
Submitted:
14 August 2025
Posted:
18 August 2025
You are already at the latest version
Abstract
Prenatal maternal stress (PNMS) increases the risk for autism, and individuals with autism inconsistently exhibit increased or decreased volumes and functional connectivity of the whole amygdala and the whole hippocampus. Given heterogeneous structures of the amygdala and hippocampus and the heterogeneity of autism symptoms, it is worth examining how their subregions contribute to different autism phenotypes. T1-weighted and resting-state functional MRI data were acquired from 32 young adults of mothers who were pregnant during, or within 3 months of, the 1998 Quebec ice storm. Their broad autism phenotype (BAP) was self-reported, including aloof personality, pragmatic language impairment and rigid personality. This sample has a wide range of scores on the BAP Questionnaire. Volumes of the amygdala nuclei and hippocampal subfields were calculated. Seed-to-voxel analysis was applied to examine functional connectivity of the amygdala nuclei and hippocampal subfields with the rest of the brain, and linear regressions were implemented to examine associations of volume and functional connectivity with the three autism phenotypes. We found that 1) pragmatic language impairment was associated with increased right medial amygdala volume; 2) rigid personality was associated with decreased left hippocampal cornu ammonis (CA)1 volume; 3) pragmatic language impairment was associated with decreased left hippocampal CA1 connectivity with the supplementary motor area, and increased right hippocampal CA4 connectivity with the left putamen; and 4) rigid personality was associated with increased right central amygdala connectivity with the left inferior lateral occipital cortex (LOC); and increased left hippocampal CA3 connectivity with the right superior parietal lobule, increased right hippocampal CA4 connectivity with the left superior LOC, and increased right hippocampal dentate gyrus connectivity with the left superior LOC. In contrast, we found no associations with aloof 3 personality. Our results suggest that, within a sample exposed to PNMS, amygdala and hippocampal structure and function contribute differently to two different autistic-like characteristics, with amygdala volume and hippocampus-motor connectivity explaining variance in communication impairment, and with hippocampal volume, amygdala- and hippocampussensory connectivity sharing the common mechanism in rigid behaviors. Given these links between brain and autistic-like traits, future research should determine the extent to which brain volumes and connectivity mediate associations between PNMS and autistic-like traits in young adulthood.
Keywords:
amygdala
; hippocampus
; autism
; volume
; prenatal stress
; functional connectivity
Introduction
Autism is a neurodevelopmental disorder characterized by social and communication deficits and restricted, repetitive behaviors [1]. Individuals with autism vary widely with respect to the defining phenotypes. The phenotypic variability likely reflects diverse etiological mechanisms, and phenotypically diverse individuals may require different treatment strategies [2]. Therefore, it is important to investigate different autistic phenotypes.
Autism can be viewed as the extreme end of the distribution of autistic-like traits in the population [3], and evidence suggests that similar etiological factors apply across the continuum[4,5]. An extension of the dimensional approach for autism concerns the autism spectrum hypothesis, which proposes a broadening of the spectrum so that not only autistic traits at the clinical level but also those observed in non-clinical populations are included [6]. This extension of the autism trait continuum into the non-clinical population implies the existence of a broad autism phenotype (BAP) that is defined as autistic-like traits that are milder but qualitatively similar to those seen in autism [7]. Compared to the rare prevalence of clinical autism [8], autistic-like traits are more common in the general population [9], making it important to study the BAP which may provide insights into the etiology of autism. The BAP includes three primary phenotypes: aloof personality, pragmatic language impairment and rigid personality. Aloof personality parallels social deficits, and is characterized by a lack of interest in or enjoyment of social interaction. Pragmatic language deficits parallel communication deficits, and are characterized by difficulties in communicating effectively or in holding a fluid, reciprocal conversation. Rigid personality, paralleling restricted, repetitive behaviors, is characterized by little interest in change or difficulty adjusting to change [10].
Despite the complicated etiology, prenatal maternal stress (PNMS) is a potential environmental risk factor of autism [11]. We know for instance that prenatal maternal depression is associated with child autism [12,13,14], and that prenatal maternal anxiety is associated with non-clinical autistic-like traits in children [15]. Apart from prenatal mood, using natural disasters to examine PNMS has several advantages, primarily because natural disasters are independent events that are not confounded by heritable parental personality traits.
In January 1998, a severe ice storm struck the province of Quebec that led to one of Canada’s worst natural disasters. To evaluate the impact of PNMS on offspring development, we launched Project Ice Storm, the world’s first prospective longitudinal natural disaster PNMS cohort, in June 1998. We recruited women who were pregnant during the crisis or became pregnant within 3 months following the ice storm, and assessed their stress experience including objective hardship, subjective distress and cognitive appraisal. The follow-up assessments of these mother-child dyads began from the age of 6 months, and continued approximately every two years, until 19 years.
Natural disaster-related PNMS research can help determine whether associations between PNMS and autistic-like traits are due more to the objective degree of exposure the pregnant woman endures, her level of subjective distress, or her cognitive appraisal of the event. More severe autistic-like traits in the Project Ice Storm children at age 6½ were associated with greater prenatal maternal objective hardship and subjective distress [16]. At age 19, we found that 21.4% of the variance in BAP total score was explained by maternal objective hardship. In the three BAP phenotypes, 6.8% of the variance in aloof personality was explained by maternal objective hardship; 15.1% of the variance in pragmatic language impairment was explained by maternal subjective distress; 20.0% of the variance in rigid personality was explained by maternal objective hardship and 14.3% by maternal cognitive appraisal [17]. Furthermore, it was shown that DNA methylation of genes related to autism – specifically the PI3K/AKT/mTOR pathway –mediated the effects of maternal objective hardship and cognitive appraisal on the total score as well as on aloof personality and pragmatic language impairment across ages 15, 16, and 19 [18].
Apart from PNMS, neuroimaging findings have partly unveiled neural correlates of autism; the amygdala and hippocampus have been promising regions of interest in autism research [19,20]. The amygdala plays a central role in emotion processing [21] and social cognition [22]. Cytoarchitectonic mapping of the amygdala in a human post-mortem brain subdivided the amygdala into the basolateral nuclei of the amygdala (BLA) and the centromedial nuclei of the amygdala (CMA) [23]. The BLA are composed of the basal nucleus, lateral nucleus, accessary basal nucleus and paralaminar nucleus [24,25], and the CMA are composed of the central nucleus and medial nucleus [23,25]. The basal nucleus and lateral nucleus receive multi-sensory inputs from widespread cortical and subcortical regions, and send major projections to the medial temporal lobe and prefrontal cortex. The central nucleus and medial nucleus send primary projections to the hypothalamus, bed nucleus of the stria terminalis and several nuclei in the midbrain. In agreement with their afferent and efferent connections, the BLA are mainly involved in social cognition and emotion generation [26]; the CMA regulate autonomic, hormonal and behavioral responses [27]
Hippocampal contribution to social cognition [28,29] makes it another candidate in the context of autism. It has been shown that social impairment, such as poor social skills and social withdrawal, often co-occur with hippocampal structural and functional abnormalities [30]. One observed hippocampal involvement is social cognitive mapping that makes predictions and supports adaptive decision-making. Specifically, maps of social relationships help one predict the actions of others and respond appropriately, just as maps of physical locations help one navigate flexibly within a changing environment [31]. Meanwhile, the traditional hippocampal role in memory and learning [32] cannot be neglected. While memory dysfunction is not the most prominent characteristic of autism, atypical memory and learning in autistic individuals have been reported; autistic individuals, for instance, have difficulties recalling episodic memories [33], have diminished false memory and reduced integration of source memory and thinking [34] and have difficulties in reversal learning [35]. Further, it has been reported that rigid memory, inflexible retrieval and less relational learning may induce atypical behaviors (e.g., restricted, repetitive behaviors) [36]. Similar to the amygdala, the hippocampus is a heterogeneous structure divided into multiple subfields with distinct functional roles. The hippocampal cornu ammonis (CA)3, CA4 and dentate gyrus (DG) are involved in learning and memory encoding while hippocampal CA1 is involved in memory retrieval [37].
Numerous structural MRI studies have inconsistently shown atypical volumes of the amygdala and hippocampus in autistic individuals compared to typically-developing controls, including larger volumes of the whole amygdala [38,39,40] and whole hippocampus [38,41,42,43], or smaller volumes of the whole amygdala [44,45] and whole hippocampus [44], or no volumetric differences in the whole amygdala [41,43] and whole hippocampus [46,47,48]. The discrepancy might be attributed to the age at which the assessments occurred. Subsequent studies [43,49] demonstrate that autistic children had larger left and right whole amygdala volumes than typically-developing children while there were no differences in left or right whole amygdala volume in late adolescence and young adulthood. Further, there are conflicting findings on associations between the three features of autism (i.e., social and communication deficits, repetitive behaviors) and whole amygdala and whole hippocampal volumes [38,40,41,42]. Considering heterogeneous structures of the amygdala and hippocampus, further investigation into their subregions may partly explain these inconclusive findings. One recent study [50], for example, reported that increased BLA volume was associated with social deficits, and that increased central nucleus volume was associated with restricted, repetitive behaviors. In terms of hippocampal subfields, abnormal CA1, CA3, CA4 and DG volumes have been seen in individuals with autism [51], while their associations with the three phenotypes of autism have yet to be examined.
In comparison, less is known about how varying degrees of autistic-like traits correspond to brain structural alterations in non-clinical samples. One study [52] reported prediction of subclinical autistic traits by large whole amygdala volume and small whole hippocampus volume during adolescence. Another study [53] showed positive correlations between autistic-like traits and the volume of whole amygdala-superior temporal sulcus white matter connectivity in young adults. These findings indicate that neural correlates of autistic traits also seem to lie on a continuum in the non-clinical population. However, there is scant knowledge about how structural changes in amygdala and hippocampal subfields contribute differently to explain variance in autistic-like traits, specific to different phenotypes. Given the fact that autism and autistic-like traits are etiologically linked, one study [50] showing positive associations between BLA volume and social deficits, and between central nucleus volume and repetitive behaviors, provides hypotheses for these two domains of subthreshold autistic characteristics.
