Submitted:
08 August 2025
Posted:
11 August 2025
You are already at the latest version
Abstract
Polychlorinated biphenyls (PCBs), a class of persistent endocrine-disrupting chemicals, continue to pose significant risks to human and environmental health due to their widespread distribution, lipophilicity, and ability to bioaccumulate. Of particular concern is exposure during critical periods of neurodevelopment, which has been associated with long-lasting effects on sensory systems, including auditory processing. This review elaborates findings from both human epidemiological studies and animal models to examine the effects of PCBs on auditory function across the peripheral and central auditory pathways. Evidence indicates that PCB exposure during gestation and early postnatal life can lead to permanent auditory deficits. At the level of the auditory periphery, reductions in distortion-product otoacoustic emissions (DPOAEs) and auditory brainstem responses (ABRs) suggest cochlear outer hair cell dysfunction, potentially mediated by oxidative stress. Centrally, alterations in the primary auditory cortex and inferior colliculus have been observed, including disrupted tonotopic organization, excitation–inhibition imbalance, and impaired neuroplasticity. Notably, such central neural changes can occur out of proportion to abnormalities in ABRs, pointing to central auditory vulnerabilities. Epidemiological data in humans further support these findings, with certain PCB congeners linked to a significantly elevated risk of hearing impairment in children. PCB exposure coupled with loud noise exposure, which is growing in prevalence, leads to cumulative deficits in auditory processing. Together, these findings highlight the multi-level impact of PCB exposure on the auditory system and highlight the need for further investigation into the molecular and circuit-level mechanisms underlying these effects. Understanding the actions of PCB congeners on the auditory system in various human population cohorts is important to inform risk assessment and guide future public health policies.
Keywords:
auditory
; hearing
; PCB
; toxicology
; cochlea
Introduction:
Polychlorinated biphenyls (PCBs) are synthetic organic chlorinated compounds used in industries for decades until their ban in 1979. PCBs were originally developed in the late 1800’s, and began to be used more widely commercially in the 1930’s (Markowitz, 2018). Their beneficial industrial properties, such as low water solubility, low ability to conduct current, low flammability, and very high chemical stability, made them ideal components of electrical insulators and plasticizers. PCBs were found in a large proportion of electrical transformers across the U.S. and contributed to the expansion of the U.S. electrical grid. Because of their low flammability, they have been incorporated into many building materials, particularly in schools and were heavily used during school building booms during the 1960s (Thomas, 2012). Most schools in the U.S. were built during the period of heavy use of PCBs in building materials (Rowand, 1999).
Concerns about human toxicity of environmental exposures to PCBs emerged in the 1960s. Two major epidemics of PCB poisoning due to the consumption of contaminated rice-bran cooking oil have been documented. The first occurred in Japan in 1968, affecting approximately 1,700 individuals, and is known as the "Yusho" incident (Kuratsune et al., 1972). A second, similar outbreak occurred in Taiwan between 1978 and 1979, with over 2,000 reported cases, referred to as the "Yu-Cheng" (oil disease) incident (Chen et al., 1980). In addition, accumulating reports of PCBs acting as carcinogens, endocrine disrupters, and causing neurodevelopmental and reproductive disorders led the Environmental Protection Agency (EPA) to ban their production in 1979 under the Toxic Substances Control Act (TSCA). Unfortunately, given their very high chemical stability (half life in sediment of aquatic ecosystems can be many decades) PCBs persist in the environment and their high lipid solubility has made them difficult to contain leading to their continued presence in air, soil, water and continued incorporation into the food chain (Beyer and Biziuk, 2009; Hens and Hens, 2017).
209 individual chemical congeners of PCBs exist, each composed of two linked benzene rings with varying numbers and positions of chlorine atoms. These congeners are broadly classified based on their structural and toxicological properties into dioxin-like (DL) and non-dioxin-like (NDL) PCBs. Dioxin-like PCBs bind the aryl hydrocarbon receptor (AhR), thereby triggering a cascade of gene regulatory events that can lead to oxidative stress, immunotoxicity, and neurotoxicity (Jin et al., 2020; Shen et al., 2016; Zhang et al., 2012). Dioxin-like activity is typically associated with a coplanar configuration. In contrast, non-dioxin-like PCBs are in a noncoplanar conformation, reducing AhR binding. These two classes of PCBs likely have different mechanisms of toxicity, though many toxic exposures involve a blend of congeners, therefore involving a mixture of both classes of PCBs (Imamoglu and Christensen, 2002; Jursa et al., 2006).
A major route of human exposure to PCBs is via fish consumption as PCBs make their way into aquatic ecosystems. PCBs bio-accumulate, and larger predator fish often consumed by humans contain higher concentrations of PCBs (Porte and Albaigés, 1994). The fetus and neonate are especially susceptible to the toxic effects of these compounds due to exposure during critical windows of nervous system development. PCBs readily cross the placenta, resulting in fetal exposure during gestation, while PCBs are additionally transferred to the neonate through breastfeeding (Aliyu et al., 2010; Rogan et al., 1986a; Rogan et al., 1986b). Their lipid solubility leads them to reach the brain leading to an impact on nervous system development. A more recently-recognized form of exposure to PCBs is the airborne route. PCBs are often spontaneously produced as byproducts during the synthesis of pigments that are still in wide use in paints. Thus, in addition to legacy industrial PCBs that continue to persist in the environment, there is continued production and release of PCBs from certain organic pigments, and high levels continue to be found in schools (Herrick et al., 2011; Marek et al., 2017). Therefore, PCB exposure continues to be an ongoing health issue.
This review is focused on the effects of PCBs on hearing. We provide a comprehensive examination of how PCBs affect the auditory system, encompassing both the peripheral components (cochlea and auditory nerve) and central auditory pathways (brainstem and auditory cortex). We discuss the possible mechanisms of PCB ototoxicity, behavioral and functional outcomes of PCB exposure on hearing. We present key findings from animal models and human studies.
Evidence for neuro- and oto-toxicity of PCBs.
Developmental PCB exposure has been associated with numerous disorders of human nervous system development. For example, children prenatally exposed to PCBs during the Taiwan incident in 1978-1979 had lower full-scale IQ scores when tested between the ages of 9 and 10 (Chen and Hsu, 1994). Jacobson and Jacobson in 1996 reported lowered IQ scores in children exposed to PCBs via maternal consumption of contaminated fish (Jacobson and Jacobson, 1996). In addition, Patandin et al. in 1999 reported across-the-board cognitive impairments in children with in utero and lactational exposures to PCBs (Patandin et al., 1999). A review of the available literature in 2008 suggested that the cognitive domain most severely affected after prenatal PCB exposure was executive function (Boucher et al., 2009; Eubig et al., 2010). There is also evidence that PCB exposure is associated with the development of autistic-like (Lyall et al., 2017; Panesar et al., 2020) and attention-deficit-hyperactivity-like phenotypes (Aguiar et al., 2010; Sagiv et al., 2012). The preclinical literature also reflects broad cognitive impairment after PCB exposure, particularly in domains requiring the frontal cortex, reviewed in (Bullert et al., 2021). Thus, epidemiological and animal model data suggest that developmental exposure to PCBs negatively impacts both cognitive and behavioral development.
The auditory system appears to be particularly vulnerable to PCB exposures. For example, a landmark study from Trnovec et al. examined peripheral auditory function in over 400 children environmentally exposed to PCBs in Eastern Slovakia. PCB levels measured in the serum of 8 and 9-year-old children were associated with elevated hearing threshold, most consistently in the lower frequencies. Importantly, PCB exposure was also associated with diminishment in transient otoacoustic emissions, which are thought to be generated by outer hair cells in the cochlea, suggesting that outer hair cells may be a locus of pathology. In addition, there was no association with thyroid size or hormone production and PCB exposure in these subjects (Trnovec et al., 2008). Similarly, a later study demonstrated that postnatal exposure to PCBs was associated with diminished childhood distortion-product otoacoustic emissions (DPOAEs), which are generated by outer hair cells (Jusko et al., 2014). There is evidence from the U.S. that hearing dysfunction observed after developmental PCB exposure can persist into adulthood (Min et al., 2014). These studies point to persistent outer hair cell pathology as responsible for hearing loss after developmental PCB exposure. Note that outer hair cells are responsible for cochlear amplification, so it would be expected that their dysfunction would be associated with elevated hearing thresholds as well as diminished otoacoustic emissions. In addition, a study conducted in the Faroe Islands, a group of North Atlantic islands where high levels of PCB environmental contamination have been described, observed elevated hearing thresholds associated with PCB exposure. Interestingly, late-wave auditory brainstem abnormalities were also found (Grandjean et al., 2001), suggesting central auditory processing deficits, which is a topic to be addressed below.
