
Submitted:
03 August 2025
Posted:
04 August 2025
You are already at the latest version
Abstract
Lassa fever (LF), a severe hemorrhagic disease endemic to West Africa, is primarily transmitted by rodents of the genus Mastomys, particularly Mastomys natalensis, which serve as the main reservoirs of Lassa virus (LASV). There have been reports of high preva-lence of LF in Nigeria, and outbreaks tend to be recurrent yet geographically restricted, implying that additional ecological or epidemiological factors influence the distribution of the disease beyond the mere presence of M. natalensis. However, national-scale data on LASV prevalence in rodent populations remain scarce. To address this gap, a targeted small mammals survey was conducted over a four-month period (May to August 2024) in Otukpo Local Government Area (LGA) of Benue State, north-central Nigeria. Rodents and other small mammals were trapped across three purposively selected wards identified as high-risk areas based on prior reports of occurrence of such small mammals in the areas as well the informal settlements in which the selected wards in Otukpo LGA. Analysis of the samples revealed no statistically significant variation in LASV prevalence among the study sites, indicating a relatively uniform, low-level exposure risk across the LGA and region. However, a marginally significant difference in LASV detection between plasma and serum samples suggests that sample type and storage conditions may influence se-rological sensitivity. These findings highlight the importance of refining diagnostic pro-tocols, broadening surveillance to include additional rodent hosts, and integrating eco-logical data with public health strategies to improve early warning systems and strengthen Lassa fever control efforts.
Keywords:
Non-Mastomys small mammals
; Mastomys natalensis
; Lassa virus (LASV)
; Lassa fever
; rodent reservoirs
; Nigeria
; West Africa
1. Introduction
Lassa fever (LF) is a serious acute viral hemorrhagic illness caused by the Lassa virus (LASV), an arenavirus endemic to several West African countries, including Nigeria, Sierra Leone, Guinea, and Liberia [1,2,3]. Clinically, the disease presents a wide spectrum, ranging from mild, non-specific symptoms to severe hemorrhagic manifestations often associated with multiorgan failure [4,5,6,7]. The LASV, the causative agent of LF, is a rodent-borne arenavirus and a significant zoonotic pathogen responsible for an estimated 5,000 deaths annually in West Africa [5,8]. Human infection typically results from direct or indirect contact with the urine or feces of infected rodents, primarily the multimammate rat Mastomys natalensis, which is historically recognized as the principal reservoir of LASV [9,10]. These rodents are widely distributed across sub-Saharan Africa and have also been reported in North Africa, including Morocco [11]. However, recent phylogenetic and ecological studies have challenged this single-host model, revealing that LASV is maintained by multiple Mastomys species—and potentially other rodent and small mammal hosts—thereby highlighting the complex and multifactorial nature of the virus’s transmission ecology [3,10,12,13,14]. This complexity is further compounded by the unresolved taxonomy of Mastomys, a genus that comprises at least eight distinct species, with differentiation based in part on diploid chromosome numbers of 32, 36, and 38, and this also further contribute to the challenges in accurately identifying reservoirs and understanding host-specific dynamics [15,16].
Ecologically, Mastomys species are highly adaptable, occupying a wide range of habitats including grasslands, savannahs, croplands, forest edges, and rural dwellings [17,18,19]. These rodents are nocturnal and among the most fecund of small mammals, with high reproductive rates that contribute to their ecological success. While the dynamics of natural LASV infection in Mastomys populations remain incompletely understood, and laboratory studies have provided important insights into their infection biology. For instance, juvenile Mastomys with a diploid number of 36 (2n = 36), when experimentally infected intraperitoneally within the first weeks of life, can become chronic carriers, maintaining persistent viremia and viruria [20,21]. In contrast, adult individuals typically develop only transient viremia, followed by robust protective immunity. Notably, despite persistent infection, LASV does not appear to cause overt disease in Mastomys, although histopathological changes have occasionally been documented [22]. Reproductive output in infected females also appears unaffected, as litter sizes are comparable to those of uninfected controls [21]. Hence, the epidemiological dynamics of the LF is quite complex due to these multifaceted routes of infection among varied species of small mammals and as well as sex-based (male or female) response of the virus.
Epidemiological evidence of human Lassa fever is most extensively documented in Nigeria, Liberia, Sierra Leone, and Guinea, as shown by numerous clinical and field investigations [2,7,17,23,24,25]. These countries represent the core endemic zones where the disease burden is highest, with an estimated 300,000 to 500,000 cases and approximately 5,000 deaths annually [7,26,27]. As mentioned earlier, the causative agent of the virus, belongs to the family Arenaviridae, genus Arenavirus, and is naturally harbored by the multimammate rat (Mastomys natalensis), recognized as the primary reservoir host [7,9,28,30]. Although M. natalensis is widely distributed across sub-Saharan Africa, LASV-infected populations have been predominantly identified in specific West African regions, particularly Nigeria, Sierra Leone, and Guinea [9,28,31,32] confirming the endemism of this virus in these countries, and Nigeria in particular the most populated country in the region and Africa as a whole, and coordinated efforts to curtail the continued spread of the virus.
Beyond these well-established endemic areas, antibody prevalence surveys and anecdotal reports have revealed evidence of LASV or related arenavirus circulation in other West African countries, pointing to a potentially wider distribution than currently confirmed [33,34,35,36,37]. Serological findings have also emerged from parts of Central Africa, reinforcing the need for broader surveillance and enhanced diagnostic capacity across the region [9,38]. Furthermore, sporadic imported cases of LF have been reported in Europe and North America, primarily involving travelers returning from endemic areas. These incidents underscore LASV’s significance as a globally relevant viral hemorrhagic fever and highlight the importance of international preparedness and response capabilities [38,39,40], which must be taken very serious as further spread of the virus resulting from these imported cases can translate into a global pandemic if care is not taken to curtail the spread.
In Nigeria, particularly in the north-central region where LF outbreaks are frequently reported, the role of non-Mastomys rodents in the maintenance and transmission of LASV remains largely understudied [41,42]. While Mastomys natalensis has traditionally been identified as the primary reservoir, growing evidence suggests that other small mammal species may also contribute to LASV persistence and spread. A comprehensive understanding of the full spectrum of potential host species is crucial for accurately predicting the dynamics of disease emergence, enhancing epidemiological surveillance, and informing the development of more effective and targeted public health interventions [43,44]. Hence, the current study in the North-central Nigeria, which helps us to unravel to multiple host dynamic of the virus in a bid to prepare for ways in which the disease can be combated to avoid national emergencies resulting from the spread of the disease.
Recent studies have provided valuable insights into the occurrence of arenaviruses among small mammals in Nigeria. Specifically, LASV lineage II and a novel Mobala-like virus have been identified in the Natal multimammate mouse (Mastomys natalensis), while LASV lineage III has been detected in the Guinea multimammate mouse (Mastomys erythroleucus), and the Kako strain of LASV in the African wood mouse (Hylomyscus pamfi) [10,45,46]. The use of antigen and antibody screening, particularly for immunoglobulin G (IgG), has proven effective in detecting both acute and past infections, thereby offering a clearer picture of arenavirus occurrence and prevalence patterns. Consequently, the present study aimed to integrate PCR-based detection results as have earlier been employed by Olayemi et al. [10,45] and Redding et al. [47] with newly acquired serological data, that are generated from the same individual animals in order to enhance our understanding of arenavirus circulation in Nigerian small mammal populations.
Furthermore, recent research in Nigeria has challenged the long-standing view that Mastomys natalensis is the sole reservoir of Lassa virus (LASV), revealing a more complex transmission ecology involving multiple rodent hosts [42]. LASV RNA and antibodies have been detected in several non-Mastomys genera, including Rattus rattus, Hylomyscus pamfi, and Tatera spp., with R. rattus showing positivity rates as high as 77.3% in some studies [42]. These findings suggest that such species may function either as independent reservoirs or as secondary hosts due to habitat overlap and ecological interactions with M. natalensis. Their frequent detection in domestic and peridomestic settings underscores the need to investigate their roles in LASV maintenance and transmission. Understanding the involvement of these alternative hosts is critical for refining rodent control strategies beyond M. natalensis, identifying new transmission pathways in high-risk human–rodent contact zones, and improving outbreak prediction models amid ongoing climate and land-use changes. However, key knowledge gaps remain, particularly regarding whether these species sustain LASV through vertical or horizontal transmission, or act merely as spillover hosts. To address these issues, this study aims to: (i) Identify and classify small mammal species (rodents and rats) inhabiting Lassa fever in selected endemic communities in North-Central Nigeria; (ii) Detect and characterize the presence of LASV RNA and antibodies in non-Mastomys rodent populations using molecular (PCR) and serological (IgG) approaches; (iii) Assess the potential role of these non-Mastomys rodents in LASV transmission through integrated ecological and virological analyses. These objectives are designed to fill critical gaps in our understanding of LASV ecology and to test the hypothesis that non-Mastomys species may significantly contribute to LASV maintenance and transmission cycles in endemic regions.
2. Materials and Methods
2.1. Ethical Approval
Ethical approval was obtained from the ethical committee of Federal University of Health Sciences Otukpo (FUHSO), Nigeria and the National Health Research Ethics Committee (NHREC) of Nigeria with approval number: NHREC/01/01/2007-01/09/2023.
2.2. Study Sites
For a period of four months (May to August 2024), small mammals (rodents and rats’) were trapped for sampling in Otukpo Local Government Area (LGA) of Benue State, North central Nigeria. Three wards in Otukpo LGA were randomly selected purposefully, namely Ewulo, Otukpo central and Otukpo town east (Figure 1). Three communities were selected per ward, and in Ewulo ward, we selected, Igbanonmanje (latitude: 7.204993, longitude: 8.148024), Otada (latitude: 7.226118, longitude: 8.14701), Upu (latitude: 7.215542, longitude: 8.150755). In Otukpo central ward, we selected Timber Depot 1 (latitude: 7.199372, longitude: 8.140525), Timber Depot 2 (latitude: 7.199690, longitude: 8.139603) and Otukpo main market axis (latitude: 8.131695, longitude: 7.201515). In Otukpo town east, we selected Tiv market axis (latitude: 8.151438, longitude: 7.199933), Ogiri Oko street (latitude: 8.138123, longitude: 7.194729) and General Hospital axis (latitude: 7.202721, longitude: 8.150125). In all a total of 150 locations were mapped out and traps were deployed to these locations. Traps were deployed per location based on the level of compliance of the households and communities in which the study was carried out. The climate of the Otukpo, Benue State, and North central Nigeria is tropical with two distinct seasons (wet and dry) with an average temperature of between 25 oC –29 oC, and an average humidity of 68% [48]. The study area is located within the guinea savanna vegetation belt of Nigeria with medium-level of grasses, shrubs and trees and very sparse forest distribution.
2.3. Traps Fabrication
Local fabricators were employed to help in the fabrication of traps for the study. The specificity of the traps and the purpose of the traps was defined and given to the local fabricators. The following specified measurements were used for the traps: i) length = 30 cm, width = 16.5 cm, height = 12 cm; ii) length = 37 cm, width = 15 cm, height = 13.5 cm; iii) length = 30 cm, width = 18.2 cm, height = 14 cm). Therefore, the average diameter of the fabricated traps is 35.79cm. Condemned fridge steels, metals and wood materials were used to fabricate the traps under strict supervision of the three of the research team members per time.

Plate 1: A fabricated trap.
