Submitted:
12 August 2025
Posted:
14 August 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
The coronavirus disease of 2019 (COVID-19) has the potential to cause or promote a number of health problems in both the short and long terms. It is often claimed that the COVID-19 messenger RNA (mRNA) vaccinations can reduce the severity of COVID-19 and its sequelae. We counter this claim with evidence and moreover hypothesize that the mRNA vaccinations are triggering mechanisms that result in an amplification of COVID-19 morbidity along with the “post-acute sequelae of COVID-19” or PASC. Since 2021, the coronavirus infections were often superimposed on a preexisting mRNA vaccine-induced milieu of toxic spike protein, inflammatory lipid nanoparticles, and residual process-related DNA impurities. Spike proteins resulting from both the mRNA vaccination and the natural SARS-CoV-2 infection have been shown to persist for extended periods, raising parallel concerns regarding potential implications for long-term safety with this vaccination. Many, if not most, of the morbidity and mortality events attributed to COVID-19 in extensively-vaccinated populations in 2022-2023 were due to the long-term background persistence of spike protein and other vaccine-associated components resulting from previous COVID-19 mRNA vaccinations. Although close temporal associations with laboratory-confirmed SARS-CoV-2 often engendered the assumption of viral causation, the morbidity and mortality events are plausibly attributed to a combination of mass vaccination and coronavirus infections. The overlapping spike-related toxicities and immunological effects of mRNA vaccinations and coronavirus infections have resulted in pronounced immune dysregulation and inflammatory cascades that likely account for near-synchronous waves of COVID-19 and all-cause mortality. We present research suggesting a potential for coronavirus infection-induced amplification of adverse events (AEs) associated with the COVID-19 mRNA vaccinations, including a diverse range of cardiovascular, hematological, autoimmune, and neurological conditions that frequently overlap with PASC. In this paper, we coin the phrase “Hybrid Harms Hypothesis”, which calls for re-examining the unique immunopathological dynamics of coronavirus infection in COVID-19 mRNA-vaccinated individuals and their implications for public health strategies.
Keywords:
modified mRNA products
; SARS-CoV-2 infections
; post-COVID vaccination syndrome
; effect modification
; serious adverse events
; immune dysfunction
1. Introduction
The emergence of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) in early 2020 eventuated in hundreds of millions of confirmed infections and fatalities globally, directly or indirectly linked with the viral infection. Public health officials attributed most of the COVID-19-related mortality in the first few years of the pandemic to uncontrolled viral replication and aberrant host immune responses in vulnerable subgroups, notably the elderly and those with comorbid conditions [1]. The more severe COVID-19 outcomes, including death, were associated with older age and specific comorbidities such as vascular diseases, chronic respiratory conditions, malignancies, obesity, and diabetes [1,2,3,4,5,6,7,8]. The risk was further compounded by pre-existing immunocompromised states and age-related comorbid conditions [9]. Combined exposure to smoking cigarettes and drinking alcohol also increases the risk of severe COVID-19 [10]. Other factors that have led to worse outcomes for COVID-19 patients included overburdened healthcare systems and a carefully coordinated campaign by the vaccine enterprise to block the adoption of effective early treatments [11,12,13,14,15]. An example of the latter was the antiparasitic drug ivermectin, which resulted in an average 31% lower COVID-19 mortality risk, based on a systematic review of the literature.16 Such findings were invariably suppressed because the proven effectiveness of such low-cost treatments would have obviated the need for a vaccine and nullified the Emergency Use Authorization.
Throughout most of 2020, COVID-19 vaccination was vigorously promoted by government agencies and the pharmaceutical industry as the only viable alternative to lockdowns and other mitigation strategies. In the two registrational trials, BNT162b2 (Comirnaty, Pfizer/BioNTech) and mRNA-1273 (Spikevax, Moderna) were reported to significantly reduce the COVID-19 symptoms in a relatively healthy population, and the novel vaccines were granted authorization after only 2-3 months of randomized trial data collection and analyses [17,18]. These were the first-ever gene transfer drugs to receive official authorization for population-wide distribution [19]. The term gene transfer (as well as its more specific application, gene therapy) refers to the ability of these agents to deliver genetic instructions to cells in the form of synthetic, modified mRNA to encode the SARS-CoV-2 spike protein and elicit its endogenous production throughout the body [20,21]. In essence, the products function as prodrugs with the antigenic target, the spike protein, synthesized within the recipient’s cells [22]. This, in turn, is intended to trigger neutralizing antibody production and T cell activation, supposedly bolstering protection against COVID-19 [23,24,25]. Because the outcome is immune stimulation, similar to the intended purpose of a vaccine, the products have also been referred to as genetic vaccines. To be scientifically accurate, however, the modified mRNA injectables should be referred to as gene transfer products. As a compromise, we will also use the terms vaccine and vaccination, as both terms have been more widely used (albeit in outmoded, hackneyed phrasings) in contemporary discussions of these products.
Many scientists still regard COVID-19 as a significant public health threat, as reflected by this opening statement by Rustagi et al. in a December 2024 systematic review on SARS-CoV-2 pathophysiology and post-vaccination severity: “Currently, COVID-19 is still striking after 4 years of prevalence, with millions of cases and thousands of fatalities being recorded every month. The virus can impact other major organ systems, including the gastrointestinal tract (GIT), cardiovascular, central nervous system, renal, and hepatobiliary systems” [26]. The authors further note that the COVID-19 mRNA vaccinations have been associated with serious health risks, including autoimmune nerve damage and increased autoimmune disease overall (likely due to molecular mimicry, as addressed later), arterial or venous thrombosis, cerebral venous sinus thrombosis, vaccine-induced thrombotic thrombocytopenia, glomerular disease, acute ischemic stroke, myocarditis and pericarditis. As of October 2024, approximately 13.58 billion doses had been distributed globally, with a daily average of about 6,959 doses. The sustained global burden of COVID-19 was echoed by other researchers, with approximately 6.8 million “COVID-19-associated deaths” by 2024 [27]. Estimates of COVID-19 mortality have been contentious, partly due to reliance on reverse transcription polymerase chain reaction (RT-PCR) testing to determine COVID-19 incidence and prevalence, rather than direct viral culture [28]. RT-PCR testing, while highly sensitive, may overestimate mortality by detecting viral RNA in asymptomatic or non-causal cases, leading to incidental positive results [29,30]. Conversely, it may underestimate true prevalence due to limitations in testing access, timing, and sensitivity to low viral loads [31].
Thus, when researchers assert that the pandemic continued to cause significant global infections and deaths throughout most of 2024, the following questions may be asked: Are these ongoing “COVID-19 fatalities” due solely to Omicron infections, or could ongoing use of the RT-PCR testing lead to spurious inflation of the estimates? Assuming that some fraction of these fatalities is indeed “COVID-related”, could the mRNA vaccinations function as an effect-modifying factor, contributing in fundamental ways to more severe morbidity (given Omicron’s otherwise mild pathogenicity) and thus to the overall mortality burden? The present paper seeks to answer this complex question.
The emergence of the Omicron variant of SARS-CoV-2 in November 2021 marked a new chapter in the COVID-19 story. Omicron exhibited notable genetic differences from earlier SARS-CoV-2 strains, leading to higher transmissibility and an elevated risk of reinfection among individuals [32]. Nevertheless, Omicron was also characterized as having relatively mild pathogenicity comparable to that of the common cold or a mild flu [33,34,35,36]. Compared to earlier variants, Omicron replicated less effectively in lung and intestinal cells, and its spike protein was not cleaved as efficiently as its predecessor, the Delta variant [37]. Infections caused by Omicron were generally regarded as mild and self-limiting, often presenting with fewer symptoms [38,39,40]. It logically follows that Omicron infections led to underestimated incidence rates, as many asymptomatic or mild reinfections went undetected [41]. Omicron did lead to a sharp increase in COVID-19 cases overall; however, initial data from South Africa did not show a corresponding increase in death rates [42,43,44] The first Omicron wave (BA.1/BA.2) had case-fatality rates half as high as those of the Delta variant, while the second Omicron wave (BA.4/BA.5) had rates three times lower [45] Data from the U.S. Centers for Disease Control (CDC) further reinforced these observations [46] Animal models and human data consistently show Omicron subvariants cause milder disease than Delta, with reduced lung pathology and hospitalization risks [47]. Given Omicron’s significantly lower intrinsic virulence compared to earlier variants, it is surprising to note that later studies paradoxically indicated elevations in both COVID-19 deaths and all-cause mortality associated with Omicron subvariants in 2022 and 2023. We discuss this research in a later section of this paper (see “Epidemiological studies from the Omicron era”).
The official rationale for endorsing the COVID-19 mRNA products was to achieve widespread immunological priming and subsequent herd immunity, thereby attenuating viral transmission and theoretically reducing the incidence of severe disease, hospitalization, and death [48]. In the past four years, many claims have been made regarding the mRNA products’ ability to achieve these outcomes, with sweeping conclusions often based on systematic reviews [49]. In this paper, we challenge the validity of these claims and highlight a crucial oversight that underlies comparisons of the severe morbidity-promoting impacts of the SARS-CoV-2 infection with those of the COVID-19 mRNA vaccination. Although these comparisons are frequently used as the basis for risk-benefit assessments, we believe such comparisons commonly rely on incorrect assumptions, all of which support a false dichotomy. The primary misassumption has been that the spike protein production following the mRNA vaccinations is relatively brief and thus has no impact on subsequent coronavirus infections. We will cite evidence showing that this spike protein production along with associated pathologies or “spikeopathies” may persist for 2-3 years at a minimum. During this timeframe, any exposure to SARS-CoV-2 or Omicron could trigger a disease or disability that would logically be attributed to the infection based on the timing. In fact, however, the persistent spike production from the mRNA vaccinations is likely to predispose individuals to more severe adverse impacts of the coronavirus infections for many months. Such interactions may have been the driving force behind the well-documented elevations in all-cause mortality that occurred in many of the most extensively mRNA-vaccinated countries in 2022-2023 [50,51,52,53,54,55].
This paper offers a new conceptual framework for addressing the potential interplay between COVID-19 mRNA vaccination and SARS-CoV-2 infections. We examine biologically plausible pathways by which either pre- or post-vaccination infections with the Omicron subvariants that have predominated since early 2022 might contribute to ongoing morbidity and mortality despite their relatively low pathogenicity. We also consider the potential epidemiological implications of overlooking this interaction. By assessing immunological interactions, epidemiological patterns, and potential adverse effects of the COVID-19 mRNA injections, the Hybrid Harms Hypothesis helps account for paradoxical increases in severe morbidity and excess mortality that have been associated with Omicron waves and that continue to serve as the primary impetus for promoting the mRNA vaccinations.
2. Waning Immunity and the “Breakthrough Infections” Phenomenon
To what degree the COVID-19 mRNA products are truly protective has been a matter of considerable debate. The discussion has been heavily biased toward the “safe and effective” narrative, with the majority of counternarrative perspectives being censored since early 2021 [56]. The controversy could easily have been predicted based on the fact that the registrational trials by Pfizer and Moderna were terminated after only 2-3 months, an insufficient timeframe for assessing safety and efficacy. The trialists concluded that the mRNA vaccinations have an efficacy of up to 95% [57] for preventing symptomatic SARS-CoV-2 infection 7–14 days after the second dose [58]. Subsequent observational studies showed, however, that the humoral immune protection is short-lived. After a substantial initial surge in IgG and neutralizing antibody levels peaking after the second mRNA dose in the primary series, the neutralizing antibody titers decline rapidly within 2-6 months, with the sharpest declines occurring in the elderly and other more vulnerable populations [59,60]. Post-vaccination antibody kinetics have consistently shown an initial rapid decay followed by a steady state [61].
Protection against symptomatic Omicron infection was observed to drop to about 10-20% by 6 months following the second dose of the COVID-19 mRNA products, with more rapid declines for the elderly and those with comorbid diseases [62]. For example, among those who had received two BNT162b2 doses, vaccine effectiveness against Omicron was 65.5% two to four weeks after the second dose, falling to 15.4% at about four months (15 to 19 weeks), and then further dropping to only 8.8% after six months [63]. Even in clinical trials, these novel products have never been shown to generate sustained immune protection against SARS-CoV-2, or protection that could be considered superior to infection-acquired immunity, also known as natural immunity [64]. Although the COVID-19 mRNA vaccine-induced neutralizing antibodies against SARS-CoV-2 are initially higher, they also decline much faster in mRNA recipients compared to non-vaccinated individuals who were previously infected with the virus [65].
Rapidly waning humoral immunity following the COVID-19 mRNA vaccination is now recognized as a major limitation of these injections. The phenomenon can be attributed to viral evolution and immune imprinting. Continuous mutations in the SARS-CoV-2 spike protein, particularly around the receptor-binding domain, facilitate the emergence of escape variants that evade neutralizing antibodies, reducing the efficacy of subsequent mRNA doses [66]. (In essence, mass vaccination with the mRNA products amidst high Omicron variant transmission exerts selective immune pressure, thereby fostering ongoing immune-escape variants.) When the immune system is repeatedly exposed to earlier SARS-CoV-2 variants, it tends to focus its antibody response on those variants, potentially limiting its effectiveness against newer, dominant strains. This phenomenon, known as immune imprinting and also “original antigenic sin,” leads to the production of cross-reactive antibodies that are less effective against new variants due to prior exposure to earlier strains [67,68]. Importantly, the immune imprinting phenomenon impacts the individual’s response to both SARS-CoV-2 infections and COVID-19 mRNA vaccinations [69]. With regard to the latter, studies demonstrate that SARS-CoV-2-naïve individuals exhibit robust humoral responses to each successive mRNA vaccine dose, whereas previously infected individuals show diminished responses to subsequent doses [70]. Similarly, the reduced effectiveness of bivalent mRNA boosters has been linked to immune systems primed for ancestral strains, highlighting the impact of imprinting on vaccine performance [71].
The early drop in immunologic protection likely accounts for the increased risk of post-mRNA-vaccination infection and reinfection by Delta and Omicron variants, even in individuals classified as “fully vaccinated” [72,73,74] These post-vaccination de novo infections and reinfections have been referred to as breakthrough infections (BTIs), the implication being that the infectious agent has “broken through” the protection theoretically afforded by the mRNA products. Because the fully-immunized individual has become infected with the target pathogen and developed the illness (in this case COVID-19), a more accurate term might be “vaccine failure”. The BTI term applies to individuals who contract COVID-19 at least two weeks post-inoculation, as vaccine-induced immunity is considered to take effect at that point, i.e., any infection taking place within the two-week post-injection period is not considered to be a BTI. Thus, the BTIs then occur within a certain period of presumed post-vaccination protection. In addition to waning immunity, other factors contributing to BTIs include the emergence of immune-evasive variants and individual differences in immune response [75]. For example, immunosuppressed patients (e.g., those undergoing cancer treatment) are more susceptible to BTIs, even after being naturally infected or after multiple vaccine doses [76]. Many biological and exogenous, largely modifiable factors (diet, lifestyle, stress, sunlight exposure) may also influence this vaccine failure phenomenon [77].
It was initially assumed that the COVID-19 vaccinations would provide sterilizing immunity and thus that BTIs would be extremely rare [78,79]. Nevertheless, numerous BTI cases involving either SARS-CoV-2 or the Omicron variants have been documented since 2021 [80,81] A cross-sectional survey (n=543) in Saudi Arabia assessed COVID-19 infection status, vaccination history, and post-vaccination side effects [82]. The findings indicated that 236 (43.5%) participants contracted COVID-19 after full vaccination. Among these cases, infections occurred after the first (46%), second (41.2%), and third (12.8%) vaccine doses. The true extent of BTI prevalence and incidence is unknown, however, mainly due to lack of screening for asymptomatic COVID-19-infected subjects [83]. Given the fact that Omicron infections often have a mild or subclinical presentation, many cases will go undetected and unreported. For instance, asymptomatic Omicron infections comprised 59.8% of test-positive cases in a Canadian study of approximately 59,000 school children [84]. In 2021, vaccinated individuals with mild or asymptomatic infections were less likely to be tested, as testing criteria often prioritized symptomatic or hospitalized cases, skewing reported BTI rates downward [85]. Given these factors, it seems highly probable that BTIs have been vastly underreported since 2022.
More severe BTI cases have also been documented. In a longitudinal study in Tunisia, approximately 11% (10.8%) of the 765 BTIs were considered either severe or critical, and those patients with a medical history of cardiovascular diseases were more than twice as likely to have severe or critical disease [86]. Rates of hospitalized BTI cases ranging from 9 to 35% have been reported [87,88]. In an observational study of 1479 BTI cases, Albtoosh et al. reported a mortality rate of 0.9%, with 92% of the deaths occurring among people older than age 45 (P-value < 0.001) [89]. The infection fatality rate (IFR) for the natural COVID-19 infection typically ranges from 0.03% to 0.05% for a population with a similar age distribution [90,91]. This mortality rate is approximately 22.5 times lower than the BTI mortality rate reported by Albtoosh et al., although the lack of a control group precludes a reliable comparison between BTIs and natural infections in this context.
In terms of susceptibility, several interrelated factors may help account for severe COVID-19 disease following the mRNA vaccinations. Risk factors for severe illness from SARS-CoV-2 infection have included advanced age, weakened immune system, specific health conditions like chronic heart, lung, kidney, liver, or neurological disorders, late-stage pregnancy, and heavy tobacco use [92]. As a result, people with these risk factors often comprise the majority of severe BTI cases. The Tunisia study showing that about one in every ten BTI cases was either severe or critical should be grounds for concern given the normally mild course of COVID-19. Other populations with lower rates of cardiovascular disease and other comorbid diseases would logically tend to have lower rates of severe morbidity associated with BTIs.
Population statistics from the U.K. and United States provide additional support for the severe morbidity and potential lethality of some BTIs. Public Health England reported that 29% of Delta variant deaths in June 2021 occurred in fully vaccinated individuals, rising to 43% (50 of 117) by June 25, with 60% of fatalities having received at least one mRNA vaccine dose [93,94]. In the U.S., by July 2021, Illinois had recorded 151 BTI deaths, representing 2.2% of total COVID-19 deaths.95 Indiana’s state health department reported a COVID-19 death toll of 531 vaccinated patients by October of that year [96]. In Massachusetts, cases doubled from 468 to 942 in three months (0.02% of vaccinated residents), with 290 additional deaths in a single week in January 2022, reaching 1,789 (0.03% of vaccinated population) [97,98]. By February 2022, Massachusetts reported 2,222 BTI deaths since December 2020, underscoring the need for continued surveillance of vaccine effectiveness and factors such as waning immunity and comorbidities [99].
The BTI death counts cited above are likely to be just the tip of the epidemiological iceberg, due to systemic biases in data collection and reporting driven mainly by biopharmaceutical industry-related financial incentives and political pressures. Due to these pressures, public health agencies in the UK and USA sought to promote vaccine efficacy in order to bolster public confidence and compliance with vaccination campaigns, particularly in terms of the childhood vaccine schedule [100]. The officially sanctioned narrative of mRNA vaccination as the sine qua non of COVID-19 control may have led to selective reporting, with agencies downplaying various risks or potential harms to avoid undermining public trust [101]. This narrative also influenced case definitions, testing protocols, and data prioritization, factors that would have led to biases and divergent patterns in reporting [102].
A study analyzing 530 “COVID-19 deaths” in seven Greek hospitals in Athens from January to August 2022 found that only 25.1% were directly caused by COVID-19, with 29.6% involving the coronavirus as a contributing factor, while 45.3% of deaths among patients who tested positive for SARS-CoV-2 were unrelated to the virus [103]. This and other studies indicate that “COVID deaths” have been overreported. By contrast, BTI-related deaths were likely underreported due to the above-mentioned external factors. Inconsistencies in classifying “fully vaccinated” status—such as excluding individuals within 14 days of their second dose or those with waning immunity—may have further contributed to this underreporting of BTI mortality [102,104]. Limited surveillance resources and the deprioritization of post-vaccination case investigations (widely assumed to be lower risk) may have hampered comprehensive data capture [92]. Finally, testing and reporting lagged during the Omicron waves, resulting in an underrepresentation of true mortality data by government health agencies [105]. All of these factors may have contributed to distortions of the true scope of BTI-related morbidity and mortality.
3. Negative Efficacy of the mRNA Products, Adverse Sequelae of the Infections
Along with the rapidly waning humoral immunity and vaccine failures described above, later studies also showed that repeated or successive doses of the modified mRNA products would lead to a further disruption of immune system functioning, resulting in a wide range of serious disease outcomes. Technically known as negative efficacy, this occurs when vaccinated individuals are more likely to contract the targeted disease—in this case, COVID-19—than those who receive either no vaccine or a placebo. This can be quantified as a negative vaccine efficacy percentage, indicating that the vaccinations result in elevated disease risk. In September 2021, the European Medicines Agency warned that frequent COVID-19 booster shots could adversely affect the immune response [106]. A Swedish study in The Lancet subsequently examined the efficacy of COVID-19 vaccinations and how immunity diminishes over time [107]. Eight months after receiving two mRNA injections, vaccinated individuals had lower immune function compared to their non-vaccinated counterparts; this effect was more significant in older adults and people with pre-existing health conditions.
The paradoxical vaccine-induced heightening of susceptibility to SARS-CoV-2 infections and/or to related health conditions was demonstrated in a large-scale retrospective cohort study at the Cleveland Clinic (n=51,017) [108]. In this study, the mRNA vaccinations initially reduced infection risk by 29% during the BA.4/5 wave, but this protection diminished to 20% in the BQ wave and was negligible (4%) by the XBB wave. Meanwhile, however, the risk of infection actually rose with successive doses, with concurrent increases of 107%, 150%, 210%, and 253% for 1, 2, 3, and >3 doses, respectively, compared to unvaccinated individuals. In striking contrast, unvaccinated Cleveland Clinic employees with prior SARS-CoV-2 exposure showed no reinfections over five months, suggesting robust natural immunity. Similarly, a Japanese case-control study reported an elevated infection risk among vaccinated individuals (adjusted OR, 1.85; 95% CI: 1.33–2.57), with a dose-dependent trend: 63% higher risk for 1-2 doses, 104% for 3-4 doses, and 121% for 5-7 doses [109].
Comparable findings from Qatar, the UK, Iceland, and four other US studies highlight increased infection rates post-vaccination, particularly after six months or additional doses [108,110,111,112,113,114,115]. In an Israeli study of 32,000 COVID-19 vaccinees, there was a 27-fold higher risk of contracting symptomatic COVID-19 when compared to non-vaccinated individuals within the same healthcare system.116 Data from the Vaccine Adverse Events Reporting System (VAERS) as of November 2024 show a dose-dependent rise in breakthrough infections, with a 30% increase after the fourth dose compared to 16% after the third.117 These VAERS findings provide further confirmation of declining vaccine efficacy with additional doses. All of these studies collectively point to the failure of the mRNA injections’ ability to protect against symptomatic infection beyond a few months, with multiple doses leading to worsening efficacy.
Beattie conducted an ecological study using Bayesian causal analysis with the R package CausalImpact and OWID COVID-19 data to evaluate the impact of vaccination programs on cumulative cases and deaths per million across 145 countries [118]. By comparing pre- and post-vaccination periods (12–16 months before and after vaccine rollout), Beattie forecasted expected COVID-19 case and death counts without vaccination and compared these to actual figures. The results showed strong statistical significance, with approximately 90% (89.84%) of countries exhibiting increased mortality rates post-vaccination. Correlational analyses suggested a causal link between vaccination programs and elevated total deaths and cases per million compared to projections without vaccines. The Bayesian analysis found that COVID-19 vaccinations significantly increased global COVID-19 cases by up to 291% (95% CI: 2.31-3.56) and COVID-19 deaths by up to 205.25% (95% CI: 1.645-2.57). Excess mortality was also significantly elevated. These increases correlated positively with the number of vaccine doses administered, with stronger correlations for cases than deaths, suggesting that higher vaccination rates were associated with greater adverse outcomes.
To further visualize how this negative efficacy might manifest in populations, consider the timeline comparisons for COVID-19 cases and deaths from the Johns Hopkins Coronavirus Resource Center (CRC; see Figure 1) [119]. Johns Hopkins University hosts two data archives for information gathered by the CRC from January 22, 2020, to March 10, 2023. The first archive includes global case and death statistics, compiled by the Center for Systems Science and Engineering (CSSE). The second contains U.S. and global vaccination records, testing data, and demographic details, managed by the Bloomberg Center for Government Excellence at the university. The CSSE hub allows researchers to access chronological time-series data, including daily data for cases and deaths in the United States and internationally.
Eight countries are represented in the time-series charts shown in Figure 1, with four Nordic countries on the left, and four Asian countries on the right. The syringe icon points to the approximate start of COVID-19 vaccinations; the blue line approximates the date of 75% vaccine uptake by each population. Strikingly, it is only after the population has attained at least 75% coverage that the major surges in COVID-19 incidence and mortality become apparent for all eight countries. (Note: The Johns Hopkins CRC visualizations often use logarithmic and/or normalized scales to compare countries with vastly different population sizes and case counts. For countries with low case numbers, daily or cumulative cases in 2020 might appear flat or near-zero when plotted against countries where cases are in the millions. This scaling can exaggerate the perception of “near-zero” cases).
One possible explanation for these patterns would be that the lockdowns were sufficiently effective to contain COVID-19, and that after the mass vaccination rollout, the lockdowns were lifted and infections escalated. The Hybrid Harms Hypothesis offers an alternative and complementary explanation. Because the waves of cases and deaths are occurring primarily in 2022, when Omicron was dominant and yet of mild pathogenicity, the elevations cannot be reasonably ascribed to Omicron infections alone. It is more biologically plausible, in light of the mechanisms outlined in the next section, that an interaction between the infections and injections is at play, particularly after >75% of the population was fully vaccinated.
The term negative efficacy, based on evidence from a COVID-19 vaccine trial or observational study, typically refers to one of the COVID-19 mRNA products paradoxically increasing COVID-19 disease risk or severity compared to a placebo. In addition, however, negative efficacy can include unexpected adverse outcomes like serious cardiac events or other diseases and disabilities that are unrelated to, or at least not definitively related to COVID-19. Such events could arise from the following: (1) direct toxic spike-related effects of the COVID-19 mRNA products; (2) immune responses and more precisely, immunological dysfunction induction (regulatory T cells or Tregs, critical for averting excessive immune responses), to the mRNA vaccine-induced spike protein; (3) interactions of the mRNA products with the coronavirus infection, both directly and indirectly; (4) specific components of the mRNA products, notably the LNPs and plasmid DNA contaminants; and (5) concurrent or underlying health issues, thus complicating attribution. Our Hybrid Harms Hypothesis primarily focuses on point #3: the interactions between the COVID-19 shots and the infections. To some extent, however, this third point implicates the other four points as well because the AEs are linked with the COVID-19 mRNA.
Numerous studies have called into question the putative safety profile of the COVID-19 mRNA products from the standpoint of serious AEs, which may include deaths, life-threatening diseases and disabilities [120,121,122,123,124,125]. Most of the safety concerns stem from the rushed authorization process and the absence of long-term clinical trial data to assess risks in a reliable way [64,126,127,128]. In a robust re-analysis of the Pfizer trial data, Fraiman et al. found a significant 36% risk (RR, 1.36; 95% CI: 1.02–1.83) of serious AEs in the mRNA-injected group compared to the placebo group [129]. The authors estimated that one out of every 800 doses of the synthetic mRNA products resulted in a serious AE [129]. Historically, vaccines were pulled from the market on the basis of one serious AE per 100,000 injections (i.e., the Fraiman et al. finding exceeded this threshold by 125 times) [130]. A forensic reanalysis of Pfizer’s interim trial data, utilizing previously undisclosed narrative reports from the trial, found a nearly four-fold higher odds (OR, 3.7; 95% CI: 1.02–13.2, p=0.03) of sudden deaths and serious cardiac events among BNT162b2 recipients compared to the unvaccinated group [131]. Confidential Pfizer documents, obtained through legal action by Public Health and Medical Professionals for Transparency after FDA resistance toward disclosure, reveal that about 1.6 million AEs were linked to the mRNA injections by August 2022, with roughly one-third involving serious injuries to cardiovascular, neurological, thrombotic, immunological, and reproductive systems, alongside a notable rise in cancers [132,133,134]. Well-adjudicated autopsy studies (which were widely discouraged during the pandemic, resulting in underreporting of mRNA-vaccination-related deaths) have provided definitive evidence that these mRNA products are causing death, often due to serious cardiac events [135,136]. The above findings have led many to conjecture that the rise in excess mortality in extensively “vaccinated” countries observed in 2021-2022 was more strongly linked with COVID-19 vaccinations than with SARS-CoV-2 infection [52,53,54,55,56].
Many of the serious AEs linked with the mRNA vaccinations have been obscured within public health data due to their relative commonality. Events such as ischemic stroke, brain hemorrhage, myocardial infarction, and acute coronary syndrome, are often observed by healthcare professionals, thus tending to diminish suspicion of vaccination-related causality when recorded in the clinical setting. The familiarity with various AEs due to high background prevalence makes it less likely that clinicians will attribute the more commonly seen conditions to the COVID-19 mRNA products. Cardiovascular diseases and serious cardiac events would be among the most commonly overlooked of the AEs. In the Fraiman et al. study, cardiovascular AEs were more frequent in the Pfizer vaccine group than the placebo group, and coagulation disorders such as pulmonary embolism exhibited the highest excess risk in both trials [129]. In July 2021, the FDA reported detecting four potential AEs of interest following the Pfizer vaccinations, based on medical claims data in older Americans: acute myocardial infarction, pulmonary embolism, immune thrombocytopenia, and disseminated intravascular coagulation [137]. Of the four serious AEs, the first two listed (acute myocardial infarction and pulmonary embolism) are commonly seen in the U.S. elderly population. Consequently, the likelihood of filing individual case safety reports in post-authorization settings is diminished, thus complicating signal detection because of underreporting in public health statistics. Indeed, there is now considerable evidence that underreporting of AEs has likely exaggerated claims of COVID-19 vaccine safety [138]. This concealment within routine disease statistics highlights a fundamental challenge within pharmacovigilance systems: common disorders, despite their severity, may evade being recorded as vaccine-related, thereby skewing risk-benefit assessments and limiting the transparency of mRNA vaccine safety profiles in public health reporting.
It is interesting to note that common chronic diseases such as diabetes, obesity, hypertension and cardiovascular disease are all associated with increased risks of both severe COVID-19 and the serious AEs associated with successive COVID-19 mRNA vaccinations. These diseases are specifically known to predispose individuals to the adverse cardiac events associated with either the coronavirus infection or the mRNA injectable products. The overlapping risks between the infections and post mRNA-vaccination AEs are most likely related to a preexisting chronic inflammatory state, as well as perhaps to a lower threshold for the development of organ dysfunction from the overactive immune response. Diabetes and hypertension are also known to accelerate the aging of the immune system (immunosenescence), which also contributes to the worsening outcomes for both the COVID-19 vaccination and the coronavirus disease itself. The question must be asked: To what extent are the adverse sequelae of the coronavirus infections due to an interaction with the COVID-19 mRNA vaccinations?
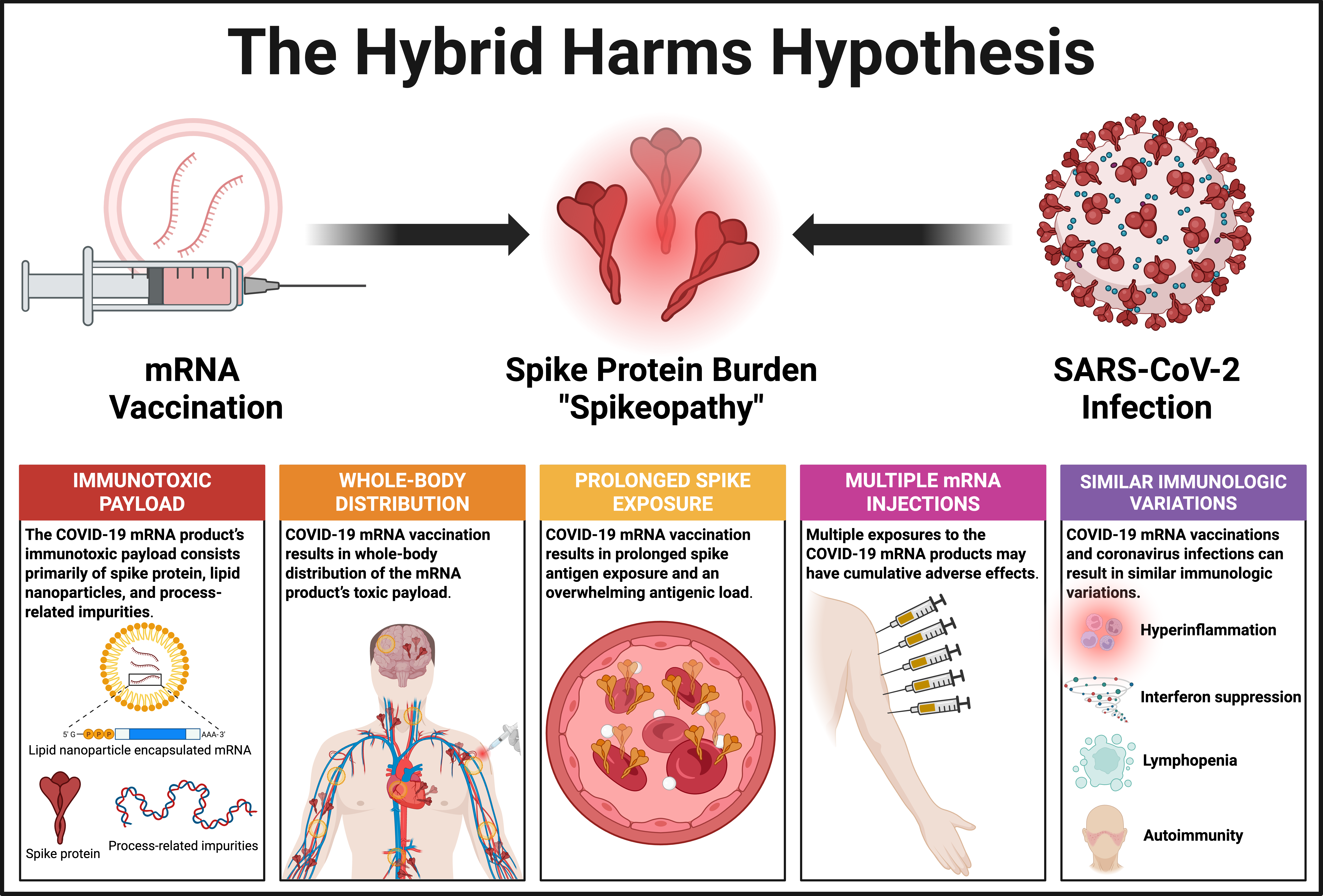
4. The Hybrid Harms Hypothesis
The Hybrid Harms Hypothesis states that repeated spike antigenic exposure via mRNA vaccination may interact with either a previous or subsequent coronavirus infection due to the long-term persistence of spike protein in the body. This interaction results in an amplification of “spikeopathy”, manifesting as chronic immunotoxicity, hyper and persistent inflammation, immune dysregulation, and diverse pathological sequelae, including many disease and disability events that have been associated with both the COVID-19 vaccinations and coronavirus infections. In the case of post-vaccination infections, the apparent temporal association between the diverse sequelae and the SARS-CoV-2 infection has led to systematic misclassification, attributing causality solely to the viral infection rather than considering the potential background noise of spike protein generated by previous mRNA vaccinations.