In addition, there is emerging evidence from resting-state functional MRI research that individuals with autism, compared to typically-developing controls, showed lower BLA functional connectivity with the superior parietal lobe [25], the nucleus accumbens [54] and the ventromedial prefrontal cortex [24], and higher CMA functional connectivity with the occipital cortex [54]. Compared to a growing body of research on functional connectivity alterations of amygdala nuclei in individuals with autism, our knowledge about functional connectivity of hippocampal subfields is limited. Further, it remains unclear the extent to which amygdala and hippocampal functional connectivity explain variance in autism phenotypes.
To address these gaps, the present study aimed to determine: 1) associations between amygdala and hippocampal subregion volumes and BAP; and 2) associations between amygdala and hippocampal subregion functional connectivity and BAP. We hypothesized that 1) increased BLA and CMA volumes would be associated with higher BAP total score; increased CA1, CA3, CA4 and DG volumes would be associated with higher BAP total score; regarding the three subdomains of BAP, increased BLA volume would be associated with aloof personality, and increased CMA volume would be associated with rigid personality; and 2) atypical functional connectivity of the BLA and CMA would be associated with higher BAP total score; atypical functional connectivity of the CA1, CA3, CA4 and DG would be associated with higher BAP total score. Regarding the three subdomains of BAP, atypical BLA functional connectivity would be associated with aloof personality, and atypical CMA functional connectivity would be associated with rigid personality. In contrast to the specific hypotheses of the BLA and CMA on the three subdomains of BAP, we were unable to propose prior specific hypotheses on the CA1, CA3, CA4 and DG.
Methods
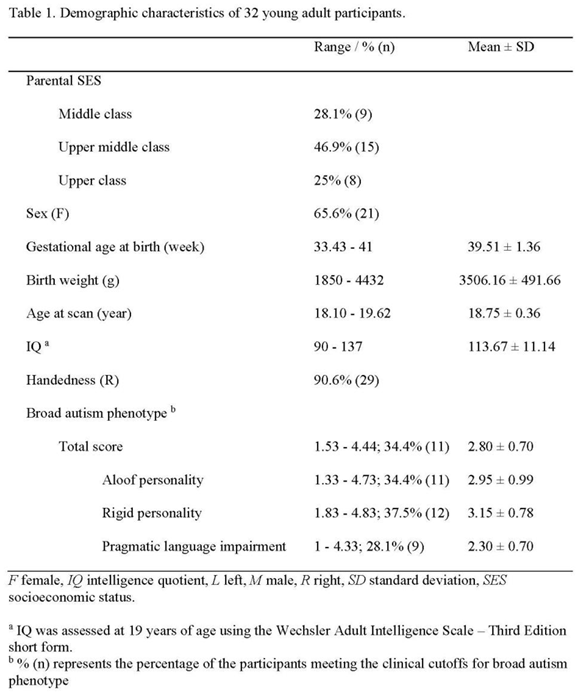
Participants
This study is part of Project Ice Storm (described in Introduction). All participants were born following a major ice storm in Quebec in January 1998. At age 19, 33 young adults self-reported their BAP score. One participant was excluded because of missing MRI data. Thirty-two participants (21 females) were included in all analyses (mean (SD) age = 18.75 (0.36) years; range 18.10-19.62 years). Table 1 presents the demographic and descriptive information. Given previous Project Ice Storm findings on autistic-like traits across different developmental ages (i.e., ages 6 [16], 15, 16 and 19 [17,18]), and given the wide range of BAP trait scores reported in Table 1, this sample is ideal for testing our hypotheses.
Ethical Approval
This study was approved by the Douglas Mental Health University Institute Research Ethics Board. We obtained written informed consent from all participants at all phases of the study.
Broad Autism Phenotype
The Broad Autism Phenotype Questionnaire (BAPQ) [55] was designed to assess BAP in non-clinical populations, and has been validated against direct clinical assessment of BAP [7]. The self-report BAPQ includes 36 questions assessing three subscales of 12 questions each: Aloof Personality, Pragmatic Language and Rigid Personality. Aloof personality is defined as a lack of interest in, or enjoyment of, social interaction (corresponding to social deficits of autism). Pragmatic language impairment is defined as deficits in social aspects of language, resulting in difficulties communicating effectively or in holding a fluid, reciprocal conversation (corresponding to communication deficits of autism). Rigid personality is defined as little interest in change or difficulty adjusting to change (corresponding to restricted, repetitive behaviors of autism) [10]. Questions are rated using a 6-point Likert scale ranging from “very rarely” (1) to “very often” (6) [7], and mean ratings are calculated for each score. Higher scores represent more severe traits. The following scores are used as clinical cutoffs for the total score (3.15), aloof personality (3.25), pragmatic language impairment (2.75) and rigid personality (3.50) [10]. Inter-item reliability for each subscale was examined: Cronbach’s α coefficient was 0.94 for the aloof subscale, 0.91 for the rigid subscale, .85 for the pragmatic language subscale and 0.95 across all 36 items [10]. Inter-item reliability did not differ between self- and informant-report versions. Sensitivity and specificity were at or above 70% for all subscales and over 80% for aloof personality and rigid personality. Sensitivity and specificity were approximately 80% for the total BAPQ score [10].
MRI Data Acquisition
MRI data were acquired using a 3.0T Siemens MAGNETOM Trio TIM Syngo MRI scanner (Siemens, Erlangen, Germany), with a 12-channel head coil. Anatomical images were obtained using a 3D, T1-weighted (T1w) Magnetization Prepared Rapid Gradient Echo sequence (192 slices, TR = 2400s, TE = 2.43ms, slice thickness = 1mm, Flip Angle = 8°, matrix = 256×256). Resting-state functional images were acquired using a T2*-weighted echo-planar imaging sequence (42 slices, TR = 2600ms, TE = 30ms, Flip Angle = 90°, slice thickness = 3.4mm, FoV = 218mm, matrix = 64×64). Throughout the 5:01-minute resting-state scan, participants were instructed to lie still with their eyes open.
MRI Data Preprocessing
fMRIPrep v.1.5.7 [56] was used for preprocessing. The T1w image was corrected for intensity non-uniformity [57], and used as T1w-reference throughout the workflow. The T1w-reference was then skull-stripped. Brain tissue segmentation of gray matter, white matter (WM) and cerebrospinal fluid (CSF) was performed on the brain-extracted T1w [58]. Volume-based spatial normalization to the montreal neurological institute (MNI) space was performed through nonlinear registration, using brain-extracted versions of both T1w reference and the T1w template. For each of the blood-oxygen-level-dependent (BOLD) runs found per subject, first, a reference volume and its skull-stripped version were generated. A deformation field to correct for susceptibility distortions was estimated based on fMRIPrep’s fieldmap-less approach. Based on the estimated susceptibility distortion, a corrected echo-planar imaging reference was calculated for a more accurate co-registration with the anatomical reference. The BOLD reference was then co-registered to the T1w reference [59]. Co-registration was configured with nine degrees of freedom to account for distortions remaining in the BOLD reference. The BOLD time-series were resampled onto their original, native space by applying a single, composite transform to correct for head-motion and susceptibility distortions. The BOLD time-series were then resampled into the MNI space. Automatic removal of motion artifacts using independent component analysis (ICA-AROMA) [60], was performed on the spatially-normalized, preprocessed BOLD on MNI space time-series after removal of non-steady state volumes and spatial smoothing with an isotropic, Gaussian kernel of 6mm FWHM. Finally, we conducted WM and CSF signal removal from the BOLD time series and temporally bandpass filtering (> 0.01 Hz).
FreeSurfer Segmentation
fMRIPrep preprocessed T1-weighted brain regions in MNI space were segmented using FreeSurfer v.7.1.1 [61] and its library tool recon-all. The segmentation of the amygdala and the hippocampus was performed using segmentHA_T1.sh [62,63]. The segmentation of 9 amygdala nuclei and 19 hippocampal subfields is presented in our previous research [64]. Given BLA and CMA have been commonly investigated in neuroimaging research of autism, AFNI 3dcalc was used to combine the BLA and CMA: the BLA are composed of lateral nucleus, basal nucleus, accessory basal nucleus and paralaminar nucleus and the CMA are composed of central nucleus and medial nucleus [23,24,25,65]. Among the 19 hippocampal subfields, only CA1, CA3, CA4 and DG were included in the current study given their common exploration in neuropathology of autism. AFNI 3dcalc was also used to combine the head and body of hippocampal CA1, CA3, CA4 and DG subfields. The BLA, CMA, CA1, CA3, CA4 and DG are shown in Figure 1. The quality of FreeSurfer segmentation was visually inspected using FreeView by two authors (X.L. and M.N.I.Q).
Volume Analysis
Volumes of the amygdala nuclei (i.e., BLA and CMA) and the hippocampus subfields (i.e., CA1, CA3, CA4 and DG) for each participant were computed with FreeSurfer v.7.1.1. All subsequent analyses were conducted using IBM SPSS v.26. Linear regression analyses were conducted to examine associations between the volumes and BAP scores (total score and three subdomain scores). Given the presence of autism or autistic-like traits in a sex-specific manner, sex was controlled for as the covariate in the volume analyses.
Functional Connectivity Analysis
Seed-to-voxel analyses were conducted using the CONN toolbox v.19b [66]. The BLA, CMA, hippocampal CA1, CA3, CA4 and DG (left and right, separately) computed with FreeSurfer v.7.1.1 were used as seeds for the amygdala nuclei and hippocampal subfields for each participant. Pearson correlation coefficients were calculated between average BOLD time series extracted from the above subregion seeds (left and right, separately) and the time courses of all voxels across the brain. The resultant correlation coefficients were converted to normally distributed mean z-values. At the group level, linear regression analyses were conducted with mean z-values as the dependent variable and with BAP scores (i.e., total and three subdomain scores) as the independent variable, and with sex as the covariate. Thresholds for the group-level results were set to p < 0.001 uncorrected for voxel, FDR-corrected p < 0.05 for cluster.
Results
Associations Between Amygdala and Hippocampal Volumes and BAP Total Score
No associations were observed between BAP total score and either BLA volume or CMA volume, nor with hippocampal subfield volumes.
