Submitted:
11 August 2025
Posted:
12 August 2025
You are already at the latest version
Abstract
During the evolutionary development of camels, circumstances of the habitat led to the emergence of various types of adaptive mechanisms in the organisms of these animals. One of them is fat accumulation. Features of fat metabolism, manifested in increased secretion and accumulation of lipids, inevitably have a direct impact on its structure. Considering the latter, the goal of this research is to establish the features of the microstructural organization of the liver of the Bactrian camel (Camelus bactrianus). The studies were conducted using liver tissue fragments. The sections were examined using a Jem-1011 electron microscope at magnifications of 2500–8000. Unlike other mammals, the camel’s liver exhibits a number of characteristics that can be linked to its lifestyle and metabolic requirements. Firstly, the significant number of hepatocytes and their special morphological characteristics indicate a high capacity for metabolism and detoxification. Secondly, the observed increase in the amount of fat deposits in hepatocytes may be a sign of adaptation to periods of food shortage. Our studies highlight the great diversity of dromedary liver structure and raise important questions about the influence of various environmental factors on its morphology.
Keywords:
1. Introduction
2. Materials and Methods
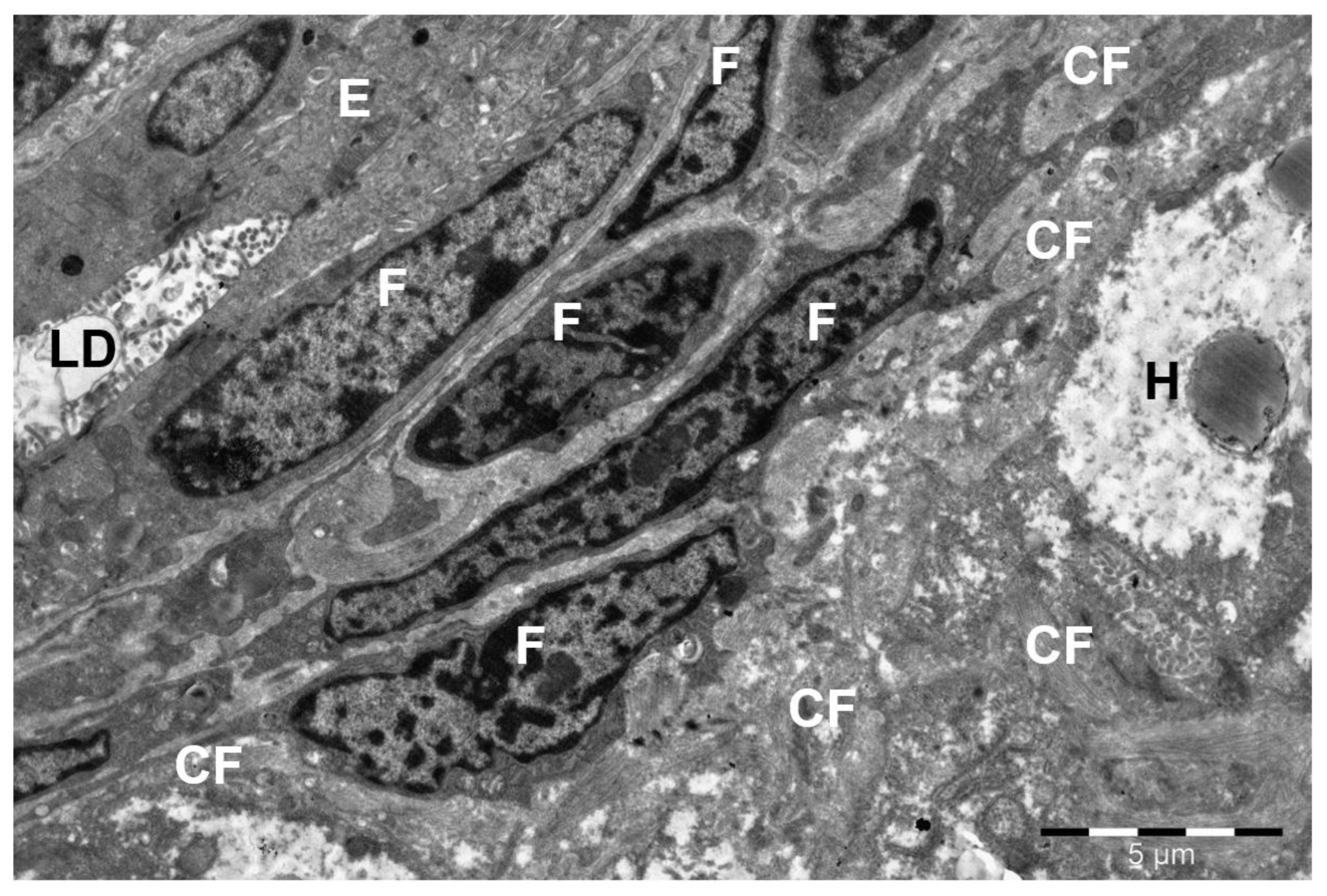
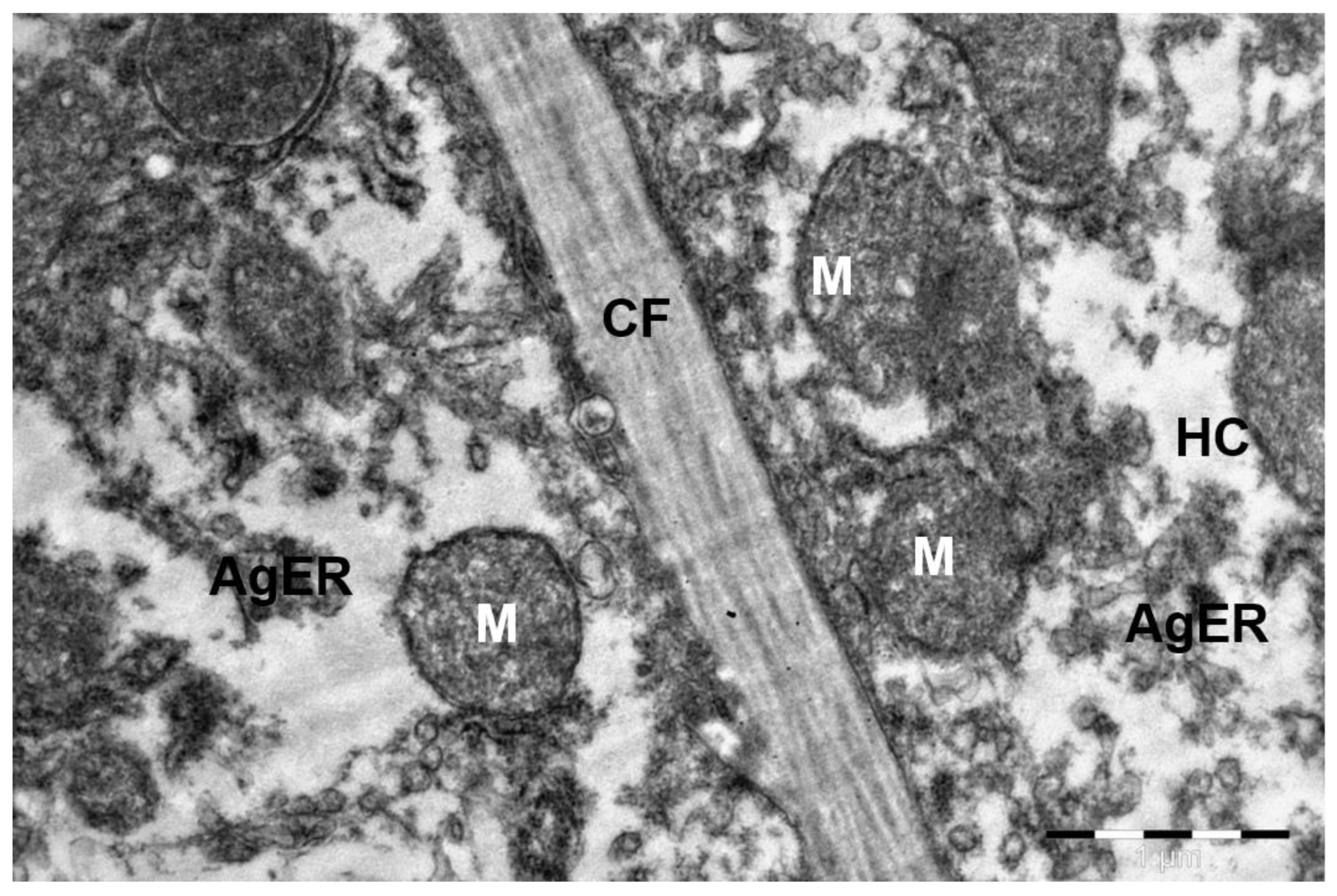
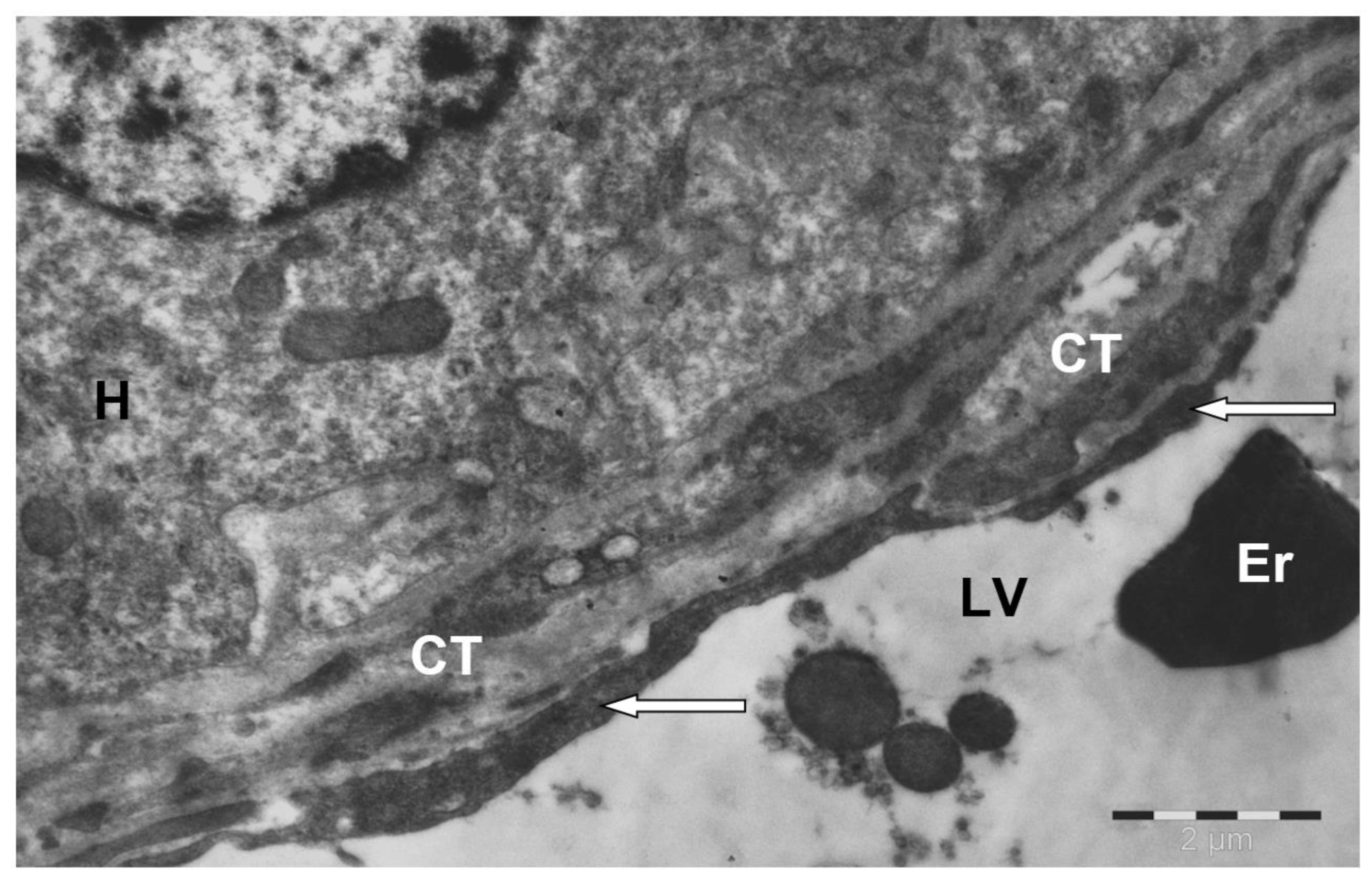
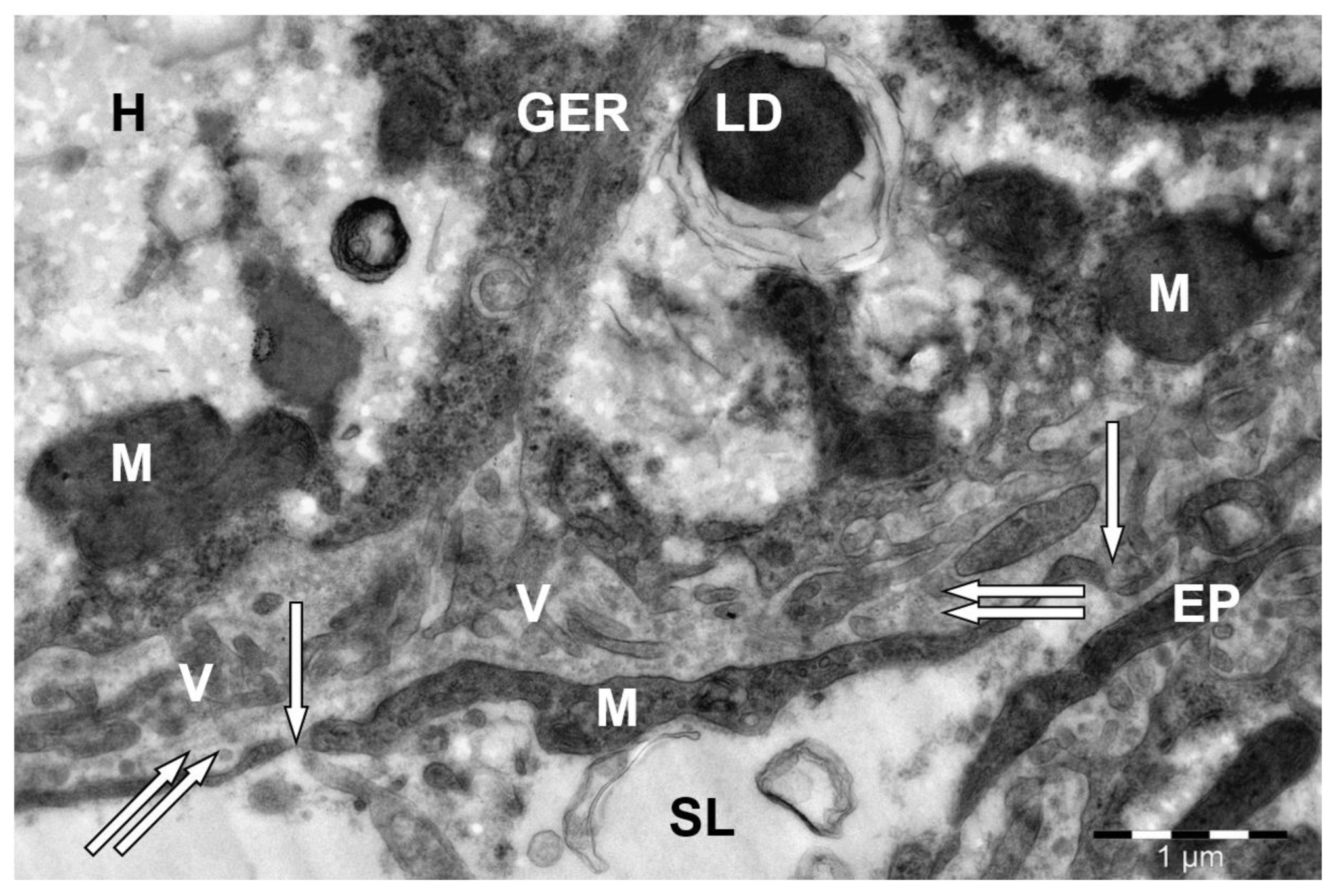
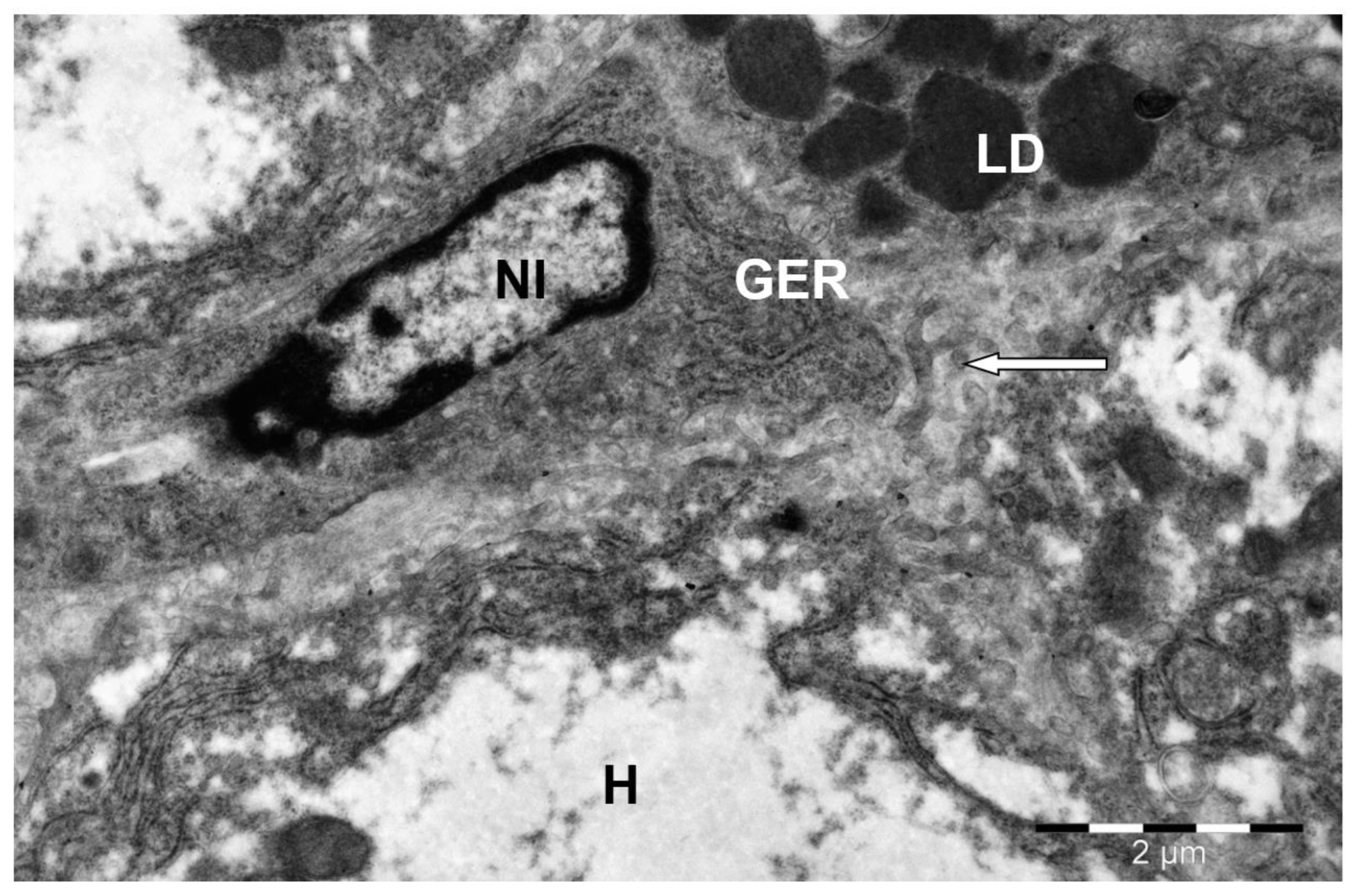
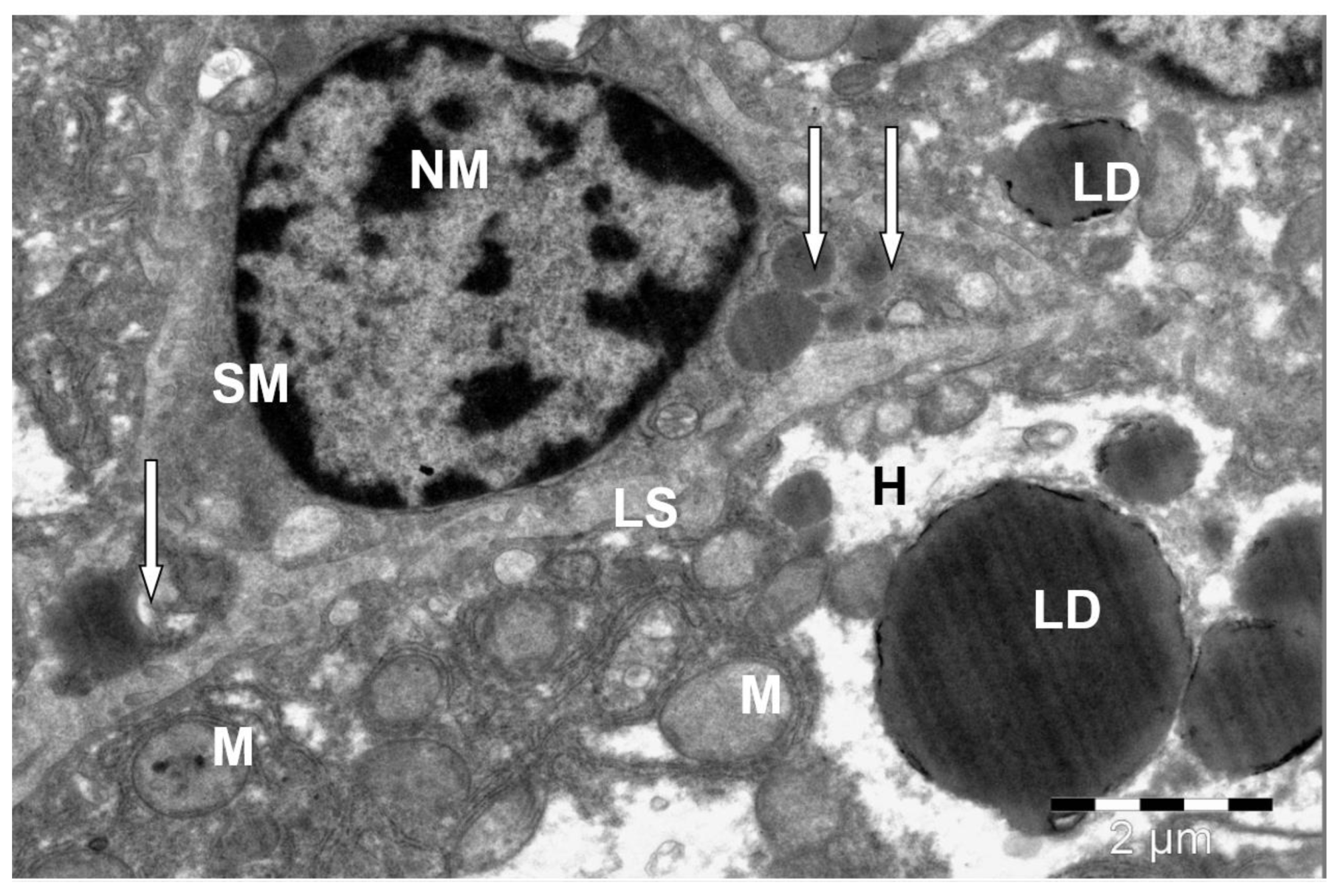
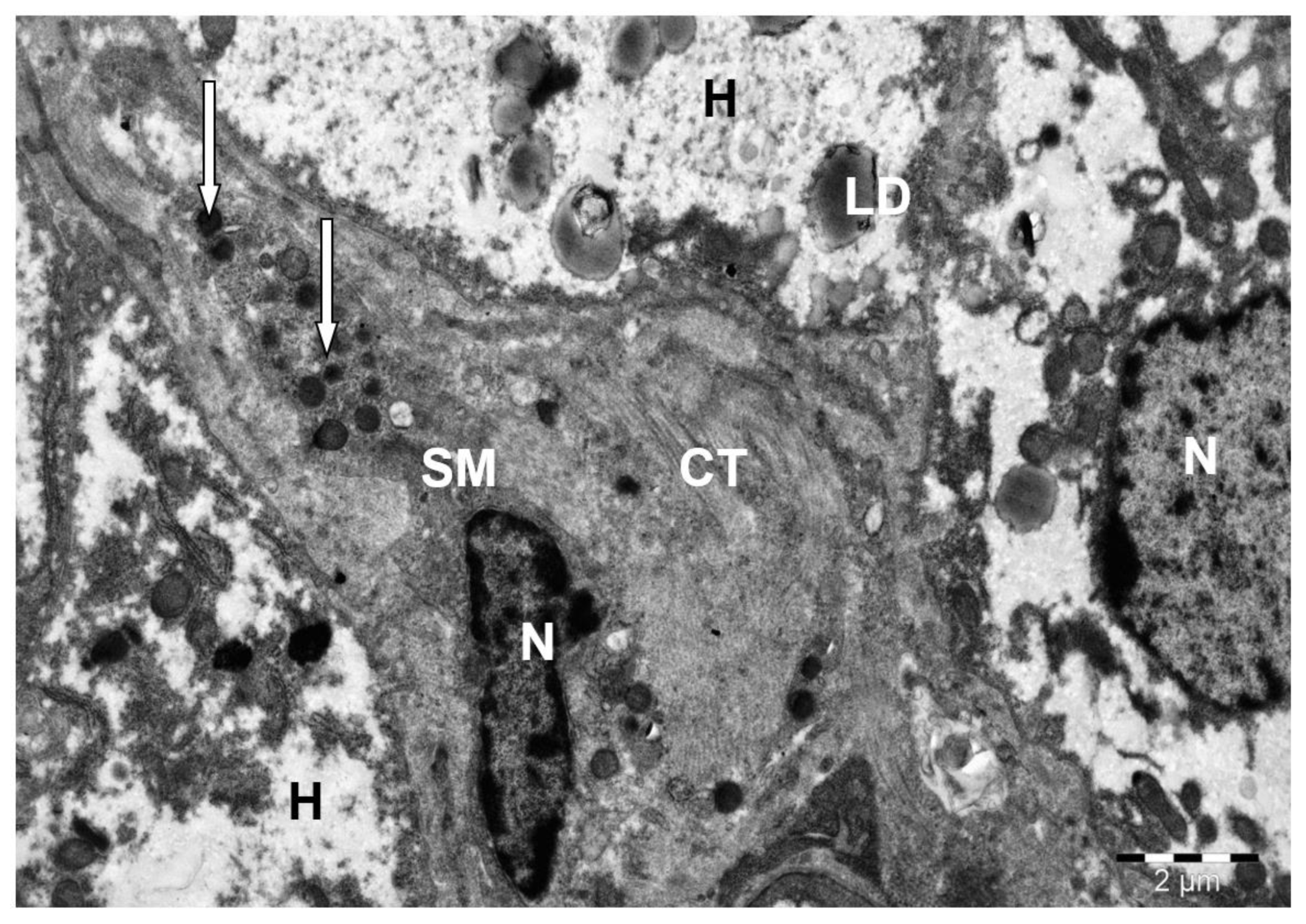
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ali MA, Abu Damir H, Adem MA, et al. Effects of long-term dehydration on stress markers, blood parameters, and tissue morphology in the dromedary camel (Camelus dromedarius). Front Vet Sci. 2023; 10: 1236425. [CrossRef]
- Riley J, Garner MM, Kiupel M, et al. Disseminated toxoplasmosis in a captive adult dromedary camel (Camelus dromedarius). J Zoo Wildl Med. 2017;48(3):937-940. [CrossRef]