Medical editorials have often claimed that the “breakthrough” SARS-CoV-2 infections are “rare and mild” in vaccinated people. The official rationale is that the vaccination is priming the body’s immune memory, theoretically enabling more effective control of viral replication. Nevertheless, as we documented in a preceding section, there is substantial evidence that BTIs can result in severe disease and death in COVID-19 mRNA-injected individuals who are considered “fully vaccinated”. The following brief synopses of media reports serve as anecdotal examples of this phenomenon:
Case 1: A robust, physically fit 21-year-old male college student, fully vaccinated against COVID-19, is subsequently diagnosed with the disease. Over a six-week period, he is hospitalized multiple times and dies after extensive medical interventions [139].
Case 2: A 36-year-old male flight attendant, known for meticulous adherence to infection prevention measures, contracts COVID-19 following full mRNA vaccination. His illness requires hospitalization, mechanical ventilation, and ultimately proves fatal [140].
Case 3: A 33-year-old female from Louisiana, fully vaccinated against COVID-19, develops a rapidly progressing illness, leading to her death just four days after symptom onset. Doctors speculate that her extremely rapid demise may be related to her obesity [141].
In each of these cases, COVID-19 was cited as the official cause of death. We hypothesize, however, that COVID-19 was only the final phase in the etiologic chain of events, and that the preceding full course of COVID-19 mRNA vaccinations represented the initial immunotoxic insult that predisposed each individual to more severe morbidity and, ultimately, to premature death.
The Hybrid Harms Hypothesis is distinguished by five fundamental features: (1) the COVID-19 mRNA product’s three-pronged toxic payload; (2) whole-body biodistribution; (3) multiple mRNA injections; (4) prolonged exposure to the mRNA product’s payload; and (5) infection with SARS-CoV-2 or Omicron variants, either sometime before or many months after the vaccination, in which case the disease symptoms may often be misattributed to the viral infection and its “Long Covid” sequelae. We will now consider each one of these and their relevance to our hypothesis. Broader implications of our hypothesis are presented in Table 1.
Table 1. Corollaries to the Hybrid Harms Hypothesis.
The following corollaries help to clarify the scope, predictions, and testable outcomes of our hypothesis:
- Both COVID-19 mRNA vaccinations and coronavirus infections contribute to the total toxic burden of spike protein, either additively or synergistically.
- Direct toxic effects in the context of these “hybrid harms” are focused on disrupting endothelial function and triggering inflammation, potentially contributing to complications like myocarditis or thrombosis.
- Indirect effects occur via the induction of autoimmunity or chronic inflammation, particularly in the context of Long COVID, whereby persistent spike protein and/or immune complexes may drive symptoms and pathogenesis.
- Many cardiac, vascular, hematologic, autoimmune, neurological, and reproductive problems can be triggered by either the modified mRNA inoculations or coronavirus infections, or both. This hypothesis focuses on the third possibility.
- COVID-19 mRNA-inoculated individuals with either prior or subsequent exposure to SARS-CoV-2 appear to face a greater risk of thromboembolism and other vascular pathologies compared to SARS-CoV-2-naïve mRNA vaccine recipients.
- D-dimer elevation may occur, reflecting possible clot formation or immune-driven coagulopathy. Elevated D-dimer levels are commonly observed in hospitalized COVID-19 patients and correlate with thrombotic complications and worse outcomes.
- C-reactive protein (CRP) elevations are also common in pathologies resulting from the interaction between mRNA inoculations and coronavirus infections.
- Antibody testing for anti-spike antibodies provides a scientifically valid, albeit indirect measure of the body’s overall spike protein exposure from previous SARS-CoV-2 infection and/or the COVID-19 mRNA vaccinations.
Hybrid Harms feature #1. The COVID-19 mRNA product’s immunotoxic payload consists primarily of spike protein, lipid nanoparticles, and process-related impurities.
The spike protein of SARS-CoV-2, a critical component of the viral envelope, facilitates cellular entry by binding to host receptors, thereby initiating infection. This protein is also a primary target of COVID-19 mRNA vaccinations, which induce its expression to elicit a transient immune response characterized by anti-spike antibody production. However, much evidence has shown that the spike protein itself exhibits toxic and pathogenic potential, regardless of whether it comes from the COVID-19 mRNA vaccination or from the coronavirus infection. The pathophysiological effects or “spikeopathy” seem to arise from mechanisms resulting in oxidative stress, inflammation, thrombogenesis, endotheliitis-related tissue damage, and prion-related dysregulation [142,143,144]. The spike protein can damage endothelial cells by downregulating ACE2 and consequently inhibiting mitochondrial function [145]. Specific single nucleotide polymorphisms may increase susceptibility to adverse reactions from COVID-19 mRNA vaccines, with these same genetic variants also connected to autoimmune conditions and certain cancers [146].
The second source of potential harm is the lipid nanoparticle (LNP) delivery system, a well-studied component of the COVID-19 mRNA products. The LNPs contribute to prolonged inflammation, as shown by the excessive production of inflammatory cytokines and chemokines [147,148]. The ionizable cationic lipids in LNPs are intrinsically immunotoxic, capable of activating a variety of pro-inflammatory and pro-apoptotic pathways [149]. Prior exposure to mRNA-LNPs in mice resulted in sustained suppression of adaptive immunity, reducing resistance to fungal infections [150]. The ionizable lipids can stimulate the NLRP3 inflammasome and toll-like receptor (TLR) pathways, including TLR4, which has been linked to cancer progression [151,152,153,154]. Pre-existing inflammation in animal models was shown to amplify the inflammatory impacts of the mRNA vaccine’s LNPs [155]. This finding could have special relevance in the context of administering the mRNA injections to individuals with chronic diseases that have an inflammatory component (e.g., obesity, diabetes, heart disease), as well as those with age-related chronic inflammation, or inflammaging.156
Process-related impurities within the COVID-19 mRNA products comprise the third source of potential harm. The production process for the COVID-19 mRNA vaccines introduces billions of plasmid-sourced DNA fragments into each dose [157,158]. Pfizer’s Comirnaty product contains plasmid DNA contaminants exceeding acceptable limits by hundreds of times, sometimes over 500-fold [159]. These DNA impurities (e.g., fragments of plasmid cDNA coding for SV40) are encapsulated within the LNPs, which deliver the contaminants alongside the synthetic mRNA into the recipient’s cells. Of utmost concern is the potential for the DNA fragments to integrate into the human genome through insertional mutagenesis [160]. Such genetic alterations could disrupt immune function, possibly leading to autoimmune conditions by triggering the production of autoantibodies that attack the body’s own cells, causing inflammation, tissue damage, and (potentially) oncogenesis [124,158,161].
A study by the Florida Department of Health indicates a potentially greater long-term risk of the Pfizer mRNA vaccination [162]. When compared to two doses of Moderna’s mRNA-1273, Florida residents who received two doses of Pfizer’s BNT162b2 had a 38% greater risk of all-cause mortality (OR:1.384; 95% CI:1.331-1.439) in the twelve months following the injections, resulting in 229 more deaths per 100,000 people. COVID-19-specific mortality was 88% higher for BNT162b2 than for mRNA-1273 (OR:1.882; 95% CI:1.596-2.220). It should be noted that the pre-vaccination infection rates were nearly identical: 3.54% for Pfizer, 3.40% for Moderna. Elevations in risk were statistically significant for all death categories examined—COVID-19, non-COVID-19, and cardiovascular. The markedly higher DNA contamination levels in the Pfizer products (see preceding paragraph) may help account for these divergent patterns, particularly when contrasted with the three-fold lower mRNA content of Pfizer versus Moderna. The findings may also suggest that BNT162b2’s protective effect against COVID-19 mortality waned more rapidly than that of mRNA-1273, consistent with earlier data from Veterans Affairs studies [163]. Of special interest is the finding that boosting raised most IgG subclass levels, with BNT162b2 vaccinees showing higher IgG2 and IgG4 levels than mRNA-1273 vaccinees, while those previously vaccinated with non-mRNA vaccines had no notable increase in IgG2 or IgG4 after boosting [164]. An IgG4 antibody class switch may increase susceptibility to infectious diseases, IgG4-related autoimmune diseases, and various cancers.
Hybrid Harms feature #2. COVID-19 mRNA vaccination results in whole-body distribution of the mRNA product’s toxic payload.
A substantial body of research indicates that both the mRNA encoding the spike protein and the protein itself can distribute to distal tissues, potentially contributing to systemic adverse effects.165 Early rodent biodistribution studies in Japan revealed that lipid nanoparticles deliver the synthetic, modified mRNA from the COVID-19 injectable products to all organs, crossing the blood-brain and blood-placenta barriers [166]. In many respects, the pharmacokinetics of these genetic vaccines resemble an invasive, blood-borne infectious agent, dispersing the spike protein’s pathogenic effects body-wide. Nonetheless, unlike the preferential affinity of viral infections for certain tissues, the mRNA product’s biohazard arises specifically from whole-body distribution. While the spleen and liver receive the highest concentrations of the synthetic mRNA, the lipid nanoparticle-transported mRNA is distributed to nearly all organs [167]. Notable accumulation occurred in the ovaries, adrenal glands, brain, eyes, heart, testes, uterus, pituitary gland, spinal cord, thymus, and bone marrow [168].
This systemic biodistribution explains why the mRNA vaccine-induced spike protein has been linked to inflammatory organ damage affecting the heart, liver, spleen, ovaries, and nervous system [169,170,171,172,173]. The spike protein can trigger a pro-inflammatory response in brain endothelial cells that may contribute to an altered functioning of the blood-brain barrier [174]. Moreover, mRNA vaccine-induced spike protein can cross that barrier and exert direct neuroinflammatory and neurotoxic effects [175]. In addition, frameshifting events due to mistranslation of the pseudouridinated mRNA can result in the production of aberrant proteins capable of forming toxic prion-like fibrils in neurons, thus potentially causing and/or accelerating neurodegenerative disorders [176,177]. Researchers have documented a startling surge in the reporting of otherwise-rare prion diseases beginning in 2021, with initial symptoms appearing within an average of 11.38 days following the gene-based COVID-19 injections [178].
Hybrid Harms feature #3. COVID-19 mRNA vaccination results in prolonged spike antigen exposure and an overwhelming antigenic load.
The Pfizer and Moderna products’ mRNA and mRNA-generated recombinant spike proteins do not immediately decay following administration of the modified mRNA vaccines and this is by design. Initially it was assumed that COVID-19 mRNA products’ components would be quite transient, due to the relative instability of the mRNA itself. Intracellular enzymatic processes were expected to break it down within days. However, the synthetic mRNA is encapsulated within the LNPs to protect it from immediate degradation, as noted above. Moreover, the persistent production of spike protein is further enabled by the replacement of uridine nitrogen bases with N1-methylpseudouridine, a more stable nitrogen base [179]. This change was made in response to RNA’s legendary instability. The addition of N1-methylpseudouridine appeared to enhance the durability of the LNP-delivered synthetic mRNA, thus ensuring the spike protein’s persistent bioavailability for an unknown period of time [180]. Of critical concern, however, the N1-methylpseudouridine modification in these COVID-19 mRNA products enhances transcriptional infidelity by promoting ribosomal miscoding, leading to elevated rates of amino acid misincorporation and a higher frequency of translation errors [181]. In principle, even minor inaccuracies in transcription can result in incorrect amino acid sequences during protein translation. The proteins synthesized in this context may be dysfunctional and/or excessively immunogenic, potentially causing adverse immune responses (e.g., inflammation or autoimmunity) or other pathological effects (e.g., cellular dysfunction or disease). Such errors also raise grave concerns about the long-term safety of the COVID-19 mRNA injections, along with the potential for catastrophic results when scaled to large populations.
Initial speculations regarding clearance, based on a combination of preclinical studies and mechanistic reviews, were that the spike protein would be found in the blood for approximately 1–2 weeks [182,183]. By 2022, emerging data indicated that spike protein could persist longer, up to 4–8 weeks post-vaccination [69,184,185]. However, spike proteins induced by the BNT162b2 injection were then found to be present on exosomes circulating throughout the body for more than 4 months [186]. When the study timeframes were further lengthened, data analyses showed detectable spike protein in circulation for 6–8 months (187-245 days) in some individuals [187,188,189]. More recent studies have further extended these timeframes. In a Japanese study of hemorrhagic stroke patients, spike protein was found in cerebral arteries up to 17 months following the COVID-19 mRNA vaccination [190]. Circulating spike protein was also elevated in individuals suffering from substantial immune dysfunction following the injections, with detectable post-vaccination levels persisting for 23.6 months (709 days) [191]. Even more concerning is a well-adjudicated case report showing that detectable Pfizer vaccine-generated mRNA was found in the blood 3.2 years after the modified mRNA injection.192
Numerous peer-reviewed studies have reported prolonged persistence of vaccine-derived mRNA and spike protein in tissues of vaccinated humans and animal models, challenging early assertions of rapid clearance by public health authorities. The best estimates to date for extended spike production post-vaccination are in the range of 24 to 38 months. The modified mRNA appears to convert transfected cells into “viral protein production factories” without an inherent mechanism for regulating the spike production. Longer-term studies may confirm that there is no regulatory brake or “off switch” mechanism for halting the generation of this target antigen [124,169]. The primary concern is that this prolonged generation of the spike protein, and the resulting cumulative antigenic load, may lead to ongoing systemic inflammation, immunologic dysfunction, and a host of immune-related disorders [147,150,193,194]. A related issue is prolonged exposure to plasmid DNA process-related impurities such as the double-stranded RNA found in the COVID-19 mRNA products [195]. This aberrant RNA may trigger dose-dependent activation of the innate immune system and subsequent inflammation, resulting in myocarditis and other serious cardiac AEs [196].
We believe that the erroneous assumptions of short-term spike protein expression in early 2021 helped engender the notion of a false dichotomy between the COVID-19 mRNA injections and coronavirus infections. We further posit that the prolonged persistence of the mRNA vaccine-encoded spike protein, potentially extending 2-3 years post-mRNA vaccination, also raises significant questions regarding the impact of any subsequent coronavirus infections. Within this period, coronavirus infections may amplify AEs linked to the ongoing presence of the vaccine-induced spike protein. This putative 2–3-year window of persistent spike protein is henceforth referred to as the “window of vulnerability” or WOV.
Hybrid Harms feature #4. Multiple exposures to the COVID-19 mRNA products may have cumulative adverse effects.
It is logical to surmise that repeated COVID-19 mRNA vaccinations will result in increased and potentially cumulative toxic impacts along with disruption of immune system functioning. This could also be called the “Triple-Hit Hypothesis” because the primary series entails two injections of the mRNA product; any additional exposure to the coronavirus infection would therefore result in a tripling of spike protein exposures, at a minimum. Multiple, successive COVID-19 mRNA vaccinations are likely to perpetuate various forms of immune dysregulation. For example, repeated administration of the modified mRNA injectables appears to increase the risk of infections as well as autoinflammatory phenomena associated with COVID-19 mRNA-induced damage to the heart, brain, and other organs [197,198,199]. Successive doses of these mRNA injections may also increase the likelihood of serious hematologic events such as acute de novo immune thrombocytopenic purpura in previously healthy individuals [200,201].
The combination of ongoing, repeated COVID-19 mRNA boosters, together with prolonged spike protein production over the course of many months (Condition #3), could translate into chronic immune dysfunction in the form of T-cell exhaustion and antibody class-switching to IgG4 [198,202]. T-cell responses against SARS-CoV-2 were significantly reduced one month following the first and second booster doses of the COVID-19 mRNA vaccine [203]. This IgG4 class switch is also linked to the reduced T-cell response after three to four doses of the COVID-19 mRNA vaccine [204,205]. These immunologic disruptions may explain the reduced protection against infections and cancers, along with an increased risk of new-onset autoimmune disease and an accelerated progression of latent autoimmune diseases [205,206]. Such immunologic shifts may also explain the findings from the U.S. and Japanese studies showing elevated rates of COVID-19 in conjunction with increasing number of doses of the COVID-19 mRNA vaccines [108,109,115,207].
Regular, successive doses of the COVID-19 mRNA product may have a more deleterious impact when the coronavirus infection follows vaccination. Kiszel et al. observed that individuals injected with the COVID-19 mRNA products displayed a distinct immune response profile depending in part on the timing of SARS-CoV-2 infection [208]. Alongside the IgG1 and IgG3 subclasses observed across all groups, a shift toward the spike-specific IgG4 and IgG2 subclasses was predominantly seen in those who either received only the mRNA vaccine or who were infected post-mRNA vaccination. In these two groups, the spike-specific IgG4 (immune-suppressive) response reached levels comparable to the spike-specific IgG1 response. These findings suggest that initial immune priming via the mRNA vaccination, if followed by SARS-CoV-2 infection, may significantly alter the humoral immune response, potentially leading to increased production of IgG4 antibodies and a heightened risk of infection, cancer, and autoimmune disease. Indeed, elevated IgG4 levels as a consequence of repeated COVID-19 mRNA vaccinations do appear to be associated with a greater risk of breakthrough infection, i.e., vaccine failure [209].
Hybrid Harms feature #5. COVID-19 mRNA vaccinations and coronavirus infections can result in similar immunologic variations, with strong potential for either additive or synergistic effects.
The COVID-19 mRNA vaccines and natural coronavirus infections both trigger robust immune responses, including the production of pro-inflammatory cytokines and activation of cell-mediated immunity, which can lead to overlapping physiologic changes in the body. These redundant phenomena suggest a strong potential for additive or synergistic effects, whereby the combined impact of vaccination and infection could amplify immune activation, potentially enhancing protection or, in some cases, increasing the risk of adverse effects. Here are the main examples from a pathophysiological perspective:
Hyperinflammation. Cytokines act as signaling molecules that regulate the immune-inflammatory response, both initiating and resolving inflammation. Elevated levels of inflammatory cytokines, such as IL-6 and IL-1β, are connected to COVID-19 mRNA vaccine-related AEs, including the new onset of autoimmune-inflammatory disorders [210,211,212]. Similarly, severe COVID-19 is often characterized by a hyperinflammation involving an excessive release of certain proinflammatory cytokines that can result in tissue damage, notably in the lungs [213]. The question must be asked: How many ICU cases involving severe COVID-19 pneumonia were exacerbated by recent COVID-19 mRNA vaccinations?
Autoimmunity. SARS-CoV-2 infections and mRNA vaccinations may promote autoimmune reactions [214,215]. Severe SARS-CoV-2 infections were linked with new-onset autoimmune conditions such as cutaneous vasculitis, polyarteritis nodosa, and immune-related hepatitis [216]. The modified mRNA inoculations can cause rheumatoid arthritis, lupus erythematosus, and autoimmune hepatitis, among other autoimmune disorders [217,218,219]. The mechanistic explanation may entail a combination of molecular mimicry (between the spike protein and human proteins), epitope spreading, and bystander activation [220]. It seems only logical to speculate that autoimmune diseases would tend to increase due to the overlapping autoimmune effects of the coronavirus infections and mRNA vaccinations.
Lymphopenia. Lymphopenia (or lymphocytopenia) can raise the risk of infections and is nearly universal in severe COVID-19 cases [221]. There is some evidence of a correlation between COVID-19 disease severity and functional exhaustion of T-lymphocytes, most often following T cell hyperactivation in earlier disease stages [222,223]. Similar T-cell reductions have been observed after multiple COVID-19 mRNA vaccinations [224]. The lymphopenia is more pronounced in immunocompromised individuals, e.g., due to recent cancer chemotherapy [225]. Paradoxically, then, the recommendation to administer the mRNA injectables to these “at risk” individuals could have serious repercussions, particularly with repeated doses and subsequent Omicron variant infections.
Interferon suppression. Interferon is a pleiotropic cytokine that regulates the immune response and inflammation. Autoantibodies against type I interferons underlie COVID-19 pneumonia and were found in about 14% of patients with life-threatening COVID-19 versus only 0.3% of healthy controls [226,227]. Single-cell profiling showed interferon signaling was strongly suppressed in COVID-19 compared to influenza, with interferon-stimulated cells entirely absent in severe COVID-19 cases [228]. Intriguingly, COVID-19 mRNA vaccinations have been shown to induce autoantibodies against type I interferons in healthy individuals [229,230]. In both the infection and injection scenarios, the effect would be to suppress antiviral and anticancer immune responses. Some research indicates, for example, that the BNT162b2 injections may alter the individual’s antiviral immune protection in ways that increase their risk of viral myocarditis, resulting in a combination of direct cardiomyocyte damage and immune-mediated cell death [231].
To summarize, COVID-19 mRNA vaccinations and coronavirus infections have been shown to evoke similar immunologic changes, including hyperinflammation, autoimmunity, lymphopenia, and type I interferon suppression. As discussed in a subsequent section, the overlapping immunologic phenomena may potentially result in many of the same disease and disability outcomes that have been ascribed to “Long Covid” but are in fact part of a post-COVID vaccination syndrome (PCVS). The convergence of immunopathological manifestations is alarming, particularly since it suggests the potential for an amplification of serious AEs in mRNA-vaccinated individuals subsequently exposed to the infection. For example, the impaired immunologic protection resulting from suppression of interferon-mediated innate immunity, if amplified by the two exposures, could result in a heightened susceptibility to infections and cancers. Such interactions could also explain the worsening of immune protection seen with successive COVID-19 mRNA inoculations, as well as the increased likelihood the infections will have an “adverse feedback effect” on the vaccinations. Again, the spike protein is the common denominator, the pivotal antigen triggering these diverse immune responses following either SARS-CoV-2 infection or mRNA vaccination. It is biologically plausible that variations in the infection- and vaccine-induced immune responses to spike protein (and other components of the mRNA products, not present in the coronavirus) are influencing severe disease morbidity and mortality in the context of COVID-19 (see Figure 2).
5. Illusions of Protection Against Severe Disease
Since December 2020, a central dogma of the vaccine enterprise has been that the COVID-19 mRNA products reduce severe COVID-19 disease and, moreover, reduce hospitalizations and death. Nevertheless, these claims have been called into question, particularly in light of the increasingly robust evidence of negative efficacy. The mRNA products’ proponents continue to contend that the mRNA vaccinations afford sufficient immunological breadth to protect against severe COVID-19 caused by any of the SARS-CoV-2 variants, including Omicron [232]. This claim suggests a separation between protection against infection and protection against severe outcomes, implying that the two sets of outcomes are independent. For example, a 2022 study from Israel reported that while protection against confirmed infection was short-lived, protection against severe illness remained stable during the study period [233].
Nevertheless, a detailed 2023 analysis of clinical trials, large-scale observational studies from Israel, and statistical dashboards revealed significant methodological flaws, including inconsistent follow-up periods, uneven exclusion criteria, varying COVID-19 testing rates, selection biases, and selective result reporting [234]. The analysis found no difference in the conditional probability of death or severe illness among infected individuals between vaccinated and unvaccinated groups in various widely cited studies. The authors concluded that there is no robust evidence to support claims that a second mRNA booster effectively prevents severe illness or mortality. Moreover, effectiveness estimates from numerous observational studies and the mRNA COVID-19 vaccine clinical trials are likely exaggerated due to methodological flaws. Many factors can skew the evaluation of COVID-19 vaccine effectiveness, resulting in a distorted perception of product efficacy. Key among these are errors in categorizing (1) COVID-19 infections (often based on a single RT-PCR test), (2) COVID-19-related deaths or “COVID deaths”, due to misattributing the death to COVID-19 simply based on the RT-PCR result; and (3) an individual’s vaccination status as “fully vaccinated” or not. Differences in defining vaccination status, testing methods, and confounding variables like disease risk factors can introduce significant biases [235,236].
Critical research attention has focused on the “case-counting window bias”, stemming from errors in diagnosing cases and assigning individuals to vaccinated or unvaccinated groups. This source of bias can greatly inflate vaccine efficacy estimates [138,237,238]. In most studies comparing vaccinated to unvaccinated groups, the observation period for counting AEs were started too late (typically 1-3 weeks after the second dose); however, the counting of AEs in the vaccinated should begin at the time of the initial dose, in order to catch the majority of serious AEs, including death [239]. This common practice results in misclassifying as “unvaccinated” anyone experiencing a serious AE and previously injected with the mRNA products up to the start of the counting window. By having the case-counting window in the vaccinated individuals begin only one or more weeks after the second dose, the study methodology can make an ineffective vaccine appear to be between 50% and 65% effective, particularly if cases in the “partially vaccinated” are given “unvaccinated” status [240]. In this way, even a negatively effective vaccine can be made to appear moderately effective.
Furthermore re-analyses have revealed persistent misclassification issues in large-scale studies assessing the safety and efficacy of mRNA-based vaccines. A systematic review by Neil et al. analyzed 39 studies that showed such miscategorization errors, where vaccinated individuals were mistakenly classified as unvaccinated for a set period post-injection; nearly one-third (31%) of the studies exhibited at least one form of bias [236]. The authors assert that most observational studies have falsely claimed that mRNA-vaccinated individuals had lower COVID-19 infection and mortality rates compared to unvaccinated individuals. Such claims have been used to justify ongoing distribution of the modified mRNA products.
Evidence that the modified mRNA injections may promote hospitalizations and mortality is highly relevant here. Some studies indicate negative efficacy in terms of increasing post-injection hospitalizations, morbidity, and mortality. A comprehensive Israeli study involving 32,000 individuals who received the COVID-19 mRNA vaccinations found a 27-fold greater risk of symptomatic COVID-19 and an 8-fold higher likelihood of hospitalization compared to unvaccinated individuals within the same healthcare system [241]. This suggests that either the unvaccinated group had superior immunity against SARS-CoV-2, or the mRNA vaccinees experienced a worsening of protection, as well as possibly more serious AEs warranting hospitalization. Hospitalization is often associated with a heightened risk of premature death. In this vein, two separate studies examining hospitalized mRNA recipients indicated that mortality rates after developing severe COVID-19 disease were higher following additional doses of the COVID-19 mRNA injectable products [242,243].
Among the central tenets of the Hybrid Harms Hypothesis is that serious AEs (including death) may be misattributed to the COVID-19 infections due to temporal biases. As an example, it is often claimed that serious cardiac outcomes (e.g., fulminant myocarditis) related to COVID-19 infection have shown a higher mortality than myocarditis related to mRNA vaccination [244,245,246]. Several of the coauthors of the present paper have recently laid out an in-depth explanation for why this assertion is incorrect [247]. In that evidenced-based narrative review, however, we did not consider a question that forms the core premise of our present inquiry: How many of those infection-associated cardiac events were preceded by exposure to COVID-19 mRNA vaccinations, which could have increased the likelihood of more serious AEs?
Along similar lines, in 2021-2022, COVID-19 was counted as the primary cause of death for over 940,000 people in the United States, including at least 1,289 children and young people aged 0–19 years [248]. At least 821 of these younger persons’ deaths occurred between August 1, 2021, and July 31, 2022. Given the low death rate in this age group, the impact of COVID-19 mortality is best evaluated by comparing it to other causes of death. This was done in a study by Flaxman and colleagues, which concluded that COVID-19 was “a leading cause of death among individuals aged 0 to 19 years in the US”, even after Omicron, a relatively benign infection, became dominant in early 2022 [248]. Remarkably, the authors did not consider COVID-19 mRNA vaccination as a potential risk factor, despite the CDC-verified myocarditis signal and association with other cardiac AEs (see section, “Hybrid harms due to co-amplification of cardiac AEs”). Instead, they evaluated COVID-19 as the underlying cause of death among young persons aged 0 to 19 years only in the context of the 10 leading causes of death that had been identified in 2019. How many of these younger persons who ostensibly died “from COVID” had previously received the COVID-19 mRNA vaccination? Again, in light of our hypothesis, such an exposure could have greatly amplified the individual’s immunologic vulnerability to the potential lethality of the infection, resulting in the misattribution. Among children and adolescents, the IFR of Omicron in 2022 was estimated to be only 0.0003%, or near zero.91 Thornley et al. estimate that, for younger generations (<40 years) overall, SARS-CoV-2 infection severity and fatality rates even as early as 2020 were comparable to those of influenza [249]. (This assessment would not apply to immunocompromised individuals, such as cancer patients undergoing intensive chemotherapy).
Hybrid Immunity Versus Hybrid Harms
Hybrid immunity, defined as the potentially synergistic immune protection derived from prior SARS-CoV-2 infection followed by COVID-19 mRNA vaccination, has been shown to confer enhanced immunological responses compared to either natural infection or vaccination alone. Studies indicate that individuals with recent SARS-CoV-2 exposure who receive mRNA vaccines exhibit a more robust and diverse antibody repertoire, characterized by elevated neutralizing antibody titers and a broader spectrum of epitope recognition [250]. For example, a large study in France (n=933,397) found that higher levels of booster-related protection against Omicron infection were associated with previous SARS-CoV-2 infection, especially when that infection was recent and involved a dominant antigenic-related variant [66]. The gain in protection with the second booster was lower than the gain observed with the first booster, at equal time points after vaccination. Overall, this hybrid-enhanced humoral immunity has been associated with a 5- to 10-fold increase in memory B cell populations compared to those induced by infection or vaccination alone [251]. In short, mRNA vaccination following infection generates a stronger humoral response than either vaccination or infection alone, with the timing of the prior infection, rather than its severity, influencing post-vaccination IgG levels [252].
It is important to emphasize, however, that most studies focusing on the benefits of hybrid immunity were concluded after only a few months. Based on a comprehensive systematic review, booster doses were associated with an approximate 91% reduction in severe Omicron infections (95% CI: 87.1–93.8) at two weeks post-vaccination, decreasing to about 78% (95% CI: 70.5–84.7) by 26 weeks.74 The authors reported that hybrid immunity offered more sustained protection than vaccination or infection alone. However, protection against Omicron reinfection declined to about 50% (95% CI: 32.5–67.8) by 26 weeks post-vaccination. Among individuals with hybrid immunity who received booster doses, protection initially reached 80.6% (95% CI: 70.0–91.2) but then dropped to 36.9% (95% CI: 19.3–54.6) within 16 weeks. The short-lived benefits suggest that many studies claiming superior protection from hybrid immunity suffered from extreme temporal bias. A study in Germany concluded that prior mRNA vaccinations or coronavirus infections provide strong protection against the Omicron variant for only a few months at most, suggesting that the coronavirus circulates seasonally, much like influenza [253].
By 2023, widespread SARS-CoV-2 exposure in developed nations suggested that most individuals likely possess some degree of natural immunity. This would tend to amplify the humoral immune stimulation afforded by the mRNA boosters. However, given the mild pathogenicity of Omicron variants, the clinical necessity of such augmented antibody responses warrants serious scrutiny. Specifically, hybrid immunity raises concerns regarding the risks associated with excessive antibody production. Overproduction of non-neutralizing antibodies may contribute to vaccine-associated enhanced disease (VAED), potentially mediated by the phenomenon of pathogenic priming (also known as antibody-dependent enhancement) [254]. To date, VAED following mRNA vaccination has been mainly observed in animal models, with few cases reported in the clinical setting [255,256,257]. A cross-sectional study using the Vaccine Safety Datalink concluded that reduced disease severity among hospitalized vaccinated versus hospitalized unvaccinated individuals supported the absence of such disease enhancement; however, this study involved the case-counting window bias, which would have miscategorized many vaccinees as unvaccinated (see section, “Illusions of protection against severe disease”) [258]. The healthy vaccinee bias may have also been a factor, with a disproportionate number of individuals with underlying diseases possibly choosing to forgo the injections. The presence of pre-existing medical comorbidities is associated with an elevated incidence and severity of BTIs as noted previously [259].
The case-counting window bias is exemplified in a large retrospective cohort study of more than 10 million people, focusing on whether COVID-19 vaccination could help prevent intensive care unit (ICU) admission for COVID-19 pneumonia and improve patient outcomes [260]. ICU patients in Lombardi, Italy, were classified as “vaccinated” 14 days after the second dose of the mRNA vaccine, either BNT162b2 or mRNA-1273. Thus, any patient who tested positive at the time of ICU admission—even if the reason for admission was a serious AE—prior to this 14-day point would have been counted as “unvaccinated”. The study concluded that the mRNA vaccinations “significantly decreased the risk of ICU admission for COVID-19 pneumonia.” (Surprisingly, despite this bias, both ICU and hospital mortality rates were similar between vaccinated and unvaccinated patients after adjustment for age, heart disease, and lung injury measurements upon ICU admission.) In light of our hypothesis, it is biologically plausible that the risk of COVID-19 pneumonia was enhanced by the previous vaccination, again even in those ICU patients misclassified as “unvaccinated”. Studies in SARS animal models revealed that spike-protein-based vaccines could worsen outcomes, including lung immunopathology and liver inflammation, following viral challenge [261,262]. Since COVID-19 pneumonia often involves a cytokine storm in the lungs, perhaps the risk of uncontrolled cytokine-mediated inflammation is amplified when the SARS-CoV-2 infection is recently preceded by the mRNA vaccination [263].