Associations Between Amygdala and Hippocampal Volumes and BAP Subdomain Scores
No associations were observed for BLA volume and BAP subdomain scores. We found a positive association for CMA volume: increased right CMA volume was associated with pragmatic language deficits (Beta = 0.365, B = 7.533, se = 3.536, p = 0.042, Figure 2a). When we further explored subnuclei of the right CMA, we found that increased right CMA volume was driven by the right medial nucleus volume, but not the right central nucleus volume, such that increased right medial nucleus volume was associated with pragmatic language impairment (Beta = 0.385, B = 3.676, se = 1.631, p = 0.032, Figure 2b). In addition, we found that decreased left hippocampal CA1 volume was associated with rigid personality (Beta = -0.406, B = -37.843, se = 15.755, p = 0.023, Figure 2c), but no associations were observed for other hippocampal subfield volumes and BAP subdomain scores.
Associations Between Amygdala and Hippocampal Functional Connectivity and BAP Total Score
No associations were observed between BAP total score and BLA functional connectivity, nor with CMA functional connectivity, nor with hippocampal subfield functional connectivity.
Associations Between Amygdala and Hippocampal Functional Connectivity and BAP Subdomain Scores
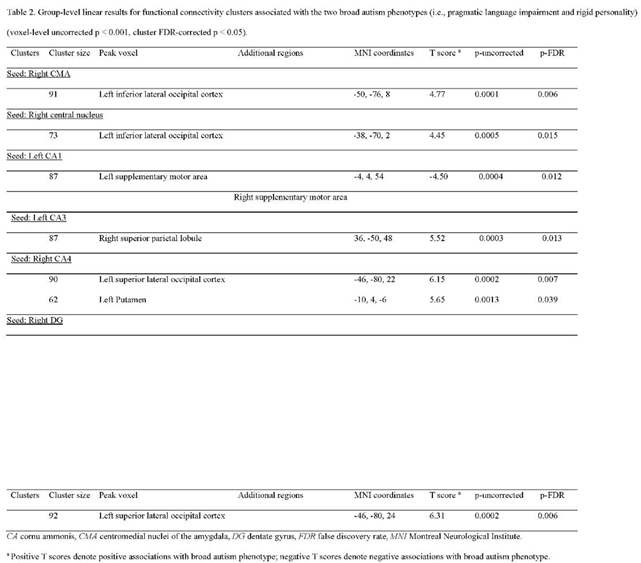
No associations were observed for BLA functional connectivity and BAP subdomain scores. No associations were found for CMA functional connectivity and scores of the two BAP subdomains (i.e., aloof personality and pragmatic language impairment). However, we found that increased functional connectivity from the right CMA to one cluster with its peak voxel located at the left inferior lateral occipital cortex (T score = 4.77, p-FDR = 0.006) was associated with rigid personality (Beta = 0.708, B = 0.109, se = 0.019, Figure 3a). When we further explored subnuclei of the right CMA, we found that increased functional connectivity from the right central nucleus to one cluster with its peak voxel located at the left inferior lateral occipital cortex (T score = 4.45, p-FDR = 0.015) was associated with rigid personality (Beta = 0.705, B = 0.106, se = 0.019, Figure 3b).
No associations were observed for hippocampal subfield functional connectivity and aloof personality scores although we found associations between hippocampal subfield functional connectivity and scores of the two BAP subdomains (i.e., pragmatic language impairment and rigid personality). Specifically, decreased functional connectivity from the left hippocampal CA1 to one cluster involving left and right supplementary motor area (SMA) with its peak voxel located at the left SMA (T score = -4.50, p-FDR = 0.012) was associated with pragmatic language impairment (Beta = -0.715, B = -0.126, se = 0.023, Figure 4a), while increased functional connectivity from the right hippocampal CA4 to one cluster with its peak voxel located at the left putamen (T score = 5.65, p-FDR = 0.039) was associated with pragmatic language impairment (Beta = 0.735, B = 0.126, se = 0.020, Figure 4b). In addition, increased functional connectivity from the right hippocampal CA4 to one cluster with its peak voxel located at the left superior lateral occipital cortex (T score = 6.15, p-FDR = 0.007) was associated with rigid personality (Beta = 0.725, B = 0.131, se = 0.024, Figure 5a); increased functional connectivity from the right hippocampal DG to one cluster with its peak voxel located at the left superior lateral occipital cortex (T score = 6.31, p-FDR = 0.006) was associated with rigid personality (Beta = 0.734, B = 0.124, se = 0.022, Figure 5b); and increased functional connectivity from the left hippocampal CA3 and one cluster with its peak voxel located at the right superior parietal lobule (T score = 5.52, p-FDR = 0.013) was associated with rigid personality (Beta = 0.724, B = 0.109, se = 0.019, Figure 5c). The details for the functional connectivity clusters are presented in Table 2.
Discussion
To our knowledge, this is the first study examining associations between the severity of BAP traits and the volumes and functional connectivity of amygdala nuclei and hippocampal subfields in young adults. All participants in this sample were born following a natural disaster and, while a nonclinical sample, there was considerable variance in the severity of BAP traits which allowed us to uncover correlational associations with amygdala and hippocampal structure and function. Our results demonstrate that it is critical to study the different subdomains of BAP traits separately since we found different patterns in their associations with amygdala and hippocampal volumes and functional connectivity, discussed as below.
First, we found no correlations between the overall BAP severity and the volumes and functional connectivity of the amygdala and hippocampal subregions. The finding that no associations between the overall BAP severity and BLA volume and functional connectivity in our study echoes previous studies showing that no associations were observed between the severity of autism total score and the BLA functional connectivity [25] while the severity of one specific subdomain of autism symptoms (e.g., social deficits) was associated with BLA volume and functional connectivity in autistic individuals [50,54]. Similarly, no associations were observed between the overall BAP severity and volumes and functional connectivity of the CMA, hippocampal CA and DG, which, taken together, emphasizes the necessity to examine the extent to which structure and function of the amygdala and hippocampal subregions are associated with the severity of specific subdomains of autistic-like traits, instead of the overall severity of autism traits.
Regarding the subdomains of BAP traits, no associations were found in our study between the severity of aloof personality and volumes or functional connectivity of the amygdala and hippocampal subregions. In contrast to our hypotheses, no associations were observed between the severity of aloof personality and BLA volume and functional connectivity, which seems inconsistent with the involvement of the BLA in social cognition [26]. A previous study [50] showed that, in 23 adolescents with autism spectrum disorder, increased BLA volume was associated with high reciprocal social interaction scores on the Autism Diagnostic Interview-Revised. The inconsistency between this previous study [50] and our study might be explained by differences in the samples (nonclinical participants with a range of autistic-like traits versus clinically-diagnosed autistic individuals). Nevertheless, both studies have the limitation of small sample size, and thus, future studies with larger sample are warranted to verify the role of the BLA in social cognition. Another possible explanation is that the BLA is heterogeneous complex composed of four different subnuclei (i.e., basal nucleus, lateral nucleus, accessary basal nucleus and paralaminar nucleus) [24,25], and that the basal nucleus and lateral nucleus might have closer involvement in social information processing [26]. Therefore, there is also a need to further explore correlations of the two subnuclei structure and function with social deficits. Less surprisingly, we found that the severity of aloof personality was not associated with the CMA volume and functional connectivity possibly due to its closer association with the regulation of autonomic, hormonal and behavioral responses [27], rather than social information processing, despite reciprocal interconnections between the CMA and BLA [26]. As with the amygdala subregions, we found no associations between the severity of aloof personality and the hippocampal CA and DG volumes and functional connectivity, which might be explained by its primary function of memory [32].
Although we found no associations between the severity of aloof personality and structure and function of the amygdala and hippocampal subregions, there were several associations observed between the severity of the two subdomains of BAP traits (i.e., pragmatic language impairment and rigid personality) and structure and function of the amygdala and hippocampal subregions.
Regarding deficits in pragmatic language, we observed that increased right medial amygdala volume was associated with pragmatic language impairment. Pragmatic language impairment primarily refers to deficits in communication, for example, individuals may be disruptive to a conversation, provide excessive detail during conversations, frequently lose track of the conversation and engage in conversational tangents [67]. The medial amygdala is a primary center for social communication processing; the medial amygdala receives chemosensory input from the main olfactory bulbs projecting to the basal forebrain to elicit appropriate behavioral and physiological responses [68]. Notably, although conventional wisdom suggests that increased volume could afford better processing, we found that larger amygdala volume was associated with more severe pragmatic language impairment. This paradoxical finding has relevance for two competing theories about brain development trajectories in autism; “Early Brain Overgrowth” proposes accelerated brain growth in autistic toddlers followed by abnormally showed growth in older autistic children and adolescents [69], while “Over-Pruning” theory suggests that overly excessive synaptic pruning is predictive of autistic traits, particularly sensory and motor abnormalities [70]. A recent study [71] using two large-sample datasets shows that substantially enlarged brain volume in autistic individuals, compared to typically-developing controls, remains from childhood through young adulthood. These results and ours align more with the overgrowth theory, and somewhat challenge the over-pruning hypothesis. Despite the clues to the origins of volumetric enlargement associated with autism, further exploration is needed to determine how these theories might apply to the non-clinical autistic-like traits described in the current study.
We also found that decreased connectivity from the left hippocampal CA1 to the left SMA and increased connectivity from the right hippocampal CA4 to the left putamen were associated with pragmatic language deficits, possibly reflecting memory-motor-language interface. Although hippocampal implication in pragmatic language impairment may appear unexpected given its classic function in memory, a growing literature reveals that the hippocampus is potentially a key contributor to the processing of language and verbal communication [72,73,74]. One view proposes the interface between memory and language, such that the hippocampal computations that are critical in memory function also meet many of the demands of flexible language use and processing [73,75,76]. For example, the theta-power in the human hippocampal complex involved in memory increases with the establishment of a meaningful context in a sentence [76], and patients with hippocampal amnesia exhibit deficits in the flexible and on-line use of language [77,78]. In support of this view, our finding of the involvement of the CA1 and CA4 subfields in language inflexibility can be explained by human studies showing that the CA1 and CA4 have sensitive roles in the process of verbal memory retrieval [79]. The hippocampus may support the ability to predict upcoming words by drawing on long-term memory representations of individual words, patterns of lexical co-occurrence and syntactic probabilities [75]. Another proposal is that the hippocampal contribution to social cognition [28] may also underlie effective communication: the hippocampus has been found to track social information in the physical environment, make predictions about the action of others and support appropriate responses and decision-making [30,80]. Compared to this relatively novel role for the hippocampus, frequently-reported brain regions closely related to social cognition (referred to as “social brain”) involve other regions such as the medial orbitofrontal cortex and the right insula [81]. Given social impairment as the core symptomatology domain of autism, future investigation is needed to gain a comprehensive picture of the neural circuits underpinning social processing.