Animal models have confirmed the human epidemiological findings that outer hair cell dysfunction likely plays a significant role in the loss of peripheral auditory function after developmental PCB exposure. Multiple studies in rats and mice have confirmed elevated auditory brainstem response thresholds after developmental PCB exposure, generally in the lower frequencies and more commonly in males (Crofton, K. et al., 2000; Crofton and Rice, 1999; Goldey et al., 1995; Ibrahim et al., 2023; Lee et al., 2021; Meerts et al., 2004; Powers et al., 2006; Sadowski et al., 2016). Similar to humans, DPOAEs are also impaired after PCB exposure (Lasky et al., 2002; Powers et al., 2006), also suggesting pathology involving outer hair cells. Loss of outer hair cells, and not inner hair cells or spiral ganglion neurons, was confirmed histologically by Crofton et al. (Crofton, K.M. et al., 2000), and this loss was seen in the low-frequency part of the cochlea, see Figure 1. Thus, there is broad consistency across human and rodent studies that indicate that developmental PCB exposure targets outer hair cells, primarily at the mid- and apical-regions of the basilar membrane (where lower frequencies are represented), leading to loss of cochlear amplification in these frequencies and thus loss of peripheral hearing.
Evidence for central auditory dysfunction after developmental PCB exposure:
Early studies in humans suggested that both central and peripheral auditory processing were impaired after developmental PCB exposure. Auditory event-related potentials, particularly the P300 component, have shown promise as non-invasive biomarkers for assessing cognitive function in children. For example, reductions in P300 amplitude have been linked to neurodevelopmental disorders such as attention-deficit disorder and reading disability (Holcomb et al., 1986), while prolonged P300 latency has been observed in children with cognitive impairments compared to typically developing peers (El-Shimy et al., 2023; Gupta et al., 2020; Tao et al., 2024).
In the context of environmental neurotoxicology, P300 has been utilized to evaluate the long-term cognitive effects of prenatal exposure to polychlorinated biphenyls (PCBs). For example, in an auditory oddball paradigm, prenatal exposure to PCBs was associated with lengthening of P300 latencies in children at the age of 9 (Vreugdenhil et al., 2004). Similar oddball responses were diminished in children who were PCB-exposed and had limited breastfeeding, suggesting that there may be interactions between PCB exposure and nutritional status (Boucher et al., 2010).
Animal studies have confirmed the presence of central auditory dysfunction after PCB exposure. Auditory brainstem responses, typically used to measure peripheral auditory function, can also provide a readout of central function when measuring the later waves. For example, Lilienthal et al. reported delays in wave IV in the auditory brainstem responses, primarily in males and at low sound frequencies (Lilienthal et al., 2011). Wave IV is thought to derive from the superior olivary complex and lateral lemniscus (Henry, 1979). Substantially more data exist regarding the effects of PCBs at the level of the auditory midbrain, or inferior colliculus. Rats treated with PCBs during development develop audiogenic seizures (Poon et al., 2015), which are epileptic seizures that are induced by loud sounds. These types of seizures are rare in humans, but when they occur, they are thought to be related to dysfunction at the level of the auditory midbrain (Faingold, 2002; Garcia-Cairasco, 2002; Ribak and Morin, 1995). Consistent with this idea, rats treated with PCBs were found to have a diminishment in the level of glutamic acid decarboxylase, the enzyme that produces the inhibitory neurotransmitter GABA, in the inferior colliculus (Bandara et al., 2016). In a recent study using multiphoton imaging to examine PCB effects on the inferior colliculus, exposed mice showed disruption of the normal tonotopic organization present on the surface of the inferior colliculus, showed altered excitatory/inhibitory balance across neurons in the inferior colliculus and evidence for elevated indicators of oxidative stress in the IC (Ibrahim et al., 2023).
Several studies have also examined PCB-related changes at the level of the auditory cortex. For example, Kenet et al. observed that rats treated with PCBs during early development displayed gross disorganization of the tonotopic map of the primary auditory cortex. In addition, similar to what was seen in the inferior colliculus, there was a disruption of the excitatory/inhibitory balance in the auditory cortex, particularly for sound frequencies that were distant from a neuron’s best frequency, altering its tuning function. Finally, there was an inability for maps to demonstrate normal experience-dependent plasticity, consistent with deficits in learning and memory seen in human studies (Kenet et al., 2007). Additional studies in rats have confirmed alterations of excitatory/inhibitory balance in the auditory cortex of PCB-treated rats (Lee et al., 2021; Sadowski et al., 2016). For example, developmental PCB exposure in rats led to an increased sensitivity to GABAergic antagonism in flavoprotein autofluoresence-based assay of auditory cortex in brain slices (Figure 2). It is important to note that peripheral hearing loss on its own can induce significant reorganization of central circuits, in particular declines in inhibitory neurotransmission (Ibrahim and Llano, 2019; Milbrandt et al., 2000). Therefore, it is important to note that the changes in inhibition observed in the auditory cortex do not appear to be correlated with peripheral hearing loss (Lee et al., 2021). These findings suggest that changes seen in the auditory cortex are not simply a consequence of loss of peripheral hearing sensitivity, but rather are likely related to effects occurring at the level of the central auditory system.
Potential mechanisms of PCB effects on the developing auditory system
Multiple mechanisms have been proposed to explain the effects of PCBs on the auditory system. PCBs are hydrophobic and typically act on intracellular targets. Many of the PCBs, particularly those with dioxin-like structures, will bind to the aryl hydrocarbon receptor, an intracellular receptor which acts as a transcription factor, and thus can modify the expression of a large number of genes. PCBs also structurally resemble thyroid hormone and can thus interfere with the production of thyroid hormone at several steps of thyroid hormone synthesis, as well as increasing hepatic clearance of thyroid hormones. Deficiencies in thyroid hormone have been documented after developmental PCB exposure (Crofton, K. et al., 2000; Little et al., 2022; Meerts et al., 2004; Mohammadparast-Tabas et al., 2024) and thyroid hormone is needed for normal development of the brain and cochlea (Bernal, 2007; Farahvar et al., 2007; Forrest et al., 1996; Knipper et al., 2000; Ng et al., 2013). Replacement of thyroid hormone can partially mitigate the impact of prenatal PCB exposure on peripheral auditory function (Goldey and Crofton, 1998), suggesting that thyroid dysfunction, as well as additional mechanisms, may be involved. No work has been done to our knowledge to determine if replacement of thyroid hormone can mitigate the central auditory system effects of PCB exposure. The aryl hydrocarbon receptor has been described throughout the brain (Kimura and Tohyama, 2017; Petersen et al., 2000), suggesting that direct effects outside of thyroid hormone abnormalities may be responsible for negative developmental effects.
PCBs, particularly non-dioxin-like forms, also bind to the ryanodine receptor (RyR), which is an intracellular calcium channel in the endoplasmic reticulum membrane. PCBs stabilize the RyR in the open state, leading to increases in cytosolic calcium levels. These increases in intracellular calcium can trigger a series of secondary events that influence neuronal function, including inducing abnormal structural plasticity in some classes of neurons while leading to excitotoxicity in others (Howard et al., 2003; Roegge et al., 2006). RyRs are also present in the cochlea, with evidence of their expression in supporting cells, spiral ganglion cells, inner hair cells and outer hair cells, including during development (Liang et al., 2009; Morton-Jones et al., 2006), though outer hair cell expression was higher than other cell types (Grant et al., 2006), which may explain the predilection of PCBs to impact outer hair cell development described above.