- Zeyadi M. Effect of organic additives on storage stability of camel liver catalase against environmental conditions. Main Group Chem. 2022;21(1):225-231. [CrossRef]
- Tharwat M, Alkheraif AA, Oikawa S. Production diseases in farm animals: A comprehensive and illustrated clinical, laboratory, and pathological overview. Open Vet J. 2025;15(1):18-34. [CrossRef]
- Yashin A, Kasatkina E, Prusakov A, et al. The T-RFLP research method in the study of rumen microbiota in dairy cows with subclinical ketosis. In: Bio Web Conf: Int Sci Pract Conf “Agrarian Science – 2023” (AgriScience2023); April 25–26, 2023; Moscow, Russia. EDP Sciences; 2023:06001. [CrossRef]
- Terab AMA, Abdel Wahab GED, Ishag HZA, et al. Pathology, bacteriology and molecular studies on caseous lymphadenitis in Camelus dromedarius in the Emirate of Abu Dhabi, UAE, 2015–2020. PLoS One. 2021;16(6):e0252893. [CrossRef]
- Tharwat M. Ultrasonography of the liver in healthy and diseased camels (Camelus dromedarius). J Vet Med Sci. 2020;82(4):399-407. [CrossRef]
- Tharwat M, Ali H, Alkheraif AA. Clinical insights on paratuberculosis in Arabian camels (Camelus dromedarius): A review. Open Vet J. 2025;15(1):8-17. [CrossRef]
- Al-Bar OAM, El-Shishtawy RM, Mohamed SA. Immobilization of camel liver catalase on nanosilver-coated cotton fabric. Catalysts. 2021;11(8). [CrossRef]
- El Saftawy EA, Abdelmoktader A, Sabry MM, et al. Histological and immunological insights to hydatid disease in camels. Vet Parasitol Reg Stud Rep. 2021;26:100635. [CrossRef]
- Panwar A, Thanvi PK, Dangi A, et al. Histochemical studies of the liver of dromedary camel (Camelus dromedarius). Int J Adv Biochem Res. 2025;9(3S):5-9. [CrossRef]
- Alekhin YN, Popova OS, Ponomarev VS, et al. Effect of farnesoid X receptor agonist on postprandial lipemia in rats fed a diet containing a supraphysiological dose of fat. Dev Regist Drugs. 2023;12(2):174-184. [CrossRef]
- Kuznetsov YuE, Lunegov AM, Ponomarev VS, et al. Correlation relationships between the content of total bile acids and the main biochemical parameters of blood in minks (Mustela vison Schreber, 1777). Agric Biol. 2022;57(6):1217-1224. [CrossRef]
- Prusakova A, Zelenevskiy N, Prusakov A, et al. Organization of histo-hematic barriers of the liver in Anglo-Nubian goat. Online J Anim Feed Res. 2023;13(4):242-245. [CrossRef]
- Ibrahim A, El-Ghareeb WR, Aljazzar A, et al. Hepatic lobe torsion in 3 dromedary camels. J Vet Diagn Invest. 2021;33(1):136-139. [CrossRef]
- Ponamarev V, Popova O, Kostrova A, et al. A new method for assessing the toxic properties of various medicinal substances on the hepatobiliary system functionality in the context of the ecopharmacology development. In: II Int Conf Sustainable Development: Agriculture, Veterinary Medicine and Ecology (VMAEE-II-2023); April 21–22, 2023; Karshi, Russia. AIP Publishing; 2023:20027. [CrossRef]
- Prusakova AV, Zelenevskiy NV, Prusakov AV, et al. Ultrastructural organization of liver hepatocytes of the Anglo-Nubian goat. Vet Glasn. 2023;77(2):176-187. [CrossRef]
- Anwar Ul-H. A Beginners’ Guide to Scanning Electron Microscopy. 1st ed. Springer Nature; 2018.
