Submitted:
11 August 2025
Posted:
13 August 2025
You are already at the latest version
Abstract
In the domain of cancer management, the intricate interplay between NK cells and cancer itself unveils compelling insights. The inhibition of NK cells can initiate cancer development, while the disease, in turn, can heighten the suppression of NK cell function. This dynamic underscores the critical need to delve into allogeneic NK cell therapeutics as a promising avenue for cancer treatment. NK cells emerge as pivotal components in cancer immunotherapy, showcasing a distinctive capacity to target and distinguish diverse cancer stem cells (CSCs). Through mechanisms such as lysis, along with the secretion of IFN-γ and TNF-α, NK cells play a crucial role in hindering tumor advancement and metastasis. As we navigate the complexities of tumorigenesis, a significant discourse arises around the decline in both the cell numbers and efficacy of NK cells during cancer progression or in maintaining a healthy lifestyle. Recognizing the indispensable attributes of NK cells, particularly their function and proliferation, highlights the importance of integrating NK cell-based therapies into the arsenal of tumor immunotherapy strategies. In the quest for effective tumor elimination, leveraging the diverse capabilities of NK cells in conjunction with innovative immunotherapeutic approaches signifies a transformative strategy in the fight against cancer.
Keywords:
NK cells
; preneoplasia
; cancer
; obesity
; gene knock-out
; osteoclasts
1. Introduction: Natural Killer Cells
Natural killer (NK) cells, essential components of innate immunity, possess unique abilities in targeting tumor cells and virally infected cells without prior sensitization [1,2]. NK cells play a crucial role in targeting tumor cells and virally-infected cells through the release of perforin and granzyme B [1,3,4]. These actions, regulated by cystatins and cathepsins, can lead to necrotic or apoptotic cell death in susceptible targets [1,3,4]. Originating from CD34+ hematopoietic stem cells in the bone marrow, NK cells are distributed throughout various body sites such as the spleen, liver, placenta, and peripheral blood [5]. These cells play a pivotal role in cancer inhibition through diverse effector functions, including direct cytotoxicity against tumors, antibody-dependent cellular cytotoxicity (ADCC), and the modulation of immune effectors by releasing inflammatory cytokines and chemokines [6,7,8]. NK cells target CSCs with altered surface marker expressions [9,10]. We have previously shown a strong association between tumor differentiation stages and their vulnerability to primary NK cell-mediated cytotoxicity [11].
NK cells offer a promising alternative to T cells in the fight against cancer. Unlike T cells, NK cells are part of the innate immune response, allowing them to target major histocompatibility complex (MHC) class I-deficient or mutated cells aggressively [12]. While tumors can develop resistance to NK cell attacks, enhancing NK cells through activation, expansion, and genetic modifications can significantly boost their anti-tumor capabilities and help them overcome drug resistance [13,14].The activation of NK cells is governed by interactions between their receptors and ligands on target cells, independent of antigen processing or presentation [15]. These versatile cells can be sourced from blood, cord blood, hematopoietic stem cells, or induced pluripotent stem cells. They can also be expanded and cryopreserved for future use, making them readily available off-the-shelf [15,16,17]. In the context of Hematopoietic stem-cell transplantation (HSCT), NK cells can be infused alone or with other cells after a pre-enrichment process. Allogeneic NK cells with inhibitory receptors like KIR can target cancer cells lacking suitable MHC ligands, leading to their elimination [18,19,20,21,22]. Clinical trials have shown promising results, with reports of complete remission and improved survival rates in cancer patients, especially in cases of acute myeloid leukemia (AML) [23,24]. The success of NK cell-based therapies highlights their potential to complement and address limitations of existing treatments, like chimeric antigen receptor T-cell therapies [25,26,27]. The lack of graft versus host disease (GVHD) in NK cells allows safe use of allogeneic NK cells in adoptive cell-based therapies [28,29]. These advancements signify a significant step forward in the fight against cancer, offering new hope for patients and clinicians alike.
In the realm of immune surveillance, the significance of NK cells cannot be overstated. When the functions of NK cells are hindered, malignant cells can elude detection, paving the way for tumor development. Studies have underscored the compromised functionalities of NK cells in both pre-neoplastic and neoplastic phases. Assessing NK cell function is crucial in the regular clinical evaluation of cancer patients, individuals at high risk, and even those in good health, as it stands as a pivotal gauge of immune vitality. This paper delves into a thorough exploration of NK cell maturation stages, particularly within the spheres of health, preneoplasia, and cancer microenvironments. It sheds light on the impacts of an unhealthy lifestyle or genetic anomalies on NK cell function. Furthermore, it delves into strategies for enhancing NK cells through the utilization of feeder cells, cytokines, and signaling activation, aiming to pave the way for the advancement of cancer immunotherapies.
2. Modulation of NK Cell Numbers and Function at the Pre-Neoplastic Stage or Due to Genetic and Environmental Factors
Genetic mutation or specific gene knock-outs could induce pre-neoplasia or cancer [30]. Studies have shown that administering a high-fat calorie diet (HFCD) to mice with a pancreatic KRAS mutation significantly impairs NK cell function during the pre-neoplastic stage of pancreatic cancer [31,32]. This impairment was observed across various tissues, with decreased NK cytotoxicity in the peripheral blood, spleen, pancreas, and peri-pancreatic tissue in mice with KRAS mutation on HFCD compared to those on a lean control diet (CD) [32]. Furthermore, mice with KRAS mutation on HFCD exhibited the most substantial reduction in NK cell expansion and function, followed by those with KRAS mutation on CD and WT mice on HFCD when compared to WT mice on CD [32]. Notably, NK cells cultured with feeder cells from mice with KRAS mutation and WT mice on HFCD showed decreased expansion, cytotoxicity, and IFN-γ secretion [32]. Conversely, NK cells from WT mice on CD, when cultured with feeder cells from KRAS mutant mice on HFCD, displayed lower cytotoxicity alongside increased cytokine secretion [32]. These results indicate that while NK cytotoxicity is consistently suppressed, the extent of this suppression determines the profiles of IFN-γ secretion. The selective loss of NK cell function is influenced by the level of suppression within the microenvironment. The decline in expansion and cytotoxicity may precede the loss of IFN-γ secretion, and in cases of significant NK cell inactivation, all key functions of NK cells can be compromised.
It was that in addition to NK cells, feeder cells known to activate and proliferate NK cells from mice with KRAS mutation fed with HFCD exhibit significantly lower levels of MHC class I inhibitory ligands and RAE1-delta activating ligands [32]. This decrease implies a reduction in both inhibitory and activating ligands crucial for NK cell signaling. Interestingly, the decline in ligand expression was notably more pronounced in feeder cells from KRAS-mutated mice on HFCD compared to those on a standard diet, with the highest levels observed in WT mice on a regular diet [32]. The diminished presence of MHC class I and RAE1-delta on feeder cells aligns with the development of pre-neoplastic lesions (PanINs) in KRAS-mutated mice, suggesting a potential link between reduced surface receptors on feeder cells, compromised NK cell activity, and PanIN formation [31,32]. These findings propose that the impairment in NK cell function, influenced by genetic and environmental factors during the pre-malignant phase of pancreatic cancer, may contribute significantly to the initiation and progression of pancreatic malignancies [31,32] (Figure 1). Pancreatic intraepithelial neoplasias (PanINs) exhibit lower CD44 expression levels, rendering them resistant to NK cell-mediated cytotoxicity, showcasing a more differentiated phenotype. In contrast, KC tumors display elevated CD44 levels, making them vulnerable to NK cell-mediated cytotoxicity, reflecting a less differentiated phenotype [32]. The distinct profiles suggest a potential decrease in NK cell inactivation during the pre-neoplastic stage compared to established cancer in mice [32]. Notably, pancreatic cancer patients and humanized mice with pancreatic and oral tumors show significant impairment in NK cell functions. However, during the pre-neoplastic phase, the suppression of NK cells may be less severe, gradually intensifying with cancer progression [33] (Figure 1). This phenomenon aligns with observations of diminished NK cell-mediated cytotoxicity and reduced IFN-γ secretion, believed to drive the advancement of oral and pancreatic tumors in humanized mice [33].
A synergistic increase in IL-6 secretion, coupled with decreased IFN-γ secretion when peri-pancreatic adipose-derived cells interact with NK cells, may facilitate the proliferation of pancreatic tumors [32]. IL-6 plays a crucial role in driving PDAC proliferation and suppressing NK cell function. Previous studies have shown that the addition of IL-6 to tumor/NK cell cultures hinders NK-mediated IFN-γ secretion [34,35,36,37,38,39]. This phenomenon suggests that adipose tissue surrounding tumors could potentially transform tumor-suppressing NK cells into tumor-promoting entities [4,40,41,42,43]. By blocking IL-6, not only could tumor growth be inhibited, but NK cells could also be rescued from the suppression induced by peri-pancreatic adipose tissue or tumor-infiltrating cells. This approach offers a promising therapeutic strategy to combat pancreatic tumors effectively.
The interplay between mutation, obesity, unhealthy lifestyle, and the compromised NK function in the development of pre-neoplastic lesions in pancreatic cancer underscores the need for innovative immunotherapeutic approaches [31,32] (Figure 1). Understanding how lifestyle factors like diet impact NK function further emphasizes the importance of public education and lifestyle modifications to counter cancer risks. Given the limited success in enhancing NK cell function in solid tumors, novel strategies are crucial to combat tumor growth and restore NK activity. The upcoming sections will delve into studies exploring the enhancement of NK functions alongside complementary therapeutic interventions.