PCBs may also impact the auditory system through their effects on the developing vascular system. Multiple studies have revealed PCB-related pathology on the vasculature (Bell, 1983; Das and Felty, 2014; Selvakumar et al., 2013) and recent findings suggest that developmental PCB expression may lead to diminished vascularity of the mouse inferior colliculus (N et al., 2024). Aryl hydrocarbon receptors are found at high levels on endothelial and other vascular cells (Guerra-Ojeda et al., 2023). Activation of these receptors leads to oxidative stress and this increase in reactive oxygen metabolites may deplete nitric oxide or angiotensin II responsiveness, leading to abnormal vasoconstriction (Agbor et al., 2011; Nakagawa et al., 2021). PCBs can also induce inflammatory responses in the vascular system and subsequent upregulation of cell adhesion molecules (Choi et al., 2003; Petriello et al., 2018; Zhang et al., 2015). Indeed, PCB exposure has been linked to chronic cardiovascular disease, in part related to their direct effects on the vasculature (Perkins, J. T. et al., 2016; Perkins, Jordan T et al., 2016).
In addition to the mechanisms outlined above, other mechanisms of PCB-induced toxicity of the cochlea and brain have been described. PCBs alter levels of several key neuromodulators in the brain, such as dopamine and acetylcholine (Choksi et al., 1997; Donahue et al., 2004), both of which have substantial effects on neural circuit development. PCBs are endocrine-disruptors and therefore may alter levels of circulating sex steroids (Dickerson et al., 2011; Lilienthal et al., 2006), which also have significant effects on the developing brain and cochlea (Peper and Koolschijn, 2012). PCBs may damage the placenta (Bäcklin et al., 1998; Tsuji et al., 2013), leading to metabolic dysregulation of the fetus. It is likely that multiple mechanisms converge to lead to the auditory (and other) toxic effects of PCBs.
The relative roles of dioxin-like vs. non-dioxin-lik PCBs in auditory toxicity is not yet known. Several studies have observed effects on the peripheral and central auditory systems after exposure to non-dioxin-like PCBs (Kenet et al., 2007; Lilienthal et al., 2011) or non-dioxin-like PCBs (Crofton and Rice, 1999). However, most other studies that assessed ototoxicity used complex mixtures of PCBs. Thus, it is likely that several mechanisms may contribute to the effects of PCBs on the auditory system. Future work will clarify the individual contributions of different types of PCB exposures.
PCB Interactions with other ototoxicants
Developmental PCB exposure can lead to lifelong elevation in hearing thresholds as well as pathological alterations in central processing of acoustic stimuli. PCBs also often co-exist in the environment along with other toxicants and outside of the auditory system can lead to cumulative pathology when combined with other environmental toxicants (Pellacani et al., 2014; Roegge et al., 2004; Smith et al., 1995). A key question is whether developmental PCB exposure increases the vulnerability of the auditory system to other ototoxicants. It is possible that by diminishing the number of outer hair cells, early PCB exposure may limit the capacity for auditory "reserve" that would normally permit functional resistance to later ototoxicant exposure. Many common ototoxicants, such as iatrogenic ones (aminglycosides, platinum-containing chemotherapeutics, certain diuretics), heavy metal exposure and noise exposure, lead to elevations in oxidative stress at the level of the cochlea (Desa et al., 2018; Henderson et al., 2006; Ramkumar et al., 2021; Roth and Salvi, 2016). PCB exposure also leads to an elevation in oxidative stress (Liu et al., 2020). Given the limited capacity of outer hair cells to combat oxidative stress (Baker and Staecker, 2012; Qi et al., 2025), depletion of normal compensatory mechanisms may increase the vulnerability of remaining hair cells.
Our group recently observed that developmental PCB exposure significantly worsens the auditory outcomes after a later noise exposure occurring during adulthood (Ibrahim et al., 2023). In this study, mice were exposed in utero to a PCB mixture and then, as adults, subjected to acute acoustic trauma for 45 minutes, designed to cause a reversible acoustic threshold shift. The findings revealed a nonlinear interaction: the PCB-exposed mice failed to recover their hearing after the noise exposure, whereas vehicle-treated (non-PCB) mice showed partial recovery over the following days. In other words, prenatal PCB exposure created a long-lasting vulnerability such that a noise-induced hearing loss became permanent when it would have been temporary in the absence of PCB exposure. A key additional finding was that mice with poorer endogenous antioxidant capacity (glutathione-related mechanisms) experienced the greatest hearing damage and central changes, implicating oxidative stress as a major driver. This finding aligns with broader observations that enhancement of antioxidant defenses can hasten recovery from noise injury (Kopke et al., 2005; Pisani et al., 2023), and any factor that weakens those defenses (such as PCB-induced chronic oxidative stress or reduced glutathione levels) can convert what would be a temporary insult into a permanent one.
A limited number of studies have examined the impact of PCBs in combination with other environmental toxicants on the auditory system. For example, exposure to PCBs in combination with polybrominated diphenyl ethers produced cumulative pathological effects on cochlear function (Poon et al., 2011). In contrast, PCB effects in the auditory system may be partially attenuated by co-exposure to methylmercury, though the mechanism for this interaction has not yet been established (Powers et al., 2009). Given the high rates of combined exposure to PCBs with other potential ototoxicants, more work is needed in this area to clarify the nature of the interactions.
Conclusions and future directions:
PCBs exert toxic effects on both peripheral and central components of the auditory system, most likely through multiple mechanisms. Although banned decades ago, PCBs persist in the environment due to their chemical stability, presence in air, soil, water, and bioaccumulation in the food chain. Human exposure remains ongoing, especially among vulnerable populations such as those living near contaminated sites or consuming tainted fish. Emerging evidence reveals that certain PCB congeners continue to form spontaneously from modern products like paints and sealants, indicating that PCB exposure is not merely historical but also contemporary and evolving.
Developmental PCB exposure impairs outer hair cell function and disrupts auditory brainstem circuit maturation, resulting in long-lasting deficits. Central nervous system studies show that PCBs alter excitatory-inhibitory balance and reduce synaptic plasticity in key auditory centers like the inferior colliculus and cortex. Furthermore, combined exposures (e.g., with noise or ototoxic drugs) suggest that PCBs may sensitize the auditory system, leading to disproportionate deficits after dual insults. These effects are troubling given the importance of auditory processing for language, learning, and cognitive function, where even small deficits in auditory function can lead to substantial educational delays (Sharma et al., 2023). Mechanistically, PCB-induced auditory dysfunction likely involves endocrine disruption, oxidative stress, vascular impairment, inflammation, and neurotransmitter imbalances. These same pathways are implicated in cognitive decline and neurodegeneration, positioning PCBs as potential environmental contributors to diseases like Alzheimer’s Disease.
Future investigations should focus on identifying the molecular underpinnings of PCB ototoxicity, including roles for thyroid hormone signaling, oxidative damage, RyR and aryl hydrocarbon receptor activation. Longitudinal studies using prenatal and early postnatal exposure models can clarify critical windows of vulnerability and help determine whether effects are permanent or reversible. Equally important is understanding how PCB exposure interacts with other ototoxicants, such as aminoglycosides, solvents, or excessive noise, to produce additive or synergistic auditory damage. These interactions may better reflect real-world exposure scenarios.
Therapeutic strategies deserve attention as well. Interventions such as antioxidant therapy, thyroid hormone supplementation, RyR or aryl hydrocarbon receptor antagonists may offer protective or restorative benefits. Additionally, research should examine whether auditory cortex reorganization following PCB exposure is a direct neurotoxic effect or secondary to reduced cochlear input. Finally, genetic and sex-based factors—including polymorphisms in detoxification enzymes or hormone receptors—may modulate susceptibility to PCB-induced hearing loss and should be systematically evaluated. In sum, PCBs pose an ongoing threat to auditory health. By deepening our understanding of their mechanisms of action and identifying protective strategies, we can develop effective public health interventions and policies to safeguard auditory and cognitive function across the lifespan.
