
Submitted:
06 August 2025
Posted:
07 August 2025
You are already at the latest version
Abstract
Bacterial Chondronecrosis with Osteomyelitis (BCO) is a significant issue affecting the animal welfare and economy of the commercial broiler industry, resulting in revenue losses of hundreds of millions of dollars annually. This metabolic disease is frequently associated with Staphylococcus spp. and Enterococcus spp. infections and necrosis of leg and vertebral bones. The typical annual lameness incidence of approximately 3-5% may increase to 30% during outbreaks. Neither the complete etiology and pathogenesis of the disease has been fully understood, nor have effective, commercially available measures of total prevention been identified. Electron beam (eBeam) technology is renowned for producing efficient whole-cell vaccines by preventing bacterial multiplication through irreversible DNA shredding while preserving the integrity of membrane proteins (immunogenic epitopes). This study aims to reduce BCO-induced lameness in broiler chickens via in ovo immunization using an eBeam-inactivated multi-strain Staphylococcus vaccine. 1080 birds were assigned to four vaccination groups: eBeam-inactivated, formalin-inactivated, combination of eBeam and formalin-inactivated, and sham (vehicle). Birds were directly exposed to aerosolized, natural BCO challenge until 56 days of age. Birds vaccinated with the eBeam-inactivated Staphylococcus vaccine showed significant reduction (>50%) in daily cumulative lameness compared to other groups and a decrease of Staphylococcus colonization in the leg-joints of treated birds.
Keywords:
electron beam
; Staphylococcus
; Chondronecrosis
; Osteomyelitis
; lameness
; broilers
1. Introduction
Bacterial Chondronecrosis with Osteomyelitis (BCO) lameness is a metabolic disease that currently poses a significant issue for animal welfare in the poultry industry, resulting in hundreds of millions of dollars in lost revenue globally. Approximately 1 billion broilers out of the global production of 74 billion [1], are produced in Arkansas alone, valued to a total of $2.82 billion annually [2]. Annually, up to 5% of BCO-related animal losses are reported globally [3]. Nevertheless, dramatic increases in lameness, ranging from 15% [4,5] to 30% [3,6,7] of flocks, have been reported worldwide during episodic outbreaks. The disease affects broilers with heavy body weight, causing discomfort and inability to walk, leading to increased mortality and economic losses from condemnation at marketing age [3]. Based on the currently accepted model of BCO pathogenesis, the heavyweight gain of broiler chickens causes initial mechanical damage to the bone joints. This is accompanied by the colonization and proliferation of opportunistic pathogens in the gastrointestinal (GI) and respiratory tracts, together with weakened immunity and damaged tight junction integrity resulting from stress. BCO subsequently progresses with bacterial translocation into the bloodstream via compromised tight junctions in the GI and respiratory tracts, which follows the subsequent hematogenous distribution of bacteria into pre-existing osteochondrotic microfractures within the mechanically stressed bone growth plates [3,8,9,10,11,12]. Necrosis and inflammation of cartilage and joints of long leg bones exacerbate the persistent infection, eventually manifesting as macroscopic bone degeneration, particularly in the proximal heads of the femurs and tibiae, resulting in visible lameness [10,12,13].
Despite research on possible interventions, such as probiotics, prebiotics, organic trace minerals, and other feed supplements to alleviate the incidence of BCO lameness in broiler chickens [10,13,14,15,16,17,18], none of the interventions were fully successful in preventing the condition. A previous practice of supplementing poultry feed and water with antibiotics for over forty years [19] was discontinued due to the spread of antibiotic resistance in pathogenic bacteria [20]. Hence, finding an efficient alternative for BCO prevention is essential, which necessitates a competent model of BCO challenge and induction in broiler chickens to evaluate various treatment effects on BCO reduction. The present study employs the aerosol transmission model of BCO induction, developed by our lab [21]. This model features 2 wire-floor pens placed in front of all other litter-floor pens, positioned closer to the cooling pads (upwind), and separated from the litter-floor pens by a buffer zone that prevents contact. Tunnel fans are placed at the rear end (downwind) of the house. Birds raised on wire floors are continuously stressed by having to walk for feed and water, which are placed at the opposite ends of the pens, making them prone to developing microfractures in leg bones, leading to accelerated lameness. These birds exhale the BCO pathogens into the air, and contaminated aerosol particles are drawn backward by the tunnel fans, infecting the birds raised in litter-floored pens. This model mimics the natural infection in a real-world scenario, as in commercial broiler chicken houses.
Staphylococcus spp. is an abundantly isolated genus of bacteria from femoral and tibial BCO lesions of broiler chicken [11,13,22,23], and hence known to be associated with the pathogenesis of BCO [24,25,26,27]. Controlling Staphylococcus infections is challenging as they have evolved several mechanisms to evade immune responses of various hosts. These include adaptations to prevent opsonization [28], phagocytosis [29], and the production of superantigens [30,31]. Whole-cell vaccines are an efficacious means of treating microbial diseases, offering several advantages over other vaccine types, including their relative time-conservation and cost-effectiveness in production, as well as being carriers of multiple immunogenic epitopes [32]. Notably, live whole-cell vaccines mimic natural infections and immune responses, but their drawbacks include reduced stability and the potential for pathogen revival. Inactivated whole-cell vaccines, typically treated with formaldehyde or other alkylating reagents [33] excel in terms of host safety and high stability. However, inactivating agents can cause damage to immunogenic epitopes by forming cross-links of surface proteins and triggering other chemical reactions, resulting in limited immunogenicity and reduced cross-presentation of antigens through the priming of CD8+ T cells [33,34,35,36]. Interestingly, electron beam (eBeam), an ionizing radiation technology, presents a promising alternative for producing inactivated whole-cell vaccines without damaging the cell surface proteins (epitopes) [19,37], while achieving irreversible reproductive death of the treated bacteria [38]. This success lies in eBeam’s property of irreparable shredding of the bacterial genome upon exposure (direct effect), coupled with short-lived free-radical attack owing to the products of radiolysis of intracellular solvents (indirect effects) [38,39]. Several studies have demonstrated eBeam’s ability to retain pathogen membrane integrity using fluorescence microscopy and electron microscopy [19,37,40,41,42]. Praveen et al. (2021) [42] have demonstrated that eBeam-inactivated Salmonella Typhimurium (ST) to be metabolically active, and to stimulate upregulation of CD40, MHC-II, CD80, and CD86 in mice immune cells, at similar levels to live ST vaccines, concluding that an eBeam vaccine is as efficient as a live vaccine but as safe as a killed vaccine. eBeam is also approved by the US Food and Drug Administration to inactivate pathogens and spoilage microorganisms in food [43]. eBeam excels above other types of ionizing radiation like gamma and X-rays in terms of its 1) safety and eco-friendliness, 2) cost-effectivity, 3) high efficiency, 4) producing substantial doses of radiation in few seconds, 5) producing planar, homogenous radiation profiles and 6) ability to switch on and off [38,44,45,46]. eBeam appears to provide a promising alternative for the efficient preparation of whole-cell vaccines against pathogenic bacteria.
Based on eBeam’s exceptional ability to produce safe and efficient whole-cell-inactivated vaccines and the dominance of Staphylococcus in the pathogenicity of BCO, it was hypothesized that efficient control of Staphylococcus in broiler chickens would lead to a reduction in lameness. Accordingly, the present study utilizes an eBeam-inactivated vaccine containing multiple Staphylococcus species to reduce BCO lameness in broiler chickens, significantly. This study was conducted as a follow-up to a previous study in our research laboratory, which tested the efficacy of BCO reduction in broilers using an eBeam inactivated vaccine containing two species of Staphylococcus [47]. The present study utilizes a modified version of the vaccine with dose optimization and incorporation of additional strains of Staphylococcus, which are frequently isolated from BCO lesions of lame broilers. Moreover, birds of the previous vaccine study were only challenged with bacterial strains included in the vaccine, but the current study closely mimics a real-world setting, where the birds were challenged with the natural intestinal and environmental microbiota. The objective of this study is to improve animal welfare and increase revenue via lameness prevention in broiler chickens. In addition, our eBeam vaccine could prevent foodborne diseases related to the consumption of poultry products contaminated with Staphylococcus spp. [48,49] thereby ensuring consumer health and well-being.
2. Materials and Methods
2.1. Bacterial Culture and Vaccine Preparation
Field-isolated strains of Staphylococcus agnetis, Staphylococcus aureus, Staphylococcus lentus, and two strains of Staphylococcus cohnii were used. The five strains were inoculated separately in 100 mL of Tryptic Soy Broth (TSB, Difco, Becton Dickinson, Franklin Lakes, NJ) and incubated at 37°C overnight. The above cultures were pooled on the following day and centrifuged to remove the spent media. The bacterial pellet was resuspended in TSB at a concentration of ~1 × 108 CFU/mL. The above mix of strains was processed as follows to prepare the types of vaccines mentioned below.
2.1.1. Electron Beam (eBeam) Vaccine
The eBeam treatment was conducted at the National Center for Electron Beam Research (NCEBR) at Texas A&M University (TAMU), College Station, Texas. The cocktails of Staphylococcus strains at ~1x108 CFU/mL in TSB were placed in plastic bags and heat-sealed to prevent leakage, for shipping in accordance with USDA-APHIS regulations. The sealed bags, each containing 20mL of culture were placed in triplicate into a second plastic bag and heat-sealed. Each secondary bag was sealed in a leak-proof 95kPa specimen transport envelope. After each sealing step, the bags were sprayed with 70% Ethanol and dried with clean paper towels. The triple-bagged samples were finally packed in a Department of Transportation and IATA-approved box for NCEBR, TAMU. A high-energy (10 MeV, 15 kW) linear accelerator was used for the eBeam treatment. Alanine pellet dosimeters, placed on the top and bottom of each sample (triple-bagged Staphylococcus mix), were used to confirm the absorbed dose by the samples. The dose-uniformity ratio was maintained as close as possible to 1.0 to ensure that the entire sample received the desired dose. The samples exposed to 8 kiloGrays (kGy) were stored at 4°C upon receipt and used as the eBeam vaccine.
2.1.2. Formalin-Inactivated (FK) Vaccine
A previously described protocol by Kaminski et al. (2014) [50] was adopted to prepare the FK vaccine. Briefly, 100 mL of the overnight-grown Staphylococcus strain cocktail in TSB at ~1x108 CFU/mL was treated with 0.6% of formaldehyde (v/v) (Electron Microscopy Sciences, Hatfield, PA). This solution was incubated at room temperature for 48 h, while stirring at 200 RPM (PC-420D Digital Hot Plate Stirrer, Corning Inc., Corning, NY). Afterwards, the vaccine solution was centrifuged at 1932 x g for 15 min at 4°C. The supernatant was discarded, and the cell suspension was resuspended in an equal volume of Phosphate-Buffered Saline (PBS) and centrifuged at 1932 x g for 15 min at 4°C. The final bacterial cell pellet was then resuspended in an equal volume of fresh TSB before storage at 4°C.
2.1.3. Combination (F+E) Vaccine
Equal proportions of both eBeam and formalin-inactivated vaccines were mixed to prepare a vaccine that contains both the eBeam and formalin-inactivated Staphylococcus cocktails.
2.2. Confirmation of Bacterial Inactivation, Membrane Integrity and Viability
2.2.1. Bacterial Inactivation
For confirmation of complete bacterial inactivation, eBeam and formalin-inactivated treated cells were inoculated into TSB. Furthermore, treated samples were serially diluted in Butterfield's Phosphate Diluent (BPD) [51], and plated onto Mannitol Salt Agar (MSA, Difco, Becton Dickinson) plates. A set of samples inoculated in TSB and plated in MSA plates were incubated overnight at 37°C, while the remaining samples inoculated in TSB were incubated overnight at room temperature. The above tubes and plates were then incubated at room temperature for a week prior to vaccine administration. Subsequently, the TSB tubes were monitored for bacterial resuscitation for up to a month post-treatment (eBeam/formalin).
2.2.2. Bacterial Membrane Integrity
For evaluating bacterial membrane integrity, the LIVE/DEAD BacLight Bacterial Viability Kit (Invitrogen Inc., Waltham, MA) was used according to the manufacturer’s guidelines. Briefly, bacterial cells (untreated, eBeam-treated, and formalin-treated) were diluted at 1:10 in BPD. Then, the cells were stained with a mixture of SYTO 9 + propidium iodide and incubated in the dark for 15 min at room temperature. Finally, stained bacterial cell samples were evaluated by fluorescence microscopy (BZ-X800 fluorescence microscope, Keyence, Osaka, Japan) in a 12-well, flat-bottom glass plate (Corning Inc. Kennebunk, ME).