Meanwhile, our results showing that the motor-related regions (i.e., the left SMA and the left putamen) are implicated in pragmatic language deficits are supported by their previously reported roles. First, apart from the traditional function in motor control, the SMA plays a critical role in processing of speech communication and language reception; for example, the use of inner speech mechanisms during language encoding, lexical disambiguation, syntax and prosody integration, and context-tracking [77]. In addition, a meta-analytic review [82] has shown that left putamen co-activates with the brain regions directly involved in language processing. Therefore, The CA functional connectivity related to pragmatic language deficits might be attributed to its functions of memory-language interface and social cognition as well as the integration of the SMA and putamen that are involved in language processing.
When it comes to rigid personality paralleling the restricted and repetitive domain of autism, our results extend previous studies by showing that increased connectivity from the right central amygdala to the left inferior lateral occipital cortex was associated with rigid personality, proposing that central amygdala-visual integration might be one potential neural circuit underlying the repetitive behaviors. Specifically, autistic individuals have been reported to exhibit increased functional connectivity from the CMA to the occipital cortex compared to typically-developing individuals [54]. One recent study [50] has shown that increased central amygdala volume is associated with more severe repetitive behaviors. It has been shown that the central amygdala participates in visual stimuli processing [83] and atypical recruitment and regulation of the visual cortex and atypical visual attention are related to restricted and repetitive behaviors [84]. Nevertheless, further investigation is needed to improve our understanding of this neural mechanism associated with the pathogenesis of repetitive behaviors.
In contrast to our hypothesis drawing on a previous finding [50] that increased central nucleus volume is linked to rigid personality, we found that decreased left hippocampal CA1 volume was associated with rigid personality. This discrepancy may be because of the aforementioned limitations of the previous study [50], but an alternative explanation may be that the neural correlates of rigid personality do not fully overlap with those of restricted and repetitive behaviors; restricted and repetitive behaviors comprise different subtypes (including restricted, stereotyped, ritualistic, sameness and self-injurious) [85] while rigid personality relates more specifically to a desire for sameness [7]. In view of hippocampal CA function, it is evident that hippocampal CA1 neurons play an important role in the retrieval of episodic memory and reversal learning [86,87]. A high-resolution MRI study [37] also shows that decreased hippocampal CA1 volume is associated with retrieval deficits.
Apart from volumetric alterations, we found that increased connectivity from the left hippocampal CA3 to the right superior parietal lobule was associated with rigid personality; the involvement of the parietal lobe might be explained by hyper-reactivity to sensory inputs seen in restricted and repetitive behaviors [1]. Another possible explanation is functional interplay between the hippocampus and the parietal lobe; for example, the hippocampal CA3 plays a role in the retrieval of episodic contexts and the parietal lobe also activates during episodic memory retrieval [88,89]. In addition, we found that increased connectivity from the right hippocampal CA4 to the left superior lateral occipital cortex and increased connectivity from the right hippocampal DG to the left superior lateral occipital cortex were associated with rigid personality. Hippocampal DG volume has been associated with visual-spatial memory retrieval [90]. Our result suggests that hippocampal CA4 and DG might share a common neural circuit underlying rigid personality, possibly due to the lack of a clear distinction between the DG and CA4 in terms of their cytoarchitecture [91]. Examples include that certain anatomists also divide the DG into fascia dentata and CA4 [92], and that cytoarchitectonic mapping of the hippocampus in a human post-mortem brain also identified hippocampal formation composed of the CA1, CA2, CA3, DG (including fascia dentata and CA4) [23].
Our results about the correlations of rigid personality with decreased CA1 volume and increased functional connectivity of the CA3, CA4, DG appear to challenge the traditional view that the hippocampus contributes exclusively to memory and learning. Rigidity, defined as difficulty adjusting to new situations or altered routines, reflects a desire for sameness and the presence of inflexibility and over-conscientiousness [7,93,94]. Although it is an unexpected finding, the involvement of the hippocampal subfields in rigid personality might be explained by the links between rigid patterns of behavior observed in autistic individuals and their atypical retrieval and learning [35,36,95]. Specifically, it has been proposed that distortion is a characteristic of human memory and that some memory errors can be adaptive for flexible behaviors; conversely, overly rigid, robust, and undistorted memory may restrict the modification of thinking and result in low relational binding and behavioral inflexibility [96]. There is evidence from human research that less flexible and less relational retrieval and learning may sometimes cause restricted and repetitive behavior, or – vice versa – that inflexible behaviors in autistic individuals may result in rigid and less connected memories [36]. Therefore, the present finding provides a clue that hippocampal CA1, CA3, CA4 and DG are implicated in rigid memory intertwining rigid behavior, suggesting a promising research direction for future studies to advance our understanding of their distinct roles.
Finally, across the results, it was somewhat surprising that a single brain region exhibited associations with different subdomains of BAP depending on its structure and function. For example, the CA1 volume, but not its functional connectivity, was associated with rigid personality while the CA1 functional connectivity, but not its volume, was associated with pragmatic language deficits. This finding indicates that hippocampal resting-state functional connectivity patterns perhaps capture the current state of social language usage better than hippocampal volume. It is possible that the contribution of the hippocampus to pragmatic language requires the functional activation of other regions that are involved in social cognition, whereas the role of the hippocampus in rigid personality may be less dependent on its coactivation with other structures and more narrowly related to hippocampal involvement in atypical memory and learning. Such divergence of the results also emphasizes the necessity to understand the relationship between brain structure and function. One view in favor of brain structure and function being related suggests that the structural properties (e.g., volume, thickness, surface area and curvature) of a region, as well as the white matter tracts, can influence the patterns and strength of its functional activation [97]. Having said that, functional connectivity is also observed between regions in the absence of structural connectivity [98]. Evidence also demonstrates that structure and function display unique features (e.g., associations with functional activity but no associations with volume or white matter tract integrity) that vary with cognition and behavior [99], in support of our results of divergent CA1 structural and functional patterns in the context of rigid personality and pragmatic language deficits.
There are several limitations that should be considered. One methodological limitation is the length of the functional MRI scan, which was 5 minutes in our protocol. Since it has been shown that the reliability of resting-state functional connectivity can increase for durations longer than 6 minutes [100], this scan duration was not ideal. The sample is also small due to loss to follow-up during the 19-year period since the birth of the participants. In addition, the socioeconomic status of the families in this study is higher than the median of the population in the region from which they were recruited in 1998, such that the current results may not generalize to lower class populations. Further, like the prevalence of autism (e.g., 4 males for every 1 female diagnosed) [101], BAP traits tend to aggregate more often in male relatives than female relatives [102,103,104,105,106], however, 65.6% of our sample were females, which may not generalize the existing BAP literature in which a large proportion are males. Finally, the analyses may have been strengthened with the addition of a control group with similar levels of BAP traits but without a prenatal stressor.
Despite the limitations, some strengths are inherent in this study. One strength is that BAP, a form of “broader” autism, was examined within a non-clinical sample that, possibly as a result of prenatal exposure to a natural disaster, was found to have a wide range of autistic-like traits [16]. Use of this sample, which endured a range of severity of prenatal stress, extends our conventional understanding of BAP from previous studies of, for example, first-degree relatives of children with autism [7,55]. A second strength of this study is our examination of the three core broad autism phenotypes separately: aloof personality, pragmatic language deficits and rigid personality [10]. A third strength is the investigation of specific amygdala nuclei (i.e., BLA and CMA) and hippocampal subfields (i.e., CA1, CA3, CA4 and DG), recognizing that the amygdala and hippocampus are composed of heterogenous structures that may be differentially associated with specific broad autism phenotypes. Although the BAP assessment and the functional MRI scanning occurred concurrently at age 19, the neural correlates observed in this non-clinical sample is one potential biomarker candidate that needs to be developed in future research; for example, a subsample of Project Ice Storm children underwent diffusion tensor imaging scanning at age 11, and our future analyses will determine whether amygdala-occipital and hippocampus-motor structural connectivity could function as predictive biomarkers for the two broad autism phenotypes at age 19. Further, since the children at ages 16 and 19 self-reported their BAP phenotypes and also underwent structural and diffusion MRI scanning, we will also examine the longitudinal changes in relationships between three BAP subdomains and amygdala and hippocampal volume and structural connectivity from age 16 to age 19.
In conclusion, our results suggest that amygdala volume and hippocampus-motor connectivity, which are responsible for emotion, memory and motor processing, possibly contribute to social language usage as seen in young adults. As well, our study suggests that hippocampal volume and amygdala-occipital and hippocampus-occipital-parietal connectivity, which are responsible for emotion, memory, visual and sensory processing, possibly contribute to rigid personality in young adulthood. Given the correlational nature of our brain imaging and BAP data, which were assessed at the same age, and the absence of any consideration of the role of prenatal stress in our analyses, we caution against any causal conclusions. Future analyses will test any mediating role of brain structure and function in explaining any possible neural mechanisms by which PNMS predicts autistic-like traits in human offspring.
Author Contributions
S.K. is the principal investigator for Project Ice Storm and devised the original protocol for the recruitment of participants and the assessment of PNMS and child development across time. X.L. contributed to the study design, all analyses and the draft of the manuscript; M.N.I.Q. contributed to methodology of segmentation functional connectivity analysis and manuscript revision. F.L. contributed to MRI data preprocessing. D.P.L. contributed to the acquisition of BAP and MRI data and manuscript revision; G.E. contributed to all statistical analyses; S.L.J. contributed to rs-fMRI data acquisition, and manuscript revision; S.K., and P.R.N contributed to the supervision of X.L. and to the final revision. All authors have approved the final manuscript.
