di Virgilio F, Giuliani AL. Purinergic signalling in autoimmunity: A role for the P2X7R in systemic lupus erythematosus? Biomed J. 2016. 39: p. 326-338. [CrossRef]
- Kahlenberg JM, Thacker SG, Berthier CC, Cohen CD, Kretzler M, Kaplan MJ. Inflammasome activation of IL-18 results in endothelial progenitor cell dysfunction in systemic lupus erythematosus. J Immunol. 2011. 187: p. 6143-56. [CrossRef]
- Hu D, Liu X, Chen S, Bao C. Expressions of IL-18 and its binding protein in peripheral blood leukocytes and kidney tissues of lupus nephritis patients. Clin Rheumatol. 2010. 29: p. 717-21. [CrossRef]
- Wang D, Drenker M, Eiz-Vesper B, Werfel T, Wittmann M. Evidence for a pathogenetic role of interleukin-18 in cutaneous lupus erythematosus. Arthritis Rheum. 2008. 58: p. 3205-15. [CrossRef]
- Li M, Yang C, Wang Y, Song W, Jia L, Peng X, Zhao R. The Expression of P2X7 Receptor on Th1, Th17, and Regulatory T Cells in Patients with Systemic Lupus Erythematosus or Rheumatoid Arthritis and Its Correlations with Active Disease. J Immunol. 2020. 205: p. 1752-1762. [CrossRef]
- Faliti CE, Gualtierotti R, Rottoli E, Gerosa M, Perruzza L, Romagnani A, Pellegrini G, De Ponte Conti B, Rossi RL, Idzko M, Mazza EMC, Bicciato S, Traggiai E, Meroni PL, Grassi F. P2X7 receptor restrains pathogenic Tfh cell generation in systemic lupus erythematosus. J Exp Med. 2019. 216: p. 317-336. [CrossRef]
- Sluyter R. The P2X7 Receptor. Adv Exp Med Biol. 2017. 1051: p. 17-53.
- He Y, Gallman AE, Xie C, Shen Q, Ma J, Wolfreys FD, Sandy M, Arsov T, Wu X, Qin Y, Zhang P, Jiang S, Stanley M, Wu P, Tan J, Ding H, Xue H, Chen W, Xu J, Criswell LA, Nititham J, Adamski M, Kitching AR, Cook MC, Cao L, Shen N, Cyster JG, Vinuesa CG. P2RY8 variants in lupus patients uncover a role for the receptor in immunological tolerance. J Exp Med. 2022. 219. [CrossRef]
- Ishikawa M, Hasanali ZS, Zhao Y, Das A, Lavaert M, Roman CJ, Londregan J, Allman D, Bhandoola A. Bone marrow plasma cells require P2RX4 to sense extracellular ATP. Nature. 2024. 626: p. 1102-1107. [CrossRef]
- Sadovnick AD, Gu BJ, Traboulsee AL, Bernales CQ, Encarnacion M, Yee IM, Criscuoli MG, Huang X, Ou A, Milligan CJ, Petrou S, Wiley JS, Vilarino-Guell C. Purinergic receptors P2RX4 and P2RX7 in familial multiple sclerosis. Hum Mutat. 2017. 38: p. 736-744.