2.2.3. Bacterial Cell Viability
To support the selection of 8 kGy as the desired eBeam dose for the vaccine, data from a previously conducted viability assay on eBeam and formalin-inactivated Staphylococcus aureus and Staphylococcus agnetis were utilized. For this purpose, the BacTiter-Glo™ Microbial Cell Viability Assay (Promega, Madison, WI) was followed according to the manufacturer’s instructions. Shortly, 100 µL of untreated/control (shipped to NCEBR but not eBeam treated), eBeam-treated (at 8, 9, and 10kGy), and formalin-treated bacteria of each of the above strains in TSB at ~1x108 CFU/mL were added to a 96-well, flat-bottom, dark-walled plate (Invitrogen Inc., Waltham, MA) in triplicate. This was followed by the addition of 100 µL of BacTiter-Glo™ Reagent to each well and incubation at room temperature on a plate shaker (VWR International, Radnor, PA) at 150 RPM for 5 min (covered by an Aluminum foil to protect from the light). Finally, luminescence was recorded using a BioTek Synergy Multi-Mode Microplate Reader (Agilent Technologies, Santa Clara, CA).
2.3. Broiler Chicken Vaccination Trial
2.3.1. Egg Placement and in Ovo Vaccination
All procedures and protocols were approved by the University of Arkansas System, Division of Agriculture, Institutional Animal Care and Use Committee (IACUC), Protocol # 23058. Figure 1 illustrates the summary of the study's timeline.
Figure 1.
Schematic illustrating the study timeline. E: age of embryos throughout incubation, D: age of birds post-hatch.
Figure 1.
Schematic illustrating the study timeline. E: age of embryos throughout incubation, D: age of birds post-hatch.
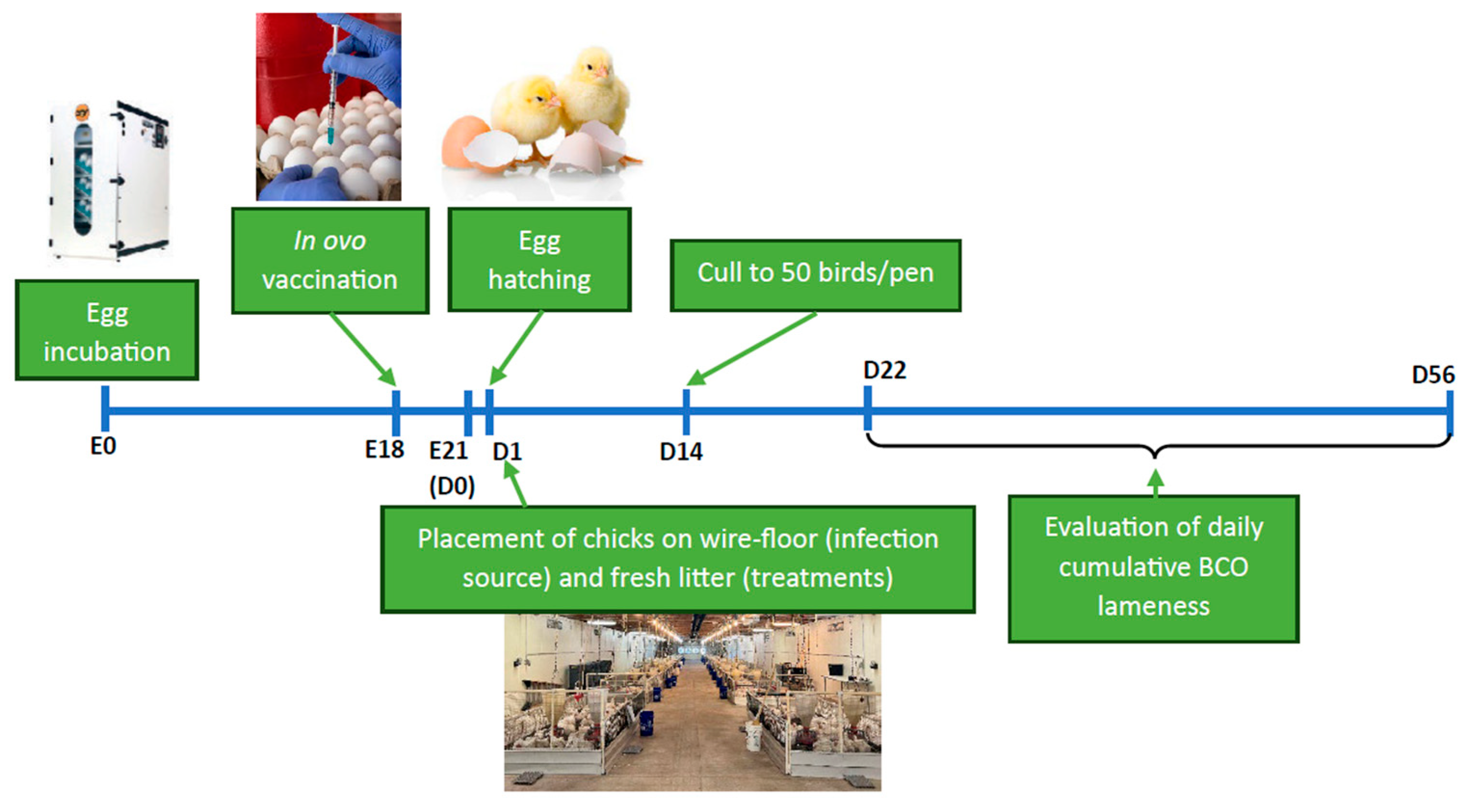
One thousand, five hundred and forty-three (1543) broiler eggs (0-day-old Cobb500) were obtained from a local hatchery and placed for incubation following IACUC-approved conditions (Temperature = 99.6°F, RH = 85%). They were candled for viability on days 10 and 18 of incubation to remove spoiled, damaged, or non-viable eggs. The in ovo vaccination was administered to the viable eggs on D18 of incubation. The eBeam and formalin-inactivated vaccines were diluted in fresh TSB to ~1x107 CFU/mL. Equal proportions of both vaccines were mixed to prepare the combination vaccine. For vaccination, the large ends of all eggs were sterilized with 70% Ethanol, wiped and gently pierced with a 0.25“ long 18-gauge needle. The vaccine or sham (TSB) was administered directly into the amniotic cavity using a 1 mL tuberculin syringe with a 1.5“ long, 25-gauge needle equipped with a modified needle guard and injected at an angle to limit all injections to a depth of 3 cm. The different treatment groups of the study are explained in Table 1 below. Following individual injections, injection sites on all eggs were sealed with melted paraffin using cotton swabs. Finally, vaccinated eggs were placed in hatching cabinets for further incubation (Temperature = 98°F, relative humidity = 85%) until hatching.

Table 1.
Treatment groups of the vaccine study.
| Treatment | Flooring | Vaccine | Number of pens allocated |
|---|---|---|---|
| WF - Infection source1 | Wire | sham | 2 |
| eB - eBeam group2 | Litter | eBeam-inactivated | 4 |
| FK - Formalin group3 | Litter | formalin-inactivated | 4 |
| F+E - Combination group4 | Litter | eBeam + formalin | 4 |
| Sham - Control group5 | Litter | sham | 4 |
1 Birds raised on a wire floor develop the lameness disease at an accelerated rate due to stress, and the house design transmits the pathogen via aerosol to all other pens. 2 Birds vaccinated with 107 CFU/mL of electron beam-inactivated vaccine. 3 Birds vaccinated with 107 CFU/mL of formalin-inactivated vaccine. 4 Birds vaccinated with a combination of electron beam and formalin-inactivated vaccine. 5 Birds vaccinated with pure culture media (Tryptic Soy Broth) without bacteria.
2.3.2. Live Bird Study and Sampling
One thousand and eighty (1080) birds were placed at 60 birds/pen on the day of hatch, according to the design in Figure 2.
Figure 2.
Schematic diagram representing the relative locations of the pens in our facility and the treatment groups of birds in each pen (WF – Wire floor pens with sham vaccinated birds, eB – Litter floor pens with birds that received the eBeam-inactivated Staphylococcus vaccine, FK – Litter floor pens with birds that received the formalin-inactivated Staphylococcus vaccine, F+E – Litter floor pens with birds that received the combination Staphylococcus vaccine, sham – Litter floor pens with birds that received the sham vaccine). Each pen had 60 birds on the day of hatch.
Figure 2.
Schematic diagram representing the relative locations of the pens in our facility and the treatment groups of birds in each pen (WF – Wire floor pens with sham vaccinated birds, eB – Litter floor pens with birds that received the eBeam-inactivated Staphylococcus vaccine, FK – Litter floor pens with birds that received the formalin-inactivated Staphylococcus vaccine, F+E – Litter floor pens with birds that received the combination Staphylococcus vaccine, sham – Litter floor pens with birds that received the sham vaccine). Each pen had 60 birds on the day of hatch.

Sixteen (16) litter floor pens and 2 wire floor pens of 5’ × 10’ were used following the aerosol transmission infection model developed by our lab [21]. The bird stocking density was maintained at 0.83 ft2/bird (D1–14) and 1 ft2/bird (D15–56), and the birds of all pens were culled down to 50 on D14. Feed and water were provided ad libitum. Standard crumble starter diet (D1-34) followed by pellet finisher diet (D34-56) were provided. Thermoneutral temperatures were maintained throughout as 32°C (90°F) for D1-3, 31°C (88°F) for D4-6, 29°C (85°F) for D7-10, 26°C (80°F) for D11-14, and 24°C (75°F) thereafter. A heat lamp was positioned in each pen to provide supplemental heat through D28 when necessary. The photoperiod was 23L:1D, with white light throughout the experiment. All remaining birds were humanely euthanized at the end of the trial on D56.
Starting from D22 of the experiment, birds of each pen were prompted to move daily (using brooms), and any bird that was reluctant to move/stand up was identified as clinically lame [52]. Upon identification as lame, the birds were humanely euthanized and necropsied to expose the proximal femoral and tibial heads, to determine the severity of the progression of bone necrosis. BCO lesion categorization, based on the severity of necrosis, was adopted from Wideman (2016) [3]. On days 16, 32, and 56, 12 birds/treatment (4 birds/pen) were randomly selected and sampled as follows. Blood was collected (ante-mortem) in EDTA vacutainers for the analysis and comparison of plasma Staphylococcus-specific antibody (IgM, IgY, and IgA) profiles via Enzyme-Linked Immunosorbent Assay (ELISA) and whole blood cell populations via immunofluorescent staining followed by flow cytometry (days 16 and 32) among treatments. Approximately 2.5 cm of tracheal tissue was collected without contamination by blood, into 1.5 mL BPD to extract tracheal mucus. Later, the lumens of tracheal tissues were flushed with the same BPD solution several times to get the mucus into the BPD prior to discarding the tissue for the analysis and comparison of IgA profiles.
On D56, BCO lesion swabs of 2 bones/bird (femur and tibia) from 4 birds/treatment (1 bird/pen) were collected aseptically and plated onto CHROMagar Orientation (CO, CHROMagar™, Paris, France) and Tryptic Soy Agar (TSA, Difco, Becton Dickinson). The above plates were incubated overnight at 37°C for bacterial enumeration. The total number of colonies and morphologies were recorded, and the isolated colonies were re-streaked on fresh media plates. Representative colonies were subjected to DNA extraction, PCR amplification of the rDNA 16S V1-V5 region, and sequencing for the identification of BCO-causing bacterial species.
2.4. Sample Processing
2.4.1. Bacterial Species Identification from BCO Lesions
Isolated pure cultures of bacteria obtained from BCO lesions, as explained above, were inoculated in TSB and incubated overnight at 37°C. 40% glycerol stocks were made for each pure culture for preservation at -80°C. DNA extraction from the cultures was performed using the NucleoSpin® Microbial DNA kit (Macherey-Nagel, Düren, Germany), following the manufacturer’s instructions.
The extracted DNA was then PCR amplified using ribosomal RNA (rRNA) as a biomarker. Conventional PCR-based amplification of the rDNA 16S V1-V5 region was performed using the protocol described by Asnayanti et al. (2024) [23]. Briefly, the PCR mixture of 50 μL was prepared using 25 μL of Phusion® High-Fidelity PCR Master Mix (New England Biolabs® Inc., Massachusetts, USA), 0.5 μM of each forward (5’-AGAGTTTGATCCTGGCTCAG-3’) and reverse (5’-GTGCGGGCCCCCGTCAATTC-3’) primers, 1.5 μL of dimethyl sulphoxide (DMSO), 2 μL of DNA samples, and 20.5 μL of nuclease-free water. A thermal cycler (Bio-Rad T100TM, California, USA) was set to 98°C for 30 s, 98°C for 10 s, 71°C for 30 s, 72°C for 30 s (35 cycles), followed by 72°C for 3 min, and an infinite hold of 4°C was used for the amplification. Afterwards, 40-60 ng/µL of the amplified DNA, along with 2-10 pmol/µL of the above primers were shipped to the Eurofins Genomics Lab (Louisville, KY, USA) for sequencing. Sequence results were visualized using the ApE Plasmid Editor software [53] and blasted (using the Blastn tool) against the database of the National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/). The species were identified from the database with a sequence similarity of greater than 98%.