3. Suppressed NK Cell Function in the Cancer-Bearing Mice Model
In the studies involving tumor-bearing humanized mice, it was observed that NK cells exhibited impaired function akin to that of cancer patients [31,33,44]. The similarities in NK cell numbers and function between cancer patients and hu-BLT mice indicate the potential of humanized mice as a suitable model for investigating NK-tumor interactions [44,45]. Studies revealed that NK cells from oral, pancreatic, melanoma, and uterine tumor-bearing humanized mice experienced a significant loss of function and reduced expansion compared to non-tumor-bearing mice [33,46,47]. However, the intravenous administration of highly potent feeder cell-activated NK cells effectively prevented tumor formation in humanized mice. Tumors resected from NK-injected humanized mice exhibited a differentiated phenotype, showing slow growth and limited expansion. In contrast, tumors from non-NK-injected tumor-bearing mice displayed faster growth, higher expansion rates, and increased susceptibility to NK cell-mediated cytotoxicity, indicating a less differentiated phenotype. Notably, tumors from mice injected with NK cells, with or without probiotic bacteria feeding, recruited 18-22 times more human CD45+ immune cells compared to tumors from mice without these interventions. This increase was observed in both the presence and absence of probiotic feeding, as detailed in sources [44]. Furthermore, the restoration of NK cytotoxicity and enhanced IFN-γ secretion in various immune cell populations within tumor-bearing mice, including PBMCs, splenocytes, bone marrow-derived immune cells, enriched NK cells, and purified T cells, was achieved by injecting NK cells with the feeding of probiotic bacteria in humanized mice [44,46,47].
4. Suppressed NK Cell Function in Human Cancer Patients
Several studies have demonstrated that peripheral blood-derived NK cells and/or tumor-infiltrating NK cells of cancer patients exhibit reduced numbers and anti-cancer activity, as well as lowered cytokine secretion levels [33,48,49,50,51,52,53,54,55]. Although the suppression of NK cell function in cancer patients is a recognized occurrence, the exact relationship between this suppression and the development of cancer remains unclear. It raises the question of whether the inhibition of NK cell function is a result of cancer induction and progression or if it occurs before cancer establishment, potentially serving as a mechanism for tumor formation (Figure 1 and Figure 2).
It is crucial to understand that in cancer patients, both cytotoxic function and IFN-γ secretion by NK cells are compromised, impacting the ability to combat aggressive tumors effectively [56]. The loss of NK cell function is attributed to various factors, including the downregulation of key receptors such as CD16 and NKG2D, as well as decreased survival, expansion, and cytokine secretion, particularly IFN-γ [33,57,58,59]. This decline in NK cell activity not only hinders the elimination of undifferentiated tumors but also contributes to the proliferation of these aggressive cancer cells [60]. Tumor microenvironment-induced suppressed function or cell death in primary NK cells was demonstrated, and reduced NK cells’ function was also seen when NK cells were co-cultured with tumors in vitro [61]. Tumor microenvironment secreted immunosuppressive molecules or factors, such as IDO, PGE2, TGF-β, prostaglandins, and IL-10, can downmodulate NK cell activating receptors and can inhibit NK cell activity [62,63]. Interestingly, research shows that even when cancer patients and healthy individuals secrete similar levels of IFN-γ, the efficacy of IFN-γ from cancer patients’ NK cells in tumor differentiation is significantly reduced compared to that from healthy individuals [33]. These findings underscore the importance of restoring IFN-γ secretion by NK cells in cancer patients and enhancing the functional capacity of this cytokine to effectively target and control tumors. In addition to NK cells, reduced function and expression levels of NK cell receptor ligands were seen on NK-activating cells in cancer patients [46] (Figure 1). Addressing these aspects holds significant translational implications for improving the outcomes of cancer treatment strategies.
Moreover, the suppressed function of peripheral blood-derived NK cells and low tumor-infiltrating NK cells has been associated with poor prognosis in cancer patients [33,50,51,56,64,65,66]. To overcome the problem, several in vitro techniques have been established to expand the number and function of NK cells to produce a higher therapeutic cell dose of NK cells for cancer therapeutics [67,68,69,70,71].
5. Natural Killer Cells’ Maturation Stages
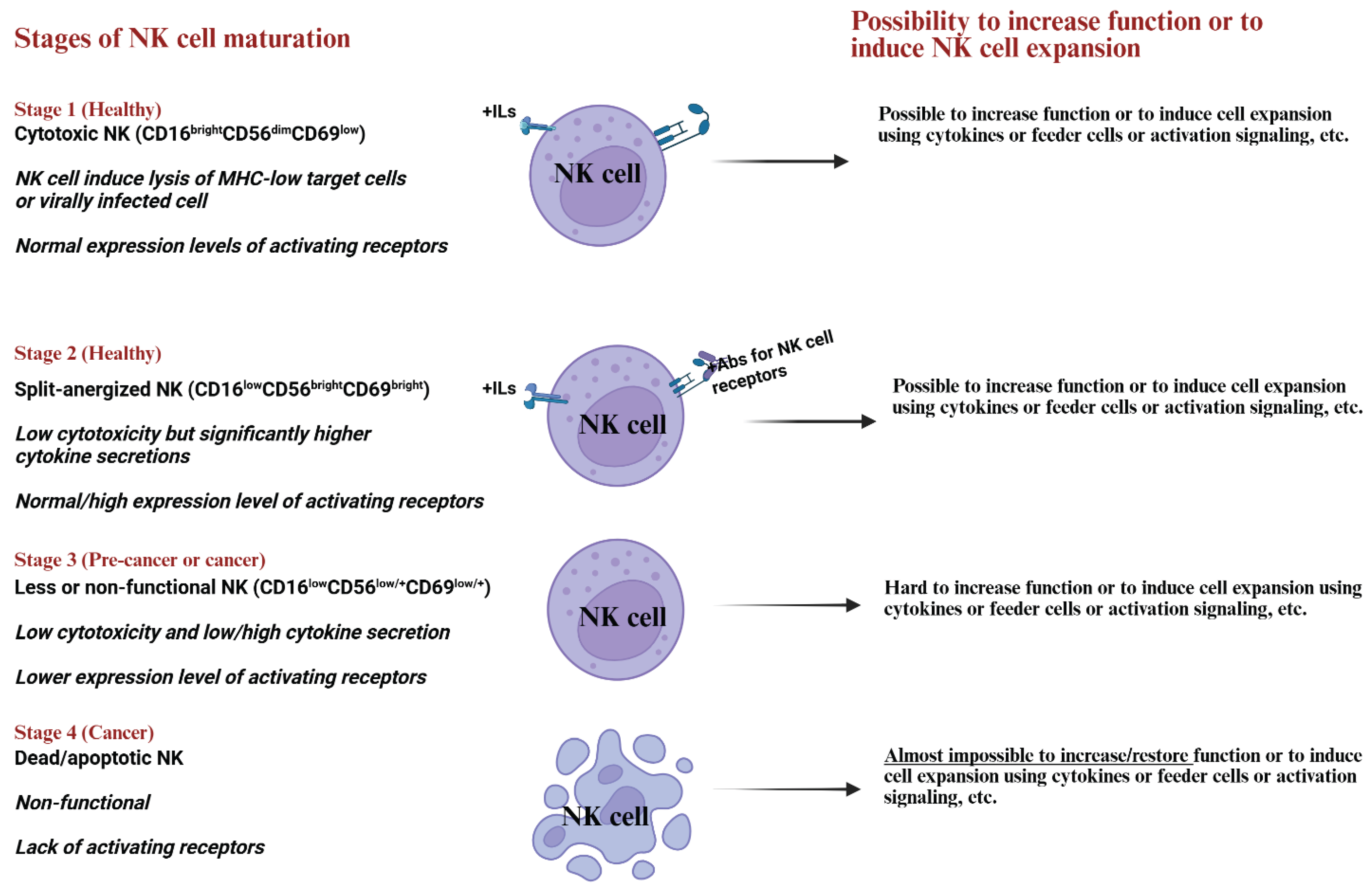
The activation of NK cells is a complex process involving various stages of maturation in humans, identified through the analysis of CD16, CD56, and CD69 surface receptors [40,72]. Human NK cells are identified by the presence of CD56 and CD16 on their surface, distinguishing them from CD3-negative cells [73]. These cells can be categorized into two subsets based on their CD56 and CD16 expression levels: the predominant subset, comprising about 90% of human NK cells, exhibits low CD56 and high CD16 levels (CD56dimCD16bright), while the minor subset, constituting approximately 10% of human NK cells, displays high CD56 and low CD16 expression (CD56brightCD16dim/-) [73,74]. Studies have shown that the CD56dimCD16bright subset demonstrates heightened cytotoxicity, whereas the CD56brightCD16dim/- subset is proficient in cytokine secretion, including IFN-γ, TNF-α, TNF-β, GM-CSF, IL-10, and IL-13 upon exposure to pro-inflammatory cytokines [73,74,75]. Notably, the production of TNF-α and IFN-γ by NK cells plays a critical role in tumor differentiation, leading to reduced growth and metastasis rates in differentiated tumors compared to cancer stem cells [11,46,76].
To comprehend the intricate workings of NK cells in various contexts, such as the tumor microenvironment or viral and bacterial infections, understanding their regulation and mechanisms is crucial. To fully understand, we have identified four stages of NK cell maturation in humans based on the expression of CD16, CD56, and CD69 surface receptors (Figure 2). Stage 1, CD16brightCD56dimCD69low NK cells, making up approximately 90% of peripheral blood NK cells, are crucial in targeting and eliminating cancer stem-like cells (CSCs). Stage 2, CD16lowCD56brightCD69bright NK cells, which are less cytotoxic or anergized, play a role in regulating other cell functions and promoting tumor cell differentiation via secreted factors. Stage 3, as NK cells progress to the third stage of maturation, they may lose their cytotoxic abilities or the capacity to produce IFN-γ. Stage 4, ultimately, NK cells may undergo apoptosis (Figure 2).