2.4. Trapping of Small Mammals
At each community, two local field workers were employed to help deploy the fabricated traps at designated locations. Traps were deployed in strategic locations known to be rodents and rats’ pathways in every house, bush paths, dumpsites and pit latrines. The coordinates (latitudes and longitudes) of all deployment locations per community were mapped out for easy tracing and proper allocation of captured small mammals to a given location. All traps set were rebaited every morning or immediately the trap catches rodent(s) or rat(s), and the deployed traps were retained for a period of 10 days per location. We used baits such as crayfish, smoke fish, cassava, roasted yam, maize and ground nut as baits in setting the traps. Deployed traps were checked by the local field workers morning and evening on a daily basis to retrieve the captured small mammals. The total number of traps set per location (e.g., houses, pit latrines, dumpsites) varied between 80 and 210, depending on the number of households willing for the traps to be deployed in houses, pit latrines and other locations in a given community. The small mammals capture success were calculated following the protocol described by Demby et al. [19] as cited in Happi et al. [42]. The trap success in a location was equal to the sum of the total catch in a location divided by the number of traps set each night in that location multiplied by the number of trap nights and multiplied by 100.
2.5. Identification, Dissection, and Sample Collection of Small Mammals
All samples of small mammals (rodents and rats) captured on the field were transported to the laboratory for the measurement of morphological parameters and blood samples extraction/collection. Captured small mammals were first sedated with chloroform by placing them inside a desiccator for a period of three minutes before other procedures commence, e.g., identification, morphological parameters measurements and extraction/collection of blood samples. Morphological parameters such as weight, entire length, length of ear, length of foot, length of tail, and length of body were measured. Other parameters observed were sex, colour of eyes, colour of fur, mammary gland arrangement (if mature female) and number of mammary glands. Standard taxonomic keys and measurements by one of the research team members who is a trained Zoologist (with the aid of pictorial guides of common small mammals in Nigeria) was the primary identification method. However, we also used iNaturalist, biota (flora and fauna) identification application for preliminary identification of the captured samples.
After measurement of the morphological parameters of the already sedated small mammal, the next step was the collection of blood samples from the heart of the organism. A dissecting scissor was used to open up the organism, while a 2 ml syringe was used to extract blood from the heart of the small mammal. Collected blood sample from the heart of the organism was put in an EDTA bottle and a plain bottle. The EDTA is for the plasma while the plain sample bottle is for the serum. Separation of plasma and serum proper by centrifugation were done at the Infectious Disease Laboratory of the Federal University of Health Sciences Teaching Hospital, Otukpo (FUHSOTH). Centrifugation was done at 1,600 RPM (revolutions per minute). Separated plasma and serum were put in cryovial tubes for proper storage. After which samples in the cryovial tubes were stored at -1oC for a period of time before onward shipment to Institute of Viral Hemorrhagic Fever and Other Emergent Pathogens (IVEP), Irrua Specialist Teaching Hospital, Irrua, Edo State for serological and PCR tests.
2.6. Serological Testing of the Blood Samples for Lassa Fever Virus (LFV) Antibodies
At the laboratory, samples of serum separated from the blood collected from the captured small mammals were screened for LFV antibodies using an indirect enzyme-linked immunosorbent assay (ELISA), following standard protocols adapted from established methods for arenavirus surveillance in rodent reservoirs [10,49]. Briefly, 96-well microplates were pre-coated with purified LFV antigens under Biosafety level-3 (BSL-3) conditions by adding 100 µL of antigen solution per well and incubated overnight at 4°C. The plates were washed with phosphate-buffered saline containing 0.05% Tween-20 (PBS-T) and blocked with 200 µL of blocking buffer (5% bovine serum albumin) at 37°C for 1 hour to prevent non-specific binding. The rodent and rat serum samples were diluted 1:100 in sample diluent, and 100 µL of each sample, alongside known positive and negative controls, was added in duplicate to the wells. After incubation at 37°C for 1 hour, plates were washed thoroughly, and 100 µL of horseradish peroxidase (HRP)-conjugated anti-rodent IgG or IgM (as appropriate) was added to detect bound antibodies. Plates were incubated for an additional hour at 37°C, washed, and developed with 100 µL of tetramethylbenzidine (TMB) substrate in the dark for 20 minutes. The enzymatic reaction was stopped with 1N sulfuric acid, and the optical density (OD) was measured at 450 nm using a microplate reader. A given sample was considered positive if its mean OD exceeded the mean OD of the negative controls plus three standard deviations, in accordance with commonly used cut-off criteria [50]. All assays were conducted in duplicate, and results were validated by including internal quality controls on each plate.
2.7. Extraction of RNA and Reverse Transcription PCR (RT-PCR)
Total RNA was extracted from rodent and rat blood samples to detect active LFV infection, following protocols adapted from Fichet-Calvet et al. [49] and Bowen et al. [51]. All extractions and subsequent manipulations were performed under BSL-3 laboratory conditions with appropriate personal protective equipment to minimize risk of laboratory-acquired infection.
Approximately 30 mg of 100 µL of whole blood was homogenized in 600 µL of lysis buffer provided in the QIAamp Viral RNA Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Tissue homogenization was performed using a sterile mechanical tissue grinder. Lysates were incubated with proteinase K at 56 °C for 10 minutes and then processed according to the spin-column protocol, including on-column DNase I treatment to remove potential genomic DNA contamination. Viral RNA was eluted in 60 µL of RNase-free water and stored at −80 °C until use.
Detection of LFV RNA was performed by one-step reverse transcription PCR (RT-PCR) targeting the nucleoprotein (NP) gene, which is conserved among LFV lineages [51]. The reaction was carried out using the SuperScript™ III One-Step RT-PCR System with Platinum® Taq DNA Polymerase (Invitrogen, USA). Each 25 µL reaction mixture contained 12.5 µL of 2× reaction buffer, 0.5 µL of SuperScript III RT/Platinum Taq Mix, 400 nM each of forward and reverse primers specific for the LFV NP gene (e.g., LFV-F: 5′-GGAAGAGGCAACTGGCATAT-3′ and LFV-R: 5′-CTCCATGTTGTTGAGGTTGT-3′), 1 µL of extracted RNA template, and nuclease-free water to volume.
Thermal cycling was performed in a GeneAmp PCR System 9700 (Applied Biosystems) under the following conditions: reverse transcription at 50 °C for 30 min; initial denaturation at 94 °C for 2 min; followed by 40 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 30 s, and extension at 68 °C for 1 min; with a final extension at 68 °C for 5 min.
PCR products were resolved by electrophoresis on a 1.5% agarose gel stained with ethidium bromide and visualized under UV illumination. Amplicons of the expected size (~300 bp) were excised and purified for Sanger sequencing to confirm LFV-specific sequences. Positive and negative extraction controls were included in each batch of samples to monitor for cross-contamination and false positives.
2.8. Data and Statistical Analyses
Results were presented in tables, charts (bar and pie charts). Datasets were subjected to various statistical tests such as one-way analysis of variance (ANOVA), Monte-Carlo permutation, t-test and chi-square test. P values greater than 0.05 were considered statistically not significant. Student t-test was used to project the level of significance in the survival rate between the males and females’ small mammals captured alive. One-way ANOVA was used ascertain the statistical significance difference in the capture rate vs survival rate of small mammals among the nine communities sampled in the three wards sampled in Otukpo. A one-way ANOVA was also to show level of statistical significance in the number of positive cases of LFV antibodies in blood samples tested among the nine communities sampled in Otukpo LGA. We conducted Chi square test to ascertain the level of significant difference in the prevalence rate of LF infection between the two blood sample types for both IgG and IgM while Monte Carlo permutation test was used to confirm the level of significance in the prevalence rate of LF infection in the blood sample types. All charts and statistical tests were constructed and computed using paleontological statistical package (PAST; version 16.0).
3. Results
3.1. Live Capture Rate vs Survival Rate of Rodents and Rats in Otukpo LGA, North-Central Nigeria
Figures 2i-iii below presents the number of individuals of rodents and rats captured live and delivered to the laboratory. In Ewulo ward, a total of 40 live captures of rodents and rats were recorded (Figure 2i), and the number of captures is distributed as follows: Igbanonmanje-A (33); Otada-B (4) and Upu-C (3). Of the 40 live captures of rodents and rats, only 26 were delivered to the laboratory alive from Igbanonmanje, while 4 and 3 were delivered alive to the laboratory from Otada and Upu, respectively (Figure 2i). In Otukpo Town Central ward, a total of 92 live captures of rodents and rats were recorded (Figure 2ii), and the number of captures is distributed as follows: Timber Depot 1 (58), Timber Depot 2 (14) and Otukpo main market axis (20). Of the 92 live captures of rodents and rats, only 24 were delivered to the Laboratory alive from Timber Depot 1, while 11 and 16 were delivered alive to the laboratory from Timber Depot 2 and Otukpo main market axis, respectively (Figure 2ii). In Otukpo Town East ward, a total of 25 live captures of rodents and rats were recorded, 16 in Tiv market axis, 4 in Ogiri Oko street, and 5 in General Hospital axis (Figure 2iii). Of the 16 live captures of rodents and rats at Tiv market, only 5 were delivered to the laboratory alive. Of the 4 live captures at Ogiri Oko street, only 1 was delivered to the laboratory alive, while 2 delivered alive from the 5 live captures in General Hospital axis (Figure 2iii).
Overall, during the sampling period, a total of 157 live captures were recorded, and only 92 rodents and rats were delivered to the laboratory alive (Figures. 2i-iii). One-way analysis of variance (ANOVA) performed showed that there is statistically significant difference in the capture rate vs survival rate of rodents and rats among the nine communities sampled in the three wards sampled in Otukpo LGA (F= 0.0047; P<0.05). Of the 92 individuals of rats and rodents recorded alive, 65 were males while 27 were females. Student t-test performed showed that there is no significant difference in the survival rate between the males and females’ rodents and rats captured alive (t=0.067; P>0.05).
3.2. Overview of the Capture Success of Rodents and Rats in the Nine Communities Sampled in Otukpo LGA, Benue State, North-Central Nigeria
Table 1 provides a summary of the number of individuals and capture success rates of rodents and rats recorded at each sampling location. In the Igbanonmaje community, four taxa were captured: Mus musculus with nine individuals (47.36% capture success), Rattus rattus with 12 individuals (63.16%), Tatera species with four individuals (10.50%), and Pallasiomys species with one individual (5.26%). In the Otada community, two taxa were identified: Mus musculus with three individuals (12.50%) and Rattus rattus with one individual (4.17%). At Upu community, two taxa were also recorded, comprising Mus musculus with one individual (5.00%) and Rattus rattus with two individuals (10.00%). For Timber Depot 1, two taxa were captured: Mus musculus with nine individuals, showing a reported capture success of 45.0%, and Rattus rattus with fifteen individuals (27.0%). Similarly, in Timber Depot 2, Mus musculus accounted for seven individuals (14.0%) and Rattus rattus for four individuals (8.0%). In the Otukpo main market axis, two taxa were found: Rattus rattus with fifteen individuals (37.5%) and Rattus norvegicus with one individual (18.8%). At the Tiv market axis community, only Rattus rattus was recorded, with five individuals and a capture success of 66.67%. In Ogori Oko Street, a single taxon, Rattus rattus, was captured with one individual (5.6%), while at the General Hospital Axis, two individuals of Rattus rattus were recorded, representing an 18.5% capture success.