2.4.2. Analysis and Comparison of Antibody (IgM, IgY, and IgA) Profiles
Relative levels of Staphylococcus-specific IgM, IgY, and IgA in blood, as well as mucosal IgA levels, were determined by ELISA. Briefly, a 1x107 CFU/mL Staphylococcus strain cocktail (comprising the same strains as the vaccine) was suspended in 0.05M Coating buffer (0.05M NaHCO3, 0.05M Na2CO3, pH 7.4). 96-well flat-bottom plates were coated with 100 µL of the inactivated bacterial cell suspension per well. The plates were then incubated at 37°C for 2 h, followed by an overnight incubation at 4 °C to allow the Staphylococcus cells to adhere. Next, the wells of all plates were washed with a washing solution (50 mM Tris, 0.14 M NaCl, 0.05% Tween 20; pH 8.0) three times (leaving the wash solution in the wells for 2 min, each cycle). The solutions in the wells were thoroughly aspirated, and the Staphylococcus-coated plates were stored at 4°C for further use. For each assay, the wells of the plates were blocked with 200 µL of blocking buffer [50 mM Tris, 0.14 M NaCl, 1% bovine serum albumin (BSA) (Sigma-Aldrich, St. Louis, MO), pH 8.0] and incubated at room temperature (RT) for 30 min, followed by 3 washes. Frozen tracheal mucus and plasma samples were thawed and diluted (titers determined via pre-tests) in sample diluent buffer (50 mM Tris, 0.14 M NaCl, 1% BSA, 0.05 % Tween 20, and pH 8.0) and added in triplicate at 100 µL/well. Sample dilutions for each assay are as follows; IgM: 1/400 (D16), 1/800 (D32), 1/3200 (D56); IgY: 1/200 (D16 and D32 samples), 1/600 (D56); IgA: 1/50 (D16 and D32), 1/400 (D56) with tracheal mucus and 1/50 (D16), 1/100 (D32), 1/200 (D56) with plasma. Aliquots of plasma and tracheal mucus samples were pooled per collection day prior to freezing. Nine doubling dilutions (100 μL/well; 2 wells/dilution) of sample pools were included in all assays as positive controls to generate a standard curve. Furthermore, each plate contained 3 ‘Blank’ wells, containing the highest concentration of the sample pool for each assay, and 3 wells of non-specific binding (NSB) control, containing diluent buffer at 100 µL/well. After adding samples and controls, the plates were incubated at 37°C for 2 h, followed by 3 washes. Horse-radish peroxidase (HRP)-conjugated, goat anti-chicken IgG (Ig γ-heavy chain), IgM, and IgA detection antibodies (Bethyl Laboratories, Montgomery, TX) were diluted in diluent buffer at 1/10,000, 1/5,000, and 1/2,500 for the detection of IgY, IgM, and IgA, respectively. Each well received the above detection antibodies (in accordance with the particular assay) at 100 µL/well, except for the ‘Blank’ wells, which received the unconjugated (no HRP enzyme) detection antibody at 100 µL/well. The plates were then incubated for 1 h at 37 °C, followed by 3 washes. Afterwards, 100 µL/well of 3,3’,5,5’-tetramethylbenzidine substrate solution (TMB, Thermofisher, Waltham, MA) was added and incubated at 37°C for 15 min. Finally, 100 µL/well of 2M sulfuric acid was added to stop the reaction. Absorbance at 450 nm was measured for each plate using a 96-well spectrophotometer (ELx800, BioTek, Winooski, VT). The absorbance units (a.u.) for the pooled sample dilutions were used to generate a standard curve for each plate, and the line equation was used to adjust relative a.u. levels of IgY, IgM, and IgA for each assay.
2.4.3. Flow Cytometry
Whole blood immunofluorescence staining and cell population analysis by flow cytometry were carried out as described [47,54,55]. Briefly, 20 µL of EDTA- blood was diluted in 980 µL of PBS+ staining buffer (PBS pH 7.2 +1% BSA + 0.1% NaN3) (1:50). Fifty µL of each diluted blood sample was mixed with 50 µL of the respective antibody mixture, as explained below, and incubated at 4°C for 45 min, protected from light. All antibodies were purchased from Southern Biotech (Birmingham, AL) unless otherwise specified. Three combinations of mouse-anti-chicken monoclonal antibodies were used: 1) FITC-CD41/61 (Biorad, Hercules, CA), PE-CD3 and SPRD-CD45 to identify leucocytes (CD45+CD41/61-), T cells (CD45+CD3+), and thrombocytes (CD45lowCD41/61+), 2) APC-CD45, FITC-CD4, PE- γδTCR and SPRD-CD8α to identify T cell subsets, and 3) FITC-KUL01, PE-Bu1 and SPRD-CD45, to identify monocytes (CD45+KUL01+Bu-1-) and B cells (CD45+KUL01-Bu-1+), respectively. Heterophils were identified based on forward- and side-scatter characteristics of CD45+KUL01-Bu-1- cells (Seliger et al., 2012). Afterwards, 150 µL of PBS+ was added to all samples (1:250 final blood dilution). Samples were analyzed using a BD C6 Accuri flow cytometer (Becton Dickinson, San Jose, California). Flow cytometry data were analyzed using FlowJo software version 10.10.0 (Becton Dickinson).
2.5. Statistical Analysis
One-way Analysis of Variance (ANOVA) followed by Tukey’s multiple means comparison test was conducted to analyze the luminescence data to compare bacterial viability between treatments on GraphPad Prism software version 10.2.0 (San Diego, CA, United States). Microsoft Excel 365 (Microsoft, Redmond, WA, USA) software was used to analyze the percentage of daily cumulative lameness and the percentage of bacterial isolates from BCO lesions. The impact of treatments on lameness reduction was examined using a logistic regression model, utilizing a Generalized Linear Model in R version 4.2.2 (R Foundation for Statistical Computing, Vienna, Austria). Kruskal-Wallis Tests followed by Dunn post-hoc tests, adapted from Yang et al. (2023)[56] were used to compare the total percentages of BCO lesions between treatment groups, separately for femoral and tibial lesions on JMP Pro software version 18.0.2. Effects of time, treatment, and time by treatment interactions for the differences in antibody responses and concentrations of blood leukocyte populations were analyzed using two-way ANOVA followed by Tukey’s multiple means comparison test on GraphPad Prism. Continuous data were presented as means ± standard error of the means (SEM). Statistical significance was considered at P values < 0.05.
3. Results
3.1. eBeam-Treated Staphylococcus Cells Were Inactivated Entirely, While Retaining Their Membrane Integrity and Higher Viability than Formalin-Treated Staphylococcus
The present study is a follow-up to a broiler vaccination trial that used an eBeam-inactivated Staphylococcus vaccine (containing Staphylococcus aureus and Staphylococcus agnetis) conducted by our laboratory, which employed an eBeam dose of 10 kGy [47]. For the present study, it was hypothesized that if complete inactivation could be achieved by a lower eBeam dose, it would be less detrimental to the bacterial cells, which would provide increased the immunogenicity to the vaccinated birds.
Figure 3.
eBeam-treated bacteria retained viability. Comparison of the luminescence (i.e., viability) of non-treated (0kGy), formalin-inactivated (FK), and eBeam-treated (8, 9 and 10 kGy) Staphylococcus aureus and Staphylococcus agnetis on the first week (D3) and the 5th week of inactivation. Graphs show Means ± SEM. a-c: Bars with different alphabets depict significant differences between treatments (P<0.05).
Figure 3.
eBeam-treated bacteria retained viability. Comparison of the luminescence (i.e., viability) of non-treated (0kGy), formalin-inactivated (FK), and eBeam-treated (8, 9 and 10 kGy) Staphylococcus aureus and Staphylococcus agnetis on the first week (D3) and the 5th week of inactivation. Graphs show Means ± SEM. a-c: Bars with different alphabets depict significant differences between treatments (P<0.05).
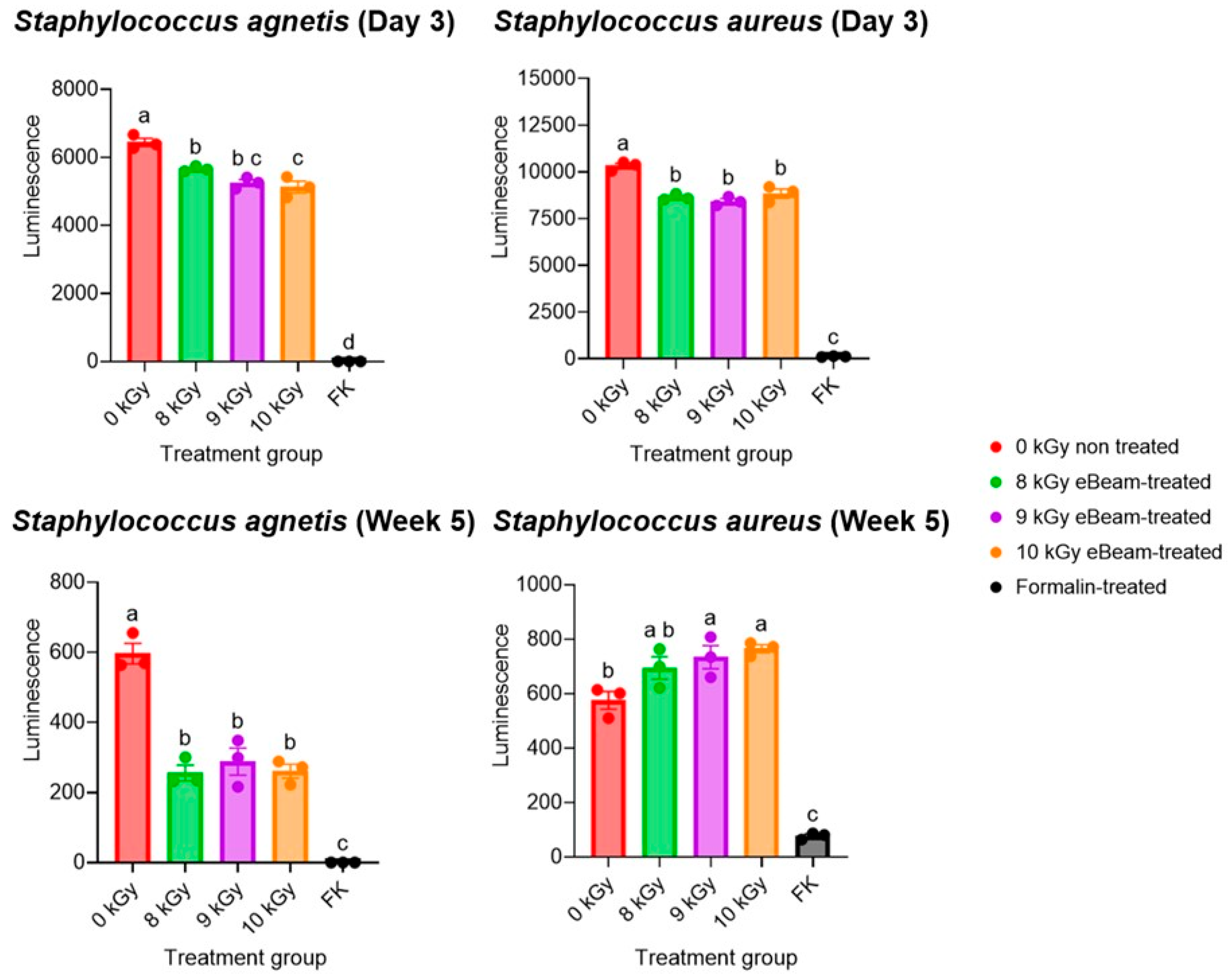
No growth was observed in the TSB tubes as well as the MSA plates inoculated with inactivated bacteria until the point of vaccine administration (within the first week of inactivation), up to a month after inactivation. These results indicate that both eBeam and formalin are capable of completely inactivating the Staphylococcus cocktail, thereby preventing both multiplication and resuscitation. This also confirms the success of the selected eBeam dose for inactivation. Accordingly, the viability was compared across Staphylococcus aureus and Staphylococcus agnetis exposed to eBeam at 8, 9, and 10 kGy and treated with formalin in preparation for this study (Figure 3). The BacTiter-Glo™ Microbial Cell Viability Assay (Promega, Madison, WI) was utilized, and the ATP and O2 in bacterial cells were quantified in terms of relative luminescence, which is a direct indication of the cells' metabolically active and viable status.
The viability of 8 kGy eBeam-treated S. agnetis was significantly higher than 10 kGy-treated cells during the week of vaccination, while the viability of eBeam-treated cells (all 3 doses) was significantly higher than the viability of formalin-treated cells in both strains. The assay was conducted weekly for 5 weeks following inactivation. Even after 5 weeks of inactivation, the viability of eBeam-treated cells (both strains) was similar between each dose but significantly different from that of formalin-treated Staphylococcus. This supported the selection of 8 kGy as the target eBeam dose for the present vaccine. Furthermore, the data suggest that the viability and metabolic activity of eBeam-treated cells are significantly higher than those of formalin-treated cells. Accordingly, despite their inability to replicate, eBeam-treated cells function as live cells, which is important for host immune response development. Therefore, this evidence supports the claim by Praveen et al. (2021) [42] that an eBeam vaccine is as efficient as a live vaccine but as safe as a killed vaccine.