- Ramezanpour H, Yousefi H, Rezaei M, et al. Effects of rotational motion in robotic needle insertion. J Biomed Phys Eng. 2015;5(4):207-216.
- Semchenko VV. International Histological Nomenclature. Kolosova; 1999.
- Ahmed A, Malik A, Jagirdar H, et al. Copper-induced inactivation of camel liver glutathione S-transferase. Biol Trace Elem Res. 2016;169(1):69-76. [CrossRef]
- Asli M, Azizzadeh M, Moghaddamjafari A, et al. Copper, iron, manganese, zinc, cobalt, arsenic, cadmium, chrome, and lead concentrations in liver and muscle in Iranian camel (Camelus dromedarius). Biol Trace Elem Res. 2020;194(2):390-400. [CrossRef]
- Chafik A, Essamadi A, Çelik SY, et al. Partial purification and some interesting properties of glutathione peroxidase from liver of camel (Camelus dromedarius). Russ J Bioorg Chem. 2018;44(1):41-51. [CrossRef]
- Maharem TM, Emam MA, Said YA. Purification and characterization of L-glutaminase enzyme from camel liver: enzymatic anticancer property. Int J Biol Macromol. 2020;150:1213-1222. [CrossRef]
- Maharem TM, Zahran WE, Hassan RE, et al. Unique properties of arginase purified from camel liver cytosol. Int J Biol Macromol. 2018;108:88-97. [CrossRef]
- Ponamarev V, Popova O, Kostrova A, et al. The concept of development of new ecologically based methods of diagnostics and pharmacocorrection in veterinary medicine (on the example of pathologies of the hepatobiliary system). In: II Int Conf Sustainable Development: Agriculture, Veterinary Medicine and Ecology (VMAEE-II-2023); April 21–22, 2023; Karshi, Russia. AIP Publishing; 2023:20028. [CrossRef]