3.3. Potential Prevalence of Lassa Fever Infection Among Small Mammals in Otukpo LGA Based on Serological and Polymerase Chain Reaction (PCR) Screening
To ascertain the level of exposure of the captured small mammals to Lassa fever previously, we carried out serological tests on all the 151-blood samples (87 plasma and 64 sera) extracted from the 92 live small mammals (rodents and rats). Four small mammals were reported positives for IgG and IgM (two positives each for both IgG and IgM; Figure 3). Of the four positive samples, three were Rattus rattus (one IgG and two IgM), and one was Mus musculus (IgG); Figure 4.
All the four small mammals’ samples that tested positive for both IgG and IgM were males. Based on the results of serology per ward/community, Ewulo ward reported the highest number of positive samples (two from Igbanonmaje community, and one from Upu community; Figure 5). The remaining one positive sample was recorded in Otukpo main market axis in Otukpo Town Central ward (Figure 5). A one-way ANOVA performed to showed that there is no significant difference number of positive cases of LFV antibodies in blood samples tested among the nine communities sampled in Otukpo LGA (F= 2, df= 16, P=0.1765, P>0.05).
The prevalence of small mammals with positive results for the serological test (IgG and IgM), four were positive (three Rattus rattus-75% and one Mus musculus-25%). The potential prevalence of Lassa fever infection in small mammals within the sampling period was calculated as follow: the total number of blood samples (plasma and sera) positive for both IgG and IgM divided by the total number of blood samples tested within the designated ward/communities. Of all 151 blood samples tested for both IgG and IgM in the three wards and nine communities’ small mammals were captured, 4 (2.65%) were positive which include one plasma (0.66%) and three sera (1.99%; Figure 6).
Of the four positive blood samples, two each for IgG and IgM, signaling a potential prevalence rate of 1.32% each. Out of the 87 plasma samples analysed, one (1.15%) was positive while out of the 64 sera samples analysed three (4.69%) were positive (Figure 7). We conducted Chi square test to ascertain if there is significant difference in the prevalence rate of LF infection between the two blood sample types for both IgG and IgM, and the it revealed that there is significant difference in the infection rate between the blood sample types (Chi square value (ɤ2) =3.2914, P=0.069644, P>0.05). We further performed a Monte Carlo permutation test to confirm the level of significant in the prevalence rate of LF infection in the blood sample types, and it confirmed that there was no significantly different (P=0.0656, P>0.05).
To further confirm the level of prevalence of Lassa fever infection among the small mammals’ blood samples, we carried out a PCR, and interestingly all the samples tested returned negative for Lassa fever infection.
4. Discussion
Our study provides preliminary insights into LASV exposure among non-Mastomys rodents in Otukpo LGA, North-Central Nigeria and highlights methodological challenges inherent in such field investigations.
4.1. Logistical Challenges and Sampling Limitations
The observed discrepancies between the live capture rates (n= 157) and the number of small mammals delivered alive to the laboratory (n=92) across the three wards in the study area underscore significant logistical and ecological challenges in field sampling. The overall survival rate of 58.6% (92/157) reflects not only the fragility of small mammals during trapping and transport, but possibly species-specific stress responses and handling-related mortality, which have been widely documented in rodent ecology and epidemiological surveillance studies [52,53]. These survival constraints are crucial because only live specimens allow for full biological sampling (e.g., blood, tissue) and accurate pathogen detection. Furthermore, the significant difference in capture vs. survival rate across the nine sampled communities (ANOVA: F = 0.0047; P < 0.05) suggests that environmental or anthropogenic factors (e.g., trap placement, handling methods, ambient temperature, or time between capture and processing) might vary considerably by location. Studies have shown that high ambient temperatures and prolonged confinement in traps can lead to increased rodent mortality [16], and optimizing these conditions is vital to enhance data quality in zoonotic disease monitoring. Although a sex-based analysis of survival did not reveal significant differences (t = 0.067; P > 0.05), the higher number of males (65) than females (27) among the survivors could reflect behavioral traits, such as greater mobility or trap susceptibility of males, which has been observed in other rodent surveillance programs elsewhere [9,54]. These patterns could influence population structure assessments and may bias pathogen prevalence estimates if one sex is underrepresented. Crucially, despite targeting areas with prior reports of small mammals, Mastomys natalensis, the primary LASV reservoir, was notably absent from our captures. This absence prevents direct comparison between known and potential reservoir species within our study and is a major limitation.
4.2. Small Mammal Community Composition
In terms of species distribution, the predominance of Rattus rattus and Mus musculus in nearly all sampling sites highlights their synanthropic tendencies and their established role in urban rodent communities [55]. Their high abundance in the capture locations on marketplaces and timber depots aligns with findings from similar environments in West Africa, where food availability and shelter contribute to rodent proliferation [16,56]. The presence of less dominant genera such as Tatera and Pallasiomys in limited numbers suggests that either they are genuine rarity in these urban habitats or a lower capture probability due to behavioral differences such as trap avoidance or divergent microhabitat preferences [45]. This composition reflects the sampled habitats but may not represent rural or sylvatic zones where Mastomys or other potential hosts might be more prevalent.
4.3. Evidence of Seropositivity of LASV in the Small Mammals’ Blood Samples Types- IgG and IgM
Regarding Lassa virus (LASV) exposure, the detection of a 2.65% seropositivity rate (4/151) based on IgG and IgM testing indicates prior LASV exposure in the sampled small mammal population, despite the absence of PCR-confirmed active infections. Such discrepancies between serological and molecular findings are not uncommon and have been reported in various LASV-endemic zones [10,13]. These inconsistencies may reflect previous exposure followed by viral clearance or persistent low viremia levels that fall below PCR detection thresholds [39,41]. Seropositivity indicates exposure but does not confirm reservoir competence (i.e., the ability to maintain the virus and transmit it). The absence of PCR positivity precludes conclusions about active infection, viral shedding, or the potential for these species to sustain independent transmission cycles. They may represent spillover hosts rather than primary reservoirs
Interestingly, all seropositive individuals were male, a pattern that could be either incidental due to sample size or biologically meaningful. Male rodents typically exhibit broader home ranges, higher intraspecific aggression, and increased exploratory behavior—all factors that could enhance contact rates with infectious conspecifics or contaminated environments [57,58]. We found no significant spatial variation in seropositivity across communities (ANOVA P>0.05), consistent with a uniformly low level of detected exposure across the sampled area.
Notably, Rattus rattus was the dominant seropositive species (75%), further supporting earlier studies [e.g., 9,59] and recent studies [e.g., 40,42] that implicate non-Mastomys rodents in LASV circulation. Although Mastomys natalensis remains the primary reservoir, emerging data suggest a broader host range, particularly in peri-urban and urban settings [18]. This highlights the importance of multi-species surveillance to capture the full epidemiological picture of LASV. The finding of no significant variation in LASV prevalence across communities (ANOVA: P > 0.05) indicates a uniform low-level exposure risk in the region. However, the significant Chi-square result (P = 0.0696) in infection rates between plasma vs. sera samples, although marginal, underscores the importance of sample type and storage conditions in serological sensitivity [60,61]. Lastly, the negative PCR results for all samples suggest that none of the small mammals were actively shedding the virus at the time of sampling. This is consistent with the low prevalence rates (<3%) reported in other endemic areas [2,19,62], and reflects the episodic nature of LASV circulation in rodent and other small mammal populations.
Compounding this complexity is the role of other commensal rodent species such as Rattus rattus and Mus musculus, which are believed to have been introduced into Africa via maritime trade routes [21,51,65]. Their presence, particularly in urban areas, appears to influence the spatial ecology of Mastomys populations. For example, in The Republic of Guinea, M. musculus was not found east of Kindia, possibly due to limited inland spread, while R. rattus was commonly observed in urbanized zones. Ecological interactions, including possible competitive exclusion, suggest that R. rattus may displace Mastomys in human dwellings [2,17]. Although the exclusion is not absolute, such interactions could partially explain the relative rarity of urban LF cases, as Mastomys are often outcompeted in densely populated urban environments. Interestingly, some authors have even speculated that the presence of R. rattus in households could offer a form of ecological “protection” from LF, as they may inhibit Mastomys colonization [60,63,64]. However, this should be interpreted with caution, as R. rattus are themselves known reservoirs for other zoonotic diseases and should not be considered benign.
Furthermore, one of the central challenges in understanding the epidemiology of LF lies in deciphering the apparent mismatch between the distribution of the disease in humans and the presence of its principal reservoir host, Mastomys natalensis, across West Africa. Although Mastomys populations are widespread, LF incidence remains highly focal. This paradox has long intrigued researchers and raises critical questions about the ecological, behavioral, and socio-environmental factors influencing transmission. Field studies in The Republic of Guinea revealed that the spatial distribution of LASV-positive Mastomys rodents closely mirrored that of human IgG seroprevalence patterns [60], suggesting shared exposure zones and patterns. However, substantial heterogeneity was observed even between closely situated villages. [60] reported that within some communities, Mastomys positive for LASV-specific antibodies (Ag or Ab) clustered in only a few households, underscoring the micro-ecological nature of rodent infections and their potential to create highly localized hotspots of human risk. More puzzling than the absence of LF in Mastomys-free regions is the variability in LASV prevalence in areas where Mastomys are abundant. In The Republic of Guinea, trap success for Mastomys was often up to 10 times higher than in hyperendemic regions of Sierra Leone, yet LASV prevalence among the rodents was markedly lower, by a factor of three to 15 [17,60]. This suggests that high rodent density does not necessarily equate to high transmission risk. Instead, human behavioral and environmental factors likely play a more decisive role in disease incidence. These include poor sanitation, overcrowding, food storage practices, housing quality, environmental disturbance, and rodent hunting and consumption, and these factors vary substantially between regions and socio-economic contexts [34,66,67]. For example, Sierra Leone’s higher LF burden relative to The Republic of Guinea may be partially attributable to years of socio-political instability, diamond mining, and displacement, which exacerbated exposure risks through habitat disruption and poor living conditions [17,68,69]. Adding to the complexity is the issue of underrecognition. Many LASV infections present with non-specific symptoms and can be misdiagnosed as malaria or typhoid fever in the absence of appropriate diagnostic infrastructure [19,59]. This diagnostic gap likely contributes to underreporting and an underestimation of LF burden in several regions, including The Republic of Guinea and parts of Nigeria.
Overall, our findings in the current study, underscore the multifactorial nature of LF epidemiology. While the presence of Mastomys natalensis remains a necessary condition for zoonotic transmission, it alone cannot account for the spatial heterogeneity observed in human LF cases. A comprehensive understanding requires the integration of of the ecology of rodents and other small mammals with human behavior, environmental transformations, and the capacity of healthcare systems. Consequently, future surveillance and control strategies must be both ecologically informed and culturally responsive, particularly in transitioning rural–urban landscapes where rodent communities, and their associated risks are dynamically evolving. In this context, Nigeria, where LF is hyperendemic and seasonal outbreaks are recurrent, the current study offers valuable lessons for future studies in exploring the ecology of small mammals that have been established to be host of LASV as well as the seroprevalence of the disease. The recent detection of novel LASV lineages in neighboring countries such as Mali highlights the urgent need for regional collaboration in surveillance, research, and early detection. Nigeria’s extensive experience in managing LF can serve as a foundation for capacity-building initiatives across at-risk West African countries. Strengthening cross-border surveillance systems, enhancing clinical awareness, and expanding access to molecular diagnostic tools are critical steps toward addressing LASV circulation as a regional public health priority.