Acknowledgements
We thank Sulantha Mathotaarachchi and Vladimir S. FONOV for programming support. Project Ice Storm was funded by grants to S.K. by the Canadian Institutes of Health Research (MOP-57849; MOP-111177; MOP125892). X.L. was supported by the China Scholarship Council (201906170056).
Conflicts of Interest
Authors declare no competing interests.
References
- Association, A. P. Diagnostic and statistical manual of mental disorders. DSM-5, 5th ed 21 (2013).
- Uljarević, M. et al. Exploring Social Subtypes in Autism Spectrum Disorder: A Preliminary Study. Autism research : official journal of the International Society for Autism Research 13, 1335-1342 (2020). [CrossRef]
- Constantino, J. N. & Todd, R. D. Autistic traits in the general population: a twin study. Archives of general psychiatry 60, 524-530 (2003). [CrossRef]
- Robinson, E. B. et al. Evidence that autistic traits show the same etiology in the general population and at the quantitative extremes (5%, 2.5%, and 1%). Archives of general psychiatry 68, 1113-1121 (2011). [CrossRef]
- Lundström, S. et al. Autism spectrum disorders and autistic like traits: similar etiology in the extreme end and the normal variation. Archives of general psychiatry 69, 46-52 (2012). [CrossRef]
- De Groot, K. & Van Strien, J. W. Evidence for a broad autism phenotype. Advances in Neurodevelopmental Disorders 1, 129-140 (2017).
- Sasson, N. J. et al. The broad autism phenotype questionnaire: prevalence and diagnostic classification. Autism research : official journal of the International Society for Autism Research 6, 134-143 (2013). [CrossRef]
- Elsabbagh, M. et al. Global prevalence of autism and other pervasive developmental disorders. Autism research : official journal of the International Society for Autism Research 5, 160-179 (2012). [CrossRef]
- Ronald, A. & Hoekstra, R. A. Autism spectrum disorders and autistic traits: a decade of new twin studies. Am J Med Genet B Neuropsychiatr Genet 156b, 255-274 (2011). [CrossRef]
- Hurley, R. S., Losh, M., Parlier, M., Reznick, J. S. & Piven, J. The broad autism phenotype questionnaire. Journal of autism and developmental disorders 37, 1679-1690 (2007). [CrossRef]
- Bhandari, R., Paliwal, J. K. & Kuhad, A. Neuropsychopathology of Autism Spectrum Disorder: Complex Interplay of Genetic, Epigenetic, and Environmental Factors. Adv Neurobiol 24, 97-141 (2020). [CrossRef]
- Chen, L. C. et al. Association of parental depression with offspring attention deficit hyperactivity disorder and autism spectrum disorder: A nationwide birth cohort study. Journal of affective disorders 277, 109-114 (2020). [CrossRef]
- Gao, L. et al. Association between Prenatal Environmental Factors and Child Autism: A Case Control Study in Tianjin, China. Biomedical and environmental sciences : BES 28, 642-650 (2015). [CrossRef]
- Say, G. N., Karabekiroğlu, K., Babadağı, Z. & Yüce, M. Maternal stress and perinatal features in autism and attention deficit/hyperactivity disorder. Pediatrics international : official journal of the Japan Pediatric Society 58, 265-269 (2016). [CrossRef]
- Sun, Y. et al. [Pregnancy-related anxiety and subthreshold autism trait in preschool children based a birth cohort study]. Zhonghua yu fang yi xue za zhi [Chinese journal of preventive medicine] 50, 118-122 (2016). [CrossRef]
- Walder, D. J. et al. Prenatal maternal stress predicts autism traits in 6½ year-old children: Project Ice Storm. Psychiatry research 219, 353-360 (2014). [CrossRef]
- Li, X., Laplante, D. P., Elgbeili, G. & King, S. Preconception and prenatal maternal stress are associated with broad autism phenotype in young adults: Project Ice Storm. J Dev Orig Health Dis, 1-9 (2023). [CrossRef]
- Cao-Lei L, E. G., Laplante D, Szyf M, King S. DNA methylation mediates the association between prenatal maternal stress and the broad autism phenotype in adolescence: Project Ice Storm. Biology. Submitted. (2024).
- Cortes Hidalgo, A. P. et al. Observed infant-parent attachment and brain morphology in middle childhood- A population-based study. Dev Cogn Neurosci 40, 100724 (2019). [CrossRef]
- Gotts, S. J. et al. Fractionation of social brain circuits in autism spectrum disorders. Brain : a journal of neurology 135, 2711-2725 (2012). [CrossRef]
- Ochsner, K. N. et al. Bottom-up and top-down processes in emotion generation: common and distinct neural mechanisms. Psychol Sci 20, 1322-1331 (2009). [CrossRef]
- Emery, N. J. et al. The effects of bilateral lesions of the amygdala on dyadic social interactions in rhesus monkeys (Macaca mulatta). Behav Neurosci 115, 515-544 (2001).
- Amunts, K. et al. Cytoarchitectonic mapping of the human amygdala, hippocampal region and entorhinal cortex: intersubject variability and probability maps. Anat Embryol (Berl) 210, 343-352 (2005). [CrossRef]
- Odriozola, P. et al. Atypical frontoamygdala functional connectivity in youth with autism. Developmental cognitive neuroscience 37, 100603 (2019). [CrossRef]
- Rausch, A. et al. Altered functional connectivity of the amygdaloid input nuclei in adolescents and young adults with autism spectrum disorder: a resting state fMRI study. Molecular autism 7, 13 (2016). [CrossRef]
- Sah, P., Faber, E. S., Lopez De Armentia, M. & Power, J. The amygdaloid complex: anatomy and physiology. Physiol Rev 83, 803-834 (2003). [CrossRef]
- Pessoa, L. Emotion and cognition and the amygdala: from "what is it?" to "what's to be done?". Neuropsychologia 48, 3416-3429 (2010). [CrossRef]
- Laurita, A. C. & Nathan Spreng, R. The hippocampus and social cognition. The hippocampus from cells to systems: Structure, connectivity, and functional contributions to memory and flexible cognition, 537-558 (2017).
- Rubin, R. D., Watson, P. D., Duff, M. C. & Cohen, N. J. The role of the hippocampus in flexible cognition and social behavior. Frontiers in human neuroscience 8, 742 (2014). [CrossRef]
- Schafer, M. & Schiller, D. The Hippocampus and Social Impairment in Psychiatric Disorders. Cold Spring Harb Symp Quant Biol 83, 105-118 (2018). [CrossRef]
- Banker, S. M., Gu, X., Schiller, D. & Foss-Feig, J. H. Hippocampal contributions to social and cognitive deficits in autism spectrum disorder. Trends Neurosci 44, 793-807 (2021). [CrossRef]
- Ergorul, C. & Eichenbaum, H. The hippocampus and memory for "what," "where," and "when". Learn Mem 11, 397-405 (2004). [CrossRef]
- Norris, J. E. & Maras, K. Supporting autistic adults' episodic memory recall in interviews: The role of executive functions, theory of mind, and language abilities. Autism : the international journal of research and practice 26, 513-524 (2022). [CrossRef]
- Naito, M., Hotta, C. & Toichi, M. Development of Episodic Memory and Foresight in High-Functioning Preschoolers with ASD. Journal of autism and developmental disorders 50, 529-539 (2020). [CrossRef]
- South, M., Newton, T. & Chamberlain, P. D. Delayed reversal learning and association with repetitive behavior in autism spectrum disorders. Autism research : official journal of the International Society for Autism Research 5, 398-406 (2012). [CrossRef]
- Hashimoto, T., Yokota, S., Matsuzaki, Y. & Kawashima, R. Intrinsic hippocampal functional connectivity underlying rigid memory in children and adolescents with autism spectrum disorder: A case-control study. Autism : the international journal of research and practice 25, 1901-1912 (2021). [CrossRef]
- Mueller, S. G., Chao, L. L., Berman, B. & Weiner, M. W. Evidence for functional specialization of hippocampal subfields detected by MR subfield volumetry on high resolution images at 4 T. Neuroimage 56, 851-857 (2011). [CrossRef]
- Groen, W., Teluij, M., Buitelaar, J. & Tendolkar, I. Amygdala and hippocampus enlargement during adolescence in autism. J Am Acad Child Adolesc Psychiatry 49, 552-560 (2010). [CrossRef]
- Nordahl, C. W. et al. Increased rate of amygdala growth in children aged 2 to 4 years with autism spectrum disorders: a longitudinal study. Archives of general psychiatry 69, 53-61 (2012). [CrossRef]
- Zhu, Z. et al. Alterations in volumes and MRI features of amygdala in Chinese autistic preschoolers associated with social and behavioral deficits. Brain Imaging Behav 12, 1814-1821 (2018). [CrossRef]
- Barnea-Goraly, N. et al. A preliminary longitudinal volumetric MRI study of amygdala and hippocampal volumes in autism. Prog Neuropsychopharmacol Biol Psychiatry 48, 124-128 (2014). [CrossRef]
- Rojas, D. C. et al. Regional gray matter volumetric changes in autism associated with social and repetitive behavior symptoms. BMC Psychiatry 6, 56 (2006). [CrossRef]
- Xu, Q., Zuo, C., Liao, S., Long, Y. & Wang, Y. Abnormal development pattern of the amygdala and hippocampus from childhood to adulthood with autism. J Clin Neurosci 78, 327-332 (2020). [CrossRef]
- Aylward, E. H. et al. MRI volumes of amygdala and hippocampus in non-mentally retarded autistic adolescents and adults. Neurology 53, 2145-2150 (1999). [CrossRef]
- Nacewicz, B. M. et al. Amygdala volume and nonverbal social impairment in adolescent and adult males with autism. Archives of general psychiatry 63, 1417-1428 (2006). [CrossRef]
- Howard, M. A. et al. Convergent neuroanatomical and behavioural evidence of an amygdala hypothesis of autism. Neuroreport 11, 2931-2935 (2000). [CrossRef]
- Haznedar, M. M. et al. Limbic circuitry in patients with autism spectrum disorders studied with positron emission tomography and magnetic resonance imaging. Am J Psychiatry 157, 1994-2001 (2000). [CrossRef]
- Piven, J., Bailey, J., Ranson, B. J. & Arndt, S. No difference in hippocampus volume detected on magnetic resonance imaging in autistic individuals. Journal of autism and developmental disorders 28, 105-110 (1998). [CrossRef]
- Schumann, C. M. et al. The amygdala is enlarged in children but not adolescents with autism; the hippocampus is enlarged at all ages. J Neurosci 24, 6392-6401 (2004). [CrossRef]
- Seguin, D. et al. Amygdala subnuclei development in adolescents with autism spectrum disorder: Association with social communication and repetitive behaviors. Brain Behav 11, e2299 (2021). [CrossRef]
- Li, G. et al. Volumetric Analysis of Amygdala and Hippocampal Subfields for Infants with Autism. Journal of autism and developmental disorders (2022). [CrossRef]
- Nees, F. et al. Global and Regional Structural Differences and Prediction of Autistic Traits during Adolescence. Brain Sci 12 (2022). [CrossRef]
- Iidaka, T., Miyakoshi, M., Harada, T. & Nakai, T. White matter connectivity between superior temporal sulcus and amygdala is associated with autistic trait in healthy humans. Neurosci Lett 510, 154-158 (2012). [CrossRef]
- Kleinhans, N. M. et al. Subregional differences in intrinsic amygdala hyperconnectivity and hypoconnectivity in autism spectrum disorder. Autism research : official journal of the International Society for Autism Research 9, 760-772 (2016). [CrossRef]
- Piven, J. The broad autism phenotype: a complementary strategy for molecular genetic studies of autism. American journal of medical genetics 105, 34-35 (2001).