NK cells at stage 3 were named as “split-anergized NK cells” as they downregulate their cytotoxic abilities while still secreting significantly higher levels of cytokines, especially IFN-γ and TNF-α [64,77,78]. This state of NK cell maturation plays a pivotal role in the differentiation of undifferentiated or stem-like tumors. Interestingly, interaction of NK cells with stem-like tumor triggers split-anergy in NK cells, but differentiated tumor cells do not trigger this split anergy in NK cells, suggesting they are not prime targets for primary NK cell activity [79,80]. Split anergy can be induced in NK cells through treatment with IL-2 and anti-CD16 monoclonal antibodies, mimicking the activation process during interactions with tumor cells [72,77,78]. Unlike T cell anergy, where all functions are suppressed, split anergy in NK cells specifically reduces cytotoxicity while increasing cytokine and chemokine secretion [64,81,82]. This selective loss in cytotoxicity is instrumental in the differentiation of tumors, potentially leading to the deactivation of NK cells upon encountering well-differentiated tumor cells [64,79,83]. Understanding these intricate mechanisms sheds light on the nuanced functions of NK cells in combating tumors and infections, offering valuable insights for future research and therapeutic interventions.
In cancer patients, there is a notable increase in the numbers and frequencies of stage 3 and 4 NK cells, leading to compromised NK cell functions (Figure 2). Reports indicate reduced NK cell cytotoxicity in the tumor microenvironment and peripheral blood of cancer patients, along with the downregulation of CD16 receptors on NK cell surfaces. The diminished functionality of NK cells correlates with higher cancer susceptibility, while enhanced NK cell activity and increased infiltration into tumor cells are linked to a more favorable prognosis.
6. Challenges and Advancements to Develop NK Cell-Based Cancer Immunotherapies
Higher efficacy of cancer immunotherapies has been shown against several cancers in recent clinical trials [28,84,85,86,87]. To address the lower frequencies and function of NK cells in cancer patients, we need to develop a method to produce abundant and highly functional NK cells for adoptive NK cell transfer therapy targeting cancers. Since the discovery of NK, scientists have attempted to develop strategies to safely use NK cells to effectively treat cancer patients. It was found that the NK cell-based therapies are extremely safe; however, the efficacy level has always been challenged due to several issues. Unlike T cells, which can comprise 60 to 40% of lymphocytes in the human peripheral blood, the percentage of NK cells is only 5-15% of total human peripheral blood lymphocytes [88]. Also, as mentioned in Section 2, Section 3, Section 4 and Section 5 of this paper, the functions of NK cells are impacted by several factors, and also during preneoplasia and cancer [31,89]. Moreover, NK cells are quick to become inactivated in the tumor microenvironment (TME), even when they are activated by cytokines or other methods. There is a lack of persistence of NK cells upon adoptive transfer to cancer patients. These qualities of NK cells have made these cells less desirable for cell therapy.
Extensive research has been conducted to determine the impact of tumor gene knockouts on NK cell function [90]. Studies have revealed significant enhancements in NK cell expansion and activation when specific genes are targeted for knockdown [90]. For instance, the knockdown of NFkB in HEp2 tumors, CD44 in breast and melanoma tumors, and COX2 in healthy myeloid cells and mouse embryonic fibroblasts resulted in notable improvements in NK cell functionality [41,43,90,91]. Interestingly, heightened NK cell responsiveness was observed in mice with gene knockouts related to inflammation and NK cell signaling, suggesting a complex interplay of genes and pathways influencing NK cell activation [92]. Furthermore, when CD44, a marker of stemness often upregulated in tumors, was suppressed in breast and melanoma tumors, a decrease in differentiation antigens and an increase in NK cell activation were noted [43]. These findings underscore the crucial role of NK cells in targeting undifferentiated cells and highlight the significance of cellular differentiation in regulating NK cell function and expansion. These studies shed light on the intricate relationship between gene modulation, cellular differentiation, and NK cell activity, emphasizing the pivotal role of NK cells in combating cells that lack optimal differentiation capabilities. Additionally, several methodologies have been found to overcome the lack of adequate expansion of NK cells ex vivo by the use of feeder cells with/without cytokines or other activation signals [67,69,70,93,94,95,96,97,98,99]. Unfortunately, few of such methodologies give rise to larger numbers of NK cells with poor quality, as a methodology to not only expand well but also retain NK cell functional capabilities [88]. Even though we are getting closer to having effective cells, we are still not entirely there. Although many studies claim that they have a significant expansion of NK cells with the use of cytokines, the majority of them have been less desirable due to the variability of the donor-derived NK cells’ expansion and functional capabilities. Thus, the search for super donors makes NK cell-based therapy very limited. Feeder layer-dependent expansion, such as K562 or OSCSCs, has been less than desirable for expansion [99,100]. Also, genetic manipulation expressing 4IBBL and IL-15 or IL-21 has gained popularity for the expansion of the NK cells [69,99,101,102,103,104,105,106,107,108]. It is not clear whether such expansion will increase the functional activation of NK cells maximally and for how long. Knockdown of genes in NK cells was shown to increase the functional activation of NK cells [109]. This characteristic is very common for the function of NK cells since either the knockdown is in the target cells or the NK cells will result in the activation of the NK cells [43,109].
Studies have demonstrated approach involves the efficient ex vivo expansion of human NK cells using osteoclasts as feeder cells. It was found that the myeloid subset plays a crucial role in activating NK cells, leading to enhanced cytotoxicity and the secretion of cytokines and chemokines compared to monocytes and dendritic cells [110]. These exciting advancements in NK cell expansion techniques have revolutionized therapeutic possibilities [69,111]. It was found that the efficacy of NK cell-based therapies could be further enhanced by combining with checkpoint inhibitors such as anti-programmed cell death-1 (anti-PD1) monoclonal antibodies as cancer therapeutics [112,113]. Efficacy of NK cell-based immunotherapies was demonstrated in vivo using clinically relevant humanized mouse models, confirming the potential to successfully translate this approach into patients [46,76,89,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132].
The expansion of purified NK cells from cancer patients presents unique challenges when compared to those from healthy individuals (Figure 1 and Figure 2). Studies indicate that the rapid proliferation of a small fraction of T cells in cancer patients hinders NK cell expansion, affecting the cytotoxic function of the expanded cells [33]. This interaction highlights the crucial interplay between NK cells and T cells, particularly Tregs and MDSCs, which are known to impede NK function [133]. In a comparative study where both healthy donors’ and cancer patients’ NK cells were expanded using the same methodology, it was observed that the expanded NK cells from cancer patients had lower cytotoxic function and IFN-γ secretion levels than those from healthy donors [118,127]. Additionally, the patient-derived expanded NK cells showed reduced expression of activating receptors such as CD16, CD56, Nkp30, Nkp44, Nkp46, NKG2D, and CD54 [33]. These findings suggest the potential use of allogeneic healthy donor-derived NK cells in cancer cell-based adoptive therapeutics clinical studies, as NK cells do not induce GVHD [28,29].
7. Conclusions
In the realm of cancer research, recent advancements have shed light on the critical role of NK cells in combating cancer. Studies have revealed a notable decline in NK cell function among preneoplastic individuals and those with cancer, emphasizing the importance of restoring NK cell activity for effective cancer treatment. Unlike T cells that target tumors with higher MHC class I expression, NK cells excel at targeting CSCs with minimal MHC class I levels. They exert control over tumor cells through direct cytolytic actions, antibody-mediated ADCC, and inducing tumor cell differentiation via IFN-γ, thereby enhancing the impact of chemotherapy and radiotherapy. The immune landscape in cancer patients underscores the significance of choosing between allogeneic and autologous NK cell immunotherapy, especially given the prevalent NK cell dysfunction in most cases. Strategies must be devised to bolster NK cell expansion and function to counter the proliferation of tumor-associated macrophages, Tregs, and MDSCs, commonly found in aggressive tumors. The integration of allogeneic NK cell-based therapies shows promise in advancing the goal of eradicating tumors effectively.