4.4. Implications and Study Constraints
While our data provide evidence of LASV exposure in Rattus rattus and Mus musculus within the study area, the key limitations are as follows: small sample size, low survival rate, absence of Mastomys natalensis, and lack of PCR confirmation—preclude definitive statements about their role as reservoirs. The logistical challenges encountered, particularly low post-capture survival, significantly impacted the scale and potential insights of this study. Optimizing trapping, handling, and transport protocols is essential for future research aiming to accurately assess pathogen prevalence and reservoir potential in small mammals.
5. Conclusions and Perspectives
This study confirms LASV exposure occurs in some non-Mastomys rodents (Rattus rattus, Mus musculus) within the sampled peri-urban environments of Otukpo LGA. It thus provides critical baseline data on rodent and other small mammals diversity, survival, and Lassa virus (LASV) exposure in Otukpo LGA, North-Central Nigeria. The findings highlight the emerging role of Rattus rattus and Mus musculus, alongside Mastomys natalensis, as potential LASV reservoirs, emphasizing the need to broaden surveillance to include synanthropic and understudied species. The complete lack of captured Mastomys natalensis within this LF-endemic region is a surprising and significant finding, underscoring the need for improved sampling strategies targeting this primary reservoir for meaningful ecological context. Future investigations require larger-scale, longitudinal designs, incorporating diverse habitats and rigorous methods to ensure adequate survival and sample sizes of both target non-Mastomys species and Mastomys natalensis to robustly evaluate the potential contribution of alternative hosts to LASV ecology.
Challenges with live trapping and post-capture survival point to the urgency of refining capture protocols and improving animal care to ensure both ethical standards and data integrity. Enhancing sampling strategies, such as microhabitat targeting, increased trap density, and optimized baiting alongside longitudinal monitoring across ecological gradients, will provide deeper insights into host turnover, transmission dynamics, and LASV prevalence. To support effective LASV control, we recommend that studies that will expand the surveillance the reservoirs of LASV as well as develop species-specific handling guidelines; integrating ecological, virological, and spatial data into predictive models. We also recommend that strengthening local research capacity and interdisciplinary collaboration, and promoting community-level awareness and rodent and other small mammals proofing initiatives will further help in curbing the imminent dangers posed by this virus ravaging mostly countries in the West Africa UN subregion. Together, these strategies can advance a more comprehensive and proactive approach to Lassa fever prevention in Nigeria and the wider West African UN subregion.
Author Contributions
AOE: Conceptualization, Funding acquisition, Methodology, Data Analysis, Writing—Original draft, Writing—review and editing, Manuscript finalization, Supervision. TOO: Methodology, Data curation, Creation of distribution map, Writing—review and editing, Manuscript finalization. JA: Conceptualization, Funding acquisition, Methodology, Writing—review and editing, Manuscript finalization. SEY: Writing—Original draft, Writing—review and editing, Manuscript finalization. OSS: Conceptualization, Funding acquisition, Methodology, Writing—review and editing, Manuscript finalization. OA: Funding acquisition, Methodology, Writing—review and editing, Manuscript finalization. EOA: Writing—review and editing, Manuscript finalization. EBA: Conceptualization, Funding acquisition, Methodology, Writing—review and editing, Manuscript finalization. JAI: Conceptualization, Funding acquisition, Methodology, Writing—review and editing, Manuscript finalization. IOAU: Conceptualization, Funding acquisition, Methodology, Writing—review and editing, Manuscript finalization. SOA: Conceptualization, Funding acquisition, Methodology, Data Analysis, Writing—Original draft, Writing—review and editing, Manuscript finalization, Supervision.
Funding
This study was funded by the Tertiary Education Trust Fund Nigeria (TETFUND NG) tagged TETFUND mega grant for the establishment of Centre of excellence with grant.
Data Availability Statement
Raw dataset collected and used in this study will be made available by the corresponding author upon request.
Acknowledgments
We acknowledge the Tertiary Education Trust Fund Nigeria (TETFUND NG) for funding this project. The communities and households in Otukpo LGA where traps were deployed for sample collections are acknowledged for allowing us carry out this research. We also acknowledge the management of the Federal University of Health Sciences Teaching Hospital, Otukpo (FUHSOTH) for allowing us preserved our samples in their laboratory before their transfer to Institute of Viral Hemorrhagic Fever and Other Emergent Pathogens (IVEP), Irrua Specialist Teaching Hospital, Irrua, Edo State for further analyses. The IVEP is also appreciated for their initial training of the project team members at the onset of the study as well as their guidance in conducting both the serological and molecular analyses. We acknowledged the efforts of the field and laboratory assistants during this project: Victor Ejeh, Augustine Okwobi, Tega Treasure Edegbene Ovie, Saturday James Okon and the Research Assistant: Clement Ameh.
Conflicts of Interest
We declare that there is no conflict of interest.
References
- Richmond, J.K.; Baglole, D.J. Lassa fever: epidemiology, clinical features, and social consequences. BMJ. 2003, 327, 1271–1275. [Google Scholar] [CrossRef]
- Ogbu, O.; Ajuluchukwu, E.; Uneke, C.J. Lassa fever in West African sub-region: an overview. J. Vector Borne Dis. 2007, 44, 1–6. [Google Scholar] [PubMed]
- Trovato, M.; Sartorius, R.; D’Apice, L.; Manco, R.; De Berardinis, P. Viral emerging diseases: challenges in developing vaccination strategies. Front. Immunol. 2020, 11, 2130. [Google Scholar] [CrossRef]
- Frame, J.D.; Baldwin, J.J.; Gocke, D.J.; Troup, J.M. Lassa fever, a new virus disease of man from West Africa. I. Clinical description and pathological findings. Am. J. Trop. Med. Hyg. 1970, 19, 670–676. [Google Scholar] [CrossRef] [PubMed]
- McCormick, J.B.; Fisher-Hoch, S.P. Lassa fever. Arenaviruses I: Epidemiol. Mol. Cell Biol. Arenaviruses 2002, 1, 75–109. [Google Scholar]
- Ogoina, D. Fever, fever patterns and diseases called ‘fever’–a review. J. Infect. Public Health 2011, 4, 108–124. [Google Scholar] [CrossRef]
- Bausch, D.G.; Mills, J.N. Arenaviruses: Lassa fever, Lujo hemorrhagic fever, lymphocytic choriomeningitis, and the South American hemorrhagic fevers. Viral Infect. Hum. 2014, 5, 147–171. [Google Scholar]
- Grubaugh, N.D.; Ladner, J.T.; Lemey, P.; Pybus, O.G.; Rambaut, A.; Holmes, E.C.; Andersen, K.G. Tracking virus outbreaks in the twenty-first century. Nat. Microbiol. 2019, 4, 10–19. [Google Scholar] [CrossRef]
- Fichet-Calvet, E.; Rogers, D.J. Risk maps of Lassa fever in West Africa. PLoS Negl. Trop. Dis. 2009, 3, e388. [Google Scholar] [CrossRef]
- Olayemi, A.; Cadar, D.; Magassouba, N.; Obadare, A.; Kourouma, F.; Oyeyiola, A.; Fasogbon, S.; Igbokwe, J.; Rieger, T.; Bockholt, S.; Günther, S.; Fichet-Calvet, E. New hosts of the Lassa virus. Sci. Rep. 2016, 6, 25280. [Google Scholar] [CrossRef]
- Echchakery, M.; Boussaa, S.; Kahime, K.; Boumezzough, A. Epidemiological role of a rodent in Morocco: case of cutaneous leishmaniasis. Asian Pac. J. Trop. Dis. 2015, 5, 589–594. [Google Scholar] [CrossRef]
- Andersen, K.G.; Shylakhter, I.; Tabrizi, S.; Grossman, S.R.; Happi, C.T.; Sabeti, P.C. Genome-wide scans provide evidence for positive selection of genes implicated in Lassa fever. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Whitmer, S.L.; Strecker, T.; Cadar, D.; Dienes, H.P.; Faber, K.; Patel, K.; Günther, S. New lineage of Lassa virus, Togo, 2016. Emerg. Infect. Dis. 2018, 24, 599–602. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, W.; Wang, Z.; Yang, W.; Li, E.; Xia, X.; Chiu, S. Emerging and reemerging infectious diseases: global trends and new strategies for their prevention and control. Signal Transduct. Target. Ther. 2024, 9, 223. [Google Scholar] [CrossRef]
- Salazar-Bravo, J.; Dragoo, J.W.; Bowen, M.D.; Peters, C.J.; Ksiazek, T.G.; Yates, T.L. Natural nidality in Bolivian hemorrhagic fever and the systematics of the reservoir species. Infect. Genet. Evol. 2002, 1, 191–199. [Google Scholar] [CrossRef]
- Meerburg, B.G.; Singleton, G.R.; Kijlstra, A. Rodent-borne diseases and their risks for public health. Crit. Rev. Microbiol. 2009, 35, 221–270. [Google Scholar] [CrossRef]
- McCormick, J.B. Epidemiology and control of Lassa fever. In Arenaviruses: Biol. Immunother. 1987, 69–78.