This brings us to a critical consideration in the context of hybrid immunity: the timing of vaccination in relation to SARS-CoV-2 infection. Most studies of hybrid immunity have emphasized the immunological benefit of pre-vaccination infection. While mRNA vaccinations were initially hypothesized to mitigate disease severity by priming the immune system for a rapid response to subsequent coronavirus infections, emerging evidence suggests potential adverse outcomes. The COVID-19 mRNA products have been associated with transient suppression of innate immunity, including impaired type I interferon signaling and dysregulated protein synthesis, which may compromise early antiviral defenses [193]. Furthermore, excessive non-neutralizing antibody production may increase the risk of autoimmune reactions through molecular mimicry, where antibodies cross-react with host tissues, potentially triggering inflammatory pathologies [264,265,266]. Such mechanisms could elevate the risk of chronic inflammatory conditions, including cardiovascular diseases, malignancies, and other autoimmune disorders [267,268]. Repeated booster administrations would theoretically increase the risk of these serious AEs, potentially compromising the long-term safety profile of mRNA vaccinations.
Safety data from clinical trials and post-marketing surveillance, particularly for Pfizer’s BNT162b2 vaccine, have reported approximately 1.6 million adverse events over a six-month period [132,133,134]. Moderna’s mRNA-1273 vaccine elicits a stronger humoral immune response, purportedly resulting in lower rates of symptomatic infection and severe COVID-19 outcomes compared to BNT162b2 [269]. However, this enhanced or “super-charged” immune response is accompanied by a higher incidence of serious AEs, as evidenced by clinical trials, observational studies, and government-sponsored surveillance data [19,20,270,271,272,273,274]. Indeed, a study in Taiwan involving 1,711 booster recipients showed that simply switching from Moderna’s mRNA1 [273] product to a different brand lowered the risk of AEs by 18% [275]. A two-year analysis of Moderna’s vaccine, covering over 772 million doses, reported that 0.7% of the more than 2.5 million AEs led to fatalities (~17,500 deaths total) [276].
Regardless of the brand, the mRNA vaccine-induced spike protein, combined with other components in the formulation, appears to elicit a far stronger and more narrow immune response compared to the spike protein of the native coronavirus. Indeed, individuals who were up-to-date with their COVID-19 vaccinations showed an average of 50 times greater antibody levels than unvaccinated, naturally infected persons [277] It is fair to describe these antibody levels as excessive relative to “natural immunity”. Moreover, as noted above, the “hybrid immunity” afforded by the injection-infection interaction would further bolster the humoral immune response, as indicated by higher antibody titers. Although this would seem to suggest an even stronger immune “benefit”, it should be noted that the amplified humoral response is associated with increased risks of hyperinflammation, severe immunopathology, and heightened reactogenicity, all of which may contribute to a wide range of AEs [278,279,280,281,282,283,284,285,286].
The biological explanation for this immune paradox has recently become clearer. Although antibody titers are strongly associated with protection against infection (primarily because they opsonize pathogens, making it easier for macrophages to recognize them), excessive antibody production can have untoward effects. Specifically, very high antibody levels will inhibit the capacity of the macrophages to phagocytize and destroy pathogens or cancer cells [277]. This hyperimmune antibody-macrophage interaction would logically help explain why the hybrid immunity afforded by the combination of the COVID-19 mRNA vaccinations and coronavirus infections may paradoxically translate into more serious AEs. For example, individuals exposed to the coronavirus shortly after mRNA vaccination may face an elevated risk of serious cardiac events, as we document in the section “Hybrid harms due to co-amplification of cardiovascular and hematologic AEs”. The trade-off between the hybrid immunity “enhanced” protection against SARS-CoV-2 and increased risk of serious AEs underscores the need for a more balanced assessment of mRNA vaccination in the context of hybrid immunity.
In conclusion, while hybrid immunity significantly enhances immune responses to SARS-CoV-2, the potential risks of excessive (and wrong type of) antibody production, innate immune suppression, and serious adverse events necessitate cautious consideration. The added theoretical benefit afforded by a greater “boost” in antibody production via hybrid immunity seems highly questionable, particularly in light of the very low IFR and relatively high risk of serious AEs for younger populations whose IFR was always near zero [91]. Further research is essential to elucidate the long-term safety and efficacy of repeated mRNA booster doses, particularly in populations with widespread natural immunity. It is disingenuous for public health officials to continue insinuating that “more is better”, or that hybrid immunity is superior to natural immunity in terms of its overall biological and public health impacts. Indeed, our research indicates the opposite to be true. Claims regarding the ability of hybrid immunity to reduce severe disease, hospitalization, and mortality, are typically made by those with blatant conflicts of interest (with funding having been obtained by Pfizer, Moderna, and other vaccine companies), as can be readily seen in many publications that make such claims.
6. Post Vaccine Syndrome Often Subsumes Post COVID Syndrome
Post-acute Sequelae of COVID-19 (PASC), commonly referred to as either Post COVID-19 Syndrome or “long COVID,” manifests in a subset of individuals following the acute phase of SARS-CoV-2 infection. This multifaceted condition is characterized by persistent symptoms, including severe fatigue, cognitive impairment, myalgia, dyspnea, paresthesia, and thoracic discomfort, often lasting for months [287]. PASC is a multisystemic disorder involving dysautonomia, neuroinflammation, immune dysregulation, cardiovascular and coagulopathic abnormalities, and multi-organ involvement [288]. Symptom-based subtypes have been proposed to refine its classification [289].
In a retrospective, multi-database cohort study, Lam et al. investigated cardiovascular, cerebrovascular, neurological, psychiatric, nephrotic, and hepatic events as long-term PASC infection [290]. The authors reported consistent increases in the incidence of PASC outcomes affecting the cardiovascular, respiratory, neurological, nephrological, and hepatic systems, as well as significant increases in cardiovascular and all-cause mortality across two populations, in Hong Kong and the United Kingdom, respectively. The diverse range of outcomes amongst patients with COVID-19 infection demonstrated the multi-organ involvement of PASC following infection.
Early in the pandemic, public health authorities suggested that mRNA-based COVID-19 vaccines would reduce the incidence of PASC, a claim that influenced vaccination uptake, particularly among younger populations. However, emerging evidence challenges this assertion, instead indicating a probable association between COVID-19 mRNA vaccination and an increased risk of PASC-like syndromes. The spike protein, a shared feature of SARS-CoV-2 infection and mRNA vaccines, is implicated in the pathogenesis of both PASC and post-vaccination syndromes [291,292]. As we noted previously, COVID-19 mRNA vaccinations induce systemic spike protein production for at least two to three years post-vaccination. The prolonged presence of spike protein has been detected in individuals with PASC, suggesting a role in perpetuating symptoms [293,294]. An exaggerated immune response to mRNA vaccine-derived spike protein may contribute to PASC-like symptoms [295]. The clinical approach to these conditions requires an integrated understanding of the underlying “spikeopathy” and associated immune dysfunction [296].
To explore the relationship between PASC and post-COVID-19 vaccine syndrome (PCVS) due to COVID-19 mRNA vaccinations, researchers measured SARS-CoV-2 Semi-Quant Spike Antibody levels in 100 consecutive patients presenting with long COVID symptoms [297]. The cohort comprised 81 vaccinated individuals with PCVS and no comorbidities, and 19 unvaccinated individuals with confirmed prior SARS-CoV-2 infection, all at least three months post-infection or vaccination. The vaccinated cohort exhibited an average spike antibody level of 11,356 U/mL (range: 1,291 to >25,000), approximately seven times higher than the unvaccinated cohort’s average of 1,632 U/mL (range: 1.5 to 4,614), despite the fact that the vaccinated group had no recent infections. This may suggest that the COVID-19 mRNA injections lead to a much higher spike protein burden than that associated with SARS-CoV-2 infections.
A retrospective analysis found that 70% of PASC cases occurred in individuals who had completed a full course of the COVID-19 mRNA vaccinations, suggesting that the genetic injectables may exacerbate or precipitate PASC symptomology in a significant proportion of these presumed “long COVID” cases [298]. Notably, unvaccinated individuals who had been exposed to Omicron showed the lowest PASC incidence, with previous Omicron infection being strongly associated with a lower PASC risk (OR 0.14, 95% CI 0.07; 0.25). No lowering of PASC risk was seen with the COVID-19 mRNA vaccinations. Thus, contrary to initial claims by the vaccine enterprise, mRNA-vaccinated individuals appear more susceptible to PASC than their unvaccinated counterparts.
In a global online survey of 7,541 individuals from 95 countries, Bhargava and Inslicht investigated PASC and COVID-19 vaccine-related AEs in vaccinated and unvaccinated individuals following their first SARS-CoV-2 infection [299]. Vaccinated males reported higher rates of severe outcomes, defined as high-grade fever (>102°F) or hospitalization, compared to unvaccinated males (13.64% vs. 8.34%; p=0.0483; HR=1.63 [95% CI: 1.008, 2.65]). Women experienced PASC more frequently than men and reported significantly higher vaccine-associated AEs after the first dose (60.85% vs. 48.79%; p<0.01). Vaccine-related hospitalizations were more common in SARS-CoV-2-naïve vaccinated individuals (6.24%) than in unvaccinated individuals post-infection (1.06%). Vaccinated women reported significantly greater menstrual cycle disturbances as reproductive PASC, while vaccinated men noted more hormonal aberrations and sexual dysfunction compared to their unvaccinated counterparts. The authors also noted that “there was no benefit of mRNA or adenovirus COVID-19 vaccines on severe disease,” and that pre-existing conditions and/or medications seemed to exert a much stronger influence.
These and related observations led Scholkmann et al. to recognize novel post-vaccination syndromes with clinical features overlapping those of PASC [300]. The authors propose the inclusion of a specific diagnostic code for “post-COVID-19 vaccination condition, unspecified” in future iterations of the International Classification of Diseases. Given the mechanistic overlap, other researchers advocate reclassifying PASC as either PCVS or “post-acute COVID-19 injection syndrome” (PACIS) to reflect the potential contribution of the modified mRNA injectables, particularly in regions with extensive coverage. As shown in Table 2, PCVS/PACIS may subsume many cases of PASC because it is difficult to disentangle the prolonged spike protein production elicited from the COVID-19 mRNA vaccinations from spike-related effects of prior or subsequent coronavirus infections (see Table 2; sources are shown in Appendix A).
To summarize this interaction, both SARS-CoV-2 infection and COVID-19 vaccination expose the human body to spike protein. This protein trimerizes and is not readily cleared from systemic circulation or tissues months to years after systemic exposure. Spike protein retention leads to immune system failure, recurrent infections, and a variety of spike protein syndromes including myocarditis, vasculopathies, venous thromboembolism, stroke, transverse myelitis, and auto-immune syndromes. Our observations call for an immediate shift in research priorities for federal agencies to focus on spike protein syndromes. Such research should fully describe the pharmacokinetics and dynamics of mRNA, adenoviral DNA, and spike protein, with an aim to clear or neutralize this dangerous pathogenic entity.
From a critical research perspective, many studies focusing on PASC may not adequately take into account the influence of COVID-19 mRNA vaccination on PASC outcomes, as well as the pathological redundancy between the injections and infections. Some PASC conditions (e.g., cardiovascular, hematological, and neurological) clearly overlap with known AEs linked with the COVID-19 mRNA products, such as myocarditis, stroke, or thromboembolism (see Table 2). The scientific community does not restrict PASC to unvaccinated individuals, mainly because COVID-19 vaccinated individuals with BTIs or reinfections can and do develop PASC. Due to the identical signs and symptoms, however, some cases counted as PASC may actually be vaccine injuries, either short term or long term. If vaccine-related AEs (e.g., myocarditis, stroke, or thromboembolism) are misclassified as PASC, this would artificially inflate the reported PASC incidence in the vaccinated group. This, in turn, would mean that many vaccine injuries could be solely and thus erroneously attributed to the adverse effects of the coronavirus infection rather than to the mRNA vaccination.
This misclassification is further complicated by the case-counting window bias described previously. As we noted, most studies comparing COVID-19 vaccinated to unvaccinated groups suffer from this fundamental flaw, which biases upward the safety and effectiveness of the mRNA vaccinations. In the context of the case-counting window bias, the unvaccinated are being grouped together with previously mRNA-injected individuals people who either (a) have not yet reached “immunologic maturity” on the first two injections (i.e., not up to 14-28 days after the second injection), (b) only partially vaccinated, or (c) or were vaccinated too long ago to be considered “up-to-date” and thus counted as having vaccine-induced immunity. Any PASC-like AEs that arise in these mRNA-injected individuals will be counted as occurring among the “unvaccinated”. Due to this additional layer of misclassification, the vaccine-related AEs are not only misclassified as PASC, but as PASC occurring among the unvaccinated. This fraudulent practice has profoundly distorted many risk-benefit analyses of COVID-19 mRNA vaccinations, resulting in gross overestimates (largely inverted) of the mRNA products’ ability to reduce cardiovascular, hematological, neuropsychiatric, and autoimmune PASC outcomes.
(Note: This common practice of miscategorizing the unvaccinated means that those who never received the modified mRNA injections—that is, the true “unvaccinated”—are unfairly characterized in the public eye. This is because the practice results in misattributing adverse health outcomes to a group that may have lower risks of many PASC outcomes that overlap with vaccine-induced AEs. We argued this convincingly in a previous paper on myocarditis.247 Whether or not this is being done deliberately, the miscategorization serves to (a) reinforce the “safe and effective” narrative, thereby ensuring more pharmaceutical industry funding of mRNA vaccination and PASC research, and (b) motivate the policymakers, commentators, and the general public to continue to embrace the vaccinations).
In conclusion, the association of COVID-19 mRNA vaccinations with PASC warrants further investigation. Ideally, we would see more studies with rigorous diagnostic criteria (e.g., requiring confirmed infection history, temporal analysis, or specific biomarkers) in order to disentangle PASC from vaccine AEs. The long-term persistence of mRNA vaccine-derived spike protein, as well as spike protein from the coronavirus infection, suggests that the interaction and overlapping contributions of both sources may underlie the vast majority of “long Covid” cases diagnosed since 2021. Both sources may contribute either additively or synergistically to the chronic immune dysfunction associated with prolonged “spikeopathy” disorders. This complex reality highlights the need for longitudinal studies to clarify the mechanisms underlying PCVS/PACIS and related conditions. Our Hybrid Harms Hypothesis posits that a significant proportion of PASC cases reported since 2021 may be attributable to mRNA vaccine-related immunological effects in conjunction with those effects associated with the coronavirus infection, rather than to the natural infection alone.
7. Support for the Hybrid Harms Hypothesis
7.1. Overlapping “Spikeopathies” from mRNA Vaccinations and Coronavirus Infections
The likelihood of either additive or synergistic interactions between the COVID-19 mRNA vaccination and coronavirus infections may be indicated by the overlapping risks associated with both exposures. As documented in the preceding section, many of the sequelae of the coronavirus infections are also known to be serious AEs linked with the COVID-19 mRNA vaccinations. In terms of potential biological causality, this was not unexpected: the spike protein is the common denominator between the COVID-19 vaccinations and the coronavirus infections, which is to say the functional basis for enabling and promulgating those infections [169]. The resultant “spikeopathy” provides a logical basis for the fact that many pathologies generated by the SARS-CoV-2 infections overlap with those associated with the mRNA injections.
A prominent pathophysiological example of this spikeopathic convergence relates to clotting disorders that can be triggered and amplified by both the mRNA injections and coronavirus infections. It is an inconvenient truth that the same coagulopathies seen in severe COVID-19 disease are mirrored by the serious thrombotic complications caused by the COVID-19 mRNA vaccinations, including myocardial infarction, disseminated intravascular coagulation, and venous thromboembolism [301,302,303]. Both the coronavirus and the mRNA vaccine can trigger endothelial dysfunction, systemic inflammation, and dysregulated coagulation cascades [169,304,305]. In COVID-19, the spike protein’s interaction with ACE2 receptors drives a hypercoagulable state through cytokine storms and upregulation of procoagulant factors, leading to microvascular thrombosis and myocardial injury [306,307]. Similarly, mRNA vaccines may induce immunopathologies such as vaccine-induced immune thrombotic thrombocytopenia (VITT), involving autoantibodies against Platelet Factor 4 or transient spike protein-mediated endothelial activation, promoting thrombosis [308,309,310].
The pathophysiology of COVID-19 involves diverse thromboinflammatory processes, such as activation of the coagulation cascade, inhibition of fibrinolysis, damage to vascular endothelium, and prothrombotic modifications in immune cells including macrophages and neutrophils.311 While thrombocytopenia does not typically manifest as an early sign of infectious coagulopathy, emerging research underscores the critical involvement of platelets in COVID-19-related coagulopathy. SARS-CoV-2 and its spike protein have been shown to directly or indirectly stimulate the secretion of prothrombotic and inflammatory mediators, thereby contributing to this coagulopathic state. VITT (mentioned above) is characterized by atypical thrombotic sites, such as cerebral venous sinuses, and it is hypothesized that prothrombotic cascades triggered by the coronavirus spike protein play a role in its etiology. Iba et al. conjecture that the spike protein—whether from the infection or the mRNA injection, or both—acts as a primary initiator, with dysregulated immune responses and inflammatory processes amplifying the generation of anti-platelet factor 4 antibodies [312].
The founding clinical trials provide robust evidence of these pathogenic virus-vaccine redundancies in the context of COVID-19. The reanalysis by Fraiman et al. found that, among the 236 serious “adverse events of special interest” (AESIs) observed in the Pfizer and Moderna founding trials, 97% (230/236) were event types that also showed a clear association with COVID-19 [129]. The authors’ definition of AESIs was based, in part, on the manifestation of “COVID-19 specific immunopathogenesis”. All redundancies aside, some key distinctions are worth noting. The mRNA vaccine-induced spike protein displays greater immunogenicity than its coronavirus infection-induced counterpart; and yet, the increased humoral response elicited by the mRNA products has also been linked with a much wider range of severe reactogenicity, immunopathology, and serious AEs overall [169,278,279,280,281,283,284,285,286]. This means that, paradoxically, the stronger the humoral response following the COVID-19 vaccinations, the stronger the potential for serious AEs that can also be demonstrated. A further potentiation of this etiopathogenesis may result from the immunological interaction with infections that occur before or after the mRNA vaccination. (We revisit this connection between AEs and hyperimmunity in the Discussion).
7.2. Hybrid Harms Due to Co-Amplification of Cardiovascular and Hematologic AEs
COVID-19 mRNA vaccine-induced myocarditis (inflammation of the myocardium or cardiac muscle) was initially reported by the CDC as a safety signal in May 2021 [313]. Public health agencies, academic institutions, and professional organizations stated emphatically that coronavirus infections were causing more myocarditis than the mRNA vaccinations [314]. This claim was soundly refuted in a recent review [247] From a biological perspective, evidence supports a stronger link between COVID-19 mRNA vaccines and severe cardiac complications compared to SARS-CoV-2 infections [315]. The mRNA vaccination and its resultant spike protein have been identified in cardiac tissue of individuals who succumbed post-vaccination with mRNA-based COVID-19 products, as well as in cases of vaccine-associated myocarditis [170,184].
Autopsies of individuals who died after SARS-CoV-2 infection indicate that myocarditis, when present, does not stem from direct viral infection of the heart [316]. In experimental models, the BNT162b2 and mRNA-1273 vaccines have demonstrated direct cardiotoxic effects on rat cardiomyocytes within 48 hours post-administration, leading to distinct pathophysiological impairments linked to cardiomyopathy [312]. Such direct cardiotoxic effects have not been observed with SARS-CoV-2 or its Omicron variant. These findings suggest a more direct mechanistic pathway for mRNA vaccine-induced myocarditis compared to SARS-CoV-2 infections, strengthening the argument that the mRNA vaccinations pose a higher risk of myocarditis than the virus itself.
Considering our hypothesis and the 2–3-year post-vaccination WOV, the injection-infection dichotomy implied in the preceding paragraph may be somewhat illusory, or at least biologically irrelevant. First, many post-vaccination SARS-CoV-2 infections or BTIs are by definition superimposed on a baseline of failed COVID-19 vaccination. Second, the mRNA injections taking place within the WOV may potentiate pathogenic processes that are subsequently triggered by the infection. These include the development and progression of myocarditis and its cardiac sequelae.
Prospective studies using cardiac function data (high-sensitivity cardiac troponins and electrocardiographic changes) indicate that COVID-19 mRNA vaccine-attributable myocardial injury occurs in approximately 2.5% of the younger population (<age 40) [317,318]. However, the cardiac troponin elevations were mild and transient. These studies demonstrate that most cases of mRNA-related myocarditis have a subclinical presentation that leads to high under-detection rates in large surveillance and observational studies. Daniels et al. studied 1,597 college athletes in the Big Ten with cardiac MRI screening following SARS-CoV-2 infection [319]. Subclinical cases in this study outnumbered clinical cases of myocarditis by threefold, with an approximate 75% incidence (28/1597 x 100% ~=1.75%) of subclinical myocarditis. The clinical incidence was approximately 0.6% (9/1597 x 100), which should not be characterized as minimal given the hundreds of millions of COVID-19 mRNA doses administered to young people. The incidence based only on initial symptom-based screening was 0.31%, about 7 times lower than what was detected by cardiac MRI. None of the cases required hospitalization.
In line with our current hypothesis, we posit that the mRNA vaccine-related subclinical myocarditis may be exacerbated as persistent vaccine-derived spike protein in cardiac tissues, combined with later exposure to infection-induced spike protein, triggers hyperimmune responses and cardiomyocyte damage. Subclinical myocarditis can lead to microscopic fibrosis in cardiac tissue; a second immune challenge (from the coronavirus infection) may then increase susceptibility to arrhythmias, contractile dysfunction, or further tissue remodeling. In this scenario, the mRNA vaccination predisposes the myocardium to heightened inflammation and tissue damage, while the subsequent viral infection serves as additional cardiac stressor, thereby increasing the likelihood that the subclinical myocarditis will be transformed into fulminant myocarditis, along with an increased likelihood of arrhythmias and heart failure. Prospective studies with appropriate testing (e.g., cardiac troponins, biopsy, and tissue morphology) are needed to confirm this etiopathogenesis.
Repeated doses of the COVID-19 mRNA product have been shown to increase the risk of myocarditis. Multi-country analyses have shown that the second mRNA dose was associated with a three- to five-fold increase in the risk of excess myocarditis and/or pericarditis, with young males being at greatest risk [320,321,322]. In a large systematic review, Moderna’s mRNA-1273 resulted in a doubling in the myocarditis/pericarditis risk when compared to Pfizer’s BNT162b2 [323]. Importantly, the primary series for Moderna’s product contains about three times the concentration of mRNA (100 micrograms) when compared to its Pfizer counterpart (30 micrograms) [324]. (Note: The decision to receive the mRNA vaccination initially involved opting for two injections, typically spaced 21-28 days apart. Therefore, getting “fully vaccinated” meant committing to both the first and second doses; and the risk of myocarditis from full vaccination essentially reflects the combined risks of both doses, i.e., a double-spike effect). Taken together, these findings suggest a dose-response relationship with regard to myocarditis and other serious cardiac AEs. This also offers indirect support for the Hybrid Harms Hypothesis, with increasing exposure to the spike protein (whether from the vaccine or virus, or both) translating into more serious pathological outcomes.
The following three case synopses offer examples of the “hybrid harms” phenomenon and confirm many clinical observations of the senior author of the present paper. It is interesting to note that all three of these published cases had received the Pfizer mRNA vaccine (Comirnaty, BNT162b2) four months prior to the coronavirus infection. In each case, the infection was cited as the cause or precipitating factor behind the myocarditis.
- A 26-year-old previously healthy male was admitted to the emergency room by his family physician after one month of worsening fatigue, palpitations, and dyspnea [325]. He had tested positive for SARS-CoV-2 eight weeks earlier, and he received a second dose of the COVID-19 mRNA vaccine four months prior to the infection. Echocardiography and cardiac MRI showed severely reduced left ventricular function and strong midmyocardial late gadolinium enhancement. Endomyocardial biopsy confirmed acute lymphocytic myocarditis.
- A 42-year-old male tested positive for Omicron and was admitted to the ICU in January 2022 with chest pain and ST-segment elevation in the inferior leads [326]. He had a history of peri-myocarditis in 2008 without recurrence or autoimmune disorder diagnoses. Four months earlier (22 August 2021), the patient had received his third dose of the Pfizer mRNA vaccine. Cardiac MRI confirmed myocarditis, with late gadolinium enhancement showing 22% left ventricular mass involvement.
- A 60-year-old male presented to the emergency department on 11 January 2022 with syncope and palpitations, testing positive for Omicron via RT-PCR [326]. He had received his third Pfizer mRNA vaccine dose four months prior, on 30 August 2021. He experienced ventricular tachycardia (250 beats/minute) requiring urgent cardioversion. No further malignant arrhythmias occurred during hospitalization. Coronary angiography showed non-obstructive disease, and cardiac MRI indicated acute myocarditis with 19% left ventricular mass involvement.
The temporal sequence in the cases outlined above can also be reversed. For instance, the peer-reviewed literature contains case reports of individuals who first contracted the SARS-CoV-2 infection, then later received the mRNA vaccination before being diagnosed with myocarditis [327,328,329]. Some case reports reflect multiple etiologies of myocarditis aside from COVID-19 vaccination [330]. In addition to SARS-CoV-2 infection, the etiologies may involve other viral infections linked with myocarditis, such as Coxsackievirus, Adenovirus, and Parvovirus B19. As part of their treatment for COVID-19, some were exposed to Remdesivir, a drug that has significant cardiotoxic effects, in some cases resulting in prolonged QT intervals and torsades de pointes, which in turn can lead to ventricular arrhythmias and sudden cardiac arrest [331,332,333].
Because both the COVID-19 vaccination and coronavirus infection can cause fulminant myocarditis, perhaps the precise sequencing is not critical in terms of causation. It is biologically plausible that a priming effect occurs with either the injection or the infection, depending on which exposure comes first. In the case synopses provided above (with infection occurring post injection), the COVID-19 mRNA vaccination may train the immune system to recognize the spike protein, creating memory T and B cells. Upon encountering the coronavirus, these cells may mount a rapid and robust immune response. In some cases, this heightened activity may inadvertently exacerbate inflammation in the myocardium, particularly if residual inflammation from the mRNA-induced myocarditis has not completely resolved. A subsequent SARS-CoV-2 infection could restimulate immune cells primed by the mRNA injection or even a preceding initial infection, and the resulting hyperimmune response might disproportionately target cardiac tissue, leading to fulminant myocarditis.
Again, in this particular context, the temporal framework for the causal chain introduces a conceptual obstacle. If a previously mRNA-vaccinated individual develops myocarditis after contracting COVID-19, one would plausibly expect the cardiac event to be attributed solely to the infection, the most recent determinant or triggering factor. Nonetheless, because the mRNA-induced spike protein production can induce inflammation in cardiac muscle, and because this production can persist for years, it seems wholly plausible that mRNA-attributable subclinical myocarditis, followed by coronavirus infection at any time during the WOV, could further amplify an inflammatory reaction in cardiomyocytes.
Our critics may point to the large self-controlled case series by Patone et al., focusing on about 43 million residents of England. The study population included just under 6 million individuals who had SARS-CoV-2 infection either before or after vaccination [334]. The authors reported that the risk of myocarditis was reduced by about half if one experienced COVID-19 disease after receiving at least 1 dose of the COVID-19 vaccine. This would seem to refute our Hybrid Harms Hypothesis. However, the study suffered from several methodological and design flaws, as we and others have documented [247,335]. Most egregiously, the infection-related myocarditis was grossly overestimated due to reliance on RT-PCR testing as a benchmark for measuring infections; the laboratory-confirmed case count captured only a fraction of true infections, greatly reducing the denominator and thereby artificially inflating the infection-related risk. As Stowe et al. cogently stated, “[T]he attributable risk estimates for COVID-19 used laboratory confirmed cases as the denominator and will be affected by the proportion of all SARS-CoV-2 infections captured by testing, precluding a direct comparison with vaccine-associated attributable risks”. [336]. Bourdon and Pantazatos analyzed the likelihood of myocarditis in males under 40 after COVID-19 vaccination or a confirmed SARS-CoV-2 infection, utilizing incident-rate ratios (IRRs). With certain plausible assumptions, they recalculated the Patone et al. IRR of 4.35 for infection-related cases to 2.75, which is below the IRRs for the second dose of Pfizer’s BNT162b2 (3.08) and the first dose of Moderna’s mRNA-1273 (3.06) [335].
It is worth noting that other investigators, utilizing RT-PCR testing, were still able to show more substantial adverse impacts of COVID-19 mRNA vaccinations compared to natural infections, at least in younger individuals. Karlstad and colleagues conducted a study estimating excess myocarditis cases after mRNA vaccination in a large cohort (n=23 million) across Denmark, Finland, Norway, and Sweden [321]. These Nordic nations provide a robust setting for analysis due to their similar demographics and reliable data collection systems. The study defined a myocarditis event as a hospital admission with a primary or secondary diagnosis of myocarditis. The highest risk of myopericarditis was observed within the first week post-vaccination, particularly among young males aged 16-24. In this group, the study reported 5.55 additional myocarditis cases per 100,000 second doses of BNT162b2 and 18.39 per 100,000 second doses of mRNA-1273. For the same age group, the researchers also noted 1.37 excess myocarditis cases per 100,000 positive SARS-CoV-2 tests over a 28-day period following a positive test. (Note: As explained above, since actual infections likely exceed reported positive tests, the true infection rate is underestimated; this would effectively inflate the denominator and lower the apparent rate of infection-related myocarditis.) Nevertheless, the data indicate that mRNA vaccine-associated myocarditis rates were significantly higher than those linked to infections based on positive tests. Among males aged 16-24, mRNA-related myocarditis cases were approximately four times higher after the second BNT162b2 dose (5.55/1.37=4.05) and over 13 times higher after the second mRNA-1273 dose (18.39/1.37=13.42) compared to infection-related myocarditis.
Other serious cardiac events may offer additional support for the Hybrid Harms Hypothesis. Blasco et al. conducted a single-center retrospective cohort study (n=949 patients) between March 1, 2020 and March 1, 2023, analyzing the humoral immune response to the COVID-19 mRNA vaccinations or the SARS-CoV2 infection [337]. Of 949 patients, 656 (69%) had ST-segment elevation MI (STEMI), a severe type of myocardial infarction characterized by a significant elevation in the ST segment on an electrocardiogram. SARS-CoV-2 spike-specific IgG titers were highest in the vaccinated group: median > 2080 AU/mL in the vaccinated versus 91 in the unvaccinated group. The combination of the mRNA injections and coronavirus infection was associated with the development of severe heart failure and cardiogenic shock in patients with STEMI. The authors speculated that this type of heart attack might be related to a hyperimmune response resulting from the interaction between the mRNA injections and SARS-CoV-2 infections.
In a South Korea study of nearly 3.4 million people who had received one or more COVID-19 mRNA vaccinations from February 2021 to March 2022, Yun and colleagues analyzed heart disease risk, including acute myocarditis, acute pericarditis, acute cardiac injury, cardiac arrest, and cardiac arrhythmia [338]. The authors extracted the date of the initial diagnosis of acute heart diseases (the primary outcome variable) and defined cardiac events within 21 days after the first vaccination date as potential AEs related to vaccination. COVID-19 mRNA recipients infected by SARS-CoV-2 showed a nearly four-fold higher heart disease risk than uninfected mRNA recipients (adjusted HR, 3.56; 95% CI, 1.15-11.04), and younger mRNA recipients had a higher heart disease risk compared to older individuals. Contracting COVID-19 at any time was among the significant covariates that increased the cardiac risk following the mRNA vaccinations.
In a retrospective study of hospitalized elderly patients diagnosed with SARS-CoV-2 infection and referred to an academic medical center in Rome (n=886), Cianci et al. evaluated clinical outcomes of COVID-19 patients diagnosed with heart failure (HF) by comparing them to a matched control group of COVID-19 patients without HF [339]. Individuals were classified as “unvaccinated” if they had never received a COVID-19 vaccine, while those who had received at least one dose of the vaccine were considered “vaccinated”. Primary outcomes included in-hospital mortality, death within 30 days of hospital admission, and ICU admission. COVID-19 with HF was strongly and independently associated with a composite outcome of death and/or need for ICU admission (OR, 6.46, 95%CI: 3.710–11.238; p= <0.0001). COVID-19 patients with HF had a significantly higher vaccination rate than those without HF: 44% of HF patients received at least one dose (p = 0.001), 18% two doses (p = 0.002), and 22% three doses (p = 0.0001). The vaccinated COVID-19 patients had twice the odds of dying (OR, 2.143, 95%CI: 1.094–4.199; p= 0.026) compared to their never-vaccinated counterparts (see Table 3 of Cianci et al.). Thus, in this hospital cohort of elderly COVID-19 patients, there was a greater than twofold elevation in the probability of death linked to the mRNA vaccinations; the relationship was statistically significant.