- Esteban, O. et al. fMRIPrep: a robust preprocessing pipeline for functional MRI. Nature methods 16, 111-116 (2019). [CrossRef]
- Avants, B. B., Epstein, C. L., Grossman, M. & Gee, J. C. Symmetric diffeomorphic image registration with cross-correlation: evaluating automated labeling of elderly and neurodegenerative brain. Medical image analysis 12, 26-41 (2008). [CrossRef]
- Zhang, Y., Brady, M. & Smith, S. Segmentation of brain MR images through a hidden Markov random field model and the expectation-maximization algorithm. IEEE transactions on medical imaging 20, 45-57 (2001). [CrossRef]
- Jenkinson, M. & Smith, S. A global optimisation method for robust affine registration of brain images. Medical image analysis 5, 143-156 (2001). [CrossRef]
- Pruim, R. H. R. et al. ICA-AROMA: A robust ICA-based strategy for removing motion artifacts from fMRI data. Neuroimage 112, 267-277 (2015). [CrossRef]
- Fischl, B. FreeSurfer. NeuroImage 62, 774-781 (2012). [CrossRef]
- Saygin, Z. M. et al. High-resolution magnetic resonance imaging reveals nuclei of the human amygdala: manual segmentation to automatic atlas. NeuroImage 155, 370-382 (2017). [CrossRef]
- Iglesias, J. E. et al. A computational atlas of the hippocampal formation using ex vivo, ultra-high resolution MRI: Application to adaptive segmentation of in vivo MRI. NeuroImage 115, 117-137 (2015). [CrossRef]
- Li, X. et al. Neural correlates of disaster-related prenatal maternal stress in young adults from Project Ice Storm: Focus on amygdala, hippocampus, and prefrontal cortex. Front Hum Neurosci 17, 1094039 (2023). [CrossRef]
- Jalbrzikowski, M. et al. Development of White Matter Microstructure and Intrinsic Functional Connectivity Between the Amygdala and Ventromedial Prefrontal Cortex: Associations With Anxiety and Depression. Biological psychiatry 82, 511-521 (2017). [CrossRef]
- Whitfield-Gabrieli, S. & Nieto-Castanon, A. Conn: a functional connectivity toolbox for correlated and anticorrelated brain networks. Brain connectivity 2, 125-141 (2012). [CrossRef]
- Landa, R. et al. Social language use in parents of autistic individuals. Psychological medicine 22, 245-254 (1992). [CrossRef]
- Biggs, L. M. & Meredith, M. Functional connectivity of intercalated nucleus with medial amygdala: A circuit relevant for chemosignal processing. IBRO Neurosci Rep 12, 170-181 (2022). [CrossRef]
- Courchesne, E. et al. Unusual brain growth patterns in early life in patients with autistic disorder: an MRI study. Neurology 57, 245-254 (2001). [CrossRef]
- Thomas, M. S., Davis, R., Karmiloff-Smith, A., Knowland, V. C. & Charman, T. The over-pruning hypothesis of autism. Dev Sci 19, 284-305 (2016). [CrossRef]
- Yankowitz, L. D. et al. Evidence against the "normalization" prediction of the early brain overgrowth hypothesis of autism. Mol Autism 11, 51 (2020). [CrossRef]
- van de Ven, V., Waldorp, L. & Christoffels, I. Hippocampus plays a role in speech feedback processing. Neuroimage 223, 117319 (2020). [CrossRef]
- Duff, M. C. & Brown-Schmidt, S. The hippocampus and the flexible use and processing of language. Frontiers in human neuroscience 6, 69 (2012). [CrossRef]
- Bonhage, C. E., Mueller, J. L., Friederici, A. D. & Fiebach, C. J. Combined eye tracking and fMRI reveals neural basis of linguistic predictions during sentence comprehension. Cortex 68, 33-47 (2015). [CrossRef]
- Covington, N. V. & Duff, M. C. Expanding the Language Network: Direct Contributions from the Hippocampus. Trends Cogn Sci 20, 869-870 (2016). [CrossRef]
- Piai, V. et al. Direct brain recordings reveal hippocampal rhythm underpinnings of language processing. Proc Natl Acad Sci U S A 113, 11366-11371 (2016). [CrossRef]
- Hertrich, I., Dietrich, S. & Ackermann, H. The role of the supplementary motor area for speech and language processing. Neuroscience and biobehavioral reviews 68, 602-610 (2016). [CrossRef]
- Kurczek, J. & Duff, M. C. Cohesion, coherence, and declarative memory: Discourse patterns in individuals with hippocampal amnesia. Aphasiology 25, 700-712 (2011). [CrossRef]
- Zammit, A. R. et al. Roles of hippocampal subfields in verbal and visual episodic memory. Behav Brain Res 317, 157-162 (2017). [CrossRef]
- Buckner, R. L. The role of the hippocampus in prediction and imagination. Annual review of psychology 61, 27-48, c21-28 (2010). [CrossRef]
- Baas, D. et al. Evidence of altered cortical and amygdala activation during social decision-making in schizophrenia. Neuroimage 40, 719-727 (2008). [CrossRef]
- Viñas-Guasch, N. & Wu, Y. J. The role of the putamen in language: a meta-analytic connectivity modeling study. Brain structure & function 222, 3991-4004 (2017). [CrossRef]
- Farley, S. J., Albazboz, H., De Corte, B. J., Radley, J. J. & Freeman, J. H. Amygdala central nucleus modulation of cerebellar learning with a visual conditioned stimulus. Neurobiol Learn Mem 150, 84-92 (2018). [CrossRef]
- McKinnon, C. J. et al. Restricted and Repetitive Behavior and Brain Functional Connectivity in Infants at Risk for Developing Autism Spectrum Disorder. Biol Psychiatry Cogn Neurosci Neuroimaging 4, 50-61 (2019). [CrossRef]
- Bishop, S. L. et al. Subcategories of restricted and repetitive behaviors in children with autism spectrum disorders. Journal of autism and developmental disorders 43, 1287-1297 (2013). [CrossRef]
- Bartsch, T., Döhring, J., Rohr, A., Jansen, O. & Deuschl, G. CA1 neurons in the human hippocampus are critical for autobiographical memory, mental time travel, and autonoetic consciousness. Proc Natl Acad Sci U S A 108, 17562-17567 (2011). [CrossRef]
- Havekes, R., Nijholt, I. M., Luiten, P. G. & Van der Zee, E. A. Differential involvement of hippocampal calcineurin during learning and reversal learning in a Y-maze task. Learn Mem 13, 753-759 (2006). [CrossRef]
- Wagner, A. D., Shannon, B. J., Kahn, I. & Buckner, R. L. Parietal lobe contributions to episodic memory retrieval. Trends Cogn Sci 9, 445-453 (2005). [CrossRef]
- Dimsdale-Zucker, H. R., Ritchey, M., Ekstrom, A. D., Yonelinas, A. P. & Ranganath, C. CA1 and CA3 differentially support spontaneous retrieval of episodic contexts within human hippocampal subfields. Nat Commun 9, 294 (2018). [CrossRef]
- Travis, S. G. et al. High field structural MRI reveals specific episodic memory correlates in the subfields of the hippocampus. Neuropsychologia 53, 233-245 (2014). [CrossRef]
- Ho Kim, J. et al. Proteome-wide characterization of signalling interactions in the hippocampal CA4/DG subfield of patients with Alzheimer's disease. Sci Rep 5, 11138 (2015). [CrossRef]
- Duvernoy, H. M. The human hippocampus: an atlas of applied anatomy. (JF Bergmann-Verlag, 2013).