- Lecompte, E.; Fichet-Calvet, E.; Daffis, S.; Koulémou, K.; Sylla, O.; Kourouma, F.; ter Meulen, J. Mastomys natalensis and Lassa fever, West Africa. Emerg. Infect. Dis. 2006, 12, 1971. [Google Scholar] [CrossRef]
- Demby, A.H.; Inapogui, A.; Kargbo, K.; Koninga, J.; Kourouma, K.; Kanu, J.; Coulibaly, M.; Wagoner, K.D.; Ksiazek, T.G.; Peters, C.J.; Rollin, P.E.; Bausch, D.G. Lassa fever in Guinea: II. Distribution and prevalence of Lassa virus infection in small mammals. Vector Borne Zoonotic Dis. 2001, 1, 283–297. [Google Scholar] [CrossRef]
- Walker, D.H.; McCormick, J.B.; Johnson, K.M.; Webb, P.A.; Komba-Kono, G.; Elliott, L.H.; Gardner, J.J. Pathologic and virologic study of fatal Lassa fever in man. Am. J. Pathol. 1982, 107, 349. [Google Scholar]
- Johnson, K.M.; McCormick, J.B.; Webb, P.A.; Smith, E.S.; Elliott, L.H.; King, I.J. Clinical virology of Lassa fever in hospitalized patients. J. Infect. Dis. 1987, 155, 456–464. [Google Scholar] [CrossRef]
- Demartini, J.C.; Green, D.E.; Monath, T.P. Lassa virus infection in Mastomys natalensis in Sierra Leone: Gross and microscopic findings in infected and uninfected animals. Bull. World Health Organ. 1975, 52, 651. [Google Scholar]
- Carey, D.E.; Kemp, G.E.; White, H.A.; Pinneo, L.; Addy, R.F.; Fom, A.L.M.D.; Henderson, B.E. Lassa fever: epidemiological aspects of the 1970 epidemic, Jos, Nigeria. Trans. R. Soc. Trop. Med. Hyg. 1972, 66, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Webb, P.A.; McCormick, J.B.; King, I.J.; Bosman, I.; Johnson, K.M.; Elliott, L.H.; O’Sullivan, R. Lassa fever in children in Sierra Leone, West Africa. Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Ter Meulen, J. Lassa fever: implications of T-cell immunity for vaccine development. J. Biotechnol. 1999, 73, 207–212. [Google Scholar] [CrossRef]
- Sogoba, N.; Feldmann, H.; Safronetz, D. Lassa fever in West Africa: evidence for an expanded region of endemicity. Zoonoses Public Health 2012, 59, 43–47. [Google Scholar] [CrossRef]
- Mbaave, T.P.; Ogbu, O.; Echekwube, P.O.; Swende, T.Z.; Igbah, I.T. Lassa Fever: Patients Profile and Treatment Outcomes at Benue State University Teaching Hospital Makurdi, North-Central Nigeria. Adv. Infect. Dis. 2023, 13, 722–734. [Google Scholar] [CrossRef]
- Monath, T.P. Lassa fever: review of epidemiology and epizootiology. Bull. World Health Organ. 1975, 52, 577. [Google Scholar]
- Capul, A.A.; de la Torre, J.C.; Buchmeier, M.J. Conserved residues in Lassa fever virus Z protein modulate viral infectivity at the level of the ribonucleoprotein. J. Virol. 2011, 85, 3172–3178. [Google Scholar] [CrossRef]
- Yun, N.E.; Walker, D.H. Pathogenesis of Lassa fever. Viruses 2012, 4, 2031–2048. [Google Scholar] [CrossRef]
- Grard, G.; Fair, J.N.; Lee, D.; Slikas, E.; Steffen, I.; Muyembe, J.J.; Leroy, E.M. A novel rhabdovirus associated with acute hemorrhagic fever in central Africa. PLoS Negl. Trop. Dis. 2012, 6, e100. [Google Scholar] [CrossRef]
- Delahoy, M.J.; Wodnik, B.; McAliley, L.; Penakalapati, G.; Swarthout, J.; Freeman, M.C.; Levy, K. Pathogens transmitted in animal feces in low-and middle-income countries. Int. J. Hyg. Environ. Health 2018, 221, 661–676. [Google Scholar] [CrossRef] [PubMed]
- Frame, J.D.; Casals, J.; Dennis, E.A. Lassa virus antibodies in hospital personnel in western Liberia. Trans. R. Soc. Trop. Med. Hyg. 1979, 73, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Ter Meulen, J.; Lukashevich, I.; Sidibe, K.; Inapogui, A.; Marx, M.; Dorlemann, A.; et al. Hunting of peridomestic rodents and consumption of their meat as possible risk factors for rodent-to-human transmission of Lassa virus in the Republic of Guinea. Am. J. Trop. Med. Hyg. 1996, 55, 661–666. [Google Scholar] [CrossRef]
- Günther, S.; Emmerich, P.; Laue, T.; Kühle, O.; Asper, M.; Jung, A.; Schmitz, H. Imported Lassa fever in Germany: molecular characterization of a new Lassa virus strain. Emerg. Infect. Dis. 2000, 6, 466. [Google Scholar] [CrossRef]
- Georges, A.J.; Gonzalez, J.P.; Abdul Wahid, S.; Saluzzo, J.F.; Meunier, D.M.; McCormick, J.B. Antibodies to Lassa and Lassa-like viruses in man and mammals in the Central African Republic. Trans. R. Soc. Trop. Med. Hyg. 1985, 79, 78–79. [Google Scholar] [CrossRef]
- Lukashevich, I.S.; Maryankova, R.; Vladyko, A.S.; Nashkevich, N.; Koleda, S.; Djavani, M.; et al. Lassa and Mopeia virus replication in human monocytes/macrophages and in endothelial cells: different effects on IL-8 and TNF-alpha gene expression. J. Med. Virol. 1999, 59, 552–560. [Google Scholar] [CrossRef]
- Macher, A.M.; Wolfe, M.S. Historical Lassa fever reports and 30-year clinical update. Emerg. Infect. Dis. 2006, 12, 835. [Google Scholar] [CrossRef]
- Safronetz, D.; Mire, C.; Rosenke, K.; Feldmann, F.; Haddock, E.; Geisbert, T.; Feldmann, H. A recombinant vesicular stomatitis virus-based Lassa fever vaccine protects guinea pigs and macaques against challenge with geographically and genetically distinct Lassa viruses. PLoS Negl. Trop. Dis. 2015, 9, e0003736. [Google Scholar] [CrossRef]
- Garry, R.F. Lassa fever—the road ahead. Nat. Rev. Microbiol. 2023, 21, 87–96. [Google Scholar]
- Ehichioya, D.U.; Asogun, D.A.; Ehimuan, J.; Okokhere, P.O.; Pahlmann, M.; Ölschläger, S.; Omilabu, S.A. Hospital-based surveillance for Lassa fever in Edo State, Nigeria, 2005–2008. Trop. Med. Int. Health 2012, 17, 1001–1004. [Google Scholar] [CrossRef]
- Happi, A.N.; Olumade, T.J.; Ogunsanya, O.A.; Sijuwola, A.E.; Ogunleye, S.C.; Oguzie, J.U.; Nwofoke, C.; Ugwu, C.A.; Okoro, S.J.; Otuh, P.I.; Ngele, L.N.; et al. Increased Prevalence of Lassa Fever Virus-Positive Rodents and Diversity of Infected Species Found during Human Lassa Fever Epidemics in Nigeria. Microbiol. Spectr. 2022, 10, e00366-22. [Google Scholar] [CrossRef]
- Mateer, E.J.; Huang, C.; Shehu, N.Y.; Paessler, S. Lassa fever–induced sensorineural hearing loss: A neglected public health and social burden. PLoS Negl. Trop. Dis. 2018, 12, e0006187. [Google Scholar] [CrossRef]
- Asogun, D.A.; Günther, S.; Akpede, G.O.; Ihekweazu, C.; Zumla, A. Lassa fever: Epidemiology, clinical features, diagnosis, management and prevention. Infect. Dis. Clin. 2019, 33, 933–951. [Google Scholar]
- Olayemi, A.; Obadare, A.; Oyeyiola, A.; Fasogbon, S.; Igbokwe, J.; Igbahenah, F.; Fichet-Calvet, E. Small mammal diversity and dynamics within Nigeria, with emphasis on reservoirs of the Lassa virus. Syst. Biodivers. 2018, 16, 118–127. [Google Scholar] [CrossRef]
- Klitting, R.; Kafetzopoulou, L.E.; Thiery, W.; Dudas, G.; Gryseels, S.; Kotamarthi, A.; Dellicour, S. Predicting the evolution of the Lassa virus endemic area and population at risk over the next decades. Nat. Commun. 2022, 13, 5596. [Google Scholar] [CrossRef] [PubMed]
- Redding, D.W.; Gibb, R.; Dan-Nwafor, C.C.; Ilori, E.A.; Yashe, R.U.; Oladele, S.H.; Ihekweazu, C. Geographical drivers and climate-linked dynamics of Lassa fever in Nigeria. Nat. Commun. 2021, 12, 5759. [Google Scholar] [CrossRef] [PubMed]
- Edegbene, A.O.; Yandev, D.; Omotehinwa, T.O.; Zakari, H.; Andy, B.O. Water quality assessment in Benue South, Nigeria: An investigation of physico-chemical and microbial characteristics. Water Sci. 2025. [Google Scholar] [CrossRef]
- Fichet-Calvet, E.; Lecompte, E.; Koivogui, L.; Soropogui, B.; Doré, A.; Kourouma, F.; Günther, S. Fluctuation of abundance and Lassa virus prevalence in Mastomys natalensis in Guinea, West Africa. Vector-Borne Zoonotic Dis. 2007, 7, 119–128. [Google Scholar] [CrossRef]
- Moses, L.M.; et al. Serological evidence of arenavirus circulation among rodents in endemic regions. Emerg. Infect. Dis. 2022, 28, 561–570. [Google Scholar]
- Bowen, M.D.; Rollin, P.E.; Ksiazek, T.G.; Hustad, H.L.; Bausch, D.G.; Demby, A.H.; Bajani, M.D.; Peters, C.J.; Nichol, S.T. Genetic diversity among Lassa virus strains. J. Virol. 2000, 74, 6992–7004. [Google Scholar] [CrossRef]
- Mills, J.N.; Childs, J.E. Rodent-borne hemorrhagic fever viruses. Infect. Dis. Wild Mamm. 2001, 254–270. [Google Scholar]
- Mariën, J.; Kourouma, F.; Magassouba, N.F.; Leirs, H.; Fichet-Calvet, E. Movement patterns of small rodents in Lassa fever-endemic villages in Guinea. EcoHealth 2018, 15, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.A.; Kenyon, M.A.; Ghersi, B.M.; Sparks, J.L.D.; Gass, J.D. Assessing the environmental drivers of Lassa fever in West Africa: A systematic review. Viruses 2025, 17, 504. [Google Scholar] [CrossRef] [PubMed]
- Aplin, K.P.; Chesser, T.; Have, J.T. Evolutionary biology of the genus Rattus: Profile of an archetypal rodent pest. ACIAR Monogr. Ser. 2003, 96, 487–498. [Google Scholar]
- Gratz, N. The burden of rodent-borne diseases in Africa south of the Sahara. Belg. J. Zool. 1997, 127, 71–84. [Google Scholar]
- Kennis, J.; Sluydts, V.; Leirs, H.; van Hooft, W.P. Polyandry and polygyny in an African rodent pest species, Mastomys natalensis. Mammalia 2008, 72, 226–245. [Google Scholar] [CrossRef]
- Madueme, P.G.U.; Chirove, F. A systematic review of mathematical models of Lassa fever. Math. Biosci. 2024, 374, 109227. [Google Scholar] [CrossRef]
- Andersen, K.G.; Shapiro, B.J.; Matranga, C.B.; Sealfon, R.; Lin, A.E.; Moses, L.M.; Sabeti, P.C. Clinical sequencing uncovers origins and evolution of Lassa virus. Cell 2015, 162, 738–750. [Google Scholar] [CrossRef]
- Bausch, D.G.; Demby, A.H.; Coulibaly, M.; Kanu, J.; Goba, A.; Bah, A.; Rollin, P.E. Lassa fever in Guinea: I. Epidemiology of human disease and clinical observations. Vector-Borne Zoonotic Dis. 2001, 1, 269–281. [Google Scholar] [CrossRef]
- Ilori, E.A.; Furuse, Y.; Ipadeola, O.B.; Dan-Nwafor, C.C.; Abubakar, A.; Womi-Eteng, O.E.; Team, N.L.F.N.R. Epidemiologic and clinical features of Lassa fever outbreak in Nigeria, January 1–May 6, 2018. Emerg. Infect. Dis. 2019, 25, 1066. [Google Scholar] [CrossRef]
- Günther, S.; Lenz, O. Lassa virus. Crit. Rev. Clin. Lab. Sci. 2004, 41, 339–390. [Google Scholar] [CrossRef]
- Monath, T.P. Lassa fever and Marburg virus disease. WHO Chron. 1974, 28, 212–219. [Google Scholar]
- Cross, R.W.; Heinrich, M.L.; Fenton, K.A.; Borisevich, V.; Agans, K.N.; Prasad, A.N.; Geisbert, T.W. A human monoclonal antibody combination rescues nonhuman primates from advanced disease caused by the major lineages of Lassa virus. Proc. Natl. Acad. Sci. USA 2023, 120, e2304876120. [Google Scholar] [CrossRef] [PubMed]
- Musa, S.S.; Zhao, S.; Gao, D.; Lin, Q.; Chowell, G.; He, D. Mechanistic modelling of the large-scale Lassa fever epidemics in Nigeria from 2016 to 2019. J. Theor. Biol. 2020, 493, 110209. [Google Scholar] [CrossRef] [PubMed]
- Isaäcson, M. The ecology of Praomys (Mastomys) natalensis in southern Africa. Bull. World Health Organ. 1975, 52, 629. [Google Scholar] [PubMed]
- Eskew, E.A.; Bird, B.H.; Ghersi, B.M.; Bangura, J.; Basinski, A.J.; Amara, E.; Nuismer, S.L. Reservoir displacement by an invasive rodent reduces Lassa virus zoonotic spillover risk. Nat. Commun. 2024, 15, 3589. [Google Scholar] [CrossRef]
- Allan, R.; Mardell, S.; Ladbury, R.; Pearce, E.; et al. The progression from endemic to epidemic Lassa fever in war-torn West Africa. In Emergence and Control of Rodent-Borne Viral Diseases; Saluzzo, J.F., Dodet, B., Eds.; Elsevier: Annecy, France, 1998; pp. 97–105. [Google Scholar]
- ter Meulen, J.; Lenz, O.; Koivogui, L.; Magassouba, N.; et al. Short communication: Lassa fever in Sierra Leone: UN peacekeepers are at risk. Trop. Med. Int. Health 2001, 6, 83–84. [Google Scholar] [CrossRef]
Figure 1.