Conflicts of Interest
The authors declare that the work reviewed in the article was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Moretta, A.; Marcenaro, E.; Parolini, S.; Ferlazzo, G.; Moretta, L. NK cells at the interface between innate and adaptive immunity. Cell Death Differ. 2007, 15, 226–233. [Google Scholar] [CrossRef]
- Vivier, E.; Raulet, D.H.; Moretta, A.; Caligiuri, M.A.; Zitvogel, L.; Lanier, L.L.; Yokoyama, W.M.; Ugolini, S. Innate or Adaptive Immunity? The Example of Natural Killer Cells. Science 2011, 331, 44–49. [Google Scholar] [CrossRef]
- Lanier, L.L. NK cell recognition. Annu. Rev. Immunol. 2005, 23, 225–274. [Google Scholar] [CrossRef]
- Magister, Š.; Tseng, H.-C.; Bui, V.T.; Kos, J.; Jewett, A. Regulation of split anergy in natural killer cells by inhibition of cathepsins C and H and cystatin F. Oncotarget 2015, 6, 22310–22327. [Google Scholar] [CrossRef]
- Colucci, F.; Caligiuri, M.A.; Di Santo, J.P. What does it take to make a natural killer? Nat. Rev. Immunol. 2003, 3, 413–425. [Google Scholar] [CrossRef]
- Palmer, J.M.; Rajasekaran, K.; Thakar, M.S.; Malarkannan, S. Clinical Relevance of Natural Killer Cells Following Hematopoietic Stem Cell Transplantation. J. Cancer 2013, 4, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Fildes, J.; Yonan, N.; Leonard, C. Natural killer cells and lung transplantation, roles in rejection, infection, and tolerance. Transpl. Immunol. 2008, 19, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Farag, S.S.; Caligiuri, M.A. Human natural killer cell development and biology. Blood Rev. 2006, 20, 123–137. [Google Scholar] [CrossRef]
- López-Soto, A.; Gonzalez, S.; Smyth, M.J.; Galluzzi, L. Control of Metastasis by NK Cells. Cancer Cell 2017, 32, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Solana, R.; Casado, J.G.; Delgado, E.; DelaRosa, O.; Marín, J.; Durán, E.; Pawelec, G.; Tarazona, R. Lymphocyte activation in response to melanoma: Interaction of NK-associated receptors and their ligands. Cancer Immunol. Immunother. 2006, 56, 101–109. [Google Scholar] [CrossRef]
- Bui, V.T.; Tseng, H.-C.; Kozlowska, A.; Maung, P.O.; Kaur, K.; Topchyan, P.; Jewett, A. Augmented IFN-γ and TNF-α Induced by Probiotic Bacteria in NK Cells Mediate Differentiation of Stem-Like Tumors Leading to Inhibition of Tumor Growth and Reduction in Inflammatory Cytokine Release; Regulation by IL-10. Front. Immunol. 2015, 6, 576. [Google Scholar] [CrossRef]
- Daher, M.; Rezvani, K. Next generation natural killer cells for cancer immunotherapy: The promise of genetic engineering. Curr. Opin. Immunol. 2018, 51, 146–153. [Google Scholar] [CrossRef]
- Page, A.; Chuvin, N.; Valladeau-Guilemond, J.; Depil, S. Development of NK cell-based cancer immunotherapies through receptor engineering. Cell. Mol. Immunol. 2024, 21, 315–331. [Google Scholar] [CrossRef]
- Sordo-Bahamonde, C.; Vitale, M.; Lorenzo-Herrero, S.; López-Soto, A.; Gonzalez, S. Mechanisms of Resistance to NK Cell Immunotherapy. Cancers 2020, 12, 893. [Google Scholar] [CrossRef]
- Liu, S.; Galat, V.; Galat, Y.; Lee, Y.K.A.; Wainwright, D.; Wu, J. NK cell-based cancer immunotherapy: From basic biology to clinical development. J. Hematol. Oncol. 2021, 14, 1–17. [Google Scholar] [CrossRef]
- Fang, F.; Wang, W.; Chen, M.; Tian, Z.; Xiao, W. Technical advances in NK cell-based cellular immunotherapy. Cancer Biol. Med. 2019, 16, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Maia, A.; Tarannum, M.; Lérias, J.R.; Piccinelli, S.; Borrego, L.M.; Maeurer, M.; Romee, R.; Castillo-Martin, M. Building a Better Defense: Expanding and Improving Natural Killer Cells for Adoptive Cell Therapy. Cells 2024, 13, 451. [Google Scholar] [CrossRef] [PubMed]
- Berrien-Elliott, M.M.; Jacobs, M.T.; Fehniger, T.A. Allogeneic natural killer cell therapy. Blood 2023, 141, 856–868. [Google Scholar] [CrossRef]
- Gao, F.; Ye, Y.; Gao, Y.; Huang, H.; Zhao, Y. Influence of KIR and NK Cell Reconstitution in the Outcomes of Hematopoietic Stem Cell Transplantation. Front. Immunol. 2020, 11, 2022. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.S.; Rezvani, K. Can we make a better match or mismatch with KIR genotyping? Hematol. Am. Soc. Hematol. Educ. Program 2016, 2016, 106–118. [Google Scholar] [CrossRef]
- Mohseni, R.; Sharif, P.M.; Behfar, M.; Shojaei, S.; Shoae-Hassani, A.; Jafari, L.; Khosravi, A.; Nikfetrat, Z.; Hamidieh, A.A. Phase I study of safety and efficacy of allogeneic natural killer cell therapy in relapsed/refractory neuroblastomas post autologous hematopoietic stem cell transplantation. Sci. Rep. 2024, 14, 1–12. [Google Scholar] [CrossRef]
- Bednarski, J.J.; Zimmerman, C.; Berrien-Elliott, M.M.; Foltz, J.A.; Becker-Hapak, M.; Neal, C.C.; Foster, M.; Schappe, T.; McClain, E.; Pence, P.P.; et al. Donor memory-like NK cells persist and induce remissions in pediatric patients with relapsed AML after transplant. Blood 2022, 139, 1670–1683. [Google Scholar] [CrossRef] [PubMed]
- Stathopoulos, G.P.; Dimitroulis, J.; Antoniou, D.; Katis, C.; Tsavdaridis, D.; Armenaki, O.; Marosis, C.; Michalopoulou, P.; Grigoratou, T.; Stathopoulos, J. Front-line paclitaxel and irinotecan combination chemotherapy in advanced non-small-cell lung cancer: A phase I–II trial. Br. J. Cancer 2005, 93, 1106–1111. [Google Scholar] [CrossRef]
- Sochacka-Ćwikła, A.; Mączyński, M.; Regiec, A. FDA-Approved Drugs for Hematological Malignancies-The Last Decade Review. Cancers 2021, 14, 87. [Google Scholar] [CrossRef]
- Heipertz, E.L.; Zynda, E.R.; Stav-Noraas, T.E.; Hungler, A.D.; Boucher, S.E.; Kaur, N.; Vemuri, M.C. Current Perspectives on “Off-The-Shelf” Allogeneic NK and CAR-NK Cell Therapies. Front. Immunol. 2021, 12, 732135. [Google Scholar] [CrossRef]
- Guillerey, C.; Huntington, N.D.; Smyth, M.J. Targeting natural killer cells in cancer immunotherapy. Nat. Immunol. 2016, 17, 1025–1036. [Google Scholar] [CrossRef]
- Lamb, M.G.; Rangarajan, H.G.; Tullius, B.P.; Lee, D.A. Natural killer cell therapy for hematologic malignancies: Successes, challenges, and the future. Stem Cell Res. Ther. 2021, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.; Marin, D.; Banerjee, P.; Macapinlac, H.A.; Thompson, P.; Basar, R.; Kerbauy, L.N.; Overman, B.; Thall, P.; Kaplan, M.; et al. Use of CAR-Transduced Natural Killer Cells in CD19-Positive Lymphoid Tumors. N. Engl. J. Med. 2020, 382, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Becker, P.S.A.; Suck, G.; Nowakowska, P.; Ullrich, E.; Seifried, E.; Bader, P.; Tonn, T.; Seidl, C. Selection and expansion of natural killer cells for NK cell-based immunotherapy. Cancer Immunol. Immunother. 2016, 65, 477–484. [Google Scholar] [CrossRef]
- Eser, S.; Schnieke, A.; Schneider, G.; Saur, D. Oncogenic KRAS signalling in pancreatic cancer. Br. J. Cancer 2014, 111, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Chang, H.-H.; Cook, J.; Eibl, G.; Jewett, A. Suppression of Gingival NK Cells in Precancerous and Cancerous Stages of Pancreatic Cancer in KC and BLT-Humanized Mice. Front. Immunol. 2017, 8, 1606. [Google Scholar] [CrossRef]
- Kaur, K.; Chang, H.-H.; Topchyan, P.; Cook, J.M.; Barkhordarian, A.; Eibl, G.; Jewett, A. Deficiencies in Natural Killer Cell Numbers, Expansion, and Function at the Pre-Neoplastic Stage of Pancreatic Cancer by KRAS Mutation in the Pancreas of Obese Mice. Front. Immunol. 2018, 9, 1229. [Google Scholar] [CrossRef]