Al-Aly et al. investigated risks for stroke, myocardial infarction, and pulmonary embolism in individuals with BTIs after a COVID-19 mRNA vaccination with either the mRNA-1273 (Moderna) or BNT162b2 (Pfizer) products [340]. They found significant increased risks for all three events in individuals who experienced a BTI compared to those with no SARS-CoV-2 infection: stroke (HR= 1.76; 95%CI: 1.53–2.03), myocardial infarction (HR= 1.93; 95%CI: 1.64–2.27), and pulmonary embolism (HR= 3.94; 95%CI: 3.39–4.58). These risks were progressively higher for all outcomes with increasing disease severity (i.e., COVID-19 cases that were non-hospitalized, hospitalized, and ICU). This suggests that fully vaccinated individuals may be at increased risk of these life-threatening conditions following SARS-CoV-2 infection. Somewhat paradoxically, however, the authors also reported that the risks were reduced in comparative analyses involving BTI versus SARS-CoV-2 infection without prior vaccination; however, that part of the analysis was seriously flawed due to classifying someone as “unvaccinated” if they suffered from the stroke, myocardial infarction, and pulmonary embolism any day prior to 14-days after the second mRNA injection. For this reason, the comparisons of vaccinated to unvaccinated infected individuals must be considered fraudulent. Due to this fundamental flaw, the study offers only partial support for the Hybrid Harms Hypothesis.
7.3. Epidemiological Studies from the Omicron Era
Analysis of the patterns of COVID-19 outbreaks and excess mortality in 2021-2022 provide additional support for the Hybrid Harms Hypothesis. By mid to late 2021, COVID-19 vaccinations were in full force in many countries, along with antiviral protocols and better clinical management. These factors should have reduced COVID-19 case fatality rates in the extensively vaccinated countries that had reliable reporting systems. In addition, the COVID-19 vaccinations may have helped lower the incidence of COVID-19 mortality in the short term; however, because of the serious AEs associated with the mRNA injections, it appears that both the COVID-19 mortality and all-cause mortality were actually higher, in many countries, after the vaccine rollouts were in full force. Again, this seems paradoxical given that the emergence of the Omicron variant (2021-2022) was predicted to result in a substantial drop in COVID-19 mortality. By 2022, with most countries’ populations being close to 80% “fully vaccinated”, the Omicron variant had become the dominant strain of SARS-CoV-2, with overall mild pathogenicity.
The COVID-19 mRNA vaccinations were widely administered throughout the second and third years of the pandemic, predominantly in the high-income nations across Europe and the Americas. Approximately 139 countries had administered COVID-19 mRNA vaccines, such as those developed by Pfizer-BioNTech and Moderna, by October 2021; the majority of vaccinated countries attained “full coverage” (primary series of the mRNA platform, two injections) for at least 75% of their populations by January 2022 [341]. As alluded to above, Omicron’s relatively mild pathogenicity meant that the infection typically resulted in either no symptoms or only very few symptoms, particularly in those without severe preexisting conditions. Despite Omicron’s less virulent nature, many countries with strong vaccination coverage experienced unexpected spikes in mortality following the Omicron waves. For example, a U.S. analysis found COVID-19 among the top 10 causes of death in younger people through July 2022, with higher rates during Omicron waves than pre-2021 [248]. In both the U.S. and U.K., age-adjusted mortality rates peaked amid 2022 Omicron surges [342]. In France, the case fatality rate (CFR) from Omicron infection was four times higher than that from influenza from week 33 of 2022 through week 12 of 2023 [343]. A study in Germany recorded Omicron infection peaks in October 2022 and March 2023, with a 12.4% in-hospital mortality rate among severe Omicron cases during the one-year observational period; cardiovascular patients with Omicron infection experienced a 4.7-fold higher CFR compared to the total population [344].
Using weekly death data from Nordic registries, Forthun et al. estimated age- and sex-standardized excess mortality rates (per 100,000, referenced to 2010–2019 baselines and standardized to the 2020 Danish population) [345]. Whereas Sweden showed excess mortality in 2020 [75 excess deaths per 100,000 population (95% prediction interval: 29–122)], Denmark, Norway, and Finland experienced excess mortality in 2022 [52 (14–90), 88 (48–128), and 130 (83–177), respectively]. In the latter three countries, the excess mortality started in mid-2021 and persisted throughout the following year, coinciding with widespread Omicron infections. Unlike Sweden, which permitted unrestricted social interactions in 2020 (potentially fostering earlier herd immunity), Denmark, Norway, and Finland implemented stricter lockdown measures, which may have delayed immunity and contributed to higher excess mortality during the Omicron waves. Notably, vaccination coverage in all three countries was stronger than in Sweden by January 2022.
Findings from two studies of the extensively mRNA-vaccinated nation of South Korea offer another compelling example of the paradoxical increase in mortality during the Omicron outbreaks. After no significant excess mortality in South Korea from January 2020 to October 2021, excess mortality rose sharply from November 2021, peaking during the Omicron BA.1/BA.2-dominant period in March (17,634) and April (11,379) 2022 [346]. Excess mortality was highest among the very elderly (≥85 years): 1,048 and 910 per 100,000 for males and females, respectively, in March 2022. A separate study found an 8.7% elevation in age-standardized excess mortality, surging from the third quarter of 2021 to June 2022, again when Omicron was dominant and South Korea had been extensively vaccinated [347].
The comparative multi-country study by Cao et al. assessed percent excess mortality (PEM, age-adjusted via national population structures) following policy shifts in six regions [348]. All six populations had previously been subjected to aggressive lockdowns in 2020-2021. By 2022, the heavily mRNA-vaccinated populations of South Korea, Hong Kong, Singapore, and Australia were showing pronounced elevations in PEM, often synchronizing with waves of Omicron infection. For example, in South Korea, PEM averaged 43.59% in early 2022 and 14.91% in late 2022 during Omicron outbreaks. In Australia, PEM was 39.85% in the first half of 2022 and 35.68% in the second half, overlapping with two Omicron waves. In Hong Kong, the average PEM was an ominous 71.14% during the Omicron wave in the first half of 2022, dropping to 9.19% in the second half. Singapore’s average PEM was 23.98% in early 2022 and 18.53% in late 2022, again corresponding with the Omicron outbreaks. In other studies, the extensively vaccinated countries of Japan and Thailand also showed all-cause mortality spikes that synchronized with Omicron waves in 2022 (see Figure 3) [349,350].
As stated by Cao et al., the governments of Australia and Southeast Asia uniformly blamed Omicron for the increases in all-cause mortality, stating that it was the end of the lockdowns that enabled Omicron outbreaks to result in increased mortality. This assertion was unreasonable, however, given Omicron’s mild infectious nature. Moreover, against a background of high vaccination coverage in these countries by early 2022 (75-90%, according to Our World In Data [341]), it is irrational to dismiss a potential interaction between COVID-19 mRNA vaccination and Omicron infections. These findings may suggest that the mortality peaks corresponding with Omicron waves in 2022 were further amplified by the preceding vaccinations, generating prolonged spike protein production in these populations.
Okoro et al. analyzed World Health Organization data on COVID-19 cases, deaths, and vaccination rates through June 7, 2023, comparing the CFR before and during vaccination across regions [351]. The authors observed a striking rise in COVID-19 deaths in tandem with vaccination coverage, from 43.3% in Africa to 1275% in the Western Pacific. Europe and the Americas comprised over 70% of global COVID-19 deaths despite high vaccination coverage. Paradoxically, COVID-19 mortality surged in 2022-2023—when the milder Omicron variant dominated—particularly in heavily vaccinated regions. Okoro et al. conclude that “COVID-19 mortality increased in the vaccination era, especially in regions with higher vaccination coverage.” Throughout Africa, where mRNA vaccination coverage has been relatively low, COVID-19-related morbidity and mortality rates have been much lower when compared with figures from other continents [352]. By October 2023, the 55 African countries (population: >1.4 billion) reported a COVID-19 mortality rate approximately 4.5 times lower than the U.S. (population: ~330 million), despite minimal public health mandates and far lower vaccination rates [353]. Similarly, Nigeria, with over 216 million people and limited vaccination uptake, exhibited a mortality rate nearly fourfold lower than Israel, a nation of fewer than 10 million with high vaccination coverage and stringent measures [354]. Haiti, with a population of ~11 million and only 5% vaccination coverage, reported negligible COVID-19 mortality [353]. Taken together, these ecological observations suggest the possibility that, with minimal vaccination, COVID-19 may result in lower mortality.
Lastly, in a comprehensive territory-wide cohort study involving 74,303 patients hospitalized with Omicron variant infection (58,894 with primary infection and 2,244 with reinfection), Yan et al. observed that patients surviving the acute phase of hospitalized reinfection faced significantly elevated risks of post-acute all-cause mortality, hospital readmission, and emergency department visits compared to those with primary infection [355]. Despite potentially milder acute outcomes in reinfection cases, the study revealed heightened post-acute risks, which persisted irrespective of COVID-19 vaccination status, with approximately two-thirds of the reinfection cohort having received at least two mRNA vaccine doses. Notably, patients with a history of hospitalization during prior COVID-19 episodes exhibited substantially worse clinical outcomes upon Omicron reinfection compared to those with primary infection, while those with non-hospitalized prior infections showed only a marginally increased risk. There were few unvaccinated individuals in the reinfection group compared to the first-time infection group. These findings may suggest that prior severe infection and potential end-organ damage, perhaps compounded by vaccination status, may contribute to increased vulnerability to adverse outcomes following hospitalized reinfection. Even individuals who develop hybrid immunity against Omicron appear to be at heightened risk of serious adverse outcomes following hospitalized re-infection.
Some researchers assert that the pathogenicity of Omicron subvariants was increasing exponentially from 2022 through 2023 [356]. In light of our hypothesis, we pose the following question: Is the intrinsic virulence of Omicron actually increasing, or is Omicron interacting with a background of prolonged vaccine-induced spike protein production? The predominant scientific consensus, based on a wide range of studies, is that Omicron and its subvariants have generally evolved toward increased transmissibility while exhibiting decreased virulence and pathogenicity compared to earlier variants. Thus, it seems more likely that extensive mRNA vaccination is the true basis for the Omicron-related elevations in excess mortality. This argument does not necessarily exclude the possibility that some Omicron subvariants did show increased pathogenicity, in tandem with the adverse impacts of the vaccinations.
8. Discussion
Epidemiologists and public health officials often pose the following question: Which factor causes more morbidity and mortality in the general population, the COVID-19 mRNA vaccination or Omicron infections? The question sets up a dichotomy that may not be biologically tenable for any population that has been extensively injected with the modified mRNA products. This is because the spike protein, the common denominator between mRNA vaccinations and COVID-19 infections, persists for many months or years after the injection, and this sets up the potential for various kinds of interaction. In this paper, we have explored the mounting evidence of complex interactions between the mRNA vaccination and the infection. These interactions may occur whether or not the infection occurs before or after the injection. This interaction may translate into a greater risk for serious AEs (cardiac, hematologic, immunologic, and neurological AEs in particular) than either exposure alone, particularly in more susceptible populations.
Clinical misunderstanding and misreporting can arise when this interaction is overlooked in assessments of pathogenesis. For example, although it is common for clinicians to rule out viral myocarditis when assessing vaccine-related myocarditis, the opposite scenario may often be overlooked, i.e., a coronavirus-infected patient whose history of COVID-19 mRNA injections may be considered secondary or even inconsequential, due to the timeframe and lack of physician knowledge concerning the 2–3-year WOV. It is highly plausible that the myocardial stress induced by the mRNA injectable products interacts additively or synergistically with the immune-inflammatory effects of SARS-CoV-2 infection on the heart. This combined impact could exacerbate cardiac injury, potentially leading to more severe outcomes than either factor alone. If an individual contracts COVID-19 within 3 years of a booster mRNA injection, it is reasonable to assume that the myocardium may already have been primed for the mRNA products’ toxic and immune-dysregulating effects. Instead of the subsequent infection being the primary cause of myocarditis, it may be viewed as a catalyst or “effect modifier” that significantly increases the risk of myocarditis and related cardiac events. (The effect modification is as follows: the previous mRNA vaccination modifies the effect of the coronavirus infection on myocarditis risk, with stronger effects in younger males [357]). Given the widespread use of mRNA vaccines and the continued circulation of Omicron variants, such interactions may be more common than currently recognized, warranting closer clinical scrutiny and further investigation into their prevalence and mechanisms.
The hybrid harms phenomenon may help account for the continued occurrence of numerous AEs among young, previously healthy individuals, even after the emergence of the Omicron variant, which ostensibly exhibited reduced virulence on its own. As we stated before, among children and adolescents, the IFR of Omicron in 2022 was exceedingly low (0.0003%). In contrast, there was a marked elevation in the incidence of post-vaccination SAEs, including sudden cardiac death among individuals under age 40. This observed increase in risk may be attributable to what may be understood as a “multi-spike phenomenon,” whereby repeated antigenic exposure via COVID-19 mRNA vaccination and coronavirus infections contributes to direct toxicities, immune dysregulation, and diverse pathological sequelae. By these means, as we have documented, the mRNA injections predispose these otherwise very low-risk individuals to contracting the infection and also raised their risk of serious AEs or PASC-like conditions. Nevertheless, the vast majority of infants and children who received COVID-19 mRNA vaccinations and then developed severe Omicron infection and were hospitalized were counted as “hospitalized for COVID-19”, rather than acknowledging the likelihood of vaccine-related predisposition. As long as the vaccine enterprise can scapegoat COVID-19 in mRNA-injected individuals, regardless of whether they are “up to date” with their vaccinations, the simplistic narrative will continue, perpetuating the call for more vaccinations.
Negative efficacy and serious AEs associated with the COVID-19 mRNA vaccinations may stem primarily from perturbations in immune system functioning that have been linked with multiple doses over time (for a graphic explanation of the larger context surrounding COVID-19 mRNA vaccine failure, see Appendix B). From the standpoint of immunological responses to the spike protein, the equivalent of an additional “dose” would be provided by the coronavirus infection itself. As discussed in the section titled “The Hybrid Hypothesis”, repeated exposures to vaccine-induced spike protein can result in elevated IgG4 antibody levels, which favor immune tolerance over viral elimination, potentially weakening antiviral defenses and increasing the risk of infectious, autoimmune, and malignant diseases [198,204]. It is notable that this switch towards spike-specific IgG4 appears almost exclusively in individuals who received either multiple COVID-19 mRNA vaccinations or who were infected after receiving the vaccinations [208]. These differences between COVID-19 naïve and non-naïve individuals who received multiple mRNA injections, along with the observed increases in the class-switch to IgG4 in this context, are highly significant. Repeat antigenic exposures from “conventional vaccination” does not result in such class-switching.
Additionally, chronic antigenic stimulation following persistent spike protein exposure—perhaps overlapping with antigenic shift in the coronavirus—may drive T-cell exhaustion, impairing T-cell responses and increasing susceptibility to reinfection [203]. These alterations in immunological protection raise concerns about the efficacy and sustainability of ongoing administration of the mRNA products, particularly as there is a high probability that the Omicron variants will persist indefinitely, thanks to its ongoing mutations, high transmissibility, and immune evasion potential (similar to influenza strains). It is therefore imperative for humans to learn to coexist with the Omicron variants in ways that do not inadvertently increase pathogenicity. Logically, in principle, the avoidance of ongoing mRNA vaccinations would contribute substantially toward meeting that objective.
Attempting to “vaccinate” an entire population during an active infectious disease epidemic, as occurred with COVID-19 on a global scale in 2021, is biologically and epidemiologically dangerous for several reasons. First, the greatest risk of selecting for mRNA vaccine-resistant viral strains arises when a significant proportion of the population has been injected with the mRNA products, yet transmission remains largely uncontrolled: it drives mutation [358]. CDC data indicate that vaccinated individuals may carry and transmit the coronavirus at levels comparable to unvaccinated individuals [85]. Additionally, in a controlled hospital setting in Vietnam, vaccinated individuals harbored significantly higher viral loads in their nasopharynx than unvaccinated individuals [359]. This could imply greater transmissibility among BTI cases, potentially prolonging community spread and complicating herd immunity.
Second, there is a very real potential for direct mRNA vaccine spike-induced endothelial damage, pathogenic priming, autoimmune-inflammatory disease processes, and other immunopathological phenomena [360,361,362,363]. Pathogenic priming (or antibody-dependent enhancement) occurs when vaccine-induced antibodies bind to the virus with insufficient neutralizing capacity, facilitating its entry into host cells, such as macrophages, via Fc receptor-mediated pathways, potentially exacerbating viral replication and disease severity, as observed in dengue infections [364]. Individuals who undergo mRNA vaccination while infected or recently exposed may generate non-neutralizing antibodies, particularly with a novel pathogen like SARS-CoV-2, where early theoretical concerns arose due to its Fc receptor interactions [365]. Importantly, Plume et al. found higher peak antibody concentrations in individuals experiencing moderate or severe COVID-19 compared to those with milder cases, reinforcing the possibility that SARS-CoV-2 antibodies (IgG, IgA and IgE) may intensify illness severity through pathogenic priming [366]. This was the first study to show that SARS-CoV-2 triggered IgE antibody production, with blood levels rising in tandem with disease intensity, hinting at a possible connection to mast cell involvement. Multiple studies indicate that mast cells are activated in severe COVID-19, suggesting that mast cell activation syndrome may contribute to excessive inflammation and PASC-related disorders [367,368]. Moreover, there is preliminary evidence that factors affecting mast cell activation may be involved in the AEs linked with the COVID-19 mRNA vaccinations [369].
Vanden Bossche has examined in depth the immunological consequences of COVID-19 vaccination, particularly in the context of the many Omicron subvariants [370]. He notes that mRNA-based vaccinations generate transient protection through polyreactive non-neutralizing antibodies (PNNAbs) and variant-nonspecific cytotoxic T lymphocytes (CTLs); however, these effects are short-lived and may not confer durable defense against emerging immune escape variants. Since Omicron’s emergence, viral neutralization has become less dependent on highly antigen-specific antibodies, and cross-reactive cytolytic activity for antigen-specific CD8+ T cells has not been demonstrated. The mRNA vaccine-related BTIs primarily induce PNNAbs and CTLs, temporarily mitigating severe disease but failing to establish long-term immunity. Vanden Bossche’s concept of steric immune refocusing suggests that the updated modified mRNA products and BTIs promote the production of broadly cross-reactive but sub-neutralizing antibodies, applying suboptimal immune pressure on viral evolution. These antibodies exhibit diminished antigen specificity and binding affinity, facilitating transient CTL-mediated viral clearance but failing to provide consistent neutralization. This process fosters immune selection pressure, favoring the emergence of highly infectious, immune-evasive variants, such as the Omicron subvariant JN.1, with increased resistance to neutralizing antibodies and enhanced replication capacity.
To summarize the biological mechanism-based rationale for the Hybrid Harms perspective, when neutralizing antibodies are relatively diminished and non-neutralizing antibodies are overabundant, the incidence of severe COVID-19 may become substantially increased in populations with extensive mRNA vaccination coverage. Moreover, as we and others have discussed in previous publications, these immunological processes may collectively increase the likelihood of various inflammatory conditions, such as cancers, cardiovascular disorders, and various other diseases rooted in chronic inflammation and immune dysfunction [193,267,371]. Immune cross-reactivity between anti-SARS-CoV-2 antibodies and human tissue antigens is likely due to molecular mimicry [372]. Vojdani et al. identified reactions between SARS-CoV-2 antibodies and 28 of 55 human tissue antigens, spanning barrier proteins, gastrointestinal, thyroid, and neural tissues [373]. The homologous peptides in the translated spike protein from the mRNA injections are 100% matched to many human proteins. The extensive cross-reactivity resulting from these homologies may contribute to the multisystem pathology of COVID-19, influence disease severity, and potentially trigger or exacerbate autoimmunity in susceptible individuals. These findings highlight a potential risk for autoimmune and multisystem disorders in COVID-19, driven by cross-reactivity between SARS-CoV-2 proteins and human tissues.
The epidemiological implications of this integrated perspective are substantial. Temporal causal reasoning, the tendency to link an outcome to the most recent or obvious preceding event, is a standard approach in both clinical practice and epidemiological studies. Against a backdrop of extensive COVID-19 vaccinations (70-80% coverage for most populations in developed countries), the coronavirus infection appears temporally proximal to the serious AE, making it a natural suspect as the presumed cause of the disease event or condition in question. Anyone with a strong bias toward the “safe and effective” vaccination narrative would be inclined to blame the infection as the exclusive cause, even if the mRNA inoculation had set the biological stage. Many of the diseases and disabilities linked with COVID-19 in previously vaccinated individuals may have been misattributed solely to the virus, overlooking the contribution of prolonged vaccine-induced spike protein, thus complicating the attribution of causality and highlighting the need for integrated risk assessments.
Most of the published case reports on COVID-19-associated cardiac and thrombotic events only provide details on the infection, omitting the individuals’ vaccination history. The obvious temporal association between SARS-CoV-2 infection and serious AEs has likely led to systematic misclassification, attributing causality solely to the viral infection rather than considering the potential interaction between prior mRNA vaccination and post-vaccination infection. By failing to consider the latter, the interpretation of these studies may be distorting the perceived relative risk of the events following infection versus vaccination. In most cases, this leads to an underestimation of the mRNA vaccination’s role in predisposing individuals to those serious AEs. Recognizing and studying the infection-injection interaction is crucial for improving public health recommendations, accurately assessing product safety though properly designed epidemiological studies, and ultimately ensuring that individuals at higher risk receive appropriate care and surveillance.
In a 5 June 2022 letter to the Virology Journal editor titled “Adverse effects of COVID-19 vaccines and measures to prevent them,” cardiologist Dr. Kenji Yamamoto stated, “As a safety measure, further booster vaccinations should be discontinued. In addition, the date of vaccination and the time since the last vaccination should be recorded in the medical record of patients” [360]. This advice was remarkably prescient. Most healthcare professionals do not consider the likelihood that the prolonged spike protein production elicited by the COVID-19 vaccinations is increasing the risk of many diseases that might otherwise never have manifested in susceptible individuals. As a matter of routine, the medical community should be taking histories on both COVID-19 vaccination and Omicron variant infection whenever assessing cardiovascular and clotting issues, just as a smoking history has become a routine part of all medical histories since the 1960s. Knowing the dates of the COVID-19 mRNA injections and coronavirus infections could provide valuable insights into the onset and progression of many contemporary diseases and disabilities. In addition, patients with a history of stroke, myocardial infarction, or pulmonary embolism who contract a COVID-19 infection and/or undergo an mRNA vaccination must be closely monitored for possible embolic events, as they are at increased risk of such events [374].
Given the very low IFR for the vast majority of people who contract COVID-19, we believe that more attention should be given to educating the general public around self-care practices for optimizing natural immunity (e.g., nutrition, exercise and stress management). These practices should be considered as viable public health strategies for minimizing the risk of COVID-19. Observational studies suggest that, even in the short term, natural immunity may confer equal or greater protection against SARS-CoV-2 infections compared to individuals receiving the two-dose COVID-19 mRNA vaccinations [250]. In these studies, natural immunity showed roughly similar effect sizes regarding protection against reinfection across different SARS-CoV-2 variants. Risk of hospitalizations and deaths was also reduced in SARS-CoV-2 reinfections versus primary infections, though the data were not consistent. The combination of a previous SARS-CoV-2 infection and a respective vaccination (hybrid immunity) seemed to confer the strongest protection against SARS-CoV-2 infections. As we have established, however, the theoretical advantage in protection afforded by such “enhanced immunity” over a short timeframe may be substantially counterbalanced by the large number of serious AEs that correlate strongly with the additional antibody production associated with hybrid immunity. This last point forms the crux of the Hybrid Harms Hypothesis.
As of 2025, the new FDA leadership recommends placebo-controlled trials for COVID-19 vaccines, using saline placebos to document AE profiles [375]. FDA guidance also states that such controls can distinguish AEs and outcomes specifically caused by the vaccine from those due to underlying disease or “background noise” [376]. However, the persistence of synthetic mRNA and vaccine-associated spike protein in some individuals may exceed typical background noise, potentially confounding the assessment of efficacy and safety [377]. Comprehensive pharmacokinetic data, including absorption, distribution, metabolism, and excretion, are critical for understanding mRNA vaccine behavior, yet such data remain incomplete [378]. Without obtaining such a robust pharmacokinetic profile, defining a true control group is challenging. Indeed, the assumption that a saline placebo group represents a neutral baseline may be flawed given the possibility of residual spike protein potentially influencing physiological responses and interacting with mRNA vaccine-induced spike protein during the trial. Establishing a “washout” period for the body’s clearance of synthetic mRNA and spike protein is essential to ensure valid trial outcomes and accurate attribution of AEs and efficacy. This washout period should be a minimum of three years.
Despite the fact that the COVID-19 mRNA vaccinations are associated with so many of the same AEs or disease outcomes that have been linked with the coronavirus infections, public health scientists and policymakers continually downplay the effects of the vaccines while emphasizing the risks of infection. Moreover, these publications invariably make the claim that “the benefits of COVID-19 vaccination far outweigh the potential risks” and encourage people to continue getting vaccinated. Whether due to editorial policy or to academic pressures with mandates still in place for some academic institutions, or both, the authors and their editors consistently refuse to question the safety profile of these genetic vaccines, even after providing definitive substantiation of their serious adverse impacts.
If we accept official government narratives that the excess mortality waves in the most extensively vaccinated countries in 2022 were driven primarily by Omicron outbreaks, there are several possible explanations. The first is that the COVID-19 vaccinations targeting the Omicron were unable to reduce the risk of infection, hospitalization and death, as reported by Ioannou et al., for the XBB.1.5 subvariant [114]. However, this would not sufficiently explain the elevations in all-cause mortality in the most extensively vaccinated countries. The second explanation, consistent with our Hybrid Harms Hypothesis, entails an interaction between the mRNA vaccinations and subsequent Omicron infection: Serious morbidity and mortality events attributed to Omicron infections in highly vaccinated populations were primarily driven by the long-term persistence of spike protein resulting from previous mRNA vaccinations. This prolonged expression may have engendered the false appearance of temporally causation tied exclusively to the viral infection, when in fact the disease and disability outcomes were more plausibly due to an interaction between Omicron infection and prior exposure to the modified mRNA injectables. The increased production of counterproductive IgG4 antibodies following repeated vaccination likely also contributed to the excess mortality statistics.
9. Conclusions
Our “Hybrid Harms Hypothesis” proposes that interactions between COVID-19 mRNA injections and later coronavirus infections may explain the manifestation and/or persistence of serious AEs in previously vaccinated individuals, even after the emergence of milder Omicron variants. In many cases, the biological impact of COVID-19 mRNA vaccination may constitute a precursor event, predisposing the individual to develop the post-COVID-19 sequelae (see Table 1) at any time during the post-injection 3-year window. Coronavirus infections may amplify the adverse effects of previous COVID-19 vaccinations over the course of years rather than months. A plausible explanation for the prolonged presence of spike protein, along with the multitude of long-term “spikeopathies” that have been documented in the context of the mRNA-LNP modality, is integration of the product’s genetic material into the genome of human cells. This latent integration would be a byproduct of the unintended plasmid-sourced foreign DNA, including the mammalian promoter SV40. If there were pockets of cells latently infected with this foreign genetic material, it could manifest as persistent spike protein production when those cells become active. It seems improbable that the persistence of spike protein production could be due to prolonged mRNA stability or protein retention in immune cells because of the years-long timeframe between introduction to the material and production itself.
Most of the administration of these synthetic, modified mRNA products took place in 2021. By 2022, Omicron had become the dominant COVID-19 viral variant, with mild pathogenicity. Even the most extensively vaccinated countries experienced substantial Omicron outbreaks in 2022. Paradoxically, despite the mild pathogenicity of this variant, the waves of Omicron infection were closely followed by spikes in all-cause mortality. Many governments attributed mortality spikes to COVID-19, even though non-COVID deaths typically accounted for a larger share of total mortality. These government bodies also failed to take into account the likely background impact of extensive vaccinations (80-90% coverage by 2022). We present the hypothesis that the infections were superimposed on a preexisting mRNA vaccine-induced milieu of toxic spike protein, inflammatory lipid nanoparticles, and residual process-related DNA impurities. As an example, subclinical myocarditis may be exacerbated as persistent vaccine-derived spike protein in cardiac tissues, combined with subsequent exposure to infection-induced spike protein, triggers hyperimmune responses, leading to myocyte damage and clinical myocarditis, along with sequelae like arrhythmias or heart failure. Spike proteins resulting from both the mRNA vaccination and the natural SARS-CoV-2 infection have been shown to persist for extended periods, raising parallel concerns regarding potential implications for long-term safety. Many if not most of the morbidity and mortality events attributed to COVID-19 in heavily vaccinated populations in 2022-2023 were likely due to the long-term background persistence of spike protein and other vaccine-associated components resulting from previous COVID-19 mRNA vaccinations.
Various immunological mechanisms (notably IgG4 class switch, T-cell exhaustion, and pathogenic priming due to excessive production of non-neutralizing antibodies) appear to account for the well-documented increase in susceptibility to coronavirus infections following repeated administration of the mRNA products. These same post-vaccination infections are more likely to trigger clinically significant health problems if the individual received a mRNA injection within the preceding 2-3 years (or longer). The appearance of a strong temporal relationship with COVID-19 (or a weak one based solely on incidental RT-PCR test findings) often leads to misclassification of the infection as the primary cause of the myocarditis and other serious cardiac events. It is biologically plausible that the Hybrid Harms Hypothesis helps explain the increased incidences of myocarditis, myocardial infarction and hemorrhagic strokes that have been observed in younger adults (<50 years old). Spike protein from either the infection or the mRNA vaccination, or both, has been found in the myocardium and in the (burst) blood vessels in the brain, thus reinforcing causation in both scenarios. Lastly, the persistence of both synthetic mRNA and mRNA product-associated spike protein in some individuals raises questions about whether these elements contribute to a level of physiological activity that exceeds typical background noise, potentially confounding safety and efficacy assessments.
The present bio-epidemiological inquiry reflects a progressive shift in understanding of the clearance of the spike protein from the body, from short-term expression to prolonged persistence. Changes in long-term persistence of the modified mRNA-encoded spike protein challenge earlier assumptions about the COVID-19 mRNA products’ kinetics and the biological implications thereof, helping to inform a new perspective on the potential impacts of the mRNA vaccinations and subsequent infections. Natural immunity, resulting in part from prior infection with respiratory viral pathogens, may well provide a superior degree of protective immunity tailored to the specific pathogen. This is due to the comprehensive immune response elicited, as well as the absence of serious AEs that have been linked with the COVID-19 mRNA vaccinations, either with or without the coronavirus infections.
Acknowledgments
We thank vaccinologist Geert vanden Bossche for insights regarding immunological aspects of the COVID-19 mRNA vaccinations. Additional thanks to Scott Sutton of the COVID Index team (https://covidindex.science) for assistance with adapting the graphics from Johns Hopkins University Coronavirus Resource Center.
Conflicts of Interest
The authors declare that they have no competing interests.
Appendix A. Literature Sources for Table 2
General
Pfizer. (2021). 5.3.6 Cumulative analysis of post-authorization adverse event reports of PF-07302048 (BNT162b2) received through 28-Feb-2021 (reissued). https://www.phmpt.org/wp-content/uploads/2022/04/reissue_5.3.6-postmarketing-experience.pdf
PASC
Bowe B, Xie Y, Al-Aly Z. Postacute sequelae of COVID-19 at 2 years. Nat Med. 2023;29(9):2347-2357. doi: 10.1038/s41591-023-02521-2.
Cheng KL, Yu WS, Wang YH, Ibarburu GH, Lee HL, Wei JC. Long-term Thyroid Outcomes After COVID-19 Vaccination: A Cohort Study of 2,333,496 Patients from the TriNetX Network. J Clin Endocrinol Metab. 2025: dgaf064. doi: 10.1210/clinem/dgaf064
Autoimmune
Sumantri S, Rengganis I. Immunological dysfunction and mast cell activation syndrome in long COVID. Asia Pac Allergy. 2023;13(1):50-53. doi: 10.5415/apallergy.0000000000000022.
Cardiovascular
Zhang T, Li Z, Mei Q, Walline JH, Zhang Z, Liu Y, Zhu H, Du B. Cardiovascular outcomes in long COVID-19: a systematic review and meta-analysis. Front Cardiovasc Med. 2025;12:1450470. doi: 10.3389/fcvm.2025.1450470.
Otolarygnological
Alzahrani M, Alshathri AH, Alduraibi K, Alshathri AH, Alanazi TF, Alandijani H, Almajed JA, Wajdi KA. Long-Term Outcomes of COVID-19 Otolaryngology Symptoms in Saudi Arabia. Cureus. 2023;15(1):e33461. doi: 10.7759/cureus.33461. PMID: 36751176; PMCID: PMC9899518.
Dermatology
Gambichler T, Boms S, Susok L, Dickel H, Finis C, Abu Rached N, Barras M, Stücker M, Kasakovski D. Cutaneous findings following COVID-19 vaccination: review of world literature and own experience. J Eur Acad Dermatol Venereol. 2022;36(2):172-180. doi: 10.1111/jdv.17744.
Endocrine
Kazakou P, Paschou SA, Psaltopoulou T, Gavriatopoulou M, Korompoki E, Stefanaki K, Kanouta F, Kassi GN, Dimopoulos MA, Mitrakou A. Early and late endocrine complications of COVID-19. Endocr Connect. 2021;10(9):R229-R239. doi: 10.1530/EC-21-0184
Gynecological/reproductive
Pollack B, von Saltza E, McCorkell L, Santos L, Hultman A, Cohen AK, Soares L. Female reproductive health impacts of Long COVID and associated illnesses including ME/CFS, POTS, and connective tissue disorders: a literature review. Front Rehabil Sci. 2023;4:1122673. doi: 10.3389/fresc.2023.1122673
Sandoval MN, Klawans MR, Bach MA, Mikhail J, Graviss EA, Cao T, Parchem JG, Husain J, Boerwinkle E. COVID-19 infection history as a risk factor for early pregnancy loss: results from the electronic health record-based Southeast Texas COVID and Pregnancy Cohort Study. BMC Med. 2025 May 9;23(1):274. doi: 10.1186/s12916-025-04094-y.