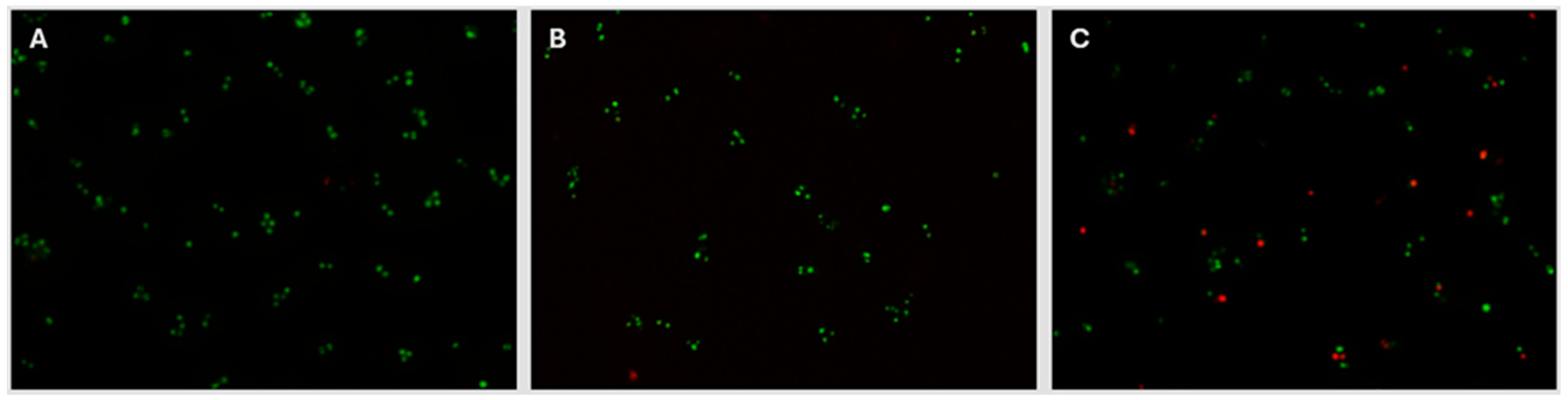
The decision to select 8 kGy as the target eBeam dose for the present Staphylococcus vaccine was further supported by the evidence that the membrane integrity of eBeam-treated cells at 8 kGy is similar to that of non-treated cells. The BacLight assay (Invitrogen Inc., Waltham, MA) was used, which utilizes a mixture of 2 fluorescent stains to identify cells with damaged membranes apart from those with intact membranes. Live cells with non-damaged membranes fluoresce green under fluorescent microscopy, while dead cells with damaged membranes fluoresce red. Figure 4 illustrates that the membrane integrity of non-treated and 8 kGy eBeam-treated Staphylococcus cells are conserved, by the presence of a majority of green (live) cells. In contrast, the membranes of formalin-treated Staphylococcus cells are damaged, resulting in considerably higher numbers of red cells.
Figure 4.
eBeam-treated cells retained their membrane integrity. Fluorescence microscopy images (100x) of Staphylococcus cells treated with A) no treatment, B) 8kGy eBeam, and C) formalin-inactivation, after being stained with the LIVE/DEAD BacLight Bacterial Viability Kit (Invitrogen Inc., Waltham, MA).
Figure 4.
eBeam-treated cells retained their membrane integrity. Fluorescence microscopy images (100x) of Staphylococcus cells treated with A) no treatment, B) 8kGy eBeam, and C) formalin-inactivation, after being stained with the LIVE/DEAD BacLight Bacterial Viability Kit (Invitrogen Inc., Waltham, MA).
3.2. eBeam-Inactivated Staphylococcus Vaccine Significantly Decreased Lameness Compared to the Other Treatment Groups
The daily cumulative BCO lameness was evaluated for all birds starting from D22 till the end of the trial. By D56, the total cumulative lameness of the WF group was 84.26%, while the control (sham) group had 67.5% cumulative lameness (Figure 5A), which illustrates the success of the wire floor pens in inducing high lameness in birds and the effectiveness of the aerosol transmission model in disseminating lameness to the litter floor pens. By D56, the daily cumulative lameness rates of the eBeam, formalin, and combination groups were 32.22%, 66.5%, and 64.5%, respectively (Figure 5A).
Figure 5.
eBeam-inactivated Staphylococcus vaccine significantly reduced lameness. 5A: Graph of variation of daily cumulative lameness of treatment groups (wire floor, eBeam, sham, formalin, and combination) through days 30-56. 5B: Percentage lameness reduction of vaccine groups (eBeam, formalin, and combination) relative to the sham group. a, b: Different alphabets denote significant differences between treatments (P<0.05).
Figure 5.
eBeam-inactivated Staphylococcus vaccine significantly reduced lameness. 5A: Graph of variation of daily cumulative lameness of treatment groups (wire floor, eBeam, sham, formalin, and combination) through days 30-56. 5B: Percentage lameness reduction of vaccine groups (eBeam, formalin, and combination) relative to the sham group. a, b: Different alphabets denote significant differences between treatments (P<0.05).
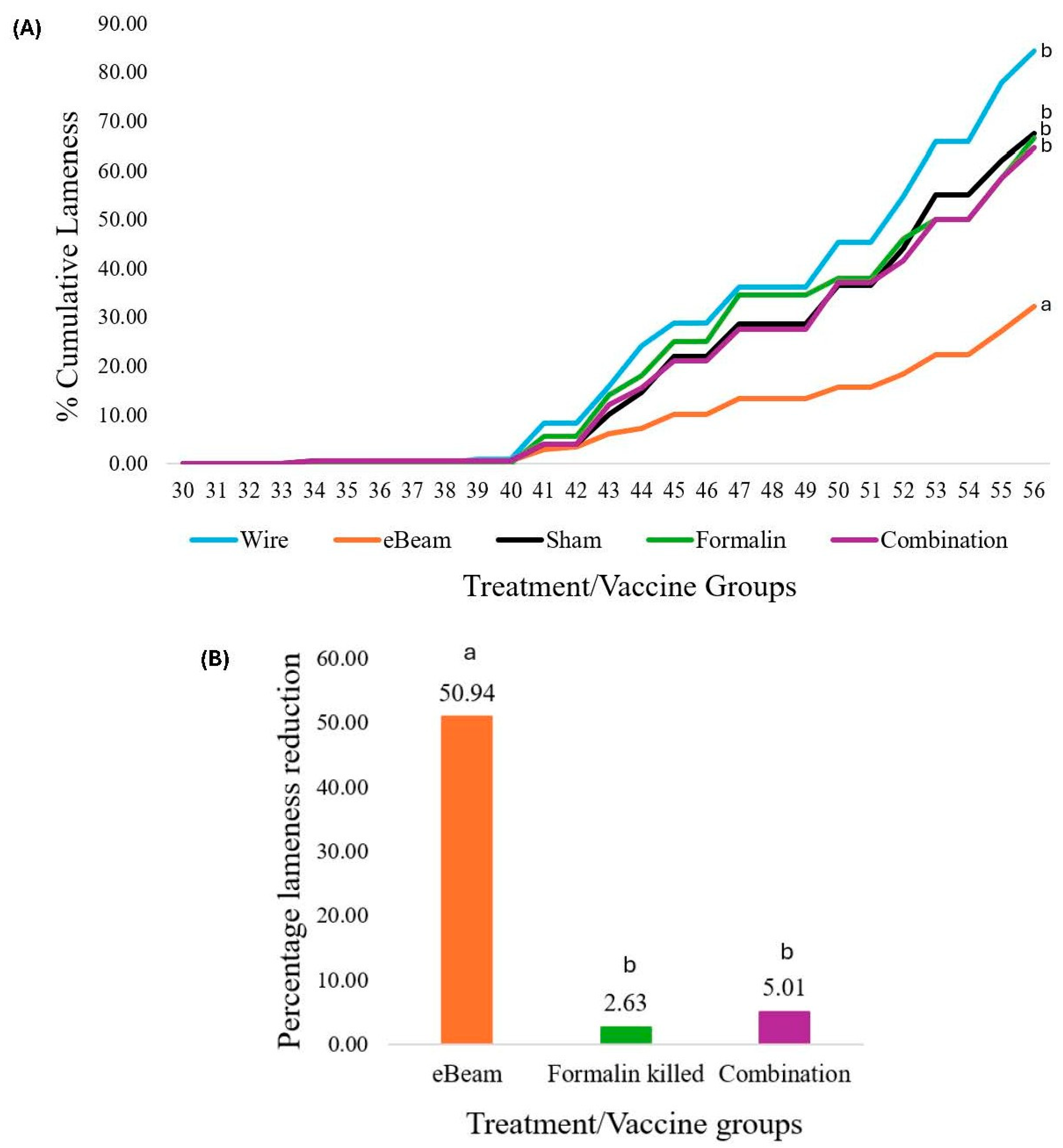
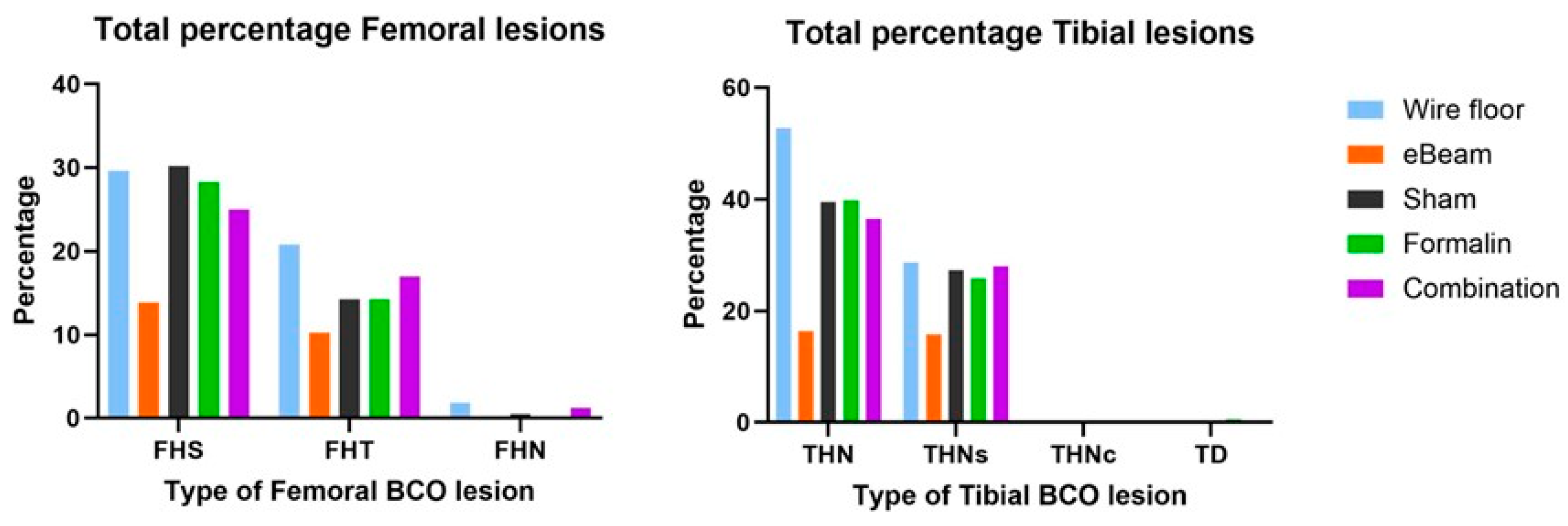
The birds that received the eBeam-inactivated Staphylococcus vaccine had a significant 50.94% reduction (P<0.05) in daily cumulative lameness compared to the birds that received the sham vaccine and were raised on a litter floor (Figure 5B). The reduction of lameness in birds that received formalin-inactivated and combination vaccines was not significant compared to the control. Moreover, numerically lower percentages of total femoral and tibial BCO lesions were recorded in lame birds of the eBeam group compared to the others (Figure 6).
Figure 6.
eBeam vaccinated birds had fewer BCO lesions. Percentages of total femoral BCO lesions (FHS: Femoral Head Separation, FHT: Femoral Head Transitional degeneration, FHN: Femoral Head Necrosis) and total tibial BCO lesions (THN: Tibial Head Necrosis, THNs: Tibial Head Necrosis severe, THNc: Tibial Head Necrosis caseous, TD: Tibial Dyschondroplasia) in lame birds across treatment groups.