References
- Agbor, L.N.; Elased, K.M.; Walker, M.K. Endothelial cell-specific aryl hydrocarbon receptor knockout mice exhibit hypotension mediated, in part, by an attenuated angiotensin II responsiveness. Biochemical pharmacology 2011, 82, 514–523. [Google Scholar] [CrossRef]
- Aguiar, A.; Eubig, P.A.; Schantz, S.L. Attention deficit/hyperactivity disorder: a focused overview for children’s environmental health researchers. Environmental health perspectives 2010, 118, 1646–1653. [Google Scholar] [CrossRef] [PubMed]
- Aliyu, M.H.; Alio, A.P.; Salihu, H.M. To breastfeed or not to breastfeed: a review of the impact of lactational exposure to polychlorinated biphenyls (PCBs) on infants. Journal of Environmental Health 2010, 73, 8–15. [Google Scholar]
- Bäcklin, B.M.; Persson, E.; Jones, C.J.; Dantzer, V. Polychlorinated biphenyl (PCB) exposure produces placental vascular and trophoblastic lesions in the mink (Mustela vison): a light and electron microscopic study. Apmis 1998, 106, 785–799. [Google Scholar] [CrossRef]
- Baker, K.; Staecker, H. Low Dose Oxidative Stress Induces Mitochondrial Damage in Hair Cells. The Anatomical Record 2012, 295, 1868–1876. [Google Scholar] [CrossRef]
- Bandara, S.B.; Eubig, P.A.; Sadowski, R.N.; Schantz, S.L. Developmental PCB exposure increases audiogenic seizures and decreases glutamic acid decarboxylase in the inferior colliculus. Toxicological Sciences 2016, 149, 335–345. [Google Scholar] [CrossRef]
- Bell, M. Ultrastructural features of the murine cutaneous microvasculature after exposure to polychlorinated biphenyl compounds (PCBs) and benzo-(a)-pyrene (BAP). Virchows Archiv B Cell Pathology Including Molecular Pathology 1983, 42, 131–142. [Google Scholar] [CrossRef]
- Bernal, J. Thyroid hormone receptors in brain development and function. Nature clinical practice Endocrinology & metabolism 2007, 3, 249–259. [Google Scholar]
- Beyer, A.; Biziuk, M. Environmental fate and global distribution of polychlorinated biphenyls. Reviews of Environmental Contamination and Toxicology Vol 2009, 201, 137–158. [Google Scholar]
- Boucher, O.; Bastien, C.H.; Saint-Amour, D.; Dewailly, É.; Ayotte, P.; Jacobson, J.L.; Jacobson, S.W.; Muckle, G. Prenatal exposure to methylmercury and PCBs affects distinct stages of information processing: an event-related potential study with Inuit children. Neurotoxicology 2010, 31, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Boucher, O.; Muckle, G.; Bastien, C.H. Prenatal exposure to polychlorinated biphenyls: a neuropsychologic analysis. Environmental health perspectives 2009, 117, 7–16. [Google Scholar] [CrossRef]
- Bullert, A.J.; Doorn, J.A.; Stevens, H.E.; Lehmler, H.-J. The effects of polychlorinated biphenyl exposure during adolescence on the nervous system: A comprehensive review. Chemical research in toxicology 2021, 34, 1948–1952. [Google Scholar] [CrossRef]
- Chen, P.; Gaw, J.; Wong, C.; Chen, C. Levels and gas chromatographic patterns of polychlorinated biphenyls in the blood of patients after PCB poisoning in Taiwan. Bull. Environ. Contam. Toxicol.;(United States), 1980; 25. [Google Scholar]
- Chen, Y.J.; Hsu, C.C. EFFECTS OF PRENATAL EXPOSURE TO PCBs ON THE NEUROLOGICAL FUNCTION OF CHILDREN: A NEUROPSYCHOLOGICAL AND NEUROPHYSIOLOGY STUDY. Developmental Medicine & Child Neurology 1994, 36, 312–320. [Google Scholar]
- Choi, W.; Eum, S.Y.; Lee, Y.W.; Hennig, B.; Robertson, L.W.; Toborek, M. PCB 104-induced proinflammatory reactions in human vascular endothelial cells: relationship to cancer metastasis and atherogenesis. Toxicological Sciences 2003, 75, 47–56. [Google Scholar] [CrossRef]
- Choksi, N.Y.; Kodavanti, P.R.S.; Tilson, H.A.; Booth, R.G. Effects of polychlorinated biphenyls (PCBs) on brain tyrosine hydroxylase activity and dopamine synthesis in rats. Fundamental and Applied Toxicology 1997, 39, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Crofton, K.; Kodavanti, P.; Derr-Yellin, E.; Casey, A.; Kehn, L. PCBs, thyroid hormones, and ototoxicity in rats: cross-fostering experiments demonstrate the impact of postnatal lactation exposure. Toxicological Sciences 2000, 57, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Crofton, K.M.; Ding, D.-L.; Padich, R.; Taylor, M.; Henderson, D. Hearing loss following exposure during development to polychlorinated biphenyls: a cochlear site of action. Hearing research 2000, 144, 196–204. [Google Scholar] [CrossRef]
- Crofton, K.M.; Rice, D.C. Low-frequency hearing loss following perinatal exposure to 3, 3′, 4, 4′, 5-pentachlorobiphenyl (PCB 126) in rats. Neurotoxicology and teratology 1999, 21, 299–301. [Google Scholar] [CrossRef]
- Das, J.K.; Felty, Q. PCB153-induced overexpression of ID3 contributes to the development of microvascular lesions. PLoS One 2014, 9, e104159. [Google Scholar] [CrossRef] [PubMed]
- Desa, D.E.; Nichols, M.G.; Smith, H.J. Aminoglycosides rapidly inhibit NAD(P)H metabolism increasing reactive oxygen species and cochlear cell demise. J Biomed Opt 2018, 24, 1–14. [Google Scholar] [CrossRef]
- Dickerson, S.M.; Cunningham, S.L.; Patisaul, H.B.; Woller, M.J.; Gore, A.C. Endocrine disruption of brain sexual differentiation by developmental PCB exposure. Endocrinology 2011, 152, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Donahue, D.A.; Dougherty, E.J.; Meserve, L.A. Influence of a combination of two tetrachlorobiphenyl congeners (PCB 47; PCB 77) on thyroid status, choline acetyltransferase (ChAT) activity, and short-and long-term memory in 30-day-old Sprague–Dawley rats. Toxicology 2004, 203, 99–107. [Google Scholar] [CrossRef]
- El-Shimy, A.A.; Nada, E.; Quriba, A.; Galhoum, D. Assessment of Higher Cognitive Function Using p300 in Children with Specific Language Impairment: Data from Zagazig University Hospital. Zagazig University Medical Journal 2023, 29, 1456–1462. [Google Scholar] [CrossRef]
- Eubig, P.A.; Aguiar, A.; Schantz, S.L. Lead and PCBs as risk factors for attention deficit/hyperactivity disorder. Environmental health perspectives 2010, 118, 1654–1667. [Google Scholar] [CrossRef]
- Faingold, C.L. Role of GABA abnormalities in the inferior colliculus pathophysiology–audiogenic seizures. Hearing research 2002, 168, 223–237. [Google Scholar] [CrossRef]
- Farahvar, A.; Darwish, N.H.; Sladek, S.; Meisami, E. Marked recovery of functional metabolic activity and laminar volumes in the rat hippocampus and dentate gyrus following postnatal hypothyroid growth retardation: a quantitative cytochrome oxidase study. Experimental neurology 2007, 204, 556–568. [Google Scholar] [CrossRef]
- Forrest, D.; Erway, L.C.; Ng, L.; Altschuler, R.; Curran, T. Thyroid hormone receptor β is essential for development of auditory function. Nature genetics 1996, 13, 354–357. [Google Scholar] [CrossRef]
- Garcia-Cairasco, N. A critical review on the participation of inferior colliculus in acoustic-motor and acoustic-limbic networks involved in the expression of acute and kindled audiogenic seizures. Hearing Research 2002, 168, 208–222. [Google Scholar] [CrossRef]