Map of Nigeria showing Benue State, Otukpo LGA and the wards/communities where small mammals were trapped.
Figure 1.
Map of Nigeria showing Benue State, Otukpo LGA and the wards/communities where small mammals were trapped.
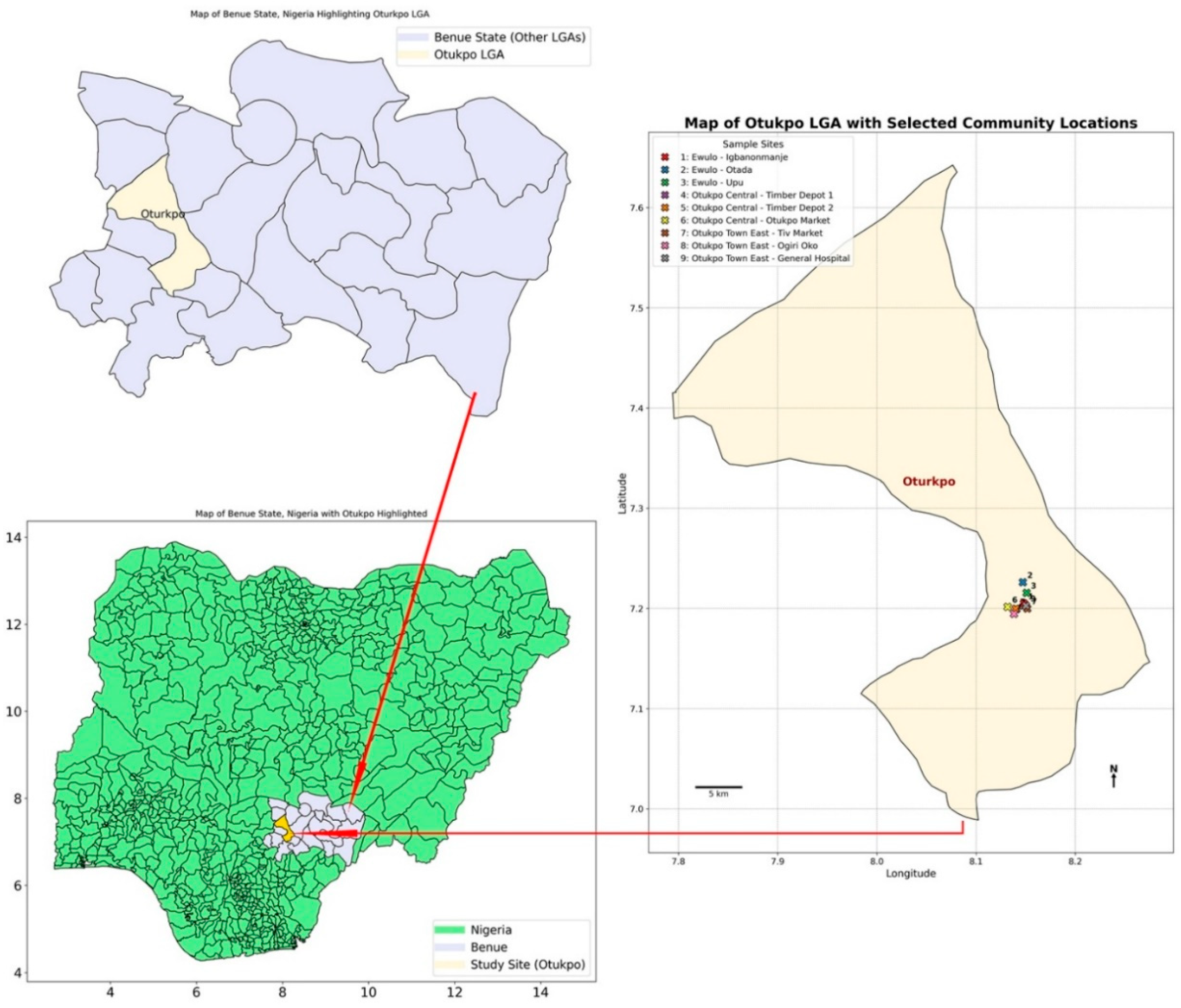
Figure 2.
(i-iii) Live capture rate vs survival rate of rodents and rats. Note: LGA wards/communities: Ewulo Ward: A= Igbanonmanje; B= Otada; C= Upu; Otukpo Town Central Ward: D = Timber Depot 1; E = Timber Depot 2; F = Otukpo main market axis; Otukpo Town East Ward: G = Tiv market axis; H = Ogiri Oko street; I = General Hospital axis.
Figure 2.
(i-iii) Live capture rate vs survival rate of rodents and rats. Note: LGA wards/communities: Ewulo Ward: A= Igbanonmanje; B= Otada; C= Upu; Otukpo Town Central Ward: D = Timber Depot 1; E = Timber Depot 2; F = Otukpo main market axis; Otukpo Town East Ward: G = Tiv market axis; H = Ogiri Oko street; I = General Hospital axis.
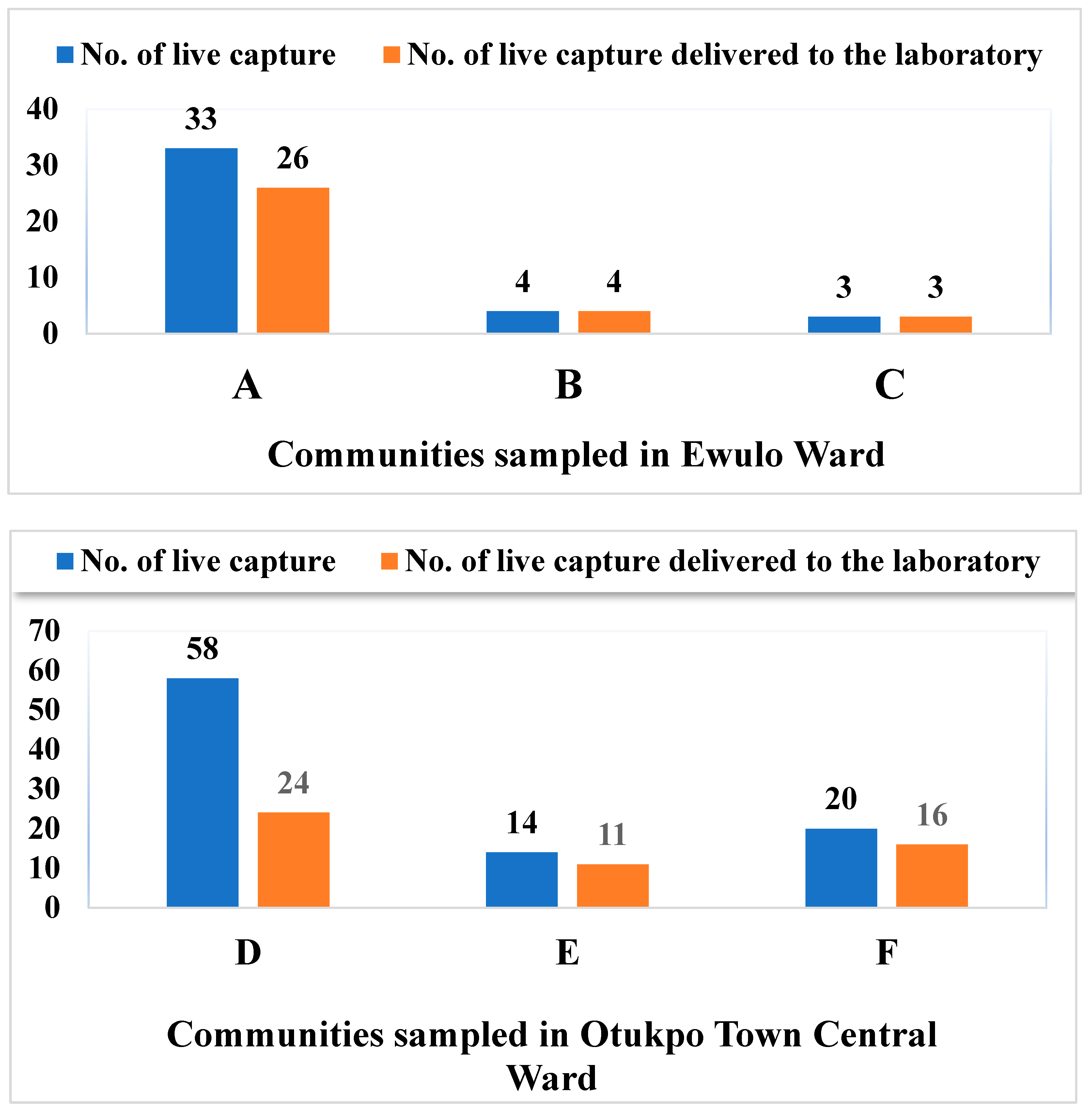
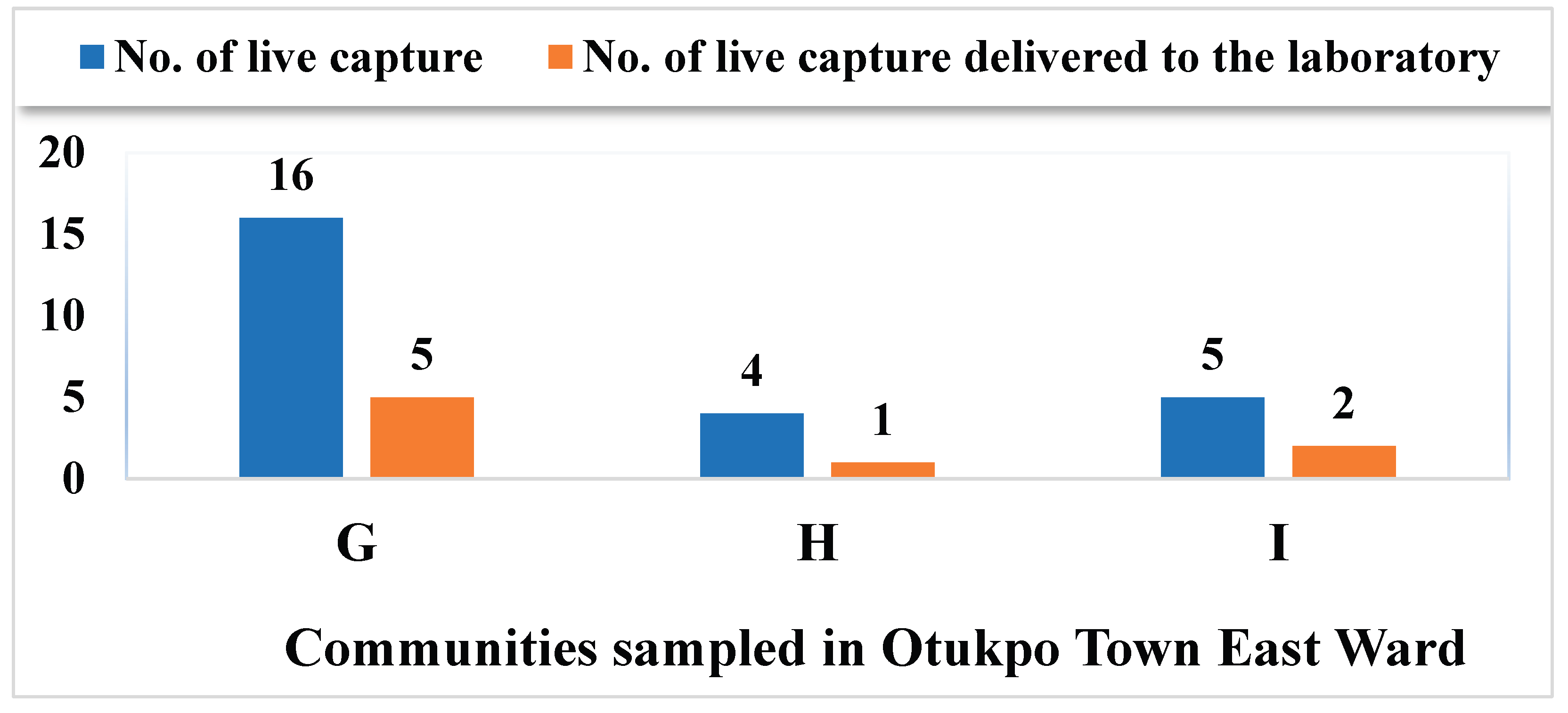
Figure 3.