Figure 6.
eBeam vaccinated birds had fewer BCO lesions. Percentages of total femoral BCO lesions (FHS: Femoral Head Separation, FHT: Femoral Head Transitional degeneration, FHN: Femoral Head Necrosis) and total tibial BCO lesions (THN: Tibial Head Necrosis, THNs: Tibial Head Necrosis severe, THNc: Tibial Head Necrosis caseous, TD: Tibial Dyschondroplasia) in lame birds across treatment groups.
Since the eBeam-inactivated multi-strain Staphylococcus vaccinated group presented significantly higher reduction of lameness and fewer BCO lesions compared to the other treatment groups, our results confirm the suitability of including Staphylococcus in the development of a vaccine against lameness, as it is among the most abundant causative agents of the disease.
3.3. Staphylococcus Was Absent in the BCO Lesions of Birds Vaccinated with the eBeam-Treated Staphylococcus Vaccine
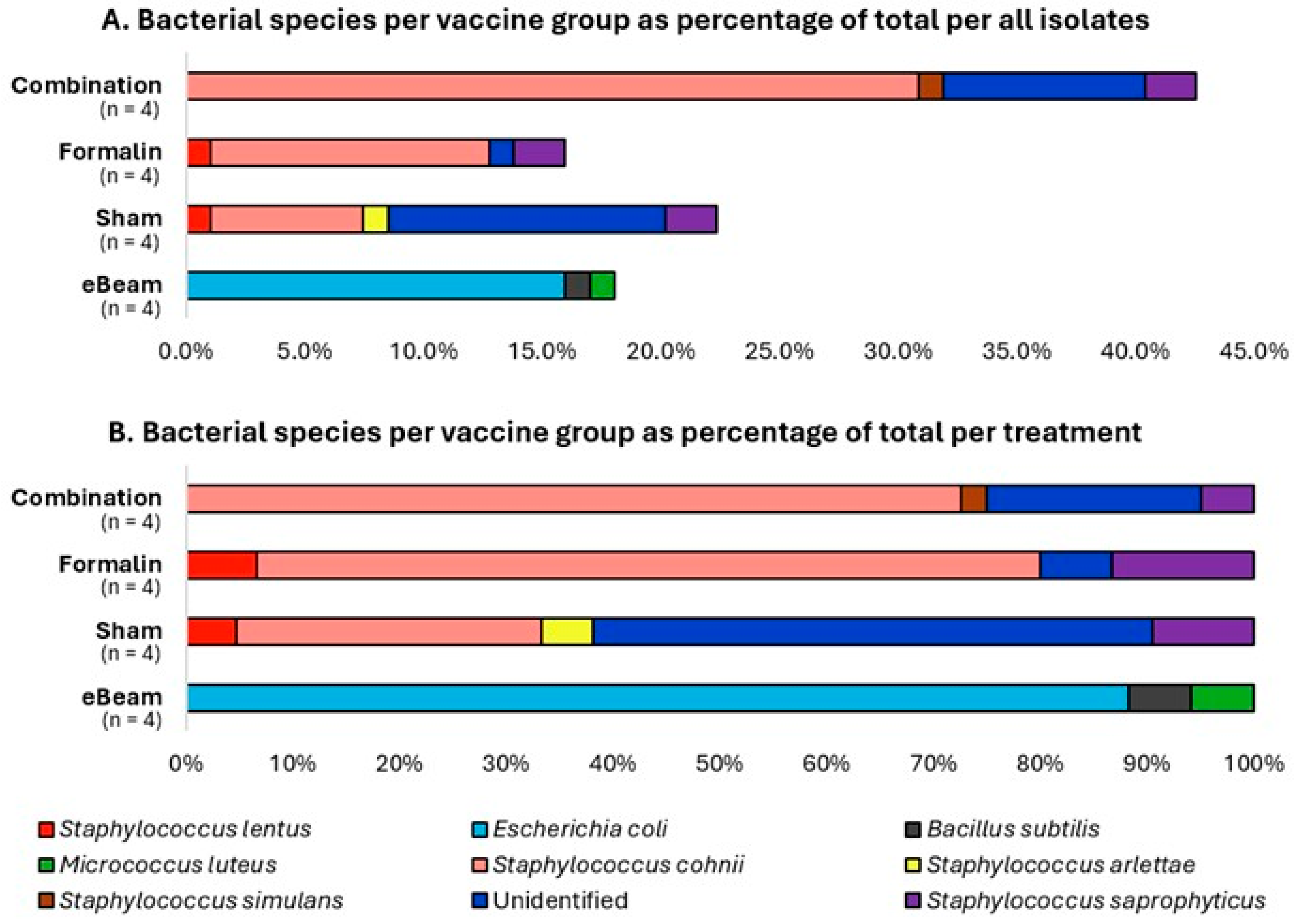
In the present study, the bacterial species recorded with the highest incidence (48.94%) in the BCO lesions of lame birds was Staphylococcus cohnii, followed by Escherichia coli (15.96%) and Staphylococcus saprophyticus (6.38%) (Figure 7A). These results are in agreement with previously obtained evidence from several studies [11,57,58] that Staphylococcus spp. and Escherichia coli are among the most frequently associated bacteria with the pathology of BCO. Additionally, the unidentified species mentioned in Figure 7 are the sum of the bacteria that grew on Agar plates initially from the BCO lesion swab samples but did not re-grow during the process of re-culturing for isolating pure cultures, plus a minority of pure-culture isolates whose DNA extraction and sequencing were unsuccessful. Most importantly, the bacterial species isolated from the lame birds of the eBeam vaccine group did not contain Staphylococcus spp., unlike the other treatments (Figure 7B). This further confirms that the eBeam vaccine not only reduced the total cumulative lameness significantly, but also effectively protected the host against the vaccine-targeted bacteria. This may be a reason behind the significant reduction in lameness, as Staphylococcus cohnii, the most abundant species in the present study, was successfully controlled in the eBeam vaccine group of birds.
Figure 7.
Staphylococcus was not identified in BCO lesions of eBeam vaccinated birds. Percentages of bacterial species identified from the BCO lesions of lame birds across treatment/vaccine groups (eBeam, sham, formalin, and combination). n represents the number of birds sampled.
Figure 7.
Staphylococcus was not identified in BCO lesions of eBeam vaccinated birds. Percentages of bacterial species identified from the BCO lesions of lame birds across treatment/vaccine groups (eBeam, sham, formalin, and combination). n represents the number of birds sampled.
3.4. Birds of the eBeam and Combination Vaccine Groups Had Higher Mucosal IgA Levels at D16 Compared to Other Groups, Suggesting Early Protection
The time-course of circulating levels of IgM, IgY, and IgA, as well as mucosal IgA, is illustrated for each treatment group in Figure 8 below.
Figure 8.
Staphylococcus-specific antibody levels changed similarly over time, increasing gradually in all treatments. Differences in blood plasma IgM (A), blood plasma IgY (B), tracheal mucous IgA (C), and blood plasma IgA (D) levels between treatment groups (sham, eBeam, formalin, and combination) on days 16, 32, and 56 of the study. Results are presented as means ± SEM; a-c: Means without a common letter within a treatment group are significantly different (P<0.05); Abbreviations: n – number of birds sampled per time point, trt – treatment (vaccine group), IxN – interactions (time x trt).
Figure 8.
Staphylococcus-specific antibody levels changed similarly over time, increasing gradually in all treatments. Differences in blood plasma IgM (A), blood plasma IgY (B), tracheal mucous IgA (C), and blood plasma IgA (D) levels between treatment groups (sham, eBeam, formalin, and combination) on days 16, 32, and 56 of the study. Results are presented as means ± SEM; a-c: Means without a common letter within a treatment group are significantly different (P<0.05); Abbreviations: n – number of birds sampled per time point, trt – treatment (vaccine group), IxN – interactions (time x trt).

While the relative levels of IgM are not significantly different between treatments at any particular time point, there is a general trend of a significant increase in IgM levels over time within all treatments. Despite changes within the early time points, D56 exhibits significantly higher IgM levels in all treatments. However, the increase in blood IgM titers from D16 to D32 is significant in the formalin and combination vaccine groups as opposed to the other treatments (Figure 8A). The time-course of blood IgY profiles (Figure 8B) closely resembles that of IgM, with no significant differences between treatments at any particular time point but with significant increments within treatments over time. However, the difference in IgY levels between D16 and D32 of any treatment is not significant. The eBeam and combination vaccine groups show numerically higher levels of IgA in tracheal mucus at D16 compared to D32, and statistically similar to D56 (Figure 8C). In contrast, the formalin and sham-vaccinated birds exhibit lower levels of IgA in tracheal mucus at D16 and D32 than at D56 (Figure 8C). Plasma IgA levels depict no significant change between treatments at any particular time point (Figure 8D). The above general trend in the change of antibody levels (of all isotypes) may be explained by the fact that all birds are continuously exposed to the BCO-causing pathogens (including Staphylococcus) via aerosol. With an increase in the overall cumulative lameness of birds in the facility over time, pathogen doses correspondingly increase, resulting in higher loads of pathogens entering the respiratory tract and crossing into the bloodstream. In response, the natural immune response may cause an elevation of antibody levels by D56 to combat the bacteria. Importantly, with exposure to the vaccine at the embryonic stage, the B lymphocytes of effectively vaccinated birds are expected to undergo affinity maturation and isotype switching to IgY and IgA isotypes. The eBeam and combination vaccine groups had higher mucosal IgA levels at D16 compared to the formalin and sham groups, which may be due to being immunized with antigens of higher quality (with undamaged epitopes) from the former vaccines. This may cause isotype switching to happen sooner, which in turn would increase in response to incoming pathogens early on. Furthermore, as BCO pathogens infect birds via aerosol in the present model, mucosal IgA of the respiratory tract would be the primary antibody to combat the invading pathogen. Higher mucosal IgA levels early in the time course suggest better protection starting at an early stage via eBeam and combination vaccines.
3.5. Leucocyte Populations Showed Diverse Trends of Concentration Changes Between Treatment Groups
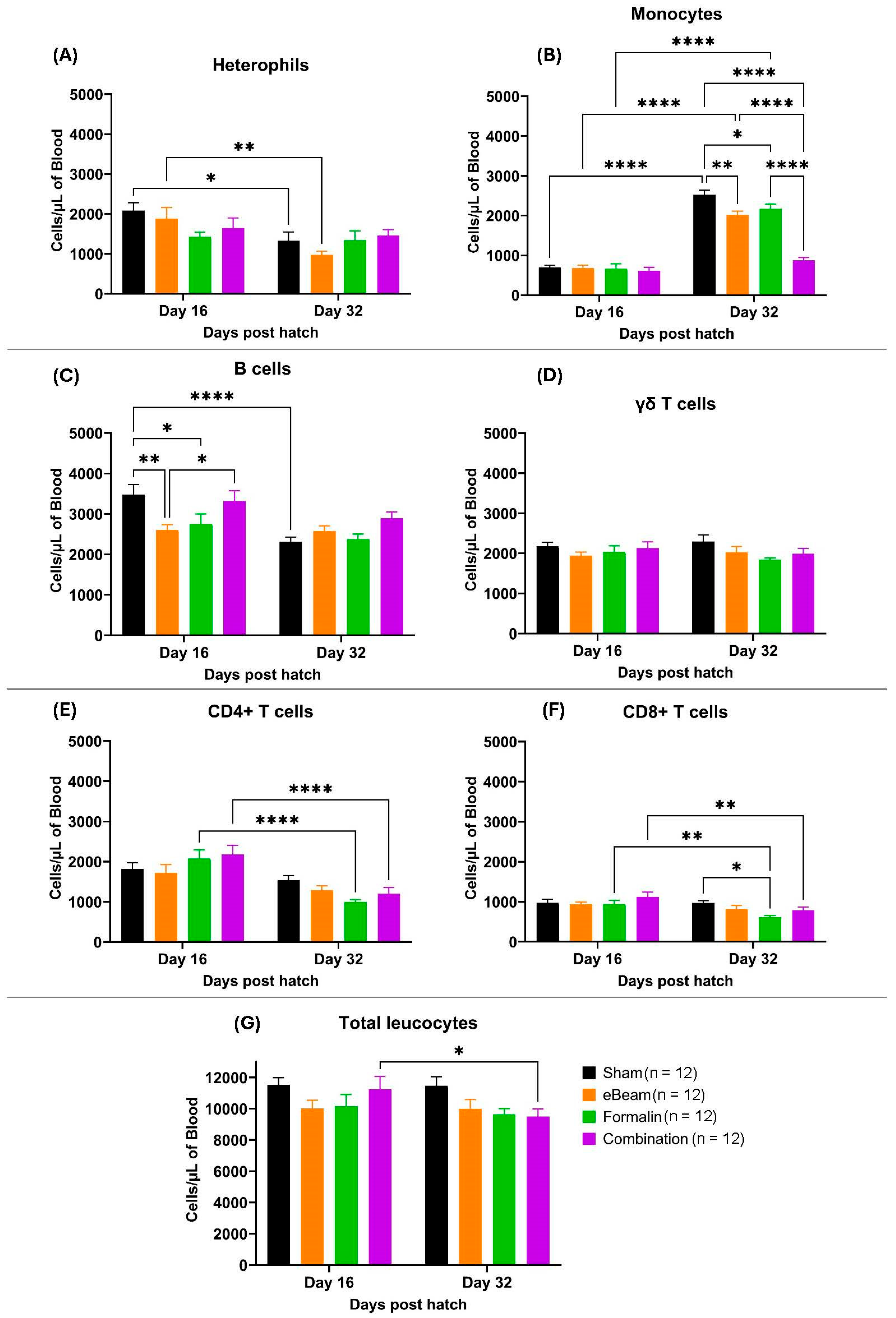
Blood cell population levels exhibit significant temporal variation before BCO clinical symptoms (D16), to when birds typically began to show symptoms of lameness (D32). The eBeam group exhibits the most significant drop in heterophil levels (cells/µL) from D16 to D32 (Figure 9A). Monocytes show a significant increase from D16 to D32 in the sham, eBeam, and formalin vaccine groups (Figure 9B), which may be due to increased production to combat the invading pathogen. However, the increase in monocyte levels from D16 to 32 in the combination group is not significant, which may be due to either lower monocyte production in the combination-vaccinated birds or early recruitment from the blood to the site of infection. The eBeam group shows significantly lower levels of B cells on day 16 compared to other treatments. B cells show a general trend of decrease from D16 to D32 in all treatments, except in the eBeam vaccine group (Figure 9C). γδ T lymphocytes (Figure 9D) and total leucocytes (Figure 9G) show almost no significant variation over time in all treatments. Significant reductions of CD4+ (Figure 9E) and CD8+ T lymphocyte (Figure 9F) levels from D16 to D32 are prominent in the formalin and combination vaccine groups. Overall, all four treatment groups presented similar blood cell population distributions at both time points. However, there was a significant increase in monocytes in the bloodstream of the birds between days 16 and 32, suggesting an immune response against an ongoing infection.