- Goldey, E.S.; Crofton, K.M. Thyroxine replacement attenuates hypothyroxinemia, hearing loss, and motor deficits following developmental exposure to Aroclor 1254 in rats. Toxicological Sciences 1998, 45, 94–105. [Google Scholar] [PubMed]
- Goldey, E.S.; Kehn, L.S.; Lau, C.; Rehnberg, G.L.; Crofton, K.M. Developmental exposure to polychlorinated biphenyls (Aroclor 1254) reduces circulating thyroid hormone concentrations and causes hearing deficits in rats. Toxicology and applied pharmacology 1995, 135, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, P.; Weihe, P.; Burse, V.W.; Needham, L.L.; Storr-Hansen, E.; Heinzow, B.; Debes, F.; Murata, K.; Simonsen, H.; Ellefsen, P. Neurobehavioral deficits associated with PCB in 7-year-old children prenatally exposed to seafood neurotoxicants. Neurotoxicology and teratology 2001, 23, 305–317. [Google Scholar] [CrossRef]
- Grant, L.; Slapnick, S.; Kennedy, H.; Hackney, C. Ryanodine receptor localisation in the mammalian cochlea: an ultrastructural study. Hearing research 2006, 219, 101–109. [Google Scholar] [CrossRef]
- Guerra-Ojeda, S.; Suarez, A.; Valls, A.; Verdu, D.; Pereda, J.; Ortiz-Zapater, E.; Carretero, J.; Mauricio, M.D.; Serna, E. The role of aryl hydrocarbon receptor in the endothelium: a systematic review. International Journal of Molecular Sciences 2023, 24, 13537. [Google Scholar] [CrossRef]
- Gupta, S.; Prasad, A.; Singh, R.; Gupta, G. Auditory and visual P300 responses in early cognitive assessment of children and adolescents with epilepsy. Journal of pediatric neurosciences 2020, 15, 9–14. [Google Scholar] [CrossRef]
- Henderson, D.; Bielefeld, E.C.; Harris, K.C.; Hu, B.H. The role of oxidative stress in noise-induced hearing loss. Ear and hearing 2006, 27, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Henry, K.R. Auditory brainstem volume-conducted responses: origins in the laboratory mouse. Ear and Hearing 1979, 4, 173–178. [Google Scholar]
- Hens, B.; Hens, L. Persistent Threats by Persistent Pollutants: Chemical Nature, Concerns and Future Policy Regarding PCBs-What Are We Heading For? Toxics 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Herrick, R.F.; Meeker, J.D.; Altshul, L. Serum PCB levels and congener profiles among teachers in PCB-containing schools: a pilot study. Environmental Health 2011, 10, 1–10. [Google Scholar] [CrossRef]
- Holcomb, P.J.; Ackerman, P.T.; Dykman, R.A. Auditory event-related potentials in attention and reading disabled boys. International Journal of Psychophysiology 1986, 3, 263–273. [Google Scholar] [CrossRef]
- Howard, A.S.; Fitzpatrick, R.; Pessah, I.; Kostyniak, P.; Lein, P.J. Polychlorinated biphenyls induce caspase-dependent cell death in cultured embryonic rat hippocampal but not cortical neurons via activation of the ryanodine receptor. Toxicology and applied pharmacology 2003, 190, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, B.A.; Llano, D.A. Aging and central auditory disinhibition: is it a reflection of homeostatic downregulation or metabolic vulnerability? Brain Sciences 2019, 9, 351. [Google Scholar] [CrossRef]
- Ibrahim, B.A.; Louie, J.J.; Shinagawa, Y.; Xiao, G.; Asilador, A.R.; Sable, H.J.; Schantz, S.L.; Llano, D.A. Developmental exposure to polychlorinated biphenyls prevents recovery from noise-induced hearing loss and disrupts the functional organization of the inferior colliculus. Journal of Neuroscience 2023, 43, 4580–4597. [Google Scholar] [CrossRef]
- Imamoglu, I.; Christensen, E.R. PCB sources, transformations, and contributions in recent Fox River, Wisconsin sediments determined from receptor modeling. Water Research 2002, 36, 3449–3462. [Google Scholar] [CrossRef]
- Jacobson, J.L.; Jacobson, S.W. Intellectual impairment in children exposed to polychlorinated biphenyls in utero. New England Journal of Medicine 1996, 335, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Wahlang, B.; Shi, H.; Hardesty, J.E.; Falkner, K.C.; Head, K.Z.; Srivastava, S.; Merchant, M.L.; Rai, S.N.; Cave, M.C.; Prough, R.A. Dioxin-like and non-dioxin-like PCBs differentially regulate the hepatic proteome and modify diet-induced nonalcoholic fatty liver disease severity. Med Chem Res 2020, 29, 1247–1263. [Google Scholar] [CrossRef] [PubMed]
- Jursa, S.; Chovancová; J; Petrík, J. ; Lokša, J. Dioxin-like and non-dioxin-like PCBs in human serum of Slovak population. Chemosphere 2006, 64, 686–691. [Google Scholar] [CrossRef]
- Jusko, T.A.; Sisto, R.; Iosif, A.-M.; Moleti, A.; Wimmerová, S; Lancz, K. ; Tihányi, J.; Šovčiková, E; Drobná, B; Palkovičová, L. Prenatal and postnatal serum PCB concentrations and cochlear function in children at 45 months of age. Environmental health perspectives 2014, 122, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Kenet, T.; Froemke, R.C.; Schreiner, C.E.; Pessah, I.N.; Merzenich, M.M. Perinatal exposure to a noncoplanar polychlorinated biphenyl alters tonotopy, receptive fields, and plasticity in rat primary auditory cortex. Proc Natl Acad Sci U S A 2007, 104, 7646–7651. [Google Scholar] [CrossRef]
- Kimura, E.; Tohyama, C. Embryonic and postnatal expression of aryl hydrocarbon receptor mRNA in mouse brain. Frontiers in neuroanatomy 2017, 11, 4. [Google Scholar] [CrossRef]
- Knipper, M.; Zinn, C.; Maier, H.; Praetorius, M.; Rohbock, K.; Köpschall, I.; Zimmermann, U. Thyroid hormone deficiency before the onset of hearing causes irreversible damage to peripheral and central auditory systems. Journal of Neurophysiology 2000, 83, 3101–3112. [Google Scholar] [CrossRef]
- Kopke, R.; Bielefeld, E.; Liu, J.; Zheng, J.; Jackson, R.; Henderson, D.; Coleman, J.K. Prevention of impulse noise-induced hearing loss with antioxidants. Acta oto-laryngologica 2005, 125, 235–243. [Google Scholar] [CrossRef]
- Kuratsune, M.; Yoshimura, T.; Matsuzaka, J.; Yamaguchi, A. Epidemiologic study on Yusho, a poisoning caused by ingestion of rice oil contaminated with a commercial brand of polychlorinated biphenyls. Environmental Health Perspectives 1972, 1, 119–128. [Google Scholar] [CrossRef]
- Lasky, R.E.; Widholm, J.J.; Crofton, K.M.; Schantz, S.L. Perinatal exposure to Aroclor 1254 impairs distortion product otoacoustic emissions (DPOAEs) in rats. Toxicological sciences 2002, 68, 458–464. [Google Scholar] [CrossRef]
- Lee, C.M.; Sadowski, R.N.; Schantz, S.L.; Llano, D.A. Developmental PCB exposure disrupts synaptic transmission and connectivity in the rat auditory cortex, independent of its effects on peripheral hearing threshold. Eneuro 2021, 8. [Google Scholar] [CrossRef]
- Liang, Y.; Huang, L.; Yang, J. Differential expression of ryanodine receptor in the developing rat cochlea. European Journal of Histochemistry: EJH 2009, 53, e30. [Google Scholar] [CrossRef]