- Kaur, K.; Cook, J.; Park, S.-H.; Topchyan, P.; Kozlowska, A.; Ohanian, N.; Fang, C.; Nishimura, I.; Jewett, A. Novel Strategy to Expand Super-Charged NK Cells with Significant Potential to Lyse and Differentiate Cancer Stem Cells: Differences in NK Expansion and Function between Healthy and Cancer Patients. Front. Immunol. 2017, 8, 297. [Google Scholar] [CrossRef]
- Kim, H.W.; Lee, J.-C.; Paik, K.-H.; Kang, J.; Kim, J.; Hwang, J.-H. Serum interleukin-6 is associated with pancreatic ductal adenocarcinoma progression pattern. Medicine 2017, 96, e5926. [Google Scholar] [CrossRef]
- Long, K.B.; Tooker, G.; Tooker, E.; Luque, S.L.; Lee, J.W.; Pan, X.; Beatty, G.L. IL6 Receptor Blockade Enhances Chemotherapy Efficacy in Pancreatic Ductal Adenocarcinoma. Mol. Cancer Ther. 2017, 16, 1898–1908. [Google Scholar] [CrossRef]
- Nagathihalli, N.S.; Castellanos, J.A.; VanSaun, M.N.; Dai, X.; Ambrose, M.; Guo, Q.; Xiong, Y.; Merchant, N.B. Pancreatic stellate cell secreted IL-6 stimulates STAT3 dependent invasiveness of pancreatic intraepithelial neoplasia and cancer cells. Oncotarget 2016, 7, 65982–65992. [Google Scholar] [CrossRef] [PubMed]
- Goumas, F.A.; Holmer, R.; Egberts, J.; Gontarewicz, A.; Heneweer, C.; Geisen, U.; Hauser, C.; Mende, M.; Legler, K.; Röcken, C.; et al. Inhibition of IL-6 signaling significantly reduces primary tumor growth and recurrencies in orthotopic xenograft models of pancreatic cancer. Int. J. Cancer 2015, 137, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Lesina, M.; Kurkowski, M.U.; Ludes, K.; Rose-John, S.; Treiber, M.; Klöppel, G.; Yoshimura, A.; Reindl, W.; Sipos, B.; Akira, S.; et al. Stat3/Socs3 activation by IL-6 transsignaling promotes progression of pancreatic intraepithelial neoplasia and development of pancreatic cancer. Cancer Cell 2011, 19, 456–469. [Google Scholar] [CrossRef]
- Xu, Y.; Sun, J.; Sheard, M.A.; Tran, H.C.; Wan, Z.; Liu, W.Y.; Asgharzadeh, S.; Sposto, R.; Wu, H.W.; Seeger, R.C. Lenalidomide overcomes suppression of human natural killer cell anti-tumor functions by neuroblastoma microenvironment-associated IL-6 and TGFβ1. Cancer Immunol. Immunother. 2013, 62, 1637–1648. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.-C.; Arasteh, A.; Paranjpe, A.; Teruel, A.; Yang, W.; Behel, A.; Alva, J.A.; Walter, G.; Head, C.; Ishikawa, T.-O.; et al. Increased Lysis of Stem Cells but Not Their Differentiated Cells by Natural Killer Cells; De-Differentiation or Reprogramming Activates NK Cells. PLoS ONE 2010, 5, e11590. [Google Scholar] [CrossRef]
- Jewett, A.; Cacalano, N.A.; Teruel, A.; Romero, M.; Rashedi, M.; Wang, M.; Nakamura, H. Inhibition of nuclear factor kappa B (NFκB) activity in oral tumor cells prevents depletion of NK cells and increases their functional activation. Cancer Immunol. Immunother. 2005, 55, 1052–1063. [Google Scholar] [CrossRef] [PubMed]
- Kozlowska, A.K.; Tseng, H.-C.; Kaur, K.; Topchyan, P.; Inagaki, A.; Bui, V.T.; Kasahara, N.; Cacalano, N.; Jewett, A. Resistance to cytotoxicity and sustained release of interleukin-6 and interleukin-8 in the presence of decreased interferon-γ after differentiation of glioblastoma by human natural killer cells. Cancer Immunol. Immunother. 2016, 65, 1085–1097. [Google Scholar] [CrossRef]
- Kozlowska, A.; Topchyan, P.; Kaur, K.; Tseng, H.-C.; Teruel, A.; Hiraga, T.; Jewett, A. Differentiation by NK cells is a prerequisite for effective targeting of cancer stem cells/poorly differentiated tumors by chemopreventive and chemotherapeutic drugs. J. Cancer 2017, 8, 537–554. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Topchyan, P.; Kozlowska, A.K.; Ohanian, N.; Chiang, J.; Maung, P.O.; Park, S.-H.; Ko, M.-W.; Fang, C.; Nishimura, I.; et al. Super-charged NK cells inhibit growth and progression of stem-like/poorly differentiated oral tumors in vivo in humanized BLT mice; effect on tumor differentiation and response to chemotherapeutic drugs. OncoImmunology 2018, 7, e1426518. [Google Scholar] [CrossRef] [PubMed]
- Jewett, A.; Kozlowska, A.K.; Kaur, K.; Topchyan, P. Novel strategies to target cancer stem cells by NK cells studies in humanized mice. Front. Biosci. 2017, 22, 370–384. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Kozlowska, A.K.; Topchyan, P.; Ko, M.-W.; Ohanian, N.; Chiang, J.; Cook, J.; Maung, P.O.; Park, S.-H.; Cacalano, N.; et al. Probiotic-Treated Super-Charged NK Cells Efficiently Clear Poorly Differentiated Pancreatic Tumors in Hu-BLT Mice. Cancers 2019, 12, 63. [Google Scholar] [CrossRef]
- Kaur, K.; Topchyan, P.; Jewett, A. Supercharged Natural Killer (sNK) Cells Inhibit Melanoma Tumor Progression and Restore Endogenous NK Cell Function in Humanized BLT Mice. Cancers 2025, 17, 2430. [Google Scholar] [CrossRef]
- Türkseven, M.R.; Oygür, T. Evaluation of natural killer cell defense in oral squamous cell carcinoma. Oral Oncol. 2010, 46, e34–e37. [Google Scholar] [CrossRef]
- Accomando, W.P.; Wiencke, J.K.; Houseman, E.A.; Butler, R.A.; Zheng, S.; Nelson, H.H.; Kelsey, K.T. Decreased NK Cells in Patients with Head and Neck Cancer Determined in Archival DNA. Clin. Cancer Res. 2012, 18, 6147–6154. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mickel, R.A.; Kessler, D.J.; Taylor, J.M.; Lichtenstein, A. Natural killer cell cytotoxicity in the peripheral blood, cervical lymph nodes, and tumor of head and neck cancer patients. Cancer Res. 1988, 48, 5017–5022. [Google Scholar] [PubMed]
- Kaur, K.; Ko, M.-W.; Chen, F.; Jewett, A. Defective NK cell expansion, cytotoxicity, and lack of ability to differentiate tumors from a pancreatic cancer patient in a long term follow-up: Implication in the progression of cancer. Cancer Immunol. Immunother. 2021, 71, 1033–1047. [Google Scholar] [CrossRef]
- Aparicio-Pagés, M.N.; Verspaget, H.W.; Peña, A.S.; Lamers, C.B. Natural killer cell activity in patients with adenocarcinoma in the upper gastrointestinal tract. J. Clin. Lab. Immunol. 1991, 35, 27–32. [Google Scholar]
- Duan, X.; Deng, L.; Chen, X.; Lu, Y.; Zhang, Q.; Zhang, K.; Hu, Y.; Zeng, J.; Sun, W. Clinical significance of the immunostimulatory MHC class I chain-related molecule A and NKG2D receptor on NK cells in pancreatic cancer. Med Oncol. 2010, 28, 466–474. [Google Scholar] [CrossRef]
- Peng, Y.-P.; Zhu, Y.; Zhang, J.-J.; Xu, Z.-K.; Qian, Z.-Y.; Dai, C.-C.; Jiang, K.-R.; Wu, J.-L.; Gao, W.-T.; Li, Q.; et al. Comprehensive analysis of the percentage of surface receptors and cytotoxic granules positive natural killer cells in patients with pancreatic cancer, gastric cancer, and colorectal cancer. J. Transl. Med. 2013, 11, 262. [Google Scholar] [CrossRef]
- Jewett, A.; Man, Y.-G.; Tseng, H.-C. Dual Functions of Natural Killer Cells in Selection and Differentiation of Stem Cells; Role in Regulation of Inflammation and Regeneration of Tissues. J. Cancer 2013, 4, 12–24. [Google Scholar] [CrossRef]
- Imai, K.; Matsuyama, S.; Miyake, S.; Suga, K.; Nakachi, K. Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: An 11-year follow-up study of a general population. Lancet 2000, 356, 1795–1799. [Google Scholar] [CrossRef] [PubMed]