Figure 9.
Concentration (cells/µL of blood) of heterophils (A), monocytes (B), B lymphocytes (C), γδ T lymphocytes (D), CD4+ T lymphocytes (E), CD8+ T lymphocytes (F), and total leucocytes (G) over sampling days (16 and 32) in differently vaccinated groups of broilers (sham, eBeam, formalin and combination). Graphs show mean ± SEM. P value; *= <0.05, **= <0.01, ****= <0.0001. n represents the number of birds sampled per time point.
Figure 9.
Concentration (cells/µL of blood) of heterophils (A), monocytes (B), B lymphocytes (C), γδ T lymphocytes (D), CD4+ T lymphocytes (E), CD8+ T lymphocytes (F), and total leucocytes (G) over sampling days (16 and 32) in differently vaccinated groups of broilers (sham, eBeam, formalin and combination). Graphs show mean ± SEM. P value; *= <0.05, **= <0.01, ****= <0.0001. n represents the number of birds sampled per time point.
4. Discussion
Bacterial Chondronecrosis with Osteomyelitis (BCO) is a primary concern currently faced by the poultry industry [22,59,60]. While the etiology and pathogenesis of the disease are incompletely understood [24,25,61], it is known to be of opportunistic bacterial origin and remains the leading cause of lameness [62,63] in broiler chickens, evidently warranting investigation into control and prevention. The aerosol transmission model developed by our lab has proven successful in assessing treatment effects in the prevention of lameness. This success greatly lies in the model’s ability to rapidly induce lameness in the birds raised on wire floor, followed by successful transfer to the birds raised on litter floor. The present study achieved approximately 85% lameness (percentage of lame birds among the total birds) in wire floor pens, followed by approximately 68% lameness in litter floor pens of the control (sham) group. This demonstrates a numerical increment of roughly 25% lameness in wire floor pens compared to the sham pens, representing the success of the infection source. Simultaneously, the difference is statistically insignificant (P > 0.05), indicating the success of lameness transfer by the current model, as demonstrated by previous trials in our lab [21,23,59,60,64].
Amidst an established stressful, lameness-stimulatory environment, the birds vaccinated with eBeam-inactivated Staphylococcus achieved a significant reduction of approximately 51% of lameness compared to the Sham group. In contrast, the formalin and combination vaccine groups presented low lameness reductions, with a similar prevalence of the disease as the control group. This success may be attributable to the retention of membrane integrity [19,37,40] and viability [41,42] of eBeam-inactivated bacteria, symbolizing a live vaccine, as opposed to cross-linking of membrane proteins [65,66,67,68] causing significant loss of membrane integrity (Figure 4) and viability (Figure 3) in formalin-inactivated bacteria, favoring better immune response against Staphylococcus in birds vaccinated with the eBeam-inactivated Staphylococcus vaccine. Moreover, the present study surpasses the lameness reduction percentages reported in other studies [17,69,70] utilizing feed additives. In addition to the success in reducing lameness, the cost and labor efficiency (one-time in ovo vaccination versus mixing feed additives with the diet and feeding birds over the entire production period) of an eBeam-inactivated vaccine makes it a potential alternative to feed additives. According to the current understanding of the pathogenicity of BCO, the frequent causative agents among a variety of other bacterial species are Staphylococcus spp., Escherichia coli, and Enterococcus spp. [11,21,57,58,71,72]. This trend is represented in the present study through the identification of Staphylococcus cohnii as the most abundant species (approximately 49%), followed by Escherichia coli (approximately 16%) as the second-most abundant bacterial species (Figure 7), isolated from the BCO lesions of lame birds. Interestingly though, birds of other treatment groups (except eBeam) contributed to the abundant discovery of S. cohnii in BCO lesions, alongside other Staphylococcus species (S. simulans, S. arlettae, S. saprophyticus, and S. lentus). eBeam vaccinated birds showed Escherichia coli in high abundance alongside Micrococcus luteus and Bacillus subtilis isolated in smaller percentages. While this certainly demonstrates the success of the eBeam vaccine in restricting Staphylococcus cohnii colonization of BCO lesions, and suggests possible cross-protection against other Staphylococcus spp., which were not detected in birds of the eBeam group, but were present in birds of the other groups. Furthermore, the fact that Staphylococcus dominated BCO lesions of the rest of the birds, as opposed to E. coli's domination in the eBeam-inactivated Staphylococcus-vaccinated birds, suggests the possible over-competition of Staphylococcus over E. coli. However, this also provides insight into the possibility that the inclusion of more bacterial species associated with the pathogenesis of BCO in an eBeam-inactivated vaccine would give even greater protection against a broad range of pathogens.
Based on previous experience, the visual onset of clinical lameness in broilers was initiated around D25-D35 [13,22,23,59,60,64,73] of the experimental model, and it occurred at D34 in the present study. The sampling time points were decided to cover periods before, during, and after the onset of visual clinical lameness. Besides the overall fluctuations in IgM (Figure 8A), IgY (Figure 8B), and IgA (Figure 8C,D) across the sampling timepoints in this study, no significant differences between treatment groups were observed at any given time-point. A possible explanation may be the absence of an actual ‘negative’ control in this study design, which does not include a group of birds entirely unexposed to the pathogen. Hence, a true background (control) level of antibodies to compare the treatment responses was not identified. The differences obtained may have been significant if compared with a truly negative population. Yet, the eBeam vaccine’s success in significantly reducing lameness compared to all other groups and the absence of Staphylococcus in BCO lesions of birds receiving the eBeam vaccine is undeniable. Despite the relative levels of antibody production by the eBeam vaccine being similar to other groups, our results suggest that the antibodies produced by eBeam-vaccinated birds are more efficient at preventing bacterial translocation, possibly due to their specificity to better-suited and/or a greater variety of epitopes to eliminate the bacteria effectively. It could be argued that the greater lameness reduction observed in the eBeam-vaccinated birds is due to a more effective stimulation of cell-mediated immunity, targeting intracellular Staphylococcus at the site of infection. This, in addition to the observed high-quality, Staphylococcus-specific antibody production would support a more efficient and targeted bacterial clearance, and protection from further infection. Although the combination vaccine group showed similar mucosal IgA and IgY responses to the eBeam vaccine, the overall protection in terms of lameness reduction is significantly lower. A possible explanation is that the combination vaccine was prepared with half of the dose of each vaccine, which may not be concentrated enough to generate the effective response seen with the full dose of the eBeam vaccine alone.
Leucocyte and antibody recruitment in response to cytokines and chemokines released by infected tissues typically occurs within hours of the onset of infection [74,75]. Furthermore, significant increases in plasma antibody profiles in eBeam-vaccinated birds compared to controls have been recorded when sampled at shorter time intervals (<16 days) between challenge and sample collection for other bacterial species [19,37]. Moreover, other studies have shown that by the 10th day post-challenge, Staphylococcus aureus mediated significant cytomorphological changes in thymus, liver, and spleen tissues were recorded in the control group chickens, compared to those that received treatments against Staphylococcus aureus [76]. Based on these findings, significant differences in antibody and blood cell population profiles between the eBeam group and other groups may have been detected prior to day 16 in the present study. Clarification of this requires further research, and it would be advisable to sample blood and tracheal mucus at earlier time points in future studies, as the onset of infection in the aerosol transmission model is inconspicuous (as opposed to providing bacterial challenge via oral gavage, drinking water, etc.). Identifying immune responses against Staphylococcus can be challenging due to its numerous adaptations to evade and misguide host immune responses. For example, their polysaccharide capsules interfere with complement-mediated phagocytosis by masking the antigens to which host-complement factors are bound [28]. Furthermore, certain strains possess anti-phagocytic properties [29] and produce exotoxins that act as superantigens, resulting in the polyclonal activation of T cells (independent of antigen-specificity) and the release of proinflammatory cytokines by T lymphocytes, thereby rendering the host vulnerable and misleading their immune system [30]. Protein A produced by S. aureus prevents antibody-mediated phagocytosis in mammals, by binding to the Fc regions of IgG, thereby preventing Fc-receptor-mediated phagocytosis of antibody-opsonized antigen [31], although it should be noted that in chickens, S. aureus protein A does not bind to IgY (chicken IgG-like immunoglobulin isotype) [77] .
Additionally, the D10 value is the dose of radiation required to achieve a 90% reduction of a microbial population [78,79]. Recent studies have demonstrated that the D10 value for inactivating Staphylococcus aureus with eBeam is approximately 0.5 kGy [80,81]. A further understanding of the D10 values of multiple Staphylococcus strains would assist in the precise optimization of the eBeam-inactivation-dose, thereby minimizing damage to membrane epitopes and maximizing viability while achieving non-reproducibility, which would increase the protection of the vaccine. Expanding this knowledge to other bacterial genera involved in the pathogenesis of BCO would facilitate the production of an eBeam-inactivated vaccine containing multiple genera of BCO causative bacteria, which may provide better protection against BCO.
5. Conclusions
Bacterial Chondronecrosis with Osteomyelitis is currently among the leading causes of lameness in broiler chickens, resulting in reduced animal welfare and significant financial losses to the poultry industry. This condition is frequently driven by multiple bacterial species, but mainly Staphylococcus species infection, which leads to the necrosis of long leg bones. The electron beam-inactivated, multi-strain Staphylococcus vaccine tested in the present study significantly reduced BCO lameness in broiler chickens and successfully reduced Staphylococcus colonization of BCO lesions on the femurs and tibiae. This study provides insight into possible means of achieving stronger protection against BCO lameness via further optimization of the vaccine.
6. Patents
University of Arkansas: Patent Pending # PCT/US25/34205: “Electron Beam (eBeam)-Killed Multi-Bacterial Vaccines”.
Author Contributions
Conceptualization, methodology, investigation, software, formal analysis, data curation, writing —original draft, R.P.; methodology, investigation, writing—review and editing, A.A.; methodology, investigation, writing—review and editing, K.A.; methodology, investigation, software, A.D.; methodology, investigation, writing—review and editing, M.B.L; methodology, investigation, writing—review and editing, A.P.A.; conceptualization, validation, supervision, writing—review and editing, A.L.F.V.A.; conceptualization, investigation, writing—review and editing, K.A.; conceptualization, investigation, writing—review and editing, G.K.P.; methodology, investigation, software, J.M.S.; validation, supervision, writing—review and editing, G.F.E.; methodology, investigation, T.K.; resources, writing—review and editing, S.D.P.; conceptualization, methodology, investigation, supervision, resources, writing—review and editing, P.J.; conceptualization, methodology, investigation, project administration, funding acquisition, writing—review and editing, A.A.K.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the US Poultry and Egg Association (Tucker, GA, USA), E. Ford Foundation, grant number AWD-103128.
Institutional Review Board Statement
All procedures and protocols were approved by the University of Arkansas System, Division of Agriculture, Institutional Animal Care and Use Committee (IACUC), Protocol # 23058.
Informed Consent Statement
Not applicable.
Data Availability Statement
Raw data supporting the conclusions of this manuscript will be available upon request.