- Lilienthal, H.; Hack, A.; Roth-Härer, A.; Grande, S.W.; Talsness, C.E. Effects of developmental exposure to 2, 2′, 4, 4′, 5-pentabromodiphenyl ether (PBDE-99) on sex steroids, sexual development, and sexually dimorphic behavior in rats. Environmental health perspectives 2006, 114, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Lilienthal, H.; Heikkinen, P.; Andersson, P.L.; van der Ven, L.T.; Viluksela, M. Auditory effects of developmental exposure to purity-controlled polychlorinated biphenyls (PCB52 and PCB180) in rats. Toxicological Sciences 2011, 122, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Little, C.C.; Barlow, J.; Alsen, M.; van Gerwen, M. Association between polychlorinated biphenyl exposure and thyroid hormones: a systematic review and meta-analysis. J Environ Sci Health C Toxicol Carcinog 2022, 40, 248–267. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tan, Y.; Song, E.; Song, Y. A critical review of polychlorinated biphenyls metabolism, metabolites, and their correlation with oxidative stress. Chemical research in toxicology 2020, 33, 2022–2042. [Google Scholar] [CrossRef] [PubMed]
- Lyall, K.; Croen, L.A.; Sjödin, A.; Yoshida, C.K.; Zerbo, O.; Kharrazi, M.; Windham, G.C. Polychlorinated biphenyl and organochlorine pesticide concentrations in maternal mid-pregnancy serum samples: association with autism spectrum disorder and intellectual disability. Environmental health perspectives 2017, 125, 474–480. [Google Scholar] [CrossRef]
- Marek, R.F.; Thorne, P.S.; Herkert, N.J.; Awad, A.M.; Hornbuckle, K.C. Airborne PCBs and OH-PCBs inside and outside urban and rural US schools. Environmental science & technology 2017, 51, 7853–7860. [Google Scholar]
- Markowitz, G. From Industrial Toxins to Worldwide Pollutants: A Brief History of Polychlorinated Biphenyls. Public Health Rep 2018, 133, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Meerts, I.A.; Lilienthal, H.; Hoving, S.; Van Den Berg, J.H.; Weijers, B.M.; Bergman, Å.; Koeman, J.H.; Brouwer, A. Developmental exposure to 4-hydroxy-2, 3, 3′, 4′, 5-pentachlorobiphenyl (4-OH-CB107): long-term effects on brain development, behavior, and brain stem auditory evoked potentials in rats. Toxicological sciences 2004, 82, 207–218. [Google Scholar] [CrossRef]
- Milbrandt, J.; Holder, T.; Wilson, M.; Salvi, R.; Caspary, D. GAD levels and muscimol binding in rat inferior colliculus following acoustic trauma. Hearing research 2000, 147, 251–260. [Google Scholar] [CrossRef]
- Min, J.-Y.; Kim, R.; Min, K.-B. Serum polychlorinated biphenyls concentrations and hearing impairment in adults. Chemosphere 2014, 102, 6–11. [Google Scholar] [CrossRef]
- Mohammadparast-Tabas, P.; Arab-Zozani, M.; Naseri, K.; Darroudi, M.; Aramjoo, H.; Ahmadian, H.; Ashrafipour, M.; Farkhondeh, T.; Samarghandian, S. Polychlorinated biphenyls and thyroid function: a scoping review. Rev Environ Health 2024, 39, 679–706. [Google Scholar] [CrossRef]
- Morton-Jones, R.; Cannell, M.; Jeyakumar, L.; Fleischer, S.; Housley, G. Differential expression of ryanodine receptors in the rat cochlea. Neuroscience 2006, 137, 275–286. [Google Scholar] [CrossRef] [PubMed]
- N, V.C.S. , M, L., P, S. Impact of combined developmental PCB and noise exposure on the cerebral microvasculature in mouse. 2024.
- Nakagawa, K.; Itoya, M.; Takemoto, N.; Matsuura, Y.; Tawa, M.; Matsumura, Y.; Ohkita, M. Indoxyl sulfate induces ROS production via the aryl hydrocarbon receptor-NADPH oxidase pathway and inactivates NO in vascular tissues. Life Sciences 2021, 265, 118807. [Google Scholar] [CrossRef]
- Ng, L.; Kelley, M.W.; Forrest, D. Making sense with thyroid hormone—the role of T3 in auditory development. Nature Reviews Endocrinology 2013, 9, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Panesar, H.K.; Kennedy, C.L.; Keil Stietz, K.P.; Lein, P.J. Polychlorinated biphenyls (PCBs): risk factors for autism spectrum disorder? Toxics 2020, 8, 70. [Google Scholar] [CrossRef] [PubMed]
- Patandin, S.; Lanting, C.I.; Mulder, P.G.H.; Boersma, E.R.; Sauer, P.J.J.; Weisglas-Kuperus, N. Effects of environmental exposure to polychlorinated biphenyls and dioxins on cognitive abilities in Dutch children at 42 months of age. The Journal of Pediatrics 1999, 134, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Pellacani, C.; Tagliaferri, S.; Caglieri, A.; Goldoni, M.; Giordano, G.; Mutti, A.; Costa, L.G. Synergistic interactions between PBDEs and PCBs in human neuroblastoma cells. Environmental Toxicology 2014, 29, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Peper, J.S.; Koolschijn, P.C.M. Sex steroids and the organization of the human brain. Journal of Neuroscience 2012, 32, 6745–6746. [Google Scholar] [CrossRef]
- Perkins, J.T.; Petriello, M.C.; Newsome, B.J.; Hennig, B. Polychlorinated biphenyls and links to cardiovascular disease. Environ Sci Pollut Res Int 2016, 23, 2160–2172. [Google Scholar] [CrossRef]
- Perkins, J.T.; Petriello, M.C.; Newsome, B.J.; Hennig, B. Polychlorinated biphenyls and links to cardiovascular disease. Environmental Science and Pollution Research 2016, 23, 2160–2172. [Google Scholar] [CrossRef]
- Petersen, S.L.; Curran, M.A.; Marconi, S.A.; Carpenter, C.D.; Lubbers, L.S.; McAbee, M.D. Distribution of mRNAs encoding the arylhydrocarbon receptor, arylhydrocarbon receptor nuclear translocator, and arylhydrocarbon receptor nuclear translocator-2 in the rat brain and brainstem. Journal of Comparative Neurology 2000, 427, 428–439. [Google Scholar] [CrossRef]
- Petriello, M.C.; Brandon, J.A.; Hoffman, J.; Wang, C.; Tripathi, H.; Abdel-Latif, A.; Ye, X.; Li, X.; Yang, L.; Lee, E. Dioxin-like PCB 126 increases systemic inflammation and accelerates atherosclerosis in lean LDL receptor-deficient mice. Toxicological Sciences 2018, 162, 548–558. [Google Scholar] [CrossRef]
- Pisani, A.; Paciello, F.; Montuoro, R.; Rolesi, R.; Galli, J.; Fetoni, A.R. Antioxidant therapy as an effective strategy against noise-induced hearing loss: From experimental models to clinic. Life 2023, 13, 1035. [Google Scholar] [CrossRef]
- Poon, E.; Bandara, S.B.; Allen, J.B.; Sadowski, R.N.; Schantz, S.L. Developmental PCB exposure increases susceptibility to audiogenic seizures in adulthood. NeuroToxicology 2015, 46, 117–124. [Google Scholar] [CrossRef]
- Poon, E.; Powers, B.E.; McAlonan, R.M.; Ferguson, D.C.; Schantz, S.L. Effects of developmental exposure to polychlorinated biphenyls and/or polybrominated diphenyl ethers on cochlear function. Toxicological Sciences 2011, 124, 161–168. [Google Scholar] [CrossRef]
- Porte, C.; Albaigés, J. Bioaccumulation patterns of hydrocarbons and polychlorinated biphenyls in bivalves, crustaceans, and fishes. Archives of Environmental Contamination and Toxicology 1994, 26, 273–281. [Google Scholar] [CrossRef]
- Powers, B.E.; Poon, E.; Sable, H.J.; Schantz, S.L. Developmental exposure to PCBs, MeHg, or both: long-term effects on auditory function. Environmental health perspectives 2009, 117, 1101–1107. [Google Scholar] [CrossRef]
- Powers, B.E.; Widholm, J.J.; Lasky, R.E.; Schantz, S.L. Auditory deficits in rats exposed to an environmental PCB mixture during development. Toxicological Sciences 2006, 89, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Gao, Z.; Qiu, Y.; Wang, R.; Tian, K.; Yue, B.; Zhang, X.; Zhang, P.; Wu, Z.; Zhu, Q. NOX2 Contributes to High-Frequency Outer Hair Cell Vulnerability in the Cochlea. Advanced Science 2025, e08830. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, V.; Mukherjea, D.; Dhukhwa, A.; Rybak, L.P. Oxidative Stress and Inflammation Caused by Cisplatin Ototoxicity. Antioxidants (Basel) 2021, 10. [Google Scholar] [CrossRef]