Manniche V, Fürst T, Schmeling M, Gilthorpe JD, Hansen PR. Rates of successful conceptions according to COVID-19 vaccination status: Data from the Czech Republic. Int J Risk Saf Med. 2025 Jun 19:9246479251353384. doi: 10.1177/09246479251353384
Thorp JA, Rogers C, Deskevich MP, Tankersley S, Benavides A, Redshaw MD, McCullough PA. COVID-19 vaccines: The impact on pregnancy outcomes and menstrual function. J Am Phys Surg. 2023; 28(1). https://www.jpands.org/vol28no1/thorp.pdf
Madaan S, Talwar D, Jaiswal A, Kumar S, Acharya N, Acharya S, Dewani D. Post-COVID-19 menstrual abnormalities and infertility: Repercussions of the pandemic. J Educ Health Promot. 2022;11:170. doi: 10.4103/jehp.jehp_1200_21
Figure A1.
The basis for the failure of COVID-19 mRNA vaccinations has been elucidated. The SARS-CoV-2 spike (S) protein interacts with ACE2 receptor on host cells, creating conditions that exert significant selective pressure on the S gene, driving mutations that facilitate viral immune escape. Most of these mRNA products were developed using the S protein sequence from the original Wuhan strain, rendering them less effective against these escape variants, as the mutants can circumvent the immune responses elicited by these vaccines. This results in diminished vaccine efficacy for formulations based on the original S protein sequence. Furthermore, repeated mRNA vaccinations may alter viral dynamics, potentially promoting the emergence of immune-evasive variants, which could further reduce the effectiveness of these biologics. Additionally, frequent booster doses may contribute to excessive spike-related toxicities as well as immune dysregulation, potentially compromising antiviral and antimicrobial immunity while increasing the risk of autoimmune conditions and accelerated oncogenesis.
Figure A1.
The basis for the failure of COVID-19 mRNA vaccinations has been elucidated. The SARS-CoV-2 spike (S) protein interacts with ACE2 receptor on host cells, creating conditions that exert significant selective pressure on the S gene, driving mutations that facilitate viral immune escape. Most of these mRNA products were developed using the S protein sequence from the original Wuhan strain, rendering them less effective against these escape variants, as the mutants can circumvent the immune responses elicited by these vaccines. This results in diminished vaccine efficacy for formulations based on the original S protein sequence. Furthermore, repeated mRNA vaccinations may alter viral dynamics, potentially promoting the emergence of immune-evasive variants, which could further reduce the effectiveness of these biologics. Additionally, frequent booster doses may contribute to excessive spike-related toxicities as well as immune dysregulation, potentially compromising antiviral and antimicrobial immunity while increasing the risk of autoimmune conditions and accelerated oncogenesis.
References
- Ejaz, H.; Alsrhani, A.; Zafar, A.; Javed, H.; Junaid, K.; Abdalla, A.E.; Abosalif, K.O.; Ahmed, Z.; Younas, S. COVID-19 and comorbidities: Deleterious impact on infected patients. J. Infect. Public Heal. 2020, 13, 1833–1839. [Google Scholar] [CrossRef]
- Zuin, M.; Rigatelli, G.; Zuliani, G.; Rigatelli, A.; Mazza, A.; Roncon, L. Arterial hypertension and risk of death in patients with COVID-19 infection: Systematic review and meta-analysis. J. Infect. 2020, 81, e84–e86. [Google Scholar] [CrossRef] [PubMed]
- Cariou, B.; for the CORONADO investigators; Hadjadj, S. ; Wargny, M.; Pichelin, M.; Al-Salameh, A.; Allix, I.; Amadou, C.; Arnault, G.; Baudoux, F.; et al. Phenotypic characteristics and prognosis of inpatients with COVID-19 and diabetes: the CORONADO study. Diabetologia 2020, 63, 1500–1515. [Google Scholar] [CrossRef]
- Lazcano, U.; Cuadrado-Godia, E.; Grau, M.; Subirana, I.; Martínez-Carbonell, E.; Boher-Massaguer, M.; Rodríguez-Campello, A.; Giralt-Steinhauer, E.; Fernández-Pérez, I.; Jiménez-Conde, J.; et al. Increased COVID-19 Mortality in People With Previous Cerebrovascular Disease: A Population-Based Cohort Study. Stroke 2022, 53, 1276–1284. [Google Scholar] [CrossRef]
- Ge, E.; Li, Y.; Wu, S.; Candido, E.; Wei, X.; Augusto, O. Association of pre-existing comorbidities with mortality and disease severity among 167,500 individuals with COVID-19 in Canada: A population-based cohort study. PLOS ONE 2021, 16, e0258154. [Google Scholar] [CrossRef]
- Phelps, M.; Christensen, D.M.; Gerds, T.; Fosbøl, E.; Torp-Pedersen, C.; Schou, M.; Køber, L.; Kragholm, K.; Andersson, C.; Biering-Sørensen, T.; et al. Cardiovascular comorbidities as predictors for severe COVID-19 infection or death. Eur. Hear. J. - Qual. Care Clin. Outcomes 2020, 7, 172–180. [Google Scholar] [CrossRef]
- Suleyman, G.; Fadel, R.A.; Malette, K.M.; Hammond, C.; Abdulla, H.; Entz, A.; Demertzis, Z.; Hanna, Z.; Failla, A.; Dagher, C.; et al. Clinical Characteristics and Morbidity Associated With Coronavirus Disease 2019 in a Series of Patients in Metropolitan Detroit. JAMA Netw. Open 2020, 3, e2012270. [Google Scholar] [CrossRef]
- Ioannou, G.N.; Locke, E.; Green, P.; Berry, K.; O’hAre, A.M.; Shah, J.A.; Crothers, K.; Eastment, M.C.; Dominitz, J.A.; Fan, V.S. Risk Factors for Hospitalization, Mechanical Ventilation, or Death Among 10 131 US Veterans With SARS-CoV-2 Infection. JAMA Netw. Open 2020, 3, e2022310–e2022310. [Google Scholar] [CrossRef]
- Witkowski, J.M.; Fulop, T.; Bryl, E. Immunosenescence and COVID-19. Mech. Ageing Dev. 2022, 204, 111672–111672. [Google Scholar] [CrossRef]
- Fang, X.; Wang, J.; Liu, Y.; Zhang, X.; Wang, T.; Zhang, H.; Liang, Z.; Luo, F.; Li, W.; Liu, D.; et al. Combined and interactive effects of alcohol drinking and cigarette smoking on the risk of severe illness and poor clinical outcomes in patients with COVID-19: a multicentre retrospective cohort study. Public Heal. 2022, 205, 6–13. [Google Scholar] [CrossRef]
- Prodromos, C.; Rumschlag, T. Hydroxychloroquine is effective, and consistently so when provided early, for COVID-19: a systematic review. New Microbes New Infect. 2020, 38, 100776–100776. [Google Scholar] [CrossRef] [PubMed]
- Kory P, McCarthy J. War on Ivermectin: The Medicine that Saved Millions and Could Have Ended the Pandemic. Skyhorse Publishing, NY. 2023. https://www.amazon.com/War-Ivermectin-Medicine-Millions-Pandemic/dp/151077386X.
- Malhotra, A. Curing the pandemic of misinformation on COVID-19 mRNA vaccines through real evidence-based medicine - Part 1. J. Metab. Heal. 2022, 5. [Google Scholar] [CrossRef]
- Quinn, G.A.; Connolly, R.; Óhaiseadha, C.; Hynds, P.; Bagus, P.; Brown, R.B.; Cáceres, C.F.; Craig, C.; Connolly, M.; Domingo, J.L.; et al. What Lessons can Be Learned From the Management of the COVID-19 Pandemic? Int. J. Public Heal. 2025, 70, 1607727. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, A. Curing the pandemic of misinformation on COVID-19 mRNA vaccines through real evidence-based medicine - Part 2. J. Metab. Heal. 2022, 5, 10. [Google Scholar] [CrossRef]
- Santin, A.; Scheim, D.; McCullough, P.; Yagisawa, M.; Borody, T. Ivermectin: a multifaceted drug of Nobel prize-honoured distinction with indicated efficacy against a new global scourge, COVID-19. New Microbes New Infect. 2021, 43, 100924. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and efficacy of the BNT162b2 mRNA COVID-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- El Sahly, H.M.; Baden, L.R.; Essink, B.; Doblecki-Lewis, S.; Martin, J.M.; Anderson, E.J.; Campbell, T.B.; Clark, J.; Jackson, L.A.; Fichtenbaum, C.J.; et al. Efficacy of the mRNA-1273 SARS-CoV-2 Vaccine at Completion of Blinded Phase. New Engl. J. Med. 2021, 385, 1774–1785. [Google Scholar] [CrossRef]
- Julian Gillespie, a prominent Australian barrister leading legal challenges against the regulatory approval of COVID-19 mRNA “vaccines”, argues that they should be classified as gene therapies and also genetically modified organisms (GMOs) under Australian law, specifically the Gene Technology Act 2000, which was bypassed during the EUA process. The Gene Technology Act 2000 requires specific gene-based risk assessments and licensing. Similar U.S. efforts to nullify the Biologics License Application contend the FDA’s waiver of National Environmental Policy Act assessments concealed their gene therapy status, evading proper scrutiny. These gene transfer technologies demand a distinct regulatory approval process; without adherence, their sale is illegal.
- Heinz, F.X.; Stiasny, K. Distinguishing features of current COVID-19 vaccines: knowns and unknowns of antigen presentation and modes of action. npj Vaccines 2021, 6, 1–13. [Google Scholar] [CrossRef]
- Oldfield PR, Gutschi M, McCullough PA, Speicher DJ. BioNTech’s COVID-19 modRNA Vaccines: Dangerous genetic mechanism of action released before sufficient preclinical testing. Journal of American Physicians and Surgeons. 2024;29(4):118-126.
- Cosentino, M.; Marino, F. Understanding the Pharmacology of COVID-19 mRNA Vaccines: Playing Dice with the Spike? Int. J. Mol. Sci. 2022, 23, 10881. [Google Scholar] [CrossRef]
- Chaudhary, J.K.; Yadav, R.; Chaudhary, P.K.; Maurya, A.; Kant, N.; Al Rugaie, O.; Haokip, H.R.; Yadav, D.; Roshan, R.; Prasad, R.; et al. Insights into COVID-19 Vaccine Development Based on Immunogenic Structural Proteins of SARS-CoV-2, Host Immune Responses, and Herd Immunity. Cells 2021, 10, 2949. [Google Scholar] [CrossRef]
- Painter, M.M.; Mathew, D.; Goel, R.R.; Apostolidis, S.A.; Pattekar, A.; Kuthuru, O.; Baxter, A.E.; Herati, R.S.; Oldridge, D.A.; Gouma, S.; et al. Rapid induction of antigen-specific CD4+ T cells is associated with coordinated humoral and cellular immunity to SARS-CoV-2 mRNA vaccination. Immunity 2021, 54, 2133–2142.e3. [Google Scholar] [CrossRef]
- Laczkó, D.; Hogan, M.J.; Toulmin, S.A.; Hicks, P.; Lederer, K.; Gaudette, B.T.; Castaño, D.; Amanat, F.; Muramatsu, H.; Oguin, T.H.; et al. A Single Immunization with Nucleoside-Modified mRNA Vaccines Elicits Strong Cellular and Humoral Immune Responses against SARS-CoV-2 in Mice. Immunity 2020, 53, 724–732.e7. [Google Scholar] [CrossRef]
- Rustagi, V.; Gupta, S.R.R.; Talwar, C.; Singh, A.; Xiao, Z.-Z.; Jamwal, R.; Bala, K.; Bhaskar, A.K.; Nagar, S.; Singh, I.K. SARS-CoV-2 pathophysiology and post-vaccination severity: a systematic review. Immunol. Res. 2024, 73, 17. [Google Scholar] [CrossRef]
- Du, J.; Lang, H.-M.; Ma, Y.; Chen, A.-W.; Qin, Y.-Y.; Zhang, X.-P.; Huang, C.-Q.; Du, \. Global trends in COVID-19 incidence and case fatality rates (2019–2023): a retrospective analysis. Front. Public Heal. 2024, 12, 1355097. [Google Scholar] [CrossRef] [PubMed]
- Binnicker, M.J.; Kraft, C.S. Challenges and Controversies to Testing for COVID-19. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef] [PubMed]
- Pujadas, E.; Chaudhry, F.; McBride, R.; Richter, F.; Zhao, S.; Wajnberg, A.; Nadkarni, G.; Glicksberg, B.S.; Houldsworth, J.; Cordon-Cardo, C. SARS-CoV-2 viral load predicts COVID-19 mortality. Lancet Respir. Med. 2020, 8, e70. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, T.; A Spencer, E.; Brassey, J.; Heneghan, C. Viral Cultures for Coronavirus Disease 2019 Infectivity Assessment: A Systematic Review. Clin. Infect. Dis. 2020, 73, e3884–e3899. [Google Scholar] [CrossRef]
- Mallett, S.; Allen, A.J.; Graziadio, S.; Taylor, S.A.; Sakai, N.S.; Green, K.; Suklan, J.; Hyde, C.; Shinkins, B.; Zhelev, Z.; et al. At what times during infection is SARS-CoV-2 detectable and no longer detectable using RT-PCR-based tests? A systematic review of individual participant data. BMC Med. 2020, 18, 1–17. [Google Scholar] [CrossRef]
- Kulkarni, D.; Lee, B.; Ismail, N.F.; Rahman, A.E.; Spinardi, J.; Kyaw, M.H.; Nair, H. Incidence, severity, risk factors and outcomes of SARS-CoV-2 reinfections during the Omicron period: a systematic review and meta-analysis. J. Glob. Heal. 2025, 15, 04032. [Google Scholar] [CrossRef]
- Mohapatra, R.K.; Sarangi, A.K.; Kandi, V.; Azam, M.; Tiwari, R.; Dhama, K. Omicron (B.1.1.529 variant of SARS-CoV-2); an emerging threat: Current global scenario. J. Med Virol. 2022, 94, 1780–1783. [Google Scholar] [CrossRef]
- Karyakarte, R.P.; Das, R.; Dudhate, S.; Agarasen, J.; Pillai, P.; Chandankhede, P.M.; Labhshetwar, R.S.; Gadiyal, Y.; Rajmane, M.V.; Kulkarni, P.P.; et al. Clinical Characteristics and Outcomes of Laboratory-Confirmed SARS-CoV-2 Cases Infected With Omicron Subvariants and the XBB Recombinant Variant. Cureus 2023, 15, e35261. [Google Scholar] [CrossRef] [PubMed]
- Tureček, P.; Kleisner, K. Symptomic Mimicry Between SARS-CoV-2 and the Common Cold Complex. Biosemiotics 2022, 15, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Joung, S.Y.; Ebinger, J.E.; Sun, N.; Liu, Y.; Wu, M.; Tang, A.B.; Prostko, J.C.; Frias, E.C.; Stewart, J.L.; Sobhani, K.; et al. Awareness of SARS-CoV-2 Omicron Variant Infection Among Adults With Recent COVID-19 Seropositivity. JAMA Netw. Open 2022, 5, e2227241–e2227241. [Google Scholar] [CrossRef] [PubMed]
- Meng, B.; Abdullahi, A.; Ferreira, I.A.T.M.; Goonawardane, N.; Saito, A.; Kimura, I.; Yamasoba, D.; Gerber, P.P.; Fatihi, S.; Rathore, S.; et al. Altered TMPRSS2 usage by SARS-CoV-2 Omicron impacts infectivity and fusogenicity. Nature 2022, 603, 706–714. [Google Scholar] [CrossRef]
- Lorenzo-Redondo, R.; A Ozer, E.; Hultquist, J.F. Covid-19: is omicron less lethal than delta? BMJ 2022, 378, o1806. [Google Scholar] [CrossRef]
- Christie, B. Covid-19: Early studies give hope omicron is milder than other variants. BMJ 2021, 375, n3144. [Google Scholar] [CrossRef]
- Zhao, H.; Lu, L.; Peng, Z.; Chen, L.-L.; Meng, X.; Zhang, C.; Ip, J.D.; Chan, W.-M.; Chu, A.W.-H.; Chan, K.-H.; et al. SARS-CoV-2 Omicron variant shows less efficient replication and fusion activity when compared with Delta variant in TMPRSS2-expressed cells. Emerg. Microbes Infect. 2022, 11, 277–283. [Google Scholar] [CrossRef]
- Mathieu E, Ritchie H, Rodés-Guirao L, Appel C, Giattino C, Hasell J, Macdonald B, et al. Coronavirus Pandemic (COVID-19). 2020. Available: https://ourworldindata.org/coronavirus. Accessed 30 April 2024.
- Abdullah, F.; Myers, J.; Basu, D.; Tintinger, G.; Ueckermann, V.; Mathebula, M.; Ramlall, R.; Spoor, S.; de Villiers, T.; Van der Walt, Z.; et al. Decreased severity of disease during the first global omicron variant covid-19 outbreak in a large hospital in tshwane, south africa. Int. J. Infect. Dis. 2021, 116, 38–42. [Google Scholar] [CrossRef]
- Maslo, C.; Friedland, R.; Toubkin, M.; Laubscher, A.; Akaloo, T.; Kama, B. Characteristics and Outcomes of Hospitalized Patients in South Africa During the COVID-19 Omicron Wave Compared With Previous Waves. JAMA 2021, 327, 583–584. [Google Scholar] [CrossRef]
- Dyer, O. COVID-19: South Africa’s surge in cases deepens alarm over omicron variant. BMJ. 2021;375:n3013.
- Jassat, W.; Karim, S.S.A.; Ozougwu, L.; Welch, R.; Mudara, C.; Masha, M.; Rousseau, P.; Wolmarans, M.; Selikow, A.; Govender, N.; et al. Trends in Cases, Hospitalizations, and Mortality Related to the Omicron BA.4/BA.5 Subvariants in South Africa. Clin. Infect. Dis. 2022, 76, 1468–1475. [Google Scholar] [CrossRef]
- Adjei, S.; Hong, K.; Molinari, N.-A.M.; Bull-Otterson, L.; Ajani, U.A.; Gundlapalli, A.V.; Harris, A.M.; Hsu, J.; Kadri, S.S.; Starnes, J.; et al. Mortality Risk Among Patients Hospitalized Primarily for COVID-19 During the Omicron and Delta Variant Pandemic Periods — United States, April 2020–June 2022. Mmwr-Morbidity Mortal. Wkly. Rep. 2022, 71, 1182–1189. [Google Scholar] [CrossRef]
- Pather, S.; Madhi, S.A.; Cowling, B.J.; Moss, P.; Kamil, J.P.; Ciesek, S.; Muik, A.; Türeci, Ö. SARS-CoV-2 Omicron variants: burden of disease, impact on vaccine effectiveness and need for variant-adapted vaccines. Front. Immunol. 2023, 14, 1130539. [Google Scholar] [CrossRef]
- Sharif, N.; Alzahrani, K.J.; Ahmed, S.N.; Dey, S.K. Efficacy, Immunogenicity and Safety of COVID-19 Vaccines: A Systematic Review and Meta-Analysis. Front. Immunol. 2021, 12, 714170. [Google Scholar] [CrossRef]
- Zheng, C.; Shao, W.; Chen, X.; Zhang, B.; Wang, G.; Zhang, W. Real-world effectiveness of COVID-19 vaccines: a literature review and meta-analysis. Int. J. Infect. Dis. 2021, 114, 252–260. [Google Scholar] [CrossRef]
- Excess Mortality Project. Excess mortality calculations for different countries. Phinance Technologies. Accessed 7/18/2025. URL: https://phinancetechnologies.com/HumanityProjects/Projects.htm.
- Kuhbandner, C.; Reitzner, M. Estimation of Excess Mortality in Germany During 2020-2022. Cureus 2023, 15, e39371. [Google Scholar] [CrossRef]
- Aarstad J, Kvitastein OA. Is there a link between the 2021 COVID-19 vaccination uptake in Europe and 2022 excess all-cause mortality? Asian Pac. J. Health Sci. 2023;10(1):25-31. https://www.apjhs.com/index.php/apjhs/article/view/3017/1610.
- Economidou, E.C.; Soteriades, E.S. Excess mortality in Cyprus during the COVID-19 vaccination campaign. Vaccine 2023, 42, 3375–3376. [Google Scholar] [CrossRef]
- Raknes, G.; Fagerås, S.J.; Sveen, K.A.; Júlíusson, P.B.; Strøm, M.S. Excess non-COVID-19 mortality in Norway 2020–2022. BMC Public Heal. 2024, 24, 1–13. [Google Scholar] [CrossRef]
- Mostert, S.; Hoogland, M.; Huibers, M.; Kaspers, G. Excess mortality across countries in the Western World since the COVID-19 pandemic: ‘Our World in Data’ estimates of January 2020 to December 2022. BMJ Public Heal. 2024, 2, e000282. [Google Scholar] [CrossRef]
- Shir-Raz, Y.; Elisha, E.; Martin, B.; Ronel, N.; Guetzkow, J. Censorship and Suppression of Covid-19 Heterodoxy: Tactics and Counter-Tactics. Minerva 2022, 61, 407–433. [Google Scholar] [CrossRef]
- This efficacy was based solely on relative risk reductions. The absolute risk reduction was only about 1%. Specifically, the absolute risk reductions for BNT162b2 and mRNA-1273 were 0.7% and 1.1%, respectively.
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Levin, E.G.; Lustig, Y.; Cohen, C.; Fluss, R.; Indenbaum, V.; Amit, S.; Doolman, R.; Asraf, K.; Mendelson, E.; Ziv, A.; et al. Waning Immune Humoral Response to BNT162b2 Covid-19 Vaccine over 6 Months. New Engl. J. Med. 2021, 385, E84–E84. [Google Scholar] [CrossRef]
- Haq MA, Roy AK, Ahmed R, Kuddusi RU, Sinha M, Hossain MS, Vandenent M, et al. Antibody longevity and waning following COVID-19 vaccination in a 1-year longitudinal cohort in Bangladesh. Sci Rep. 2024;14(1):11467. [CrossRef]
- Srivastava, K.; Carreño, J.M.; Gleason, C.; Monahan, B.; Singh, G.; Abbad, A.; Tcheou, J.; Raskin, A.; Kleiner, G.; van Bakel, H.; et al. SARS-CoV-2-infection- and vaccine-induced antibody responses are long lasting with an initial waning phase followed by a stabilization phase. Immunity 2024, 57, 587–599.e4. [Google Scholar] [CrossRef]
- Menegale, F.; Manica, M.; Zardini, A.; Guzzetta, G.; Marziano, V.; D'ANdrea, V.; Trentini, F.; Ajelli, M.; Poletti, P.; Merler, S. Evaluation of Waning of SARS-CoV-2 Vaccine–Induced Immunity: A Systematic Review and Meta-analysis. JAMA Netw. Open 2023, 6, e2310650–e2310650. [Google Scholar] [CrossRef]
- Andrews N, Stowe J, Kirsebom F, Toffa S, Rickeard T, Gallagher E, Gower C, Kall M, Groves N, O'Connell AM, Simons D, Blomquist PB, Zaidi A, Nash S, Iwani Binti Abdul Aziz N, Thelwall S, Dabrera G, Myers R, Amirthalingam G, Gharbia S, Barrett JC, Elson R, Ladhani SN, Ferguson N, Zambon M, Campbell CNJ, Brown K, Hopkins S, Chand M, Ramsay M, Lopez Bernal J. Covid-19 Vaccine Effectiveness against the Omicron (B.1.1.529) Variant. N Engl J Med. 2022 Apr 21;386(16):1532-1546. [CrossRef]
- Mead, M.N.; Seneff, S.; Wolfinger, R.; Rose, J.; Denhaerynck, K.; Kirsch, S.; A McCullough, P. COVID-19 Modified mRNA “Vaccines”: Lessons Learned from Clinical Trials, Mass Vaccination, and the Bio-Pharmaceutical Complex, Part 1. Int. J. Vaccine Theory, Pr. Res. 2024, 3, 1112–1178. [Google Scholar] [CrossRef]
- Abou-Saleh, H.; Abo-Halawa, B.Y.; Younes, S.; Younes, N.; Al-Sadeq, D.W.; Shurrab, F.M.; Liu, N.; Qotba, H.; Al-Dewik, N.; Ismail, A.; et al. Neutralizing antibodies against SARS-CoV-2 are higher but decline faster in mRNA vaccinees compared to individuals with natural infection. J. Travel Med. 2022, 29. [Google Scholar] [CrossRef] [PubMed]
- Tamandjou, C.; Auvigne, V.; Schaeffer, J.; Vaux, S.; du Châtelet, I.P. Effectiveness of second booster compared to first booster and protection conferred by previous SARS-CoV-2 infection against symptomatic Omicron BA.2 and BA.4/5 in France. Vaccine 2023, 41, 2754–2760. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.W. Original antigen sin and COVID-19: implications for seasonal vaccination. Expert Opin. Biol. Ther. 2022, 22, 1353–1358. [Google Scholar] [CrossRef]
- Noori, M.; Nejadghaderi, S.A.; Rezaei, N. “Original antigenic sin”: A potential threat beyond the development of booster vaccination against novel SARS-CoV-2 variants. Infect. Control. Hosp. Epidemiology 2021, 43, 1091–1092. [Google Scholar] [CrossRef]
- Röltgen, K.; Nielsen, S.C.; Silva, O.; Younes, S.F.; Zaslavsky, M.; Costales, C.; Yang, F.; Wirz, O.F.; Solis, D.; Hoh, R.A.; et al. Immune imprinting, breadth of variant recognition, and germinal center response in human SARS-CoV-2 infection and vaccination. Cell 2022, 185, 1025–1040.e14. [Google Scholar] [CrossRef]
- Samanovic, M.I.; Cornelius, A.R.; Gray-Gaillard, S.L.; Allen, J.R.; Karmacharya, T.; Wilson, J.P.; Hyman, S.W.; Tuen, M.; Koralov, S.B.; Mulligan, M.J.; et al. Robust immune responses are observed after one dose of BNT162b2 mRNA vaccine dose in SARS-CoV-2–experienced individuals. Sci. Transl. Med. 2022, 14, eabi8961. [Google Scholar] [CrossRef]
- Offit, P.A. Bivalent Covid-19 Vaccines — A Cautionary Tale. New Engl. J. Med. 2023, 388, 481–483. [Google Scholar] [CrossRef]
- Wang, Q.; Feng, L.; Zhang, H.; Gao, J.; Mao, C.; Landesman-Bollag, E.; Mostoslavsky, G.; Lunderberg, J.M.; Zheng, W.; Hao, S.; et al. Longitudinal waning of mRNA vaccine-induced neutralizing antibodies against SARS-CoV-2 detected by an LFIA rapid test. Antib. Ther. 2022, 5, 55–62. [Google Scholar] [CrossRef]
- Afshar, Z.M.; Barary, M.; Hosseinzadeh, R.; Alijanpour, A.; Hosseinzadeh, D.; Ebrahimpour, S.; Nazary, K.; Sio, T.T.; Sullman, M.J.M.; Carson-Chahhoud, K.; et al. Breakthrough SARS-CoV-2 infections after vaccination: a critical review. Hum. Vaccines Immunother. 2022, 18, 2051412. [Google Scholar] [CrossRef]
- Moore, M.; Anderson, L.; Schiffer, J.T.; Matrajt, L.; Dimitrov, D. Durability of COVID-19 vaccine and infection induced immunity: A systematic review and meta-regression analysis. Vaccine 2025, 54, 126966. [Google Scholar] [CrossRef]
- Gopinath, S.; Ishak, A.; Dhawan, N.; Poudel, S.; Shrestha, P.S.; Singh, P.; Xie, E.; Tahir, P.; Marzaban, S.; Michel, J.; et al. Characteristics of COVID-19 Breakthrough Infections among Vaccinated Individuals and Associated Risk Factors: A Systematic Review. Trop. Med. Infect. Dis. 2022, 7, 81. [Google Scholar] [CrossRef]
- Zilla, M.L.; Keetch, C.; Mitchell, G.; McBreen, J.; Shurin, M.R.; E Wheeler, S. SARS-CoV-2 Serologic Immune Response in Exogenously Immunosuppressed Patients. J. Appl. Lab. Med. 2020, 6, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Jafarzadeh, A.; Ghadimi-Moghadam, A.; Mortazavi, S.M.J.; Haghani, M.; Sihver, L. Breakthrough Infection and Death after COVID-19 Vaccination: A Physics Perspective. J. Biomed. Phys. Eng. 2023, online, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.S.; Jones, F.K.; Nodoushani, A.; Kelly, M.; Becker, M.; Slater, D.; Mills, R.; Teng, E.; Kamruzzaman, M.; Garcia-Beltran, W.F.; et al. Persistence and decay of human antibody responses to the receptor binding domain of SARS-CoV-2 spike protein in COVID-19 patients. Sci. Immunol. 2020, 5. [Google Scholar] [CrossRef]
- Wajnberg, A.; Amanat, F.; Firpo, A.; Altman, D.R.; Bailey, M.J.; Mansour, M.; McMahon, M.; Meade, P.; Mendu, D.R.; Muellers, K.; et al. Robust neutralizing antibodies to SARS-CoV-2 infection persist for months. Science 2020, 370, 1227–1230. [Google Scholar] [CrossRef]
- Miyamoto, S.; Suzuki, T. Infection-mediated immune response in SARS-CoV-2 breakthrough infection and implications for next-generation COVID-19 vaccine development. Vaccine 2024, 42, 1401–1406. [Google Scholar] [CrossRef]
- Huang, W.; Gao, C.X.; Luo, D.; Wang, Y.; Zheng, X.; Liu, C.; Wang, Y.; Li, Y.; Qian, H. Risk evaluation of venue types and human behaviors of COVID-19 outbreaks in public indoor environments: A systematic review and meta-analysis. Environ. Pollut. 2023, 341, 122970. [Google Scholar] [CrossRef] [PubMed]
- Jamous, Y.F.; Alnakhli, M.; Alshaibi, A.; Alhawsawi, M.; Binsalman, A.; Uduman, M.S.T.S. The Incidence and Severity of COVID-19 Infection Post Vaccination in Saudi Arabia. Cureus 2023, 15, e39766. [Google Scholar] [CrossRef]
- Shahid, I.; Alzahrani, A.R.; Jabeen, Q.; Al-Ghamdi, S.S.; Shahzad, N.; Rehman, S.; Algarni, A.S.; Bamagous, G.A.; AlanazI, I.M.M.; Ibrahim, I.A.A. SARS-CoV-2 Detection and COVID-19 Diagnosis: A Bird’s Eye View. Rev. Recent Clin. Trials 2023, 18, 1–25. [Google Scholar] [CrossRef]
- Martignoni, M.M.; Mohammadi, Z.; Loredo-Osti, J.C.; Hurford, A. Extensive SARS-CoV-2 testing reveals BA.1/BA.2 asymptomatic rates and underreporting in school children. Can. Commun. Dis. Rep. 2023, 49, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.M. Outbreak of SARS-CoV-2 Infections, Including COVID-19 Vaccine Breakthrough Infections, Associated with Large Public Gatherings — Barnstable County, Massachusetts, July 2021. Mmwr-Morbidity Mortal. Wkly. Rep. 2021, 70, 1059–1062. [Google Scholar] [CrossRef]
- Ben Fredj, S.; Ghammem, R.; Zammit, N.; Maatouk, A.; Haddad, N.; Haddad, N.; Kachroudi, M.; Rebai, S.; Laadhari, H.; Ghodhbani, M.M.; et al. Risk factors for severe Covid-19 breakthrough infections: an observational longitudinal study. BMC Infect. Dis. 2022, 22, 1–11. [Google Scholar] [CrossRef]
- Moreno-Perez, O.; Ribes, I.; Boix, V.; Martinez-García, M.Á.; Otero-Rodriguez, S.; Reus, S.; Sánchez-Martínez, R.; Ramos, J.M.; Chico-Sánchez, P.; Merino, E. Hospitalized patients with breakthrough COVID-19: Clinical features and poor outcome predictors. Int. J. Infect. Dis. 2022, 118, 89–94. [Google Scholar] [CrossRef]
- Christensen, P.A.; Olsen, R.J.; Long, S.W.; Snehal, R.; Davis, J.J.; Saavedra, M.O.; Reppond, K.; Shyer, M.N.; Cambric, J.; Gadd, R.; et al. Signals of Significantly Increased Vaccine Breakthrough, Decreased Hospitalization Rates, and Less Severe Disease in Patients with Coronavirus Disease 2019 Caused by the Omicron Variant of Severe Acute Respiratory Syndrome Coronavirus 2 in Houston, Texas. Am. J. Pathol. 2022, 192, 642–652. [Google Scholar] [CrossRef]
- Albtoosh, A.S.; Farah, R.; Al Oweidat, K.; Hussein, O.M.; Obeid, A.A.; Hamila, H.M.; Radwan, M.N.M.; Ahmad, R.F.; Masadeh, H.M.; Hammad, A.I.; et al. Presenting clinical symptoms of post-COVID-19 breakthrough infection: Predictors of mortality in a Middle Eastern population. Vaccine: X 2024, 18, 100495. [Google Scholar] [CrossRef]
- Ioannidis, J.P.A. Infection fatality rate of COVID-19 inferred from seroprevalence data. Bull. World Health Organ. 2021, 99, 19–33F. [Google Scholar] [CrossRef]
- Pezzullo, A.M.; Axfors, C.; Contopoulos-Ioannidis, D.G.; Apostolatos, A.; Ioannidis, J.P. Age-stratified infection fatality rate of COVID-19 in the non-elderly population. Environ. Res. 2022, 216, 114655–114655. [Google Scholar] [CrossRef]
- Lipsitch, M.; Krammer, F.; Regev-Yochay, G.; Lustig, Y.; Balicer, R.D. SARS-CoV-2 breakthrough infections in vaccinated individuals: measurement, causes and impact. Nat. Rev. Immunol. 2021, 22, 57–65. [Google Scholar] [CrossRef]
- Robinson, J. Are these the numbers scaring Boris? Study shows 29% of the 42 people who have died after catching the new strain had BOTH vaccinations as cases soar another 40%. Daily Mail News. Published June 13, 2021. Accessed June 20, 2025. https://www.dailymail.co.uk/news/article-9681613/Study-shows-29-people-died-catching-new-strain-vaccinations.html.