- Gogali, F.; Paterakis, G.; Rassidakis, G.Z.; Liakou, C.I.; Liapi, C. CD3(−)CD16(−)CD56(bright) immunoregulatory NK cells are increased in the tumor microenvironment and inversely correlate with advanced stages in patients with papillary thyroid cancer. Thyroid 2013, 23, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- López-Cobo, S.; Pieper, N.; Campos-Silva, C.; García-Cuesta, E.M.; Reyburn, H.T.; Paschen, A.; Valés-Gómez, M. Impaired NK cell recognition of vemurafenib-treated melanoma cells is overcome by simultaneous application of histone deacetylase inhibitors. OncoImmunology 2017, 7, e1392426. [Google Scholar] [CrossRef]
- Ciszak, L.; Kosmaczewska, A.; Werynska, B.; Szteblich, A.; Jankowska, R.; Frydecka, I. Impaired zeta chain expression and IFN-gamma production in peripheral blood T and NK cells of patients with advanced lung cancer. Oncol. Rep. 2009, 21, 173–184. [Google Scholar] [PubMed]
- Bui, V.T.; Tseng, H.-C.; Kozlowska, A.; Maung, P.O.; Kaur, K.; Topchyan, P.; Jewett, A. Augmented IFN-γ and TNF-α Induced by Probiotic Bacteria in NK Cells Mediate Differentiation of Stem-Like Tumors Leading to Inhibition of Tumor Growth and Reduction in Inflammatory Cytokine Release; Regulation by IL-10. Front. Immunol. 2015, 6, 576. [Google Scholar] [CrossRef]
- Jewett, A.; Tseng, H.-C. Tumor Induced Inactivation of Natural Killer Cell Cytotoxic Function; Implication in Growth, Expansion and Differentiation of Cancer Stem Cells. J. Cancer 2011, 2, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Pietra, G.; Manzini, C.; Rivara, S.; Vitale, M.; Cantoni, C.; Petretto, A.; Balsamo, M.; Conte, R.; Benelli, R.; Minghelli, S.; et al. Melanoma Cells Inhibit Natural Killer Cell Function by Modulating the Expression of Activating Receptors and Cytolytic Activity. Cancer Res. 2012, 72, 1407–1415. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Kato, S.; Nesline, M.K.; Conroy, J.M.; DePietro, P.; Pabla, S.; Kurzrock, R. Indoleamine 2,3-dioxygenase (IDO) inhibitors and cancer immunotherapy. Cancer Treat. Rev. 2022, 110, 102461. [Google Scholar] [CrossRef]
- Tseng, H.-C.; Cacalano, N.; Jewett, A. Split anergized natural killer cells halt inflammation by inducing stem cell differentiation, resistance to NK cell cytotoxicity and prevention of cytokine and chemokine secretion. Oncotarget 2015, 6, 8947–8959. [Google Scholar] [CrossRef]
- Kaur, K.; Ko, M.-W.; Ohanian, N.; Cook, J.; Jewett, A. Osteoclast-expanded super-charged NK-cells preferentially select and expand CD8+ T cells. Sci. Rep. 2020, 10, 1–17. [Google Scholar] [CrossRef]
- Tartter, P.I.; Steinberg, B.; Barron, D.M.; Martinelli, G. The Prognostic Significance of Natural Killer Cytotoxicity in Patients With Colorectal Cancer. Arch. Surg. 1987, 122, 1264–1268. [Google Scholar] [CrossRef]
- Igarashi, T.; Wynberg, J.; Srinivasan, R.; Becknell, B.; McCoy, J.P.; Takahashi, Y.; Suffredini, D.A.; Linehan, W.M.; Caligiuri, M.A.; Childs, R.W. Enhanced cytotoxicity of allogeneic NK cells with killer immunoglobulin-like receptor ligand incompatibility against melanoma and renal cell carcinoma cells. Blood 2004, 104, 170–177. [Google Scholar] [CrossRef]
- Alici, E.; Sutlu, T.; Björkstrand, B.; Gilljam, M.; Stellan, B.; Nahi, H.; Quezada, H.C.; Gahrton, G.; Ljunggren, H.-G.; Dilber, M.S. Autologous antitumor activity by NK cells expanded from myeloma patients using GMP-compliant components. Blood 2008, 111, 3155–3162. [Google Scholar] [CrossRef]
- Fujisaki, H.; Kakuda, H.; Shimasaki, N.; Imai, C.; Ma, J.; Lockey, T.; Eldridge, P.; Leung, W.H.; Campana, D. Expansion of highly cytotoxic human natural killer cells for cancer cell therapy. Cancer Res. 2009, 69, 4010–4017. [Google Scholar] [CrossRef] [PubMed]
- Berg, M.; Lundqvist, A.; McCoy, P.; Samsel, L.; Fan, Y.; Tawab, A.; Childs, R. Clinical-grade ex vivo-expanded human natural killer cells up-regulate activating receptors and death receptor ligands and have enhanced cytolytic activity against tumor cells. Cytotherapy 2009, 11, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; Martín-Antonio, B.; Yang, H.; Ku, S.; Lee, D.A.; Cooper, L.J.N.; Decker, W.K.; Li, S.; Robinson, S.N.; Sekine, T.; et al. Antigen Presenting Cell-Mediated Expansion of Human Umbilical Cord Blood Yields Log-Scale Expansion of Natural Killer Cells with Anti-Myeloma Activity. PLoS ONE 2013, 8, e76781. [Google Scholar] [CrossRef]
- Jewett, A.; Cacalano, N.A.; Head, C.; Teruel, A. Coengagement of CD16 and CD94 Receptors Mediates Secretion of Chemokines and Induces Apoptotic Death of Naive Natural Killer Cells. Clin. Cancer Res. 2006, 12, 1994–2003. [Google Scholar] [CrossRef]
- Cooper, M.A.; Fehniger, T.A.; Caligiuri, M.A. The biology of human natural killer-cell subsets. Trends Immunol. 2001, 22, 633–640. [Google Scholar] [CrossRef]
- Farag, S.S.; Caligiuri, M.A. Human natural killer cell development and biology. Blood Rev. 2006, 20, 123–137. [Google Scholar] [CrossRef]
- Sun, H.; Sun, C.; Tian, Z.; Xiao, W. NK cells in immunotolerant organs. Cell. Mol. Immunol. 2013, 10, 202–212. [Google Scholar] [CrossRef]
- Kaur, K.; Topchyan, P.; Kozlowska, A.K.; Ohanian, N.; Chiang, J.; Maung, P.O.; Park, S.-H.; Ko, M.-W.; Fang, C.; Nishimura, I.; et al. Super-charged NK cells inhibit growth and progression of stem-like/poorly differentiated oral tumors in vivo in humanized BLT mice; effect on tumor differentiation and response to chemotherapeutic drugs. OncoImmunology 2018, 7, e1426518. [Google Scholar] [CrossRef] [PubMed]
- Jewett, A.; Teruel, A.; Romero, M.; Head, C.; Cacalano, N. Rapid and potent induction of cell death and loss of NK cell cytotoxicity against oral tumors by F(ab′)2 fragment of anti-CD16 antibody. Cancer Immunol. Immunother. 2008, 57, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Jewett, A.; Cavalcanti, M.; Bonavida, B. Pivotal role of endogenous TNF-alpha in the induction of functional inactivation and apoptosis in NK cells. J. Immunol. 1997, 159, 4815–4822. [Google Scholar] [CrossRef] [PubMed]
- Jewett, A.; Bonavida, B. Target-induced inactivation and cell death by apoptosis in a subset of human NK cells. J. Immunol. 1996, 156, 907–915. [Google Scholar] [CrossRef]
- Jewett, A.; Bonavida, B. Target-induced anergy of natural killer cytotoxic function is restricted to the NK—Target conjugate subset. Cell. Immunol. 1995, 160, 91–97. [Google Scholar] [CrossRef]
- Woroniecka, K.; Chongsathidkiet, P.; Rhodin, K.; Kemeny, H.; Dechant, C.; Farber, S.H.; Elsamadicy, A.A.; Cui, X.; Koyama, S.; Jackson, C.; et al. T-Cell Exhaustion Signatures Vary with Tumor Type and Are Severe in Glioblastoma. Clin. Cancer Res. 2018, 24, 4175–4186. [Google Scholar] [CrossRef]
- Schwartz, R.H. T cell anergy. Annu. Rev. Immunol. 2003, 21, 305–334. [Google Scholar] [CrossRef] [PubMed]
- Bonavida, B.; Lebow, L.T.; Jewett, A. Natural killer cell subsets: Maturation, differentiation and regulation. Nat. Immun. 1993, 12, 194–208. [Google Scholar]
- Terranova-Barberio, M.; Pawlowska, N.; Dhawan, M.; Moasser, M.; Chien, A.J.; Melisko, M.E.; Rugo, H.; Rahimi, R.; Deal, T.; Daud, A.; et al. Exhausted T cell signature predicts immunotherapy response in ER-positive breast cancer. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Chen, E.X.; Jonker, D.J.; Loree, J.M.; Kennecke, H.F.; Berry, S.R.; Couture, F.; Ahmad, C.E.; Goffin, J.R.; Kavan, P.; Harb, M.; et al. Effect of Combined Immune Checkpoint Inhibition vs Best Supportive Care Alone in Patients With Advanced Colo-rectal Cancer: The Canadian Cancer Trials Group CO.26 Study. JAMA Oncol. 2020, 6, 831–838. [Google Scholar] [CrossRef]