- Ribak, C.E.; Morin, C.L. The role of the inferior colliculus in a genetic model of audiogenic seizures. Anatomy and embryology 1995, 191, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Roegge, C.S.; Morris, J.R.; Villareal, S.; Wang, V.C.; Powers, B.E.; Klintsova, A.Y.; Greenough, W.T.; Pessah, I.N.; Schantz, S.L. Purkinje cell and cerebellar effects following developmental exposure to PCBs and/or MeHg. Neurotoxicology and teratology 2006, 28, 74–85. [Google Scholar] [CrossRef]
- Roegge, C.S.; Wang, V.C.; Powers, B.E.; Klintsova, A.Y.; Villareal, S.; Greenough, W.T.; Schantz, S.L. Motor impairment in rats exposed to PCBs and methylmercury during early development. Toxicol Sci 2004, 77, 315–324. [Google Scholar] [CrossRef]
- Rogan, W.J.; Gladen, B.C.; McKinney, J.D.; Carreras, N.; Hardy, P.; Thullen, J.; Tingelstad, J.; Tully, M. Polychlorinated biphenyls (PCBs) and dichlorodiphenyl dichloroethene (DDE) in human milk: effects of maternal factors and previous lactation. American journal of public health 1986, 76, 172–177. [Google Scholar] [CrossRef]
- Rogan, W.J.; Gladen, B.C.; McKinney, J.D.; Carreras, N.; Hardy, P.; Thullen, J.; Tinglestad, J.; Tully, M. Neonatal effects of transplacental exposure to PCBs and DDE. The Journal of pediatrics 1986, 109, 335–341. [Google Scholar] [CrossRef]
- Roth, J.A.; Salvi, R. Ototoxicity of divalent metals. Neurotoxicity research 2016, 30, 268–282. [Google Scholar] [CrossRef]
- Rowand, C. How old are America's public schools. Education Statistics Quarterly 1999, 1, 53–56. [Google Scholar]
- Sadowski, R.N.; Stebbings, K.A.; Slater, B.J.; Bandara, S.B.; Llano, D.A.; Schantz, S.L. Developmental exposure to PCBs alters the activation of the auditory cortex in response to GABAA antagonism. Neurotoxicology 2016, 56, 86–93. [Google Scholar] [CrossRef]
- Sagiv, S.K.; Thurston, S.W.; Bellinger, D.C.; Altshul, L.M.; Korrick, S.A. Neuropsychological measures of attention and impulse control among 8-year-old children exposed prenatally to organochlorines. Environ Health Perspect 2012, 120, 904–909. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, K.; Prabha, R.L.; Saranya, K.; Bavithra, S.; Krishnamoorthy, G.; Arunakaran, J. Polychlorinated biphenyls impair blood–brain barrier integrity via disruption of tight junction proteins in cerebrum, cerebellum and hippocampus of female Wistar rats: neuropotential role of quercetin. Human & experimental toxicology 2013, 32, 706–720. [Google Scholar]
- Sharma, R.K.; Chern, A.; Golub, J.S.; Lalwani, A.K. Subclinical hearing loss and educational performance in children: A national study. Frontiers in Audiology and Otology 2023, 1, 1214188. [Google Scholar] [CrossRef]
- Shen, H.; Robertson, L.W.; Ludewig, G. Regulatory effects of dioxin-like and non-dioxin-like PCBs and other AhR ligands on the antioxidant enzymes paraoxonase 1/2/3. Environmental Science and Pollution Research 2016, 23, 2108–2118. [Google Scholar] [CrossRef]
- Smith, A.; Carthew, P.; Clothier, B.; Constantin, D.; Francis, J.; Madra, S. Synergy of iron in the toxicity and carcinogenicity of polychlorinated biphenyls (PCBs) and related chemicals. Toxicology letters 1995, 82, 945–950. [Google Scholar] [CrossRef]
- Tao, M.; Sun, J.; Liu, S.; Zhu, Y.; Ren, Y.; Liu, Z.; Wang, X.; Yang, W.; Li, G.; Wang, X.; Zheng, W.; Zhang, J.; Yang, J. An event-related potential study of P300 in preschool children with attention deficit hyperactivity disorder. Front Pediatr 2024, 12, 1461921. [Google Scholar] [CrossRef]
- Thomas, K. 2012. Polychlorinated biphenyls (PCBs) in school buildings: sources, environmental levels, and exposures. US Environmental Protection Agency, National Exposure Research Laboratory ….
- Trnovec, T.; Šovčíková; E; Hust’ák, M. ; Wimmerová; S; Kočan, A.; Jurečková; D; Langer, P.; Palkovičová; Lu, D.r.o.b.n.á.; B Exposure to polychlorinated biphenyls and hearing impairment in children. Environmental Toxicology and Pharmacology 2008, 25, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, M. , Aiko, Y., Kawamoto, T., Hachisuga, T., Kooriyama, C., Myoga, M., Tomonaga, C., Matsumura, F., Anan, A., Tanaka, M., Yu, H.S., Fujisawa, Y., Suga, R., Shibata, E., 2013. Polychlorinated biphenyls (PCBs) decrease the placental syncytiotrophoblast volume and increase Placental Growth Factor (PlGF) in the placenta of normal pregnancy. Placenta 34. [CrossRef]
- Vreugdenhil, H.; Van Zanten, G.; Brocaar, M.; Mulder, P.G.; Weisglas-Kuperus, N. Prenatal exposure to polychlorinated biphenyls and breastfeeding: opposing effects on auditory P300 latencies in 9-year-old Dutch children. Developmental Medicine and Child Neurology 2004, 46, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Feng, S.; Bai, H.; Zeng, P.; Chen, F.; Wu, C.; Peng, Y.; Zhang, Q.; Zhang, Q.; Ye, Q. Polychlorinated biphenyl quinone induces endothelial barrier dysregulation by setting the cross talk between VE-cadherin, focal adhesion, and MAPK signaling. American Journal of Physiology-Heart and Circulatory Physiology 2015, 308, H1205–H1214. [Google Scholar] [CrossRef]
- Zhang, W.; Sargis, R.M.; Volden, P.A.; Carmean, C.M.; Sun, X.J.; Brady, M.J. PCB 126 and other dioxin-like PCBs specifically suppress hepatic PEPCK expression via the aryl hydrocarbon receptor. PloS one 2012, 7, e37103. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Example illustrating selective loss of low-frequency outer hair cells after developmental PCB exposure. Light micrographs from the cochlea of rats showing outer hair cells (top of each image – large bracket) and inner hair cells (bottom of each image – small bracket). Tissue is from a control rat from the apical turn (A), a PCB-exposed rat from the apical turn (B), a control rat from the basal turn (C) and a control rat from the apical turn. Note that there is only loss of outer hair cells and that the loss is seen in the apical turn (B) which is the site of low-frequency tuning of the basilar membrane. Scale bar = 250 mm. Image from Crofton et al. 2000, with permission from publisher.
Figure 1.
Example illustrating selective loss of low-frequency outer hair cells after developmental PCB exposure. Light micrographs from the cochlea of rats showing outer hair cells (top of each image – large bracket) and inner hair cells (bottom of each image – small bracket). Tissue is from a control rat from the apical turn (A), a PCB-exposed rat from the apical turn (B), a control rat from the basal turn (C) and a control rat from the apical turn. Note that there is only loss of outer hair cells and that the loss is seen in the apical turn (B) which is the site of low-frequency tuning of the basilar membrane. Scale bar = 250 mm. Image from Crofton et al. 2000, with permission from publisher.