- UK Government. Public Health England. Research and analysis. Investigation of SARS-CoV-2 variants of concern: technical briefings. Published 21 December 2020. Last updated 17 September 2021. URL: https://www.gov.uk/government/publications/investigation-of-novel-sars-cov-2-variant-variant-ofconcern-20201201.
- Anonymous. 151 Dead, 563 Hospitalized in Illinois Breakthrough COVID Cases. NBC News Chicago. Published July 16, 2021. Accessed June 20, 2025. https://www.nbcchicago.com/news/coronavirus/151-dead-563-hospitalized-in-illinois-breakthrough-covid-cases/2556408/.
- Ong, D. 277 Fully Vaccinated Indiana Residents Have Died Of COVID-19 In 3 Weeks. International Business Times. Published January 23, 2022. Accessed June 20, 2025. https://www.ibtimes.com/277-fully-vaccinatedindiana-residents-have-died-covid-19-3-weeks-3381687.
- Anonymous. COVID-19 Cases in Fully Vaccinated Individuals. Massachusetts Department of Public Health. Published January 4, 2021. Accessed June 20, 2025. https://www.mass.gov/doc/weekly-reportcovid-19-cases-in-vaccinated-individuals-january-4-2022/download.
- Ong, D. 290 Fully Vaccinated Massachusetts Residents Died Of COVID-19 Over 1 Week. International Business Times. Published February 2, 2022. Accessed June 20, 2025. https://www.ibtimes.com/290-fullyvaccinated-massachusetts-residents-died-covid-19-over-1-week-3389234.
- Ong, D. 2,222 Fully Vaccinated Massachusetts Residents Have Died Of COVID-19. International Business Times. Published February 16, 2022. Accessed June 20, 2025. https://www.ibtimes.com/2222-fullyvaccinated-massachusetts-residents-have-died-covid-19-3403439.
- Attwell, K.; Hannah, A. Convergence on Coercion: Functional and Political Pressures as Drivers of Global Childhood Vaccine Mandates. Int. J. Heal. Policy Manag. 2022, 11, 2660–2671. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.K.; Alleaume, C.; Peretti-Watel, P.; theCOCONEL Group. The French public's attitudes to a future COVID-19 vaccine: The politicization of a public health issue. Soc. Sci. Med. 2020, 265, 113414. [Google Scholar] [CrossRef] [PubMed]
- Fenton N, Neil M. The Very Best Cheap Trick. In: Fighting Goliath: Exposing the flawed science and statistics behind the COVID-19 event. Sovereign Rights Publishing, United Kingdom. 2024. pp. 202-212.
- Basoulis, D.; Logioti, K.; Papaodyssea, I.; Chatzopoulos, M.; Alexopoulou, P.; Mavroudis, P.; Rapti, V.; Poulia, V.; Samara, S.; Georgakopoulou, V.E.; et al. Deaths “due to” COVID-19 and deaths “with” COVID-19 during the Omicron variant surge, among hospitalized patients in seven tertiary-care hospitals, Athens, Greece. Sci. Rep. 2025, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chemaitelly, H.; Tang, P.; Hasan, M.R.; AlMukdad, S.; Yassine, H.M.; Benslimane, F.M.; Al Khatib, H.A.; Coyle, P.; Ayoub, H.H.; Al Kanaani, Z.; et al. Waning of BNT162b2 Vaccine Protection against SARS-CoV-2 Infection in Qatar. New Engl. J. Med. 2021, 385, E83–E83. [Google Scholar] [CrossRef]
- Chitwood, M.H.; Russi, M.; Gunasekera, K.; Havumaki, J.; Klaassen, F.; Pitzer, V.E.; Salomon, J.A.; Swartwood, N.A.; Warren, J.L.; Weinberger, D.M.; et al. Reconstructing the course of the COVID-19 epidemic over 2020 for US states and counties: Results of a Bayesian evidence synthesis model. PLOS Comput. Biol. 2022, 18, e1010465. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Interim public health considerations for the provision of additional COVID-19 vaccine doses. https://www.ecdc.europa.eu/en/publications-data/covid-19-public-health-considerations-additional-vaccine-doses Accessed 22 June 2025.
- Nordström, P.; Ballin, M.; Nordström, A. Risk of infection, hospitalisation, and death up to 9 months after a second dose of COVID-19 vaccine: a retrospective, total population cohort study in Sweden. Lancet 2022, 399, 814–823. [Google Scholar] [CrossRef]
- Shrestha, N.K.; Burke, P.C.; Nowacki, A.S.; Simon, J.F.; Hagen, A.; Gordon, S.M. Effectiveness of the Coronavirus Disease 2019 Bivalent Vaccine. Open Forum Infect. Dis. 2023, 10. [Google Scholar] [CrossRef]
- Nakatani, E.; Morioka, H.; Kikuchi, T.; Fukushima, M. Behavioral and Health Outcomes of mRNA COVID-19 Vaccination: A Case-Control Study in Japanese Small and Medium-Sized Enterprises. Cureus 2024, 16, e75652. [Google Scholar] [CrossRef]
- Chemaitelly, H.; Ayoub, H.H.; AlMukdad, S.; Coyle, P.; Tang, P.; Yassine, H.M.; Al-Khatib, H.A.; Smatti, M.K.; Hasan, M.R.; Al-Kanaani, Z.; et al. Duration of mRNA vaccine protection against SARS-CoV-2 Omicron BA.1 and BA.2 subvariants in Qatar. Nat. Commun. 2022, 13, 1–12. [Google Scholar] [CrossRef]
- UK Health Security Agency. COVID-19 vaccine surveillance report, Week 8 Feb. 24, 2022. https://assets.publishing.service.gov.uk/media/621c91c0d3bf7f4f04b2b648/Vaccine_surveillance_report_-_week-8.pdf.
- Eythorsson, E.; Runolfsdottir, H.L.; Ingvarsson, R.F.; Sigurdsson, M.I.; Palsson, R. Rate of SARS-CoV-2 Reinfection During an Omicron Wave in Iceland. JAMA Netw. Open 2022, 5, e2225320–e2225320. [Google Scholar] [CrossRef] [PubMed]
- Feldstein, L.R.; Ruffin, J.; Wiegand, R.; Grant, L.; Babu, T.M.; Briggs-Hagen, M.; Burgess, J.L.; Caban-Martinez, A.J.; Chu, H.Y.; Ellingson, K.D.; et al. Protection From COVID-19 Vaccination and Prior SARS-CoV-2 Infection Among Children Aged 6 Months–4 Years, United States, September 2022–April 2023. J. Pediatr. Infect. Dis. Soc. 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, G.N.; Berry, K.; Rajeevan, N.; Li, Y.; Yan, L.; Huang, Y.; Lin, H.-M.; Bui, D.; Hynes, D.M.; Rowneki, M.; et al. Effectiveness of the 2023-to-2024 XBB.1.5 COVID-19 Vaccines Over Long-Term Follow-up. Ann. Intern. Med. 2025, 178, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.K.; Burke, P.C.; Nowacki, A.S.; Gordon, S.M.; Martellucci, C.A. Risk of Coronavirus Disease 2019 (COVID-19) among those up-to-date and not up-to-date on COVID-19 vaccination by US CDC criteria. PLOS ONE 2023, 18, e0293449. [Google Scholar] [CrossRef]
- Gazit, S.; Shlezinger, R.; Perez, G.; Lotan, R.; Peretz, A.; Ben-Tov, A.; Cohen, D.; Muhsen, K.; Chodick, G.; Patalon, T. Comparing SARS-CoV-2 natural immunity to vaccine-induced immunity: Reinfections versus breakthrough infections. MedRxiv 2021. [Google Scholar] [CrossRef]
- Rose, J. Breakthrough Infection Signal in VAERS Corroborates IgG4-Mediated Increased Susceptibility to SARS-CoV-2. Sci Public Health Policy Law. 2025;6:2019-2025. Available from: https://publichealthpolicyjournal.com.
- Beattie, KA. Worldwide Bayesian causal impact analysis of vaccine administration on deaths and dases associated with COVID-19: A BigData analysis of 145 countries. Department of Political Science University of Alberta, Alberta, Canada. 2021. [CrossRef]
- Johns Hopkins University School of Medicine, Coronavirus Resource Center. Accessed on March 15, 2025. https://coronavirus.jhu.edu/.
- Faksova, K.; Walsh, D.; Jiang, Y.; Griffin, J.; Phillips, A.; Gentile, A.; Kwong, J.; Macartney, K.; Naus, M.; Grange, Z.; et al. COVID-19 vaccines and adverse events of special interest: A multinational Global Vaccine Data Network (GVDN) cohort study of 99 million vaccinated individuals. Vaccine 2024, 42, 2200–2211. [Google Scholar] [CrossRef]
- Montano, D. Frequency and Associations of Adverse Reactions of COVID-19 Vaccines Reported to Pharmacovigilance Systems in the European Union and the United States. Front. Public Heal. 2022, 9, 756633. [Google Scholar] [CrossRef]
- Yan, M.-M.; Zhao, H.; Li, Z.-R.; Chow, J.-W.; Zhang, Q.; Qi, Y.-P.; Wu, S.-S.; Zhong, M.-K.; Qiu, X.-Y. Serious adverse reaction associated with the COVID-19 vaccines of BNT162b2, Ad26.COV2.S, and mRNA-1273: Gaining insight through the VAERS. Front. Pharmacol. 2022, 13, 921760. [Google Scholar] [CrossRef]
- Classen, B. US COVID-19 vaccines proven to cause more harm than good based on pivotal clinical trial data analyzed using the proper scientific endpoint, “all cause severe morbidity”. Trends in Internal Medicine 2021; 1:1-6. URL: https://scivisionpub.com/pdfs/us-covid19-vaccines-proven-to-cause-more-harm-than-good-based-on-pivotal-clinical-trial-data-analyzed-using-the-proper-scientific--1811.pdf.
- Acevedo-Whitehouse, K.; Bruno, R. Potential health risks of mRNA-based vaccine therapy: A hypothesis. Med Hypotheses 2023, 171, 111015. [Google Scholar] [CrossRef]
- A Abdulkader, M.; A Merza, M. Immediate and Long-Term Adverse Events of COVID-19 Vaccines: A One-Year Follow-Up Study From the Kurdistan Region of Iraq. Cureus 2023, 15, e47670. [Google Scholar] [CrossRef]
- Doshi, P. Covid-19 vaccines: In the rush for regulatory approval, do we need more data? BMJ 2021, 373, n1244. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S. Don’t rush to deploy COVID-19 vaccines and drugs without sufficient safety guarantees. Nature 2020, 579, 321–321. [Google Scholar] [CrossRef] [PubMed]
- Gøtzsche PC, Demasi M. Serious harms of the COVID-19 vaccines: a systematic review. medRxiv Preprint. 2022.
- Fraiman, J.; Erviti, J.; Jones, M.; Greenland, S.; Whelan, P.; Kaplan, R.M.; Doshi, P. Serious adverse events of special interest following mRNA COVID-19 vaccination in randomized trials in adults. Vaccine 2022, 40, 5798–5805. [Google Scholar] [CrossRef] [PubMed]
- U.S. Centers for Disease Control. Historical Vaccine Safety Concerns. Accessed 7/10/2023. https://www.cdc.gov/vaccinesafety/concerns/concerns-history.html.
- Michels, C.; Perrier, D.; Kunadhasan, J.; Clark, E.; Gehrett, J.; Gehrett, B.; Kwiatek, K.; Adams, S.; Chandler, R.; Stagno, L.; et al. Forensic analysis of the 38 subject deaths in the 6-Month Interim Report of the Pfizer/BioNTech BNT162b2 mRNA Vaccine Clinical Trial. Int. J. Vaccine Theory, Pr. Res. 2023, 3, 973–1008. [Google Scholar] [CrossRef]
- Pfizer, Inc. Appendix 2.2 Cumulative and Interval Summary Tabulation of Serious and Non-serious Adverse Reactions From Post-marketing Data Sources (BNT162B2). New York, NY. 2022. Accessed: December 9, 2024: https://www.globalresearch.ca/wp-content/uploads/2023/05/pfizer-report.pdf.
- Pfizer, Inc. Periodic safety update report #3 for active substance: COVID-19 mRNA vaccine, BNT162b2. BioNTech Manufacturing GmbH, Mainz, Germany. 2022. Accessed: December 9, 2024: https://tkp.at/wp-content/uploads/2023/03/3.PSUR-1.pdf.
- Horowitz, D. Confidential Pfizer document shows the company observed 1.6 million adverse events covering nearly every organ system. 2023. Accessed: December 9, 2024: https://www.conservativereview.com/horowitz-confidential-pfizer-document-shows-the-company-observed-1-6-million-adverse-events-covering-nearly-every-organ-system-2661316948.html.
- Hulscher, N.; Hodkinson, R.; Makis, W.; McCullough, P.A. Autopsy findings in cases of fatal COVID-19 vaccine-induced myocarditis. ESC Hear. Fail. 2024. [Google Scholar] [CrossRef]
- Hulscher N, Alexander PE, Amerling R, Gessling H, Hodkinson R, Makis W, Risch HA, et al. A systematic review of autopsy findings in deaths after COVID-19 vaccination. Science, Public Health Policy and the Law. 17 November 2024. v5.2019-2024. https://publichealthpolicyjournal.com/a-systematic-review-ofautopsy-findings-in-deaths-after-covid-19-vaccination/.
- Food and Drug Administration. Initial Results of Near Real-Time Safety Monitoring COVID-199 Vaccines in Persons Age 65 Years and Older [Internet] 2021. Accessed 11 May 2025. https://www.fda.gov/vaccinesblood-biologics/safety-availability-biologics/initial-results-near-real-time-safety-monitoring-covid-19-vaccines-persons-aged-65-years-and-older.
- Lataster, R. Reply to Fung et al. on COVID-19 vaccine case-counting window biases overstating vaccine effectiveness. J. Evaluation Clin. Pr. 2023, 30, 82–85. [Google Scholar] [CrossRef]
- HHJ News, Obituaries: Shawn Thomas Kuhn. Houston Home Journal. 10-16-2021. https://hhjonline.com/shawn-thomas-kuhn/.
- Gilbertson, D. ‘I’m going to miss my friend’: Southwest Airlines flight attendant, 36, dies from COVID-19. USA Today. Airline News. 8-12-2021. https://www.usatoday.com/story/travel/airlinenews/ 2021/08/11/southwest-airlines-flight-attendant-covid-maurice-reggie-shepperson/8100532002/.
- Suleyman, A. Fully Vaccinated New Orleans Woman Dies of COVID Aged 33 in Rare Breakthrough Case. Newsweek. 7-27-2021. https://www.newsweek.com/fully-vaccinated-new-orleans-woman-dies-covidaged- 33-rare-breakthrough-case-1613379.
- Parry, P.I.; Lefringhausen, A.; Turni, C.; Neil, C.J.; Cosford, R.; Hudson, N.J.; Gillespie, J. ‘Spikeopathy’: COVID-19 Spike Protein Is Pathogenic, from Both Virus and Vaccine mRNA. Biomedicines 2023, 11, 2287. [Google Scholar] [CrossRef]
- Schwartz, L.; Aparicio-Alonso, M.; Henry, M.; Radman, M.; Attal, R.; Bakkar, A. Toxicity of the spike protein of COVID-19 is a redox shift phenomenon: A novel therapeutic approach. Free. Radic. Biol. Med. 2023, 206, 106–110. [Google Scholar] [CrossRef]
- Theoharides, T. Be aware of SARS-CoV-2 spike protein: There is more than meets the eye. J. Biol. Regul. Homeost. AGENTS 2021, 35, 833–838. [Google Scholar] [CrossRef]
- Lei, Y.; Zhang, J.; Schiavon, C.R.; He, M.; Chen, L.; Shen, H.; Zhang, Y.; Yin, Q.; Cho, Y.; Andrade, L.; et al. SARS-CoV-2 Spike Protein Impairs Endothelial Function via Downregulation of ACE 2. Circ. Res. 2021, 128, 1323–1326. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-P.; Wen, Y.-H.; Lin, W.-T.; Hsu, F.-P. Association between the side effect induced by COVID-19 vaccines and the immune regulatory gene polymorphism. Front. Immunol. 2022, 13, 941497. [Google Scholar] [CrossRef] [PubMed]
- Ndeupen, S.; Qin, Z.; Jacobsen, S.; Bouteau, A.; Estanbouli, H.; Igyártó, B.Z. The mRNA-LNP platform's lipid nanoparticle component used in preclinical vaccine studies is highly inflammatory. iScience 2021, 24, 103479. [Google Scholar] [CrossRef]
- Di Gioacchino M, Petrarca C, Lazzarin F, Di Giampaolo L, Sabbioni E, Boscolo P, Mariani-Costantini R, et al. Immunotoxicity of nanoparticles. Int J Immunopathol Pharmacol. 2011;24(1 Suppl):65S-71S.
- Cui, S.; Wang, Y.; Gong, Y.; Lin, X.; Zhao, Y.; Zhi, D.; Zhou, Q.; Zhang, S. Correlation of the cytotoxic effects of cationic lipids with their headgroups. Toxicol. Res. 2018, 7, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Bouteau, A.; Herbst, C.; Igyártó, B.Z.; Klein, S.L. Pre-exposure to mRNA-LNP inhibits adaptive immune responses and alters innate immune fitness in an inheritable fashion. PLOS Pathog. 2022, 18, e1010830. [Google Scholar] [CrossRef]
- Verbeke, R.; Lentacker, I.; De Smedt, S.C.; Dewitte, H. The dawn of mRNA vaccines: The COVID-19 case. J. Control. Release 2021, 333, 511–520. [Google Scholar] [CrossRef]
- Lonez, C.; Bessodes, M.; Scherman, D.; Vandenbranden, M.; Escriou, V.; Ruysschaert, J.-M. Cationic lipid nanocarriers activate Toll-like receptor 2 and NLRP3 inflammasome pathways. Nanomedicine: Nanotechnology, Biol. Med. 2014, 10, 775–782. [Google Scholar] [CrossRef]
- Kedmi, R.; Ben-Arie, N.; Peer, D. The systemic toxicity of positively charged lipid nanoparticles and the role of Toll-like receptor 4 in immune activation. Biomaterials 2010, 31, 6867–6875. [Google Scholar] [CrossRef]
- Kashani, B.; Zandi, Z.; Pourbagheri-Sigaroodi, A.; Bashash, D.; Ghaffari, S.H. The role of toll-like receptor 4 (TLR4) in cancer progression: A possible therapeutic target? J. Cell. Physiol. 2020, 236, 4121–4137. [Google Scholar] [CrossRef]
- Parhiz, H.; Brenner, J.S.; Patel, P.N.; Papp, T.E.; Shahnawaz, H.; Li, Q.; Shi, R.; Zamora, M.E.; Yadegari, A.; Marcos-Contreras, O.A.; et al. Added to pre-existing inflammation, mRNA-lipid nanoparticles induce inflammation exacerbation (IE). J. Control. Release 2021, 344, 50–61. [Google Scholar] [CrossRef]
- Soegiarto, G.; Purnomosari, D. Challenges in the Vaccination of the Elderly and Strategies for Improvement. Pathophysiology 2023, 30, 155–173. [Google Scholar] [CrossRef]
- McKernan K, Helbert Y, Kane LT, McLaughlin S. Sequencing of bivalent Moderna and Pfizer mRNA vaccines reveals nanogram to microgram quantities of expression vector dsDNA per dose. OSF Prepr. 2023. [CrossRef]
- Speicher DJ, Rose J, Gutschi LM, McKernan K. DNA Fragments detected in monovalent and bivalent Pfizer/BioNTech and Moderna modRNA COVID-19 vaccines from Ontario, Canada: Exploratory dose response relationship with SERIOUS adverse events. OSF Preprints. Oct 19, 2023. [(accessed on 26 February 2024)]. Available online: https://osf.io/preprints/osf/mjc97.
- König, B.; Kirchner, J.O. Methodological Considerations Regarding the Quantification of DNA Impurities in the COVID-19 mRNA Vaccine Comirnaty®. Methods Protoc. 2024, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Aldén, M.; Olofsson Falla, F.; Yang, D.; Barghouth, M.; Luan, C.; Rasmussen, M.; De Marinis, Y. Intracellular Reverse Transcription of Pfizer BioNTech COVID-19 mRNA Vaccine BNT162b2 In Vitro in Human Liver Cell Line. Curr. Issues Mol. Biol. 2022, 44, 1115–1126. [Google Scholar] [CrossRef] [PubMed]
- Caforio, A.L.P.; Tona, F.; Bottaro, S.; Vinci, A.; Dequal, G.; Daliento, L.; Thiene, G.; Iliceto, S. Clinical implications of anti-heart autoantibodies in myocarditis and dilated cardiomyopathy. Autoimmunity 2008, 41, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Levi R, Mansuri F, Jordan MM, Ladapo JA. Twelve-Month All-Cause Mortality after Initial COVID-19 Vaccination with Pfizer-BioNTech or mRNA-1273 among Adults Living in Florida. medRxiv [preprint]. Posted 29 April 2025. [CrossRef]
- Weiler, JL. Florida Study Reveals Elevated Mortality Risk Following Pfizer COVID-19 Vaccine Compared to Moderna. Popular Rationalism [Substack]. 6 May 2025. https://popularrationalism.substack.com/p/florida-study-reveals-elevatedmortality?utm_source=substack&utm_campaign=post_embed&utm_medium=web.
- Siebner, A.S.; Griesbaum, J.; E Huus, K.; Flügge, J.; Hopfensperger, K.; Michel, T.; Schneiderhan-Marra, N.; Sauter, D.; Kremsner, P.G.; E Ley, R.; et al. Class switch towards IgG2 and IgG4 is more pronounced in BNT162b2 compared to mRNA-1273 COVID-19 vaccinees. . 2025, 107990. [Google Scholar] [CrossRef]
- Sass, E. COVID-19 mRNA “vaccine” harms research collection. In Toxic Shot: Facing the Dangers of the COVID “Vaccines”. Zenodo. 2 July. [CrossRef]
- Japanese Pharmaceuticals and Medical Devices Agency (PMDA) SARS-CoV-2 mRNA Vaccine (BNT162, PF-07302048) 2021. [(accessed on 7 April 2023)]. Available online: https://www.pmda.go.jp/drugs/2021/P20210212001/672212000_30300AMX00231_I100_1.pdf.
- Judicial Watch Pfizer/BioNTech Study Found Lipid Nanoparticles Materials Outside Injection Site in Test Animals. judicialwatch.org. 2022. [(accessed on 12 July 2023)]. Available online: https://www.judicialwatch.org/nanoparticles-materials-outside-injection-site/.
- Di, J.; Du, Z.; Wu, K.; Jin, S.; Wang, X.; Li, T.; Xu, Y. Biodistribution and Non-linear Gene Expression of mRNA LNPs Affected by Delivery Route and Particle Size. Pharm. Res. 2022, 39, 105–114. [Google Scholar] [CrossRef]
- Trougakos, I.P.; Terpos, E.; Alexopoulos, H.; Politou, M.; Paraskevis, D.; Scorilas, A.; Kastritis, E.; Andreakos, E.; Dimopoulos, M.A. Adverse effects of COVID-19 mRNA vaccines: the spike hypothesis. Trends Mol. Med. 2022, 28, 542–554. [Google Scholar] [CrossRef]
- Baumeier, C.; Aleshcheva, G.; Harms, D.; Gross, U.; Hamm, C.; Assmus, B.; Westenfeld, R.; Kelm, M.; Rammos, S.; Wenzel, P.; et al. Intramyocardial Inflammation after COVID-19 Vaccination: An Endomyocardial Biopsy-Proven Case Series. Int. J. Mol. Sci. 2022, 23, 6940. [Google Scholar] [CrossRef]
- Sriwastava, S.; Sharma, K.; Khalid, S.H.; Bhansali, S.; Shrestha, A.K.; Elkhooly, M.; Srivastava, S.; Khan, E.; Jaiswal, S.; Wen, S. COVID-19 Vaccination and Neurological Manifestations: A Review of Case Reports and Case Series. Brain Sci. 2022, 12, 407. [Google Scholar] [CrossRef]
- Vogrig, A.; Tartaglia, S.; Dentoni, M.; Fabris, M.; Bax, F.; Belluzzo, M.; Verriello, L.; Bagatto, D.; Gastaldi, M.; Tocco, P.; et al. Central nervous system immune-related disorders after SARS-CoV-2 vaccination: a multicenter study. Front. Immunol. 2024, 15, 1344184. [Google Scholar] [CrossRef] [PubMed]
- Schinas, G.; Polyzou, E.; Dimakopoulou, V.; Tsoupra, S.; Gogos, C.; Akinosoglou, K. Immune-mediated liver injury following COVID-19 vaccination. World J. Virol. 2023, 12, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Buzhdygan, T.P.; DeOre, B.J.; Baldwin-Leclair, A.; Bullock, T.A.; McGary, H.M.; Khan, J.A.; Razmpour, R.; Hale, J.F.; Galie, P.A.; Potula, R.; et al. The SARS-CoV-2 spike protein alters barrier function in 2D static and 3D microfluidic in-vitro models of the human blood–brain barrier. Neurobiol. Dis. 2020, 146, 105131–105131. [Google Scholar] [CrossRef]
- Posa, A. Spike protein-related proteinopathies: A focus on the neurological side of spikeopathies. Ann. Anat. - Anat. Anz. 2025, 260, 152662. [Google Scholar] [CrossRef]
- Mulroney, T.E.; Pöyry, T.; Yam-Puc, J.C.; Rust, M.; Harvey, R.F.; Kalmar, L.; Horner, E.; Booth, L.; Ferreira, A.P.; Stoneley, M.; et al. N1-methylpseudouridylation of mRNA causes +1 ribosomal frameshifting. Nature 2023, 625, 189–194. [Google Scholar] [CrossRef]
- Seneff, S.; Kyriakopoulos, A.M.; Nigh, G.; A McCullough, P.; McCullough, P.A. A Potential Role of the Spike Protein in Neurodegenerative Diseases: A Narrative Review. Cureus 2023, 15, e34872. [Google Scholar] [CrossRef]
- Perez, J.-C.; Moret-Chalmin, C.; Montagnier, L. Emergence of a New Creutzfeldt-Jakob Disease: 26 Cases of the Human Version of Mad-Cow Disease, Days After a COVID-19 Injection. Int. J. Vaccine Theory, Pr. Res. 2023, 3, 727–770. [Google Scholar] [CrossRef]
- Nance, K.D.; Meier, J.L. Modifications in an Emergency: The Role of N1-Methylpseudouridine in COVID-19 Vaccines. ACS Central Sci. 2021, 7, 748–756. [Google Scholar] [CrossRef]
- Morais, P.; Adachi, H.; Yu, Y.-T. The Critical Contribution of Pseudouridine to mRNA COVID-19 Vaccines. Front. Cell Dev. Biol. 2021, 9, 789427. [Google Scholar] [CrossRef]
- Kim, K.Q.; Burgute, B.D.; Tzeng, S.-C.; Jing, C.; Jungers, C.; Zhang, J.; Yan, L.L.; Vierstra, R.D.; Djuranovic, S.; Evans, B.S.; et al. N1-methylpseudouridine found within COVID-19 mRNA vaccines produces faithful protein products. Cell Rep. 2022, 40, 111300–111300. [Google Scholar] [CrossRef]
- Corbett, K.S.; Edwards, D.K.; Leist, S.R.; Abiona, O.M.; Boyoglu-Barnum, S.; Gillespie, R.A.; Himansu, S.; Schäfer, A.; Ziwawo, C.T.; DiPiazza, A.T.; et al. SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature 2020, 586, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.J.; Gychka, S.G. SARS-CoV-2 Spike Protein Elicits Cell Signaling in Human Host Cells: Implications for Possible Consequences of COVID-19 Vaccines. Vaccines 2021, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Krauson, A.J.; Casimero, F.V.C.; Siddiquee, Z.; Stone, J.R. Duration of SARS-CoV-2 mRNA vaccine persistence and factors associated with cardiac involvement in recently vaccinated patients. npj Vaccines 2023, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ogata, A.F.; Cheng, C.-A.; Desjardins, M.; Senussi, Y.; Sherman, A.C.; Powell, M.; Novack, L.; Von, S.; Li, X.; Baden, L.R.; et al. Circulating Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Vaccine Antigen Detected in the Plasma of mRNA-1273 Vaccine Recipients. Clin. Infect. Dis. 2021, 74, 715–718. [Google Scholar] [CrossRef]
- Bansal, S.; Perincheri, S.; Fleming, T.; Poulson, C.; Tiffany, B.; Bremner, R.M.; Mohanakumar, T. Cutting Edge: Circulating Exosomes with COVID Spike Protein Are Induced by BNT162b2 (Pfizer–BioNTech) Vaccination prior to Development of Antibodies: A Novel Mechanism for Immune Activation by mRNA Vaccines. J. Immunol. 2021, 207, 2405–2410. [Google Scholar] [CrossRef]
- Boros, L.G.; Kyriakopoulos, A.M.; Brogna, C.; Piscopo, M.; McCullough, P.A.; Seneff, S. Long-lasting, biochemically modified mRNA, and its frameshifted recombinant spike proteins in human tissues and circulation after COVID-19 vaccination. Pharmacol. Res. Perspect. 2024, 12, e1218. [Google Scholar] [CrossRef]
- Brogna, C.; Cristoni, S.; Marino, G.; Montano, L.; Viduto, V.; Fabrowski, M.; Lettieri, G.; Piscopo, M. Detection of recombinant Spike protein in the blood of individuals vaccinated against SARS-CoV-2: Possible molecular mechanisms. Proteom. – Clin. Appl. 2023, 17, e2300048. [Google Scholar] [CrossRef]
- Patterson, B.K.; Yogendra, R.; Francisco, E.B.; Guevara-Coto, J.; Long, E.; Pise, A.; Osgood, E.; Bream, J.; Kreimer, M.; Jeffers, D.; et al. Detection of S1 spike protein in CD16+ monocytes up to 245 days in SARS-CoV-2-negative post-COVID-19 vaccine syndrome (PCVS) individuals. Hum. Vaccines Immunother. 2025, 21, 2494934. [Google Scholar] [CrossRef]
- Ota, N.; Itani, M.; Aoki, T.; Sakurai, A.; Fujisawa, T.; Okada, Y.; Noda, K.; Arakawa, Y.; Tokuda, S.; Tanikawa, R. Expression of SARS-CoV-2 spike protein in cerebral Arteries: Implications for hemorrhagic stroke Post-mRNA vaccination. J. Clin. Neurosci. 2025, 136, 111223. [Google Scholar] [CrossRef]
- Bhattacharjee B, Lu P, Monteiro VS, Tabachnikova A, Wang K, Hooper WB, Bastos V, et al. Immunological and Antigenic Signatures Associated with Chronic Illnesses after COVID-19 Vaccination. medRxiv preprint 2025. [CrossRef]
- Personal communications with epidemiologist Nicolas Hulscher, McCullough Foundation. Article in press. 25 July 2025.