Summary of blood samples positive for both IgG and IgM .
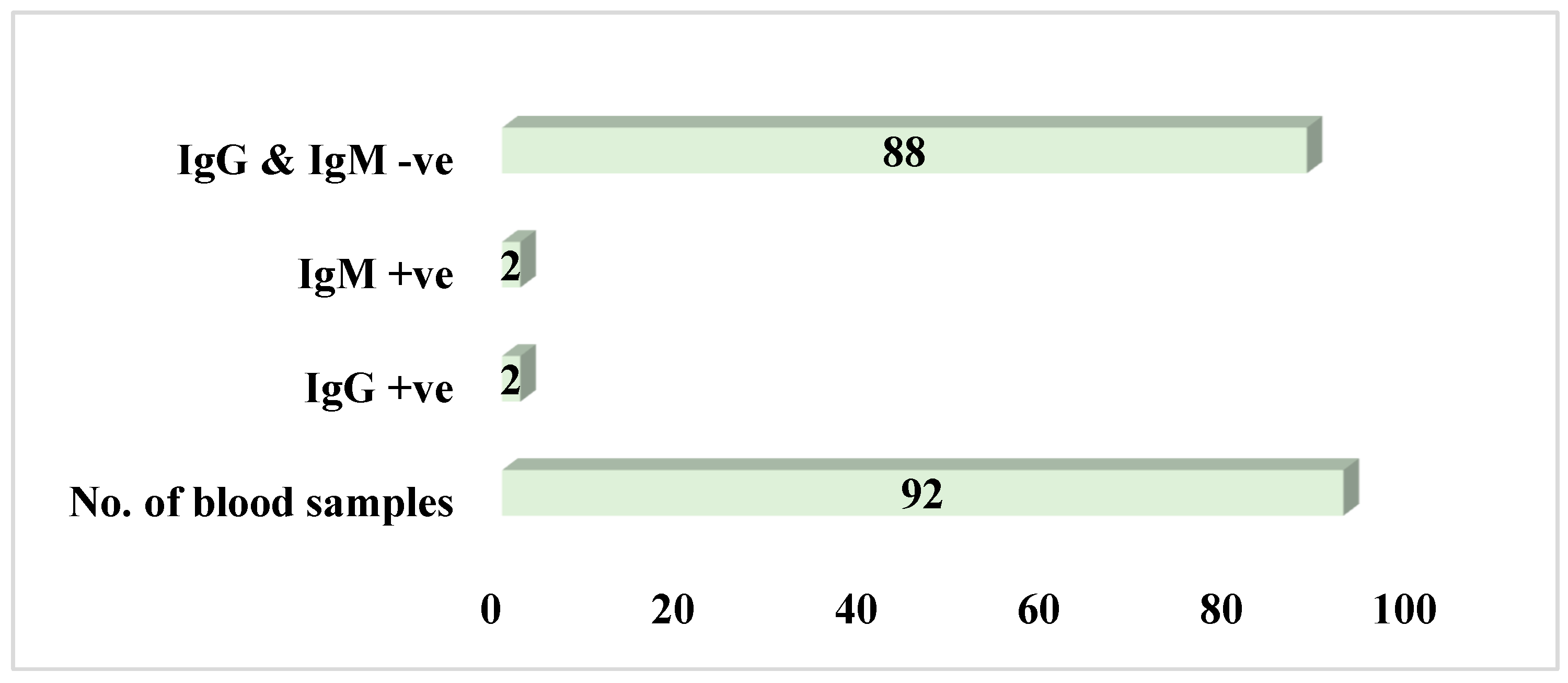
Figure 4.
Percentage potential prevalence of Lassa fever infection among captured small mammals .
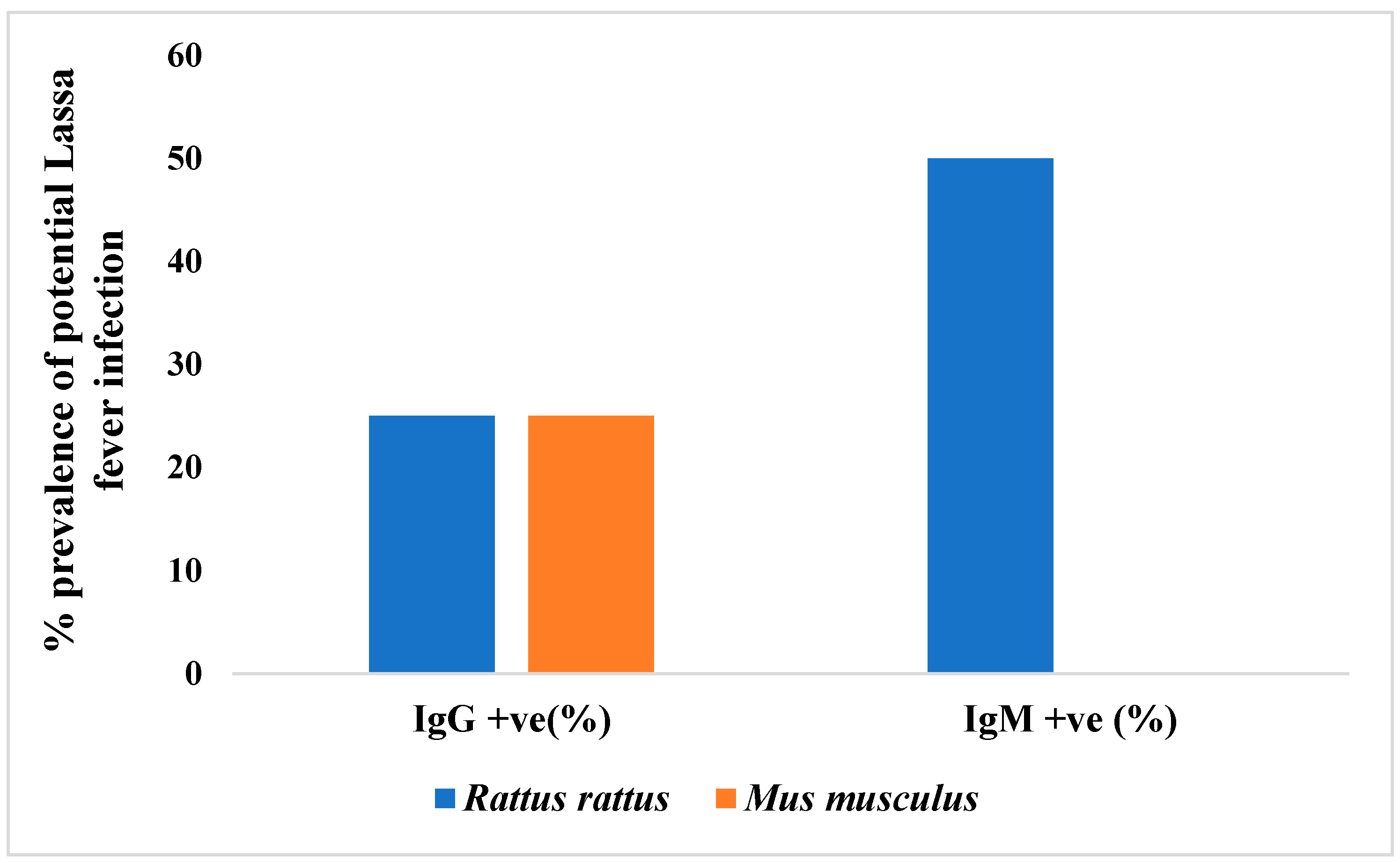
Figure 5.
Number of positives cases of LFV antibodies in the blood samples of the small mammals among the communities sampled. Note: LGA wards/communities: Ewulo Ward: A = Igbanonmanje; B = Otada; C= Upu; Otukpo Town Central Ward: D = Timber Depot 1; E = Timber Depot 2; F = Otukpo main market axis; Otukpo Town East Ward: G = Tiv market axis; H = Ogiri Oko street; I= General Hospital axis.
Figure 5.
Number of positives cases of LFV antibodies in the blood samples of the small mammals among the communities sampled. Note: LGA wards/communities: Ewulo Ward: A = Igbanonmanje; B = Otada; C= Upu; Otukpo Town Central Ward: D = Timber Depot 1; E = Timber Depot 2; F = Otukpo main market axis; Otukpo Town East Ward: G = Tiv market axis; H = Ogiri Oko street; I= General Hospital axis.