- Zabala A, Vazquez-Villoldo N, Rissiek B, Gejo J, Martin A, Palomino A, Perez-Samartin A, Pulagam KR, Lukowiak M, Capetillo-Zarate E, Llop J, Magnus T, Koch-Nolte F, Rassendren F, Matute C, Domercq M. P2X4 receptor controls microglia activation and favors remyelination in autoimmune encephalitis. EMBO Mol Med. 2018. 10. [CrossRef]
- Qin C, Zhou J, Gao Y, Lai W, Yang C, Cai Y, Chen S, Du C. Critical Role of P2Y12 Receptor in Regulation of Th17 Differentiation and Experimental Autoimmune Encephalomyelitis Pathogenesis. J Immunol. 2017. 199: p. 72-81. [CrossRef]
- Zhang J, Li Z, Hu X, Su Q, He C, Liu J, Ren H, Qian M, Liu J, Cui S, Jiang W. Knockout of P2Y12 aggravates experimental autoimmune encephalomyelitis in mice via increasing of IL-23 production and Th17 cell differentiation by dendritic cells. Brain Behav Immun. 2017. 62: p. 245-255. [CrossRef]
- Van Olst L, Rodriguez-Mogeda C, Picon C, Kiljan S, James RE, Kamermans A, Van Der Pol SMA, Knoop L, Michailidou I, Drost E, Franssen M, Schenk GJ, Geurts JJG, Amor S, Mazarakis ND, Van Horssen J, De Vries HE, Reynolds R, Witte ME. Meningeal inflammation in multiple sclerosis induces phenotypic changes in cortical microglia that differentially associate with neurodegeneration. Acta Neuropathol. 2021. 141: p. 881-899. [CrossRef]
- Saber S, Youssef ME, Sharaf H, Amin NA, El-Shedody R, Aboutouk FH, El-Galeel YA, El-Hefnawy A, Shabaka D, Khalifa A, Saleh RA, Osama D, El-Zoghby G, Gobba NA. BBG enhances OLT1177-induced NLRP3 inflammasome inactivation by targeting P2X7R/NLRP3 and MyD88/NF-kappaB signaling in DSS-induced colitis in rats. Life Sci. 2021. 270: p. 119123. [CrossRef]
- Neves AR, Castelo-Branco MT, Figliuolo VR, Bernardazzi C, Buongusto F, Yoshimoto A, Nanini HF, Coutinho CM, Carneiro AJ, Coutinho-Silva R, De Souza HS. Overexpression of ATP-activated P2X7 receptors in the intestinal mucosa is implicated in the pathogenesis of Crohn's disease. Inflamm Bowel Dis. 2014. 20: p. 444-57. [CrossRef]
- Kurashima Y, Amiya T, Nochi T, Fujisawa K, Haraguchi T, Iba H, Tsutsui H, Sato S, Nakajima S, Iijima H, Kubo M, Kunisawa J, Kiyono H. Extracellular ATP mediates mast cell-dependent intestinal inflammation through P2X7 purinoceptors. Nat Commun. 2012. 3: p. 1034. [CrossRef]
- Figliuolo VR, Savio LEB, Safya H, Nanini H, Bernardazzi C, Abalo A, De Souza HSP, Kanellopoulos J, Bobe P, Coutinho C, Coutinho-Silva R. P2X7 receptor promotes intestinal inflammation in chemically induced colitis and triggers death of mucosal regulatory T cells. Biochim Biophys Acta Mol Basis Dis. 2017. 1863: p. 1183-1194. [CrossRef]
- Grbic DM, Degagne E, Langlois C, Dupuis AA, Gendron FP. Intestinal inflammation increases the expression of the P2Y6 receptor on epithelial cells and the release of CXC chemokine ligand 8 by UDP. J Immunol. 2008. 180: p. 2659-68. [CrossRef]
- Placet M, Molle CM, Arguin G, Geha S, Gendron FP. The expression of P2Y6 receptor promotes the quality of mucus in colitic mice. FEBS J. 2021. 288: p. 5459-5473. [CrossRef]
- Salem M, El Azreq MA, Pelletier J, Robaye B, Aoudjit F, Sévigny J. Exacerbated intestinal inflammation in P2Y6 deficient mice is associated with Th17 activation. Biochim Biophys Acta Mol Basis Dis. 2019. 1865: p. 2595-2605. [CrossRef]
- Vuerich M, Mukherjee S, Robson SC, Longhi MS. Control of Gut Inflammation by Modulation of Purinergic Signaling. Front Immunol. 2020. 11: p. 1882. [CrossRef]
- Grubisic V, Perez-Medina AL, Fried DE, Sévigny J, Robson SC, Galligan JJ, Gulbransen BD. NTPDase1 and -2 are expressed by distinct cellular compartments in the mouse colon and differentially impact colonic physiology and function after DSS colitis. Am J Physiol Gastrointest Liver Physiol. 2019. 317: p. G314-G332. [CrossRef]