- Seneff, S.; Nigh, G.; Kyriakopoulos, A.M.; McCullough, P.A. Innate immune suppression by SARS-CoV-2 mRNA vaccinations: The role of G-quadruplexes, exosomes, and MicroRNAs. Food Chem. Toxicol. 2022, 164, 113008–113008. [Google Scholar] [CrossRef]
- Klingel, H.; Krüttgen, A.; Imöhl, M.; Kleines, M. Humoral immune response to SARS-CoV-2 mRNA vaccines is associated with choice of vaccine and systemic adverse reactions DMD TNR. Clin. Exp. Vaccine Res. 2023, 12, 60–69. [Google Scholar] [CrossRef]
- Ouranidis, A.; Vavilis, T.; Mandala, E.; Davidopoulou, C.; Stamoula, E.; Markopoulou, C.K.; Karagianni, A.; Kachrimanis, K. mRNA Therapeutic Modalities Design, Formulation and Manufacturing under Pharma 4.0 Principles. Biomedicines 2021, 10, 50. [Google Scholar] [CrossRef]
- Milano, G.; Gal, J.; Creisson, A.; Chamorey, E. Myocarditis and COVID-19 Mrna Vaccines: A Mechanistic Hypothesis Involving dsRNA. Futur. Virol. 2021, 17, 191–196. [Google Scholar] [CrossRef]
- Uversky, V.N.; Redwan, E.M.; Makis, W.; Rubio-Casillas, A. IgG4 Antibodies Induced by Repeated Vaccination May Generate Immune Tolerance to the SARS-CoV-2 Spike Protein. Vaccines 2023, 11, 991. [Google Scholar] [CrossRef] [PubMed]
- Irrgang, P.; Gerling, J.; Kocher, K.; Lapuente, D.; Steininger, P.; Habenicht, K.; Wytopil, M.; Beileke, S.; Schäfer, S.; Zhong, J.; et al. Class switch toward noninflammatory, spike-specific IgG4 antibodies after repeated SARS-CoV-2 mRNA vaccination. Sci. Immunol. 2023, 8, eade2798. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.K.; Paliwal, V.K. Spectrum of neurological complications following COVID-19 vaccination. Neurol. Sci. 2021, 43, 3–40. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Kobayashi, S.; Hayashi, T.; Tachibana, M.; Saito, T.; Ogura, K.; Miyakoshi, S. Immune thrombocytopenic purpura in an elderly patient with cerebral hemorrhage after the fourth mRNA-1273 COVID-19 vaccination. Geriatr. Gerontol. Int. 2023, 23, 969–970. [Google Scholar] [CrossRef]
- Lee, E.-J.; Beltrami-Moreira, M.; Al-Samkari, H.; Cuker, A.; DiRaimo, J.; Gernsheimer, T.; Kruse, A.; Kessler, C.; Kruse, C.; Leavitt, A.D.; et al. SARS-CoV-2 vaccination and ITP in patients with de novo or preexisting ITP. Blood 2021, 139, 1564–1574. [Google Scholar] [CrossRef]
- Liu, J.; Wang, J.; Xu, J.; Xia, H.; Wang, Y.; Zhang, C.; Chen, W.; Zhang, H.; Liu, Q.; Zhu, R.; et al. Comprehensive investigations revealed consistent pathophysiological alterations after vaccination with COVID-19 vaccines. Cell Discov. 2021, 7, 1–15. [Google Scholar] [CrossRef]
- Chevaisrakul, P.; Lumjiaktase, P.; Kietdumrongwong, P.; Chuatrisorn, I.; Chatsangjaroen, P.; Phanuphak, N. Hybrid and herd immunity 6 months after SARS-CoV-2 exposure among individuals from a community treatment program. Sci. Rep. 2023, 13, 1–9. [Google Scholar] [CrossRef]
- Espino, A.M.; Armina-Rodriguez, A.; Alvarez, L.; Ocasio-Malavé, C.; Ramos-Nieves, R.; Martinó, E.I.R.; López-Marte, P.; Torres, E.A.; Sariol, C.A. The Anti-SARS-CoV-2 IgG1 and IgG3 Antibody Isotypes with Limited Neutralizing Capacity against Omicron Elicited in a Latin Population a Switch toward IgG4 after Multiple Doses with the mRNA Pfizer–BioNTech Vaccine. Viruses 2024, 16, 187. [Google Scholar] [CrossRef] [PubMed]
- Motta, R.V.; Culver, E.L. IgG4 autoantibodies and autoantigens in the context of IgG4-autoimmune disease and IgG4-related disease. Front. Immunol. 2024, 15, 1272084. [Google Scholar] [CrossRef] [PubMed]
- Collier, J.L.; Weiss, S.A.; Pauken, K.E.; Sen, D.R.; Sharpe, A.H. Not-so-opposite ends of the spectrum: CD8+ T cell dysfunction across chronic infection, cancer and autoimmunity. Nat. Immunol. 2021, 22, 809–819. [Google Scholar] [CrossRef]
- Shrestha, N.K.; Burke, P.C.; Nowacki, A.S.; Gordon, S.M. Effectiveness of the 2023–2024 Formulation of the COVID-19 Messenger RNA Vaccine. Clin. Infect. Dis. 2024, 79, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Kiszel, P.; Sík, P.; Miklós, J.; Kajdácsi, E.; Sinkovits, G.; Cervenak, L.; Prohászka, Z. Class switch towards spike protein-specific IgG4 antibodies after SARS-CoV-2 mRNA vaccination depends on prior infection history. Sci. Rep. 2023, 13, 1–12. [Google Scholar] [CrossRef]
- Pérez, C.M.; Ruiz-Rius, S.; Ramírez-Morros, A.; Vidal, M.; Opi, D.H.; Santamaria, P.; Blanco, J.; Vidal-Alaball, J.; Beeson, J.G.; Molinos-Albert, L.M.; et al. Post-vaccination IgG4 and IgG2 class switch associates with increased risk of SARS-CoV-2 infections. J. Infect. 2025, 90, 106473. [Google Scholar] [CrossRef]
- Awaya, T.; Moroi, M.; Enomoto, Y.; Kunimasa, T.; Nakamura, M. What Should We Do after the COVID-19 Vaccination? Vaccine-Associated Diseases and Precautionary Measures against Adverse Reactions. Vaccines 2022, 10, 866. [Google Scholar] [CrossRef]
- Zagorec, N.; Horvatić, I.; Šenjug, P.; Horaček, M.; Ljubanović, D.G.; Galešić, K. Immune-mediated diseases after coronavirus disease 2019 vaccination: rare but important complication. Croat. Med J. 2022, 63, 389–393. [Google Scholar] [CrossRef]
- Franchini, M.; Liumbruno, G.M.; Pezzo, M. COVID-19 vaccine-associated immune thrombosis and thrombocytopenia (VITT): Diagnostic and therapeutic recommendations for a new syndrome. Eur. J. Haematol. 2021, 107, 173–180. [Google Scholar] [CrossRef]
- Leisman, D.E.; Ronner, L.; Pinotti, R.; Taylor, M.D.; Sinha, P.; Calfee, C.S.; Hirayama, A.V.; Mastroiani, F.; Turtle, C.J.; Harhay, M.O.; et al. Cytokine elevation in severe and critical COVID-19: a rapid systematic review, meta-analysis, and comparison with other inflammatory syndromes. Lancet Respir. Med. 2020, 8, 1233–1244. [Google Scholar] [CrossRef]
- Safary, A.; Esalatmanesh, K.; Eftekharsadat, A.T.; Nakjavani, M.-R.J.; Khabbazi, A. Autoimmune inflammatory rheumatic diseases post-COVID-19 vaccination. Int. Immunopharmacol. 2022, 110, 109061–109061. [Google Scholar] [CrossRef]
- Keijzer, S.; Oskam, N.; Heer, P.O.-D.; Steenhuis, M.; Keijser, J.B.; Wieske, L.; van Dam, K.P.; Stalman, E.W.; Kummer, L.Y.; Boekel, L.; et al. Longitudinal rheumatoid factor autoantibody responses after SARS-CoV-2 vaccination or infection. Front. Immunol. 2024, 15, 1314507. [Google Scholar] [CrossRef]
- Hileman, C.O.; Malakooti, S.K.; Patil, N.; Singer, N.G.; McComsey, G.A. New-onset autoimmune disease after COVID-19. Front. Immunol. 2024, 15, 1337406. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, Z.; Wang, P.; Li, X.; Shuai, Z.; Ye, D.; Pan, H. New-onset autoimmune phenomena post-COVID-19 vaccination. Immunology 2022, 165, 386–401. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, H.; Xia, S.-L. New-Onset Arthritis Following COVID-19 Vaccination: A Systematic Review of Case Reports. Vaccines 2023, 11, 665. [Google Scholar] [CrossRef] [PubMed]
- Sagy, I.; Zeller, L.; Raviv, Y.; Porges, T.; Bieber, A.; Abu-Shakra, M. New-onset systemic lupus erythematosus following BNT162b2 mRNA COVID-19 vaccine: a case series and literature review. Rheumatol. Int. 2022, 42, 2261–2266. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Liu, X.; Chen, X.; Li, Q. Insights into new-onset autoimmune diseases after COVID-19 vaccination. Autoimmun. Rev. 2023, 22, 103340–103340. [Google Scholar] [CrossRef]
- Mehta, P.; Fajgenbaum, D.C. Is severe COVID-19 a cytokine storm syndrome: a hyperinflammatory debate. Curr. Opin. Rheumatol. 2021, 33, 419–430. [Google Scholar] [CrossRef]
- Diao, B.; Wang, C.; Tan, Y.; Chen, X.; Liu, Y.; Ning, L.; Chen, L.; Li, M.; Liu, Y.; Wang, G.; et al. Reduction and Functional Exhaustion of T Cells in Patients With Coronavirus Disease 2019 (COVID-19). Front. Immunol. 2020, 11, 827. [Google Scholar] [CrossRef]
- Mathew, D.; Giles, J.R.; Baxter, A.E.; Oldridge, D.A.; Greenplate, A.R.; Wu, J.E.; Alanio, C.; Kuri-Cervantes, L.; Pampena, M.B.; D’aNdrea, K.; et al. Deep immune profiling of COVID-19 patients reveals distinct immunotypes with therapeutic implications. Science 2020, 369, 1209-+. [Google Scholar] [CrossRef]
- DiPiazza, A.T.; Graham, B.S.; Ruckwardt, T.J. T cell immunity to SARS-CoV-2 following natural infection and vaccination. Biochem. Biophys. Res. Commun. 2021, 538, 211–217. [Google Scholar] [CrossRef]
- Olszewska, B.; Zaryczańska, A.; Nowicki, R.J.; Sokołowska-Wojdyło, M. Rare COVID-19 vaccine side effects got lost in the shuffle. Primary cutaneous lymphomas following COVID-19 vaccination: a systematic review. Front. Med. 2024, 11, 1325478. [Google Scholar] [CrossRef]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.-H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V.; et al. Auto-antibodies against type I IFNs in patients with life-threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef] [PubMed]
- Puel, A.; Bastard, P.; Bustamante, J.; Casanova, J.-L. Human autoantibodies underlying infectious diseases. J. Exp. Med. 2022, 219. [Google Scholar] [CrossRef]
- Combes, A.J.; Courau, T.; Kuhn, N.F.; Hu, K.H.; Ray, A.; Chen, W.S.; Chew, N.W.; Cleary, S.J.; Kushnoor, D.; Reeder, G.C.; et al. Global absence and targeting of protective immune states in severe COVID-19. Nature 2021, 591, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Ning, W.; Xu, W.; Cong, X.; Fan, H.; Gilkeson, G.; Wu, X.; Hughes, H.; Jiang, W. COVID-19 mRNA vaccine BNT162b2 induces autoantibodies against type I interferons in a healthy woman. J. Autoimmun. 2022, 132, 102896–102896. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Wen, X.; Cong, X.; Jiang, W. COVID-19 mRNA vaccine, but not a viral vector-based vaccine, promotes neutralizing anti-type I interferon autoantibody production in a small group of healthy individuals. J. Med Virol. 2023, 95, e29137. [Google Scholar] [CrossRef]
- Kim, H.; Ahn, H.-S.; Hwang, N.; Huh, Y.; Bu, S.; Seo, K.J.; Kwon, S.H.; Lee, H.-K.; Kim, J.-W.; Yoon, B.K.; et al. Epigenomic landscape exhibits interferon signaling suppression in the patient of myocarditis after BNT162b2 vaccination. Sci. Rep. 2023, 13, 1–11. [Google Scholar] [CrossRef]
- Wahl, I.; Wardemann, H. Sterilizing immunity: Understanding COVID-19. Immunity 2022, 55, 2231–2235. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Goldberg, Y.; Mandel, M.; Bodenheimer, O.; Amir, O.; Freedman, L.; Alroy-Preis, S.; Ash, N.; Huppert, A.; Milo, R. Protection by a Fourth Dose of BNT162b2 against Omicron in Israel. New Engl. J. Med. 2022, 386, 1712–1720. [Google Scholar] [CrossRef]
- Ophir Y, Shira-Raz Y, Zakov S, McCullough PA. The efficacy of COVID-19 vaccine boosters against severe illness and deaths scientific fact or wishful myth? J Am Phys Surg. 2023; 28: 20-7. https://www.jpands.org/search-results.htm.
- Ioannidis, J.P.A. Estimating conditional vaccine effectiveness. Eur. J. Epidemiology 2022, 37, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Neil M, Fenton NE, McLachlan S. The extent and impact of vaccine status miscategorisation on covid-19 vaccine efficacy studies. MedRxiv Preprint. 2024. [CrossRef]
- Fung, K.; Jones, M.; Doshi, P. Sources of bias in observational studies of covid-19 vaccine effectiveness. J. Evaluation Clin. Pr. 2023, 30, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Doshi, P.; Fung, K. How the case counting window affected vaccine efficacy calculations in randomized trials of COVID-19 vaccines. J. Evaluation Clin. Pr. 2023, 30, 105–106. [Google Scholar] [CrossRef] [PubMed]
- Lataster, R. How the adverse effect counting window affected vaccine safety calculations in randomised trials of COVID-19 vaccines. J. Evaluation Clin. Pr. 2024, 30, 453–458. [Google Scholar] [CrossRef]
- Lataster, R. Science summary: COVID-19 vaccines’ effectiveness and safety exaggerated in clinical trials & observational studies, academics find. Okay Then News Substack. 28 Feb. 2024. URL: https://okaythennews.substack.com/p/science-summary-covid-19-vaccines.
- Gazit, S.; Shlezinger, R.; Perez, G.; Lotan, R.; Peretz, A.; Ben-Tov, A.; Herzel, E.; Alapi, H.; Cohen, D.; Muhsen, K.; et al. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Naturally Acquired Immunity versus Vaccine-induced Immunity, Reinfections versus Breakthrough Infections: A Retrospective Cohort Study. Clin. Infect. Dis. 2022, 75, e545–e551. [Google Scholar] [CrossRef]
- Rzymski, P.; Pazgan-Simon, M.; Simon, K.; Łapiński, T.; Zarębska-Michaluk, D.; Szczepańska, B.; Chojnicki, M.; Mozer-Lisewska, I.; Flisiak, R. Clinical Characteristics of Hospitalized COVID-19 Patients Who Received at Least One Dose of COVID-19 Vaccine. Vaccines 2021, 9, 781. [Google Scholar] [CrossRef]
- Adhikari, B.; Bednash, J.S.; Horowitz, J.C.; Rubinstein, M.P.; Vlasova, A.N. Brief research report: impact of vaccination on antibody responses and mortality from severe COVID-19. Front. Immunol. 2024, 15, 1325243. [Google Scholar] [CrossRef]
- Heymans, S.; Dawson, D.; Fuster, V.; Metra, M.; Tocchetti, C.G. Myocarditis Following SARS-CoV2 mRNA Vaccination Against COVID-19. Circ. 2022, 80, 1363–1365. [Google Scholar] [CrossRef]
- Block, J.P. Cardiac Complications After SARS-CoV-2 Infection and mRNA COVID-19 Vaccination — PCORnet, United States, January 2021–January 2022. Mmwr-Morbidity Mortal. Wkly. Rep. 2022, 71, 517–523. [Google Scholar] [CrossRef]
- Holland, D.J.; Blazak, P.L.; Martin, J.; Broom, J.; Poulter, R.S.; Stanton, T. Myocarditis and Cardiac Complications Associated With COVID-19 and mRNA Vaccination: A Pragmatic Narrative Review to Guide Clinical Practice. Hear. Lung Circ. 2022, 31, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Mead, M.N. Myocarditis after SARS-CoV-2 infection and COVID-19 vaccination: Epidemiology, outcomes, and new perspectives. Int J Cardiovasc Res Innov. 2025, 3, 1–43. [Google Scholar] [CrossRef]
- Flaxman, S.; Whittaker, C.; Semenova, E.; Rashid, T.; Parks, R.M.; Blenkinsop, A.; Unwin, H.J.T.; Mishra, S.; Bhatt, S.; Gurdasani, D.; et al. Assessment of COVID-19 as the Underlying Cause of Death Among Children and Young People Aged 0 to 19 Years in the US. JAMA Netw. Open 2023, 6, e2253590–e2253590. [Google Scholar] [CrossRef] [PubMed]
- Thornley S, Morris AJ, Sundborn G, S Bailey. How fatal is COVID-19 compared with seasonal influenza? The devil is in the detail [Rapid Response]. BMJ 2020; 371: m3883. [CrossRef]
- Pilz, S.; Theiler-Schwetz, V.; Trummer, C.; Krause, R.; Ioannidis, J.P. SARS-CoV-2 reinfections: Overview of efficacy and duration of natural and hybrid immunity. Environ. Res. 2022, 209, 112911–112911. [Google Scholar] [CrossRef]
- Spinardi, J.R.; Srivastava, A. Hybrid Immunity to SARS-CoV-2 from Infection and Vaccination—Evidence Synthesis and Implications for New COVID-19 Vaccines. Biomedicines 2023, 11, 370. [Google Scholar] [CrossRef] [PubMed]
- Epsi, N.J.; A Richard, S.; A Lindholm, D.; Mende, K.; Ganesan, A.; Huprikar, N.; Lalani, T.; Fries, A.C.; Maves, R.C.; E Colombo, R.; et al. Understanding “Hybrid Immunity”: Comparison and Predictors of Humoral Immune Responses to Severe Acute Respiratory Syndrome Coronavirus 2 Infection (SARS-CoV-2) and Coronavirus Disease 2019 (COVID-19) Vaccines. Clin. Infect. Dis. 2022, 76, e439–e449. [Google Scholar] [CrossRef]
- Klee, B.; Diexer, S.; Xu, C.; Gottschick, C.; Hartmann, C.; Meyer-Schlinkmann, K.M.; Kuhlmann, A.; Rosendahl, J.; Binder, M.; Gekle, M.; et al. Household transmission of Omicron variant of SARS-CoV-2 under conditions of hybrid immunity—a prospective study in Germany. Infection 2024, 53, 221–230. [Google Scholar] [CrossRef]
- Bigay, J.; Le Grand, R.; Martinon, F.; Maisonnasse, P. Vaccine-associated enhanced disease in humans and animal models: Lessons and challenges for vaccine development. Front. Microbiol. 2022, 13, 932408. [Google Scholar] [CrossRef]
- Gartlan, C.; Tipton, T.; Salguero, F.J.; Sattentau, Q.; Gorringe, A.; Carroll, M.W. Vaccine-Associated Enhanced Disease and Pathogenic Human Coronaviruses. Front. Immunol. 2022, 13, 882972. [Google Scholar] [CrossRef]
- Ebenig, A.; Muraleedharan, S.; Kazmierski, J.; Todt, D.; Auste, A.; Anzaghe, M.; Gömer, A.; Postmus, D.; Gogesch, P.; Niles, M.; et al. Vaccine-associated enhanced respiratory pathology in COVID-19 hamsters after TH2-biased immunization. Cell Rep. 2022, 40, 111214–111214. [Google Scholar] [CrossRef]
- Tunjungputri, R.N.; Tetrasiwi, E.N.; Veronica, M.; Pandelaki, J.; Ibrahim, F.; Nelwan, E.J.; Supinski, G.S. Vaccine-Associated Disease Enhancement (VADE): Considerations in Postvaccination COVID-19. Case Rep. Med. 2021, 2021, 1–5. [Google Scholar] [CrossRef]
- Boyce, T.G.; McClure, D.L.; Hanson, K.E.; Daley, M.F.; DeSilva, M.B.; Irving, S.A.; Jackson, L.A.; Klein, N.P.; Lewin, B.; Williams, J.T.B.; et al. Lack of Evidence for Vaccine-Associated Enhanced Disease From COVID-19 Vaccines Among Adults in the Vaccine Safety Datalink. Pharmacoepidemiol. Drug Saf. 2024, 33, e5863. [Google Scholar] [CrossRef]
- Parameswaran, A.; Apsingi, S.; Eachempati, K.K.; Dannana, C.S.; Jagathkar, G.; Iyer, M.; Aribandi, H. Incidence and severity of COVID-19 infection post-vaccination: a survey among Indian doctors. Infection 2022, 50, 889–895. [Google Scholar] [CrossRef]
- Grasselli, G.; Zanella, A.; Carlesso, E.; Florio, G.; Canakoglu, A.; Bellani, G.; Bottino, N.; Cabrini, L.; Castelli, G.P.; Catena, E.; et al. Association of COVID-19 Vaccinations With Intensive Care Unit Admissions and Outcome of Critically Ill Patients With COVID-19 Pneumonia in Lombardy, Italy. JAMA Netw. Open 2022, 5, e2238871–e2238871. [Google Scholar] [CrossRef]
- Chatterjee, S.; Nalla, L.V.; Sharma, M.; Sharma, N.; Singh, A.A.; Malim, F.M.; Ghatage, M.; Mukarram, M.; Pawar, A.; Parihar, N.; et al. Association of COVID-19 with Comorbidities: An Update. ACS Pharmacol. Transl. Sci. 2023, 6, 334–354. [Google Scholar] [CrossRef]
- Cao, C.; Cai, Z.; Xiao, X.; Rao, J.; Chen, J.; Hu, N.; Yang, M.; Xing, X.; Wang, Y.; Li, M.; et al. The architecture of the SARS-CoV-2 RNA genome inside virion. Nat. Commun. 2021, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hamedi, K.R.; Loftus, G.; Traylor, L.; Goodwin, R.; Arce, S. Comparison of COVID-19 Vaccine-Associated Myocarditis and Viral Myocarditis Pathology. Vaccines 2023, 11, 362. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, Y.; Rojas, M.; Beltrán, S.; Polo, F.; Camacho-Domínguez, L.; Morales, S.D.; Gershwin, M.E.; Anaya, J.-M. Autoimmune and autoinflammatory conditions after COVID-19 vaccination. New case reports and updated literature review. J. Autoimmun. 2022, 132, 102898–102898. [Google Scholar] [CrossRef] [PubMed]
- Rojas, M.; Herrán, M.; Ramírez-Santana, C.; Leung, P.S.; Anaya, J.-M.; Ridgway, W.M.; Gershwin, M.E. Molecular mimicry and autoimmunity in the time of COVID-19. J. Autoimmun. 2023, 139, 103070–103070. [Google Scholar] [CrossRef]
- Talotta, R. Do COVID-19 RNA-based vaccines put at risk of immune-mediated diseases? In reply to “potential antigenic cross-reactivity between SARS-CoV-2 and human tissue with a possible link to an increase in autoimmune diseases”. Clin. Immunol. 2021, 224, 108665–108665. [Google Scholar] [CrossRef]
- Akinosoglou, K.; Tzivaki, I.; Marangos, M. Covid-19 vaccine and autoimmunity: Awakening the sleeping dragon. Clin Immunol. 2021, 226, 108721–108721. [Google Scholar] [CrossRef]
- Polykretis, P.; Donzelli, A.; Lindsay, J.C.; Wiseman, D.; Kyriakopoulos, A.M.; Mörz, M.; Bellavite, P.; Fukushima, M.; Seneff, S.; McCullough, P.A. Autoimmune inflammatory reactions triggered by the COVID-19 genetic vaccines in terminally differentiated tissues. Autoimmunity 2023, 56, 2259123. [Google Scholar] [CrossRef]
- Wang, L.; Davis, P.B.; Kaelber, D.C.; Volkow, N.D.; Xu, R. Comparison of mRNA-1273 and BNT162b2 Vaccines on Breakthrough SARS-CoV-2 Infections, Hospitalizations, and Death During the Delta-Predominant Period. JAMA 2022, 327, 678–680. [Google Scholar] [CrossRef]
- Beatty, A.L.; Peyser, N.D.; Butcher, X.E.; Cocohoba, J.M.; Lin, F.; Olgin, J.E.; Pletcher, M.J.; Marcus, G.M. Analysis of COVID-19 Vaccine Type and Adverse Effects Following Vaccination. JAMA Netw. Open 2021, 4, e2140364–e2140364. [Google Scholar] [CrossRef]
- Kitagawa, H.; Kaiki, Y.; Sugiyama, A.; Nagashima, S.; Kurisu, A.; Nomura, T.; Omori, K.; Akita, T.; Shigemoto, N.; Tanaka, J.; et al. Adverse reactions to the BNT162b2 and mRNA-1273 mRNA COVID-19 vaccines in Japan. J. Infect. Chemother. 2022, 28, 576–581. [Google Scholar] [CrossRef]
- Valera-Rubio, M. (.; Sierra-Torres, M.I.(.; García, R.(.C.; Cordero-Ramos, J.(.; López-Márquez, M.R.(.; Cruz-Salgado, Ó.(.; Calleja-Hernández, M.Á.(. Adverse events reported after administration of BNT162b2 and mRNA-1273 COVID-19 vaccines among hospital workers: a cross-sectional survey-based study in a Spanish hospital. Expert Rev. Vaccines 2022, 21, 533–540. [Google Scholar] [CrossRef]
- Chapin-Bardales, J.; Gee, J.; Myers, T. Reactogenicity Following Receipt of mRNA-Based COVID-19 Vaccines. JAMA 2021, 325, 2201–2202. [Google Scholar] [CrossRef]
- Chapin-Bardales, J.; Myers, T.; Gee, J.; Shay, D.K.; Marquez, P.; Baggs, J.; Zhang, B.; Licata, C.; Shimabukuro, T.T. Reactogenicity within 2 weeks after mRNA COVID-19 vaccines: Findings from the CDC v-safe surveillance system. Vaccine 2021, 39, 7066–7073. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-H.; Chen, T.-A.; Chiang, P.-H.; Hsieh, A.-R.; Wu, B.-J.; Chen, P.-Y.; Lin, K.-C.; Tsai, Z.-S.; Lin, M.-H.; Chen, T.-J.; et al. Incidence and Nature of Short-Term Adverse Events following COVID-19 Second Boosters: Insights from Taiwan’s Universal Vaccination Strategy. Vaccines 2024, 12, 149. [Google Scholar] [CrossRef] [PubMed]
- Urdaneta, V.; Esposito, D.B.; Dharia, P.; Moraga, M.S.; Anteyi, K.; Oduyebo-Omotosho, T.; Rossi, M.; Burton, P.; Vega, J.M.; Dawson, R.; et al. Global Safety Assessment of Adverse Events of Special Interest Following 2 Years of Use and 772 Million Administered Doses of mRNA-1273. Open Forum Infect. Dis. 2024, 11, ofae067. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Casillas, A.; Redwan, E.M.; Uversky, V.N. More antibodies are not always better: Fc effector functions play a critical role in SARS-CoV-2 infection and protection. Prog Mol Biol Transl Sci. 2025;213:413-447. [CrossRef]
- Brisotto, G.; Montico, M.; Turetta, M.; Zanussi, S.; Cozzi, M.R.; Vettori, R.; Boschin, R.B.; Vinante, L.; Matrone, F.; Revelant, A.; et al. Integration of Cellular and Humoral Immune Responses as an Immunomonitoring Tool for SARS-CoV-2 Vaccination in Healthy and Fragile Subjects. Viruses 2023, 15, 1276. [Google Scholar] [CrossRef]
- Debes, A.K.; Xiao, S.; Colantuoni, E.; Egbert, E.R.; Caturegli, P.; Gadala, A.; Milstone, A.M. Association of Vaccine Type and Prior SARS-CoV-2 Infection With Symptoms and Antibody Measurements Following Vaccination Among Health Care Workers. JAMA Intern. Med. 2021, 181, 1660–1662. [Google Scholar] [CrossRef]
- Kobashi, Y.; Shimazu, Y.; Kawamura, T.; Nishikawa, Y.; Omata, F.; Kaneko, Y.; Kodama, T.; Tsubokura, M.; Yunihastuti, E. Factors associated with anti-severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) spike protein antibody titer and neutralizing activity among healthcare workers following vaccination with the BNT162b2 vaccine. PLOS ONE 2022, 17, e0269917. [Google Scholar] [CrossRef]
- Levy, I.; Levin, E.G.; Olmer, L.; Regev-Yochay, G.; Agmon-Levin, N.; Wieder-Finesod, A.; Indenbaum, V.; Herzog, K.; Doolman, R.; Asraf, K.; et al. Correlation between Adverse Events and Antibody Titers among Healthcare Workers Vaccinated with BNT162b2 mRNA COVID-19 Vaccine. Vaccines 2022, 10, 1220. [Google Scholar] [CrossRef] [PubMed]
- Naaber, P.; Tserel, L.; Kangro, K.; Sepp, E.; Jürjenson, V.; Adamson, A.; Haljasmägi, L.; Rumm, A.P.; Maruste, R.; Kärner, J.; et al. Dynamics of antibody response to BNT162b2 vaccine after six months: a longitudinal prospective study. Lancet Reg. Health-Eur. 2021, 10, 100208. [Google Scholar] [CrossRef] [PubMed]
- Pozdnyakova, V.; Weber, B.; Cheng, S.; Ebinger, J.E. Review of Immunologic Manifestations of COVID-19 Infection and Vaccination. Hear. Fail. Clin. 2023, 19, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Rechavi, Y.; Shashar, M.; Lellouche, J.; Yana, M.; Yakubovich, D.; Sharon, N. Occurrence of BNT162b2 Vaccine Adverse Reactions Is Associated with Enhanced SARS-CoV-2 IgG Antibody Response. Vaccines 2021, 9, 977. [Google Scholar] [CrossRef]
- Takeuchi, M.; Higa, Y.; Esaki, A.; Nabeshima, Y.; Nakazono, A.; Moreland, N.J. Does reactogenicity after a second injection of the BNT162b2 vaccine predict spike IgG antibody levels in healthy Japanese subjects? PLOS ONE 2021, 16, e0257668. [Google Scholar] [CrossRef]
- Uwamino, Y.; Kurafuji, T.; Sato, Y.; Tomita, Y.; Shibata, A.; Tanabe, A.; Yatabe, Y.; Noguchi, M.; Arai, T.; Ohno, A.; et al. Young age, female sex, and presence of systemic adverse reactions are associated with high post-vaccination antibody titer after two doses of BNT162b2 mRNA SARS-CoV-2 vaccination: An observational study of 646 Japanese healthcare workers and university staff. Vaccine 2022, 40, 1019–1025. [Google Scholar] [CrossRef]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: major findings, mechanisms and recommendations. Nat. Rev. Microbiol. 2023, 21, 133–146. [Google Scholar] [CrossRef]
- Yong, S.J.; Liu, S. Proposed subtypes of post-COVID-19 syndrome (or long-COVID) and their respective potential therapies. Rev. Med Virol. 2021, 32, e2315. [Google Scholar] [CrossRef]
- Raveendran, A.V.; Jayadevan, R.; Sashidharan, S. Long COVID: An overview. Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Lam, I.C.H.; Wong, C.K.H.; Zhang, R.; Chui, C.S.L.; Lai, F.T.T.; Li, X.; Chan, E.W.Y.; Luo, H.; Zhang, Q.; Man, K.K.C.; et al. Long-term post-acute sequelae of COVID-19 infection: a retrospective, multi-database cohort study in Hong Kong and the UK. eClinicalMedicine 2023, 60, 102000. [Google Scholar] [CrossRef] [PubMed]
- Arjun, M.C.; Singh, A.K.; Pal, D.; Das, K.; G. , A.; Venkateshan, M.; Mishra, B.; Patro, B.K.; Mohapatra, P.R.; Subba, S.H.; et al. Characteristics and predictors of Long COVID among diagnosed cases of COVID-19. PLOS ONE 2022, 17, e0278825. [Google Scholar] [CrossRef] [PubMed]
- Bocquet-Garçon, A. Impacts of the SARS-CoV-2 Spike Protein on the Innate Immune System: A Review. Cureus 2024, 16, e57008. [Google Scholar] [CrossRef]
- Craddock, V.; Mahajan, A.; Spikes, L.; Krishnamachary, B.; Ram, A.K.; Kumar, A.; Chen, L.; Chalise, P.; Dhillon, N.K. Persistent circulation of soluble and extracellular vesicle-linked Spike protein in individuals with postacute sequelae of COVID-19. J. Med Virol. 2023, 95, e28568–e28568. [Google Scholar] [CrossRef]
- Dhuli, K.; Medori, M.C.; Micheletti, C.; Donato, K.; Fioretti, F.; Calzoni, A.; Praderio, A.; De Angelis, M.G.; Arabia, G.; Cristoni, S.; et al. Presence of viral spike protein and vaccinal spike protein in the blood serum of patients with long-COVID syndrome (Retracted Article). 2023, 27, 13–19. [CrossRef]
- Vogel G, Couzin-Frankel J. Rare link between coronavirus vaccines and Long Covid-like illness starts to gain acceptance. Science 2023; 381: 6653. [CrossRef]
- Hulscher, N.; Procter, B.C.; Wynn, C.; A McCullough, P. Clinical Approach to Post-acute Sequelae After COVID-19 Infection and Vaccination. Cureus 2023, 15, e49204. [Google Scholar] [CrossRef]
- White JR, Abraham RL, Coleman WT, Pitre E, Stevenson MM, Kaplan JL, Goldberg AG, et al. SARS-CoV-2 Semi-Quantitative Total Antibody Correlates with Symptoms of Long COVID in Both Vaccinated and Unvaccinated Subjects. Preprints.org. Posted:. 16 July 2025. [CrossRef]
- Diexer, S.; Klee, B.; Gottschick, C.; Xu, C.; Broda, A.; Purschke, O.; Binder, M.; Frese, T.; Girndt, M.; Hoell, J.I.; et al. Association between virus variants, vaccination, previous infections, and post-COVID-19 risk. Int. J. Infect. Dis. 2023, 136, 14–21. [Google Scholar] [CrossRef]
- Bhargava A, Inslicht, S. Postacute sequelae of SARS-CoV-2 in the population: Risk factors and vaccines. Preprint. [CrossRef]
- . [CrossRef]
- Scholkmann, F.; May, C.-A. COVID-19, post-acute COVID-19 syndrome (PACS, “long COVID”) and post-COVID-19 vaccination syndrome (PCVS, “post-COVIDvac-syndrome”): Similarities and differences. Pathol. - Res. Pr. 2023, 246, 154497–154497. [Google Scholar] [CrossRef]
- See, I.; Lale, A.; Marquez, P.; Streiff, M.B.; Wheeler, A.P.; Tepper, N.K.; Woo, E.J.; Broder, K.R.; Edwards, K.M.; Gallego, R.; et al. Case Series of Thrombosis With Thrombocytopenia Syndrome After COVID-19 Vaccination—United States, December 2020 to August 2021. Ann. Intern. Med. 2022, 175, 513–522. [Google Scholar] [CrossRef]
- Sangli, S.; Virani, A.; Cheronis, N.; Vannatter, B.; Minich, C.; Noronha, S.; Bhagavatula, R.; Speredelozzi, D.; Sareen, M.; Kaplan, R.B. Thrombosis With Thrombocytopenia After the Messenger RNA–1273 Vaccine. Ann. Intern. Med. 2021, 174, 1480–1482. [Google Scholar] [CrossRef]
- Jabagi, M.J.; Botton, J.; Bertrand, M.; Weill, A.; Farrington, P.; Zureik, M.; Dray-Spira, R. Myocardial Infarction, Stroke, and Pulmonary Embolism After BNT162b2 mRNA COVID-19 Vaccine in People Aged 75 Years or Older. JAMA 2021, 327, 80–82. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Y.; Wang, X.; Yang, L.; Li, H.; Wang, Y.; Liu, M.; Zhao, X.; Xie, Y.; Yang, Y.; et al. SARS-CoV-2 binds platelet ACE2 to enhance thrombosis in COVID-19. J. Hematol. Oncol. 2020, 13, 120. [Google Scholar] [CrossRef]
- Liu, R.; Pan, J.; Zhang, C.; Sun, X. Cardiovascular Complications of COVID-19 Vaccines. Front. Cardiovasc. Med. 2022, 9, 840929. [Google Scholar] [CrossRef]
- Conway, E.M.; Mackman, N.; Warren, R.Q.; Wolberg, A.S.; Mosnier, L.O.; Campbell, R.A.; Gralinski, L.E.; Rondina, M.T.; van de Veerdonk, F.L.; Hoffmeister, K.M.; et al. Understanding COVID-19-associated coagulopathy. Nat. Rev. Immunol. 2022, 22, 639–649. [Google Scholar] [CrossRef]
- Hoteit, L.; Deeb, A.-P.; Andraska, E.A.; Kaltenmeier, C.; Yazdani, H.O.; Tohme, S.; Neal, M.D.; Mota, R.I. The Pathobiological Basis for Thrombotic Complications in COVID-19: a Review of the Literature. Curr. Pathobiol. Rep. 2021, 9, 107–117. [Google Scholar] [CrossRef]
- Fan, B.E.; Shen, J.Y.; Lim, X.R.; Tu, T.M.; Chang, C.C.R.; Khin, H.S.W.; Koh, J.S.; Rao, J.P.; Lau, S.L.; Tan, G.B.; et al. Cerebral venous thrombosis post BNT162b2 mRNA SARS-CoV -2 vaccination: A black swan event. Am. J. Hematol. 2021, 96, E357–E361. [Google Scholar] [CrossRef]
- Bilotta, C.; Perrone, G.; Adelfio, V.; Spatola, G.F.; Uzzo, M.L.; Argo, A.; Zerbo, S. COVID-19 Vaccine-Related Thrombosis: A Systematic Review and Exploratory Analysis. Front. Immunol. 2021, 12, 729251. [Google Scholar] [CrossRef]
- Padilla-Flores, T.; Sampieri, A.; Vaca, L. Incidence and management of the main serious adverse events reported after COVID-19 vaccination. Pharmacol. Res. Perspect. 2024, 12, e1224. [Google Scholar] [CrossRef]
- Iba, T.; Levy, J.H. The roles of platelets in COVID-19-associated coagulopathy and vaccine-induced immune thrombotic thrombocytopenia. Trends Cardiovasc. Med. 2021, 32, 1–9. [Google Scholar] [CrossRef]
- Schreckenberg, R.; Woitasky, N.; Itani, N.; Czech, L.; Ferdinandy, P.; Schulz, R. Cardiac side effects of RNA-based SARS-CoV-2 vaccines: Hidden cardiotoxic effects of mRNA-1273 and BNT162b2 on ventricular myocyte function and structure. Br. J. Pharmacol. 2023, 181, 345–361. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. COVID-19 VaST Work Group Report – May 17, 2021. 2021. [cited 2023 January 5]; Available from: https://www.cdc.gov/vaccines/acip/work-groups-vast/report-2021-05-17.html. 17 May.