- Piven, J. et al. Personality characteristics of the parents of autistic individuals. Psychological medicine 24, 783-795 (1994). [CrossRef]
- Losh, M. et al. Defining genetically meaningful language and personality traits in relatives of individuals with fragile X syndrome and relatives of individuals with autism. Am J Med Genet B Neuropsychiatr Genet 159b, 660-668 (2012). [CrossRef]
- Blundell, J. et al. Neuroligin-1 deletion results in impaired spatial memory and increased repetitive behavior. J Neurosci 30, 2115-2129 (2010). [CrossRef]
- Schacter, D. L., Guerin, S. A. & St Jacques, P. L. Memory distortion: an adaptive perspective. Trends Cogn Sci 15, 467-474 (2011). [CrossRef]
- Litwińczuk, M. C., Muhlert, N., Cloutman, L., Trujillo-Barreto, N. & Woollams, A. Combination of structural and functional connectivity explains unique variation in specific domains of cognitive function. Neuroimage 262, 119531 (2022). [CrossRef]
- Uddin, L. Q. Complex relationships between structural and functional brain connectivity. Trends Cogn Sci 17, 600-602 (2013). [CrossRef]
- Mansour, L. S., Tian, Y., Yeo, B. T. T., Cropley, V. & Zalesky, A. High-resolution connectomic fingerprints: Mapping neural identity and behavior. Neuroimage 229, 117695 (2021). [CrossRef]
- Birn, R. M. et al. The effect of scan length on the reliability of resting-state fMRI connectivity estimates. Neuroimage 83, 550-558 (2013). [CrossRef]
- Nag, H. E., Nordgren, A., Anderlid, B. M. & Nærland, T. Reversed gender ratio of autism spectrum disorder in Smith-Magenis syndrome. Mol Autism 9, 1 (2018). [CrossRef]
- Bolton, P. et al. A case-control family history study of autism. J Child Psychol Psychiatry 35, 877-900 (1994). [CrossRef]
- Piven, J., Palmer, P., Jacobi, D., Childress, D. & Arndt, S. Broader autism phenotype: evidence from a family history study of multiple-incidence autism families. Am J Psychiatry 154, 185-190 (1997). [CrossRef]
- Pickles, A. et al. Variable expression of the autism broader phenotype: findings from extended pedigrees. J Child Psychol Psychiatry 41, 491-502 (2000).
- Schwichtenberg, A. J., Young, G. S., Sigman, M., Hutman, T. & Ozonoff, S. Can family affectedness inform infant sibling outcomes of autism spectrum disorders? J Child Psychol Psychiatry 51, 1021-1030 (2010). [CrossRef]
- Wheelwright, S., Auyeung, B., Allison, C. & Baron-Cohen, S. Defining the broader, medium and narrow autism phenotype among parents using the Autism Spectrum Quotient (AQ). Mol Autism 1, 10 (2010). [CrossRef]
Figure 1.
Segmentation of the hippocampus and the amygdala of one participant. The column from left to right represents coronal, axial and sagittal views, respectively. The amygdala nuclei (BLA and CMA) and hippocampal subfields (CA1, CA3, CA4 and DG) (left and right) were labeled with different colors. BLA, basolateral nuclei of the amygdala; CA, cornu ammonis; CMA, centromedial nuclei of the amygdala; DG, dentate gyrus; L, left; R, right.
Figure 1.
Segmentation of the hippocampus and the amygdala of one participant. The column from left to right represents coronal, axial and sagittal views, respectively. The amygdala nuclei (BLA and CMA) and hippocampal subfields (CA1, CA3, CA4 and DG) (left and right) were labeled with different colors. BLA, basolateral nuclei of the amygdala; CA, cornu ammonis; CMA, centromedial nuclei of the amygdala; DG, dentate gyrus; L, left; R, right.
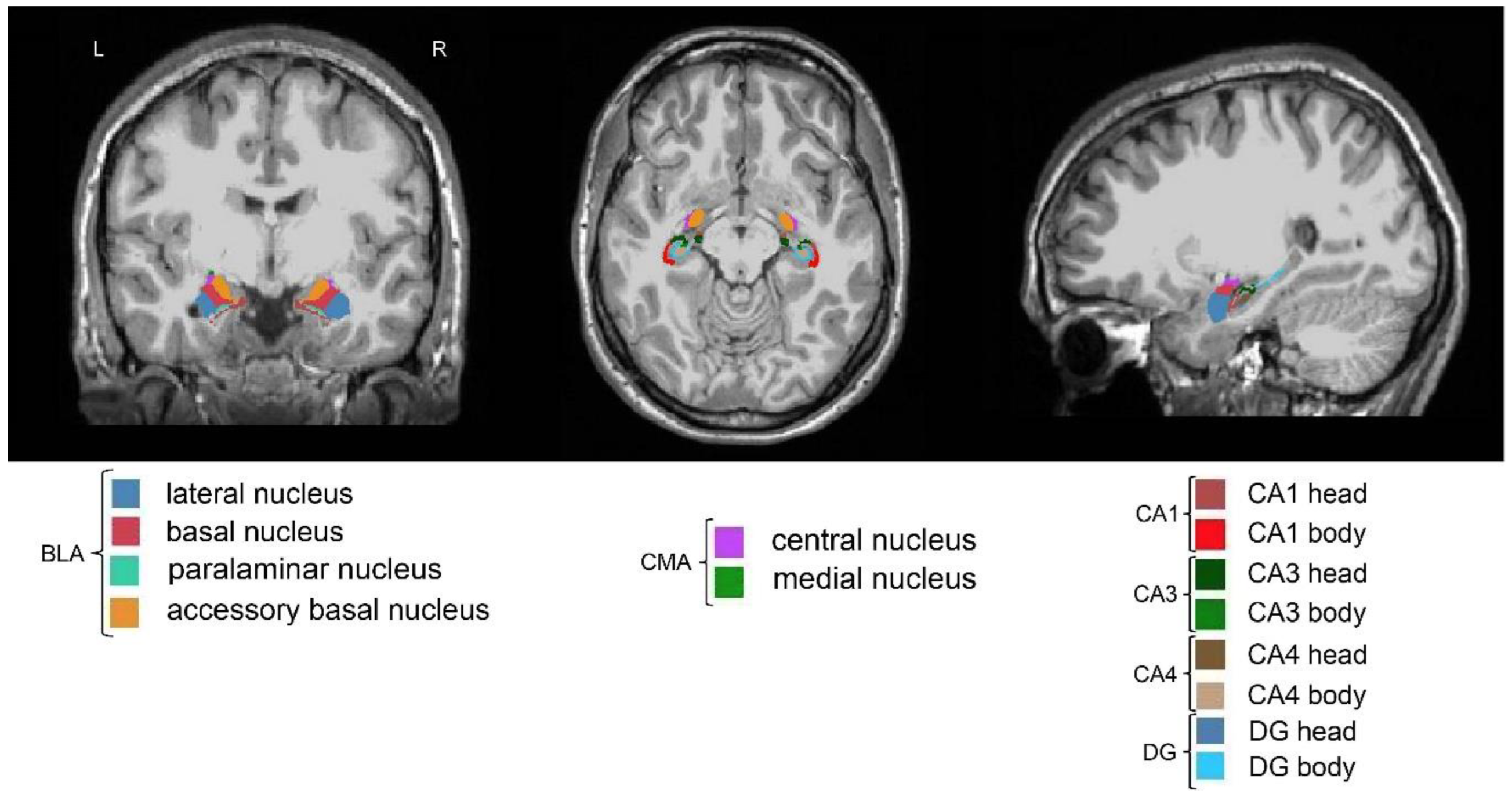
Figure 2.
Correlations of amygdala and hippocampal subregion volumes with two broad autism phenotypes. (a) The left three columns from left to right represents axial, coronal and sagittal views of the right CMA. The right column of the scatterplot represents positive association between the right CMA volume and pragmatic language impairment. (b) The left three columns from left to right represents axial, coronal and sagittal views of the right MeA. The right column of the scatterplot represents positive association between the right MeA volume and pragmatic language impairment. (c) The left three columns from left to right represents axial, coronal and sagittal views of the left CA1. The right column of the scatterplot represents negative association between the left CA1 volume and rigid personality. CA, cornu ammonis; CMA, centromedial nuclei of the amygdala; MeA, medial nucleus of the amygdala; L, left; R, right.
Figure 2.
Correlations of amygdala and hippocampal subregion volumes with two broad autism phenotypes. (a) The left three columns from left to right represents axial, coronal and sagittal views of the right CMA. The right column of the scatterplot represents positive association between the right CMA volume and pragmatic language impairment. (b) The left three columns from left to right represents axial, coronal and sagittal views of the right MeA. The right column of the scatterplot represents positive association between the right MeA volume and pragmatic language impairment. (c) The left three columns from left to right represents axial, coronal and sagittal views of the left CA1. The right column of the scatterplot represents negative association between the left CA1 volume and rigid personality. CA, cornu ammonis; CMA, centromedial nuclei of the amygdala; MeA, medial nucleus of the amygdala; L, left; R, right.
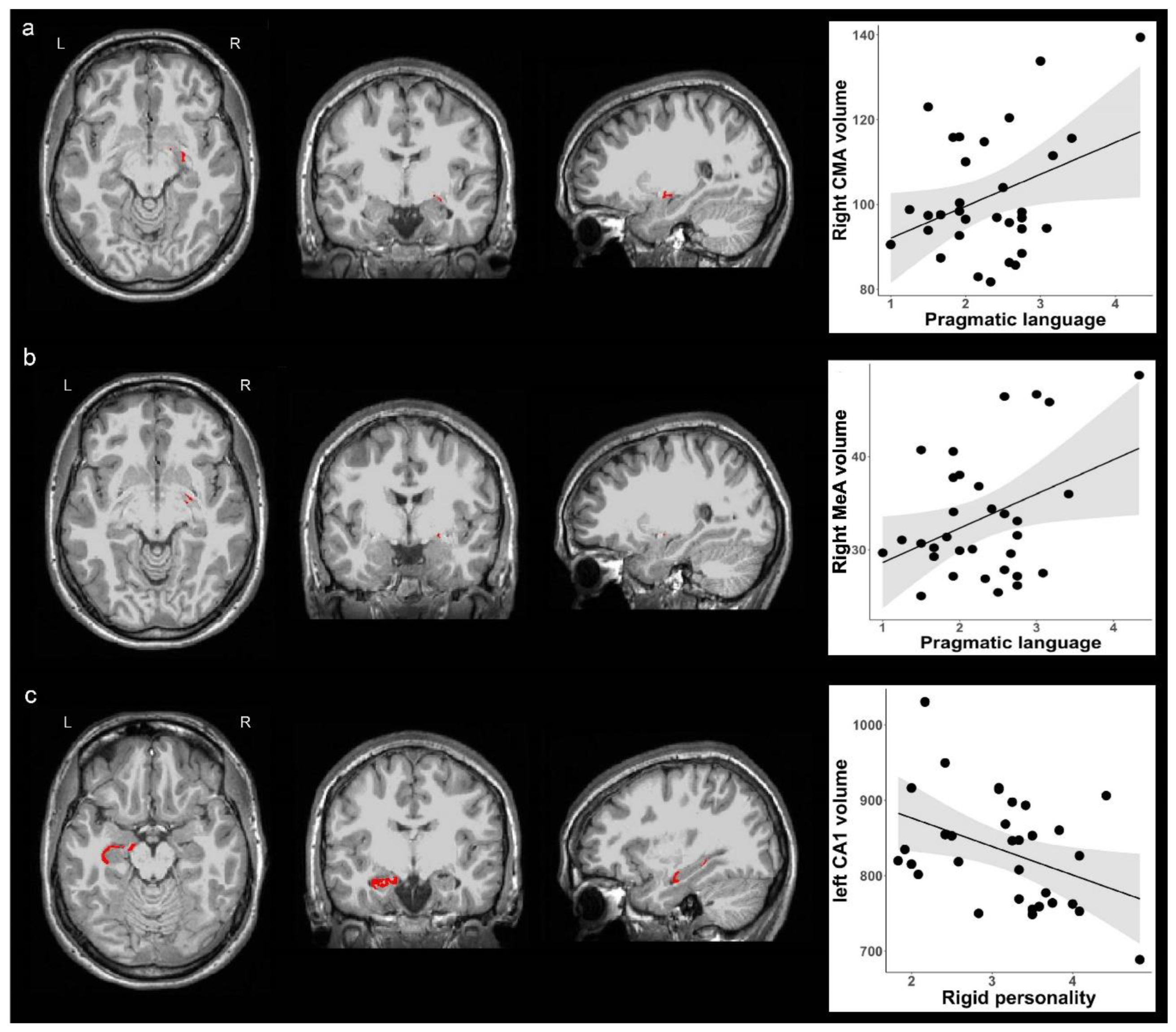
Figure 3.