- Ponamarev V, Yashin A, Prusakov A, et al. Influence of modern probiotics on morphological indicators of pigs’ blood in toxic dyspepsia. In: Agriculture Digitalization and Organic Production: Proc Second Int Conf; June 6–8, 2022; St. Petersburg, Russia. Springer; 2022:133-142. [CrossRef]
- Prusakova AV, Zelenevskiy NV, Prusakov AV, et al. Ultra-structural organization of the gallbladder mucous membrane of Anglo-Nubian goat. Vet Res Forum. 2024;15(3):165-169. [CrossRef]
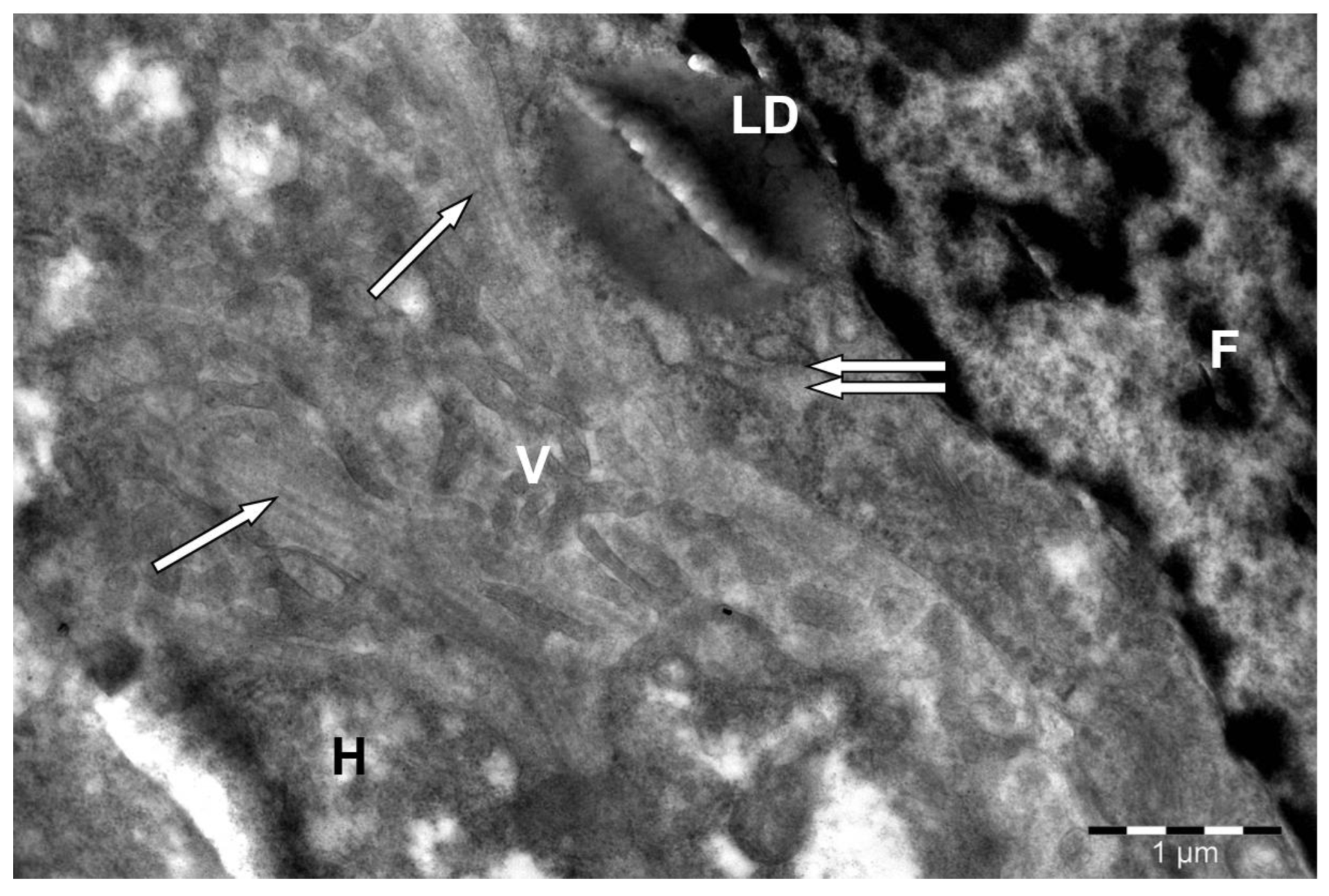
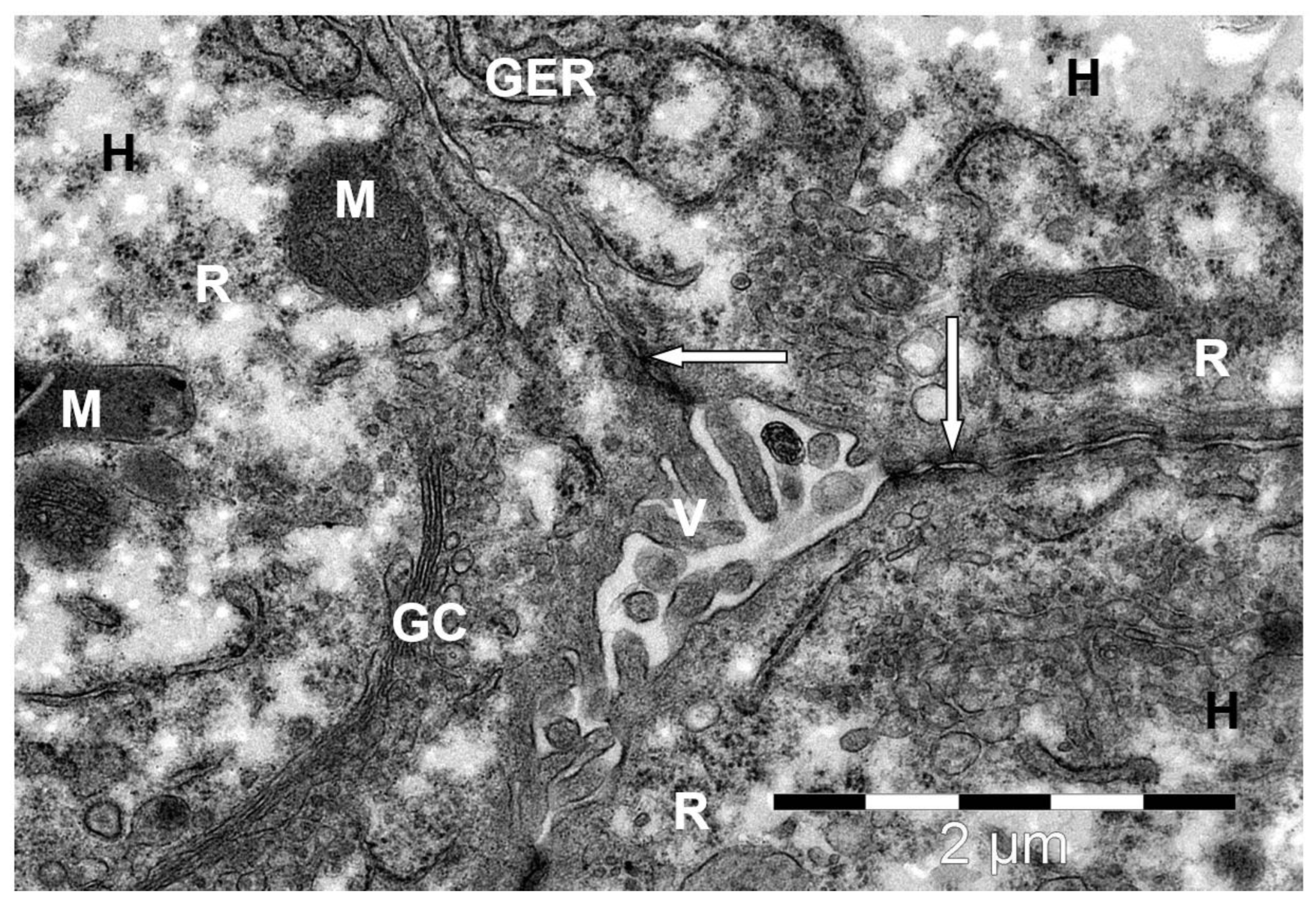
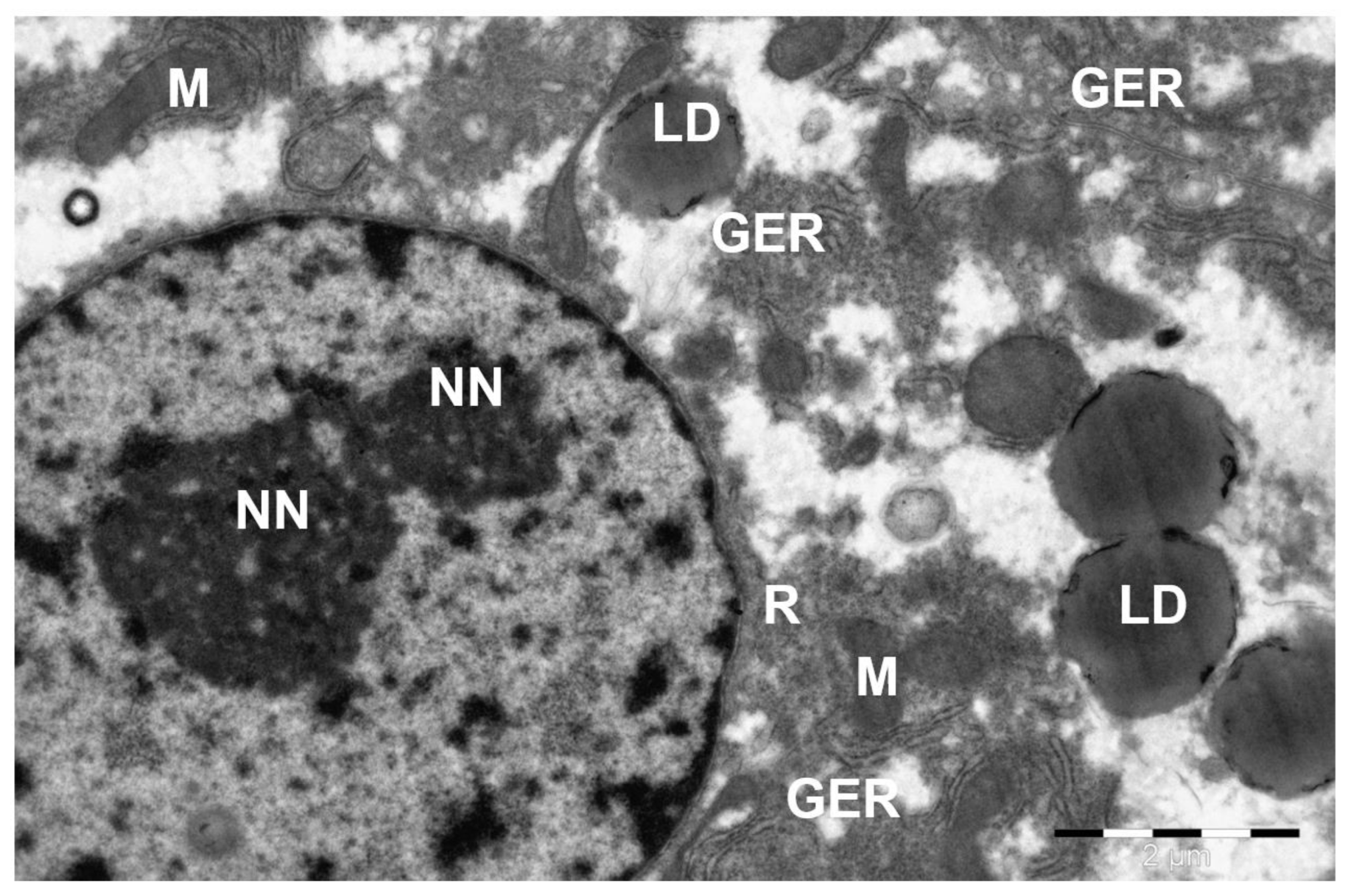
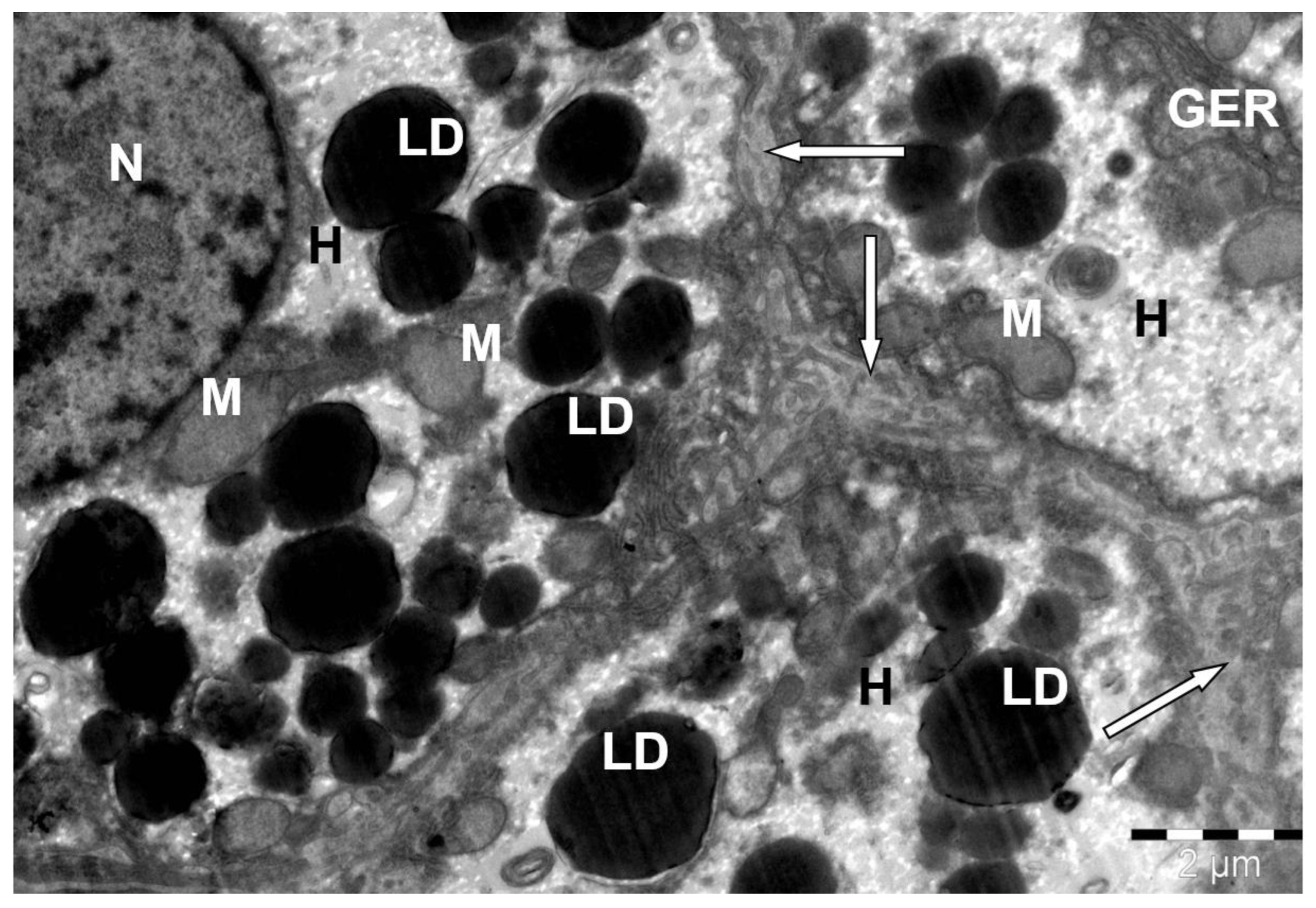
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