Acknowledgments
Our utmost appreciation is extended to the honor’s undergraduate students of Dr. Alrubaye’s research group (Hannah McCarver, Kara Hoover, Alexa Thomas, Wesley Vaught, and Dhruvkumar Patel) and the staff of the Poultry Research Farm, University of Arkansas, Fayetteville for their tremendous technical support, which was invaluable for the successful completion of the project. .
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Which are the world’s 10 largest chicken companies? https://www.wattagnet.com/broilers-turkeys/broilers/article/15660706/which-are-the-worlds-10-largest-chicken-companies (accessed on 11 July 2024).
- National Chicken Council. Top Broiler Producing States https://www.nationalchickencouncil.org/industry/broiler-industry-today/ (accessed on 11 July 2024).
- Wideman, R.F. Bacterial Chondronecrosis with Osteomyelitis and Lameness in Broilers: A Review. Poult Sci 2016, 95, 325–344. [Google Scholar] [CrossRef]
- Siegel, P.B.; Barger, K.; Siewerdt, F. Limb Health in Broiler Breeding: History Using Genetics to Improve Welfare. Journal of Applied Poultry Research 2019, 28, 785–790. [Google Scholar] [CrossRef]
- Thorp, B.H. Skeletal Disorders in the Fowl: A Review. Avian Pathology 1994, 23, 203–236. [Google Scholar] [CrossRef] [PubMed]
- Granquist, E.G.; Vasdal, G.; De Jong, I.C.; Moe, R.O. Lameness and Its Relationship with Health and Production Measures in Broiler Chickens. Animal 2019, 13, 2365–2372. [Google Scholar] [CrossRef] [PubMed]
- Szafraniec, G.M.; Szeleszczuk, P.; Dolka, B. Review on Skeletal Disorders Caused by Staphylococcus Spp. in Poultry. Veterinary Quarterly 2022, 42, 21–40. [Google Scholar] [CrossRef]
- McNamee, P.T.; Smyth, J.A. Bacterial Chondronecrosis with Osteomyelitis ('femoral Head Necrosis’) of Broiler Chickens: A Review. Avian Pathology 2000, 29, 477–495. [Google Scholar] [CrossRef]
- Nairn, M.E.; Watson, A.R.A. Leg Weakness of Poultry—A Clinical and Pathological Characterisation. Aust Vet J 1972, 48, 645–656. [Google Scholar] [CrossRef]
- Wideman Jr, R.F.; Prisby, R.D. Bone Circulatory Disturbances in the Development of Spontaneous Bacterial Chondronecrosis with Osteomyelitis: A Translational Model for the Pathogenesis of Femoral Head Necrosis. Front Endocrinol (Lausanne) 2013, 3, 183. [Google Scholar] [CrossRef]
- Al-Rubaye, A.A.K.; Couger, M.B.; Ojha, S.; Pummill, J.F.; Koon, J.A.; Wideman Jr, R.F.; Rhoads, D.D. Genome Analysis of Staphylococcus Agnetis, an Agent of Lameness in Broiler Chickens. PLoS One 2015, 10, e0143336. [Google Scholar] [CrossRef]
- Ramser, A.; Greene, E.; Alrubaye, A.A.K.; Wideman, R.; Dridi, S. Role of Autophagy Machinery Dysregulation in Bacterial Chondronecrosis with Osteomyelitis (BCO): AUTOPHAGY & BROILER LAMENESS. Poult Sci 2022, 101750. [Google Scholar] [CrossRef]
- Alrubaye, A.A.K.; Ekesi, N.S.; Hasan, A.; Elkins, E.; Ojha, S.; Zaki, S.; Dridi, S.; Wideman, R.F.; Rebollo, M.A.; Rhoads, D.D. Chondronecrosis with Osteomyelitis in Broilers: Further Defining Lameness-Inducing Models with Wire or Litter Flooring to Evaluate Protection with Organic Trace Minerals. Poult Sci 2020, 99, 5422–5429. [Google Scholar] [CrossRef] [PubMed]
- Junior, A.M.B.; Fernandes, N.L.M.; Snak, A.; Fireman, A.; Horn, D.; Fernandes, J.I.M. Arginine and Manganese Supplementation on the Immune Competence of Broilers Immune Stimulated with Vaccine against Salmonella Enteritidis. Poult Sci 2019, 98, 2160–2168. [Google Scholar]
- Kidd, M.T. Nutritional Modulation of Immune Function in Broilers. Poult Sci 2004, 83, 650–657. [Google Scholar] [CrossRef] [PubMed]
- McKnight, L.L.; Page, G.; Han, Y. Effect of Replacing In-Feed Antibiotics with Synergistic Organic Acids, with or without Trace Mineral and/or Water Acidification, on Growth Performance and Health of Broiler Chickens under a Clostridium Perfringens Type A Challenge. Avian Dis 2020, 64, 374–378. [Google Scholar] [CrossRef]
- Wideman, R.F.; Al-Rubaye, A.; Kwon, Y.M.; Blankenship, J.; Lester, H.; Mitchell, K.N.; Pevzner, I.Y.; Lohrmann, T.; Schleifer, J. Prophylactic Administration of a Combined Prebiotic and Probiotic, or Therapeutic Administration of Enrofloxacin, to Reduce the Incidence of Bacterial Chondronecrosis with Osteomyelitis in Broilers. Poult Sci 2015, 94, 25–36. [Google Scholar] [CrossRef]
- Wideman, R.F.; Hamal, K.R.; Stark, J.M.; Blankenship, J.; Lester, H.; Mitchell, K.N.; Lorenzoni, G.; Pevzner, I. A Wire-Flooring Model for Inducing Lameness in Broilers: Evaluation of Probiotics as a Prophylactic Treatment. Poult Sci 2012, 91, 870–883. [Google Scholar] [CrossRef]
- Jesudhasan, P.R.; Bhatia, S.S.; Sivakumar, K.K.; Praveen, C.; Genovese, K.J.; He, H.L.; Droleskey, R.; McReynolds, J.L.; Byrd, J.A.; Swaggerty, C.L. Controlling the Colonization of Clostridium Perfringens in Broiler Chickens by an Electron-Beam-Killed Vaccine. Animals 2021, 11, 671. [Google Scholar] [CrossRef]
- Thompson, D.R.; Parreira, V.R.; Kulkarni, R.R.; Prescott, J.F. Live Attenuated Vaccine-Based Control of Necrotic Enteritis of Broiler Chickens. Vet Microbiol 2006, 113, 25–34. [Google Scholar] [CrossRef]
- Asnayanti, A.; Do, A.D.T.; Alharbi, K.; Alrubaye, A. Inducing Experimental Bacterial Chondronecrosis with Osteomyelitis Lameness in Broiler Chickens Using Aerosol Transmission Model. Poult Sci 2024, 103, 103460. [Google Scholar] [CrossRef]
- Asnayanti, A.; Hasan, A.; Alharbi, K.; Hassan, I.; Bottje, W.; Rochell, S.J.; Rebollo, M.A.; Kidd, M.T.; Alrubaye, A.A.K. Assessing the Impact of Spirulina Platensis and Organic Trace Minerals on the Incidence of Bacterial Chondronecrosis with Osteomyelitis Lameness in Broilers Using an Aerosol Transmission Model. Journal of Applied Poultry Research 2024, 100426. [Google Scholar] [CrossRef]
- Asnayanti, A.; Alharbi, K.; Do, A.D.T.; Al-Mitib, L.; Bühler, K.; Van der Klis, J.D.; Gonzalez, J.; Kidd, M.T.; Alrubaye, A.A.K. Early 1, 25-Dihydroxyvitamin D3-Glycosides Supplementation: An Efficient Feeding Strategy against Bacterial Chondronecrosis with Osteomyelitis Lameness in Broilers Assessed by Using an Aerosol Transmission Model. Journal of Applied Poultry Research 2024, 33, 100440. [Google Scholar] [CrossRef]
- Ekesi, N.S. Examining Pathogenesis and Preventatives in Spontaneous and Staphylococcus-Induced Bacterial Chondronecrosis with Osteomyelitis in Broilers; University of Arkansas, 2020.
- Wijesurendra, D.S.; Chamings, A.N.; Bushell, R.N.; Rourke, D.O.; Stevenson, M.; Marenda, M.S.; Noormohammadi, A.H.; Stent, A. Pathological and Microbiological Investigations into Cases of Bacterial Chondronecrosis and Osteomyelitis in Broiler Poultry. Avian pathology 2017, 46, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Choppa, V.S.R.; Kim, W.K. A Review on Pathophysiology, and Molecular Mechanisms of Bacterial Chondronecrosis and Osteomyelitis in Commercial Broilers. Biomolecules 2023, 13, 1032. [Google Scholar] [CrossRef] [PubMed]
- Ferver, A.; Dridi, S. Bacterial Chondronecrosis with Osteomyelitis (BCO) in Modern Broilers: Impacts, Mechanisms, and Perspectives. CABI Reviews, 2020. [Google Scholar]
- Peterson, P.K.; Wilkinson, B.J.; Kim, Y.; Schmeling, D.; Douglas, S.D.; Quie, P.G.; Verhoef, J. The Key Role of Peptidoglycan in the Opsonization of Staphylococcus Aureus. J Clin Invest 1978, 61, 597–609. [Google Scholar] [CrossRef]
- Watson, D.L. Vaccination against Experimental Staphylococcal Mastitis in Dairy Heifers. Res Vet Sci 1992, 53, 346–353. [Google Scholar] [CrossRef]
- Shinefield, H.R.; Black, S. Prevention of Staphylococcus Aureus Infections: Advances in Vaccine Development. Expert Rev Vaccines 2005, 4, 669–676. [Google Scholar] [CrossRef]
- Prescott, J.F.; MacInnes, J.I.; Van Immerseel, F.; Boyce, J.D.; Rycroft, A.N.; Vázquez-Boland, J.A. Pathogenesis of Bacterial Infections in Animals; Wiley Online Library, 2022.
- Walker, R.I. Considerations for Development of Whole Cell Bacterial Vaccines to Prevent Diarrheal Diseases in Children in Developing Countries. Vaccine 2005, 23, 3369–3385. [Google Scholar] [CrossRef]
- Osterloh, A. Vaccination against Bacterial Infections: Challenges, Progress, and New Approaches with a Focus on Intracellular Bacteria. Vaccines (Basel) 2022, 10, 751. [Google Scholar] [CrossRef]
- Ellis, R.W.; Rappuoli, R.; Ahmed, S. Technologies for Making New Vaccines. In Vaccines; Elsevier, 2013; pp 1182–1199.
- Strugnell, R.; Zepp, F.; Cunningham, A.; Tantawichien, T. Vaccine Antigens. Perspect Vaccinol 2011, 1, 61–88. [Google Scholar] [CrossRef]
- Vetter, V.; Denizer, G.; Friedland, L.R.; Krishnan, J.; Shapiro, M. Understanding Modern-Day Vaccines: What You Need to Know. Ann Med 2018, 50, 110–120. [Google Scholar] [CrossRef]
- Jesudhasan, P.R.; McReynolds, J.L.; Byrd, A.J.; He, H.; Genovese, K.J.; Droleskey, R.; Swaggerty, C.L.; Kogut, M.H.; Duke, S.; Nisbet, D.J. Electron-Beam–Inactivated Vaccine against Salmonella Enteritidis Colonization in Molting Hens. Avian Dis 2015, 59, 165–170. [Google Scholar] [CrossRef]
- Bhatia, S.S.; Pillai, S.D. Ionizing Radiation Technologies for Vaccine Development-A Mini Review. Front Immunol 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.; Pillai, S.D.; Alrubaye, A.; Jesudhasan, P. Leveraging Electron Beam (EBeam) Technology for Advancing the Development of Inactivated Vaccines. Vaccines (Basel) 2025, 13, 179. [Google Scholar] [CrossRef] [PubMed]
- Hieke, A.-S. C.; Pillai, S.D. Escherichia Coli Cells Exposed to Lethal Doses of Electron Beam Irradiation Retain Their Ability to Propagate Bacteriophages and Are Metabolically Active. Front Microbiol 2018, 2138. [Google Scholar] [CrossRef] [PubMed]
- Praveen, C. Electron Beam as a next Generation Vaccine Platform: Microbiological and Immunological Characterization of an Electron Beam Based Vaccine against Salmonella Typhimurium; Texas A&M University, 2014.