Correlations of amygdala subregion functional connectivity with rigid personality. (a) The left three columns from left to right represents axial, coronal and sagittal views of the left inferior lateral occipital cortex. The right column of the scatter plot represents positive association between right CMA functional connectivity with the left inferior lateral occipital cortex and rigid personality. (b) The left three columns from left to right represents axial, coronal and sagittal views of the left inferior lateral occipital cortex. The right column of the scatter plot represents positive association between right CeA functional connectivity with the left inferior lateral occipital cortex and rigid personality. Clusters displaying positive associations are depicted in yellow colors. CMA, centromedial nuclei of the amygdala; CeA, central nucleus of the amygdala; iLOC, inferior lateral occipital cortex, L, left; R, right.
Figure 3.
Correlations of amygdala subregion functional connectivity with rigid personality. (a) The left three columns from left to right represents axial, coronal and sagittal views of the left inferior lateral occipital cortex. The right column of the scatter plot represents positive association between right CMA functional connectivity with the left inferior lateral occipital cortex and rigid personality. (b) The left three columns from left to right represents axial, coronal and sagittal views of the left inferior lateral occipital cortex. The right column of the scatter plot represents positive association between right CeA functional connectivity with the left inferior lateral occipital cortex and rigid personality. Clusters displaying positive associations are depicted in yellow colors. CMA, centromedial nuclei of the amygdala; CeA, central nucleus of the amygdala; iLOC, inferior lateral occipital cortex, L, left; R, right.
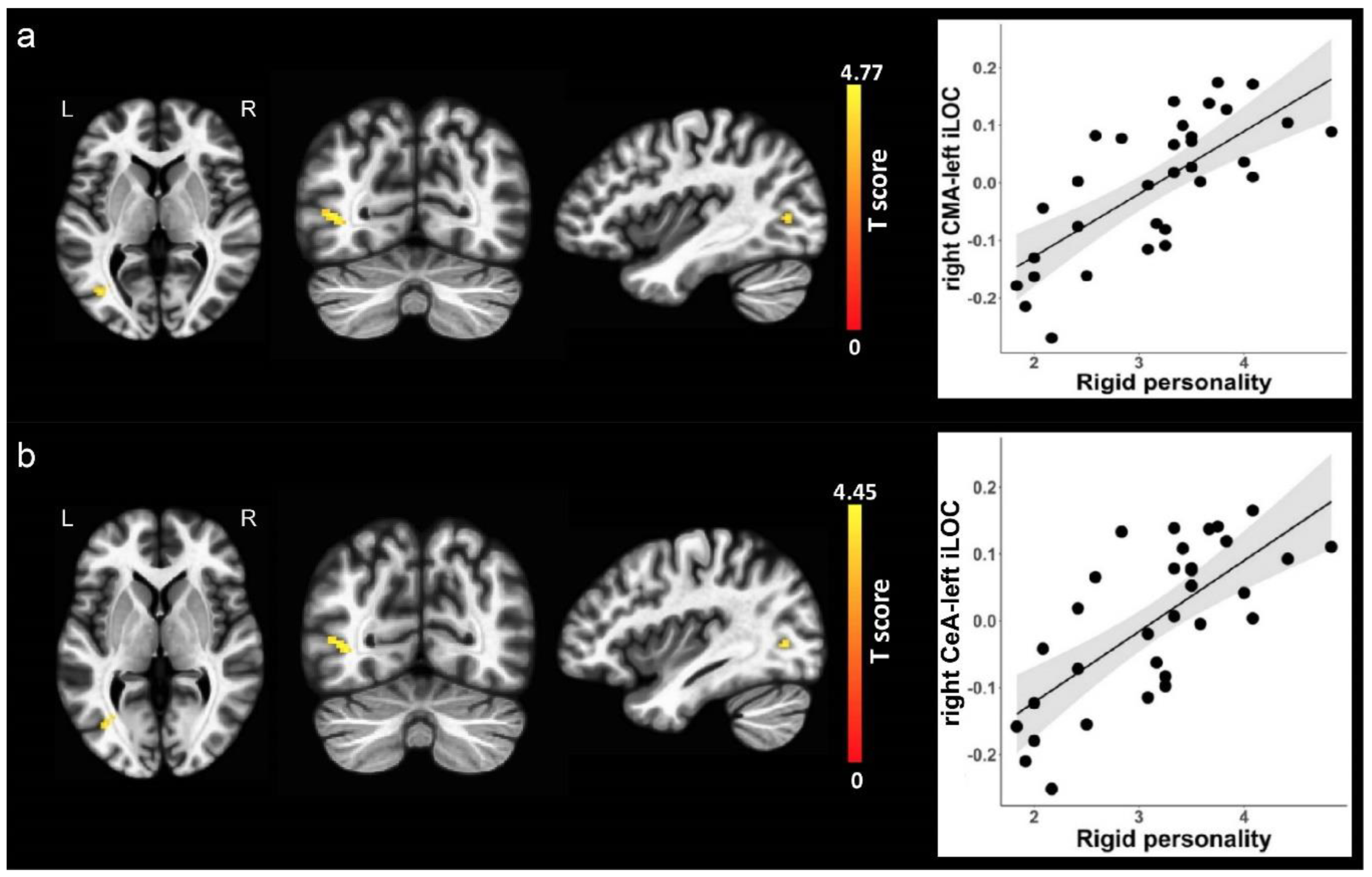
Figure 4.
Correlations of hippocampal subregion functional connectivity with pragmatic language impairment. (a) The left three columns from left to right represents axial, coronal and sagittal views of the left supplementary motor area. The right column of the scatter plot represents negative association between left CA1 functional connectivity with the left SMA and pragmatic language impairment. (b) The left three columns from left to right represents axial, coronal and sagittal views of the left putamen. The right column of the scatter plot represents positive association between right CA4 functional connectivity with the left putamen and pragmatic language impairment. Clusters displaying positive associations are depicted in yellow colors, and clusters displaying negative associations are depicted in purple colors. CA, cornu ammonis; L, left; R, right; SMA, supplementary motor area.
Figure 4.
Correlations of hippocampal subregion functional connectivity with pragmatic language impairment. (a) The left three columns from left to right represents axial, coronal and sagittal views of the left supplementary motor area. The right column of the scatter plot represents negative association between left CA1 functional connectivity with the left SMA and pragmatic language impairment. (b) The left three columns from left to right represents axial, coronal and sagittal views of the left putamen. The right column of the scatter plot represents positive association between right CA4 functional connectivity with the left putamen and pragmatic language impairment. Clusters displaying positive associations are depicted in yellow colors, and clusters displaying negative associations are depicted in purple colors. CA, cornu ammonis; L, left; R, right; SMA, supplementary motor area.

Figure 5.
Correlations of hippocampal subregion functional connectivity with rigid personality. (a) The left three columns from left to right represents axial, coronal and sagittal views of the left superior lateral occipital cortex. The right column of the scatter plot represents positive association between right CA4 functional connectivity with left superior lateral occipital cortex and rigid personality. (b) The left three columns from left to right represents axial, coronal and sagittal views of the left superior lateral occipital cortex. The right column of the scatter plot represents positive association between right DG functional connectivity with the left superior lateral occipital cortex and rigid personality. (c) The left three columns from left to right represents axial, coronal and sagittal views of the right superior parietal lobule. The right column of the scatter plot represents positive association between left CA3 functional connectivity with the right superior parietal lobule and rigid personality. Clusters displaying positive associations are depicted in yellow colors. CA, cornu ammonis; DG, dentate gyrus; L, left; R, right; sLOC, superior lateral occipital cortex; SPL, superior parietal lobule.
Figure 5.
Correlations of hippocampal subregion functional connectivity with rigid personality. (a) The left three columns from left to right represents axial, coronal and sagittal views of the left superior lateral occipital cortex. The right column of the scatter plot represents positive association between right CA4 functional connectivity with left superior lateral occipital cortex and rigid personality. (b) The left three columns from left to right represents axial, coronal and sagittal views of the left superior lateral occipital cortex. The right column of the scatter plot represents positive association between right DG functional connectivity with the left superior lateral occipital cortex and rigid personality. (c) The left three columns from left to right represents axial, coronal and sagittal views of the right superior parietal lobule. The right column of the scatter plot represents positive association between left CA3 functional connectivity with the right superior parietal lobule and rigid personality. Clusters displaying positive associations are depicted in yellow colors. CA, cornu ammonis; DG, dentate gyrus; L, left; R, right; sLOC, superior lateral occipital cortex; SPL, superior parietal lobule.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