- Rosenberg, S.A.; Restifo, N.P. Adoptive cell transfer as personalized immunotherapy for human cancer. Science 2015, 348, 62–68. [Google Scholar] [CrossRef]
- Sharma, P.; Allison, J.P. The future of immune checkpoint therapy. Science 2015, 348, 56–61. [Google Scholar] [CrossRef]
- Fang, F.; Xie, S.; Chen, M.; Li, Y.; Yue, J.; Ma, J.; Shu, X.; He, Y.; Xiao, W.; Tian, Z. Advances in NK cell production. Cell. Mol. Immunol. 2022, 19, 460–481. [Google Scholar] [CrossRef] [PubMed]
- Jewett, A.; Kos, J.; Kaur, K.; Safaei, T.; Sutanto, C.; Chen, W.; Wong, P.; Namagerdi, A.K.; Fang, C.; Fong, Y.; et al. Natural Killer Cells: Diverse Functions in Tumor Immunity and Defects in Pre-neoplastic and Neoplastic Stages of Tumorigenesis. Mol. Ther.-Oncolytics 2020, 16, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.C.; Arasteh, A.; Kaur, K.; Kozlowska, A.; Topchyan, P.; Jewett, A. Differential Cytotoxicity but Augmented IFN-gamma Secretion by NK Cells after Interaction with Monocytes from Humans, and Those from Wild Type and Myeloid-Specific COX-2 Knockout Mice. Front. Immunol. 2015, 6, 259. [Google Scholar] [CrossRef]
- Jewett, A.; Wang, M.-Y.; Teruel, A.; Poupak, Z.; Bostanian, Z.; Park, N.-H. Cytokine dependent inverse regulation of CD54 (ICAM1) and major histocompatibility complex class I antigens by nuclear factor κB in HEp2 tumor cell line: Effect on the function of natural killer cells. Hum. Immunol. 2003, 64, 505–520. [Google Scholar] [CrossRef]
- Wu, J.; Cherwinski, H.; Spies, T.; Phillips, J.H.; Lanier, L.L. Dap10 and Dap12 Form Distinct, but Functionally Cooperative, Receptor Complexes in Natural Killer Cells. J. Exp. Med. 2000, 192, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Perussia, B.; Ramoni, C.; Anegon, I.; Cuturi, M.C.; Faust, J.; Trinchieri, G. Preferential proliferation of natural-killer-cells among peripheral-blood mononuclear-cells cocultured with b-lymphoblastoid cell-lines. Nat. Immun. Cell Growth Regul. 1987, 6, 171–188. [Google Scholar] [PubMed]
- Rabinowich, H.; Sedlmayr, P.; Herberman, R.B.; Whiteside, T.L. Increased proliferation, lytic activity, and purity of human natural killer cells cocultured with mitogen-activated feeder cells. Cell. Immunol. 1991, 135, 454–470. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Lundqvist, A.; Childs, R. Natural killer cell immunotherapy for cancer: A new hope. Cytotherapy 2008, 10, 775–783. [Google Scholar] [CrossRef]
- Gras Navarro, A.; Björklund, A.T.; Chekenya, M. Therapeutic Potential and Challenges of Natural Killer Cells in Treatment of Solid Tumors. Front. Immunol. 2015, 6, 202. [Google Scholar] [CrossRef]
- Alici, E.; Sutlu, T.; Björkstrand, B.; Gilljam, M.; Stellan, B.; Nahi, H.; Quezada, H.C.; Gahrton, G.; Ljunggren, H.-G.; Dilber, M.S. Autologous antitumor activity by NK cells expanded from myeloma patients using GMP-compliant components. Blood 2008, 111, 3155–3162. [Google Scholar] [CrossRef]
- Garg, T.K.; Szmania, S.M.; Khan, J.A.; Hoering, A.; Malbrough, P.A.; Moreno-Bost, A.; Greenway, A.D.; Lingo, J.D.; Li, X.; Yaccoby, S.; et al. Highly activated and expanded natural killer cells for multiple myeloma immunotherapy. Haematologica 2012, 97, 1348–1356. [Google Scholar] [CrossRef]
- Kaur, K.; Cook, J.; Park, S.-H.; Topchyan, P.; Kozlowska, A.; Ohanian, N.; Fang, C.; Nishimura, I.; Jewett, A. Novel Strategy to Expand Super-Charged NK Cells with Significant Potential to Lyse and Differentiate Cancer Stem Cells: Differences in NK Expansion and Function between Healthy and Cancer Patients. Front. Immunol. 2017, 8, 297. [Google Scholar] [CrossRef]
- Kaur, K.; Chen, P.-C.; Ko, M.-W.; Mei, A.; Senjor, E.; Malarkannan, S.; Kos, J.; Jewett, A. Sequential therapy with supercharged NK cells with either chemotherapy drug cisplatin or anti-PD-1 antibody decreases the tumor size and significantly enhances the NK function in Hu-BLT mice. Front. Immunol. 2023, 14, 1132807. [Google Scholar] [CrossRef]
- Voskens, C.J.; Watanabe, R.; Rollins, S.; Campana, D.; Hasumi, K.; Mann, D.L. Ex-vivo expanded human NK cells express activating receptors that mediate cytotoxicity of allogeneic and autologous cancer cell lines by direct recognition and antibody directed cellular cytotoxicity. J. Exp. Clin. Cancer Res. 2010, 29, 134. [Google Scholar] [CrossRef]
- Yang, H.; Tang, R.; Li, J.; Liu, Y.; Ye, L.; Shao, D.; Jin, M.; Huang, Q.; Shi, J. A New Ex Vivo Method for Effective Expansion and Activation of Human Natural Killer Cells for Anti-Tumor Immunotherapy. Cell Biochem. Biophys. 2015, 73, 723–729. [Google Scholar] [CrossRef]
- Fujisaki, H.; Kakuda, H.; Imai, C.; Mullighan, C.G.; Campana, D. Replicative potential of human natural killer cells. Br. J. Haematol. 2009, 145, 606–613. [Google Scholar] [CrossRef]
- Chang, Y.H.; Connolly, J.; Shimasaki, N.; Mimura, K.; Kono, K.; Campana, D. A chimeric receptor with NKG2D specificity enhances natural killer cell activation and killing of tumor cells. Cancer Res. 2013, 73, 1777–1786. [Google Scholar] [CrossRef] [PubMed]
- Lapteva, N.; Durett, A.G.; Sun, J.; Rollins, L.A.; Huye, L.L.; Fang, J.; Dandekar, V.; Mei, Z.; Jackson, K.; Vera, J.; et al. Large-scale ex vivo expansion and characterization of natural killer cells for clinical applications. Cytotherapy 2012, 14, 1131–1143. [Google Scholar] [CrossRef]
- Moseman, J.E.; Foltz, J.A.; Sorathia, K.; Heipertz, E.L.; Lee, D.A. Evaluation of serum-free media formulations in feeder cell–stimulated expansion of natural killer cells. Cytotherapy 2020, 22, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Y.; Jiang, Q.; Jiang, H.; Hu, L.J.; Zhao, T.; Yu, X.X.; Huang, X.J. Expanded clinical-grade membrane-bound IL-21/4-1BBL NK cell products exhibit activity against acute myeloid leu-kemia in vivo. Eur. J. Immunol. 2020, 50, 1374–1385. [Google Scholar] [CrossRef]
- Zhang, C.; Kadu, S.; Xiao, Y.; Johnson, O.; Kelly, A.; O’COnnor, R.S.; Lai, M.; Kong, H.; Srivatsa, S.; Tai, V.; et al. Sequential Exposure to IL21 and IL15 During Human Natural Killer Cell Expansion Optimizes Yield and Function. Cancer Immunol. Res. 2023, 11, 1524–1537. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.-C.; Arasteh, A.; Kaur, K.; Kozlowska, A.; Topchyan, P.; Jewett, A. Differential cytotoxicity but augmented IFN-γ secretion by NK cells after interaction with monocytes from hu-mans, and those from wild type and myeloid specific COX-2 knockout mice. Front. Immunol. 2015, 6, 259. [Google Scholar] [CrossRef]
- Tseng, H.C.; Kanayama, K.; Kaur, K.; Park, S.H.; Park, S.; Kozlowska, A.; Sun, S.; McKenna, C.E.; Nishimura, I.; Jewett, A. Bisphosphonate-induced differential modulation of immune cell function in gingiva and bone marrow in vivo: Role in osteoclast-mediated NK cell activation. Oncotarget 2015, 6, 20002–20025. [Google Scholar] [CrossRef]
- Leivas, A.; Perez-Martínez, A.; Blanchard, M.J.; Martín-Clavero, E.; Fernández, L.; Lahuerta, J.J.; Martinez-Lopez, J. Novel treatment strategy with autologous activated and expanded natural killer cells plus anti-myeloma drugs for multiple myeloma. OncoImmunology 2016, 5, e1250051. [Google Scholar] [CrossRef]
- Cichocki, F.; Bjordahl, R.; Gaidarova, S.; Mahmood, S.; Abujarour, R.; Wang, H.; Tuininga, K.; Felices, M.; Davis, Z.B.; Bendzick, L.; et al. iPSC-derived NK cells maintain high cytotoxicity and enhance in vivo tumor control in concert with T cells and anti–PD-1 therapy. Sci. Transl. Med. 2020, 12, eaaz5618. [Google Scholar] [CrossRef]