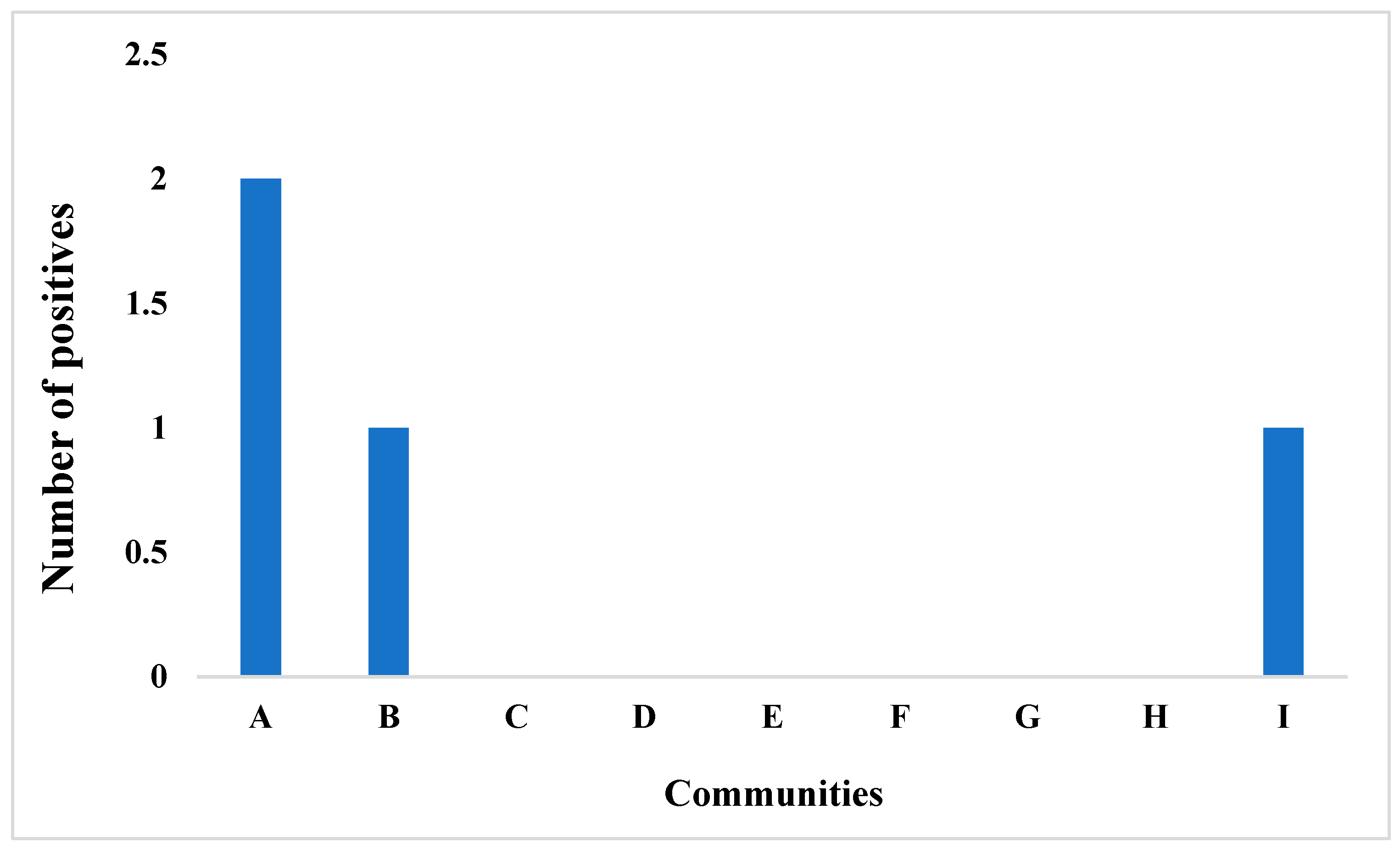
Figure 6.
Percentage prevalence of LFV in the types of blood samples tested for LFV antibodies.
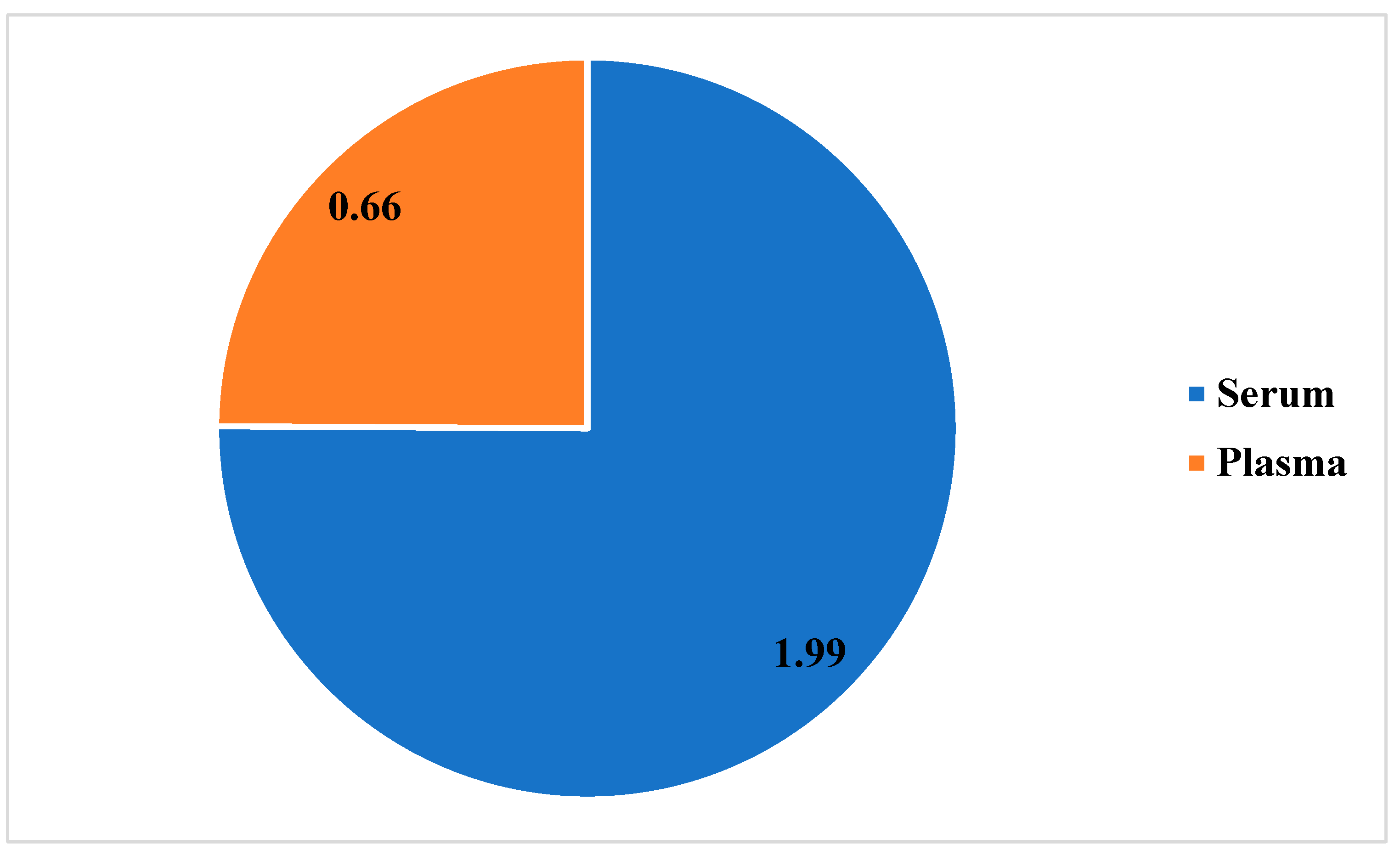
Figure 7.
Absolute number of blood sample types tested and % prevalence of LF infection positive cases recorded.
Figure 7.
Absolute number of blood sample types tested and % prevalence of LF infection positive cases recorded.
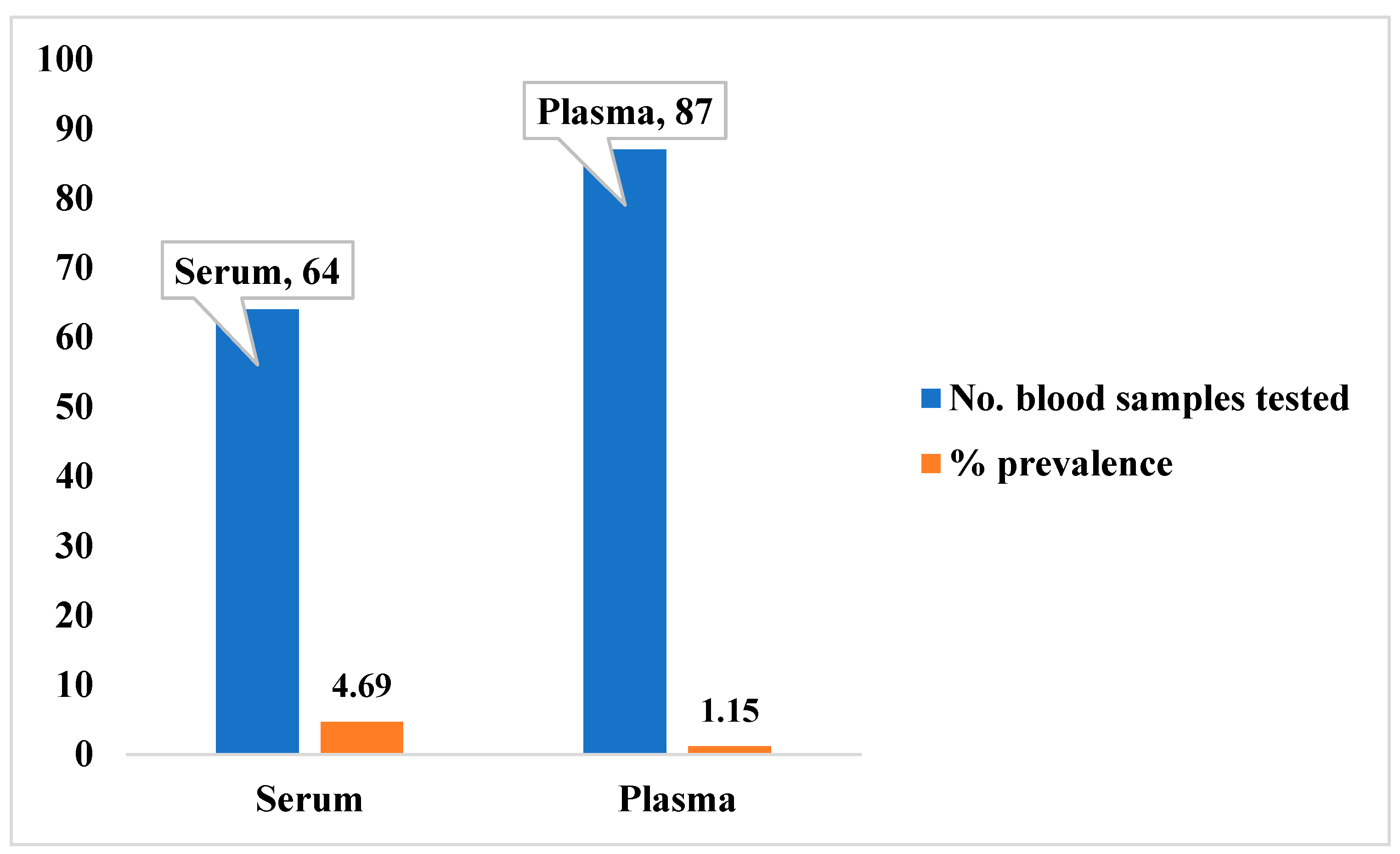

Table 1.
Number of individuals of rodents and rats and capture success rate per sampling community .
Table 1.
Number of individuals of rodents and rats and capture success rate per sampling community .
| Capture rate (%) for: | |||||||||
| Communities in Ewulo Ward | Communities in Otukpo Central Ward | Communities in Otukpo Town East Ward | |||||||
| Rodents and rats’ taxa | A | B | C | D | E | F | G | H | I |
| Mus musculus | (9) 47.37% | (3) 12.50% | (1) 5.0% | (9) 45.0% | (7) 14.0% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% |
| Rattus rattus | (12) 63.16% | (1) 4.17% | (2) 10.0% | (15) 27.0% | (4) 80% | (15) 37.5% | (5) 66.67% | (1)5.6% | (2) 18.5% |
| Rattus norvegicus | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (1) 18.8% | (0) 0.00% | (0) 0.00% | (0) 0.00% |
| Tatera spp. | (4) 10.50% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% |
| Pallasiomys spp. | (1) 5.26% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% | (0) 0.00% |
Note: LGA wards/communities: Ewulo Ward: A = Igbanonmanje; B = Otada; C = Upu; Otukpo Town Central Ward: D = Timber Depot 1; E = Timber Depot 2; F = Otukpo main market axis; Otukpo Town East Ward: G = Tiv market axis; H = Ogiri Oko street; I = General Hospital axis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