- Anonymous. ACC Underscores Safety of COVID-19 Vaccine. ACC News Story.
- Oct 14, 2022. https://www.acc.org/Latest-in-Cardiology/Articles/2022/10/14/15/13/ACC-Underscores-Safety-of-COVID-19-Vaccine. 2022.
- Rose, J.; Hulscher, N.; McCullough, P.A. Determinants of COVID-19 vaccine-induced myocarditis. Ther. Adv. Drug Saf. 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Lindner, D.; Fitzek, A.; Bräuninger, H.; Aleshcheva, G.; Edler, C.; Meissner, K.; Scherschel, K.; Kirchhof, P.; Escher, F.; Schultheiss, H.-P.; et al. Association of Cardiac Infection With SARS-CoV-2 in Confirmed COVID-19 Autopsy Cases. JAMA Cardiol. 2020, 5, 1281–1285. [Google Scholar] [CrossRef] [PubMed]
- Buergin, N.; Lopez-Ayala, P.; Hirsiger, J.R.; Mueller, P.; Median, D.; Glarner, N.; Rumora, K.; Herrmann, T.; Koechlin, L.; Haaf, P.; et al. Sex-specific differences in myocardial injury incidence after COVID-19 mRNA-1273 booster vaccination. Eur. J. Hear. Fail. 2023, 25, 1871–1881. [Google Scholar] [CrossRef] [PubMed]
- Mansanguan, S.; Charunwatthana, P.; Piyaphanee, W.; Dechkhajorn, W.; Poolcharoen, A.; Mansanguan, C. Cardiovascular Manifestation of the BNT162b2 mRNA COVID-19 Vaccine in Adolescents. Trop. Med. Infect. Dis. 2022, 7, 196. [Google Scholar] [CrossRef]
- Daniels, C.J.; Rajpal, S.; Greenshields, J.T.; Rosenthal, G.L.; Chung, E.H.; Terrin, M.; Jeudy, J.; Mattson, S.E.; Law, I.H.; Borchers, J.; et al. Prevalence of Clinical and Subclinical Myocarditis in Competitive Athletes With Recent SARS-CoV-2 Infection. JAMA Cardiol. 2021, 6, 1078–1087. [Google Scholar] [CrossRef]
- Foltran, D.; Delmas, C.; Flumian, C.; De Paoli, P.; Salvo, F.; Gautier, S.; Drici, M.-D.; Karsenty, C.; Montastruc, F. Myocarditis and pericarditis in adolescents after first and second doses of mRNA COVID-19 vaccines. Eur. Hear. J. - Qual. Care Clin. Outcomes 2021, 8, 99–103. [Google Scholar] [CrossRef]
- Karlstad, Ø.; Hovi, P.; Husby, A.; Härkänen, T.; Selmer, R.M.; Pihlström, N.; Hansen, J.V.; Nohynek, H.; Gunnes, N.; Sundström, A.; et al. SARS-CoV-2 Vaccination and Myocarditis in a Nordic Cohort Study of 23 Million Residents. JAMA Cardiol. 2022, 7, 600–612. [Google Scholar] [CrossRef]
- Buchan, S.A.; Seo, C.Y.; Johnson, C.; Alley, S.; Kwong, J.C.; Nasreen, S.; Calzavara, A.; Lu, D.; Harris, T.M.; Yu, K.; et al. Epidemiology of Myocarditis and Pericarditis Following mRNA Vaccination by Vaccine Product, Schedule, and Interdose Interval Among Adolescents and Adults in Ontario, Canada. JAMA Netw. Open 2022, 5, e2218505–e2218505. [Google Scholar] [CrossRef]
- Gao, J.; Feng, L.; Li, Y.; Lowe, S.; Guo, Z.; Bentley, R.; Xie, C.; Wu, B.; Xie, P.; Xia, W.; et al. A Systematic Review and Meta-analysis of the Association Between SARS-CoV-2 Vaccination and Myocarditis or Pericarditis. Am. J. Prev. Med. 2022, 64, 275–284. [Google Scholar] [CrossRef]
- Munro, A.P.S.; Feng, S.; Janani, L.; Cornelius, V.; Aley, P.K.; Babbage, G.; Baxter, D.; Bula, M.; Cathie, K.; Chatterjee, K.; et al. Safety, immunogenicity, and reactogenicity of BNT162b2 and mRNA-1273 COVID-19 vaccines given as fourth-dose boosters following two doses of ChAdOx1 nCoV-19 or BNT162b2 and a third dose of BNT162b2 (COV-BOOST): a multicentre, blinded, phase 2, randomised trial. Lancet Infect. Dis. 2022, 22, 1131–1141. [Google Scholar] [CrossRef]
- Bohné, M.; Bohnen, S.; Willems, S.; Klingel, K.; Kivelitz, D.; Bahlmann, E.; Shirotani, M. Acute Lymphocytic Myocarditis in a Young Male Post-COVID-19. Case Rep. Cardiol. 2023, 2023, 1–7. [Google Scholar] [CrossRef]
- Fishman, B.; Goitein, O.; Berkovitch, A.; Rahav, G.; Matetzky, S.; Vandenbriele, C.; Abela, M.; Alvarez, C.; Liu, Z.; Vervaat, F. First report of myocarditis in two patients with COVID-19 Omicron variant: case report. Eur. Hear. J. - Case Rep. 2022, 6, ytac407. [Google Scholar] [CrossRef] [PubMed]
- Patrignani, A.; Schicchi, N.; Calcagnoli, F.; Falchetti, E.; Ciampani, N.; Argalia, G.; Mariani, A. Acute myocarditis following Comirnaty vaccination in a healthy man with previous SARS-CoV-2 infection. Radiol. Case Rep. 2021, 16, 3321–3325. [Google Scholar] [CrossRef] [PubMed]
- Shime, M.; Nozaki, Y.; Morita, A.; Ishiodori, T.; Murakami, T.; Yamasaki, H.; Yamamoto, M.; Takada, H. Life-threatening severe acute respiratory syndrome coronavirus-2 mRNA vaccine-associated myocarditis after COVID-19 myocarditis. J. Paediatr. Child Heal. 2023, 59, 1319–1322. [Google Scholar] [CrossRef] [PubMed]
- Etuk, A.S.; Jackson, I.N.; Panayiotou, H. A Rare Case of Myocarditis After the First Dose of Moderna Vaccine in a Patient With Two Previous COVID-19 Infections. Cureus 2022, 14, e24802. [Google Scholar] [CrossRef]
- Zaveri, S.; Tagliaferri, A.R.; Woldemariam, S.; Aron, P.; Palacios, C.; Melki, G.; Michael, P. A Case of Multifactorial Viral Myocarditis. Cureus 2021, 13. [Google Scholar] [CrossRef]
- Gholipour, M.; Samidoost, P.; Moayerifar, M.; Ghasemzadeh, G. A case report of QTc prolongation: Drug induced or myocarditis in Severe Acute Respiratory Syndrome Coronavirus 2. SAGE Open Med Case Rep. 2024, 12. [Google Scholar] [CrossRef]
- Al Harbi, S.; AlFaifi, M.; Al-Dorzi, H.M.; Aljuhani, O.; Alenazi, A.A.; Alalawi, M.; Al Sulaiman, K. A case report on the association between QTc prolongation and remdesivir therapy in a critically ill patient. IDCases 2022, 29, e01572. [Google Scholar] [CrossRef]
- Nabati, M.; Parsaee, H. Potential Cardiotoxic Effects of Remdesivir on Cardiovascular System: A Literature Review. Cardiovasc. Toxicol. 2021, 22, 268–272. [Google Scholar] [CrossRef]
- Patone, M.; Mei, X.W.; Handunnetthi, L.; Dixon, S.; Zaccardi, F.; Shankar-Hari, M.; Watkinson, P.; Khunti, K.; Harnden, A.; Coupland, C.A.; et al. Risk of Myocarditis After Sequential Doses of COVID-19 Vaccine and SARS-CoV-2 Infection by Age and Sex. Circulation 2022, 146, 743–754. [Google Scholar] [CrossRef]
- Bourdon, P.S.; Pantazatos, S.P. Why a major study on myocarditis risk following COVID vaccination should not influence public-health policy. Front. Med. 2023, 10, 1126945. [Google Scholar] [CrossRef] [PubMed]
- Stowe, J.; Miller, E.; Andrews, N.; Whitaker, H.J. Risk of myocarditis and pericarditis after a COVID-19 mRNA vaccine booster and after COVID-19 in those with and without prior SARS-CoV-2 infection: A self-controlled case series analysis in England. PLOS Med. 2023, 20, e1004245. [Google Scholar] [CrossRef] [PubMed]
- Blasco, A.; Royuela, A.; García-Gómez, S.; Gómez-Lozano, N.; Sánchez-Arjona, A.; de la Fuente, J.; Anel, J.; Sánchez-Galarraga, I.; Pérez-Redondo, M.; González, E.; et al. Association of SARS-CoV-2 immunoserology and vaccination status with myocardial infarction severity and outcome. Vaccine 2024, 42, 126305. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.; Lee, Y.; Heo, S.-J.; Kim, N.; Jung, I. The impact of COVID-19 status and vaccine type following the first dose on acute heart disease: A nationwide retrospective cohort study in South Korea. Epidemiology Infect. 2024, 152, e134. [Google Scholar] [CrossRef]
- Cianci, R.; Caldarelli, M.; Rio, P.; Pignataro, G.; Fernandez, M.S.; Ocarino, F.; Della Polla, D.A.; Franceschi, F.; Gasbarrini, A.; Gambassi, G.; et al. Outcomes of Patients with Heart Failure Hospitalized for COVID-19—A Study in a Tertiary Italian Center. Diseases 2024, 12, 337. [Google Scholar] [CrossRef]
- Al-Aly, Z.; Bowe, B.; Xie, Y. Long COVID after breakthrough SARS-CoV-2 infection. Nat. Med. 2022, 28, 1461–1467. [Google Scholar] [CrossRef]
- Our World In Data (OWID). Accessed 8 May 2025. https://ourworldindata.org/grapher/covid-vaccinationdoses-per-capita.
- Ndugga N, Hill L, Artiga S. COVID-19 Cases and Deaths, Vaccinations, and Treatments by Race/Ethnicity as of Fall 2022. KFF. 17 November 2022.
- https://www.kff.org/racial-equity-and-health-policy/issue-brief/covid-19-cases-and-deaths-vaccinationsand-treatments-by-race-ethnicity-as-of-fall-2022/.
- Goldstein, E. Mortality associated with Omicron and influenza infections in France before and during the COVID-19 pandemic. Epidemiology Infect. 2023, 151, 1–22. [Google Scholar] [CrossRef]
- Müller, S.; Schmetz, A.; Knaul, J.K.; Wilke, T.; Yang, J.; Dornig, S.; Lehmann, C.; Spinner, C.D. COVID-19 Disease Burden in the Omicron Variant-Dominated Endemic Phase: Insights from the ROUTINE-COV19 Study Using Real-World German Statutory Health Insurance Data. Viruses 2025, 17, 424. [Google Scholar] [CrossRef]
- Forthun, I.; Madsen, C.; Emilsson, L.; Nilsson, A.; Kepp, K.P.; Björk, J.; Vollset, S.E.; Lallukka, T.; Knudsen, A.K.S. Excess mortality in Denmark, Finland, Norway and Sweden during the COVID-19 pandemic 2020–2022. Eur. J. Public Heal. 2024, 34, 737–743. [Google Scholar] [CrossRef]
- Jang, M.; Kim, S.; Choi, S.; Ryu, B.; Choi, S.Y.; Choi, S.; An, M.; Kim, S.-S. Estimating Excess Mortality During the COVID-19 Pandemic Between 2020–2022 in Korea. J. Korean Med Sci. 2024, 39, e267. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Jang, H.; Oh, J. Excess mortality during the Coronavirus disease pandemic in Korea. BMC Public Heal. 2023, 23, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Li, Y.; Zi, Y.; Zhu, Y. The shift of percent excess mortality from zero-COVID policy to living-with-COVID policy in Singapore, South Korea, Australia, New Zealand and Hong Kong SAR. Front. Public Heal. 2023, 11, 1085451. [Google Scholar] [CrossRef]
- Scherb, H.; Hayashi, K.; Nat. , 1.D.-M.D.R.; (Md), H.C.C.2.M.D. Annual All-Cause Mortality Rate in Germany and Japan (2005 to 2022) With Focus on The Covid-19 Pandemic: Hypotheses And Trend Analyses. Med. Clin. Sci. 2023, 5. [Google Scholar] [CrossRef]
- Durante, A.C.D.P.; Lacaza, R.; Lapitan, P.; Kochhar, N.; Tan, E.S.; Thomas, M. Mixed effects modelling of excess mortality and COVID-19 lockdowns in Thailand. Sci. Rep. 2024, 14, 1–15. [Google Scholar] [CrossRef]
- Okoro, E.; A Ikoba, N.; E Okoro, B.; Akpila, A.S.; O Salihu, M. Paradoxical increase in global COVID-19 deaths with vaccination coverage: World Health Organization estimates (2020–2023). Int. J. Risk Saf. Med. 2025. [Google Scholar] [CrossRef]
- Röltgen, K.; Boyd, S.D. Antibody and B Cell Responses to SARS-CoV-2 Infection and Vaccination: The End of the Beginning. Annu. Rev. Pathol. Mech. Dis. 2024, 19, 69–97. [Google Scholar] [CrossRef]
- Kelleni, M.T. Evolution of SARS CoV-2 Omicron subvariants BF.7 and XBB.1.5: Time to follow Africa and abort all COVID restrictions. J. Infect. 2023, 86, 405–405. [Google Scholar] [CrossRef]
- Kelleni, M.T. COVID-19 mortality paradox (United States vs Africa): Mass vaccination vs early treatment. World J. Exp. Med. 2024, 14, 88674. [Google Scholar] [CrossRef]
- Yan, V.K.C.; Zhang, Y.; Yang, D.; Li, X.; Mak, L.Y.; Lai, F.T.T.; Chui, C.S.L.; Wan, E.Y.F.; Wong, C.; Chan, S.C.W.; et al. Post-acute sequelae of hospitalised COVID-19 re-infection compared with hospitalised first-time infection: a population-based retrospective cohort study in Hong Kong. BMJ Public Heal. 2025, 3, e000833. [Google Scholar] [CrossRef]
- DeGrasse, D.C.; Black, S.D. The Rise of SARS-CoV-2 (COVID-19) Omicron Subvariant Pathogenicity. Cureus 2023, 15, e40148. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Sa, S.; Hong, M.; Kim, J.; Shim, S.R.; Han, H.W. Adverse Events and Safety Profile of the COVID-19 Vaccines in Adolescents: Safety Monitoring for Adverse Events Using Real-World Data. Vaccines 2022, 10, 744. [Google Scholar] [CrossRef] [PubMed]
- Rella, S.A.; Kulikova, Y.A.; Dermitzakis, E.T.; Kondrashov, F.A. Rates of SARS-CoV-2 transmission and vaccination impact the fate of vaccine-resistant strains. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chau, N.V.V.; Ngoc, N.M.; Nguyet, L.A.; Quang, V.M.; Ny, N.T.H.; Khoa, D.B.; Phong, N.T.; Toan, L.M.; Hong, N.T.T.; Tuyen, N.T.K.; et al. An observational study of breakthrough SARS-CoV-2 Delta variant infections among vaccinated healthcare workers in Vietnam. eClinicalMedicine 2021, 41, 101143. [Google Scholar] [CrossRef]
- Yamamoto, K. Adverse effects of COVID-19 vaccines and measures to prevent them. Virol. J. 2022, 19, 1–3. [Google Scholar] [CrossRef]
- Lamprinou, M.; Sachinidis, A.; Stamoula, E.; Vavilis, T.; Papazisis, G. COVID-19 vaccines adverse events: potential molecular mechanisms. Immunol. Res. 2023, 71, 356–372. [Google Scholar] [CrossRef]
- Lyons-Weiler, J. Pathogenic priming likely contributes to serious and critical illness and mortality in COVID-19 via autoimmunity. J. Transl. Autoimmun. 2020, 3. [Google Scholar] [CrossRef]
- Kim, S.-J.; Rhee, T.G.; Shim, S.R. Autoimmune and auto-inflammatory adverse events after COVID-19 vaccination in the United States. Clin. Immunol. 2023, 259, 109882. [Google Scholar] [CrossRef]
- Xu, L.; Ma, Z.; Li, Y.; Pang, Z.; Xiao, S. Antibody dependent enhancement: Unavoidable problems in vaccine development. Adv Immunol. 2021;151:99-133. [CrossRef]
- Ajmeriya, S.; Kumar, A.; Karmakar, S.; Rana, S.; Singh, H. Neutralizing Antibodies and Antibody-Dependent Enhancement in COVID-19: A Perspective. J. Indian Inst. Sci. 2022, 102, 671–687. [Google Scholar] [CrossRef]
- Plūme, J.; Galvanovskis, A.; Šmite, S.; Romanchikova, N.; Zayakin, P.; Linē, A. Early and strong antibody responses to SARS-CoV-2 predict disease severity in COVID-19 patients. J. Transl. Med. 2022, 20, 1–12. [Google Scholar] [CrossRef]
- Afrin, L.B.; Weinstock, L.B.; Molderings, G.J. Covid-19 hyperinflammation and post-Covid-19 illness may be rooted in mast cell activation syndrome. Int. J. Infect. Dis. 2020, 100, 327–332. [Google Scholar] [CrossRef]
- Sumantri, S.; Rengganis, I. Immunological dysfunction and mast cell activation syndrome in long COVID. Asia Pac. Allergy 2023, 13, 50–53. [Google Scholar] [CrossRef]
- Gorman, R.d.S.; Syed, I.U. Connecting the Dots in Emerging Mast Cell Research: Do Factors Affecting Mast Cell Activation Provide a Missing Link between Adverse COVID-19 Outcomes and the Social Determinants of Health? Med Sci. 2022, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Bossche, GV. The inescapable immune escape pandemic. Pierucci Publishing, Aspen, CO. 2023. https://www.boswellbooks.com/book/9781956257809.
- Mead, M.N.; Seneff, S.; Rose, J.; Wolfinger, R.; Hulscher, N.; McCullough, P.A. COVID-19 Modified mRNA “Vaccines”: Lessons Learned from Clinical Trials, Mass Vaccination, and the Bio-Pharmaceutical Complex, Part 2. Int. J. Vaccine Theory, Pr. Res. 2024, 3, 1275–1344. [Google Scholar] [CrossRef]
- Kanduc, D.; Shoenfeld, Y. Molecular mimicry between SARS-CoV-2 spike glycoprotein and mammalian proteomes: implications for the vaccine. Immunol. Res. 2020, 68, 310–313. [Google Scholar] [CrossRef]
- Vojdani, A.; Vojdani, E.; Kharrazian, D. Reaction of Human Monoclonal Antibodies to SARS-CoV-2 Proteins With Tissue Antigens: Implications for Autoimmune Diseases. Front. Immunol. 2021, 11, 617089. [Google Scholar] [CrossRef]
- Güven, O.; Karakurt, G.; Naser, A.; Selçuk, H.; Keleş, D.V.; Gedik, E.; Avsever, M.; Köse, F.F. The Impact of COVID-19 Infection on the Development of Stroke, Pulmonary Embolism, and Myocardial Infarction: A Retrospective Study. Cureus 2025, 17. [Google Scholar] [CrossRef] [PubMed]
- Prasad, V.; Makary, M.A. An Evidence-Based Approach to Covid-19 Vaccination. New Engl. J. Med. 2025, 392, 2484–2486. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food & Drug Administration; Health & Human Services. Guidance for Industry. E 10 Choice of Control Group and Related Issues in Clinical Trials. May 2001. URL: https://www.fda.gov/media/71349/download.
- Taccetta, C. The “Noise” of mRNA. The Daily Clout. 9 July 2025. URL: https://dailyclout.io/the-noise-ofmrna/.
- Le, J. Vivian EM. Overview of Pharmacokinetics. Merck Manual, Professional Version. Reviewed/Revised Nov 2024. URL: https://www.merckmanuals.com/professional/clinical-pharmacology/pharmacokinetics/overview-of-pharmacokinetics.
Figure 1.
Timeline Comparisons of COVID-19 Cases & Mortality: Nordic & Asian Countries. Timeline comparisons for COVID-19 cases (orange) and deaths (red) for four Nordic countries and four Southeast Asian countries. The syringe icon shows approximate start of mRNA vaccinations. The blue vertical line approximates date of 75% vaccine coverage for each country’s population, based on data from Our World In Data (https://ourworldindata.org/). Charts adapted from Johns Hopkins Coronavirus Resource Center.
Figure 1.
Timeline Comparisons of COVID-19 Cases & Mortality: Nordic & Asian Countries. Timeline comparisons for COVID-19 cases (orange) and deaths (red) for four Nordic countries and four Southeast Asian countries. The syringe icon shows approximate start of mRNA vaccinations. The blue vertical line approximates date of 75% vaccine coverage for each country’s population, based on data from Our World In Data (https://ourworldindata.org/). Charts adapted from Johns Hopkins Coronavirus Resource Center.
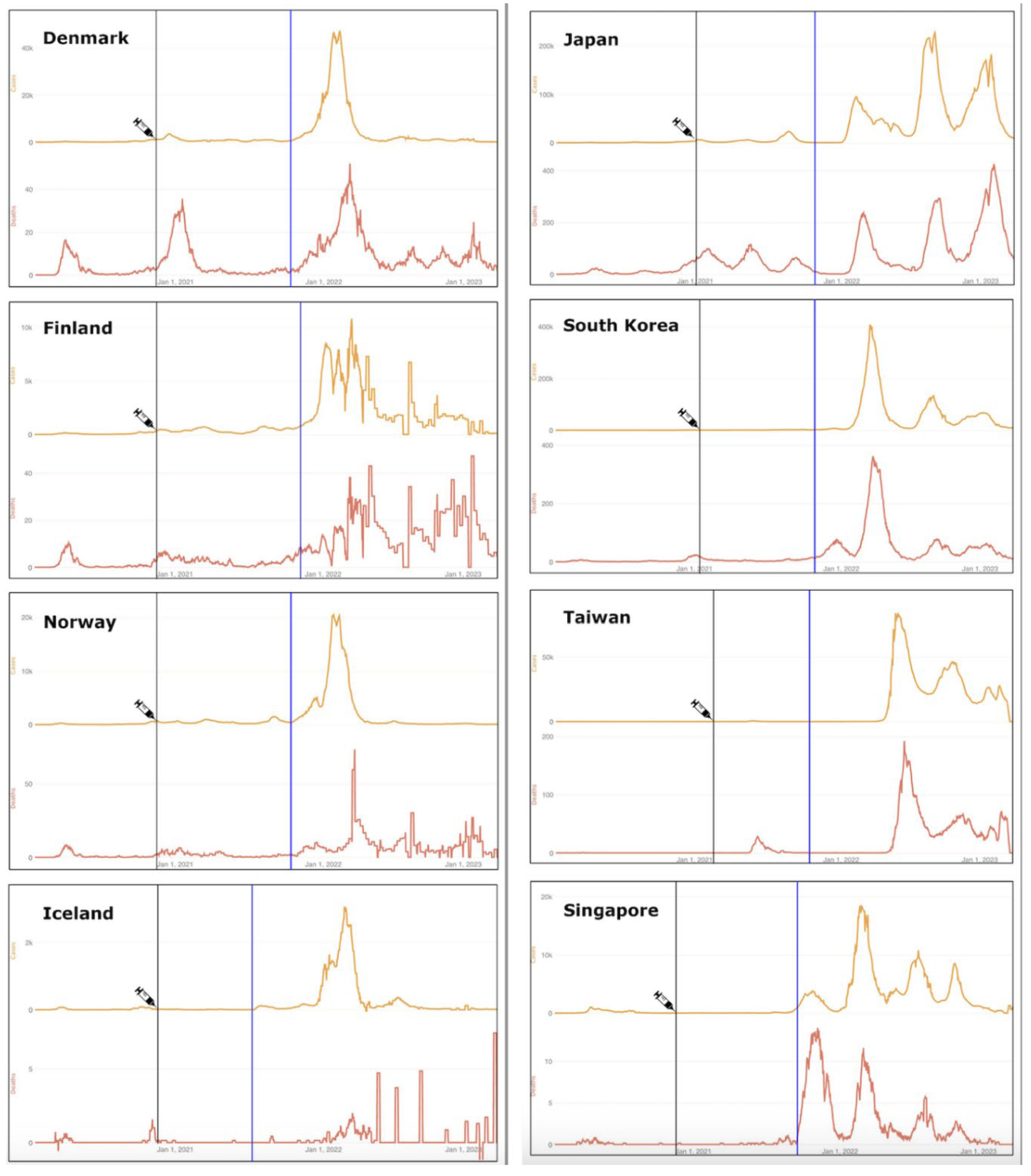
Figure 2.
Central figure, 5 features of Hybrid Harms Hypothesis. Created with BioRender.com.
Figure 3.
Timeline comparisons for Australia, So. Korea, and Singapore, Jan. 2020 to Sept. 2022. For each country, the top chart shows the COVID cases (black line) and COVID-associated deaths (blue); bottom chart shows the percent excess mortality (PEM, monthly) from Jan. 2022 to Sept. 2022. The dotted line is the 10% PEM/ percent COVID-excess mortality line. The pink area, “Living with COVID”, coincides with the height of mass vaccination. Graphs from Cao et al. [348] Front Public Health. 2023;11:1085451. Permission to use this figure has been granted in accordance with the open access Creative Common CC BY 4.0 license.
Figure 3.
Timeline comparisons for Australia, So. Korea, and Singapore, Jan. 2020 to Sept. 2022. For each country, the top chart shows the COVID cases (black line) and COVID-associated deaths (blue); bottom chart shows the percent excess mortality (PEM, monthly) from Jan. 2022 to Sept. 2022. The dotted line is the 10% PEM/ percent COVID-excess mortality line. The pink area, “Living with COVID”, coincides with the height of mass vaccination. Graphs from Cao et al. [348] Front Public Health. 2023;11:1085451. Permission to use this figure has been granted in accordance with the open access Creative Common CC BY 4.0 license.
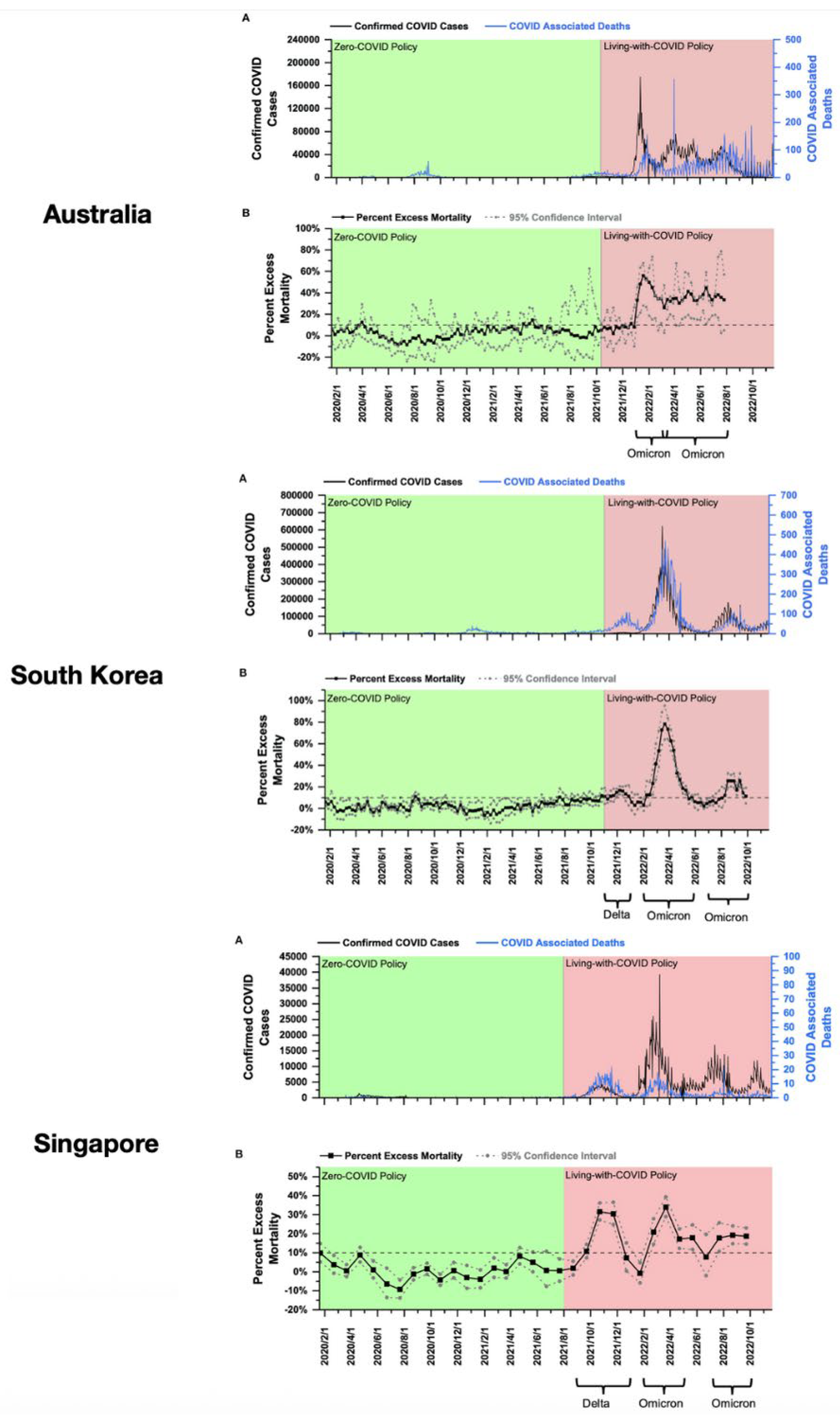

Table 2.
Overlapping Sequelae of Post-Acute Sequelae of COVID-19 (PASC) and Post-COVID-19 Vaccine Syndrome (PCVS).
Table 2.
Overlapping Sequelae of Post-Acute Sequelae of COVID-19 (PASC) and Post-COVID-19 Vaccine Syndrome (PCVS).
| Organ system | PASC/PCVS Combined | Organ system | PASC/PCVS Combined |
|---|---|---|---|
| Cardiovascular |
|
Gastrointestinal |
|
| Coagulation/ Hematological |
|
Gynecological |
|
| Dermatological |
|
Mental Health |
|
| Endocrine |
|
Musculoskeletal |
|
| Immunological/ Autoimmune |
|
Neurological |
|
| Otolaryngological |
|
Pulmonary |
|
| Renal |
|
||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