- Benson, D.M., Jr.; Bakan, C.E.; Mishra, A.; Hofmeister, C.C.; Efebera, Y.; Becknell, B.; Baiocchi, R.A.; Zhang, J.; Yu, J.; Smith, M.K.; et al. The PD-1/PD-L1 axis modulates the natural killer cell versus multiple myeloma effect: A therapeutic target for CT-011, a novel monoclonal anti–PD-1 antibody. Blood 2010, 116, 2286–2294. [Google Scholar] [CrossRef]
- Kaur, K.; Chen, P.-C.; Ko, M.-W.; Mei, A.; Senjor, E.; Malarkannan, S.; Kos, J.; Jewett, A. Sequential therapy with supercharged NK cells with either chemotherapy drug cisplatin or anti-PD-1 antibody decreases the tumor size and significantly enhances the NK function in Hu-BLT mice. Front. Immunol. 2023, 14, 1132807. [Google Scholar] [CrossRef]
- Sadeghi, S.; Chen, P.-C.; Jewett, A.; Kaur, K. Combination of NK cell immunotherapy with chemotherapy and radiation enhances NK cell therapy and provides improved prognosis in cancer patients and in humanized BLT mouse model system. In NK Cells in Cancer Immunotherapy: Successes and Challenges; Academic Press: Cambridge, MA, USA, 2023; pp. 301–320. [Google Scholar] [CrossRef]
- Senjor, E.; Ko, M.-W.; Kaur, K.; Chen, P.-C.; Breznik, B.; Chovatiya, N.; Kos, J.; Jewett, A. Multifaceted nature of natural killer cells: Potential mode of interaction and shaping of stem cells. In NK Cells in Cancer Immunotherapy: Successes and Challenges; Academic Press: Cambridge, MA, USA, 2023; pp. 3–25. [Google Scholar] [CrossRef]
- Ko, M.-W.; Kaur, K.; Chen, P.-C.; Breznik, B.; Senjor, E.; Chovatiya, N.; Wong, P.; Turnsek, T.L.; Kos, J.; Jewett, A. Diagnostic methods to assess the numbers, phenotype, and function of primary and engineered NK cells: Methods to predict prognosis and treatment outcome. In NK Cells in Cancer Immunotherapy: Successes and Challenges; Academic Press: Cambridge, MA, USA, 2023; pp. 281–297. [Google Scholar] [CrossRef]
- Kaur, K.; Ko, M.-W.; Chen, F.; Jewett, A. Defective NK cell expansion, cytotoxicity, and lack of ability to differentiate tumors from a pancreatic cancer patient in a long term follow-up: Implication in the progression of cancer. Cancer Immunol. Immunother. 2021, 71, 1033–1047. [Google Scholar] [CrossRef]
- Kaur, K.; Safaie, T.; Ko, M.W.; Wang, Y.; Jewett, A. ADCC against MICA/B Is Mediated against Differentiated Oral and Pancreatic and Not Stem-Like/Poorly Differentiat-ed Tumors by the NK Cells; Loss in Cancer Patients due to Down-Modulation of CD16 Receptor. Cancers 2021, 13, 239. [Google Scholar] [CrossRef] [PubMed]
- Jewett, A.; Kos, J.; Turnsek, T.L.; Chen, P.-C.; Breznik, B.; Senjor, E.; Chovatiya, N.; Kaur, K.; Ko, M.-W. Novel strategies to expand supercharged NK cells with augmented capacity to withstand inactivation by tumors. In Successes and Challenges of NK Immunotherapy; Academic Press: Cambridge, MA, USA, 2021; pp. 101–119. [Google Scholar] [CrossRef]
- Kaur, K.; Ko, M.-W.; Chen, P.-C.; Breznik, B.; Senjor, E.; Wong, P.; Wang, Y.; Chovatiya, N.; Jewett, A. Probiotics in Health and Disease: Distinct Roles of Different Strains in Natural Killer Cell Activation and Regulation. Crit. Rev. Immunol. 2021, 41, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Ko, M.-W.; Ohanian, N.; Cook, J.; Jewett, A. Osteoclast-expanded super-charged NK-cells preferentially select and expand CD8+ T cells. Sci. Rep. 2020, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Jewett, A.; Kos, J.; Kaur, K.; Turnsek, T.L.; Breznik, B.; Senjor, E.; Wong, P.; Nguyen, K.Y.; Ko, M.-W. Multiple Defects of Natural Killer Cells in Cancer Patients: Anarchy, Dysregulated Systemic Immunity, and Immunosuppression in Metastatic Cancer. Crit. Rev. Immunol. 2020, 40, 93–133. [Google Scholar] [CrossRef]
- Jewett, A.; Kos, J.; Fong, Y.; Ko, M.-W.; Safaei, T.; Nanut, M.P.; Kaur, K. NK cells shape pancreatic and oral tumor microenvironments; role in inhibition of tumor growth and metastasis. Semin. Cancer Biol. 2018, 53, 178–188. [Google Scholar] [CrossRef]
- Kaur, K.; Nanut, M.P.; Ko, M.-W.; Safaie, T.; Kos, J.; Jewett, A. Natural killer cells target and differentiate cancer stem-like cells/undifferentiated tumors: Strategies to optimize their growth and expansion for effective cancer immunotherapy. Curr. Opin. Immunol. 2018, 51, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Chang, H.-H.; Cook, J.; Eibl, G.; Jewett, A. Suppression of Gingival NK Cells in Precancerous and Cancerous Stages of Pancreatic Cancer in KC and BLT-Humanized Mice. Front. Immunol. 2017, 8, 1606. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Cook, J.; Park, S.-H.; Topchyan, P.; Kozlowska, A.; Ohanian, N.; Fang, C.; Nishimura, I.; Jewett, A. Novel Strategy to Expand Super-Charged NK Cells with Significant Potential to Lyse and Differentiate Cancer Stem Cells: Differences in NK Expansion and Function between Healthy and Cancer Patients. Front. Immunol. 2017, 8, 297. [Google Scholar] [CrossRef] [PubMed]
- Kozlowska, A.K.; Kaur, K.; Topchyan, P.; Jewett, A. Novel strategies to target cancer stem cells by NK cells; studies in humanized mice. Front. Biosci. 2017, 22, 370–384. [Google Scholar] [CrossRef]
- Kozlowska, A.K.; Kaur, K.; Topchyan, P.; Jewett, A. Adoptive transfer of osteoclast-expanded natural killer cells for immunotherapy targeting cancer stem-like cells in humanized mice. Cancer Immunol. Immunother. 2016, 65, 835–845. [Google Scholar] [CrossRef]
- Breznik, B.; Novak, M.; Majc, B.; Habič, A.; Jewett, A. Natural killer cells in the treatment of glioblastoma: Diverse antitumor functions and potential clinical applications. In NK Cells in Cancer Immunotherapy: Successes and Challenges; Academic Press: Cambridge, MA, USA, 2023; pp. 335–367. [Google Scholar] [CrossRef]
- Breznik, B.; Ko, M.; Chen, P.; Senjor, E.; Majc, B.; Novak, M.; Habič, A.; Jewett, A. P06.07.A Natural killer cells lyse glioblastoma stem cells and increase their sensitivity to chemotherapy. Neuro-Oncology 2022, 24, ii39. [Google Scholar] [CrossRef]
- Breznik, B.; Ko, M.-W.; Tse, C.; Chen, P.-C.; Senjor, E.; Majc, B.; Habič, A.; Angelillis, N.; Novak, M.; Župunski, V.; et al. Infiltrating natural killer cells bind, lyse and increase chemotherapy efficacy in glioblastoma stem-like tumorospheres. Commun. Biol. 2022, 5, 1–15. [Google Scholar] [CrossRef]
- Cekic, C.; Day, Y.J.; Sag, D.; Linden, J. Myeloid expression of adenosine A2A receptor suppresses T and NK cell responses in the solid tumor microenviron-ment. Cancer Res. 2014, 74, 7250–7259. [Google Scholar] [CrossRef]
Figure 1.
The illustration shows the effects of obesity, gene mutation, and unhealthy diet on the phenotype of NK cells and NK activating cells to induce preneoplastic conditions, and further inhibition of NK cells and NK activating cells in cancer.
Figure 1.
The illustration shows the effects of obesity, gene mutation, and unhealthy diet on the phenotype of NK cells and NK activating cells to induce preneoplastic conditions, and further inhibition of NK cells and NK activating cells in cancer.
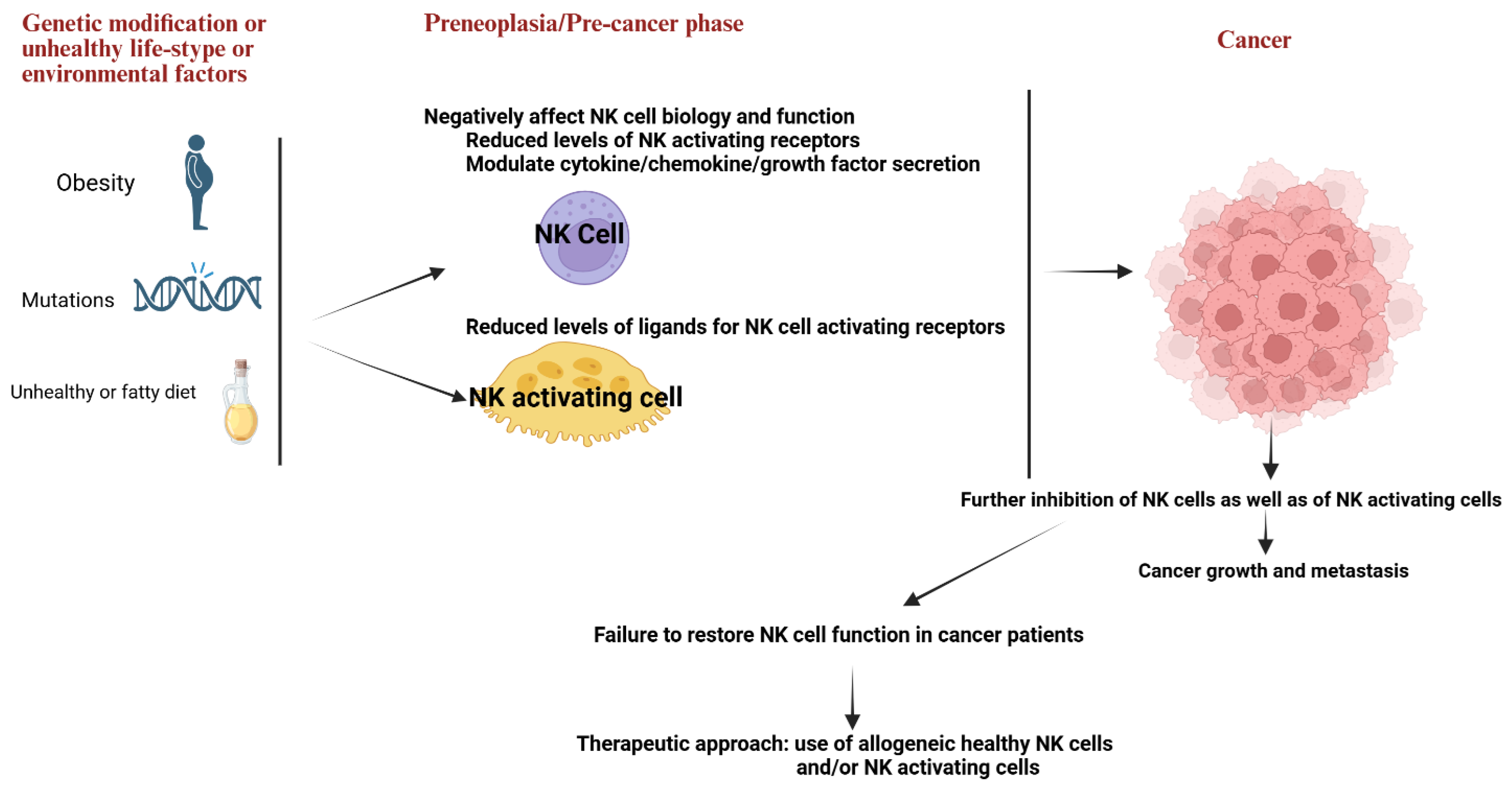
Figure 2.
The illustration shows four NK cell maturation stages and characteristics of NK cell subsets at each stage.
Figure 2.
The illustration shows four NK cell maturation stages and characteristics of NK cell subsets at each stage.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