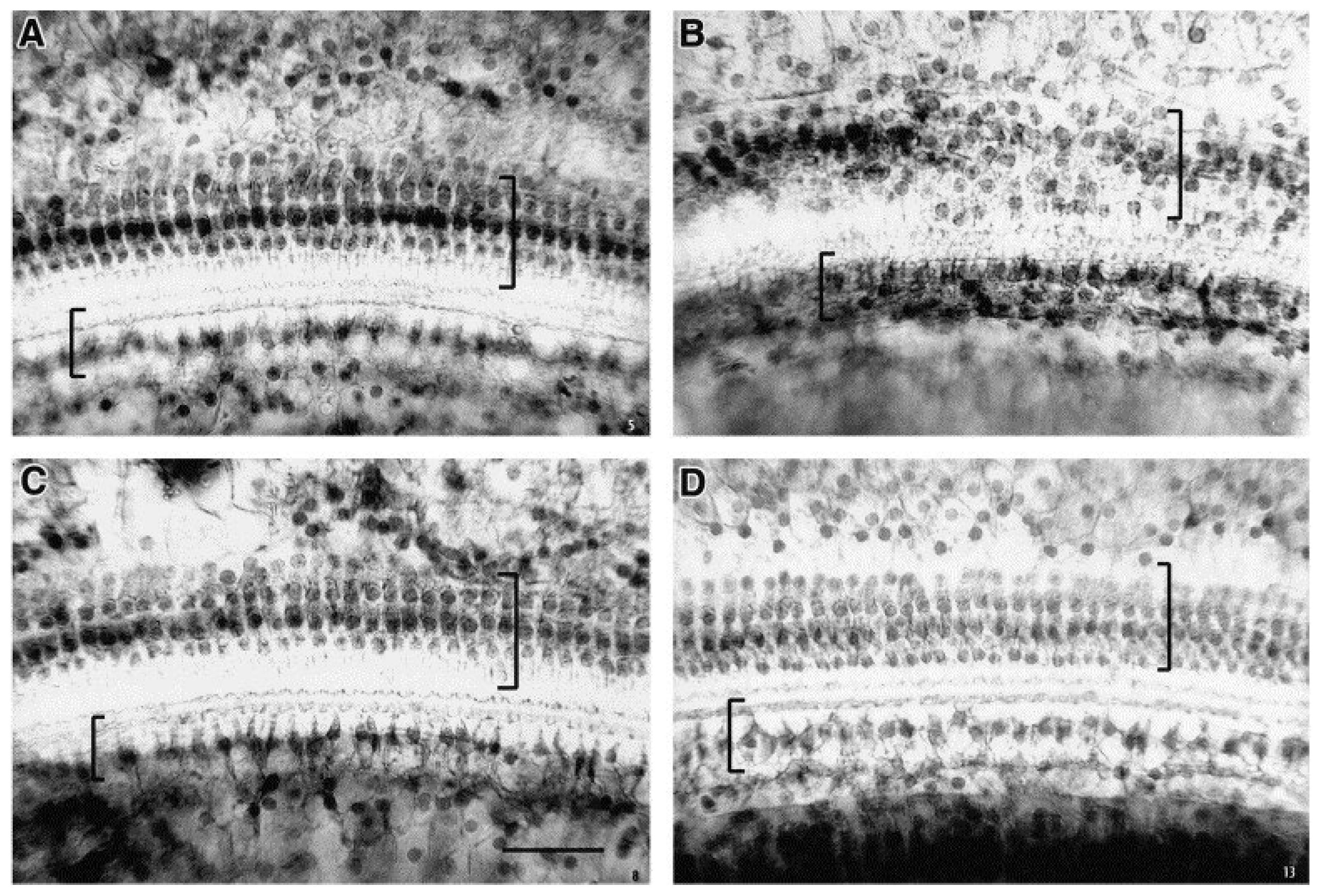
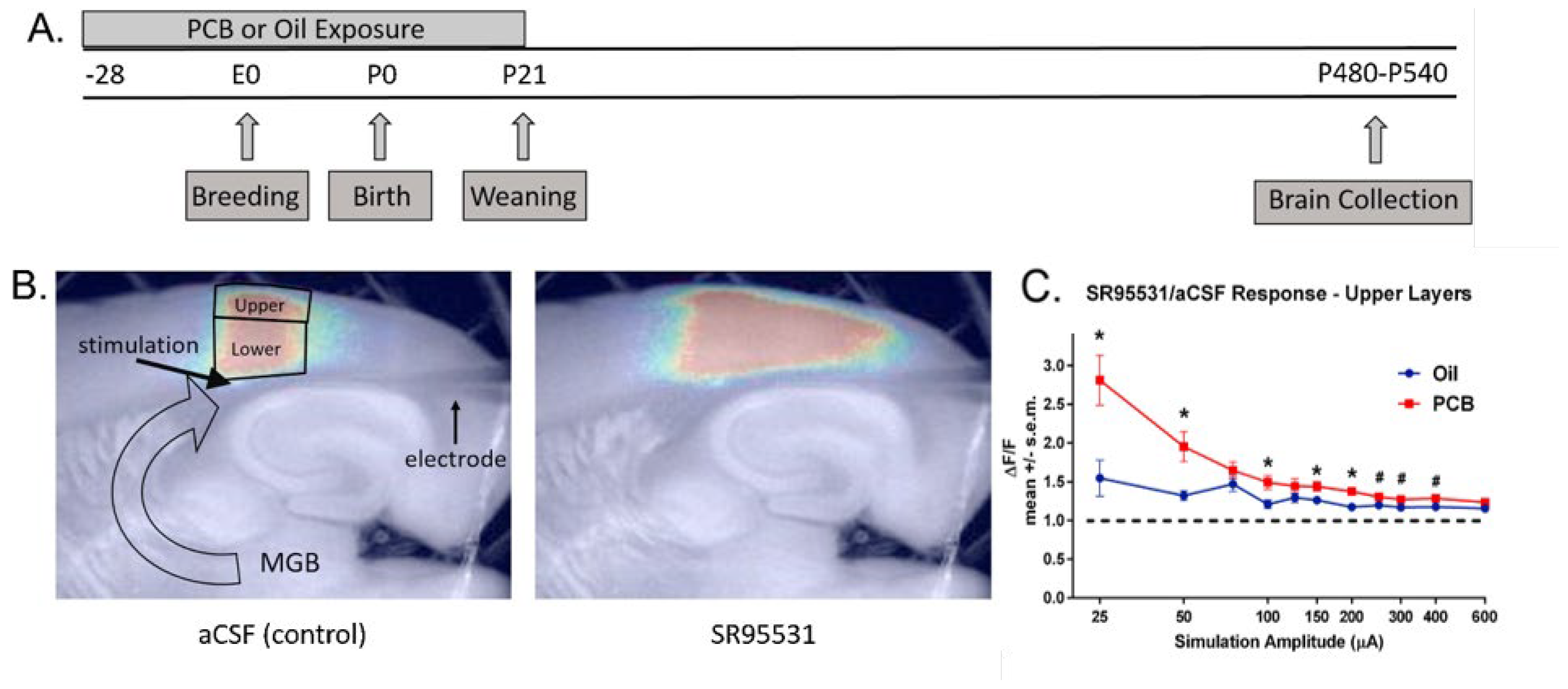
Figure 2.
Example of increased sensitivity to GABAergic blockade at the level of the auditory cortex after developmental PCB exposure. A) Experimental design. Rat dams were treated with PCB or oil vehicle for 28 days prior to breeding, throughout gestation and 21 days after birth. Rats were assessed at ages P480-P540. B) Brain slice flavoprotein autofluoresence imaging was done after stimulation of auditory thalamocortical afferents under normal artificial cerebrospinal fluid (aCSF) conditions and after blocking GABAergic transmission with SR95531. C) Ratio of the response under SR95531 to normal aCSF is elevated across most amplitudes, indicating an increased sensitivity to GABAergic blockade. Modified from Sadowski et al. 2016. Permission obtained from publisher.
Figure 2.
Example of increased sensitivity to GABAergic blockade at the level of the auditory cortex after developmental PCB exposure. A) Experimental design. Rat dams were treated with PCB or oil vehicle for 28 days prior to breeding, throughout gestation and 21 days after birth. Rats were assessed at ages P480-P540. B) Brain slice flavoprotein autofluoresence imaging was done after stimulation of auditory thalamocortical afferents under normal artificial cerebrospinal fluid (aCSF) conditions and after blocking GABAergic transmission with SR95531. C) Ratio of the response under SR95531 to normal aCSF is elevated across most amplitudes, indicating an increased sensitivity to GABAergic blockade. Modified from Sadowski et al. 2016. Permission obtained from publisher.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