- Praveen, C.; Bhatia, S.S.; Alaniz, R.C.; Droleskey, R.E.; Cohen, N.D.; Jesudhasan, P.R.; Pillai, S.D. Assessment of Microbiological Correlates and Immunostimulatory Potential of Electron Beam Inactivated Metabolically Active yet Non Culturable (MAyNC) Salmonella Typhimurium. PLoS One 2021, 16, e0243417. [Google Scholar] [CrossRef]
- Liu, Y.; Shao, Y.; Wang, L.; Lu, W.; Li, S.; Xu, D.; Fu, Y.V. Inactivation of Porcine Epidemic Diarrhea Virus with Electron Beam Irradiation under Cold Chain Conditions. Environ Technol Innov 2022, 27, 102715. [Google Scholar] [CrossRef]
- Tahergorabi, R.; Matak, K.E.; Jaczynski, J. Application of Electron Beam to Inactivate Salmonella in Food: Recent Developments. Food Research International 2012, 45, 685–694. [Google Scholar] [CrossRef]
- Sabbaghi, A.; Miri, S.M.; Keshavarz, M.; Zargar, M.; Ghaemi, A. Inactivation Methods for Whole Influenza Vaccine Production. Rev Med Virol 2019, 29, e2074. [Google Scholar] [CrossRef]
- Miller, R.B. Introduction to Food Irradiation. Electronic irradiation of foods: an introduction to the technology, 2005; 1–15. [Google Scholar]
- Assumpcao, A.L.F. V.; Arsi, K.; Asnayanti, A.; Alharbi, K.S.; Do, A.D.T.; Read, Q.D.; Perera, R.; Shwani, A.; Hasan, A.; Pillai, S.D. Electron-Beam-Killed Staphylococcus Vaccine Reduced Lameness in Broiler Chickens. Vaccines (Basel) 2024, 12, 1203. [Google Scholar] [CrossRef]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler Jr, V.G. Staphylococcus Aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin Microbiol Rev 2015, 28, 603–661. [Google Scholar] [CrossRef]
- Abd El Tawab, A.A.; El-Hofy, F.I.; Maarouf, A.A.; El-Said, A.A. Bacteriological Studies on Some Food Borne Bacteria Isolated from Chicken Meat and Meat Products in Kaliobia Governorate. Benha Vet Med J 2015, 29, 47–59. [Google Scholar] [CrossRef]
- Kaminski, R.W.; Wu, M.; Turbyfill, K.R.; Clarkson, K.; Tai, B.; Bourgeois, A.L.; Van De Verg, L.L.; Walker, R.I.; Oaks, E.V. Development and Preclinical Evaluation of a Trivalent, Formalin-Inactivated Shigella Whole-Cell Vaccine. Clinical and Vaccine Immunology 2014, 21, 366–382. [Google Scholar] [CrossRef]
- FDA. BAM R11: Butterfield’s Phosphate-Buffered Dilution Water https://www.fda.gov/food/laboratory-methods-food/bam-r11-butterfields-phosphate-buffered-dilution-water (accessed Jun 25, 2024).
- Al-Rubaye, A.A.K.; Ekesi, N.S.; Zaki, S.; Emami, N.K.; Wideman, R.F.; Rhoads, D.D. Chondronecrosis with Osteomyelitis in Broilers: Further Defining a Bacterial Challenge Model Using the Wire Flooring Model. Poult Sci 2017, 96, 332–340. [Google Scholar] [CrossRef]
- Davis, M.W.; Jorgensen, E.M. ApE, a Plasmid Editor: A Freely Available DNA Manipulation and Visualization Program. Frontiers in Bioinformatics 2022, 2, 818619. [Google Scholar] [CrossRef] [PubMed]
- Seliger, C.; Schaerer, B.; Kohn, M.; Pendl, H.; Weigend, S.; Kaspers, B.; Härtle, S. A Rapid High-Precision Flow Cytometry Based Technique for Total White Blood Cell Counting in Chickens. Vet Immunol Immunopathol 2012, 145, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.; Assumpcao, A.L.F. V.; Arsi, K.; Erf, G.F.; Donoghue, A.; Jesudhasan, P.R.R. Effect of Salmonella Typhimurium Colonization on Microbiota Maturation and Blood Leukocyte Populations in Broiler Chickens. Animals 2022, 12, 2867. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhao, Y.; Gan, H.; Hawkins, S.; Eckelkamp, L.; Prado, M.; Burns, R.; Purswell, J.; Tabler, T. Modeling Gait Score of Broiler Chicken via Production and Behavioral Data. animal 2023, 17, 100692. [Google Scholar] [CrossRef]
- Ekesi, N.S.; Dolka, B.; Alrubaye, A.A.K.; Rhoads, D.D. Analysis of Genomes of Bacterial Isolates from Lameness Outbreaks in Broilers. Poult Sci 2021, 100, 101148. [Google Scholar] [CrossRef]
- Shwani, A.; Adkins, P.R.F.; Ekesi, N.S.; Alrubaye, A.; Calcutt, M.J.; Middleton, J.R.; Rhoads, D.D. Whole-Genome Comparisons of Staphylococcus Agnetis Isolates from Cattle and Chickens. Appl Environ Microbiol 2020, 86, e00484–20. [Google Scholar] [CrossRef]
- Alharbi, K.; Asnayanti, A.; Do, A.D.T.; Perera, R.; Al-Mitib, L.; Shwani, A.; Rebollo, M.A.; Kidd, M.T.; Alrubaye, A.A.K. Identifying Dietary Timing of Organic Trace Minerals to Reduce the Incidence of Osteomyelitis Lameness in Broiler Chickens Using the Aerosol Transmission Model. Animals 2024, 14, 1526. [Google Scholar] [CrossRef]
- Do, A.D.T.; Anthney, A.; Alharbi, K.; Asnayanti, A.; Meuter, A.; Alrubaye, A.A.K. Assessing the Impact of Spraying an Enterococcus Faecium-Based Probiotic on Day-Old Broiler Chicks at Hatch on the Incidence of Bacterial Chondronecrosis with Osteomyelitis Lameness Using a Staphylococcus Challenge Model. Animals 2024, 14, 1369. [Google Scholar] [CrossRef] [PubMed]
- Petry, B.; Savoldi, I.R.; Ibelli, A.M.G.; Paludo, E.; de Oliveira Peixoto, J.; Jaenisch, F.R.F.; de Córdova Cucco, D.; Ledur, M.C. New Genes Involved in the Bacterial Chondronecrosis with Osteomyelitis in Commercial Broilers. Livest Sci 2018, 208, 33–39. [Google Scholar] [CrossRef]
- Weimer, S.L.; Wideman, R.F.; Scanes, C.G.; Mauromoustakos, A.; Christensen, K.D.; Vizzier-Thaxton, Y. The Utility of Infrared Thermography for Evaluating Lameness Attributable to Bacterial Chondronecrosis with Osteomyelitis. Poult Sci 2019, 98, 1575–1588. [Google Scholar] [CrossRef] [PubMed]
- Wideman Jr, R.F.; Al-Rubaye, A.; Gilley, A.; Reynolds, D.; Lester, H.; Yoho, D.; Hughes, J.M.; Pevzner, I. Susceptibility of 4 Commercial Broiler Crosses to Lameness Attributable to Bacterial Chondronecrosis with Osteomyelitis. Poult Sci 2013, 92, 2311–2325. [Google Scholar] [CrossRef]
- Alharbi, K.; Ekesi, N.; Hasan, A.; Asnayanti, A.; Liu, J.; Murugesan, R.; Ramirez, S.; Rochell, S.; Kidd, M.T.; Alrubaye, A. Deoxynivalenol and Fumonisin Predispose Broilers to Bacterial Chondronecrosis with Osteomyelitis Lameness. Poult Sci 2024, 103598. [Google Scholar] [CrossRef]
- Bouveret, E.; Derouiche, R.; Rigal, A.; Lloubès, R.; Lazdunski, C.; Bénédetti, H. Peptidoglycan-Associated Lipoprotein-TolB Interaction: A Possible Key to Explaining the Formation of Contact Sites between the Inner and Outer Membranes of Escherichia Coli. Journal of Biological Chemistry 1995, 270, 11071–11077. [Google Scholar] [CrossRef]
- Prossnitz, E.; Nikaido, K.; Ulbrich, S.J.; Ames, G.F. Formaldehyde and Photoactivatable Cross-Linking of the Periplasmic Binding Protein to a Membrane Component of the Histidine Transport System of Salmonella Typhimurium. Journal of biological chemistry 1988, 263, 17917–17920. [Google Scholar] [CrossRef]
- Klockenbusch, C.; O’Hara, J.E.; Kast, J. Advancing Formaldehyde Cross-Linking towards Quantitative Proteomic Applications. Anal Bioanal Chem 2012, 404, 1057–1067. [Google Scholar] [CrossRef]
- Saggers, E.J.; Waspe, C.R.; Parker, M.L.; Waldron, K.W.; Brocklehurst, T.F. Salmonella Must Be Viable in Order to Attach to the Surface of Prepared Vegetable Tissues. J Appl Microbiol 2008, 105, 1239–1245. [Google Scholar] [CrossRef]
- Tan, J.; Wideman Jr, R.F.; Blankenship, J.; Pevzner, I. Effects of a Well-Defined Multi-Species Probiotic Feed Additive on Lameness in Broiler Chickens [Conference Poster]. 2014.
- Chen, J.; Wedekind, K.J.; Dibner, J.J.; Richards, J.D. Effects of Nutrition and Gut Barrier Function on the Development of Osteomyelitis Complex and Other Forms of Lameness in Poultry. 2013.
- Mandal, R.K.; Jiang, T.; Al-Rubaye, A.A.; Rhoads, D.D.; Wideman, R.F.; Zhao, J.; Pevzner, I.; Kwon, Y.M. An Investigation into Blood Microbiota and Its Potential Association with Bacterial Chondronecrosis with Osteomyelitis (BCO) in Broilers. Sci Rep 2016, 6, 25882. [Google Scholar] [CrossRef]
- Jiang, T.; Mandal, R.K.; Wideman Jr, R.F.; Khatiwara, A.; Pevzner, I.; Min Kwon, Y. Molecular Survey of Bacterial Communities Associated with Bacterial Chondronecrosis with Osteomyelitis (BCO) in Broilers. PLoS One 2015, 10, e0124403. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.; Alharbi, K.; Hasan, A.; Asnayanti, A.; Do, A.; Shwani, A.; Murugesan, R.; Ramirez, S.; Kidd, M.; Alrubaye, A.A.K. Evaluating the Impact of the PoultryStar® Bro Probiotic on the Incidence of Bacterial Chondronecrosis with Osteomyelitis Using the Aerosol Transmission Challenge Model. Microorganisms 2024, 12, 1630. [Google Scholar] [CrossRef] [PubMed]
- Bayona, J.A.M.; Karuppannan, A.K.; Barreda, D.R. Contribution of Leukocytes to the Induction and Resolution of the Acute Inflammatory Response in Chickens. Dev Comp Immunol 2017, 74, 167–177. [Google Scholar] [CrossRef] [PubMed]
- French, C.E.; Sales, M.A.; Rochell, S.J.; Rodriguez, A.; Erf, G.F. Local and Systemic Inflammatory Responses to Lipopolysaccharide in Broilers: New Insights Using a Two-Window Approach. Poult Sci 2020, 99, 6593–6605. [Google Scholar] [CrossRef]
- Pavlova, A.V.; Pimenov, N.V.; Konstantinov, A.V.; Bordyugova, S.S. Immunomorphological Changes in Bird Staphylococcus. IOP Conf. Ser. Earth Environ. Sci. 2020, 548, 042033. [Google Scholar] [CrossRef]
- Kota, R.K.; Reddy, P.N.; Sreerama, K. Application of IgY Antibodies against Staphylococcal Protein A (SpA) of Staphylococcus Aureus for Detection and Prophylactic Functions. Appl Microbiol Biotechnol 2020, 104, 9387–9398. [Google Scholar] [CrossRef]
- Kortei, N.K.; Odamtten, G.T.; Obodai, M.; Wiafe-Kwagyan, M. Mycofloral Profile and the Radiation Sensitivity (D10 Values) of Solar Dried and Gamma Irradiated Pleurotus Ostreatus (Jacq. Ex. Fr. ) Kummer Fruitbodies Stored in Two Different Packaging Materials. Food Sci Nutr 2018, 6, 180–188. [Google Scholar]
- Jeong, S.-G.; Kang, D.-H. Inactivation of Escherichia Coli O157: H7, Salmonella Typhimurium, and Listeria Monocytogenes in Ready-to-Bake Cookie Dough by Gamma and Electron Beam Irradiation. Food Microbiol 2017, 64, 172–178. [Google Scholar] [CrossRef]
- Zheng, Q.; Chen, Z.; Yan, W.; Wang, H.; Tian, W.; Feng, D.; Yue, L.; Qi, W.; He, X.; Kong, Q. TMT-Based Quantitative Proteomic and Scanning Electron Microscopy Reveals Biological and Morphological Changes of Staphylococcus Aureus Irradiated by Electron Beam. LWT 2023, 184, 114977. [Google Scholar] [CrossRef]
- Ebrahim, H.; Abou ElNour, S.; Hammad, A.A.; Abouzeid, M.; Abdou, D. Comparative Effect of Gamma and Electron Beam Irradiation on Some Food Borne Pathogenic Bacteria Contaminating Meat Products. Egyptian Journal of Pure and Applied Science 2022, 60, 62–72. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