- Harshe RP, Xie A, Vuerich M, Frank LA, Gromova B, Zhang H, Robles RJ, Mukherjee S, Csizmadia E, Kokkotou E, Cheifetz AS, Moss AC, Kota SK, Robson SC, Longhi MS. Endogenous antisense RNA curbs CD39 expression in Crohn's disease. Nat Commun. 2020. 11: p. 5894. [CrossRef]
- Goettel JA, Gandhi R, Kenison JE, Yeste A, Murugaiyan G, Sambanthamoorthy S, Griffith AE, Patel B, Shouval DS, Weiner HL, Snapper SB, Quintana FJ. AHR Activation Is Protective against Colitis Driven by T Cells in Humanized Mice. Cell Rep. 2016. 17: p. 1318-1329. [CrossRef]
- Longhi MS, Vuerich M, Kalbasi A, Kenison JE, Yeste A, Csizmadia E, Vaughn B, Feldbrugge L, Mitsuhashi S, Wegiel B, Otterbein L, Moss A, Quintana FJ, Robson SC. Bilirubin suppresses Th17 immunity in colitis by upregulating CD39. JCI Insight. 2017. 2. [CrossRef]
- Weinhage T, Dabritz J, Brockhausen A, Wirth T, Bruckner M, Belz M, Foell D, Varga G. Granulocyte Macrophage Colony-Stimulating Factor-Activated CD39+/CD73+ Murine Monocytes Modulate Intestinal Inflammation via Induction of Regulatory T Cells. Cell Mol Gastroenterol Hepatol. 2015. 1: p. 433-449 e1. [CrossRef]
- Heneghan MA, Yeoman AD, Verma S, Smith AD, Longhi MS. Autoimmune hepatitis. Lancet. 2013. 382: p. 1433-44.
- Vuerich M, Robson SC, Longhi MS. Ectonucleotidases in Intestinal and Hepatic Inflammation. Front Immunol. 2019. 10: p. 507. [CrossRef]
- Liu Z, Sun M, Liu W, Feng F, Li X, Jin C, Zhang Y, Wang J. Deficiency of purinergic P2X4 receptor alleviates experimental autoimmune hepatitis in mice. Biochem Pharmacol. 2024. 221: p. 116033. [CrossRef]
- Grant CR, Liberal R, Holder BS, Cardone J, Ma Y, Robson SC, Mieli-Vergani G, Vergani D, Longhi MS. Dysfunctional CD39(POS) regulatory T cells and aberrant control of T-helper type 17 cells in autoimmune hepatitis. Hepatology. 2014. 59: p. 1007-15. [CrossRef]

Table 1.
P2 receptors in autoimmune diseases.
| P2 receptor | Function | Autoimmune Disease | References |
|---|---|---|---|
| P2X4 | Pathogenic: Increased IL-17 production by effector/memory CD4+ T cells Activation and differentiation of Th17 cells |
Rheumatoid Arthritis | [34] |
| Pathogenic: Promotion of Th17 activation and increased levels of IL-17 in arthritic joints | Collagen-induced Arthritis mouse model | [34] | |
| Pathogenic: Increased levels of serum of IL-1β, TNF-α, IL-6, and IL-17 | Collagen-induced Arthritis mouse model | [74] | |
| Pathogenic: Increased Proteinuria (kidney disease) and autoantibody titers | NZB/W Mouse model of Systemic Lupus Erythematosus | [89] | |
| Pathogenic: Increased levels of serum inflammatory mediators IL-1β, IL-6, IL-17A, IFN-γ, and TNF-α. | Concanavalin-induced mouse model of Liver Autoimmunity | [110] | |
| Protective: Promotion of microglia remyelination | Myelin oligodendrocyte glycoprotein (MOG35-55)-induced mouse model of EAE | [91] | |
| P2X7 | Pathogenic: Upregulation of pro-inflammatory cytokines (Myd88, NF-κB, IL-6, IL-1β, and TNF-α) production | DSS-induced rat colitis | [95] |
| Pathogenic: Promotion of Th17 differentiation and expression of Th17 polarizing cytokines (IL-1β, TGF-β1, IL-23p19, and IL-6) | Collagen-induced Arthritis mouse model | [22] | |
| P2Y6 | Protective: Reduction of Th1 and Th17 cells in the colon | DSS-induced mouse colitis | [101] |
| Protective: Promotes mucus quality | DSS-induced mouse colitis | [100] | |
| Protective: Inhibition of the production of Th1 and Th17 polarizing cytokines (IL-12 and IL-23) | MOG35-55-induced mouse model of EAE | [49] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



